
Minimal Recruitment and Activation of Dendritic Cells Within Renal Cell Carcinoma
10
1998;4:585-593. Published online March 1, 1998. Clin Cancer Res A J Troy, K L Summers, P J Davidson, et al. renal cell carcinoma. Minimal recruitment and activation of dendritic cells within Updated Version http://clincancerres.aacrjournals.org/content/4/3/585 Access the most recent version of this article at: Citing Articles http://clincancerres.aacrjournals.org/content/4/3/585#related-urls This article has been cited by 20 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications American Association for Cancer Research Copyright © 1998 on July 17, 2011 clincancerres.aacrjournals.org Downloaded from
Transcript of Minimal Recruitment and Activation of Dendritic Cells Within Renal Cell Carcinoma

1998;4:585-593. Published online March 1, 1998.Clin Cancer Res A J Troy, K L Summers, P J Davidson, et al. renal cell carcinoma.Minimal recruitment and activation of dendritic cells within
Updated Version http://clincancerres.aacrjournals.org/content/4/3/585
Access the most recent version of this article at:
Citing Articles http://clincancerres.aacrjournals.org/content/4/3/585#related-urls
This article has been cited by 20 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
[email protected] atTo order reprints of this article or to subscribe to the journal, contact the AACR Publications
To request permission to re-use all or part of this article, contact the AACR Publications
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

Vol. 4, 585-593, March 1998 Clinical Cancer Research 585
Minimal Recruitment and Activation of Dendritic Cells within Renal
Cell Carcinoma’
Andrew J. Troy, Kelly L. Summers,
Peter J. T. Davidson, Christopher H. Atkinson,
and Derek N. J. Hart2
Hematology/immunology Research Group [A. J. T., K. L. S.,D. N. J. H.], Department of Urology [A. J. I., P. J. I. D.], OncologyService [C. H. A.], Christchurch Hospital, Christchurch, New Zealand
ABSTRACT
Dendritic cells (DCs) are predicted to participate innatural tumor immunity by migrating into tumors, where
they acquire antigen, undergo activation, and migrate tolymph nodes to initiate a T-lymphocyte response against
tumor-associated antigens. The presence of DCs using de-fined lineage markers and their function in human tumorshas not been assessed previously. The monocbonal antibodiesagainst CMRF-44 and CD83, which are differentiation/ac-tivation antigens on DCs, were used in immunohistobogicaland flow cytometry studies to analyze the DC subtypesinfiltrating 14 cases of human renal cell carcinoma (RCC).The functional immunocompetence of the DCs isolated fromRCC was assessed by testing their ability to stimulate an
allogeneic mixed leukocyte reaction. The majority of leuko-
cytes present within the RCC were macrophages (62% ±
14.7) or T lymphocytes (19% ± 9.5), with CD45” HLA-DR�
lineage-negative putative DCs accounting for less than 10%of the leukocytes present. Of these, a subset, comprising lessthan 1 % of total leukocytes, had an activated CMRF-44” orCD83� DC phenotype. Activated CMRF-44� and CD83�DCs were more evident outside the tumor in association withT-lymphocyte clusters. The number of CMRF-44� DCs cor-related closely with the number of 5-100-positive DCs. Iso-lation of DCs from eight RCCs was achieved, and flowcytometry studies confirmed the small proportion of acti-vated CMRF-44� DCs. The CMRF-44� DCs stimulated anallogeneic mixed leukocyte reaction, but the CMRF-44
DCs (normal tissue DC precursors and other cells) failed todo so. These results suggest that RCCs recruit few DCs intothe tumor substance, and the tumor environment fails toinitiate the expected protective activation of DCs. These two
Received 8/4/97; revised I 1/18/97; accepted 12/1/97.The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to
indicate this fact.I A. J. I. was supported by a Schering Plow Fellowship from the Uro-logical Society of Australasia. Running costs for the project were
provided by the generosity of Urology Associates and the CancerSociety of New Zealand.2 To whom requests for reprints should be addressed, at HematologyDepartment, Christchurch Hospital, P. 0. Box 151, Christchurch, NewZealand. Phone: 64 3 3640 300; Fax 64 3 3640 750.
mechanisms, amongst others, may contribute to tumor es-
cape from immunosurveillance. In vitro loading of DCs with
tumor-associated antigens may be a useful therapeutic ma-neuver.
INTRODUCTION
DCs3 are specialist antigen-presenting cells that are found
in the interstitium of tissues and within epithelial surfaces (1, 2).
If DCs are to play a part in natural tumor immunity, DCs would
be expected to migrate into tumors, derive antigen, undergo
activation, and migrate to central lymphoid tissue to initiate any
T-lymphocyte response against TAAs. The failure of DCs to
migrate from the blood into malignant tissue or to be activated
and migrate out into draining lymph nodes represents a potential
tumor escape mechanism. Little is known about the migration of
DCs into human tumors, their state of activation and their
interactions, if any, with responding T lymphocytes. The immu-
nohistobogical studies performed to date using S-bOO staining
have shown that an increased number of DCs located within
tumors correlate with a better prognosis in coborectal adenocar-
cinoma (3); adenocarcinoma of the lung (4, 5); gastric (6),
esophageal (7), and nasopharyngeal (8) carcinomas; and papil-
lary carcinoma of the thyroid (9). These studies have not been
correlated with more detailed immunohistobogical analyses us-
ing mAbs. Furthermore, the widespread interest in using blood
DCs as “nature’s adjuvant” for tumor immunotherapy (10)
makes an understanding of how DCs function within tumors
vital for evolving strategies to use DCs in effective tumor
vaccines.
RCC is the most common malignancy of the kidney and
accounts for approximately 2-3% of all adult neoplasms (1 1).
The observation that large numbers of lymphocytes are often
found in RCC has encouraged investigators to develop immu-
notherapeutic treatments but with limited success (12, 13). The
presence or absence of DCs in RCC is unknown, despite the fact
that interstitial DCs were first defined in normal kidney some
time ago (14, 15). A recent study (16) described infiltration of
S-bOO-positive cells into RCC but not into benign adenomas.
Difficulties with the specificity of S- 100 expression and the
solubility of S-bOO has limited its use for characterizing DCs
further.
DCs have been identified in blood (17) and normal tissues
(14, 15) by demonstrating their high density expression of MHC
class II antigens and their lack of other mature hematopoietic
lineage markers (lin) using immunocytobogical techniques.
The recent availability of the relatively more DC-specific, mAbs
3 The abbreviations used are: DC, dendritic cell: TAA, tumor-associatedantigen; mAb, monocbonal antibody; RCC, renal cell carcinoma: NK,natural killer; HLA, human leukocyte antigen: FACS. fluorescence-activated cell sorter; MLR, mixed leukocyte reaction.
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

586 Dendritic Cells in Renal Cell Carcinoma
CMRF-44 (18) and HBlSa (19), combined with double-labeling
techniques. allows more accurate identification of DCs, as well
as providing information on their state of differentiation/activa-
tion. The CMRF-44 antibody recognizes an early activation
antigen, which is expressed on blood DCs, only after a short
period of culture ( 18, 20). HB1Sa detects the CD83 antigen, a
member of the immunogbobulin superfamily with an as yet
unknown function (21), which is expressed by activated DCs
and monocyte-derived DCs generated in culture with cytokines
(Mo-DC: Ref. 22).
In this study on DCs in RCC, we identified the CD45�/
HLA-DR�/lin� DC population in RCC and adjacent normal
kidney. and used the CMRF-44 and HBl5a mAbs to define the
activated DC subsets. Expression of the CDla antigen (23), a
marker of the Langerhans cell subset of DCs as well as the
up-regulation of the critical costimulator molecules, CD8O and
CD86 (24), were also assessed. These results and the functional
data on the DCs isolated from RCC suggest that deficient DC
activation and migration may contribute to RCC escape from
immune surveillance.
MATERIALS AND METHODS
Tissue Samples. Fresh tumor tissue was collected, fol-
lowing informed consent, from 14 radical nephrectomy speci-
mens, all removed for RCC. All patients were well at the time
of surgery with no evidence of metastases, and only one case (2)
had invasion of the resection margin of Gerotas fascia. There
were nine clear cell tumors, three papillary tumors, and two
granular tumors. After initial assessment by a histopathobogist,
samples were processed for routine histopathobogy. Samples for
immunohistobogy were mounted in OCT compound (Tissue
Tek), snap-frozen in liquid nitrogen. and stored at -80#{176}Cuntil
required for sectioning. Additional fresh tumor tissue was pro-
cessed immediately when available for further studies on the
isolated cell populations. For 5-100 staining, tissues obtained
from the same specimens were embedded in paraffin.
mAbs. The anti-CD3 (OKT3, IgG2a), anti-CD19
(FMC63, IgG2a), and anti-HLA-DR (L243, IgG2a) antibodies
were produced from hybridomas obtained from the American
Type Culture Collection (Rockville, MD). The antibodies
CMRF-l2 (CD45, IgGl), CMRF-lS (negative control, 1gM),
CMRF-3l (CD14, IgG2a), and CMRF-44 (activation antigen.
1gM: Ref. 18) were produced and characterized in this labora-
tory. The anti-CD83 mAb (Hbl5a, IgG2a; Ref. 19) was a gift
from Prof. T. F. Tedder (Durham, NC), and the anti p55 (K-2,
IgG I ) mAb was provided by Dr. E. Langhoff (Boston, MA; Ref.
25). The anti-CD16 mAb (HuNK-2, IgG2a) was a gift from
Prof. I. F. C. McKenzie (Melbourne, Australia), and the Na 1/34
mAb (CDIa, IgG2a) was a gift from Prof. A. J. McMichael
(Oxford, United Kingdom). The anti-CD8O (IgGla) mAb was
purchased from Becton Dickinson (Australia), and the anti-
CD86 (IgG2a) mAb was purchased from Serotek (United King-
dom). The anti-S-lOO polyclonal antibody was purchased from
Dako Corp. (Carpinteria, CA). The negative control mAb SalS
(IgG2a) and X63 (IgGI) were gifts from Prof. H. Zola (Ade-
laide, Australia). Phycoerythrin-conjugated CD14, CD19, and
negative control mAbs (IgG 1 , IgG2a) were purchased from
Becton Dickinson (Australia).
Immunoperoxidase (Single-Label) Staining. Cryostat-
cut sections (-20#{176}C; 4-6 p.m) were placed on gelatin-coated
slides and air-dried overnight, prior to fixing for 10 mm in cold
acetone (4#{176}C).Sections were prebbocked with 10% human AB
serum for 30 mm, followed by the application of the primary
mAb for 30 mm at room temperature. Sections were then
washed three times with PBS and then incubated with peroxi-
dase-conjugated goat anti-mouse IgG (Dako) for an additional
30 mm. After washing twice in PBS and once in TrisfHCl buffer
(pH 7.6), the sections were developed with 0.1% 3,3’-diamino-
benzidine for 3-10 mm before counterstaining with hematoxy-
bin.
S-100 staining was performed on sections cut from forma-
bin-fixed, paraffin-embedded tissues. These were dewaxed and
rehydrated before staining as for fresh tissue, except that rabbit
anti-S-bOO and peroxidase-conjugated goat anti-rabbit IgG (Bio-
source International, CA) were used.
Immunoalkaline Phosphatase/Peroxidase (Double-Label)
Staining. Sections were treated as for single staining up to the
first primary mAb incubation. Following this, sections were
incubated with biotinylated goat anti-mouse IgG (Dako) diluted
1:200 for 30 mm, washed three times in PBS, and then incu-
bated for 30 mm in Extravidin#{174} alkaline phosphatase (Sigma
Chemical Co., St. Louis, MO) at 1 :200 dilution. After washing
two times in PBS and once in TrisIHCL buffer (pH 7.6), the
sections were developed with Fast Blue (Sigma) for 5-10 mm.
Sections were then washed with PBS before a 30-mm incuba-
tion in 10% mouse serum. Incubation with the second mAb was
carried out as for single immunoperoxidase staining, including
developing the color reaction with 3,3’-diaminobenzidine.
Immunohistobogical Assessment. T lymphocytes (CD3).B lymphocytes (CD19), NK cells (CD16), and monocyte/macro-
phages (CD14 and CD1 Ib) were identified and counted as cells
when identifiable nuclei were present. Potential DCs were identi-
fled as CD45�. lin (CD3, CDI9, CDI6, CD14, and CD1 lb)
using double-labeling and further confirmed to be HLA-DR posi-
tive. CMRF-44� DC and CDla� LC were identified by double-
labeling with CD14 to exclude macrophages, which may also
express these antigens. CMRF-44� and CD83� DCs were consid-
ered on the basis of data published previously (18, 19) to be
partially activated.
Sections were examined at X400 for cell types present in
large numbers and at X 100 for cell types present in low num-
bers through an eyepiece graticube, giving a field of view of
0.044 and 0.772 mm2, respectively. All counts were converted
to number per mm2 for comparison. Positively stained cells
were counted in 5-10 random, nonoverlapping fields of tumor
tissue, excluding areas containing dense stroma or large lymph-
oid aggregates. Cell numbers are expressed as a percentage of
the total leukocyte count (mean % ± SD), as determined by
CD4S staining. Separate counts were made within the area of
lymphoid aggregates when present.
Isolation of Tumor Containing Leukocyte Populations.
Tissues were minced into small pieces in sterile 10% FCS/RPMI
media (RPMI 1640 supplemented with 100 units/mb penicillin,
1 m�i glutamine, and 100 units/mI streptomycin) containing
colbagenase (0.5 mg/mI) and DNase (1 .0 pg/mb) and incubated
at 37#{176}Cfor 1-2 h with gentle agitation. The resulting cell
suspension was washed in PBS twice before filtering through a
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

Clinical Cancer Research 587
Table I DC response to each tumor
Cell counts are expressed as the number of cells per mm2 and are also expressed as a percentage of total CD4S leukocytes (boldface). S- 100counts were not converted to percentages, because these were obtained from different samples of the same tumor. Counts were averaged over S fields
for CD45 and CDI4 and 10 fields for CMRF-44 and CD83. Individual I-lymphocyte and NK cell counts have been omitted for clarity (theyrepresented 8-40% of leukocytes; see Fig. 3A).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 Mean±SD
Histology” CC CC CC P CC CC CC CC CC” p CC P G G
Stage (TNM)’ T� T4 13b T3� T3� T35 Tib T� T2 T3� 1, 13b 12 T�b
Total leukocytes 531 603 349 413 363 640 1152 739 631 619 513 925 308 354 581 ± 239(CD45)
Total lin 82 82 41 45 23 122 91 127 45 102 73 127 36 68 76 ± 35
leukocytes 15 14 12 11 6 19 8 17 7 16 14 13 12 19 13.1±4.1%lin DCs 17.9
3.420.0
3.321.1
6.0
12.1
2.9
15.2
4.2
24.6
3.8
41.4
3.6
19.0
2.6
19.6
3.1
16.1
2.6
21.8
4.2
25.3
2.7
8.0
2.6
11.5
3.2
19.5 ± 8.0
3.4 ± 0.9%
CMRF44� DCs 0.500.9
0.300.5
1.20.35
0.700.17
0.28
0.081.6
0.250.70
0.061.0
0.140.0
0.00.69
0.112.50.49
0.97
0.100.35
0.110.28
0.080.79 ± 0.650.15 ± 0.13%
CD83 DCs 1.7
0.320.7
0.121.6
0.461.4
0.340.55
0.153.50.55
3.5
0.301.6
0.220.18
0.030.18
0.031.1
0.212.3
0.250
0
0.69
0.19
1.4 ± 1.1
0.23 ± 0.16%
CDIa LCs 0.35
0.070.20.04
2.20.43
7.21.7
0.280.08
0.820.13
1.80.16
0.55
0.076.51.03
3.20.52
0.14
0.03
1.2
0.13
0.0
0.0
1.9
0.54
1.8 ± 2.3
0.35 ± 0.48%CD86� DCs 0.0
00.0
00.0
0
0.0
0
0.0
0
0.0
0
0.14
0.010.0
00.0
00.0
00.0
00.0
00.0
00.0
00.01 ± 0.04
0.00%
S-l00 NA” NA NA 9.5 0.14 0 1.1 1.9 3.6 0.7 0.3 NA 0.7 0.8 1.87 ± 2.9
“ CC, clear cell: P. papillary clear cell; G. granular clear cell.h Cystic variety of clear cell.
C. ThM, Tumor-Node-Metastasis classification.d NA, not available.
70 �im nylon cell strainer (Falcon; Becton Dickinson, Australia)
to remove undigested material. This single-cell suspension was
layered over a Ficoll-Hypaque density gradient (density, 1.077;
Pharmacia, Uppsala, Sweden) and centrifuged at 400 X g for 20
mm to obtain mononuclear cells. This step excluded granubo-
cytes but retained some tumor cells (identified as CD45 cells).
Cell viability was always >80%, as confirmed by trypan blue
exclusion.
Flow Cytometry Analysis and Sorting. Mononuclearcells were labeled with primary mAb for 30 mm at 4#{176}C.Fob-
bowing one wash in PBS, labeled cells were detected by incu-
bation with FITC-conjugated sheep anti-mouse IgG (Silenus
Laboratories, Australia) for 30 mm at 4#{176}C.Following blocking
for 10 mm in 10% mouse serum, cells were double-labeled with
phycoerythrin-conjugated CD14 mAb. Appropriate gating and
flow cytometry analysis were performed using Lysis software
on a FACS (FACS Vantage; Becton Dickinson). Contaminating
CMRF-44�-activated B lymphocytes were not considered, be-
cause B lymphocytes were not present in the mononuclear cell
populations. For analysis of DC phenotype, mononuclear cells
were labeled with a mix of lineage markers (CD3, CDI lb,
CD14, CD16, and CD19), and the lin population was sorted
and then reanalyzed for the expression of DC activation anti-
gens.
For functional studies, the mononuclear cell suspension
was sorted after labeling, as described in the results. Single
labeling was used for three experiments. In an additional five
double-labeling studies, the four different populations consist-
ing of CMRF-44�/CDl4 (activated DCs), CMRF-44�/CD l4�
(activated monocytes), CMRF-44/CDl4� (resting mono-
cytes), and CMRF-441CD14 (other cells, including resting
DCs) were isolated.
MLR. Tumor cell suspension and the sorted leukocyte
populations were cocultured with 1 X l0� allogeneic normal
peripheral blood T lymphocytes. Cultures were maintained at
37#{176}Cand 5% CO2 in 96-microwell, round-bottomed plates for 6
days. Wells were pulsed with [3Hjthymidine (5 Ci/mmol; Am-
ersham) in the last 16-18 h of culture. [3HjThymidine incorpo-
ration was measured on a liquid scintillation analyzer to give a
direct readout of T-lymphocyte proliferation. Responses were
reported as the mean value ± SE of triplicate wells.
RESULTS
Immunohistological Analysis of the Leukocytes InfiltratingRCC
Identification of Leukocyte Types within Cryostat
Sections. A summary of the results of the immunostaining of
the 14 tumors analyzed are shown in Tables 1 and 2 and Figs.
1 and 2. The majority of the leukocytes infiltrating the RCCs
were either CDl4� macrophages or CD3� T lymphocytes,
accounting for 62% ± 14.7 and 19% ± 9.5, respectively, of the
leukocytes present. NK cells (CDl6� lymphoid cells) were
present in small numbers (1 1% ± 8. 1), whereas B lymphocytes
were virtually never detected. Total CD45�/lin cells, which
include the DCs, comprised 13. 1% ± 4. 1 of the total leukocyte
population in RCCs. These numbers were not significantly
different from the proportion of CD45�/lin leukocytes seen in
normal kidney (12.7% ± 3.0). Morphobogically, at least 25% of
these were large cells with dendritic processes, suggesting a
degree of activation (Fig. 1A). When only these cells were
counted, the potential DCs were found to comprise 3.4% ± 0.9
in RCCs and 4.4% ± 2. 1 in normal kidney. Double-labeling to
identify lin and HLA-DR� cells, although difficult to analyze
accurately due to HLA-DR expression on endothelium and the
tumor cells, gave cell counts similar to the CD45�fbin counts,
with a minimum estimate of 50% of these cells having a
CD45�fl-ILA-DR�/lin phenotype. CDla�/CDl4 LC ac-
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

588 Dendritic Cells in Renal Cell Carcinoma
Table 2 DC numbers in T-bymphocyte aggregates at tumor margins
All counts were made at X400 and converted to numbers per mm2 on matched lymphoid areas (three to five, depending on sample) for CMRF-44and CD83. Direct comparison of 5-100 counts was not possible because different samples of the tumor are required for processing.
Case number
DC phenotype 2 5 6 7 8 9 10 1 1 14
CMRF-44� CDI4 136 556 447 415 513 386 284 358 449
CD83� CD14 113 426 295 243 333 147 159 352 277
5-100 NA” NA 95 177 150 132 95 NA 159
a NA, not available.
counted for 0.35% ± 0.48 of leukocytes in RCCs and 0.02% ±
0.03 of leukocytes in normal kidney and was elevated in RCCs
due to the increased counts in the cases with papillary and cystic
RCCs.
Identification of 5-100 Cells in Paraffin Sections. The
results for the 5- 100 antigen staining in paraffin sections cannot
be compared directly with the frozen sections because the for-
mer are necessarily cut from a slightly different area of the
tumor. Nonetheless, staining for S-bOO antigen identified only
small numbers of positive cells in 9 of 10 cases (Fig. 1D; Table
1 ). They were present in greater numbers where T lymphocytes
had accumulated. In over one-half the cases, weak S- 100 stain-
ing of tumor cells was noted.
Identification of Activated DCs. CD14 beukocytes,
which expressed the CMRF-44 antigen, i.e., activated DCs,
accounted for only 0.15% ± 0.13 of the total leukocytes infib-
trating the tumors. The CD83 antigen, which is induced after the
CMRF-44 antigen (20) and may undergo some subsequent
down-regulation (26), was expressed by a similar number
(0.23% ± 0. 16) of tumor leukocytes (Fig. 1C). These data
suggested that only a small proportion (5-10%) of the lin/
HLA-DR� putative DCs identified within the RCCs are acti-
vated. The number of activated DCs in the adjacent normal
kidney was much less, with only 0.04% ± 0.04 CMRF-44�/
CD14 cells and 0.04% ± 0.04 CD83� cells present. No
specific localization of DCs was found within the tumors with
respect to necrotic or inflammatory areas.
Staining for the p55 antigen (25) was also performed, but
accurate assessment was difficult because of tumor cell staining
in close to one-half of the cases. In those cases that could be
assessed, the number of p55-positive cells was similar to the
number of CMRF-44- and CD83-staining cells.
The expression of the CD8O and CD86 costimulator mob-
ecules was examined. No CD80� cells were detected in either
normal or RCC tissue. Small numbers of CD86� cells were
present in many of the cases, but when double staining with
CD14 was performed to exclude CD86� macrophages, only one
case was found to have CD86�/CDl4 cells present.
Although the number of activated DCs within the RCCs
was limited, CMRF-44 � or CD83 � DCs were more evident just
outside the tumor in the pseudocapsule. Here, relatively large
numbers of activated DCs were seen in the T-lymphocyte clus-
ters that develop. In nine tumors, in which sufficiently well-
defined T-cell aggregates had formed, it was possible to count
the number of DCs in the clusters (Fig. 1, E and F; Table 2).
Perhaps consistent with the earlier induction of the CMRF-44
antigen on blood DCs in vitro (20), more CMRF-44�/CDl4
cells than CD83�/CD14 cells were present in the bymphoid
areas.
FACS Analysis of Leukocytes in Single-Cell TumorSuspensions
Flow cytometric analyses on the mononuclear cell sus-
pensions isolated from digested tumor tissue were performed
on eight tumors (Table 3). Side scatter and forward scatter
were used to define typical “bymphoid” and “monocytic-
macrophage” electronic gates. The bymphoid gate contained
>95% CD4S’ leukocytes and consisted predominantly of
CD3� cells. The monocytic-macrophage gate, containing
>85% CD4S’ leukocytes, was analyzed more extensively to
identify potential DC populations. CD14� cells accounted for
over 50% of the monocytic-macrophage gate. A variable
number of these macrophages expressed the CMRF-44 anti-
gen and accounted for the bulk of the CMRF-44�-positive
cells within the tumors (Fig. 3A). Activated CMRF-44’/
CD14 DC comprised only 1.5% ± 1.02 of the total mono-
cyte-macrophage gated cell population. The contaminating
CD45 tumor cells did not express the CMRF-44 or CD14
antigens. The remaining population of CD14/CMRF-44
cells was presumed to contain unactivated DCs, contaminat-
ing tumor cells, and occasional CD14 lymphoid cells. Over-
all, these leukocyte populations occurred in similar propor-
tions to that observed in direct tissue staining.
To facilitate a more direct comparison of the flow cytom-
etry data with the immunohistobogical results, lin cells were
sorted, and the CD4S’ cells were analyzed in five cases (Fig.
3B). These represented 8.1% ± 2.4 of the total leukocytes
(lymphoid and monocyte-macrophage gates). Approximately
one-half these cells were HLA-DR�, i.e., approaching the pro-
portion of total DCs identified by tissue staining. The CMRF-44
antigen was expressed on only a subpopulation of lin cells
(15.6% ± 9.0), i.e., - 1% of total leukocytes, a figure that
relates closely to the bow percentage of CMRF-44� DCs esti-
mated by direct staining. The CD83 antigen was not detected by
this method, possibly because of its sensitivity to proteases.
Functional Studies
To examine the functional capacity of these tumor leuko-
cyte populations as antigen-presenting cells, various cell popu-
lations were tested as stimubators in an albogeneic MLR. Limited
cells were available in each case; therefore, only certain sorted
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

C�
.t
4 s.C.
�;j� -�
C D
E
,�.-. C. ..,;&.
.�: .t
F
. � e’
#{149}�:-:��
Clinical Cancer Research 589
1, � /(.�
)i� �Fig. I Immunohistobogical analysis identifies a CD45�fHLA-DR�ilin population of putative DCs, a subset of which express the CMRF-44 CD83
differentiation/activation antigens within RCC. A, a putative DC (arrow) stained for CD45 (golden brown) but not CD3, CD1 lb. CDI4, CDI6, or
CDI9 (lin. blue black) compared with the CD14-positive macrophage (arrowhead, X400). B, single CMRF-44�/CDl4 DC (arrow) surroundedby many CDl4� macrophages (blue, X400). C. CD83� DCs were seen among RCC cells (single-label immunoperoxidase). D, S-lO0- DC (white
arrow) in RCC (single-label immunoperoxidase). Note weak S-l00� tumor cells. DCs are found in lymphoid aggregates at the margins of RCCs. E.
CMRF-44�/CDl4 DC (yellow) and CDl4� macrophages (blue) are identified (no counterstain). F, CD8Y DCs are seen among the lymphocytes
(counterstained) in a lymphoid aggregate.
populations could be tested. The first two analyses established 4447CD14 DCs were found to be at least three to five times
that the RCC leukocyte population was albostimulatory (data not more potent stimulators of the abbogeneic T-lymphocyte re-
shown) and that the lin population contained the albostimula- sponse than the CD14� macrophages (Fig. 4B). The CMRF-44
tory cells (data not shown). An additional case was analyzed cells, i.e. , the population that included the presumed resting or
after sorting the tumor mononuclear cell suspension into unactivated tumor-associated DCs (and some contaminating tu-
CMRF-44� and CMRF-44 populations. The CMRF-44� pop- mor cells), failed to stimulate T lymphocytes in all five cases
ubation was clearly the most effective at stimulating the albo- tested. Blood DCs, sorted from two additional patients, were
MLR (Fig. 4A). Five additional cases were sorted using double- found to fully stimulate T lymphocytes in an albogeneic MLR
labeling to identify the CMRF-44�/CDl4 DCs. The CMRF- (Fig. 4D).
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from
Leukocyte subsets in RCC
U)
EC
U)
U0
.�
.2
Is
0
05)
Is
C5)
C.)
a
U)5)
>.U0
.�
.2
Is
0
0
CU)U
U)0.
590 Dendritic Cells in Renal Cell Carcinoma
75
50
25
0
U
A
#{149}.
U..
U
:U
U
#{163}
�
--�4�#{163} .#{163}AA . 0
.. 00 � �
w�w I
CDI4 CD3 CDI9 CDI6 TotalS, IinDC 44DC 83�DC CDIa
cell type
DC subtypes in normal kidney and RCC
20
15
10
5.
n,
U BU
U.
o UU
0 UU
-#{176}n-- 0 normal kidney0 � .RCC0
0 U 0
U0
U 0 U
U.
�0i�
U
�Total un- lin’ DC 44�DC CD83� DC CD1a�LC
cell type
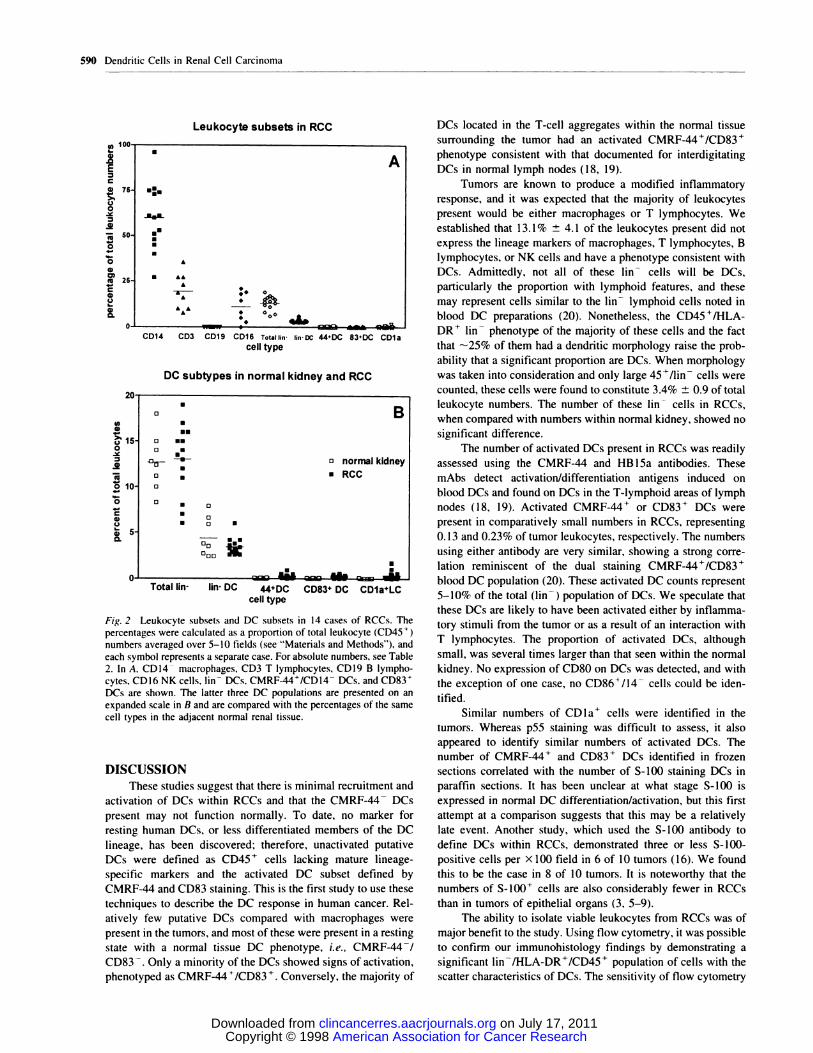
Fig. 2 Leukocyte subsets and DC subsets in 14 cases of RCCs. The
percentages were calculated as a proportion of total leukocyte (CD45 �)
numbers averaged over 5-10 fields (see “Materials and Methods”), and
each symbol represents a separate case. For absolute numbers, see Table
2. In A. CD14 macrophages. CD3 T lymphocytes, CD19 B lympho-cytes, CDI6 NK cells, lin DCs, CMRF-44�/CDI4 DCs. and CD83�
DCs are shown. The latter three DC populations are presented on anexpanded scale in B and are compared with the percentages of the same
cell types in the adjacent normal renal tissue.
DISCUSSION
These studies suggest that there is minimal recruitment and
activation of DCs within RCCs and that the CMRF-4�F DCs
present may not function normally. To date, no marker for
resting human DCs, or less differentiated members of the DC
lineage, has been discovered; therefore, unactivated putative
DCs were defined as CD4S’ cells backing mature lineage-
specific markers and the activated DC subset defined by
CMRF-44 and CD83 staining. This is the first study to use these
techniques to describe the DC response in human cancer. Rel-
ativeby few putative DCs compared with macrophages were
present in the tumors, and most of these were present in a resting
state with a normal tissue DC phenotype, i.e., CMRF-44/
CD83 - . Only a minority of the DCs showed signs of activation,
phenotyped as CMRF-44�/CD83�. Conversely, the majority of
DCs located in the T-cell aggregates within the normal tissue
surrounding the tumor had an activated CMRF-44�/CD83�
phenotype consistent with that documented for interdigitating
DCs in normal lymph nodes (18, 19).
Tumors are known to produce a modified inflammatory
response, and it was expected that the majority of leukocytes
present would be either macrophages or T lymphocytes. We
established that 13.1% ± 4.1 of the leukocytes present did not
express the lineage markers of macrophages, T lymphocytes, B
lymphocytes, or NK cells and have a phenotype consistent with
DCs. Admittedly, not all of these lin cells will be DCs,
particularly the proportion with lymphoid features, and these
may represent cells similar to the lin lymphoid cells noted in
blood DC preparations (20). Nonetheless, the CD45�fHLA-
DR� lin phenotype of the majority of these cells and the fact
that -25% of them had a dendritic morphology raise the prob-
ability that a significant proportion are DCs. When morphology
was taken into consideration and only large 45�/lin cells were
counted, these cells were found to constitute 3.4% ± 0.9 of total
beukocyte numbers. The number of these lin� cells in RCCs,
when compared with numbers within normal kidney, showed no
significant difference.
The number of activated DCs present in RCCs was readily
assessed using the CMRF-44 and HB 1Sa antibodies. These
mAbs detect activation/differentiation antigens induced on
blood DCs and found on DCs in the T-lymphoid areas of lymph
nodes (18, 19). Activated CMRF-44� or CD83� DCs were
present in comparatively small numbers in RCCs, representing
0. 13 and 0.23% of tumor leukocytes, respectively. The numbers
using either antibody are very similar, showing a strong corre-
lation reminiscent of the dual staining CMRF-44’7CD83�
blood DC population (20). These activated DC counts represent
5-10% of the total (bin) population of DCs. We speculate that
these DCs are likely to have been activated either by inflamma-
tory stimuli from the tumor or as a result of an interaction with
T lymphocytes. The proportion of activated DCs, although
small, was several times larger than that seen within the normal
kidney. No expression of CD8O on DCs was detected, and with
the exception of one case, no CD86�/l4 cells could be iden-
rifled.
Similar numbers of CDla� cells were identified in the
tumors. Whereas p55 staining was difficult to assess, it also
appeared to identify similar numbers of activated DCs. The
number of CMRF-44� and CD83� DCs identified in frozen
sections correlated with the number of 5-100 staining DCs in
paraffin sections. It has been unclear at what stage 5-100 is
expressed in normal DC differentiation/activation, but this first
attempt at a comparison suggests that this may be a relatively
late event. Another study, which used the 5-100 antibody to
define DCs within RCCs, demonstrated three or less S-bOO-
positive cells per X 100 field in 6 of 10 tumors (16). We found
this to be the case in 8 of 10 tumors. It is noteworthy that the
numbers of S-l0O� cells are also considerably fewer in RCCs
than in tumors of epitheliab organs (3. 5-9).
The ability to isolate viable beukocytes from RCCs was of
major benefit to the study. Using flow cytometry, it was possible
to confirm our immunohistobogy findings by demonstrating a
significant bin�fHLA-DR�/CD45� population of cells with the
scatter characteristics of DCs. The sensitivity of flow cytometry
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

A
ri�
B
‘I.
c)
CONTROL
5b’5� (bit)) ‘4’)lo (I 175) 15% (236)
Clinical Cancer Research 591
Table 3 FACS a nalysis of tumor mon onuclear cell suspensi on (myeloi d gate)”
DC phenotype
Case number
Mean ± SD2 3 4 6 8 10 1 1 13
CMRF-44�/CDl4 DCs 0.0 1.0 1.4 1.3 3.5 2.3 1.2 1.2 1.5 ± 1.02
CMRF-44�/CDl4� macrophages 19.6 25.9 31.5 23.8 41.7 3.3 5.3 41.0 24.0 ± 14.4
CMRF-44/CDI4� macrophages 65.1 42.7 24.4 21.5 39.8 9.8 42.6 41.0 35.9 ± 16.9
CMRF-44�/CD14 cellsb 15.9 30.3 42.7 52.6 14.7 78.9 46.4 17.0 37.3 ± 22.4
a Results are expressed as percentages of each cell type in the myeloid gate. The myeloid gate included between S and 30% of total cells, and
when analyzed, >85% of CD45� cells. The nonleukocytes fall in the CMRF-44 CDI4 population.b Heterogeneous population including DCs (-50% HLA-DR� cells).
FSC
7.06% .� . ,�
�:
.- .
� � .....,.rt...
. . ). . 1.59%
CMRF44
CD3,llb,14,16,19 CD45 HLA-DR CMRF44
Fig. 3 FACS analysis of tumor leukocyte suspension. In A, the analysis was performed on the gated mononuclear population. Within the
mononuclear gate, the majority of the cells are CDl4� macrophages. Only 1.59% of this sample are CDl4/CMRF-44� DC (one of eightrepresentative analyses; see Table 3). B, analysis of sorted lin cells sorted from the mononuclear gate. In this sample, lin cells account for 7.06%.
The majority are CD45 leukocytes, with close to 50% of these being HLA-DR�, probable DCs. Only a small proportion (15%) of lin cells areactivated DCs expressing CMRF-44. Solid line, the test mAb: broken line, the control mAb labeling. Parentheses, mean channel fluorescence.
excludes the possibility that the lin cells included low density
CDl4� or CD3� cells, which were not detected by immuno-
histological techniques. Flow cytometry, likewise, confirmed
that CMRF�44tactivated DCs comprise a very small portion of
the tumor leukocyte infiltrate. We also compared the stimulatory
abilities of DCs and macrophages, albeit with difficulty, because
only limited numbers of cells were obtained from finite tissue
samples.
It is of some significance that the CMRF-44 DCs ob-
tamed from RCCs were relatively poor stimubators in our ex-
periments, although an additional effect from contaminating
tumor cells in the MLR cannot be readily excluded. Interstitial
DCs isolated from mouse and rat require a period of in vitro
culture to induce allostimubatory (costimubatory) activity (27).
However, in humans, functional albostimulatory activity in the
DC lineage arises early with the CD34� precursor (28) and
blood DCs (including the CMRF-44, freshly isolated DCs;
Ref. 29) stimulate a strong MLR. Isolated human Langerhans
cells also act as potent albostimubatory cells (30). Thus, the
failure of RCC-associated interstitial CMRF-44 DCs to stim-
ulate an albogeneic MLR is remarkable. Indeed, these results
have parallels with the situation in rheumatoid arthritis, where it
appears that despite an inflammatory environment, inhibitory
factors prevent full DC activation and expression of a costimu-
bator phenotype (31). Any effect on the DCs within RCCs
appears to be localized to the tumor because fresh blood DCs
from RCC patients were able to stimulate T lymphocytes max-
imally. Both interleukin 10 (32) and transforming growth factor
1� (33) have negative effects on DCs, and both have been shown
to be produced by RCCs (34).
The CDl4/CMRF-44� DCs were considerably more po-
tent stimubators of albogeneic T-bymphocyte proliferation than
were isolated CD l4� tumor macrophages, and this suggests that
at least some DCs avoid active tumor suppression of their
function. A predominantly activated CMRF-44� and CD83�
DC population was noted in the presence of T-lymphoid aggre-
gates. This population may represent the DCs induced to mi-
grate out of the tumor. We speculate that the interaction between
these CMRF-44� DCs and the corresponding T lymphocytes
may involve recognition of TAA. It is possible that these ag-
gregates represent a peripheral version of DC-T-lymphocyte
interactions normally seen in the lymph node. Equally, these T
lymphocytes (specific or nonspecific) may represent an efferent
response reacting to the DCs that have not yet reached regional
lymph nodes. Whether the relatively few CMRF-44� DC or T
lymphocytes noted in these clusters are capable of trafficking
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

E
� 20000
i� 10000
I.
B-.- CMRF44+
-‘- unsortedcells
-*-CMRF44-
0 T cells alone
A
C
E0.C.)
C
.2
a0C.)C5)
C
:2E>.
10 100 1000 10000
number of APC
1000C-U- RCC 44+ DC-e’- RCC44-M_,_ unsorted cells
7500 -*- RCC 44- DC
:: �aRC���:: ,
10 100 1000 10000 100000 100
number of APC
100000 1001 000
D
000
...- RCC 44+DC
-A-. RCC 44- DC
-0- Blood DC
0 1 �lIs�ne�f
10 loo 11*10 10000 100000 1000000
number of APC number of APC
592 Dendritic Cells in Renal Cell Carcinoma
.�. 500000.
C.)
.� 40000
.t�#{176}#{176}#{176}#{176}.� 20000
E
:�‘ 10000
Fig. 4 Analysis of tumor-infiltrating DC abbostimulatory potential. Individual tumor cell mononuclear cell suspensions were prepared, and the
electronically gated lymphoid and monocyte-macrophage cell populations were sorted after labeling. A, CMRF-44� cells but not CMRF-44 cells
are allostimulatory (one experiment). B, CMRF-447CD14 DCs are potent allostimulatory cells. CMRF-44� CD14’ cells stimulated minimally inthis experiment only. CMRF-44 cells were not allostimulatory (one of two experiments). Results are presented as [3H]thymidine uptake (mean cpm)
using 2 x l0� responders and the absolute stimulator cell numbers as indicated. C. CMRF-44� tumor DCs stimulate as potently as blood DCs, whereas
CMRF-44 tumor DCs stimulate poorly. In D, fresh blood DCs from two patients with stage T, tumors were able to stimulate T lymphocytesmaximally (as determined by phorbol myristate acetate and ionomycin stimulation). A-D, bars, SE.
normally is another important question. It has to be conceded
that little is known about the reaction of DCs in pathological
circumstances and to what extent these results reflect normal or
abnormal DC responses to tissue damage.
It seems that RCCs do not recruit DCs into the tumor above
the numbers seen in normal tissue: (a) these first clinical data
reinforce a similar concept proposed as a result of rat studies in
vivo (27), and recent experiments demonstrate that a tumor cell
line secreting vascubar-endothelial growth factor suppressed in
vitro DC production (35); (b) our functional studies on the
CMRF-44 tumor DC population also raise the possibility that
there is a component of the tumor environment that suppresses
their normal costimubatory function. This is supported by the
finding of the lack of CD8O or CD86 up-regulation; and (c) the
paucity of CMRF-44� DCs in RCCs also prompts us to spec-
ulate that RCCs lack the inflammatory/noxious signals, such as
lipopolysaccharide, tumor necrosis factor-a, or chemokines,
required to initiate significant DC activation/migration (36). If
DC function is compromised at these three levels, tumor escape
from immunosurveibbance is likely to result.
These data establish that the new reagents CMRF-44 and
CDS3 can be exploited to study DCs in tumor biology, and
extended studies on breast cancer are now in progress. Further-
more, it encourages the concept that in vitro loading of autobo-
gous DCs with TAA and subsequent vaccination of patients
with these DCs may reinforce one phase of the immune re-
sponse to RCCs sufficiently to initiate a therapeutic immune
response.
ACKNOWLEDGMENTS
We thank the urobogists and pathologists at Christchurch Hospitaland Medlab South for help in obtaining nephrectomy specimens.
REFERENCES
1 . Steinman, R. M. The dendritic cell system and its role in immuno-
genicity. Annu. Rev. Immunob., 9: 271-296, 1991.
2. Hart, D. N. J. Dendritic cells: unique leucocyte populations which
control the primary immune response. Blood, 90: 3245-3287, 1997.
3. Ambe, K., Mori, M., and Enjoji, M. S-l00 protein-positive dendriticcells in coborectal adenocarcinomas. Distribution and relation to the
clinical prognosis. Cancer (Phiba.). 63: 496-503, 1989.
4. Furukawa, T., Watanabe. S., Kodama, T., Sato, Y., Shimosato, Y.,
and Suemasu, K. T-zone histiocytes in adenocarcinoma of the lung in
relation to postoperative prognosis. Cancer (Phila.), 56: 265 1-2656,
1985.
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from

Clinical Cancer Research 593
5. Fox, S. B., Jones, M., Dunnill, M. S., Gatter, K. C., and Mason, D. Y.Langerhans cells in human lung tumors: immunohistological study.Histopathobogy, 14: 269-275, 1989.
6. Tsujitani, S., Kakeji, Y., Watanabe, A., Kohnoe, S., Maehara, Y., andSugimachi, K. Infiltration of dendritic cells in relation to tumor invasion
and lymph node metastasis in human gastric cancer. Cancer (Phila.), 66:2012-2016, 1990.
7. Imai, Y., and Yamakawa, M. Dendritic cells in esophageal cancerand lymph node tissues. In Vivo, 7: 239-248, 1993.
8. Nomori, H., Watanabe, S., Nakajima, T., Shiosata, Y., and Kameya,
T. Histiocytes in nasopharyngeal carcinoma in relation to prognosis.Cancer (Phila.), 57: 100-105, 1986.
9. Schroder, S., Schwarz, W., Rehpenning, W., Loning, T., and Bocker,w. Dendritic/Langerhans cells and prognosis in patients with papillarycarcinoma. Immunohistochemical study of 106 thyroid neoplasms cor-
related to follow up data. Am. J. Clin. Pathol., 99: 295-300, 1988.
10. Young, J. W., and Inaba, K. Dendritic cells as adjuvants for class Imajor histocompatibility complex-restricted antitumor immunity. J.Exp. Med., 183: 7-11, 1996.
1 1 . New Zealand Ministry of Health. Cancer: New registrations and
deaths, 1992. 6, 1995.
12. Goedegebuure, P. 5., Douville, L. M., Li, H., Richmond, G. C.,Schoof, D. D., Scavone, M., and Eberlein, T. J. Adoptive immunother-apy with tumor-infiltrating lymphocytes and interleukin-2 in patientswith metastatic malignant melanoma and renal cell carcinoma: a pilotstudy. J. Clin. Oncol., 13: 1939-1949, 1995.
13. Belldegrun, A., Pierce, W., Kaboo, R., Tso, C., Shau, H., Turcilbo,P., Moldawer, N., Golub, S., deKernion, J., and Figlin, R. Interferon-aprimed tumor-infiltrating lymphocytes combined with interleukin-2 and
interferon-a as therapy for metastatic renal cell carcinoma. J. Urol., 150:1384-1390, 1993.
14. Hart, D. N. J., Fuggle, S. V., Williams, K. A., Fabre, J. W., Ting, A.,and Morris, P. J. Localization of HLA-ABC and DR antigens in humankidney. Transplantation, 31: 428-433, 1981.
15. Daar, A. S., Fuggle, S. V., Hart, D. N. J., Dalchau, R., Abdulaziz,Z., Fabre, J. W., Ting, A., and Morris, P. J. Demonstration and pheno-typic characterization of HLA-DR positive interstitial dendritic cells
widely distributed in human connective tissues. Transplant Proc., 1:
311-315, 1983.
16. Ohmori, T., Okada, K., Arita, N., Watanabe, Y., Miyazaki, T.Characteristics of MHC antigen expression and tumor-infiltrating mono-
nuclear cells in renal cell adenomas and carcinomas. Histol. His-topathol., 10: 789-794, 1995.
17. Van Voorhis, W. C., Hair, L. S., Steinman, R. M., and Kaplan, G.Human dendritic cells. Enrichment and characterisation from peripheralblood.J.Exp.Med., /55: 1172-1187, 1982.
18. Hock, B. D., Starling, G. C., Daniel, P. B., and Hart, D. N. J.
Characterisation of CMRF-44, a novel monoclonal antibody to anactivation antigen expressed by the allostimulatory cells within periph-
eral blood, including dendritic cells. Immunology, 83: 573-581, 1994.
19. Zhou, L., Schwarting, R., Smith, H. M., and Tedder, T. F. A novelcell-surface molecule expressed by human interdigitating reticulum
cells, Langerhans cells, and activated lymphocytes is a new member of
the Ig superfamily. J. Immunol., 149: 735-742, 1992.
20. Fearnley, D. B., McLellan, A. D., Mannering, S. I., Hock, B. D., and
Hart, D. N. J. Isolation of human blood dendritic cells using the CMRF-44
monocbonal antibody: implications for studies on antigen presenting cell
function and immunotherapy. Blood, 89: 3708-3726, 1997.
21. Zhou, L. J., and Tedder, T. F. Human blood dendritic cells selec-tively express CD83, a member of the immunoglobulin superfamily.
J. Immunol., 154: 3821-3835, 1995.
22. Zhou, L., and Tedder, T. F. CDl4� blood monocytes can differen-tiate into functionally mature CD83� dendritic cells. Proc. Natl. Acad.
Sci. USA, 93: 2588-2592, 1996.
23. Nestle, F. 0., Zheng, X., Thompson, C. B.. Turka, L. A., and
Nickoboff, B. J. Characterization ofdermal dendritic cells obtained from
normal human skin reveals phenotypic and functionally distinctive
subsets. J. Immunol., 151: 6535-6545, 1993.
24. Guinan, E. C., Gribben, J. G., Boussiotis, V. A., Freeman, G. J., andNadler, L. M. Pivotal role of the B7:CD28 pathway in transplantation
tolerance and tumor immunity. Blood, 84: 3261-3282, 1994.
25. Mosiabos, G., Birkenbach, M., Ayehunie, S., Matsumura, F.,
Pinkus, G. S., Kieff, E., and Langhoff, E. Circulating human dendritic
cells differentially express high levels of a 55-kd actin-bundling protein.
Am. J. Pathol., 148: 593-600, 1996.
26. Armitage, R. J., Macduff, B. M., Ulrich, D. T., Zappone. J.. Otten,
C., and Fansbow, W. C. Evidence for a functional role of CD83 in T- and
B- cell responses. Tissue Antigens, 48: 453, 1996.
27. Matsuno, K., Ezaki, T., Kudo, S., and Uehara, Y. A life stage ofparticle-laden rat dendritic cells in vivo: their terminal division, activephagocytosis and translocation from the liver to the draining lymph. J.
Exp. Med., 183: 1865-1878, 1996.
28. Egner, W., and Hart, D. N. J. The phenotype of freshly isolated and
cultured human bone marrow albostimulatory cells: heterogeneity in
dendritic cell populations. Immunology, 85: 61 1-620, 1995.
29. McLellan, A. D., Starling, G. C., Williams, L. A., Hock, B. D., andHart. D. N. J. Activation of human peripheral blood dendritic cellsinduces the CD86 costimulatory molecule. Eur. J. Immunol., 25: 2064-
2068, 1995.
30. Inaba, K., Schuler, G., Witmer, M. D., Valinsky, J., Atassi, B., and
Steinman, R. M. Immunologic properties of purified epidermal Lange-
rhans cells. J. Exp. Med., 164: 605-613, 1986.
31. Summers, K., O’Donnell, J., Williams, L. A., and Hart, D. N. J.Expression and function of CD8O and CD86 costimulator molecules on
synovial dendritic cells in chronic arthritic disease. Arthritis Rheum.,
39: 1287-1291, 1996.
32. Mitra, R. S., Judge, T. A., Nestle, F. 0., Turka, L. A., and Nickoloff,
B. J. Psoriatic skin-derived dendritic cell function is inhibited by exog-
enous IL-b. J. Immunol., 154: 2668-2677, 1995.
33. Lipscomb, M. F., Pollard, A. M., and Yates, J. L. A role for TGF-�3
in the suppression by murine bronchoalveolar cells of lung dendritic cell
initiated immune responses. Reg. Immunol., 5: 151-157, 1993.
34. Knoefel, B., Nuske, K., Steiner, T., Junker, K., Kosmehl, H.,
Rebstock, K., Heinhold, D., and Junker, U. Renal cell carcinomasproduce IL-6, 1-10, IL-l 1, and TGF-�3l in primary cultures and modu-late T lymphocyte blast information. J. Interferon Cytokine Res., 17:
95-102, 1997,.
35. Gabribovich, D. I., Chen, H. L., Girgis, K. R., Cunningham. H. T.,
Meny, G. M., Nadaf, S., Kavanaugh, D., and Carbone, D. P. Production
of vascular endothelial growth factor by human tumors inhibits thefunctional maturation ofdendntic cells. Nat. Med., 2: 1096-1 103, 1996.
36. Austyn, J. M. New insights into the mobilization and phagocytic
activity of dendritic cells. J. Exp. Med., 183: 1287-1292, 1996.
American Association for Cancer Research Copyright © 1998 on July 17, 2011clincancerres.aacrjournals.orgDownloaded from




















