
Vasoactive intestinal peptide generates CD4+CD25+ regulatory T cells in vivo
12
Vasoactive intestinal peptide generates CD4 CD25 regulatory T cells in vivo Mario Delgado,* ,† Alejo Chorny, † Elena Gonzalez-Rey, † and Doina Ganea* ,1 *Department of Biological Sciences, Rutgers University, Newark, New Jersey; and † Instituto de Parasitologia y Biomedicina, Instituto del Consejo Superior de Investigaciones Cientı ´ficas, Granada, Spain Abstract: CD4 CD25 regulatory T (Treg) cells control the immune response to a variety of anti- gens, including self-antigens, and several models support the idea of the peripheral expansion of CD4 CD25 Treg cells. Although hormones such as estrogen and -melanocyte-stimulating hor- mone have been recently reported to expand the CD4 CD25 Foxp3-expressing Treg cell com- partment, little is known about the endogenous factors and mechanisms controlling the peripheral expansion of CD4 CD25 Treg cells. In this study, we report on the capacity of the vasoactive intestinal peptide (VIP), an immunosuppressive neuropeptide, to induce functional Treg cells in vivo. The administration of VIP together with spe- cific antigen to T cell receptor (TCR)-transgenic (Tg) mice results in the expansion of the CD4 CD25 , Foxp-3/neuropilin 1-expressing T cells, which inhibit responder T cell proliferation through direct cellular contact. In addition to the increase in the number of CD4 CD25 Treg cells, VIP induces more efficient suppressors on a per- cell basis. The VIP-generated CD4 CD25 Treg cells transfer suppression, inhibit delayed-type hy- persensitivity in TCR-Tg hosts, and prevent graft- versus-host disease in irradiated hosts reconsti- tuted with allogeneic bone marrow. J. Leukoc. Biol. 78: 1327–1338; 2005. Key Words: neuropeptides tolerance collagen-induced arthritis graft-versus-host disease INTRODUCTION The immune system mounts a protective response against pathogen-derived antigens and maintains tolerance for self- antigens. Various mechanisms, including T cell apoptosis, anergy, changes in the balance of effector T cells, and the generation and activation of regulatory T (Treg) cells, contrib- ute to the control of ongoing immune responses and tolerance. Two major types of Treg cells have been characterized in the CD4 population, i.e., the naturally occurring, thymus-gener- ated Treg cells and the peripherally induced, interleukin (IL)-10 or transforming growth factor- (TGF-)-secreting Treg cells [1–3]. The CD4 CD25 Foxp3-expressing, naturally oc- curring Treg cells generated in the thymus migrate and are maintained in the periphery. The signals for their thymic generation and maintenance in the periphery are not defined entirely, although CD28 stimulation and IL-2 appear to be required [4, 5]. Several experimental models support the idea of peripheral generation of CD4 CD25 Treg cells from CD4 CD25 – T cells [6]. However, in contrast to the Tr1/T helper cell type 3 (Th3; Tr2) Treg cells induced by IL-10, TGF-, or through infectious tolerance [7–9], the CD4 CD25 Treg cells developing de novo in the periphery exert their suppressive function in a cell contact-dependent manner [10]. The endogenous factors and mechanisms controlling the pe- ripheral expansion of CD4 CD25 Treg cells are mostly un- known. Recently, Polanczyk et al. [11] reported that estrogen promotes tolerance by expanding the CD4 CD25 Foxp3 - expressing regulatory compartment. Vasoactive intestinal peptide (VIP) and pituitary adenylate cyclase-activating polypeptide (PACAP) are two neuropeptides with profound immunoregulatory activities. We have shown previously that VIP/PACAP inhibit macrophage/dendritic cell/ microglia activation and promote Th2-type responses at the expense of proinflammatory Th1 immunity [12–14]. As Treg cells are major players in limiting the immune response, we investigated the possibility that VIP and PACAP promote Treg cell development and/or activation. In this study, we charac- terized the functional Treg cells generated in vivo following VIP administration. The major Treg cell population induced in vivo following VIP and antigen administration is CD4 CD25 , Foxp-3 hi , neuropilin (Nrp) hi and inhibits responder T cell pro- liferation through direct cellular contact. These cells exhibit increased suppressive activity compared with Treg cells gen- erated in response to antigen alone. The in vivo VIP-generated Treg cells transfer suppression, inhibit delayed-type hypersen- sitivity (DTH) in T cell receptor (TCR)-transgenic (Tg) hosts and prevent graft-versus-host disease (GVHD) in irradiated hosts reconstituted with allogeneic bone marrow (BM). MATERIALS AND METHODS Reagents Synthetic VIP was purchased from Calbiochem-Novabiochem (Laufelfingen, Switzerland). Capture and biotinylated antibodies against murine IL-4, IL-5, 1 Correspondence: Rutgers University, Department of Biological Sciences, 101 Warren St., Newark, NJ 07102. E-mail: [email protected] Received June 3, 2005; revised July 29, 2005; accepted August 1, 2005; doi: 10.1189/jlb.0605299. 0741-5400/05/0078-1327 © Society for Leukocyte Biology Journal of Leukocyte Biology Volume 78, December 2005 1327
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Vasoactive intestinal peptide generates CD4+CD25+ regulatory T cells in vivo

Vasoactive intestinal peptide generates CD4�CD25�
regulatory T cells in vivoMario Delgado,*,† Alejo Chorny,† Elena Gonzalez-Rey,† and Doina Ganea*,1
*Department of Biological Sciences, Rutgers University, Newark, New Jersey; and †Instituto de Parasitologia yBiomedicina, Instituto del Consejo Superior de Investigaciones Cientı́ficas, Granada, Spain
Abstract: CD4�CD25� regulatory T (Treg) cellscontrol the immune response to a variety of anti-gens, including self-antigens, and several modelssupport the idea of the peripheral expansion ofCD4�CD25� Treg cells. Although hormones suchas estrogen and �-melanocyte-stimulating hor-mone have been recently reported to expand theCD4�CD25� Foxp3-expressing Treg cell com-partment, little is known about the endogenousfactors and mechanisms controlling the peripheralexpansion of CD4�CD25� Treg cells. In thisstudy, we report on the capacity of the vasoactiveintestinal peptide (VIP), an immunosuppressiveneuropeptide, to induce functional Treg cells invivo. The administration of VIP together with spe-cific antigen to T cell receptor (TCR)-transgenic(Tg) mice results in the expansion of theCD4�CD25�, Foxp-3/neuropilin 1-expressing Tcells, which inhibit responder T cell proliferationthrough direct cellular contact. In addition to theincrease in the number of CD4�CD25� Treg cells,VIP induces more efficient suppressors on a per-cell basis. The VIP-generated CD4�CD25� Tregcells transfer suppression, inhibit delayed-type hy-persensitivity in TCR-Tg hosts, and prevent graft-versus-host disease in irradiated hosts reconsti-tuted with allogeneic bone marrow. J. Leukoc.Biol. 78: 1327–1338; 2005.
Key Words: neuropeptides � tolerance � collagen-induced arthritis� graft-versus-host disease
INTRODUCTION
The immune system mounts a protective response againstpathogen-derived antigens and maintains tolerance for self-antigens. Various mechanisms, including T cell apoptosis,anergy, changes in the balance of effector T cells, and thegeneration and activation of regulatory T (Treg) cells, contrib-ute to the control of ongoing immune responses and tolerance.
Two major types of Treg cells have been characterized in theCD4� population, i.e., the naturally occurring, thymus-gener-ated Treg cells and the peripherally induced, interleukin(IL)-10 or transforming growth factor-� (TGF-�)-secreting Tregcells [1–3]. The CD4�CD25� Foxp3-expressing, naturally oc-curring Treg cells generated in the thymus migrate and are
maintained in the periphery. The signals for their thymicgeneration and maintenance in the periphery are not definedentirely, although CD28 stimulation and IL-2 appear to berequired [4, 5]. Several experimental models support the ideaof peripheral generation of CD4�CD25� Treg cells fromCD4�CD25– T cells [6]. However, in contrast to the Tr1/Thelper cell type 3 (Th3; Tr2) Treg cells induced by IL-10,TGF-�, or through infectious tolerance [7–9], the CD4�CD25�
Treg cells developing de novo in the periphery exert theirsuppressive function in a cell contact-dependent manner [10].The endogenous factors and mechanisms controlling the pe-ripheral expansion of CD4�CD25� Treg cells are mostly un-known. Recently, Polanczyk et al. [11] reported that estrogenpromotes tolerance by expanding the CD4�CD25� Foxp3�-expressing regulatory compartment.
Vasoactive intestinal peptide (VIP) and pituitary adenylatecyclase-activating polypeptide (PACAP) are two neuropeptideswith profound immunoregulatory activities. We have shownpreviously that VIP/PACAP inhibit macrophage/dendritic cell/microglia activation and promote Th2-type responses at theexpense of proinflammatory Th1 immunity [12–14]. As Tregcells are major players in limiting the immune response, weinvestigated the possibility that VIP and PACAP promote Tregcell development and/or activation. In this study, we charac-terized the functional Treg cells generated in vivo followingVIP administration. The major Treg cell population induced invivo following VIP and antigen administration is CD4�CD25�,Foxp-3hi, neuropilin (Nrp)hi and inhibits responder T cell pro-liferation through direct cellular contact. These cells exhibitincreased suppressive activity compared with Treg cells gen-erated in response to antigen alone. The in vivo VIP-generatedTreg cells transfer suppression, inhibit delayed-type hypersen-sitivity (DTH) in T cell receptor (TCR)-transgenic (Tg) hostsand prevent graft-versus-host disease (GVHD) in irradiatedhosts reconstituted with allogeneic bone marrow (BM).
MATERIALS AND METHODS
ReagentsSynthetic VIP was purchased from Calbiochem-Novabiochem (Laufelfingen,Switzerland). Capture and biotinylated antibodies against murine IL-4, IL-5,
1 Correspondence: Rutgers University, Department of Biological Sciences,101 Warren St., Newark, NJ 07102. E-mail: [email protected]
Received June 3, 2005; revised July 29, 2005; accepted August 1, 2005;doi: 10.1189/jlb.0605299.
0741-5400/05/0078-1327 © Society for Leukocyte Biology Journal of Leukocyte Biology Volume 78, December 2005 1327

IL-2, TGF-�1, and interferon-� (IFN-�); monoclonal antibodies (mAb) tocytotoxic T-lymphocyte antigen (CTLA)-4, CD4, V�3 (clone KJ25), I-Ek, I-Ed
[major histocompatibility complex type II (MHC-II)], CD25, CD69, B220,CD103, CD62 ligand (CD62L), and CD45RB; and recombinant murine(rm)IFN-� and rmIL-4 were purchased from BD PharMingen (San Diego, CA).Capture and biotinylated antibodies against murine IL-10 and rmIL-2 werepurchased from PreproTech Inc. (Rocky Hill, NJ). Pigeon cytochrome cfragment (PCCF) was synthesized and purified by Research Genetics (Hunts-ville, AL). Ovalbumin (OVA) and avidin-peroxidase were purchased fromSigma Chemical Co. (St. Louis, MO). Rat anti-glucocorticoid-induced tumornecrosis factor receptor (GITR) antibody was purchased from R&D Systems(Minneapolis, MN).
Animals
B10.A (I-Ek), C57Bl/6 (H-2b), BALB/c (H-2d), and TCR-Cyt-5CC7-I/Rag1 Tg(I-Ek) mice were obtained from Jackson Laboratory (Bar Harbor, ME) andTaconic Farms (Germantown, NY). All mice used were between 7 and 12weeks of age.
Cell isolation and cultures
Purified, naı̈ve CD4 T cells from Tg mice were isolated by positive immuno-magnetic selection by treating spleen cells with magnetic beads conjugatedwith anti-CD4 mAb [magnetic cell sorter (MACS), Miltenyi Biotec, Auburn,CA]. The purified T cells were �98% CD4� by fluorescein-activated cellsorter (FACS) analysis.
Antigen-presenting cells (APC) were prepared by T cell depletion of B10.A(I-Ek) spleen cells by treating the cells with a mixture of magnetic beadsconjugated with anti-CD8 and anti-CD4 mAb (MACS) at 15°C for 20 min,followed by magnetic separation, and treatment with 50 �g/ml mitomycin C(mitC; Sigma Chemical Co.) for 20 min at 37°C.
CD4�CD25� and CD4�CD25– cell populations were isolated with anAutoMACS magnetic cell sorter (Miltenyi Biotec). Briefly, PCCF TCR-Tgspleen and draining lymph node (LN; popliteal, inguinal, and mesenteric LN)cells were depleted of red blood cells, of adherent cells, and CD8� T cells byincubation for 2 h at 37°C in Petri dishes followed by negative magneticselection with anti-CD8 microbeads (Miltenyi Biotec). Resulting cells werelabeled with biotinylated anti-CD25 mAb for 45 min at 4°C, washed, andincubated with a 1:10 dilution of streptavidin-conjugated magnetic beads(Miltenyi Biotec) at a concentration of 108 cells/ml in phosphate-bufferedsaline (PBS) containing 0.5% bovine serum albumin (BSA) and 1 mM EDTAfor 20 min at 8°C. The cells were washed once, passed through a nytex filterto remove clumps, pelleted, and resuspended at a concentration of 2 � 108
cells/ml. CD25� T cells were isolated with the AutoMACS, and CD4� T cellswere isolated from the CD25– fraction with anti-CD4 microbeads (MiltenyiBiotec). Contaminating MHC-II� cells were removed from the CD25� andCD4�CD25– fractions by panning with anti-I-Ek-coated plates for 12 h at37°C. The CD25� populations were �95% CD4�CD25�, and theCD4�CD25– populations were 98% pure, as determined by flow cytometry.
In some of the in vivo experiments, the cells were labeled with 1 mlPBS-0.1% BSA-5 �M carboxyfluorescein succinimidyl ester (CFSE) for 10min at 37°C, washed three times in RPMI-10% fetal bovine serum, andinjected intravenously (i.v.; 100 �l) as described below.
To determine T cell anergy, isolated CD4 T cells (5�104 cells/well) werestimulated with mitC-treated B10.A spleen APC (105 cells/well) and varyingconcentrations of PCCF. Proliferation was determined following pulsing withbromodeoxyuridine (BrdU) for the last 16 h of a 4-day culture period. BrdUincorporation was measured by using a peroxidase-labeled anti-BrdU mAbdetected with tetramethylenbenzidine (TMB) substrate and quantified at opti-cal density (OD)450 in an enzyme-linked immunosorbent assay (ELISA) reader,according to the manufacturer’s (Roche Applied Science, Mannheim, Ger-many) protocol. Results are expressed as arbitrary units of OD450. To deter-mine the in vitro Treg cell function, different numbers of CD4�, CD4�CD25�,or CD4�CD25– T cells isolated from naı̈ve mice or mice inoculated with PCCFor PCCF plus VIP were cocultured with responder CD4� T cells (rCD4; 5�104
cells/well), isolated from naı̈ve PCCF-Tg mice and stimulated in the presenceof mitC-treated B10.A spleen APC (105 cells/well) and PCCF (5 �M). Prolif-eration was determined as described above. Culture supernatants were col-lected and assayed for cytokine determination as described below. In someexperiments, cocultures were performed in the presence of blocking mouse
anti-IL-10 (10 �g/ml), anti-TGF-�1 (40 �g/ml), and/or anti-CTLA-4 (10�g/ml) mAb or of rmIL-2 (100 U/ml). Transwell experiments were done in24-well plates (Millicell, 0.4 �m, Millipore, Bedford, MA). Responder PCCF-specific-Tg CD4 T cells (5�105 cells), together with mitC-treated B10.Aspleen APC (2�104 cells) and PCCF (5 �M), were placed in the bottom wells.CD4� T cells or CD4�CD25� T cells (5�105 cells) isolated from PCCF- orPCCF � VIP-treated Tg mice were placed together with mitC-treated B10.Aspleen APC (2�104 cells) and PCCF (5 �M) in the upper wells. Three dayslater, the basket was removed, and the proliferation of the responder T cellswas measured.
FACS analysis
Spleen and LN (popliteal, inguinal, and mesenteric LN) cells (106 cells/ml)were harvested in ice-cold RPMI complete medium and washed twice with PBScontaining 0.1% sodium azide plus 2% heat-inactivated fetal calf serum (washbuffer). Cells were incubated with various mAb [phycoerythrin (PE)-anti-CD25, fluorescein isothiocyanate (FITC)-anti-CD62L, FITC-anti-CD69, FITC-anti-CD103, FITC-anti-V�3, FITC-anti-CD45RB, FITC-anti-GITR, peridininchlorophyll protein (PerCP)-anti-CD4, 2.5 �g/ml final concentration] at 4°C for1 h. Isotype-matched antibodies were used as controls, and immunogloublin G(IgG) block (Sigma Chemical Co.) was used to block the nonspecific binding toFc receptors. After extensive washing, the cells were fixed in 1% paraformal-dehyde. Stained T cells, gated accordingly to CD4 staining, were analyzedusing a FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA). Sam-ples with isotype-matched antibodies instead of specific antibodies were usedto determine the proper region or window setting. Fluorescence data wereexpressed as mean channel fluorescence and as percentage of positive cellsafter subtraction of background, isotype-matched values.
For analysis of intracellular CTLA-4, spleen and LN T cells (106 cells/ml)were harvested in ice-cold RPMI complete medium, washed twice with washbuffer, and stained with PerCP-anti-CD4 and FITC-anti-CD25 mAb for 30 minat 4°C, followed by fixation with Cytofix/Cytoperm solution (BD PharMingen)and incubation for 45 min at 4°C with PE-anti-CTLA-4 mAb diluted in PBS �1% BSA � 0.5% saponin. After extensive washing, cells were analyzed usinga FACSCalibur flow cytometer.
Cytokine assays
Cytokine contents in the supernatants of T cell cultures or APC-CD4� T cellcocultures were determined by specific sandwich ELISAs. The cytokineamount in each supernatant was extrapolated from the standard curve. Theantibody pairs used were as follows, listed by capture/biotinylated detectionantibodies (BD PharMingen): IL-4, BVD4-1D11/BVD6-24G2; IFN-�, R4-6A2/XMG1.2; IL-5, TRFK5/TRFK4; IL-2, JES6-1A12/JES6-5H4.
Determination of antibody responses
Specific antibody responses in the PCCF or OVA-immunized mice weredetermined by ELISA. Serum was obtained by cardiac puncture. Maxisorbplates (Millipore) were coated overnight at 4°C with 100 �l soluble PCCF orOVA (10 �g/ml). After washing with PBS containing 0.05% v/v Tween-20, theplates were blocked with 3% BSA and incubated with serial dilutions of serumfor 2 h at 37°C. After washing, biotinylated anti-IgG, anti-IgG1, or anti-IgG2a(2.5 �g/ml; Serotec, Oxford, UK) was added for 1 h at 37°C. The plates werewashed, followed by incubation with streptavidin-horseradish peroxidase. Thebound enzyme was detected with the TMB substrate and quantified at OD450
in an ELISA reader (Bio-Tek Instrument, Inc., Winooski, VT). Standard curveswere constructed for each assay by coating wells with an isotype-specificanti-mouse Ig, followed by addition of known concentrations of the mouse Igisotype.
Real-time reverse transcriptase-polymerasechain reaction (RT-PCR)
Total RNA was isolated from CD4� T cells or sorted CD4�CD25� (106 cells)using the Ultraspec RNA reagent (Biotecx, Houston, TX). Total RNA (2 �g)was reverse-transcribed with oligo-dT primers and Moloney murine leukemiavirus-RT polymerase (Invitrogen, Carlsbad, CA). Quantitative real-time RT-PCR was performed in an ABI PRISM cycler (Applied Biosystems, Foster City,CA) using a SYBR Green PCR kit from Applied Biosystems. A threshold wasset in the linear part of the amplification curve, and the number of cycles
1328 Journal of Leukocyte Biology Volume 78, December 2005 http://www.jleukbio.org

needed to reach the threshold was calculated for each gene. Relative mRNAlevels were determined by using standard curves for each individual gene andfurther normalization to hypoxanthine guanine phosphoribosyl transferase(HPRT). Melting curves established the purity of the amplified band. Primersequences are: Nrp1 (5�-GCCTGCTTTCTTCTCTTGGTTTCA-3�, 5�-GCTCT-GGGCACTGGGCTACA-3�); Foxp3 (5�-CTGGCGAAGGGCTCGGTAGTCCT-3�, 5�-CTCCCAGAGCCCATGGCAGAAGT-3�); HPRT (5�-TGGAAAGAAT-GTCTTGATTGTTGAA-3�, 5�-AGCTTGCAACCTTAACCATTTTG-3�).
Acute GVHD and graft-versus-leukemia (GVL)
GVHD
BALB/c mice (H-2d) were irradiated lethally (8Gy TBI with a 200-Kv X-raysource) and reconstituted i.v. with 100 �l T cell-depleted (TCD), allogeneicBM cells (5�106 cells) from B10.A mice (H-2k) plus CD4 T cells (106 cells)and CD4�CD25– or CD4�CD25� cells (0.5�106 cells), isolated from naı̈vePCCF-Tg mice or from mice immunized with PCCF or PCCF plus VIP asdescribed above. The survival and appearance of the BALB/c hosts weremonitored daily, and body weight was measured weekly.
GVL
Irradiated BALB/c mice were reconstituted with TCD allogeneic BM cells(B10.A, 106 cells). The recipients also received a single i.v. injection of CD4T cells (106 cells) isolated from PCCF-Tg mice immunized with PCCF or PCCFplus VIP and A20 leukemic cells (derived from BALB/c mice, 104 cells). Therecipients were monitored once every day from the day of transplantation untilthey succumbed naturally to GVHD and/or tumor burden. Tumor growth/elimination was assessed by the presence of A20 cells in blood and detectedbased on coexpression of B220 and H-2Kd and size by flow cytometry.
Data analysis
All values are expressed as mean � SD of mice/experiment. The differencesbetween groups were analyzed by Mann-Whitney U test and if appropriate, byKruskal-Wallis ANOVA test.
RESULTS
In vivo administration of VIP increases thenumber of CD4�CD25� T cells
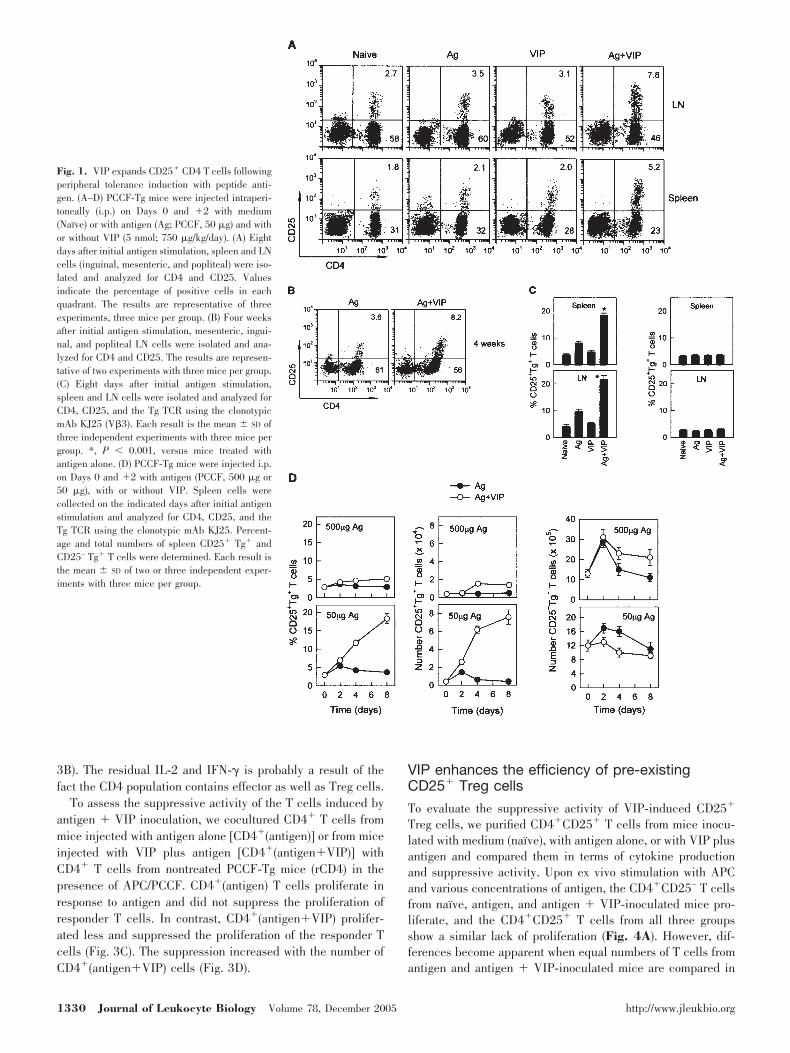
Previous reports from our laboratory indicated that in thepresence of immunogenic doses of antigen, VIP promotes Th2-type immune responses in vivo and in vitro. In vivo inoculationof VIP into PCCF-TCR-Tg mice results in long-term survival ofTh2 effectors and predominant development of Th2 memorycells. As antigen-specific tolerance can be induced with lowdoses of antigen, we assessed the ability of VIP to facilitateCD4�CD25� Treg cell induction in vivo in the presence of lowdoses of antigen. PCCF-TCR-Tg mice were inoculated with lowdoses of antigen (50 �g PCCF) on Days 0 and �2, with orwithout VIP (5 nmol, 750 �g/kg/day). Eight days later, FACSdetermined the numbers of CD4�CD25� T cells from spleenand mesenteric LN. Administration of antigen plus VIP resultsin the doubling of CD4�CD25� cells in spleen and LN (Fig.1A). Similar numbers were observed 4 weeks after immuniza-tion, indicating a long-term effect of VIP on the expansion ofthe CD4�CD25� T cell population (Fig. 1B). The VIP-inducedincrease was observed only in the antigen-specific T cellpopulation, as determined by triple staining for CD4, CD25,and the TCR/V�3 (Fig. 1C). The VIP-induced increase inCD25�V�3� T cells was observed only in the presence of lowdoses of antigen (Fig. 1D, left and middle panels), and VIP didnot increase the number of CD25–V�3� T cells, which pre-
sumably represent effector T cells whose CD25 expression istransient (Fig. 1D, right panel).
High expression of Foxp3 and more recently, of Nrp1characterizes CD4�CD25� Treg cells. We determinedwhether VIP induces or contributes to the levels of Foxp3and Nrp1 expression in CD4� T cells. PCCF-Tg mice wereinoculated with medium (naı̈ve), PCCF (antigen), or PCCFplus VIP (antigen�VIP), and splenic CD4� as well aspurified CD4�CD25– and CD4�CD25� T cells were sub-jected to real-time RT-PCR for Foxp3 and Nrp1 expression.CD4� T cells express low levels of Foxp3 and Nrp1, andPCCF administration slightly increased Foxp3 and Nrp1expression. In contrast, Foxp3 and Nrp1 were inducedstrongly by PCCF � VIP inoculation (Fig. 2, left panels).PCCF � VIP administration also increased the levels ofexpression for surface GITR, CD103, CD62L, and CD69 anddecreased surface CD45RB in the CD4� T cell population(Fig. 2, right panel). These changes could be a result of aVIP-induced increase in the number of CD4�CD25� Tregcells or to changes in levels of expression per cell. Todistinguish between these possibilities, we determined theeffect of VIP on separated CD4�CD25– and CD4�CD25� Tcells. Foxp3 and Nrp1 were expressed at high levels inCD4�CD25� T cells but not in the CD4�CD25– population.VIP did not affect Foxp3/Nrp1 expression in either popula-tion (Fig. 2, left panels). CD4�CD25� T cells expresshigher levels of GITR and CD103 and lower levels ofCD45RB than the CD4�CD25– population. VIP administra-tion did not affect the levels of expression in either popu-lation (Fig. 2, right panel). Taken together, these resultssuggest that the VIP effect is mediated through an increasein the number of CD4�CD25� Treg cells.
In vivo administration of VIP induces CD4�
T cells that exhibit low proliferative capacity,secrete IL-10 and TGF-�, and suppress theproliferation of responder T cells
All Treg cell subsets are characterized by low in vitro prolif-eration in response to antigen. We investigated the prolifera-tive capacity of splenic CD4� T cells harvested 8 days afterantigen � VIP inoculation. The CD4� T cells were restimu-lated ex vivo with mitC-treated APC and various concentra-tions of PCCF. The T cells obtained from animals injected withantigen � VIP proliferate less than those obtained from ani-mals injected with antigen alone or from controls injected withmedium (naı̈ve; Fig. 3A).
Although all Treg cell subsets are poor producers of IL-2and IFN-�, the Tr1 cell subset secretes predominantly IL-10with or without TGF-�, and the Tr2 (Th3) cell subset ischaracterized by TGF-� and IL-4 production. We analyzed thecytokine profile of CD4� T cells from antigen � VIP-inocu-lated mice following ex vivo restimulation. T cells from animalsinjected with antigen alone had a profile similar to controls(naı̈ve), i.e., high levels of IL-2 and IFN-� and low levels ofIL-4, IL-10, and TGF-�. In contrast, T cells from antigen �VIP-inoculated mice produce less IL-2 and IFN-�, slightlymore IL-4, and significantly higher levels of IL-10 and TGF-�than T cells from controls (naı̈ve) or antigen-injected mice (Fig.
Delgado et al. VIP induces Treg cells in vivo 1329
3B). The residual IL-2 and IFN-� is probably a result of thefact the CD4 population contains effector as well as Treg cells.
To assess the suppressive activity of the T cells induced byantigen � VIP inoculation, we cocultured CD4� T cells frommice injected with antigen alone [CD4�(antigen)] or from miceinjected with VIP plus antigen [CD4�(antigen�VIP)] withCD4� T cells from nontreated PCCF-Tg mice (rCD4) in thepresence of APC/PCCF. CD4�(antigen) T cells proliferate inresponse to antigen and did not suppress the proliferation ofresponder T cells. In contrast, CD4�(antigen�VIP) prolifer-ated less and suppressed the proliferation of the responder Tcells (Fig. 3C). The suppression increased with the number ofCD4�(antigen�VIP) cells (Fig. 3D).
VIP enhances the efficiency of pre-existingCD25� Treg cells
To evaluate the suppressive activity of VIP-induced CD25�
Treg cells, we purified CD4�CD25� T cells from mice inocu-lated with medium (naı̈ve), with antigen alone, or with VIP plusantigen and compared them in terms of cytokine productionand suppressive activity. Upon ex vivo stimulation with APCand various concentrations of antigen, the CD4�CD25– T cellsfrom naı̈ve, antigen, and antigen � VIP-inoculated mice pro-liferate, and the CD4�CD25� T cells from all three groupsshow a similar lack of proliferation (Fig. 4A). However, dif-ferences become apparent when equal numbers of T cells fromantigen and antigen � VIP-inoculated mice are compared in
Fig. 1. VIP expands CD25� CD4 T cells followingperipheral tolerance induction with peptide anti-gen. (A–D) PCCF-Tg mice were injected intraperi-toneally (i.p.) on Days 0 and �2 with medium(Naı̈ve) or with antigen (Ag; PCCF, 50 �g) and withor without VIP (5 nmol; 750 �g/kg/day). (A) Eightdays after initial antigen stimulation, spleen and LNcells (inguinal, mesenteric, and popliteal) were iso-lated and analyzed for CD4 and CD25. Valuesindicate the percentage of positive cells in eachquadrant. The results are representative of threeexperiments, three mice per group. (B) Four weeksafter initial antigen stimulation, mesenteric, ingui-nal, and popliteal LN cells were isolated and ana-lyzed for CD4 and CD25. The results are represen-tative of two experiments with three mice per group.(C) Eight days after initial antigen stimulation,spleen and LN cells were isolated and analyzed forCD4, CD25, and the Tg TCR using the clonotypicmAb KJ25 (V�3). Each result is the mean � SD ofthree independent experiments with three mice pergroup. *, P 0.001, versus mice treated withantigen alone. (D) PCCF-Tg mice were injected i.p.on Days 0 and �2 with antigen (PCCF, 500 �g or50 �g), with or without VIP. Spleen cells werecollected on the indicated days after initial antigenstimulation and analyzed for CD4, CD25, and theTg TCR using the clonotypic mAb KJ25. Percent-age and total numbers of spleen CD25� Tg� andCD25– Tg� T cells were determined. Each result isthe mean � SD of two or three independent exper-iments with three mice per group.
1330 Journal of Leukocyte Biology Volume 78, December 2005 http://www.jleukbio.org

terms of cytokine production. The CD4�CD25–(antigen�VIP)T cells secrete less IFN-� and more IL-10 and TGF-� than theCD4�CD25– (antigen) or the CD4�CD25– (naı̈ve; Fig. 4B),which probably reflects the presence of developing Tr1/Th3-likeTreg cells within the CD25– population. The difference is evengreater within the three CD4�CD25� populations. AlthoughCD4�CD25�(antigen�VIP), CD4�CD25�(antigen), and CD4�-CD25�(naı̈ve) do not secrete IL-2, the CD4�CD25�-(antigen�VIP) cells produce much less IFN-�, more IL-10, andmuch more TGF-� than the CD4�CD25�(antigen) or theCD4�CD25�(naı̈ve) populations (Fig. 4B).
In addition, we compared the suppressive activity ofCD4�CD25�(naı̈ve),CD4�CD25�(antigen),andCD4�CD25�-(antigen�VIP) T cells by coculturing these cells with re-sponder T cells at different ratios in the presence of APC/antigen. Although at high ratios (one-half and one-fourth ofTreg/Tresp), CD4�CD25�(naı̈ve), CD4�CD25�(antigen), andCD4�CD25�(antigen�VIP) appear to be equally effective insuppressing the proliferation of responder T cells, at lowerratios, the CD4�CD25�(antigen�VIP) proved significantlymore efficient (Fig. 4C). This indicates that on a per-cell basis,the Treg cells generated in the presence of VIP are moreefficient. Also, whereas CD4�CD25– (antigen) or CD4�CD25–
(naı̈ve) did not have suppressive activity, the CD4�CD25–
(antigen�VIP) exhibited some suppressive activity at highratios (Fig. 4C, bottom panel), suggesting again the presence ofdeveloping Treg cells within the CD25– population.
It has been reported previously that Treg cells need TCRactivation to function as suppressors. To assess the require-ment for TCR activation in the CD4�CD25�(antigen�VIP)population, we isolated CD4�CD25�(antigen�VIP) fromPCCF-immunized mice and cocultured them with CD4�CD25–
T cells from OVA-immunized MHC II-compatible mice in thepresence of OVA, OVA plus PCCF, or anti-CD3. Suppressionoccurred only in the cultures containing PCCF or anti-CD3(Fig. 4D), indicating that the CD4�CD25�(antigen�VIP) Tregcells need to be activated through the TCR to function assuppressors.
The in vivo VIP-induced Treg cells act primarilythrough cell-cell contact
Naturally occurring CD4�CD25� Treg cells exert their sup-pressive activity primarily through direct cellular contact,whereas Tr1/Tr2(Th3) suppressors act primarily through cyto-kines. To evaluate the requirement for direct cellular contact inthe suppressive activity of CD4�CD25�(antigen�VIP), wegenerated CD4�(antigen�VIP; containing CD25� and CD25–
T cells) and purified CD4�CD25�(antigen�VIP) and cocul-tured them with rCD4� T cells in the presence of APC/PCCF.For the cultures containing CD4�(antigen�VIP), addition ofsaturating amounts of anti-IL-10 or anti-TGF-�-neutralizingantibodies slightly reversed the inhibitory effect on responderT cell proliferation, and a combination of anti-IL-10, anti-TGF-�, and anti-CTLA-4 antibodies completely reversed
Fig. 2. VIP induces the in vivo emergence of CD4� T cells with a regulatory phenotype. PCCF-Tg mice were injected i.p. with medium (Naı̈ve) and antigen(PCCF), with or without VIP. Eight days after initial antigen stimulation, spleen CD4 and CD4�CD25� and CD4�CD25– T cells were isolated. Cells from naı̈vePCCF-Tg mice were used as control. Foxp3 and Nrp1 mRNA expression was analyzed by real-time RT-PCR and normalized using HPRT expression. GITR, CD103,CD45RB, CD62L, and CD69 expression was determined by flow cytometry. Results are representative of three independent experiments. *, P 0.001, versusmice treated with antigen alone.
Delgado et al. VIP induces Treg cells in vivo 1331

suppression (Fig. 5A, left panel). Also, when CD4�-(antigen�VIP) and responder T cells were separated in Trans-well experiments, there was a slight reduction in proliferation(P0.05 for Transwell vs. rCD4�APC samples; Fig. 5A, leftpanel). This suggests that the CD4�(antigen�VIP) cells con-sist of two Treg cell populations: a major, contact-dependent,cytokine-independent population and a minor population, ex-erting suppression through soluble cytokines. In contrast, thesuppressive activity of the CD4�CD25�(antigen�VIP) wasentirely contact-dependent, with no reversal upon anti-IL-10 oranti-TGF-� administration, complete reversal in the presenceof anti-CTLA-4 antibodies, and no inhibition in the Transwellexperiments (Fig. 5A, right panel). As expected, exogenousIL-2 reversed the suppressive effect for CD4�(antigen�VIP)and CD4�CD25�(antigen�VIP) Treg cells.
As CTLA-4 appears to play an essential role in the suppres-sion exerted by the CD4�CD25�(antigen�VIP) T cells, weassessed CTLA-4 protein expression in spleen cells from naı̈vemice, mice inoculated with PCCF (antigen), and mice inocu-lated with VIP plus PCCF (antigen�VIP). VIP increasedCTLA-4 expression in a significant manner in the CD4�CD25–
and particularly in the CD4�CD25� cells (Fig. 5B).
Transfer of antigen-specific, suppressive activitywith VIP-generated Treg cells
Treg cells retain their suppressive activity upon adoptive trans-fer. To assess the capacity of the in vivo VIP-generated Tregcells, we purified splenic CD4� and CD4�CD25� T cells fromPCCF-Tg mice injected with PCCF (antigen) or with PCCF plusVIP (antigen�VIP). CD4� T cells were injected i.v. intoPCCF-Tg naı̈ve hosts, followed 24 h later by immunization withPCCF/complete Freund’s adjuvant (CFA) in the footpads. Thespleen, draining LN (DLN), and peripheral nondraining LN(PLN) were obtained. The spleen and DLN from mice trans-ferred with CD4�(antigen) were significantly larger thanthe counterparts from animals transferred with CD4�-(antigen�VIP; Fig. 6A). This is presumably because of theTreg cells present in the CD4�(antigen�VIP) population,which upon transfer, suppress the host’s response to PCCF.
We assessed the suppressive activity of the transferredCD4�(antigen�VIP) and CD4�CD25�(antigen�VIP) in amodel of DTH. CD4� and CD4�CD25� T cells from PCCF-injected (antigen) or PCCF � VIP-injected (antigen�VIP)donors were labeled with CFSE and injected i.v. into PCCF-Tgnaı̈ve hosts. A control group was also injected with naı̈ve CD4�
T cells. The mice were immunized 24 h later with PCCF/CFAin the footpads, followed by a challenge 2 weeks later withPCCF in the left ear pinna. PBS instead of PCCF was used ascontrol in the right ear. The DTH reaction was assessed 24 hlater. In addition, we determined the proliferation of spleencells in response to PCCF, and the levels of anti-PCCF anti-
4Fig. 3. VIP induces the in vivo emergence of functional Treg CD4� cells.PCCF-Tg mice were injected i.p. on Days 0 and �2 with medium (Naı̈ve) orantigen (PCCF), with or without VIP. Eight days after initial antigen stimula-tion, spleen CD4� T cells were isolated. Spleen cells from naı̈ve PCCF-Tgmice were used as control. (A) VIP induces CD4� T cell hyporesponsiveness.Isolated CD4� T cells (5�104 cells/well) were stimulated with mitC-treatedB10.A spleen APC (1�105 cells/well) and varying concentrations of PCCF.BrdU was added for the last 16–18 h of a 4-day culture, and T cell proliferationwas assayed as described in Materials and Methods. Each result is the mean �SD of three experiments performed in duplicate. A450, Absorbance at 450 nm.(B) VIP induces IL-10/TGF-�1-producing T cells. Isolated CD4� T cells(2�105 cells/well) were stimulated with mitC-treated B10.A spleen APC(5�105 cells/well) and PCCF. After 48 h culture, cytokine secretion wasdetermined by ELISA. Cells cultured with an irrelevant antigen (OVA, 10�g/ml) did not secrete cytokines. Each result is the mean � SD of threeexperiments performed in duplicate. (C and D) In vivo treatment with VIPinduces functional Treg cells. Different numbers (5�104 cells/well in C) ofCD4� T cells isolated from mice inoculated with PCCF [CD4(Ag)] or PCCFplus VIP [CD4(Ag�VIP)] were cocultured with responder CD4� T cells(rCD4�, 5�104 cells/well), isolated from naı̈ve PCCF-Tg mice and stimulatedin the presence of mitC-treated B10.A spleen APC (105 cells/well) and PCCF(5 �M). Proliferation was determined by using BrdU. Secreted IL-2 wasquantified by ELISA. Each result is the mean � SD of three experimentsperformed in duplicate. *, P 0.001, versus mice treated with antigen alone.
1332 Journal of Leukocyte Biology Volume 78, December 2005 http://www.jleukbio.org

bodies. CD4�(antigen) exhibited little, if any, suppressive ac-tivity, whereas CD4�(antigen�VIP) T cells were suppressive(Fig. 6B). The CD4�(antigen) exhibited a similar lack of sup-pressive activity as the CD4� control (from naı̈ve mice; resultsnot shown). The suppressive effect of the CD4�(antigen�VIP)T cells could be a result of the VIP-induced increase in thenumber of CD25� Treg cells and/or to an increase in theefficiency of the CD25� Treg cell activity. As expected,
CD4�CD25�(antigen) and CD4�CD25�(antigen�VIP) T cellshave a suppressive effect; however, the CD4�CD25�-(antigen�VIP) T cells exert a much more pronounced suppres-sion than the same number of CD4�CD25�(antigen; Fig. 6B),suggesting that VIP increases the efficiency of CD4�CD25�
Treg cells.The presence and proliferative state of the transferred T
cells were ascertained by FACS analysis for CFSE, CD4, and
Fig. 4. VIP induces highly efficient CD4�CD25� Treg cells. PCCF-Tg mice were injected i.p. with medium (Naı̈ve) or with PCCF (Ag), with or without VIP (5nmol; 750 �g/kg). Eight days after initial antigen stimulation, spleen CD4�CD25� and CD4�CD25– T cells were isolated. (A) Isolated CD4�CD25� andCD4�CD25– T cells (5�104 cells/well) were stimulated with mitC-treated B10.A spleen APC (105 cells/well) and varying concentrations of PCCF. Proliferationwas determined by using BrdU. Each result is the mean � SD of three experiments performed in duplicate. (B) Isolated CD4�CD25� and CD4�CD25– T cells(2�105 cells/well) were stimulated with mitC-treated B10.A spleen APC and PCCF. After 48 h culture, cytokine secretion was determined by ELISA. Cells culturedwith an irrelevant antigen (OVA, 10 �g/ml) did not secrete cytokines. Each result is the mean � SD of three experiments performed in duplicate. (C) rCD4� Tcells (5�104 cells/well) isolated from naı̈ve PCCF-Tg mice were cocultured with different numbers of CD4�CD25� and CD4�CD25– T cells (ratios of Treg/Trespfrom 1:2 to 1:10) in the presence of mitC-treated B10.A spleen APC plus PCCF or soluble anti-CD3 antibody (5 �g/ml). Proliferation was determined by usingBrdU. Each result is the mean � SD of three experiments performed in duplicate. *, P 0.001, versus mice treated with antigen alone. (D) Functional CD4�CD25�
Treg cells generated by VIP require TCR stimulation. CD4�CD25� T cells (105 cells/well) from PCCF-Tg mice treated with PCCF plus VIP for 8 days werecocultured with CD4�CD25– T cells (5�105 cells/well) from OVA-immunized B10.A mice in the presence of mitC-treated B10.A spleen APC and antigen (5 �MPCCF and/or 200 �g/ml OVA) or soluble anti-CD3 antibody (5 �g/ml). Proliferation was determined by using BrdU. Each result is the mean � SD of twoexperiments performed in duplicate.
Delgado et al. VIP induces Treg cells in vivo 1333

CD25. CD25– and CD25� CFSE-labeled cells from antigenand antigen � VIP-injected donors were present in the spleenof recipients 2 weeks after inoculation (Fig. 6C). Although bothpopulations proliferated, CD25� cells proliferated less thanCD25– T cells.
Effects of VIP-generated Treg cells in GVHDand GVL
The treatment of choice for hematological malignancies con-sists of allogeneic BM transplantation (BMT) following hostirradiation or chemotherapy. The donor T cells responsible forthe elimination of the surviving malignant cells are also thereason for the major complication, i.e., GVHD. Recently, it hasbeen reported that Treg cells play an essential role in estab-lishing tolerance to alloantigens and that Treg cells can beused to control GVHD. The question remains whether Treg
cells, whereas limiting GVHD, still allow a graft-versus-tumor(GVT) response. We assessed first the potential use of VIP-induced Treg cells in a model of GVHD. Irradiated BALB/crecipients (H-2d) were reconstituted with allogeneic (H-2k)TCD BM cells together with CD4� splenic T cells obtainedfrom Tg-PCCF mice (H-2k), previously inoculated with PCCF[CD4�(antigen)] or with PCCF � VIP [CD4�(antigen�VIP)]or from naı̈ve mice (control). Mice that received CD4�(antigen)developed severe GVHD, including weight loss, reduced mo-bility, hunched posture, diarrhea, and ruffled fur and diedwithin 30 days. In contrast, mice that received CD4�-(antigen�VIP) were protected, and more than 60% survivedfor more than 75 days (Fig. 7A). Control mice that receivedCD4� T cells from naı̈ve Tg PCCF mice developed GVHDsimilar to those receiving CD4�(antigen; results not shown).When the CD4� T cells were separated into CD25– and
Fig. 5. In vivo VIP-induced Treg cells act through direct cell contact. PCCF-Tg mice were injected i.p. with medium (Naive) or PCCF with or without VIP. Eightdays later, spleen CD4�, CD4�CD25�, and CD4�CD25– T cells were isolated. (A) Total CD4� T cells [CD4(Ag�VIP)] or sorted CD4�CD25� T cells[CD25(Ag�VIP); 2.5�105 cells/well] were cocultured with rCD4� T cells from naı̈ve-Tg mice (5�104 cells/well) in the presence of mitC-treated B10.A spleenAPC and PCCF. Coculture experiments were also performed in the presence of blocking mouse anti-IL-10 (10 �g/ml), anti-TGF-�1 (40 �g/ml), and/or anti-CTLA-4(10 �g/ml) mAb or of rmIL-2 (100 U/ml). Proliferation was determined by using BrdU. In addition, rCD4� T cells (5�104 cells) were placed with mitC-treatedB10.A spleen APC and PCCF in the bottom wells of a Transwell system, and total CD4� or sorted CD4�CD25� Treg cells (2�104 cells) from PCCF � VIP-treatedmice were placed with mitC-treated B10.A spleen APC and PCCF in the upper wells. Three days later, the basket was removed, and the proliferative responseof the bystander rCD4� T cells was measured after a pulse with BrdU during the final 16 h of the 72-h culture. Results represent mean � SD of triplicates fromone representative experiment out of three. *, P 0.001, versus untreated samples (None). (B) Spleen cells from mice treated with medium (Naı̈ve), PCCF, orPCCF plus VIP were analyzed for CD4, CD25, and intracellular CTLA-4 by flow cytometry. (Left panels) Double-staining for CD25 and CTLA-4 expression of gatedCD4 T cells. Values refer to percentage of positive cells in each quadrant. (Right panels) Intracellular CTLA-4 expression was analyzed for the CD4�CD25� andCD4�CD25– populations. Values refer to the percentage of CTLA-4� cells. One representative experiment out of three is shown.
1334 Journal of Leukocyte Biology Volume 78, December 2005 http://www.jleukbio.org

CD25� subpopulations, the CD25�(antigen�VIP) cells weremore protective than the CD25�(antigen) cells (Fig. 7B). Next,we determined the effect of CD4�(antigen) and CD4�
(antigen�VIP) cells on anti-tumor activity. Host-irradiatedBALB/c mice (H-2d) were reconstituted with TCD allogeneicBM cells (H-2k) plus A20 leukemic cells (H-2d) andCD4�(antigen) or CD4�(antigen�VIP) T cells (H-2k). BloodA20 leukemic cells were identified as B220�H-2d by FACSanalysis. Mice receiving only BM cells and A20 died betweenDays 20 and 30, and A20 cells were present in high numbersin the blood. Mice receiving BM and CD4�(antigen) T cellsdied of GVHD between Days 5 and 24, although the A20 tumorcells had been eliminated. In contrast, 80% of the mice thatreceived BM and CD4�(antigen�VIP) T cells survived, and noA20 cells were present in blood (Fig. 7C). These resultssuggest that the CD4�(antigen�VIP) T cells consist of apopulation that eliminates the allogeneic tumor cells and asecond population that prevents GVHD. The anti-tumor T cellsresided within the CD4�CD25– population, as purifiedCD4�CD25� T cells from the antigen � VIP-injected micedid not eliminate the tumor cells (Fig. 7C).
To finally demonstrate that the CD25� Treg cells generatedby antigen � VIP protect against GVHD and still allow GVTto occur, we reconstituted BALB/c-irradiated hosts (H-2d) withallogeneic TCD BM cells (H-2k), plus A20 leukemic cells
(H-2d), CD4� T cells from B10.A mice (H-2k), andCD25�(antigen) or CD25�(antigen�VIP) T cells (H-2k). Asexpected, the B10.A CD4� T cells eliminated the leukemiccells but killed the hosts through GVHD (between Days 5 and20; Fig. 7D). The CD25�(antigen�VIP) T cells protectedagainst GVHD (80% survival at 70 days) without affectingGVT (no A20 cells were detected). The CD25�(antigen) T cellsalso had a protective effect but to a lesser extent (50% survivalat 70 days; Fig. 7D).
DISCUSSION
VIP and PACAP are potent, immunosuppressive agents thatproved to be protective in models of autoimmune diseases suchas collagen-induced arthritis and inflammatory bowel disease[15–17]. Until now, the VIP/PACAP were described as deac-tivators of macrophages, dendritic cells, and microglia andpromoters of Th2 effector differentiation and survival [12–14].This is the first report about the in vivo effect of VIP onCD4�CD25� Treg cell activity.
We compared the effects of antigen and antigen � VIPadministration in TCR-Tg mice and concluded that VIP con-tributes primarily to the expansion of the CD4�CD25� Tregcell population. The increase in the numbers of CD4�CD25�
Fig. 6. In vivo VIP-induced Treg cells are capableof transferring tolerance. PCCF-Tg mice were in-jected i.p. with PCCF (Ag) or with PCCF plus VIP(Ag�VIP). Eight days later, spleen CD4� orCD4�CD25� T cells were isolated. (A) Total CD4�
T cells (1�106 cells; 100 �l) were injected i.v. intoPCCF-Tg mice. Twenty-four hours later, mice wereimmunized subcutaneously (s.c.) with 100 �gPCCF/CFA in the footpads. Two weeks after immu-nization, spleen, PLN, and DLN were isolated. Re-sults are representative of three experiments. (B)Total CD4� or sorted CD4�CD25� T cells (106
cells) were injected i.v. into PCCF-Tg mice. Miceinjected with medium instead of cells were used ascontrols. Twenty-four hours later, mice were immu-nized with 100 �g PCCF/CFA in the footpads (s.c.).Two weeks after immunization, the mice were chal-lenged intradermally with 50 �g PCCF in 20 �lPBS in the left ear or with 20 �l PBS in the rightear. Ear thickness was measured 24 h later. earthickness � thickness of left ear – thickness ofright ear. In addition, 2 weeks after immunization,mice were analyzed individually for splenic T cellproliferation in response to antigen and serum an-tibody responses. Spleen cells (4�105 cells/well)were incubated with PCCF (5 �M), and the prolif-erative response was determined by BrdU incorpo-ration. Serum was collected and assayed for PCCF-specific IgG antibodies by ELISA. Results repre-sent mean � SD of triplicates from onerepresentative experiment out of three. *, P 0.001, versus mice treated with antigen alone. (C)The relative presence and the proliferative state ofthe transferred CFSE-stained CD4� or CD25� cellsin the spleen were determined by flow cytometry.One representative experiment out of three isshown.
Delgado et al. VIP induces Treg cells in vivo 1335

cells occurred only in the antigen-specific T cells for low dosesof antigen and was maintained long-term. CD4�CD25� Tregcells have been characterized by high expression of the tran-scription repressor Foxp3, high surface expression of GITR,Nrp1, CD103, CD62L, and CD69, and low expression ofCD45RB [18–22]. The CD4 population from antigen � VIP-
inoculated mice showed a decrease in CD45RB and increasesin the expression of all the other markers, as compared with theCD4� T cells from antigen-inoculated mice. However, thepurified CD4�CD25� T cells from antigen- or antigen �VIP-inoculated mice expressed similar levels of the markerscharacteristic for CD4�CD25� Treg cells. These results sug-
Fig. 7. In vivo VIP-induced Treg cells inhibit acute lethal GVHD. PCCF-Tg mice were injected i.p. with PCCF (Ag) or with PCCF plus VIP (Ag�VIP). Eightdays later, spleen CD4�, CD4�CD25–, and CD4�CD25� T cells were isolated. (A) BALB/c mice (groups of 10, H-2d) were irradiated and injected i.v. with TCDBM cells (5�106 cells), isolated from B10.A mice (H-2k), with or without CD4� T cells (106 cells) from PCCF-Tg mice inoculated with PCCF or PCCF plus VIPas described above. Survival and appearance were monitored daily, and body weight was measured weekly after BMT. (B) BALB/c hosts (groups of 10) wereirradiated and injected i.v. with B10.A TCD BM (5�106 cells) and B10.A spleen CD4� T cells (0.5�106 cells), plus CD4�CD25– or CD4�CD25� cells (0.5�106
cells) from PCCF-Tg mice inoculated with PCCF (Ag) or PCCF plus VIP (Ag�VIP) as described above. (C) Irradiated BALB/c hosts (groups of eight) wereinoculated with BMT (106 cells) and A20 leukemic cells (1�104 cells). CD4� T cells or CD4�CD25� cells (106 cells) from PCCF-Tg mice treated with PCCF(Ag) or PCCF plus VIP (Ag�VIP) were also injected. Survival was monitored, and tumor growth/elimination was assessed by the presence of A20 cells in blooddetected by size and coexpression of B220 and H-2Kd. (D) GVT is maintained in the presence of VIP-induced CD25� Treg cells. Irradiated BALB/c hosts (groupsof 10) were inoculated with BMT (B10.A TCD BM cells, 106 cells) and A20 leukemic cells (derived from BALB/c mice, 1�104 cells). B10.A splenic CD4� T cells(0.5�106 cells) and CD4�CD25� cells (0.5�106 cells) from PCCF-Tg mice treated with PCCF (Ag) or PCCF plus VIP (Ag�VIP) were injected. Survival and tumorgrowth/elimination were assessed as in C. *, P 0.001, versus mice treated with antigen alone.
1336 Journal of Leukocyte Biology Volume 78, December 2005 http://www.jleukbio.org

gest that VIP expands the CD25� population without inducingincreases in Foxp3 or Nrp1 (or other markers) on a per-cellbasis. In addition to expanding the CD4�CD25� population,VIP also induced more efficient Treg cells in terms of suppres-sive activity. On a per-cell basis, the VIP-inducedCD4�CD25� Treg cells are stronger suppressors of responderT cell proliferation, particularly at low Treg/Tresp cell ratios.
Several subsets of Treg cells with different phenotypes andsuppressive mechanisms have been described [7, 23]. In con-trast to Tr1/Tr2(Th3), which suppress mostly through cytokinerelease (IL-10 and TGF-�), CD4�CD25� naturally occurringTreg cells suppress through direct cellular contact. The in vivoVIP-generated CD4� Treg cells appear to consist of two pop-ulations: a major CD4�CD25� population, whose suppressivemechanism is mediated through direct cellular contact anddoes not involve IL-10 or TGF-�, and a minor CD4� popula-tion, which produces and uses IL-10 and/or TGF-� as sup-pressive molecules. The major population of Treg cells inducedby VIP indeed resembles the recently reported CD25� cyto-kine-independent suppressors recruited from the peripheralCD25– population by CD4�CD25� T cells stimulated withIL-2 and TGF-� [24].
Whether VIP acts directly on T cells inducing the generationor expansion of CD4�CD25� Treg cells remains to be estab-lished. Another neuropeptide, the � melanocyte-stimulatinghormone, has been shown to induce CD4�CD25� Treg cells invitro in conjunction with antigen stimulation or TGF-� [25,26]. The role of TGF-� in the expansion of CD4�CD25� Tregcells has been demonstrated in vivo and in vitro [27, 28].Experiments are in progress to determine whether the in vitrotreatment with VIP and TGF-� converts antigen-primed T cellsto CD25�Foxp3� Treg cells.
The in vivo VIP-induced CD4�CD25� Treg cells transferantigen-specific suppression to naı̈ve hosts, inhibiting DTH, aTh1-dependent immune reaction, and antibody formation, aTh2-dependent response. The in vivo induction of Treg by VIPis physiologically significant from at least two points of view.First, our results suggest that VIP is one of the endogenousfactors controlling the peripheral expansion of theCD4�CD25� Treg cells. This is important, as little is presentlyknown about the factors and mechanisms controlling Treg cellexpansion. Second, the induction of Treg cells adds an addi-tional dimension to the general anti-inflammatory role of VIP,which was reported previously to occur through macrophage/microglia deactivation and bias toward Th2 immunity.
The possible therapeutic use of the VIP-induced Treg cellshas been assessed in a model of allogeneic BMT. AllogeneicBMT is a treatment of choice in many hematopoietic malig-nancies. Following high-dose chemotherapy or irradiation, thehost is reconstituted with BM cells, and the donor T cells areresponsible for the GVT effects, which eliminate the remainingmalignant cells in the host. However, the same donor T cellsinitiate a GVHD reaction, which represents the major compli-cation following allogeneic stem cell transplantation. There-fore, a desirable therapy will eliminate GVHD without affect-ing the GVT response. In mice, Treg cells have been shown toprevent lethal GVHD in lethally irradiated hosts reconstitutedwith allogeneic BM [29, 30]. Recent studies have also demon-strated that although controlling GVHD, CD4�CD25� Treg
cells maintain the GVL response [31]. Similar to these reports,the VIP-induced CD4�CD25� Treg cells prevented GVHDand maintained GVT in a murine model for allogeneic BMT.These studies suggest the possible use of VIP-induced Tregcells in the survival after hematopoietic stem cell transplanta-tion.
ACKNOWLEDGMENTS
This work was supported by Grants AI52306 and AI47325(D. G.), Johnson and Johnson Neuroimmunology Fellowships(M. D.), the Spanish Ministry of Health PI04/0674, and RamonAreces Foundation (M. D.). The authors have no conflictingfinancial interests.
REFERENCES
1. Fehervari, Z., Sakaguchi, S. (2004) Development and function ofCD25�CD4� regulatory T cells. Curr. Opin. Immunol. 16, 203–208.
2. Wraith, D. C., Nicolson, K. S., Whitlet, N. T. (2004) Regulatory CD4� Tcells and the control of autoimmune disease. Curr. Opin. Immunol. 16,695–701.
3. Piccirillo, C. A., Shevach, E. M. (2004) Naturally-occurringCD4�CD25� immunoregulatory T cells: central players in the arena ofperipheral tolerance. Semin. Immunol. 16, 81–88.
4. Tang, Q., Henriksen, K. J., Boden, E. K., Tooley, A. J., Ye, J., Subudhi,S. K., Zheng, X. X., Strom, T. B., Bluestone, J. A. (2003) Cutting edge:CD28 controls peripheral homeostasis of CD4�CD25� regulatory T cells.J. Immunol. 171, 3348–3352.
5. Malek, T. R., Yu, A., Vincek, V., Scibelli, P., Kong, L. (2002) CD4regulatory T cells prevent lethal autoimmunity in IL-2R�-deficient mice:implications for the nonredundant function of IL-2. Immunity 17, 167–178.
6. Akbar, A. N., Taams, L. S., Salmon, M., Vukmanovic-Stejic, M. (2003)The peripheral generation of CD4�CD25� regulatory T cells. Immunol-ogy 109, 319–325.
7. Thompson, C., Powrie, F. (2004) Regulatory T cells. Curr. Opin. Phar-macol. 4, 408–414.
8. Jonuleit, H., Schmitt, E., Kakirman, H., Stassen, M., Knop, J., Enk, A. H.(2002) Infectious tolerance: human CD25� regulatory T cells conveysuppressor activity to conventional CD4� T helper cells. J. Exp. Med.196, 255–260.
9. Dieckmann, D., Bruett, C. H., Ploettner, H., Lutz, M. B., Schuler, G.(2002) Human CD4�CD25� regulatory, contact-dependent T cells in-duce interleukin 10-producing, contact-independent type 1-like regula-tory T cells. J. Exp. Med. 196, 247–253.
10. Shevach, E. M. (2002) CD4�CD25� suppressor T cells: more questionsthan answers. Nat. Rev. Immunol. 2, 389–400.
11. Polanczyk, M. J., Carson, B. D., Subramanian, S., Afentoulis, M., Van-denbark, A. A., Ziegler, S. F., Offner, H. (2004) Cutting edge: estrogendrives expansion of the CD4�CD25� regulatory T cell compartment.J. Immunol. 173, 2227–2230.
12. Delgado, M., Pozo, D., Ganea, D. (2004) The significance of vasoactiveintestinal peptide in immunomodulation. Pharmacol. Rev. 56, 249–290.
13. Ganea, D., Rodriguez, R., Delgado, M. (2003) Vasoactive intestinal pep-tide and pituitary adenylate cyclase-activating polypeptide: players ininnate and adaptive immunity. Cell. Mol. Biol. 49, 127–142.
14. Ganea, D., Delgado, M. (2003) The neuropeptides VIP/PACAP and Tcells: inhibitors or activators? Curr. Pharm. Des. 9, 997–1004.
15. Delgado, M., Abad, C., Martinez, C., Leceta, J., Gomariz, R. P. (2001)Vasoactive intestinal peptide prevents experimental arthritis by down-regulating both autoimmune and inflammatory components of the disease.Nat. Med. 7, 563–568.
16. Abad, C., Martinez, C., Leceta, J., Gomariz, R. P., Delgado, M. (2001)Pituitary adenylate cyclase-activating polypeptide inhibits collagen-in-duced arthritis: an experimental immunomodulatory therapy. J. Immunol.167, 3182–3189.
17. Abad, C., Martinez, C., Juarranz, M. G., Arranz, A., Leceta, J., Delgado,M., Gomariz, R. P. (2003) Therapeutic effects of vasoactive intestinal
Delgado et al. VIP induces Treg cells in vivo 1337

peptide in the trinitrobenzene sulfonic acid mice model of Crohn’s disease.Gastroenterology 124, 961–971.
18. McHugh, R. S., Whitters, M. J., Piccirillo, C., Young, D. A., Shevach,E. M., Collins, M., Byrne, M. C. (2002) CD4�CD25� immunoregulatoryT cells: gene expression analysis reveals a functional role for the glu-cocorticoid-induced TNF receptor. Immunity 16, 311–323.
19. Shimizu, J., Yamazaki, S., Takahashi, T., Ishida, Y., Sakaguchi, S. (2002)Stimulation of CD25�CD4� regulatory T cells through GITR breaksimmunological self-tolerance. Nat. Immunol. 3, 135–142.
20. Hori, S., Nomura, T., Sakaguchi, S. (2003) Control of regulatory T celldevelopment by the transcription factor Foxp3. Science 299, 1057–1061.
21. Fontenot, J. D., Gavin, M. A., Rudenski, A. Y. (2003) Foxp3 programs thedevelopment and function of CD4�CD25� regulatory T cells. Nat. Im-munol. 4, 330–336.
22. Bruder, D., Probst-Kepper, M., Westendorf, A. M., Geffers, R., Beissert,S., Loser, K., von Boehmer, H., Buer, J., Hansen, W. (2004) Neuropilin-1:a surface marker of regulatory T cells. Eur. J. Immunol. 34, 623–630.
23. von Boehmer, H. (2005) Mechanisms of suppression by suppressor T cells.Nat. Immunol. 6, 338–344.
24. Zheng, S. G., Wang, J. H., Gray, J. D., Soucier, H., Horwitz, D. A. (2004)Natural and induced CD4�CD25� cells educate CD4�CD25– cells todevelop suppressive activity: the role of IL-2, TGF-�, and IL-10. J. Im-munol. 172, 5213–5221.
25. Namba, K., Kitaichi, N., Nishida, T., Taylor, A. W. (2002) Induction ofregulatory T cells by the immunomodulating cytokines �-melanocyte-
stimulating hormone and transforming growth factor-�2. J. Leukoc. Biol.72, 946–952.
26. Taylor, A., Namba, K. (2001) In vitro induction of CD25�CD4� regula-tory T cells by the neuropeptide �-melanocyte stimulating hormone (�-MSH). Immunol. Cell Biol. 79, 358–367.
27. Huber, S., Schramm, C., Lehr, H. A., Mann, A., Schmitt, S., Becker, C.,Prostschka, M., Galle, P. R., Neurath, M., Blessing, M. (2004) Cuttingedge: TGF-� signaling is required for the in vivo expansion and immu-nosuppressive capacity of regulatory CD4�CD25� T cells. J. Immunol.173, 6526–6531.
28. Chen, W., Jin, W., Hardegen, N., Lei, K., Li, L., Marinos, N., McGrady,G., Wahl, S. M. (2003) Conversion of peripheral CD4�CD25– naı̈ve Tcells to CD4�CD25� regulatory T cells by TGF-� induction of transcrip-tion factor Foxp3. J. Exp. Med. 198, 1875–1886.
29. Cohen, J. L., Trenado, A., Vasey, D., Klatzmann, D., Salomon, B. I. (2002)CD4(�)CD25(�) immunoregulatory T cells: new therapeutics for graft-versus-host disease. J. Exp. Med. 196, 401–406.
30. Hoffmann, P., Ermann, J., Edinger, M., Fathman, C. G., Strober, S. (2002)Donor type CD4(�)CD25(�) regulatory T cells suppress lethal acutegraft-versus-host disease after allogeneic bone marrow transplantation. J.Exp. Med. 196, 389–399.
31. Trenado, A., Charlotte, F., Fisson, S., Yagello, M., Klatzmann, D.,Salomon, B. L., Cohen, J. L. (2003) Recipient-type specific CD4�CD25�regulatory T cells favor immune reconstitution and control graft-versus-host disease while maintaining graft-versus-leukemia. J. Clin. Invest.112, 1688–1696.
1338 Journal of Leukocyte Biology Volume 78, December 2005 http://www.jleukbio.org




















