
Complete Regression of Experimental Solid Tumors by Combination LEC/chTNT-3 Immunotherapy and CD25...
10
[CANCER RESEARCH 63, 8384 – 8392, December 1, 2003] Complete Regression of Experimental Solid Tumors by Combination LEC/chTNT-3 Immunotherapy and CD25 T-Cell Depletion Jiali Li, Peisheng Hu, Leslie A. Khawli, and Alan L. Epstein Department of Pathology, University of Southern California, Keck School of Medicine, Los Angeles, California ABSTRACT LEC/chTNT-3, a chemokine fusion protein generated previously in our laboratory, produces a 40 – 60% reduction in well-established solid tumors of the BALB/c mouse. In this study, CD25 T-cell depletion was used in combination with LEC/chTNT-3 treatment to enhance the therapeutic value of this approach. In two tumor models (Colon 26 and RENCA), this combination immunotherapy produced complete regression of established s.c. tumors after 5 consecutive days of i.v. treatment. To show that targeted LEC is critical to these results, similar combination studies were performed with chTNT-3/cytokine fusion proteins consisting of human interleukin 2, murine IFN-, and murine granulocyte macrophage colony- stimulating factor using identical treatment regimens. These studies showed no significant improvement indicating that combination therapy with anti-CD25 antisera requires LEC localization to tumor to produce complete regression. To study the mechanism of this remarkable response, immunotherapeutic studies were repeated in knockout mice and showed that successful treatment with CD25 depletion was dependent on the presence of IFN- but not perforin. Other studies using real-time PCR, ex vivo proliferation, and intracellular cytokine staining with lymphocytes from tumor draining lymph nodes, suggested that this combination treat- ment was associated with increased T-helper 1 cytokine expression, en- hanced T-cell activation, and increased IFN- production by T cells. Rechallenge experiments showed that combination LEC/chTNT-3 treat- ment and CD25 depletion produced long-acting memory cells capable of preventing re-engraftment of the same but not different tumor cell lines. These studies suggest that LEC/monoclonal antibody fusion proteins, when used in combination with CD25 T-cell depletion, is a viable method of immunotherapy for the treatment of solid tumors. INTRODUCTION T-reg. 1 or suppressor T cells have been described for 20 years (1–3), but only recently have their roles in autoimmune disease and cancer been recognized. CD4 CD25 cells constitutively express CD25 (IL-2 receptor chain) on their surface and constitute 5–10% of CD4 T cells in humans and rodents. Although a number of studies on CD4 CD25 T-cell subsets have appeared in the literature, most have focused on their role in autoimmune diseases. Recent studies indicate that there may be two mechanisms regarding the function of T-reg., one that is cell-cell contact-dependent, and the other that is cytokine (IL-4 and IL-10) -dependent (4). Most recently, it was found that the cytotoxic T lymphocyte-associated antigen 4, which is con- stitutively expressed on CD4 CD25 T cells, also plays a key role in T cell-mediated dominant immunological self-tolerance (5). Another suppressive cell population, CD8 -suppressive T cells, has been shown to be related to oral tolerance (6, 7), but these cells have only been shown to suppress local intestinal immune function (8). Finally, CD8 CD25 T cells have been identified recently in human thymus (9). Although these cells appear to share a similar phenotype and regulatory functions as CD4 CD25 T cells, their role in peripheral tissues remains to be demonstrated. It is clear that T-reg. is essential in immune system homeostasis (10 –12), and the manipulation of its function should have potential therapeutic effects in the clinic (13). Specifically, the enhancement of T-reg. may be beneficial for autoimmune diseases, and the removal of T-reg. should result in increased immune responses that can facilitate the induction or augmentation of tumor immunity (14 –16). Indeed, recent studies have demonstrated that in vivo injection of anti-CD25 antibody caused the regression of leukemia and solid tumors in animal models (14, 17). In most of these studies, CD4 or CD4 CD25 T-cell depletion was tumor suppressive, but T-reg. depletion alone resulted in either incomplete tumor reduction or a delay in the growth of well-established tumor implants. Combination therapy seems to be more effective as exemplified by studies performed by Shimizu et al. (14), which revealed a synergistic effect of cytotoxic T lymphocyte- associated antigen 4 blockage and the suppression of CD4 CD25 T cells in tumor therapy. In the current investigation, a new combination treatment is pro- posed, which uses the depletion of CD4 CD25 T-reg. and the use of a chemokine/mAb fusion protein to attract and activate the immune response at the tumor site. It has been shown previously that there is a strong correlation between the infiltration of lymphocytes into tumor sites and increased survival, suggesting that cytotoxic T-cells contrib- ute to tumor remission (18). Methods to increase the penetration of lymphocytes into the tumor microenvironment would be very bene- ficial for active immunotherapy. To accomplish this, chemokine/ antibody fusion proteins are logical candidates due to the chemoat- tractant properties of their chemokine moiety on different populations of lymphocytes and dendritic cells by receptor-ligand interactions (19 –21). LEC/chTNT-3 is a fusion protein generated in our laboratory (22), which was genetically engineered to link the liver expression chemo- kine (LEC, also named HCC-4, NCC-4, and CCL-16; Ref. 23) to mAb chTNT-3, which targets the necrotic regions of tumors (24). We demonstrated previously that LEC/chTNT-3 attracts different sub- populations of lymphocytes including CD4 and CD8 T cells, neutrophils, dendritic cells, and B cells (22). Although dramatic lymphocyte migration has been observed in the tumors of treated mice, LEC/chTNT-3 treatment alone resulted in a modest 40 –50% reduction of tumor growth. We hypothesize that the infiltrated lym- phocytes might be suppressed by normal regulatory pathways. To test this hypothesis, we combined the use of LEC/chTNT-3 with methods to deplete T-reg. in an effort to optimize the immunotherapy of experimental solid tumors of the mouse. MATERIALS AND METHODS Antibodies and Cell Lines Hybridomas, including rat antimouse L3T4 (anti-CD4) mAb GK1.5, anti- lyt-2.3 (anti-CD8) mAb 2.43, and anti- IL-2 (anti-CD25) receptor mAb 7D4 Received 7/17/03; revised 9/15/03; accepted 9/25/03. Grant support: California Cancer Research Program #00-00749v-20244, the Philip Morris External Research Program, Cancer Therapeutics Laboratories (Los Angeles, CA), and MediPharm, Inc. (Los Angeles, CA). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Requests for reprints: Alan L. Epstein, Department of Pathology, University of Southern California, Keck School of Medicine, 2011 Zonal Avenue, Los Angeles, CA 90033. 1 The abbreviations used are: T-reg., T-regulatory cell; IL, interleukin; mAb, mono- clonal antibody; MAD109, Madison 109; TDLN, tumor draining lymph node; FACS, fluorescence-activated cell sorter; PE, phycoerythrin; CFSE, 5-(and-6)-carboxyfluores- cein diacetate, succinimidyl ester; TNF, tumor necrosis factor; NK, natural killer; PMN, polymorphonuclear; Th, T-helper. 8384 Research. on July 29, 2015. © 2003 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Complete Regression of Experimental Solid Tumors by Combination LEC/chTNT-3 Immunotherapy and CD25...

[CANCER RESEARCH 63, 8384–8392, December 1, 2003]
Complete Regression of Experimental Solid Tumors by Combination LEC/chTNT-3Immunotherapy and CD25� T-Cell Depletion
Jiali Li, Peisheng Hu, Leslie A. Khawli, and Alan L. EpsteinDepartment of Pathology, University of Southern California, Keck School of Medicine, Los Angeles, California
ABSTRACT
LEC/chTNT-3, a chemokine fusion protein generated previously in ourlaboratory, produces a 40–60% reduction in well-established solid tumorsof the BALB/c mouse. In this study, CD25� T-cell depletion was used incombination with LEC/chTNT-3 treatment to enhance the therapeuticvalue of this approach. In two tumor models (Colon 26 and RENCA), thiscombination immunotherapy produced complete regression of establisheds.c. tumors after 5 consecutive days of i.v. treatment. To show thattargeted LEC is critical to these results, similar combination studies wereperformed with chTNT-3/cytokine fusion proteins consisting of humaninterleukin 2, murine IFN-�, and murine granulocyte macrophage colony-stimulating factor using identical treatment regimens. These studiesshowed no significant improvement indicating that combination therapywith anti-CD25� antisera requires LEC localization to tumor to producecomplete regression. To study the mechanism of this remarkable response,immunotherapeutic studies were repeated in knockout mice and showedthat successful treatment with CD25� depletion was dependent on thepresence of IFN-� but not perforin. Other studies using real-time PCR, exvivo proliferation, and intracellular cytokine staining with lymphocytesfrom tumor draining lymph nodes, suggested that this combination treat-ment was associated with increased T-helper 1 cytokine expression, en-hanced T-cell activation, and increased IFN-� production by T cells.Rechallenge experiments showed that combination LEC/chTNT-3 treat-ment and CD25� depletion produced long-acting memory cells capable ofpreventing re-engraftment of the same but not different tumor cell lines.These studies suggest that LEC/monoclonal antibody fusion proteins,when used in combination with CD25� T-cell depletion, is a viable methodof immunotherapy for the treatment of solid tumors.
INTRODUCTION
T-reg.1 or suppressor T cells have been described for �20 years(1–3), but only recently have their roles in autoimmune disease andcancer been recognized. CD4�CD25� cells constitutively expressCD25 (IL-2 receptor � chain) on their surface and constitute 5–10%of CD4� T cells in humans and rodents. Although a number of studieson CD4�CD25� T-cell subsets have appeared in the literature, mosthave focused on their role in autoimmune diseases. Recent studiesindicate that there may be two mechanisms regarding the function ofT-reg., one that is cell-cell contact-dependent, and the other that iscytokine (IL-4 and IL-10) -dependent (4). Most recently, it was foundthat the cytotoxic T lymphocyte-associated antigen 4, which is con-stitutively expressed on CD4�CD25� T cells, also plays a key role inT cell-mediated dominant immunological self-tolerance (5). Anothersuppressive cell population, CD8�-suppressive T cells, has been
shown to be related to oral tolerance (6, 7), but these cells have onlybeen shown to suppress local intestinal immune function (8). Finally,CD8�CD25� T cells have been identified recently in human thymus(9). Although these cells appear to share a similar phenotype andregulatory functions as CD4�CD25� T cells, their role in peripheraltissues remains to be demonstrated.
It is clear that T-reg. is essential in immune system homeostasis(10–12), and the manipulation of its function should have potentialtherapeutic effects in the clinic (13). Specifically, the enhancement ofT-reg. may be beneficial for autoimmune diseases, and the removal ofT-reg. should result in increased immune responses that can facilitatethe induction or augmentation of tumor immunity (14–16). Indeed,recent studies have demonstrated that in vivo injection of anti-CD25antibody caused the regression of leukemia and solid tumors in animalmodels (14, 17). In most of these studies, CD4� or CD4�CD25�
T-cell depletion was tumor suppressive, but T-reg. depletion aloneresulted in either incomplete tumor reduction or a delay in the growthof well-established tumor implants. Combination therapy seems to bemore effective as exemplified by studies performed by Shimizu et al.(14), which revealed a synergistic effect of cytotoxic T lymphocyte-associated antigen 4 blockage and the suppression of CD4�CD25� Tcells in tumor therapy.
In the current investigation, a new combination treatment is pro-posed, which uses the depletion of CD4�CD25� T-reg. and the use ofa chemokine/mAb fusion protein to attract and activate the immuneresponse at the tumor site. It has been shown previously that there isa strong correlation between the infiltration of lymphocytes into tumorsites and increased survival, suggesting that cytotoxic T-cells contrib-ute to tumor remission (18). Methods to increase the penetration oflymphocytes into the tumor microenvironment would be very bene-ficial for active immunotherapy. To accomplish this, chemokine/antibody fusion proteins are logical candidates due to the chemoat-tractant properties of their chemokine moiety on different populationsof lymphocytes and dendritic cells by receptor-ligand interactions(19–21).
LEC/chTNT-3 is a fusion protein generated in our laboratory (22),which was genetically engineered to link the liver expression chemo-kine (LEC, also named HCC-4, NCC-4, and CCL-16; Ref. 23) to mAbchTNT-3, which targets the necrotic regions of tumors (24). Wedemonstrated previously that LEC/chTNT-3 attracts different sub-populations of lymphocytes including CD4� and CD8� T cells,neutrophils, dendritic cells, and B cells (22). Although dramaticlymphocyte migration has been observed in the tumors of treatedmice, LEC/chTNT-3 treatment alone resulted in a modest 40–50%reduction of tumor growth. We hypothesize that the infiltrated lym-phocytes might be suppressed by normal regulatory pathways. To testthis hypothesis, we combined the use of LEC/chTNT-3 with methodsto deplete T-reg. in an effort to optimize the immunotherapy ofexperimental solid tumors of the mouse.
MATERIALS AND METHODS
Antibodies and Cell Lines
Hybridomas, including rat antimouse L3T4 (anti-CD4) mAb GK1.5, anti-lyt-2.3 (anti-CD8) mAb 2.43, and anti- IL-2 (anti-CD25) receptor mAb 7D4
Received 7/17/03; revised 9/15/03; accepted 9/25/03.Grant support: California Cancer Research Program #00-00749v-20244, the Philip
Morris External Research Program, Cancer Therapeutics Laboratories (Los Angeles, CA),and MediPharm, Inc. (Los Angeles, CA).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Requests for reprints: Alan L. Epstein, Department of Pathology, University ofSouthern California, Keck School of Medicine, 2011 Zonal Avenue, Los Angeles, CA90033.
1 The abbreviations used are: T-reg., T-regulatory cell; IL, interleukin; mAb, mono-clonal antibody; MAD109, Madison 109; TDLN, tumor draining lymph node; FACS,fluorescence-activated cell sorter; PE, phycoerythrin; CFSE, 5-(and-6)-carboxyfluores-cein diacetate, succinimidyl ester; TNF, tumor necrosis factor; NK, natural killer; PMN,polymorphonuclear; Th, T-helper.
8384
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

and PC61 were purchased from American Type Culture Collection (Manassas,VA). To obtain sufficient quantities of reagents, hybridoma cells were grownin Integra CL 1000 culture chambers (IBS Integra Biosciences, Wallisellen,Switzerland) and purified by ammonium sulfate precipitation following byQ-Sepharose ion-exchange chromatography (Bio-Rad Laboratories, Hercules,CA). Anti-asialo GM1 (anti-NK) was purchased from Wako Pure ChemicalIndustries Ltd. (Osaka, Japan). The Colon 26 murine colon carcinoma and theRENCA murine renal cell carcinoma were obtained from the American TypeCulture Collection. The MAD109, a murine lung carcinoma, was purchasedfrom the National Cancer Institute (Frederick, MD).
Animals
Six-week-old female BALB/c mice were obtained from Harlan SpragueDawley (Indianapolis, IN). Perforin and IFN-� knockout mice with H-2d
backgrounds were generously provide by Dr. Stephen Stohlman (Departmentof Neurology, University of Southern California Keck School of Medicine,Los Angeles, CA; Ref. 25).
Depletion of Lymphocytes Subsets in Vivo
Antibodies were administrated 1 day before tumor implantation for CD25�
T-cell depletion studies or 6 days after tumor implantation for the depletion ofthe other T-cell subsets. For CD4�, CD8�, and CD25� T-cell depletion, 0.5mg of anti-CD4 antibody (GK1.5), anti-CD8 antibody (2.43), or anti-CD25(PC61) were injected i.p. in a 1-ml inoculum, and repeated every 5 days. ForNK cell depletion, 0.35 mg of anti-asialo GM1 was injected i.p. every 7 days.Depletion of specific T-cell subsets was confirmed by flow cytometric anal-yses of splenocytes using normal mice (data not shown). Each of these studieswas repeated two or three times, and each group contained 5 mice each.
Immunotherapy Studies
Groups (n � 5) of 6-week-old female BALB/c mice were injected s.c. in theleft flank with a 0.2-ml inoculum containing �107 Colon 26, RENCA, orMAD109 cells under a University Animal Care Committee-approved protocol.Treatments were started when tumors reached �0.5 cm in diameter. Groups oftumor-bearing mice (with or without lymphocyte subset depletion) weretreated i.v. with a 0.1-ml inoculum containing LEC/chTNT-3 (20 �g) orcontrol chTNT-3 (20 �g). In addition, LEC/chTNT-3 was replaced with threeother fusion proteins developed in our laboratory, including chTNT-3/IL-2(26), chTNT-3/IFN-� (27), muTNT-3/mu-granulocyte macrophage colony-stimulating factor2 using the same dosage level. All of the groups were treateddaily times 5, and tumor growth was monitored every other day by calipermeasurement in three dimensions. Tumor volumes were calculated by theformula: length � width � height. The results were expressed as themean � SD, and the significance levels (Ps) were determined usingthe Wilcoxon rank-sum test. Each of these studies was repeated twice.
Rechallenge Experiments
One month (n � 7) and 6 months (n � 10) after the completion of treatment,tumor-free mice from previous studies and control naı̈ve mice (n � 5) werechallenged with 107 cells of Colon 26 and MAD109 or Colon 26 and RENCAin the left and right flanks, respectively. The injection sites were observed for3–4 weeks. To study the presence of CD4�CD25� T cells in different groups,TDLNs were removed, and T-reg. were stained with PE-anti-CD25 and FITC-anti-CD4 for FACS analysis.
Mechanistic Studies
Infiltration of Lymphocytes by Flow Cytometric Analysis. Tumors fromcontrol and treated mice were aseptically removed on days 9, 15, and 20 aftertumor implantation and manually cut into 2–3-mm pieces in a culture Petridish. The small tissue fragments were then digested with 0.01% DNase, 0.01%hyaluranidase, and 0.1% collagenase (all from Sigma Chemical Co.) in RPMI1640 for 2–3 h at 37°C with continuous stirring. The resulting single cellsuspensions were then washed twice with 0.1% FCS in PBS and stained bystandard flow cytometry methods. To detect subpopulations of lymphocytes
infiltrating these tissues, the following conjugated antibodies were used forFACS: PE-anti-CD4, FITC-anti-CD8, PE-anti-PMN, FITC-anti-CD25, APC-anti-CD11b, FITC-anti-CD11c, and FITC-Pan NK (BD Biosciences PharM-ingen, San Diego, CA). Experiments were repeated three times, and the mostrepresentative data are shown below.
Intracellular IFN-� Production. TDLNs from control and treated micewere removed from mice on days 15 and 20 after tumor implantation. Singlecell suspensions were obtained as described above, and 2 � 106 viablecells/well were plated into a 24-well plate. Intracellular IFN-� production wasperformed by first stimulating the cells for 4 h in complete RPMI 1640containing 10 ng/ml PMA (Sigma, Aldrich) and 1000 ng/ml of ionomycin(Sigma) in the presence of GolgiStop (BD PharMingen). Cells were thenwashed, and mouse Fc receptors were blocked with 1 �g Fc Blocking antibody(CD16/CD32) per 106 cells in 100 �l of Staining Buffer (1% fetal bovineserum in PBS) for 15 min at 4°C. Cells were then stained with a PE-conjugatedanti-CD3e antibody for 30 min at 4°C, fixed/permeabilized with 100 �lCytofix/Cytoperm (BD PharMingen) for 15 min at 4°C, and washed with 300�l of Perm/Wash (BD PharMingen). The fixed cells were then resuspended in50 �l Perm/Wash containing of FITC-conjugated anti-IFN-� antibody (BDPharMingen) for 30 min at 4°C in the dark. Cells were washed and resus-pended in FACS buffer, and the intracellular production of IFN-� was ana-lyzed by FACS.
Knockout Mice Immunotherapy Studies. For the perforin knockoutmouse studies, 107 Colon 26 cells were implanted on day 1 in the left flank,and mice were depleted of CD4� T cells on day 6 with 0.5 mg/mouse ofGK1.5, which was repeated every 5 days. Treatment began on day 7 whentumors reached 0.5 cm in diameter. Mice were divided into three groups andtreated with: (a) PBS; (b) chTNT-3 (20 �g/mouse); or (c) LEC/chTNT-3 (20�g/mouse) i.v. with a 0.1-ml inoculum. All of the groups were treated daily for5 days, and tumor volumes were monitored every other day. For the IFN-�knockout mice studies, Colon 26 tumor-bearing mice were divided into fourgroups and treated for 5 consecutive days i.v. with: (a) control chTNT-3 (20�g/mouse); (b) LEC/chTNT-3 (20 �g/mouse); (c) CD25� depletion andcontrol chTNT-3 (20 �g/mouse); or (d) CD25� depletion and LEC/chTNT-3(20 �g/mouse). Tumor volumes were monitored by caliper as described above.
T-Cell Proliferation Assay. The proliferation of T cells was measured bya modified flow cytometry method. Briefly, TDLNs from control and treatedmice were removed from tumor-bearing mice on days 15 and 20 after tumorimplantation. Single cell suspensions obtained by mincing the lymph nodes ina Petri dish were labeled with CFSE (Molecular Probes, Eugene, OR) with thefollowing modification (28). Briefly, cells were washed with PBS twice,resuspended in PBS containing 1–5 �M of CFSE, incubated at 37°C for 5–10min, and the reaction stopped by addition of 1 ml of prewarmed 10% FBS inPBS into each tube to remove any unbound CFSE. Two � 106 cells/well werethen washed twice with 1% FBS in PBS and plated in a 24-well plate. Tumorlysates obtained previously by four repeated freeze/thaw cycles using liquidnitrogen and a 37°C water bath, and stored frozen in 100 �l aliquot at �80°C,were thawed and centrifuged at 1,200 rpm as the source of tumor antigen.Tumor lysate was added to each well at a final concentration of 10 �g/ml ofprotein, and the cells were collected at 20 and 50 h after incubation. Cells werestained with PE-conjugated anti-CD3e to stain CD3� T cells. The effect oftumor cell lysate stimulation on the proliferation of CD3� T cells was deter-mined by FACS analysis. If T-cell proliferation was observed, the CFSE vitaldye was decreased in the cell progeny.
Detection of Cytokines by Real-Time PCR. Tumors were removed atdays 9, 12, and 15 after tumor implantation for real-time PCR analysis ofinfiltrating lymphocytes. For these studies, single cell suspensions of tumorswere incubated in tissue culture flasks containing RPMI 1640 and 10% FCSfor 3 h at 37°C to separate nonattached lymphocytes from tumor cells. Primersfor selected cytokine were designed by software provided by the ABI PRISM7900HT Sequence Detection System (Applied Biosystems, Foster City, CA).Briefly, total lymphocyte RNA was extracted by Trizol (Life Technologies,Inc., Rockville, MD) following by treated with DNase I (Ambion, Inc, Austin,TX) to eliminate possible genomic DNA contamination, and 1 �g of total RNAwas reverse transcribed into cDNA using a SuperScript cDNA synthesis kit(Life Technologies). The remaining DNA was removed by a DNA-free kit(Ambion, Inc.) according to the manufacturer’s protocol. The real-time PCRreaction mixture (20 �l reaction) consisted of 5 �l of cDNA, 10 �l of SYBRGreen Master Mix (Applied Biosystems), 2 �l of primers (3.3 �M), and 1 �l2 Unpublished observations.
8385
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
of water. The PCR reaction was performed for 30 cycles, and the quantity ofcytokines (IL-2, IL-10, IL-4, IFN�, transforming growth factor b1, and TNF�)was detected by the ABI PRISM 7900HT Sequence Detection System (Ap-plied Biosystems).
RESULTS
Combination LEC/chTNT-3 Immunotherapy and T-Cell SubsetDepletion in Colon 26 Tumor-Bearing Mice
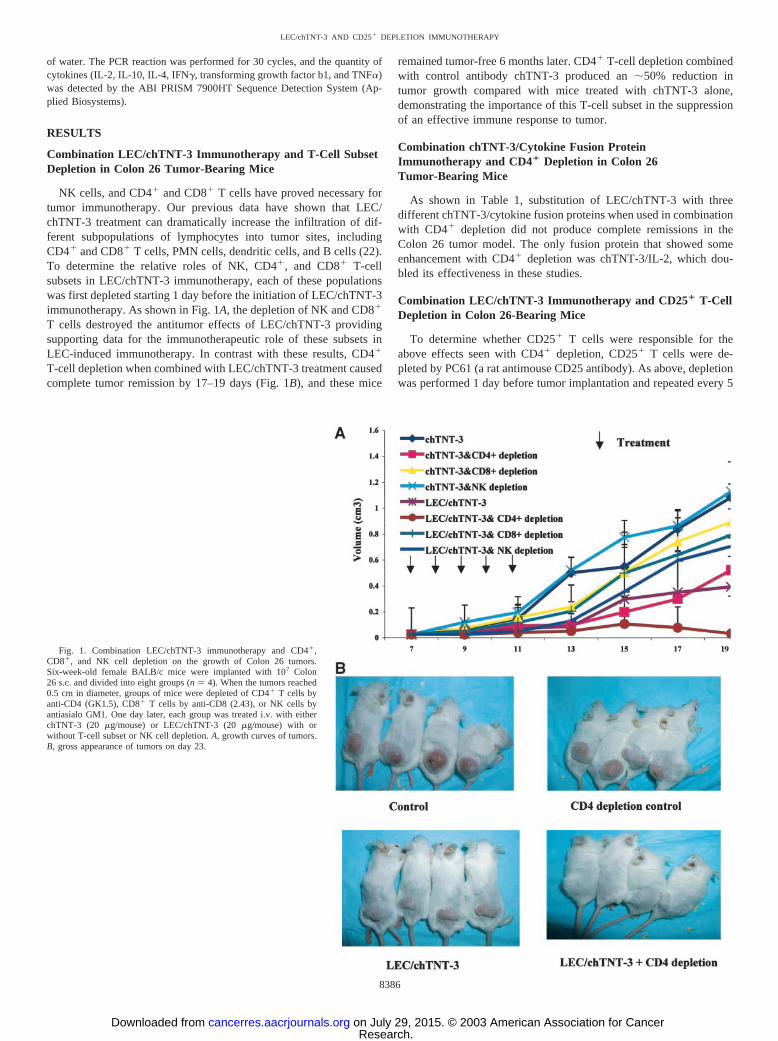
NK cells, and CD4� and CD8� T cells have proved necessary fortumor immunotherapy. Our previous data have shown that LEC/chTNT-3 treatment can dramatically increase the infiltration of dif-ferent subpopulations of lymphocytes into tumor sites, includingCD4� and CD8� T cells, PMN cells, dendritic cells, and B cells (22).To determine the relative roles of NK, CD4�, and CD8� T-cellsubsets in LEC/chTNT-3 immunotherapy, each of these populationswas first depleted starting 1 day before the initiation of LEC/chTNT-3immunotherapy. As shown in Fig. 1A, the depletion of NK and CD8�
T cells destroyed the antitumor effects of LEC/chTNT-3 providingsupporting data for the immunotherapeutic role of these subsets inLEC-induced immunotherapy. In contrast with these results, CD4�
T-cell depletion when combined with LEC/chTNT-3 treatment causedcomplete tumor remission by 17–19 days (Fig. 1B), and these mice
remained tumor-free 6 months later. CD4� T-cell depletion combinedwith control antibody chTNT-3 produced an �50% reduction intumor growth compared with mice treated with chTNT-3 alone,demonstrating the importance of this T-cell subset in the suppressionof an effective immune response to tumor.
Combination chTNT-3/Cytokine Fusion ProteinImmunotherapy and CD4� Depletion in Colon 26Tumor-Bearing Mice
As shown in Table 1, substitution of LEC/chTNT-3 with threedifferent chTNT-3/cytokine fusion proteins when used in combinationwith CD4� depletion did not produce complete remissions in theColon 26 tumor model. The only fusion protein that showed someenhancement with CD4� depletion was chTNT-3/IL-2, which dou-bled its effectiveness in these studies.
Combination LEC/chTNT-3 Immunotherapy and CD25� T-CellDepletion in Colon 26-Bearing Mice
To determine whether CD25� T cells were responsible for theabove effects seen with CD4� depletion, CD25� T cells were de-pleted by PC61 (a rat antimouse CD25 antibody). As above, depletionwas performed 1 day before tumor implantation and repeated every 5
Fig. 1. Combination LEC/chTNT-3 immunotherapy and CD4�,CD8�, and NK cell depletion on the growth of Colon 26 tumors.Six-week-old female BALB/c mice were implanted with 107 Colon26 s.c. and divided into eight groups (n � 4). When the tumors reached0.5 cm in diameter, groups of mice were depleted of CD4� T cells byanti-CD4 (GK1.5), CD8� T cells by anti-CD8 (2.43), or NK cells byantiasialo GM1. One day later, each group was treated i.v. with eitherchTNT-3 (20 �g/mouse) or LEC/chTNT-3 (20 �g/mouse) with orwithout T-cell subset or NK cell depletion. A, growth curves of tumors.B, gross appearance of tumors on day 23.
8386
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
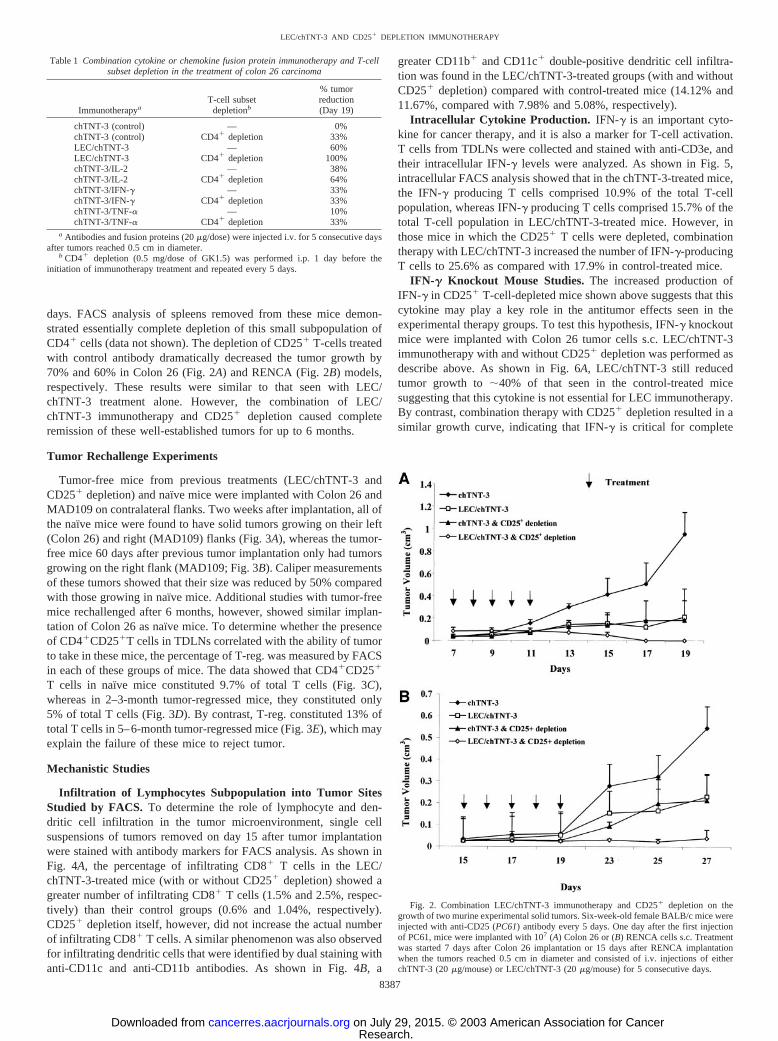
days. FACS analysis of spleens removed from these mice demon-strated essentially complete depletion of this small subpopulation ofCD4� cells (data not shown). The depletion of CD25� T-cells treatedwith control antibody dramatically decreased the tumor growth by70% and 60% in Colon 26 (Fig. 2A) and RENCA (Fig. 2B) models,respectively. These results were similar to that seen with LEC/chTNT-3 treatment alone. However, the combination of LEC/chTNT-3 immunotherapy and CD25� depletion caused completeremission of these well-established tumors for up to 6 months.
Tumor Rechallenge Experiments
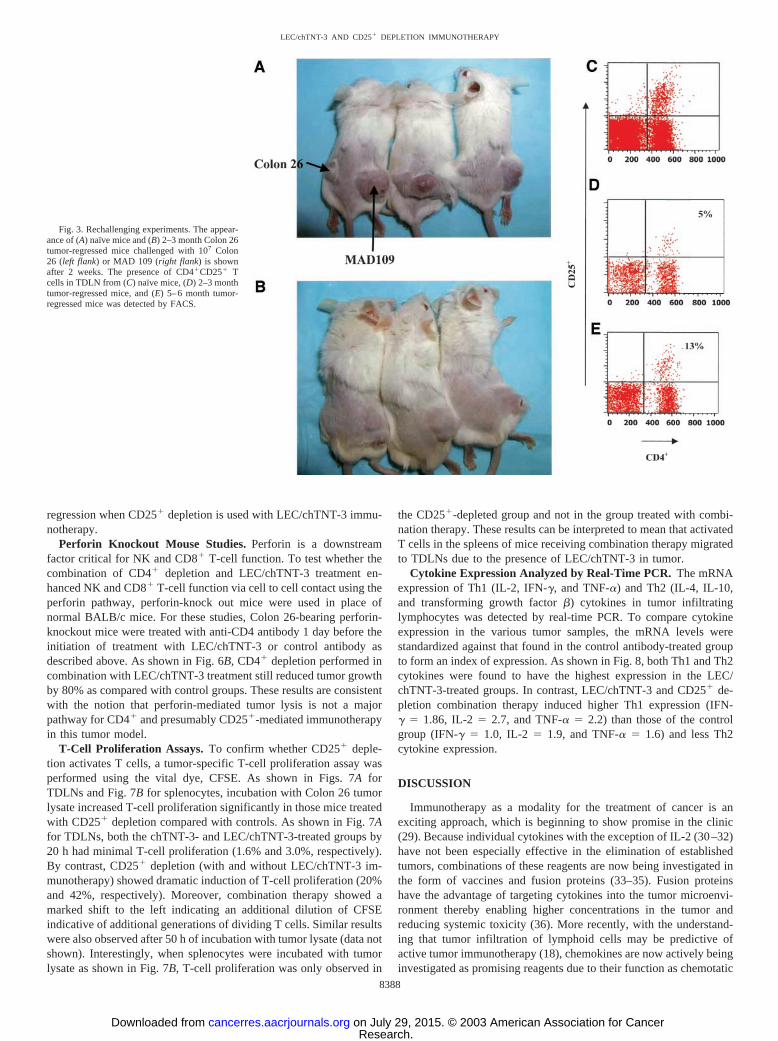
Tumor-free mice from previous treatments (LEC/chTNT-3 andCD25� depletion) and naı̈ve mice were implanted with Colon 26 andMAD109 on contralateral flanks. Two weeks after implantation, all ofthe naı̈ve mice were found to have solid tumors growing on their left(Colon 26) and right (MAD109) flanks (Fig. 3A), whereas the tumor-free mice 60 days after previous tumor implantation only had tumorsgrowing on the right flank (MAD109; Fig. 3B). Caliper measurementsof these tumors showed that their size was reduced by 50% comparedwith those growing in naı̈ve mice. Additional studies with tumor-freemice rechallenged after 6 months, however, showed similar implan-tation of Colon 26 as naı̈ve mice. To determine whether the presenceof CD4�CD25�T cells in TDLNs correlated with the ability of tumorto take in these mice, the percentage of T-reg. was measured by FACSin each of these groups of mice. The data showed that CD4�CD25�
T cells in naı̈ve mice constituted 9.7% of total T cells (Fig. 3C),whereas in 2–3-month tumor-regressed mice, they constituted only5% of total T cells (Fig. 3D). By contrast, T-reg. constituted 13% oftotal T cells in 5–6-month tumor-regressed mice (Fig. 3E), which mayexplain the failure of these mice to reject tumor.
Mechanistic Studies
Infiltration of Lymphocytes Subpopulation into Tumor SitesStudied by FACS. To determine the role of lymphocyte and den-dritic cell infiltration in the tumor microenvironment, single cellsuspensions of tumors removed on day 15 after tumor implantationwere stained with antibody markers for FACS analysis. As shown inFig. 4A, the percentage of infiltrating CD8� T cells in the LEC/chTNT-3-treated mice (with or without CD25� depletion) showed agreater number of infiltrating CD8� T cells (1.5% and 2.5%, respec-tively) than their control groups (0.6% and 1.04%, respectively).CD25� depletion itself, however, did not increase the actual numberof infiltrating CD8� T cells. A similar phenomenon was also observedfor infiltrating dendritic cells that were identified by dual staining withanti-CD11c and anti-CD11b antibodies. As shown in Fig. 4B, a
greater CD11b� and CD11c� double-positive dendritic cell infiltra-tion was found in the LEC/chTNT-3-treated groups (with and withoutCD25� depletion) compared with control-treated mice (14.12% and11.67%, compared with 7.98% and 5.08%, respectively).
Intracellular Cytokine Production. IFN-� is an important cyto-kine for cancer therapy, and it is also a marker for T-cell activation.T cells from TDLNs were collected and stained with anti-CD3e, andtheir intracellular IFN-� levels were analyzed. As shown in Fig. 5,intracellular FACS analysis showed that in the chTNT-3-treated mice,the IFN-� producing T cells comprised 10.9% of the total T-cellpopulation, whereas IFN-� producing T cells comprised 15.7% of thetotal T-cell population in LEC/chTNT-3-treated mice. However, inthose mice in which the CD25� T cells were depleted, combinationtherapy with LEC/chTNT-3 increased the number of IFN-�-producingT cells to 25.6% as compared with 17.9% in control-treated mice.
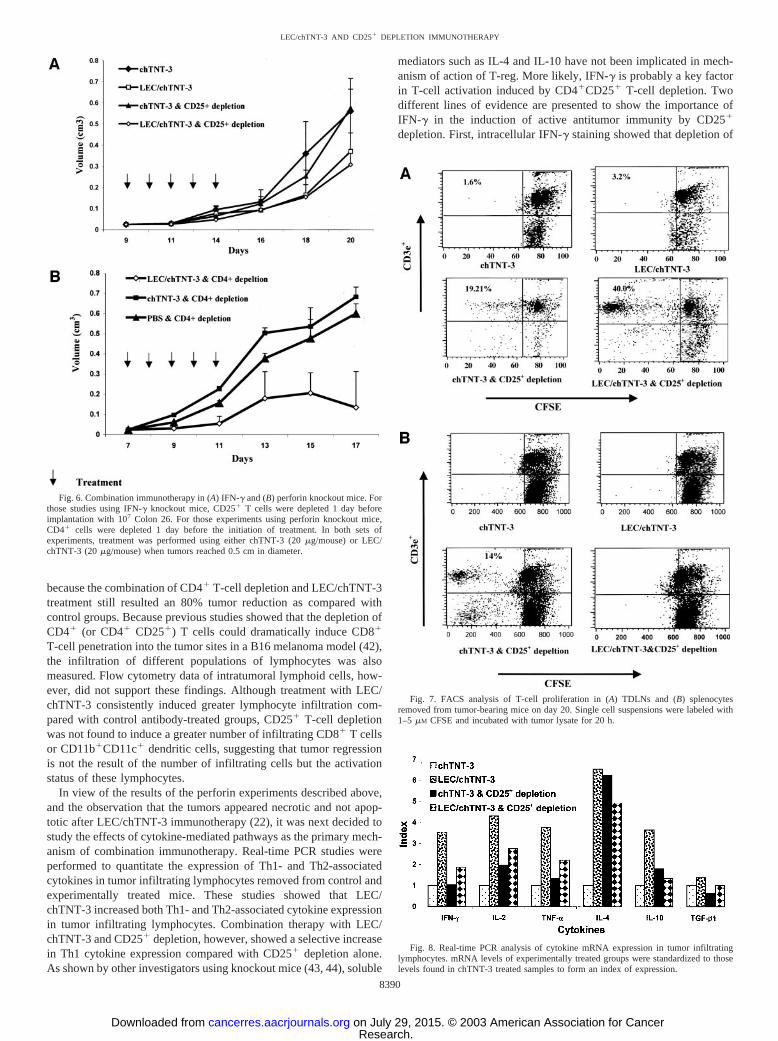
IFN-� Knockout Mouse Studies. The increased production ofIFN-� in CD25� T-cell-depleted mice shown above suggests that thiscytokine may play a key role in the antitumor effects seen in theexperimental therapy groups. To test this hypothesis, IFN-� knockoutmice were implanted with Colon 26 tumor cells s.c. LEC/chTNT-3immunotherapy with and without CD25� depletion was performed asdescribe above. As shown in Fig. 6A, LEC/chTNT-3 still reducedtumor growth to �40% of that seen in the control-treated micesuggesting that this cytokine is not essential for LEC immunotherapy.By contrast, combination therapy with CD25� depletion resulted in asimilar growth curve, indicating that IFN-� is critical for complete
Table 1 Combination cytokine or chemokine fusion protein immunotherapy and T-cellsubset depletion in the treatment of colon 26 carcinoma
ImmunotherapyaT-cell subset
depletionb
% tumorreduction(Day 19)
chTNT-3 (control) — 0%chTNT-3 (control) CD4� depletion 33%LEC/chTNT-3 — 60%LEC/chTNT-3 CD4� depletion 100%chTNT-3/IL-2 — 38%chTNT-3/IL-2 CD4� depletion 64%chTNT-3/IFN-� — 33%chTNT-3/IFN-� CD4� depletion 33%chTNT-3/TNF-� — 10%chTNT-3/TNF-� CD4� depletion 33%
a Antibodies and fusion proteins (20 �g/dose) were injected i.v. for 5 consecutive daysafter tumors reached 0.5 cm in diameter.
b CD4� depletion (0.5 mg/dose of GK1.5) was performed i.p. 1 day before theinitiation of immunotherapy treatment and repeated every 5 days.
Fig. 2. Combination LEC/chTNT-3 immunotherapy and CD25� depletion on thegrowth of two murine experimental solid tumors. Six-week-old female BALB/c mice wereinjected with anti-CD25 (PC61) antibody every 5 days. One day after the first injectionof PC61, mice were implanted with 107 (A) Colon 26 or (B) RENCA cells s.c. Treatmentwas started 7 days after Colon 26 implantation or 15 days after RENCA implantationwhen the tumors reached 0.5 cm in diameter and consisted of i.v. injections of eitherchTNT-3 (20 �g/mouse) or LEC/chTNT-3 (20 �g/mouse) for 5 consecutive days.
8387
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
regression when CD25� depletion is used with LEC/chTNT-3 immu-notherapy.
Perforin Knockout Mouse Studies. Perforin is a downstreamfactor critical for NK and CD8� T-cell function. To test whether thecombination of CD4� depletion and LEC/chTNT-3 treatment en-hanced NK and CD8� T-cell function via cell to cell contact using theperforin pathway, perforin-knock out mice were used in place ofnormal BALB/c mice. For these studies, Colon 26-bearing perforin-knockout mice were treated with anti-CD4 antibody 1 day before theinitiation of treatment with LEC/chTNT-3 or control antibody asdescribed above. As shown in Fig. 6B, CD4� depletion performed incombination with LEC/chTNT-3 treatment still reduced tumor growthby 80% as compared with control groups. These results are consistentwith the notion that perforin-mediated tumor lysis is not a majorpathway for CD4� and presumably CD25�-mediated immunotherapyin this tumor model.
T-Cell Proliferation Assays. To confirm whether CD25� deple-tion activates T cells, a tumor-specific T-cell proliferation assay wasperformed using the vital dye, CFSE. As shown in Figs. 7A forTDLNs and Fig. 7B for splenocytes, incubation with Colon 26 tumorlysate increased T-cell proliferation significantly in those mice treatedwith CD25� depletion compared with controls. As shown in Fig. 7Afor TDLNs, both the chTNT-3- and LEC/chTNT-3-treated groups by20 h had minimal T-cell proliferation (1.6% and 3.0%, respectively).By contrast, CD25� depletion (with and without LEC/chTNT-3 im-munotherapy) showed dramatic induction of T-cell proliferation (20%and 42%, respectively). Moreover, combination therapy showed amarked shift to the left indicating an additional dilution of CFSEindicative of additional generations of dividing T cells. Similar resultswere also observed after 50 h of incubation with tumor lysate (data notshown). Interestingly, when splenocytes were incubated with tumorlysate as shown in Fig. 7B, T-cell proliferation was only observed in
the CD25�-depleted group and not in the group treated with combi-nation therapy. These results can be interpreted to mean that activatedT cells in the spleens of mice receiving combination therapy migratedto TDLNs due to the presence of LEC/chTNT-3 in tumor.
Cytokine Expression Analyzed by Real-Time PCR. The mRNAexpression of Th1 (IL-2, IFN-�, and TNF-�) and Th2 (IL-4, IL-10,and transforming growth factor �) cytokines in tumor infiltratinglymphocytes was detected by real-time PCR. To compare cytokineexpression in the various tumor samples, the mRNA levels werestandardized against that found in the control antibody-treated groupto form an index of expression. As shown in Fig. 8, both Th1 and Th2cytokines were found to have the highest expression in the LEC/chTNT-3-treated groups. In contrast, LEC/chTNT-3 and CD25� de-pletion combination therapy induced higher Th1 expression (IFN-� � 1.86, IL-2 � 2.7, and TNF-� � 2.2) than those of the controlgroup (IFN-� � 1.0, IL-2 � 1.9, and TNF-� � 1.6) and less Th2cytokine expression.
DISCUSSION
Immunotherapy as a modality for the treatment of cancer is anexciting approach, which is beginning to show promise in the clinic(29). Because individual cytokines with the exception of IL-2 (30–32)have not been especially effective in the elimination of establishedtumors, combinations of these reagents are now being investigated inthe form of vaccines and fusion proteins (33–35). Fusion proteinshave the advantage of targeting cytokines into the tumor microenvi-ronment thereby enabling higher concentrations in the tumor andreducing systemic toxicity (36). More recently, with the understand-ing that tumor infiltration of lymphoid cells may be predictive ofactive tumor immunotherapy (18), chemokines are now actively beinginvestigated as promising reagents due to their function as chemotatic
Fig. 3. Rechallenging experiments. The appear-ance of (A) naı̈ve mice and (B) 2–3 month Colon 26tumor-regressed mice challenged with 107 Colon26 (left flank) or MAD 109 (right flank) is shownafter 2 weeks. The presence of CD4�CD25� Tcells in TDLN from (C) naı̈ve mice, (D) 2–3 monthtumor-regressed mice, and (E) 5–6 month tumor-regressed mice was detected by FACS.
8388
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

agents. In many regards, chemokine fusion proteins are ideal thera-peutics because they target the chemoattractant into the tumor, therebyfacilitating the intratumoral infiltration of immune cells. To investi-gate the therapeutic potential of this approach, Challita-Eid et al. (37)and our laboratory (Li et al.; Ref. 22) have generated RANTES andLEC fusion proteins, respectively, using different tumor targetingmAbs. Although the RANTES fusion protein was not tested in vivo(37), our studies with LEC/chTNT-3 showed marked reduction intumor size recently in three experimental solid tumor models of theBALB/c mouse (22). In the present study, we now show that completeremission can be obtained when LEC/chTNT-3 immunotherapy isused in combination with either CD4� or CD4�CD25� T-cell deple-tion. Treatment with control antibody chTNT-3 and CD25� T-celldepletion produced only a 20% remission rate in Colon 26-bearingmice and a 0% remission rate in RENCA-bearing mice. Combinationsof CD4� depletion and other cytokine fusion proteins such as thoseconsisting of IL-2, TNF�, and IFN-�, were next tested to determinewhether these treatments had synergistic effects on tumor growth. Asshown in Table 1, two of the three fusion proteins, chTNT-3/IL-2 andchTNT-3/TNF�, showed some improvement with CD4� depletion,but these results were not as dramatic as those with LEC/chTNT-3,suggesting that complete remission required LEC/chTNT-3 localiza-tion to tumor. One difference between the LEC fusion protein and thecytokine fusion proteins is the location of the cytokine moiety on the
antibody molecule. As described in “Materials and Methods” above,LEC was attached to the NH2 terminus near the variable region of theheavy chain, whereas the other cytokines were attached to the COOHterminus distal to the Fc region. These fusion proteins were con-structed in this manner to insure that the active binding sites of thesecytokines or chemokines were distal to the linkage site enabling themto bind to their respective receptors. Construction in the reversemanner might compromise the full activity of the chemokine orcytokine making it difficult to determine whether this difference maybe in some way responsible for observed results. Finally, it shouldalso be noted that studies conducted by Golgher et al. (38) showed amuch higher frequency of Colon 26 tumor rejection (70%) by CD25�
T-cell depletion alone, a finding that may be explained by their use ofa much smaller inoculum of tumor cells compared with our studies(5 � 104/mouse versus 107 cells tumor cells) 1 day after the initiationof T-cell depletion.
T-reg. was first identified by Hill et al. (2), who showed that theadministration of anti-CD4 antibody significantly increased the anti-tumor effects by mAb therapy, radiation therapy, and chemotherapy invarious tumor models (3). Sakaguchi et al. (39) later showed that theCD25� subset of CD4� was responsible for the immunoregulation ofthe antitumor response in immunocompetent mice. Our studies verifythese results and show for the first time that the combination of achemokine fusion protein and CD25� depletion has the potential toproduce complete remissions in well-established solid tumors. Re-challenge experiments in naı̈ve and cured mice showed that memorywas intact for up to 3 months after completion of immunotherapy andthat the return of CD25� T-reg. coincided with the loss of tumorrecognition as shown by successful implantation of tumor at 6 months.During CD25� depletion, we found that the immune response wasboth tumor-specific and nonspecific because implanted MAD109tumors in the contralateral flank were found to grow much slower thanthose implanted in naı̈ve controls. These results were consistent withthose of Murakami et al. and Shevach et al. (40, 41) who showed thatthe presence of CD4�CD25� T cells inhibits the proliferation ofCD8� memory cells in the C57BL/6 mouse model.
The suppressive mechanisms mediated through CD4�CD25� Tcells still remain unknown. Our data using perforin knockout miceindicate that combination therapy was not associated with perforin
Fig. 5. FACS analysis of intracellular IFN-� expression in TDLNs. Single cellsuspensions of TDLNs were obtained 20 days after tumor implantation from treated andcontrol groups of mice. The cells were stimulated with 10 ng/ml of PMA and 100 ng/mlof ionomycine in the presence of GolgiStop for 4–6 h before being double stained withPE-anti-CD3e and FITC-anti-IFN-� for FACS analysis.
Fig. 4. FACS analysis of tumor infiltrating lymphocytes in control and experimentallytreated tumors. Single cell suspensions of tumors removed on day 15 after tumorimplantation were stained with (A) anti-CD8 and anti-PMN or (B) anti-CD11b andanti-CD11c to quantitate the presence of T-cells, PMNs, and dendritic cells by flowcytometry.
8389
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
because the combination of CD4� T-cell depletion and LEC/chTNT-3treatment still resulted an 80% tumor reduction as compared withcontrol groups. Because previous studies showed that the depletion ofCD4� (or CD4� CD25�) T cells could dramatically induce CD8�
T-cell penetration into the tumor sites in a B16 melanoma model (42),the infiltration of different populations of lymphocytes was alsomeasured. Flow cytometry data of intratumoral lymphoid cells, how-ever, did not support these findings. Although treatment with LEC/chTNT-3 consistently induced greater lymphocyte infiltration com-pared with control antibody-treated groups, CD25� T-cell depletionwas not found to induce a greater number of infiltrating CD8� T cellsor CD11b�CD11c� dendritic cells, suggesting that tumor regressionis not the result of the number of infiltrating cells but the activationstatus of these lymphocytes.
In view of the results of the perforin experiments described above,and the observation that the tumors appeared necrotic and not apop-totic after LEC/chTNT-3 immunotherapy (22), it was next decided tostudy the effects of cytokine-mediated pathways as the primary mech-anism of combination immunotherapy. Real-time PCR studies wereperformed to quantitate the expression of Th1- and Th2-associatedcytokines in tumor infiltrating lymphocytes removed from control andexperimentally treated mice. These studies showed that LEC/chTNT-3 increased both Th1- and Th2-associated cytokine expressionin tumor infiltrating lymphocytes. Combination therapy with LEC/chTNT-3 and CD25� depletion, however, showed a selective increasein Th1 cytokine expression compared with CD25� depletion alone.As shown by other investigators using knockout mice (43, 44), soluble
mediators such as IL-4 and IL-10 have not been implicated in mech-anism of action of T-reg. More likely, IFN-� is probably a key factorin T-cell activation induced by CD4�CD25� T-cell depletion. Twodifferent lines of evidence are presented to show the importance ofIFN-� in the induction of active antitumor immunity by CD25�
depletion. First, intracellular IFN-� staining showed that depletion of
Fig. 6. Combination immunotherapy in (A) IFN-� and (B) perforin knockout mice. Forthose studies using IFN-� knockout mice, CD25� T cells were depleted 1 day beforeimplantation with 107 Colon 26. For those experiments using perforin knockout mice,CD4� cells were depleted 1 day before the initiation of treatment. In both sets ofexperiments, treatment was performed using either chTNT-3 (20 �g/mouse) or LEC/chTNT-3 (20 �g/mouse) when tumors reached 0.5 cm in diameter.
Fig. 7. FACS analysis of T-cell proliferation in (A) TDLNs and (B) splenocytesremoved from tumor-bearing mice on day 20. Single cell suspensions were labeled with1–5 �M CFSE and incubated with tumor lysate for 20 h.
Fig. 8. Real-time PCR analysis of cytokine mRNA expression in tumor infiltratinglymphocytes. mRNA levels of experimentally treated groups were standardized to thoselevels found in chTNT-3 treated samples to form an index of expression.
8390
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

T-reg. could significantly induce the production of IFN-� producing Tcells in TDLNs. Secondly, studies performed in IFN-� knockout miceshowed that the therapeutic effects of CD25� depletion no longerenhanced LEC/chTNT-3 immunotherapy. By contrast, LEC/chTNT-3immunotherapy alone was only slightly affected in IFN-� knockoutmice (40% tumor reduction versus 60% in normal mice) in theseexperiments.
Studies conducted by others (16, 45) indicate that T-reg. cells arepotent suppressors of activated T cells, including CD4�CD25� andCD8� T cells. In these studies, nonspecific in vitro stimulation withCD3 and IL-2 was used to demonstrate this suppression. Contrast-ingly, our investigations used specific stimulation (tumor lysates) toactivate TDLNs and splenocytes ex vivo. By these methods, weobserved that T-cell proliferation in TDLNs was markedly enhancedin those mice receiving CD25� T-cell depletion with and withoutLEC/chTNT-3 immunotherapy. These results are consistent with thefinding that CD4�CD25� T-reg. from cancer patients suppresses theproliferation of T cells (46). In contrast, splenocytes from treated miceshowed enhanced T-cell proliferation when mice were treated withCD25� depletion alone. These data provide convincing evidence thatCD25� depletion causes T-cell proliferation in all of the lymphoidorgans of the mouse but when used in combination with LEC/chTNT-3, T-cell proliferation occurs specifically in tumor and TDLNswhere tumor antigens are present or sequestered, respectively.
Because T-reg. has been shown to be important for the suppressionof autoimmune disease (47–51), it is a significant finding that CD25�
depletion caused T-cell proliferation only in TDLNs and presumablytumor when used in combination with LEC/chTNT-3. In fact, none ofthe treated mice showed any signs of toxicity by either LEC/chTNT-3therapy or combination therapy with CD25� depletion. In support ofthese findings, increased levels of CD4�CD25� T cells are now beingreported in the circulation of melanoma, pancreas, and breast adeno-carcinoma cancer patients (52–54). These early studies suggest thatT-reg. may be suppressing an effective immune response against thesetumors. If so, then depletion of T-reg. may be useful for the immu-notherapy of cancer if it can be directed to the tumor site and notnonspecifically to normal tissues. Chemokines in general are idealcandidates for controlling tissue-specific lymphocyte trafficking andhoming, because chemokines and their receptors help control thespecific migration of memory lymphocyte subsets by CCL17/CCR4and CCL27/CCR10 interactions (55–58). LEC is a particularly attrac-tive immune modulator, because it has been shown that CCR1 andCCR8 are the active receptors for LEC, both of which are expressedon resting and activated lymphocytes (59). In this regard, LEC/chTNT-3 may be an effective treatment reagent for attracting themigration and activation of immune effector cells into tumor andTDLNs where an effective and specific antitumor response can bedirected in the absence of T-reg.
ACKNOWLEDGMENTS
We thank Drs. Stephen Stohlman, Norman Martin, and Dixon Gray atUniversity of Southern California Keck School of Medicine for their technicalsupport and discussion of the data. We also acknowledge the expert technicalassistance of Aoyun Yun, Jingzhong Pang, and Sam Kim.
REFERENCES
1. Mills, C. D., and North, R. J. Ly-1�2- suppressor T cells inhibit the expression ofpassively transferred antitumor immunity by suppressing the generation of cytolyticT cells. Transplantation, 39: 202–208, 1985.
2. Hill, J. O., Awwad, M., and North, R. J. Elimination of CD4� suppressor T cells fromsusceptible BALB/c mice releases CD8� T lymphocytes to mediate protective im-munity against Leishmania. J. Exp. Med., 169: 1819–1827, 1989.
3. Awwad, M., and North, R. J. Radiosensitive barrier to T-cell-mediated adoptiveimmunotherapy of established tumors. Cancer Res., 50: 2228–2233, 1990.
4. Shevach, E. M. CD4� CD25� suppressor T cells: more questions than answers. Nat.Rev. Immunol., 2: 389–400, 2002.
5. Takahashi, T., Tagami, T., Yamazaki, S., Uede, T., Shimizu, J., Sakaguchi, N., Mak,T. W., and Sakaguchi, S. Immunologic self-tolerance maintained byCD25(�)CD4(�) regulatory T cells constitutively expressing cytotoxic T lympho-cyte-associated antigen 4. J. Exp. Med., 192: 303–310, 2000.
6. Weiner, H. L. Oral tolerance for the treatment of autoimmune diseases. Ann. Rev.Med., 48: 341–351, 1997.
7. Ke, Y., and Kapp, J. A. Oral antigen inhibits priming of CD8� CTL, CD4� T cells,and antibody responses while activating CD8� suppressor T cells. J. Immunol., 156:916–921, 1996.
8. Grdic, D., Hornquist, E., Kjerrulf, M., and Lycke, N. Y. Lack of local suppression inorally tolerant CD8-deficient mice reveals a critical regulatory role of CD8� T cellsin the normal gut mucosa. J. Immunol., 160: 754–762, 1998.
9. Cosmi, L., Liotta, F., Lazzeri, E., Francalanci, M., Angeli, R., Mazzinghi, B.,Santarlasci, V., Manetti, R., Vanini, V., Romagnani, P., Maggi, E., Romagnani, S.,and Annunziato, F. Human CD8�CD25� thymocytes sharing phenotypic and func-tional features with CD4�CD25� regulatory thymocytes. Blood, in press, 2003.
10. McHugh, R. S., and Shevach, E. M. The role of suppressor T cells in regulation ofimmune responses. J. Allergy Clin. Immunol., 110: 693–702, 2002.
11. Annunziato, F., Cosmi, L., Liotta, F., Lazzeri, E., Manetti, R., Vanini, V., Romagnani,P., Maggi, E., and Romagnani, S. Phenotype, localization, and mechanism of sup-pression of CD4(�)CD25(�) human thymocytes. J. Exp. Med., 196: 379–387, 2002.
12. Shevach, E. M., McHugh, R. S., Thornton, A. M., Piccirillo, C., Natarajan, K., andMargulies, D. H. Control of autoimmunity by regulatory T cells. Adv. Exp. Med.Biol., 490: 21–32, 2001.
13. Antony, P. A., and Restifo, N. P. Do CD4� CD25� immunoregulatory T cells hindertumor immunotherapy? J. Immunother., 25: 202–206, 2002.
14. Shimizu, J., Yamazaki, S., and Sakaguchi, S. Induction of tumor immunity byremoving CD25�CD4� T cells: a common basis between tumor immunity andautoimmunity. J. Immunol., 163: 5211–5218, 1999.
15. Tanaka, H., Tanaka, J., Kjaergaard, J., and Shu, S. Depletion of CD4� CD25�
regulatory cells augments the generation of specific immune T cells in tumor-draininglymph nodes. J. Immunother., 25: 207–217, 2002.
16. Piccirillo, C. A., and Shevach, E. M. Cutting edge: control of CD8� T cell activationby CD4�CD25� immunoregulatory cells. J. Immunol., 167: 1137–1140, 2001.
17. Onizuka, S., Tawara, I., Shimizu, J., Sakaguchi, S., Fujita, T., and Nakayama, E.Tumor rejection by in vivo administration of anti-CD25 (interleukin-2 receptor �)monoclonal antibody. Cancer Res., 59: 3128–3133, 1999.
18. Clark, W. H., Jr., Elder, D. E., Guerry, D. t., Braitman, L. E., Trock, B. J., Schultz,D., Synnestvedt, M., and Halpern, A. C. Model predicting survival in stage Imelanoma based on tumor progression. J. Natl. Cancer Inst., 81: 1893–1904, 1989.
19. Warnock, R. A., Campbell, J. J., Dorf, M. E., Matsuzawa, A., McEvoy, L. M., andButcher, E. C. The role of chemokines in the microenvironmental control of T versusB cell arrest in Peyer’s patch high endothelial venules. J. Exp. Med., 191: 77–88,2000.
20. Reiss, Y., Proudfoot, A. E., Power, C. A., Campbell, J. J., and Butcher, E. C. CCchemokine receptor (CCR)4 and the CCR10 ligand cutaneous T cell-attractingchemokine (CTACK) in lymphocyte trafficking to inflamed skin. J. Exp. Med., 194:1541–1547, 2001.
21. Johnston, B., and Butcher, E. C. Chemokines in rapid leukocyte adhesion triggeringand migration. Semin. Immunol., 14: 83–92, 2002.
22. Li, J., Hu, P., Kwali, L., and Epstein, A. LEC/chTNT-3 fusion protein for theimmunotherapy of experimental solid tumors. J. Immunother., 26: 320–331, 2003.
23. Naruse, K., Ueno, M., Satoh, T., Nomiyam, H., Tei, H., Takeda, M., Ledbetter, D.,Coillie, V., Opdenakker, G., Gunge, N., Sakaki, Y., and Mura, R. A YAC contig ofthe human CC chemokine genes clustered on chromosome 17q11.2. Genomics, 34:236–240, 1996.
24. Hornick, J. L., Hu, P., Khawli, L. A., Biola, B. H., Yun, A., Sharifi, J., Taylor, C. R.,and Epstein, A. L. chTNT-3/B, a new chemically modified chimeric monoclonalantibody directed against DNA for the tumor necrosis treatment of solid tumors.Cancer Biother. Radiopharm., 13: 255–268, 1998.
25. Bergmann, C. C., Parra, B., Hinton, D. R., Chandran, R., Morrison, M., and Stohlman,S. A. Perforin-mediated effector function within the central nervous system requiresIFN-�-mediated MHC up-regulation. J. Immunol., 170: 3204–3213, 2003.
26. Hornick, J. L., Khawli, L. A., Hu, P., Sharifi, J., Khanna, C., and Epstein, A. L.Pretreatment with a monoclonal antibody/interleukin-2 fusion protein directed againstDNA enhances the delivery of therapeutic molecules to solid tumors. Clin. CancerRes., 5: 51–60, 1999.
27. Mizokami, M. M., Hu, P., Khawli, L. A., and Epstein, A. L. Chimeric TNT-3antibody-murine-interferon-g fusion protein for the immunotherapy of solid malig-nancies. Hybridoma Hybridomics, 22: 197–207, 2003.
28. Weston, S. A., and Parish, C. R. New fluorescent dyes for lymphocyte migrationstudies. Analysis by flow cytometry and fluorescence microscopy. J. Immunol.Methods, 133: 87–97, 1990.
29. Lode, H. N., and Reisfeld, R. A. Targeted cytokines for cancer immunotherapy.Immunol. Res., 21: 279–288, 2000.
30. Rosenberg, S. A. Interleukin-2 and the development of immunotherapy for thetreatment of patients with cancer. Cancer J. Sci. Am., 6(Suppl. 1): S2–S7, 2000.
31. Atkins, M. B. Interleukin-2 in metastatic melanoma: what is the current role? CancerJ. Sci. Am., 6(Suppl. 1): S8–S10, 2000.
32. Atkins, M. B., Kunkel, L., Sznol, M., and Rosenberg, S. A. High-dose recombinantinterleukin-2 therapy in patients with metastatic melanoma: long-term survival up-date. Cancer J. Sci. Am., 6(Suppl. 1): S11–S14, 2000.
8391
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

33. Gillies, S. D., Lan, Y., Brunkhorst, B., Wong, W. K., Li, Y., and Lo, K. M.Bi-functional cytokine fusion proteins for gene therapy and antibody-targeted treat-ment of cancer. Cancer Immunol. Immunother., 51: 449–460, 2002.
34. Lode, H. N., Moehler, T., Xiang, R., Jonczyk, A., Gillies, S. D., Cheresh, D. A., andReisfeld, R. A. Synergy between an antiangiogenic integrin �v antagonist and anantibody-cytokine fusion protein eradicates spontaneous tumor metastases. Proc.Natl. Acad. Sci. USA, 96: 1591–1596, 1999.
35. Penichet, M. L., and Morrison, S. L. Antibody-cytokine fusion proteins for thetherapy of cancer. J. Immunol. Methods, 248: 91–101, 2001.
36. Schrama, D., Straten, P., Brocker, E. B., Reisfeld, R. A., and Becker, J. C. Cytokinefusion protein treatment. Recent Results Cancer Res., 160: 185–194, 2002.
37. Challita-Eid, P., Abboud, C., Morrison, S., Penichet, M., Rosell, K., Poles, T.,Hilchey, S., Planelles, V., and Rosenblatt, J. A RANTES-antibody fusion proteinretains antigen specificity and chemokine function. J. Immunol., 161: 3729–3736,1998.
38. Golgher, D., Jones, E., Powrie, F., Elliot, T., and Gallimore, A. Depletion of CD25�
regulatory cells uncovers immune responses to shared murine tumor rejection anti-gens. Eur. J. Immunol., 32: 3267–3275, 2002.
39. Sakaguchi, S., Sakaguchi, N., Asano, M., Itoh, M., and Toda, M. Immunologicself-tolerance maintained by activated T cells expressing IL-2 receptor �-chains(CD25). Breakdown of a single mechanism of self-tolerance causes various autoim-mune diseases. J. Immunol., 155: 1151–1164, 1995.
40. Murakami, M., Sakamoto, A., Bender, J., Kappler, J., and Marrack, P. CD25�CD4�
T cells contribute to the control of memory CD8� T cells. Proc. Natl. Acad. Sci. USA,99: 8832–8837, 2002.
41. Shevach, E. M., McHugh, R. S., Piccirillo, C. A., and Thornton, A. M. Control ofT-cell activation by CD4�CD25� suppressor T cells. Immunol. Rev., 182: 58–67,2001.
42. Nagai, H., Hara, I., Horikawa, T., Oka, M., Kamidono, S., and Ichihashi, M.Elimination of CD4(�) T cells enhances anti-tumor effect of locally secreted inter-leukin-12 on B16 mouse melanoma and induces vitiligo-like coat color alteration.J. Investig. Dermatol., 115: 1059–1064, 2000.
43. Takahashi, T., Kuniyasu, Y., Toda, M., Sakaguchi, N., Itoh, M., Iwata, M., Shimizu,J., and Sakaguchi, S. Immunologic self-tolerance maintained by CD25�CD4� natu-rally anergic and suppressive T cells: Induction of autoimmune disease by breakingtheir anergic/suppressive state. Int. Immunol., 10: 1969–1980, 1998.
44. Kuhn, R., Lohler, J., Rennick, D., Rajewsky, K., and Muller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell, 75: 263–274, 1993.
45. Thornton, A. M., and Shevach, E. M. CD4�CD25� immunoregulatory T cellssuppress polyclonal T cell activation in vitro by inhibiting interleukin-2 production.J. Exp. Med., 188: 287–296, 1998.
46. Woo, E. Y., Yeh, H., Chu, C. S., Schlienger, K., Carroll, R. G., Riley, J. L., Kaiser,L. R., and June, C. H. Cutting edge: Regulatory T cells from lung cancer patientsdirectly inhibit autologous T cell proliferation. J. Immunol., 168: 4272–4276, 2002.
47. McHugh, R. S., Shevach, E. M., and Thornton, A. M. Control of organ-specificautoimmunity by immunoregulatory CD4(�)CD25(�) T cells. Microbes Infect., 3:919–927, 2001.
48. Han, H-S., Jun, H-S., Utsugi, T., and Yoon, J-W. A new type of CD4� suppressor Tcell completely prevents spontaneous autoimmune diabetes and recurrent diabetes insyngeneic islet-transplanted NOD mice. J. Autoimmunity, 9: 331–339, 1996.
49. Van de Keere, F., and Tonegawa, S. CD4� T cells prevent spontaneous experimentalautoimmune encephalomyelitis in anti-myelin basic protein T cell receptor transgenicmice. J. Exp. Med., 188: 1875–1882, 1998.
50. Itoh, M., Takahashi, T., Sakaguchi, N., Kuniyasu, Y., Shimizu, J., Otsuka, F., andSakaguchi, S. Thymus and autoimmunity: production of CD25�CD4� naturallyanergic and suppressive T cells as a key function of the thymus in maintainingimmunologic self-tolerance. J. Immunol., 162: 5317–5326, 1999.
51. Morrissey, P., Charrier, K., Braddy, S., Liggitt, D., and Watson, J. CD4� T cells thatexpress high levels of CD45RB induce wasting disease when transferred into con-genic severe combined immunodeficient mice. Disease development is prevented bycotransfer of purified CD4� T cells. J. Exp. Med., 178: 237–244, 1993.
52. Javia, L. R., and Rosenberg, S. A. CD4�CD25� suppressor lymphocytes in thecirculation of patients immunized against melanoma antigens. J. Immunother., 26:85–93, 2003.
53. Liyanage, U. K., Moore, T. T., Joo, H. G., Tanaka, Y., Herrmann, V., Doherty, G.,Drebin, J. A., Strasberg, S. M., Eberlein, T. J., Goedegebuure, P. S., and Linehan,D. C. Prevalence of regulatory T cells is increased in peripheral blood and tumormicroenvironment of patients with pancreas or breast adenocarcinoma. J. Immunol.,169: 2756–2761, 2002.
54. Woo, E. Y., Chu, C. S., Goletz, T. J., Schlienger, K., Yeh, H., Coukos, G., Rubin,S. C., Kaiser, L. R., and June, C. H. Regulatory CD4�CD25� T cells in tumors frompatients with early-stage non-small cell lung cancer and late-stage ovarian cancer.Cancer Res., 61: 4766–4772, 2001.
55. Campbell, J. J., and Butcher, E. C. Chemokines in tissue-specific and microenviron-ment-specific lymphocyte homing. Curr. Opin. Immunol., 12: 336–341, 2000.
56. Kunkel, E. J., and Butcher, E. C. Chemokines and the tissue-specific migration oflymphocytes. Immunity, 16: 1–4, 2002.
57. Campbell, J. J., Brightling, C. E., Symon, F. A., Qin, S., Murphy, K. E., Hodge, M.,Andrew, D. P., Wu, L., Butcher, E. C., and Wardlaw, A. J. Expression of chemokinereceptors by lung T cells from normal and asthmatic subjects. J. Immunol., 166:2842–2848, 2001.
58. Morales, J., Homey, B., Vicari, A. P., Hudak, S., Oldham, E., Hedrick, J., Orozco, R.,Copeland, N. G., Jenkins, N. A., McEvoy, L. M., and Zlotnik, A. CTACK, askin-associated chemokine that preferentially attracts skin-homing memory T cells.Proc. Natl. Acad. Sci. (USA), 96: 14470–14475, 1999.
59. Howard, O. M. Z., Dong, H. F., Shirakawa, A-K., and Oppenheim, J. J. LEC induceschemotaxis and adhesion by interacting with CCR1 and CCR8. Blood, 96: 840–845,2000.
8392
LEC/chTNT-3 AND CD25� DEPLETION IMMUNOTHERAPY
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

2003;63:8384-8392. Cancer Res Jiali Li, Peisheng Hu, Leslie A. Khawli, et al. T-Cell Depletion
+Combination LEC/chTNT-3 Immunotherapy and CD25Complete Regression of Experimental Solid Tumors by
Updated version
http://cancerres.aacrjournals.org/content/63/23/8384
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/63/23/8384.full.html#ref-list-1
This article cites 50 articles, 29 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/63/23/8384.full.html#related-urls
This article has been cited by 13 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on July 29, 2015. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from




















