
Targeting of Embryonic and Postnatal Autonomic and Enteric Neurons with a Vasoactive Intestinal...
10
Targeting of Embryonic and Postnatal Autonomic and Enteric Neurons with a Vasoactive Intestinal Peptide Transgene James A. Waschek, Dawn T. Bravo, Matthew Sena, Robert Casillas, Williams Rodriguez, Thinh Nguyen, and Steve Colburn Department of Psychiatry and Mental Retardation Research Center, University of California at Los Angeles, Los Angeles, California, U.S.A. Abstract: The neuropeptide vasoactive intestinal peptide (VIP) is expressed in several distinct sites in the CNS, in cholinergic and enteric ganglia, and in a small subpopu- lation of neurons within sympathetic ganglia. Previous studies on the human VIP gene indicate that transcription in neural crest-derived neuroblastoma and pheochromo- cytoma cell lines is controlled in part by multiple regula- tory elements located along 4.5 kb of upstream 59 flank- ing sequence. In the current studies, transgenic mice were created with a chimeric gene consisting of 16.5 kb of the mouse VIP gene fused to the b-galactosidase reporter. In situ hybridization analysis in adult mice indi- cated that reporter gene expression was correctly tar- geted to neurons in the esophagus, stomach, small in- testine, and colon. No expression was observed in the brain, including regions that contain abundant VIP-ex- pressing cells, such as the thalamus, amygdala, cerebral cortex, hippocampus, and suprachiasmatic nucleus. Analysis of transgene expression in neonatal and embry- onic day 13.5 mice revealed a near perfect correlation between VIP and b-galactosidase gene expression in cranial cholinergic ganglia and the superior cervical gan- glia, and lack of transgene expression in sensory ganglia and in nonneuronal tissue. Potential ectopic transgene expression was observed in neonates, in the cerebellar external granule layer and in a small subpopulation of neurons in the olfactory epithelium. We conclude that the 16.5 kb of VIP gene used in these studies contains se- quences sufficient for directing expression specifically to VIP neurons in the PNS, and that sequences located elsewhere on the gene are required for proper CNS ex- pression. The VIP gene sequences used here should be capable of targeting other gene products to specific pop- ulations of embryonic and adult peripheral neurons with- out causing significant expression in the CNS. Key Words: Vasoactive intestinal peptide —Transgenic— Promoter—Cholinergic—Embryo—Neonate. J. Neurochem. 73, 1739 –1748 (1999). Recently evolved transgenic approaches to study the nervous system in mice rely on the ability to target gene expression to precise locations. One strategy to do this is to identify promoter sequences on a gene that is normally expressed in the desired site, and then use the promoter sequences to drive the expression of the gene of interest. For example, the role of CA1 hippocampal NMDA re- ceptors in learning and memory was examined recently using this strategy (Tsien et al., 1996). In this work, the NMDA receptor was specifically disrupted in CA1 neu- rons using a Cre/Lox excision system in which the Cre recombinase was targeted to CA1 neurons using the a-calcium/calmodulin-dependent protein kinase II gene promoter. Despite the potential value of such an ap- proach, it is limited by the availability of promoter se- quences that can target gene expression specifically to the cells of interest. Moreover, it is sometimes difficult to identify effective tissue-specific promoters, because reg- ulatory sequences that enhance or restrict expression may be located far upstream or downstream from coding sequences (for review, see Waschek, 1995). On the other hand, the inclusion or exclusion of tissue-specific regu- latory elements on a gene can potentially allow expres- sion to be directed to a specific subset of cells in which the gene is normally expressed (Yuh et al., 1998). The gene encoding the neuropeptide vasoactive intes- tinal peptide (VIP) is expressed in subpopulations of neurons in several structures of the CNS, including the suprachiasmatic nucleus (SCN), cerebral cortex, hip- pocampus, amygdala, and thalamus (Dussaillant et al., 1992). VIP gene transcripts and/or VIP immunoreactiv- ity have been detected in a high percentage of myenteric and submucosal neurons throughout the gastrointestinal tract (Ho ¨kfelt et al., 1982; Fink et al., 1986), in many cholinergic ganglia (Lundberg et al., 1984; Leblanc et al., 1987), and within specific cholinergic neurons resid- ing in the sympathetic ganglia that innervate sweat Received May 17, 1999; revised manuscript received June 9, 1999; accepted June 10, 1999. Address correspondence and reprint requests to Dr. J. A. Waschek at 68-225 NPI, University of California at Los Angeles, 760 Westwood Plaza, Los Angeles, CA 90024-1759, U.S.A. Abbreviations used: E, embryonic day; P, postnatal day; SCG, su- perior cervical ganglia; SCN, suprachiasmatic nucleus; VIP, vasoactive intestinal peptide. 1739 Journal of Neurochemistry Lippincott Williams & Wilkins, Inc., Philadelphia © 1999 International Society for Neurochemistry
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Targeting of Embryonic and Postnatal Autonomic and Enteric Neurons with a Vasoactive Intestinal...

Targeting of Embryonic and Postnatal Autonomic and EntericNeurons with a Vasoactive Intestinal Peptide Transgene
James A. Waschek, Dawn T. Bravo, Matthew Sena, Robert Casillas, Williams Rodriguez,Thinh Nguyen, and Steve Colburn
Department of Psychiatry and Mental Retardation Research Center, University of California at Los Angeles,Los Angeles, California, U.S.A.
Abstract: The neuropeptide vasoactive intestinal peptide(VIP) is expressed in several distinct sites in the CNS, incholinergic and enteric ganglia, and in a small subpopu-lation of neurons within sympathetic ganglia. Previousstudies on the human VIP gene indicate that transcriptionin neural crest-derived neuroblastoma and pheochromo-cytoma cell lines is controlled in part by multiple regula-tory elements located along 4.5 kb of upstream 59 flank-ing sequence. In the current studies, transgenic micewere created with a chimeric gene consisting of 16.5 kbof the mouse VIP gene fused to the b-galactosidasereporter. In situ hybridization analysis in adult mice indi-cated that reporter gene expression was correctly tar-geted to neurons in the esophagus, stomach, small in-testine, and colon. No expression was observed in thebrain, including regions that contain abundant VIP-ex-pressing cells, such as the thalamus, amygdala, cerebralcortex, hippocampus, and suprachiasmatic nucleus.Analysis of transgene expression in neonatal and embry-onic day 13.5 mice revealed a near perfect correlationbetween VIP and b-galactosidase gene expression incranial cholinergic ganglia and the superior cervical gan-glia, and lack of transgene expression in sensory gangliaand in nonneuronal tissue. Potential ectopic transgeneexpression was observed in neonates, in the cerebellarexternal granule layer and in a small subpopulation ofneurons in the olfactory epithelium. We conclude that the16.5 kb of VIP gene used in these studies contains se-quences sufficient for directing expression specifically toVIP neurons in the PNS, and that sequences locatedelsewhere on the gene are required for proper CNS ex-pression. The VIP gene sequences used here should becapable of targeting other gene products to specific pop-ulations of embryonic and adult peripheral neurons with-out causing significant expression in the CNS. KeyWords: Vasoactive intestinal peptide—Transgenic—Promoter—Cholinergic—Embryo—Neonate.J. Neurochem. 73, 1739–1748 (1999).
Recently evolved transgenic approaches to study thenervous system in mice rely on the ability to target geneexpression to precise locations. One strategy to do this isto identify promoter sequences on a gene that is normally
expressed in the desired site, and then use the promotersequences to drive the expression of the gene of interest.For example, the role of CA1 hippocampal NMDA re-ceptors in learning and memory was examined recentlyusing this strategy (Tsien et al., 1996). In this work, theNMDA receptor was specifically disrupted in CA1 neu-rons using a Cre/Lox excision system in which the Crerecombinase was targeted to CA1 neurons using thea-calcium/calmodulin-dependent protein kinase II genepromoter. Despite the potential value of such an ap-proach, it is limited by the availability of promoter se-quences that can target gene expression specifically tothe cells of interest. Moreover, it is sometimes difficult toidentify effective tissue-specific promoters, because reg-ulatory sequences that enhance or restrict expressionmay be located far upstream or downstream from codingsequences (for review, see Waschek, 1995). On the otherhand, the inclusion or exclusion of tissue-specific regu-latory elements on a gene can potentially allow expres-sion to be directed to a specific subset of cells in whichthe gene is normally expressed (Yuh et al., 1998).
The gene encoding the neuropeptide vasoactive intes-tinal peptide (VIP) is expressed in subpopulations ofneurons in several structures of the CNS, including thesuprachiasmatic nucleus (SCN), cerebral cortex, hip-pocampus, amygdala, and thalamus (Dussaillant et al.,1992). VIP gene transcripts and/or VIP immunoreactiv-ity have been detected in a high percentage of myentericand submucosal neurons throughout the gastrointestinaltract (Hokfelt et al., 1982; Fink et al., 1986), in manycholinergic ganglia (Lundberg et al., 1984; Leblanc etal., 1987), and within specific cholinergic neurons resid-ing in the sympathetic ganglia that innervate sweat
Received May 17, 1999; revised manuscript received June 9, 1999;accepted June 10, 1999.
Address correspondence and reprint requests to Dr. J. A. Waschek at68-225 NPI, University of California at Los Angeles, 760 WestwoodPlaza, Los Angeles, CA 90024-1759, U.S.A.
Abbreviations used:E, embryonic day; P, postnatal day; SCG, su-perior cervical ganglia; SCN, suprachiasmatic nucleus; VIP, vasoactiveintestinal peptide.
1739
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 1999 International Society for Neurochemistry

glands and other targets (Ho¨kfelt et al., 1982; Landis,1983; Morales et al., 1995; Schutz et al., 1998).
We and others have studied the regulatory regions onthe human VIP gene using neural crest-derived neuro-blastoma and pheochromocytoma cell lines (Tsukada etal., 1987; Waschek et al., 1988, 1992; Agoston et al.,1990; Symes et al., 1995; Hahm and Eiden, 1996, 1998).These studies indicate that VIP gene transcription inthese cells is controlled by multiple regulatory elementsdistributed along.4.5 kb of 59 flanking sequence. In thisreport, we show that 16.5 kb of the mouse VIP gene,including 12 kb of 59 flanking sequence and 4.5 kb ofintragenic sequence, efficiently targetedb-galactosidasegene expression to peripheral VIP neurons, but not toCNS sites of VIP expression. Expression of the transgenein the PNS, like that of VIP, was detected in the embryoas early as embryonic day (E) 13.5.
EXPERIMENTAL PROCEDURES
Construction of the VIP/b-galactosidase fusion geneThe isolation and partial characterization of a genomic clone
containing the 59 portion of the VIP gene from a C57Bl/6mouse library were described previously (Waschek et al., 1992;Sena et al., 1994). A promoterlessb-galactosidase expressionvector (pGAL5; Lem et al., 1991), obtained from Dr. LeroyHood, was used as the parent vector. To construct the VIPtransgene, the polylinker and;200 bp of sequence upstreamfrom theb-galactosidase gene were removed from pGAL5 bydigesting withAsp718, and replaced with a synthetic polylinkermade with complementary oligonucleotides containing selected
restriction sites. The resulting vector was called pllacZ. Ap-proximately 16.5 kb of VIP gene sequences was then insertedinto pllacZ in a three-fragment ligation. The ligation reactioncontained pllacZ digested withSalI and SphI, a 12.5-kbSalI/Asp718 VIP gene fragment (consisting of 12-kb VIP gene 59flanking sequences, exon 1, and the 59 part of intron 1), and a4.0-kbAsp718/SphI fragment (containing the 39 part of intron1 and all VIP gene sequences downstream to theSphI site inexon 4) (Fig. 1). The resulting plasmid (pmVIPlacZ12) con-tained 16.5-kb sequences of the mouse VIP gene fused in frameto theb-galactosidase reporter gene (mVIPlacZ12; Fig. 1). Theligation mixture was transfected by electroporation into DH5acells (Bethesda Research Laboratories, Gaithersburg, MD,U.S.A.). Three colonies that hybridized to an oligonucleotidesequence in exon 1 (Lamperti et al., 1991) were selected andfound to contain identical 16.5-kb inserts. These clones werecharacterized further as described in Results.
Production of transgenic miceTo prepare the DNA fragment for injection, pmVIPlacZ12
was digested withSalI andNotI to release the vector sequences.The transgene (mVIPlacZ12) was then isolated by gel electro-phoresis and electroelution, purified by chromatography(Schleicher & Schuell Elutip), and ethanol precipitated. Thefragment was shipped to the National Transgenic DevelopmentFacility (a National Institute of Child Health and Human De-velopment–subsidized facility operated by DNX, Inc., Prince-ton, NJ, U.S.A.), where the fragment was injected into C57Bl/63 SJL F2 hybrid fertilized oocytes. Mice were screened for thepresence of the transgene by Southern blot analysis using a32Prandom primer-labeled 0.7-kbAatII/PstI fragment of pllacZ[AatII is within the b-galactosidase gene (Fig. 1),PstI is in thesynthetic polylinker of pllacZ].
FIG. 1. Exon/intron structure and restriction site map of the mouse VIP/peptide histidine isoleucine (PHI) gene and the chimerictransgene mVIPlacZ12. The seven exons on the endogenous mouse VIP/PHI are denoted by black boxes. As indicated, PHI and VIP areencoded on exons 4 and 5, respectively. The transgene contained ;12 kb of 59 flanking sequence of the mouse VIP/PHI gene, plus thefirst three exons and introns, and part of exon 4 fused in frame to the reporter gene b-galactosidase (lacZ). The sites on the mouse VIPgene denoted with asterisks represent the 59 and 39 limits of the mouse VIP gene in the original phage isolate. The asterisk at the 59 endof the mouse VIP gene corresponds to the SalI site in mVIPlacZ12. The SalI site was derived from the phage isolate and is not presenton the mouse VIP gene. All sequences between the SalI and SphI sites on mVIPlacZ12 are contiguous VIP gene sequences derived fromthe original phage isolate (see Experimental Procedures for details on construction). Sequences corresponding to the cDNA probes andriboprobes for the Southern analysis and in situ hybridization, respectively, are shown.
J. Neurochem., Vol. 73, No. 4, 1999
1740 J. A. WASCHEK ET AL.

Analysis of b-galactosidase mRNA expression intransgenic mice
Brain, intestine, and other tissues were dissected and eitherfrozen in tubes in liquid nitrogen for RNA extraction or frozenin OCT (Miles Laboratories, Elkhart, IN, U.S.A.) on dry ice forin situ hybridization. Samples were stored at270°C. Fornorthern analysis, total RNA and poly(A)-selected RNA wereprepared as described (Waschek et al., 1996). Theb-galacto-sidase probe used for northern blots was the same as that usedfor the Southern analysis described above. The VIP probe wasthe same as that described below for in situ hybridizationassays and corresponded to 39 coding sequences not present onthe transgene (Fig. 1). For in situ hybridization, adult tissuesand whole E13.5 and postnatal day (P) 1 mice frozen in OCTwere sectioned by cryostat (10–12mm), mounted on slides, andprocessed for in situ hybridization with digoxigenin-labeledprobes as described (Waschek et al., 1998).b-Galactosidasegene transcripts were detected using a digoxigenin-labeled an-tisense riboprobe corresponding to the entire 3.6 kb of theb-galactosidase gene in pllacZ (Fig. 1). This probe was pre-pared by digesting pllacZ withSalI (located in the upstreamsynthetic polylinker in pllacZ) and incubating with T7 RNApolymerase as described below. The senseb-galactosidaseprobe was prepared by digesting pllacZ withSstII (at the 39 endof the lacZ gene) and incubating with T3 RNA polymerase.Endogenous VIP mRNA transcripts were detected in alternatesections using a digoxigenin-labeled antisense riboprobe thatcorresponded to the 39-most sequences on the VIP mRNA (onexon 7, Fig. 1). To obtain this probe, the mouse VIP cDNA wasfirst amplified from a mouse brain cDNA library by polymerasechain reaction (Waschek et al., 1996). A 39 portion of this,containing nearly all sequences downstream from theHindIIIsite on exon seven (Fig. 1), was subcloned into Bluescript IISK. Because the VIP sequences in this probe were external tothe transgene, this probe detected only transcripts from theendogenous VIP gene.
After linearization of the plasmids, digoxigenin-labeled ri-boprobes were prepared using the kit protocol provided byBoehringer Mannheim. Before hybridization, probes were hy-drolyzed to an average size of;200 nucleotides. After hybrid-ization and wash steps, hybridizing transcripts were detectedusing an antibody to digoxigenin that was coupled to alkalinephosphatase (Boehringer Mannheim). Antibody and color sub-strates were incubated overnight as suggested by the manufac-turer’s protocol.
RESULTS
Construction of the mouse VIP/b-galactosidasefusion gene and production of transgenic mice
A phage clone was isolated from a mouse genomiclibrary, partially characterized, and found to contain an;17-kb insert, including;12-kb VIP gene 59 flankingsequences (Waschek et al., 1992; Sena et al., 1994).Further restriction analysis demonstrated the presence ofunique restriction sites on this genomic fragment thatallowed the subcloning of nearly the entire insert into aplasmid containing theb-galactosidase reporter gene(see Experimental Procedures and Fig. 1). The chimericVIP/b-galactosidase gene thus contained;12 kb of themouse VIP gene, plus the first three exons and introns,and part of exon 4 fused in frame to the reporter geneb-galactosidase. Subsequent restriction analysis demon-
strated a 16.5-kb reconstituted VIP gene fragment withthe expected hybridization pattern. Figure 2 shows thatthe full 12 kb of VIP gene 59 flanking sequences in thetransgene gave the same pattern of hybridization as theendogenous VIP gene. The VIP/b-galactosidase fusiongene (mVIPlacZ12) was then isolated from vector se-quences and injected into fertilized mouse oocytes asdescribed in Experimental Procedures. Six founder micecontaining nonrearranged transgene were obtained. Fromthese, we were able to establish three transgenic mousepedigrees, 481, 482, and 491, that produced sufficientoffspring for the analysis in adult animals. One pedigree,482, was carried for an extended period of time so thatmultiple analysis could be done on embryos and postna-tal transgenic offspring.
Analysis of transgene expressionThe endogenous VIP gene is expressed at high levels
in intestinal neurons, and in certain areas of the rat brain,most prominently the SCN, the cerebral cortex (in widelyspaced nonpyramidal neurons in layers II–V), severalnuclei of the amygdala and thalamus, and in discreteneurons of the hippocampus (Dussaillant et al., 1992).Initial analysis of the transgene was carried out on adultmice 2–6 months old. Histochemical analysis ofb-ga-lactosidase activity showed no detectable transgene ex-pression in the CNS of any of the animals. Analysis inthe intestine was equivocal, because variableb-galacto-sidase-like activity was present in the intestine of wild-type and transgenic mice. To circumvent these problems,subsequent analyses were carried out at the level of themRNA using a sensitive in situ hybridization protocol.This used a partially hydrolyzed digoxigenin-labeled ri-boprobe. Hybridizing transcripts were detected with anantidigoxigenin antibody coupled to alkaline phospha-tase. Like that seen with the histochemical assay, no
FIG. 2. Southern blot showing that mVIPlacZ12 (A) and genomic(B) DNA from C57Bl/6 mouse gave identical hybridization patternsusing an oligonucleotide hybridization probe (GGGGAACGGTTG-GCTGTAGCTGGCTAGTGCTCTGCT) directed against the 59 partof exon 1 of the VIP gene (Sena et al., 1994). A and B are 2-h and10-day autoradiographic exposures, respectively. The high-molec-ular-weight band in the EcoRI digest of the genomic blot (B) ispresumed to represent an artifact (possibly due to incompletedigestion), because there are no restriction sites within the exon 1hybridization probe (Lamperti et al., 1991).
J. Neurochem., Vol. 73, No. 4, 1999
1741VIP TRANSGENE EXPRESSION
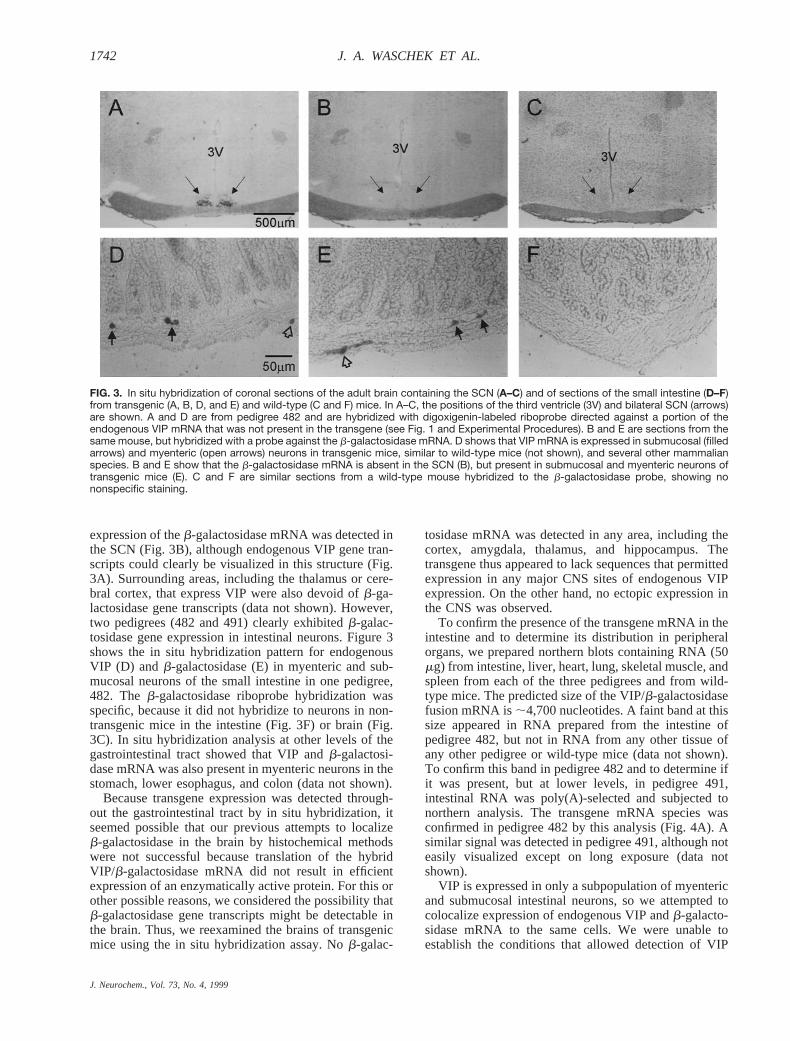
expression of theb-galactosidase mRNA was detected inthe SCN (Fig. 3B), although endogenous VIP gene tran-scripts could clearly be visualized in this structure (Fig.3A). Surrounding areas, including the thalamus or cere-bral cortex, that express VIP were also devoid ofb-ga-lactosidase gene transcripts (data not shown). However,two pedigrees (482 and 491) clearly exhibitedb-galac-tosidase gene expression in intestinal neurons. Figure 3shows the in situ hybridization pattern for endogenousVIP (D) and b-galactosidase (E) in myenteric and sub-mucosal neurons of the small intestine in one pedigree,482. Theb-galactosidase riboprobe hybridization wasspecific, because it did not hybridize to neurons in non-transgenic mice in the intestine (Fig. 3F) or brain (Fig.3C). In situ hybridization analysis at other levels of thegastrointestinal tract showed that VIP andb-galactosi-dase mRNA was also present in myenteric neurons in thestomach, lower esophagus, and colon (data not shown).
Because transgene expression was detected through-out the gastrointestinal tract by in situ hybridization, itseemed possible that our previous attempts to localizeb-galactosidase in the brain by histochemical methodswere not successful because translation of the hybridVIP/b-galactosidase mRNA did not result in efficientexpression of an enzymatically active protein. For this orother possible reasons, we considered the possibility thatb-galactosidase gene transcripts might be detectable inthe brain. Thus, we reexamined the brains of transgenicmice using the in situ hybridization assay. Nob-galac-
tosidase mRNA was detected in any area, including thecortex, amygdala, thalamus, and hippocampus. Thetransgene thus appeared to lack sequences that permittedexpression in any major CNS sites of endogenous VIPexpression. On the other hand, no ectopic expression inthe CNS was observed.
To confirm the presence of the transgene mRNA in theintestine and to determine its distribution in peripheralorgans, we prepared northern blots containing RNA (50mg) from intestine, liver, heart, lung, skeletal muscle, andspleen from each of the three pedigrees and from wild-type mice. The predicted size of the VIP/b-galactosidasefusion mRNA is;4,700 nucleotides. A faint band at thissize appeared in RNA prepared from the intestine ofpedigree 482, but not in RNA from any other tissue ofany other pedigree or wild-type mice (data not shown).To confirm this band in pedigree 482 and to determine ifit was present, but at lower levels, in pedigree 491,intestinal RNA was poly(A)-selected and subjected tonorthern analysis. The transgene mRNA species wasconfirmed in pedigree 482 by this analysis (Fig. 4A). Asimilar signal was detected in pedigree 491, although noteasily visualized except on long exposure (data notshown).
VIP is expressed in only a subpopulation of myentericand submucosal intestinal neurons, so we attempted tocolocalize expression of endogenous VIP andb-galacto-sidase mRNA to the same cells. We were unable toestablish the conditions that allowed detection of VIP
FIG. 3. In situ hybridization of coronal sections of the adult brain containing the SCN (A–C) and of sections of the small intestine (D–F)from transgenic (A, B, D, and E) and wild-type (C and F) mice. In A–C, the positions of the third ventricle (3V) and bilateral SCN (arrows)are shown. A and D are from pedigree 482 and are hybridized with digoxigenin-labeled riboprobe directed against a portion of theendogenous VIP mRNA that was not present in the transgene (see Fig. 1 and Experimental Procedures). B and E are sections from thesame mouse, but hybridized with a probe against the b-galactosidase mRNA. D shows that VIP mRNA is expressed in submucosal (filledarrows) and myenteric (open arrows) neurons in transgenic mice, similar to wild-type mice (not shown), and several other mammalianspecies. B and E show that the b-galactosidase mRNA is absent in the SCN (B), but present in submucosal and myenteric neurons oftransgenic mice (E). C and F are similar sections from a wild-type mouse hybridized to the b-galactosidase probe, showing nononspecific staining.
J. Neurochem., Vol. 73, No. 4, 1999
1742 J. A. WASCHEK ET AL.

mRNA with 35S-labeled riboprobes without losing theability to detectb-galactosidase mRNA. We thus con-ducted an analysis on consecutive 10-mm sections withdigoxigenin-labeled probes specific forb-galactosidaseand endogenous VIP mRNA. Preliminary work showed
that many VIP intestinal neurons could be detected ontwo and occasionally three consecutive sections. Thisfinding is consistent with the observation that manyintestinal neurons have cell bodies larger than 10mm(Furness et al., 1990). As shown in Fig. 5, neuronsfrequently appeared to express both endogenous VIP andb-galactosidase mRNA. Staining on consecutive sec-tions, however, was not always observed, a finding that ispredictable unless all neuron bodies are significantlygreater than 10mm and optimally oriented to the plane ofsection. Thus, to obtain additional evidence for colocal-ization, we counted labeled neurons on 48 consecutivesections. Because VIP is expressed in 30–50% of intes-tinal neurons (Furness et al., 1990),b-galactosidase-labeled cells might be expected to be more abundant thanVIP-labeled neurons if transgene expression occurrednondiscriminately. The mean number of neurons labeledper section (6SD) was approximately the same for theVIP and b-galactosidase probes (9.66 4.4 versus 8.96 5.5, respectively), indicating that transgene expressionwas restricted to an appropriate number of neurons.Taken together with the data in Fig. 5, it seems likely thattransgene expression was targeted to the appropriate
FIG. 4. Northern blot showingtransgene and endogenous VIPmRNA expression in the intestinefrom two mVIPlacZ12 transgenicpedigrees. Approximately 10 mg ofpoly(A)-selected intestinal RNAwas loaded in each lane, and thenorthern blot was prepared as de-scribed (Waschek et al., 1996). Theblot was first hybridized to a b-ga-lactosidase cDNA probe, washed,and exposed for 40 h at 270°C (A)(see Experimental Procedures). On longer exposure (13 days;not shown), a band at ;4,400 nucleotides could be better visu-alized in pedigree 491. The blot was then stripped and hybrid-ized to a VIP cDNA probe corresponding to sequences on exon7 (see Experimental Procedures) and then exposed for 24 h atroom temperature (B). Arrows show location of residual 28S and18S ribosomal RNA on blot, visualized by ethidium bromidestaining.
FIG. 5. Localization of VIP and reporter gene tran-scripts in three consecutive 10-mm sections of thecolon. Arrows 1 and 3 indicate positions of neuroncell bodies expressing b-galactosidase and VIPmRNA in two or three consecutive sections. Arrow 2indicates position of a neuron expressing VIP andb-galactosidase in the middle and bottom sections,respectively. The neuron indicated by arrow 4 ex-pressed b-galactosidase mRNA in the bottom, butnot VIP mRNA in the preceding or subsequent sec-tion (latter not shown), possibly because this neuroncell body was fully contained within this section.Size bar 5 50 mm.
J. Neurochem., Vol. 73, No. 4, 1999
1743VIP TRANSGENE EXPRESSION
subpopulation of neurons in the myenteric and submu-cosal plexus.
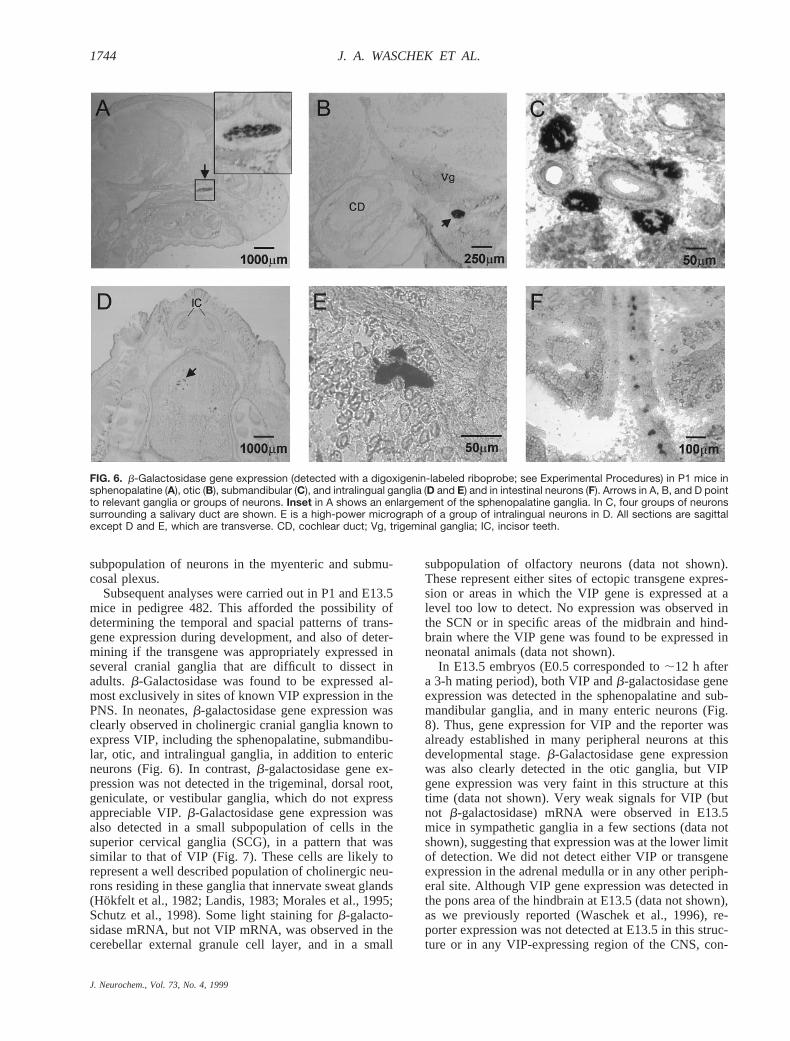
Subsequent analyses were carried out in P1 and E13.5mice in pedigree 482. This afforded the possibility ofdetermining the temporal and spacial patterns of trans-gene expression during development, and also of deter-mining if the transgene was appropriately expressed inseveral cranial ganglia that are difficult to dissect inadults.b-Galactosidase was found to be expressed al-most exclusively in sites of known VIP expression in thePNS. In neonates,b-galactosidase gene expression wasclearly observed in cholinergic cranial ganglia known toexpress VIP, including the sphenopalatine, submandibu-lar, otic, and intralingual ganglia, in addition to entericneurons (Fig. 6). In contrast,b-galactosidase gene ex-pression was not detected in the trigeminal, dorsal root,geniculate, or vestibular ganglia, which do not expressappreciable VIP.b-Galactosidase gene expression wasalso detected in a small subpopulation of cells in thesuperior cervical ganglia (SCG), in a pattern that wassimilar to that of VIP (Fig. 7). These cells are likely torepresent a well described population of cholinergic neu-rons residing in these ganglia that innervate sweat glands(Hokfelt et al., 1982; Landis, 1983; Morales et al., 1995;Schutz et al., 1998). Some light staining forb-galacto-sidase mRNA, but not VIP mRNA, was observed in thecerebellar external granule cell layer, and in a small
subpopulation of olfactory neurons (data not shown).These represent either sites of ectopic transgene expres-sion or areas in which the VIP gene is expressed at alevel too low to detect. No expression was observed inthe SCN or in specific areas of the midbrain and hind-brain where the VIP gene was found to be expressed inneonatal animals (data not shown).
In E13.5 embryos (E0.5 corresponded to;12 h aftera 3-h mating period), both VIP andb-galactosidase geneexpression was detected in the sphenopalatine and sub-mandibular ganglia, and in many enteric neurons (Fig.8). Thus, gene expression for VIP and the reporter wasalready established in many peripheral neurons at thisdevelopmental stage.b-Galactosidase gene expressionwas also clearly detected in the otic ganglia, but VIPgene expression was very faint in this structure at thistime (data not shown). Very weak signals for VIP (butnot b-galactosidase) mRNA were observed in E13.5mice in sympathetic ganglia in a few sections (data notshown), suggesting that expression was at the lower limitof detection. We did not detect either VIP or transgeneexpression in the adrenal medulla or in any other periph-eral site. Although VIP gene expression was detected inthe pons area of the hindbrain at E13.5 (data not shown),as we previously reported (Waschek et al., 1996), re-porter expression was not detected at E13.5 in this struc-ture or in any VIP-expressing region of the CNS, con-
FIG. 6. b-Galactosidase gene expression (detected with a digoxigenin-labeled riboprobe; see Experimental Procedures) in P1 mice insphenopalatine (A), otic (B), submandibular (C), and intralingual ganglia (D and E) and in intestinal neurons (F). Arrows in A, B, and D pointto relevant ganglia or groups of neurons. Inset in A shows an enlargement of the sphenopalatine ganglia. In C, four groups of neuronssurrounding a salivary duct are shown. E is a high-power micrograph of a group of intralingual neurons in D. All sections are sagittalexcept D and E, which are transverse. CD, cochlear duct; Vg, trigeminal ganglia; IC, incisor teeth.
J. Neurochem., Vol. 73, No. 4, 1999
1744 J. A. WASCHEK ET AL.

sistent with the results obtained in the CNS of adultanimals. Ectopic transgene expression was not observedin any site in E13.5 animals. Overall the data in embry-onic, postnatal, and adult animals indicate that the VIPsequences on the transgene specifically and effectivelytargetedb-galactosidase gene expression to the correctenteric and autonomic ganglia in a developmental stage-appropriate manner.
DISCUSSION
The results reported here show that 16.5 kb of themouse VIP gene contained sequences that specificallytargeted expression of a reporter gene to myenteric andsubmucosal gastrointestinal neurons, as well as to severalautonomic ganglia where the VIP gene is expressed. Incontrast, reporter gene expression was absent in the adultbrain, indicating that critical sequences conferring geneexpression to VIP neurons in the CNS were missing fromthe transgene and are therefore distinct. This, togetherwith the fact that ectopic expression was very limited,indicates that the VIP gene targeting sequences used hereshould be able to target the expression of other genesspecifically to VIP neurons in the PNS. These includeperhaps a majority of cholinergic autonomic neurons, butalso inhibitory myenteric intestinal neurons, and a subsetof submucosal intestinal neurons.
The VIP sequences targeted PNS neurons in a devel-opmental stage-appropriate manner. For example, VIPgene expression in the gastrointestinal tract and in sev-eral cholinergic ganglia was established by E13.5. Trans-gene expression was observed specifically in these samesites at this time (Fig. 8). In contrast, VIP gene expres-sion in the CNS is detectable in the mouse hindbrain atE11.5 (Waschek et al., 1996) and in the SCN, thalamus,cortex, and brainstem of rats just before birth (Graberand Burgunder, 1996). Transgene expression was unde-tectable in these CNS sites at any stage, again showing
that the VIP gene sequences used in this study almostexclusively target peripheral VIP neurons.
VIP gene expression and VIP immunoreactivity havebeen reported in a subpopulation of cholinergic neuronswithin certain sympathetic ganglia in rats (Landis, 1983;Morales et al., 1995; Schutz et al., 1998). During devel-opment, an initial peak of VIP expression around E14.5has been observed, with levels decreasing to;5% of thisby birth (Tyrrell and Landis, 1994). Analysis of singlecells in cryostat sections of stellate ganglia in that workindicated that the number of VIP neurons declines inparallel with immunoreactivity, and that neurons stillexpressing VIP at birth did not express tyrosine hydrox-ylase immunoreactivity. The neurons still expressingVIP at birth may represent cholinergic neurons withinthe ganglia that persists at least until the postnatal period.Our data in the mouse show that VIP andb-galactosidasewere expressed in a small subpopulation of cells in theP1 SCG (6–12 cells/12-mm section) and might thusrepresent this subset of SCG neurons that are cholinergic.In contrast, we could not detect appreciable transgeneexpression in sympathetic ganglia at E13.5 (correspond-ing to E14.5 in rats), and VIP gene expression was barelydetectable at this time. Thus, it is possible that thisunique early aspect of VIP gene regulation was notrecapitulated by the transgene, or that it differs signifi-cantly in mice versus rats.
The relative lack of ectopic expression observed in thebrain and elsewhere in these mice suggests that VIP genesilencer elements on the transgene may have activelysuppressed expression in the CNS and in other sites. AVIP transgene consisting of;2.0 kb of human VIP 59flanking sequence fused tob-galactosidase was variablyexpressed in certain areas of the brain that express VIP(Tsuruda et al., 1996). However, even within these brainregions, expression of this transgene reportedly did notcolocalize well with VIP in individual neurons, and little
FIG. 7. b-Galactosidase (A and C) and VIP (B) gene expression in P1 mouse SCG (sagittal sections). The SCG in A and B are encircledwith a dashed line for better visualization. C is a high-power micrograph of the b-galactosidase mRNA-expressing cell boxed in A.
J. Neurochem., Vol. 73, No. 4, 1999
1745VIP TRANSGENE EXPRESSION

or no transgene expression was observed in some VIPsites, most notably the SCN. Further, significant ectopicexpression of this transgene was observed in certainperipheral tissues, such as vascular smooth muscle andSchwann cells (Tolentino et al., 1995). Although theresults obtained with the human and mouse VIP trans-genes cannot be strictly compared, it seems possible thatthe much larger mouse VIP transgene contained se-quences that prevented significant ectopic expression.Studies on the vasopressin and other genes stronglyimply that mechanisms that restrict gene expression op-erate in the nervous system (for review, see Waschek,1995). For example, a vasopressin transgene containingthe intact bovine gene and;12 kb of 59 flanking se-quences gave a highly appropriate pattern of hypotha-lamic expression, whereas smaller vasopressin trans-genes showed ectopic hypothalamic and CNS transgeneexpression (Murphy et al., 1987; Russo et al., 1988; Anget al., 1993). Vasopressin transgenes containing only 0.2
kb of promoter showed high and inappropriate levels ofexpression in all organs analyzed (Ang et al., 1993). Themuch larger vasopressin transgene may thus have con-tained sequences that restrict expression, or that shieldedthe gene from regulatory sequences of other genes nearthe point of transgene insertion or from effects of chro-matin structure. Whether or not the VIP gene sequencesused in the current studies contain specific elements thatrestrict expression will require further study. However,analysis of the human VIP promoter indicates the exis-tence of sequences more than 4 kb upstream from thetranscription start site that inhibit expression (unpub-lished observations).
We have shown previously that 5.2 kb of the humanVIP gene promoter fused to the chloramphenicol acetyl-transferase was sufficient to target chloramphenicolacetyltransferase activity to the intestine, but not to VIPsites in the brain (Agoston et al., 1990). Thus, the humantransgene, like the mouse gene in the current studies,
FIG. 8. b-Galactosidase (A, C, andE) and VIP (B, D, and F) gene ex-pression in E13.5 mice (sagittalsections). A and B show positive-staining cells in sphenopalatine(open arrows) and submandibular(filled arrows) ganglia. The spheno-palatine ganglion in B appears astwo ganglia, because the section ison the medial edge of the ganglia.C–F show b-galactosidase (C andE) and VIP (D and F) gene expres-sion in an intestinal loop. The ar-rows in C and D denote approxi-mate edges of a cross section ofthe intestinal loop. E and F arehigh-power micrographs of the in-dicated areas in C and D, respec-tively. NC, nasal cavity; J, jaw.
J. Neurochem., Vol. 73, No. 4, 1999
1746 J. A. WASCHEK ET AL.

appeared to contain specific elements required for intes-tinal expression. Although not examined in those studies,it seems likely that the 5.2-kb human gene fragmentcontained enhancer sequences required for appropriateexpression in other PNS sites, because a transgene con-taining only 2.0 kb of the human VIP gene promoterclearly targeted expression to sphenopalatine as well asintestinal neurons (Tolentino et al., 1995). The lattertransgene also showed appropriately regulated expres-sion in the SCG after axotomy (Tsuruda et al., 1996).The increase in VIP in the SCG after axotomy appears tobe mediated at least in part by the cytokine leukemiainhibitory factor (Sun and Zigmond, 1996; Zigmond,1997) and a leukemia inhibitory factor-response DNAelement on the human VIP gene, located;1.1 kb up-stream from the transcription start site (Symes et al.,1995). A sequence corresponding to a cytokine-responseelement resides at a similar location on the mouse VIPgene (Sena et al., 1994), so it seems likely that the mouseVIP transgene would also respond to the axotomy. A210-bp sequence (which encompasses this cytokine-re-sponse element) is conserved 91% between mice andhumans (Sena et al., 1994), and may contribute to estab-lishing and/or maintaining basal expression in some neu-rons, in addition to conferring cytokine responsiveness.In addition, a DNA sequence on the human gene locatedmore than 4 kb upstream of the transcription start sitecontains complex enhancer and silencer elements thatinteract with downstream sequences to control cell-spe-cific expression (Hahm and Eiden, 1998; unpublishedobservations). Further analysis of the VIP gene promoterusing in vitro and in vivo approaches should eventuallyreveal how the very distinct patterns of VIP expressionare established and maintained, and how the gene isregulated in response to injury and to hormonal andenvironmental signals (for reviews, see Waschek, 1994;Zigmond, 1997).
Acknowledgment: This work was supported by NIHHD06576 and HD04612, the SHARE Foundation, and a Pilotand Feasibility Grant from CURE Digestive Diseases ResearchCenter, NIH-DK41301. We thank Dr. Babette Fuss for help indeveloping in situ hybridization protocols.
REFERENCES
Agoston D. A., Bravo D. T., and Waschek J. A. (1990) Expression ofa chimeric VIP gene is targeted to the intestine in transgenic mice.J. Neurosci. Res.27, 479–486.
Ang H.-L., Carter D. A., and Murphy D. (1993) Neuron-specificexpression and physiological regulation of bovine vasopressintransgenes in mice.EMBO J.12, 2397–2409.
Dussaillant M., Sarrieau A., Gozes I., Berod A., and Rostene W. (1992)Distribution of cells expressing vasoactive intestinal peptide/pep-tide histidine isoleucine-amide precursor messenger RNA in therat brain.Neuroscience50, 519–530.
Fink J., Montminy M. R., Tsukada T., Hoefler H., Specht L. A., LechanR. M., Wolfe H., Mandel G. and Goodman R. H. (1986)In situhybridization of somatostatin and VIP mRNA in the rat nervoussystem, inIn Situ Hybridization in Brain(Uhl G., ed), pp. 181–191. Plenum Press, New York.
Furness J. B., Bornstein J. C., and Smith T. K. (1990) The normalstructure of gastrointestinal innervation.J. Gastroenterol. Hepa-tol. 1, 1–9.
Graber M. and Burgunder J. M. (1996) Ontogeny of vasoactive intes-tinal peptide gene expression in rat brain.Anat. Embryol. (Berl.)194,595–605.
Hahm S. H. and Eiden L. E. (1996) Tissue-specific expression of thevasoactive intestinal peptide gene requires both an upstream tissuespecifier element and the 59 proximal cyclic AMP-responsiveelement.J. Neurochem.67, 1872–1881.
Hahm S. H. and Eiden L. E. (1998) Five discrete cis-active domainsdirect cell type-specific transcription of the vasoactive intestinalpeptide (VIP) gene.J. Biol. Chem.273,17086–17094.
Hokfelt T., Schultzberg M., Lundberg J. M., Fuxe K., Mutt V., Fahr-enkrug J., and Said S. I. (1982) Distribution of vasoactive intes-tinal polypeptide in the central and peripheral nervous systems asrevealed by immunocytochemistry, inVasoactive Intestinal Pep-tide (Said S., ed), pp. 65–90. Raven Press, New York.
Lamperti E. D., Rosen K. M., and Villa-Komaroff L. (1991) Charac-terization of the gene and messages for vasoactive intestinalpolypeptide (VIP) in rat and mouse.Brain Res. Mol. Brain Res.9,217–231.
Landis S. C. (1983) Development of cholinergic sympathetic neurons:evidence for transmitter plasticity in vivo.Fed. Proc.42, 1633–1638.
Leblanc G. G., Trimmer B. A., and Landis S. C. (1987) NeuropeptideY-like immunoreactivity in rat cranial parasympathetic neurons:coexistence with vasoactive intestinal peptide and choline acetyl-transferase.Proc. Natl. Acad. Sci. USA84, 3511–3515.
Lem J., Applebury M. L., Falk J. D., Flannery J. G., and Simon M. I.(1991) Tissue-specific and developmental regulation of rod opsinchimeric genes in transgenic mice.Neuron6, 201–210.
Lundberg J. M., Fahrenkrug J., Ho¨kfelt T., Martling C. R., Larsson O.,Tatemoto K., and Anggard A. (1984) Co-existence of peptide HI(PHI) and VIP in nerves regulating blood flow and bronchialsmooth muscle tone in various mammals including man.Peptides5, 593–606.
Morales M. A., Holmberg K., Xu Z. Q., Cozzari C., Hartman B. K.,Emson P., Goldstein M., Elfvin L. G., and Ho¨kfelt T. (1995)Localization of choline acetyltransferase in rat peripheral sympa-thetic neurons and its coexistence with nitric oxide synthase andneuropeptides.Proc. Natl. Acad. Sci. USA92, 11819–11823.
Murphy D., Bishop A., Rindi G., Murphy M. N., Stamp G. W. H., HansonJ., Polak J. M., and Hogan B. (1987) Mice transgenic for a vasopres-sin-SV40 hybrid oncogene develop tumors of the endocrine pancreasand the anterior pituitary. A possible model for human multipleendocrine neoplasia type 1.Am. J. Pathol.129,552–556.
Russo A. F., Crenshaw E. B. III, Lira S. A., Simmons D. M., SwansonL. W., and Rosenfeld M. G. (1988) Neuronal expression of chi-meric genes in transgenic mice.Neuron1, 311–320.
Schutz B., Schafer M. K., Eiden L. E., and Weihe E. (1998) VIP andNPY expression during differentiation of cholinergic and norad-renergic sympathetic neurons.Ann. NY Acad. Sci.865,537–541.
Sena M., Agoston D., and Waschek J. (1994) High conservation ofupstream regulatory sequence on the mouse and human vasoactiveintestinal peptide genes.DNA Seq.5, 25–29.
Sun Y. and Zigmond R. E. (1996) Involvement of leukemia inhibitoryfactor in the increases in galanin and vasoactive intestinal peptidemRNA and the decreases in neuropeptide Y and tyrosine hydrox-ylase mRNA in sympathetic neurons after axotomy.J. Neuro-chem.67, 1751–1760.
Symes A. J., Corpus L., and Fink J. S. (1995) Differences in nuclearsignaling by leukemia inhibitory factor and interferon-g: the roleof STAT proteins in regulating vasoactive intestinal peptide geneexpression.J. Neurochem.65, 1926–1933.
Tolentino P. J., Dikkes P., Tsuruda L., Ebert K., Fink J. S., Villa-KomaroffL., and Lamperti E. D. (1995) Quantitative analysis of the expressionof a VIP transgene.Brain Res. Mol. Brain Res.33, 47–60.
Tsien J. Z., Huerta P. T., and Tonegawa S. (1996) The essential role ofhippocampal CA1 NMDA receptor-dependent synaptic plasticityin spatial memory.Cell 87, 1327–1338.
J. Neurochem., Vol. 73, No. 4, 1999
1747VIP TRANSGENE EXPRESSION

Tsukada T., Fink J. S., Mandel G., and Goodman R. H. (1987) Iden-tification of a region in the human vasoactive intestinal polypep-tide gene responsible for regulation by cyclic AMP.J. Biol. Chem.262,8743–8747.
Tsuruda L. M., Lamperti E. D., Lewis S. E., Tolentino P. J., Dikkes P.,Villa-Komaroff L., Ebert K. M., and Fink J. S. (1996) Region-specific central nervous system expression and axotomy-inducedregulation in sympathetic neurons of a VIP–beta-galactosidase fusiongene in transgenic mice.Brain Res. Mol. Brain Res.42, 181–192.
Tyrrell S. and Landis S. C. (1994) The appearance of NPY and VIP insympathetic neuroblasts and subsequent alterations in their ex-pression.J. Neurosci.14, 4529–4547.
Waschek J. A. (1994) Environmental and hormonal control of vasoac-tive intestinal peptide (VIP) gene expression and changes inpathological conditions, inFrontiers in Neurobiology, Vol. 1:Neuropeptide Gene Expression(Turner A. J., ed), pp. 107–132.Portland Press, London.
Waschek J. (1995) Transgenic targeting of neuroendocrine peptide genesin the hypothalamic–pituitary axis.Mol. Neurobiol.10, 205–217.
Waschek J. A., Hsu C.-M., and Eiden L. E. (1988) Lineage-specific
regulation of the vasoactive intestinal peptide gene in neuroblas-toma cells is conferred by 5.2 kilobases of 59-flanking sequence.Proc. Natl. Acad. Sci. USA85, 9547–9551.
Waschek J. A., Krajniak K. G., Bravo D. T., and Agoston D. A. (1992)Regulation of human and mouse vasoactive intestinal peptidegenes.Biomed. Res.13 (Suppl. 2), 1–6.
Waschek J. A., Ellison J., Bravo D. T., and Hadley V. (1996) Embry-onic expression of vasoactive intestinal peptide (VIP) and VIPreceptor genes.J. Neurochem.66, 1762–1765.
Waschek J. A., Casillas R. A., Nguyen T. B., DiCicco-Bloom E. M.,Carpenter E. M., and Rodriguez W. I. (1998) Neural tube expres-sion of pituitary adenylate cyclase-activating peptide (PACAP)and receptor: potential role in patterning and neurogenesis.Proc.Natl. Acad. Sci. USA95, 9602–9607.
Yuh C. H., Bolouri H., and Davidson E. H. (1998) Genomic cis-regulatory logic: experimental and computational analysis of a seaurchin gene.Science279,1896–1902.
Zigmond R. E. (1997) LIF, NGF, and the cell body response toaxotomy.Neuroscientist3, 176–185.
J. Neurochem., Vol. 73, No. 4, 1999
1748 J. A. WASCHEK ET AL.




















