
CD8+ regulatory T cells induced by T cell vaccination protect against autoimmune nephritis
10
BASIC RESEARCH www.jasn.org CD8+ Regulatory T Cells Induced by T Cell Vaccination Protect Against Autoimmune Nephritis Yuan Min Wang,* Geoff Yu Zhang,* Min Hu,* Tania Polhill,* Andrew Sawyer,* Jimmy Jianheng Zhou,* Mitsuru Saito,* Debbie Watson,* † Huiling Wu, ‡ Ya Wang, § Xin Maggie Wang, | Yiping Wang, § David C.H. Harris, § and Stephen I. Alexander* *Centre for Kidney Research, Children’s Hospital at Westmead, Westmead, Australia; † Centre for Medical Bioscience, University of Wollongong, Wollongong, Australia; ‡ Collaborative Transplant Research Group, Royal Prince Alfred Hospital, Sydney, Australia; § Centre for Transplantation and Renal Research, University of Sydney at Westmead Millennium Institute, Sydney, Australia; and | Flow Cytometry Core Facility, Westmead Millennium Institute, University of Sydney, Sydney, Australia ABSTRACT Autoreactive T cells play a pivotal role in the pathogenesis of autoimmune kidney disease. T cell vaccination (TCV) may limit autoimmune disease and induce CD8+ regulatory T cells (Tregs). We used Heymann nephritis (HN), a rat model of human membranous nephritis, to study the effects of TCV on autoimmune kidney disease. We harvested CD4+ T cells from renal tubular antigen (Fx1A) -immunized rats and activated these cells in vitro to express the MHC Class Ib molecule Qa-1. Vaccination of Lewis rats with these autoreactive Fx1A-induced T cells protected against HN, whereas control-primed T cells did not. Rats that underwent TCV had lower levels of proteinuria and serum creatinine and significantly less glomerulosclerosis, tubular damage, and interstitial infiltrates. Furthermore, these rats expressed less IFN-g and IL-6 in splenocytes, whereas the numbers of Tregs and the expression of Foxp3 were unchanged. In vitro cytotoxicity assays showed CD8+ T cell-mediated elimination of Qa-1–expressing CD4+ T cells. In vivo, TCV abrogated the increase in Qa-1–expressing CXCR5+ TFH cells observed in HN compared with controls. Taken together, these results suggest that TCV protects against autoimmune kidney disease by targeting Qa-1–expressing autoreactive CD4+ cells. J Am Soc Nephrol 23: 1058–1067, 2012. doi: 10.1681/ASN.2011090914 CD4+ regulatory T cells (Tregs) have been shown to be critically involved in limiting autoimmune dis- eases such as multiple sclerosis, diabetes, and auto- immune renal disease. 1 The importance of CD8+ T cells with regulatory function has been less studied until recently. However, the recognition that T cell vaccination (TCV) induces regulatory CD8 T cells and that these CD8 T cells seem to use the MHC class Ib molecule Qa-1 (or HLA-E in humans) has reignited interest in this subset. 2–5 Furthermore, the identification that T follicular helper cells, a subset of splenic CD4 T cells that is responsible for assisting B cell antibody production and class switching, ex- press high levels of Qa-1 suggests that CD8 Tregs may play a role in limiting antibody-mediated disease. 6 Mouse Qa-1 and its human equivalent (HLA-E) are non-classic MHC class I molecules that bind hy- drophobic peptides, including the leader sequences of class I HLA molecules and components of the T cell receptor (TCR). These molecules are tran- siently upregulated on the surface of activated T and antigen-presenting cells. 7,8 Earlier studies of CD8+ suppressor cells suggested an important role for Qa-1 in mediating the function of these cells. 9 Qa-1 gene-deficient mice showed enhanced CD4+ T cell responses to self and foreign antigens and increased susceptibility to experimental autoimmune enceph- alomyelitis (EAE), the mouse model of multiple sclerosis. 10,11 Mice with mutations in Qa-1 to limit Received September 14, 2011. Accepted February 14, 2012. Published online ahead of print. Publication date available at www.jasn.org. Correspondence: Dr. Stephen I. Alexander, Centre for Kidney Research, Children’s Hospital at Westmead, Westmead, Sydney, NSW 2145, Australia. Email: [email protected] Copyright © 2012 by the American Society of Nephrology 1058 ISSN : 1046-6673/2306-1058 J Am Soc Nephrol 23: 1058–1067, 2012
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of CD8+ regulatory T cells induced by T cell vaccination protect against autoimmune nephritis

BASIC RESEARCH www.jasn.org
CD8+ Regulatory T Cells Induced by T Cell VaccinationProtect Against Autoimmune Nephritis
Yuan Min Wang,* Geoff Yu Zhang,* Min Hu,* Tania Polhill,* Andrew Sawyer,*Jimmy Jianheng Zhou,* Mitsuru Saito,* Debbie Watson,*† Huiling Wu,‡ Ya Wang,§
Xin Maggie Wang,| Yiping Wang,§ David C.H. Harris,§ and Stephen I. Alexander*
*Centre for Kidney Research, Children’s Hospital at Westmead, Westmead, Australia; †Centre for Medical Bioscience,University of Wollongong, Wollongong, Australia; ‡Collaborative Transplant Research Group, Royal Prince AlfredHospital, Sydney, Australia; §Centre for Transplantation and Renal Research, University of Sydney at WestmeadMillennium Institute, Sydney, Australia; and |Flow Cytometry Core Facility, Westmead Millennium Institute,University of Sydney, Sydney, Australia
ABSTRACTAutoreactive T cells play a pivotal role in the pathogenesis of autoimmune kidney disease. T cell vaccination(TCV)may limit autoimmune disease and induceCD8+ regulatory T cells (Tregs).We usedHeymann nephritis(HN), a ratmodel of humanmembranous nephritis, to study the effects of TCVonautoimmunekidneydisease.We harvestedCD4+T cells from renal tubular antigen (Fx1A) -immunized rats and activated these cells in vitroto express theMHCClass IbmoleculeQa-1. Vaccination of Lewis ratswith these autoreactive Fx1A-induced Tcells protected against HN, whereas control-primed T cells did not. Rats that underwent TCV had lower levelsof proteinuria and serum creatinine and significantly less glomerulosclerosis, tubular damage, and interstitialinfiltrates. Furthermore, these rats expressed less IFN-g and IL-6 in splenocytes, whereas the numbers ofTregs and the expression of Foxp3were unchanged. In vitro cytotoxicity assays showedCD8+ T cell-mediatedelimination of Qa-1–expressing CD4+ T cells. In vivo, TCV abrogated the increase in Qa-1–expressing CXCR5+TFH cells observed in HN compared with controls. Taken together, these results suggest that TCV protectsagainst autoimmune kidney disease by targeting Qa-1–expressing autoreactive CD4+ cells.
J Am Soc Nephrol 23: 1058–1067, 2012. doi: 10.1681/ASN.2011090914
CD4+ regulatory T cells (Tregs) have been shown tobe critically involved in limiting autoimmune dis-eases such as multiple sclerosis, diabetes, and auto-immune renal disease.1 The importance of CD8+T cells with regulatory function has been less studieduntil recently. However, the recognition that T cellvaccination (TCV) induces regulatory CD8 T cellsand that these CD8 T cells seem to use the MHCclass Ib molecule Qa-1 (or HLA-E in humans) hasreignited interest in this subset.2–5 Furthermore, theidentification that T follicular helper cells, a subsetof splenic CD4T cells that is responsible for assistingB cell antibody production and class switching, ex-press high levels of Qa-1 suggests that CD8 Tregsmayplay a role in limiting antibody-mediated disease.6
Mouse Qa-1 and its human equivalent (HLA-E)are non-classicMHC class Imolecules that bind hy-drophobic peptides, including the leader sequencesof class I HLA molecules and components of the
T cell receptor (TCR). These molecules are tran-siently upregulated on the surface of activated Tandantigen-presenting cells.7,8 Earlier studies of CD8+suppressor cells suggested an important role forQa-1 in mediating the function of these cells.9 Qa-1gene-deficient mice showed enhanced CD4+ T cellresponses to self and foreign antigens and increasedsusceptibility to experimental autoimmune enceph-alomyelitis (EAE), the mouse model of multiplesclerosis.10,11 Mice with mutations in Qa-1 to limit
Received September 14, 2011. Accepted February 14, 2012.
Published online ahead of print. Publication date available atwww.jasn.org.
Correspondence: Dr. Stephen I. Alexander, Centre for KidneyResearch, Children’s Hospital at Westmead, Westmead, Sydney,NSW 2145, Australia. Email: [email protected]
Copyright © 2012 by the American Society of Nephrology
1058 ISSN : 1046-6673/2306-1058 J Am Soc Nephrol 23: 1058–1067, 2012

CD8 binding but with inhibitory natural killer binding have se-vere autoimmune disease, including nephritis.6 TCV with acti-vated CD4+ T cells expressing Qa-1 can induce Qa-1–restrictedCD8+Tregs.4,10,12 In addition, CD8aa+TCRаb+Treg clonesrecognize a TCR-derived peptide in the context of Qa-1 mole-cules.13 More recently, a number of markers of CD8 Tregs thattarget Qa-1–expressing T follicular helper (TFH) cells have beenidentified, including CD44, inducible T cell costimulator ligand,CXCR5, very late antigen-4, and CD122.6
CD8+ Tregs specific for Qa-1–expressing, intermediate-affinity CD4 effector T cells have been found in EAE andtype 1 diabetes models induced by vaccination with antigenbut targeted at T cells with intermediate avidity distinct fromantigen specificity in keeping with a nonantigen-driven mech-anism of suppression.14 In humans, increased expression ofHLA-E is found on effector cells in multiple sclerosis. Mice canbe protected from EAE by TCV.15 This finding has led to TCV inclinical trials in patients with multiple sclerosis.16–18 In glomer-ulonephritis, the work by Trivedi et al.19 using an HLA DR4transgenic humanizedmousemodelwith peptide immunizationhas induced nephritis that is limited by TCV. However, the pro-tection against disease in thismodel is limited by its variability.19
Human idiopathic membranous nephritis is an autoim-mune renal disease characterized by severe proteinuria andprogression to renal failure.20 It is frequently caused by sensi-tization to phospholipase A2 receptor, an antigen expressed onglomerular podocytes,21 and associated with antibody deposi-tion in the glomerulus. It is a major cause of ESRD world-wide.22 Active Heymann nephritis (HN), an experimental ratmodel of human autoimmune-mediatedmembranous nephri-tis, is used to study this disease and its potential therapies.HN isinduced in Lewis rats by immunization with a crude renal tu-bular antigen (Fx1A), and it reproduces clinical features ofhuman idiopathic membranous glomerulonephritis.23 Al-though autoantibodies particularly against megalin are ob-served in HN, the disease also involves T cell-mediated injurywith a major role of CD4+ and CD8+ T cells.24–26 Althoughsome studies have suggested that complement is important inHN,27 recent studies find that C6-deficient rats developequivalent passive HN, excluding antibody-bound comple-ment as a major component of injury.28
We have identified TCR restriction to TCRVb-2 and -16 inHN and have shown that TCR DNA vaccines directed againstthe Vb-sequence of the TCR can protect against nephritis29–31
A slowly progressivemodel of autoimmune disease with tissue-specific antibody production and the involvement of specificTCRs by pathogenic T cells thatmay be presented on their surfaceby Qa-1 to CD8 make Tregs HN an excellent candidate for TCVand evaluation of its mechanism of protection. Here, for the firsttime, we evaluate the role of CD8 Tregs induced by TCV inautoimmune renal disease, evaluate the role of Qa-1, and assessthe potential role of TFH cells in autoimmune renal disease.
Our studies showed that TCV protects rats from develop-ing active HN, with a major reduction in renal pathology andFx1A antibodies. Qa-1 was expressed in vaccinating CD4+
cells and TFH cells, which were both increased in HN. Qa-1–restricted CD8+ cytotoxicity was shown in TCVrats. These find-ings suggest a therapeutic role forTCVandCD8+Tregs in limitingautoimmune renal disease by deleting autoreactive CD4 T cells.
RESULTS
Autoreactive CD4+ T Cells from HN Rats Used forCellular Vaccination Express Qa-1HN is induced in Lewis rats by immunization with a renaltubular antigen (Fx1A), leading to the induction of autoanti-bodies and immune injuries to the kidney. It has been shownpreviously in other disease models that TCV limits autoimmunedisease through suppressive CD8+ T cells that target Qa-1–expressing CD4+ cells. We, therefore, sought to determine ifTCV would be protective in the HN model. CD4+ T cellsfrom draining lymph nodes (LNs) of the hind feet of rats im-munized with Fx1A or CFA 10 days before were isolated andcultured in vitro with concanavalin A (conA), and then, theywere irradiated and used to vaccinate Lewis rats (Figure 1A).For the control group, CD4+ T cells from rats immunizedwith CFA were cultured under the same conditions as the Fx1Agroup. CD4+ T cells used for TCVand controls were examinedfor surface expression of Qa-1 after conA stimulation. Increasedexpression of Qa-1 was shown (Figure 1 B and C). Qa-1 expres-sion on CD4+ T cells was increased from 13% (PBS) to 18%(conA) in CFA-immunized rats (Figure 1B). However, therewas a larger increase in Qa-1 expression on CD4 T cells from11% to 27% in Fx1A-immunized rats after conA stimulationcompared with the control group (Figure 1C, P,0.05, n=6).
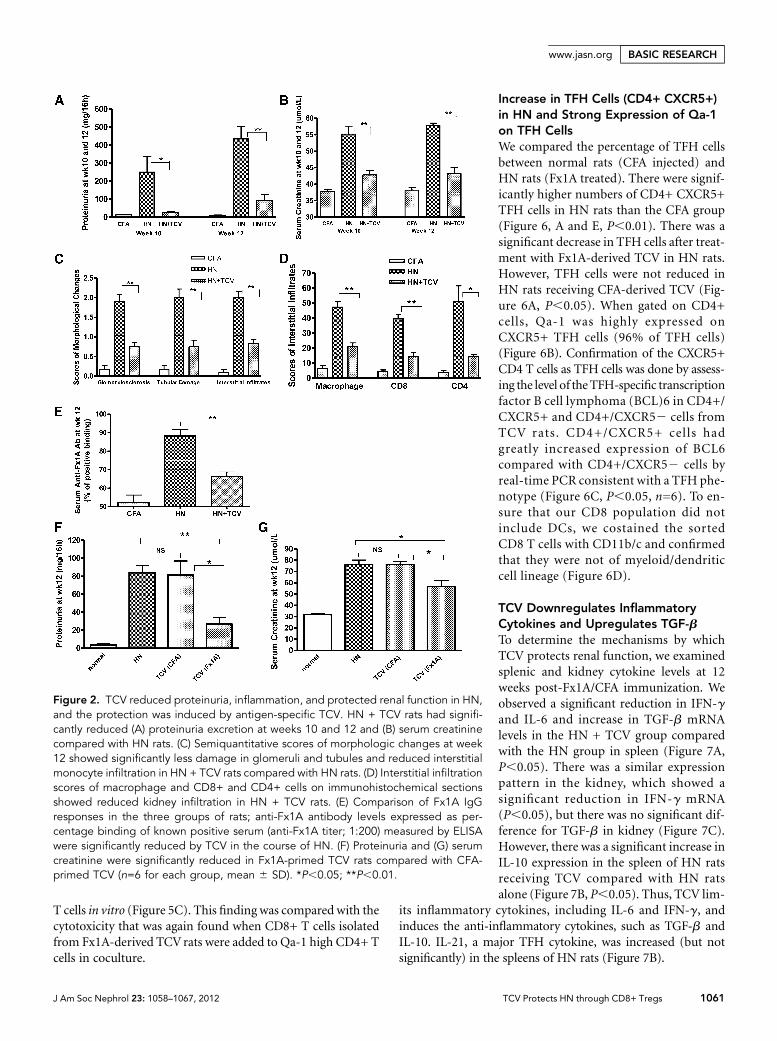
TCV Reduces Proteinuria, Renal Dysfunction, andHistologic Damage in HNT cell-vaccinated HN rats (HN + TCV) showed significantprotection against disease at weeks 10 and 12 compared withnonvaccinated HN rats that had already developed severe HNwith more severe proteinuria and elevation of serum creati-nine (P,0.05 or P,0.01) (Figure 2, A and B). Quantitativeanalysis showed significantly less glomerulosclerosis, tubulardamage, and interstitial monocyte infiltration in the HN +TCV group compared with the HN group (P,0.05 orP,0.01) (Figure 2, C and D). Anti-Fx1A antibody levels ex-pressed as percentage binding of known positive serum weremeasured by ELISA. The level of serum anti-Fx1A antibody(total IgG) inHN rats was significantly suppressed by TCV (Fig-ure 2E). However, although the TCV (Fx1A) group signifi-cantly reduced proteinuria and serum creatinine, the controlTCV (CFA) group was not protected and was equivalent tostandard HN (Figure, 2 F and G), and the histologic renal injuryreflected the same specific protectionbyTCV(Fx1A) (Figure 3C).
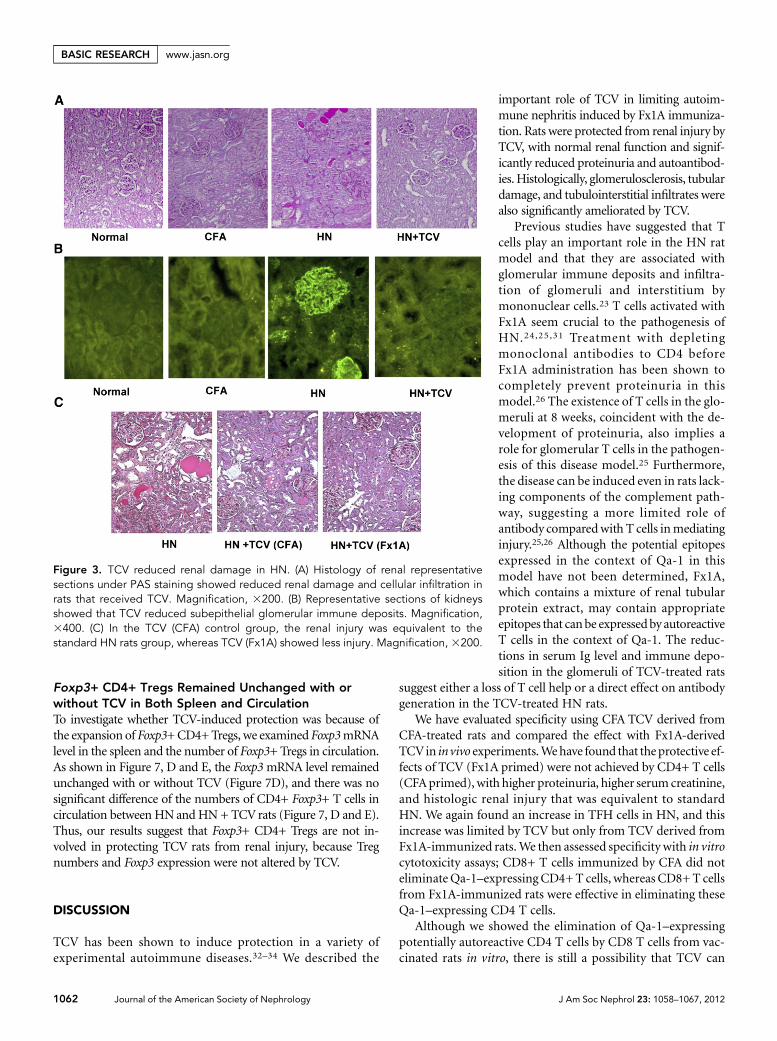
Histologic renal injury at week 12 was significantly amelio-rated in HN + TCV compared with HN rats (Figure 3A). Fur-thermore, the HN group showed significant immune deposition
J Am Soc Nephrol 23: 1058–1067, 2012 TCV Protects HN through CD8+ Tregs 1059
www.jasn.org BASIC RESEARCH

along the glomerular basement membrane,which was not detected in the HN + TCVorcontrol group (Figure 3B).
TCV Reduces Infiltration ofMacrophages and CD4+ and CD8+T CellsTo assess the cellular infiltrations, we per-formed immunohistochemistry of week 12kidneysections.Therewasamarkedreductionof infiltrating macrophages, CD8+ T cells,and CD4+ T cells in the HN + TCV groupcompared with the HN group (Figure 4).
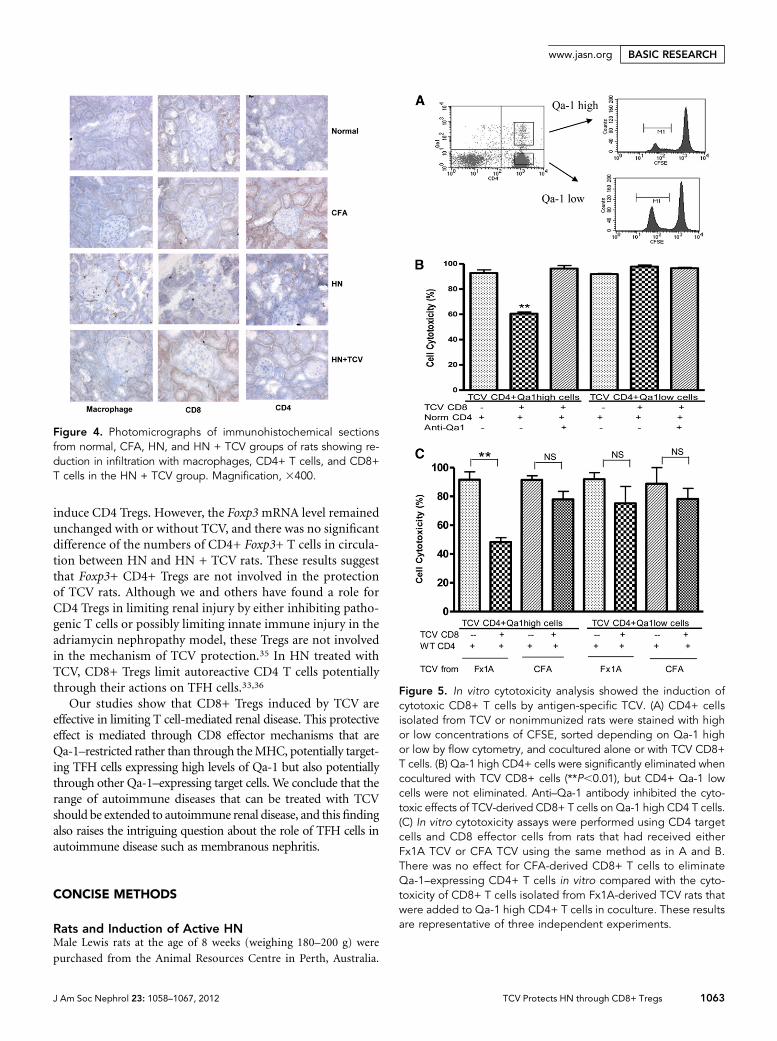
Qa-1–Expressing CD4+ T Cells CanBe Eliminated by CD8+ T Cells In VitroTo investigate the possible role of CD8+Tregs in protection of renal function byTCV, we conducted in vitro cytotoxicityassays. The studies were carried out tomore directly define the potential inhibi-tory interaction between CD4+ and CD8+cells (Concise Methods). CD4+ splenicT cells isolated from HN + TCV rats weresorted by flow cytometry into Qa-1 highand low populations and stained with alow concentration of carboxyfluoresceinsuccinimidyl ester (CFSE); an equal numberof control CD4 T cells from nonimmunizedrats were stained with a high concentrationof CFSE to act as a reference number of cells(Figure 5A). CFSE staining (high and low)was used to distinguish between the controland test populations. The relative CD4T cell survival was reduced to about 60%when CD8+ T cells isolated from TCV ratswere added to Qa-1 high and normal CD4+T cells in coculture (P,0.01). The effect ofCD8+ T cells was abrogated by the additionof blocking mouse anti-rat Qa-1 antibody(Figure 5B). These results suggest that CD8+T cells specifical ly el iminate Qa-1–expressing CD4+ T cells. This result is alsoshown by the absence of effect on Qa-1 lowCD4 cells. Thus, the induced CD8 T cellsseem restricted byQa-1 and able to eliminateQa-1–expressing autoreactive CD4 T cells.
To assess specificity, we conducted invitro cytotoxicity assays using CD4 targetcells and CD8 effector cells from rats thathad received either Fx1A TCVor CFA TCVusing the same method as described above.We found no effect for CFA-derived CD8+T cells to eliminate Qa-1–expressing CD4+
Figure 1. TCV through autoreactive CD4+ T cells increased the expression of Qa-1. (A)Fx1A was generated from Sprague–Dawley rat kidneys as previously described. Lewisrats were immunized with Fx1A as the experimental group and CFA as the controlgroup. CD4+ T cells were purified from draining LNs of the two groups of rats 10 daysafter footpad injection of Fx1A or CFA and cultured with conA for 40 hours. The CD4+T cells from Fx1A- and CFA-immunized rats were irradiated at 30 Gy and injected intotwo groups of Lewis rats through the tail vein (53106 cells/rat). Fx1A was injected intoeach of their hind footpads 3 weeks after TCV. (B and C) CD4+ T cells were isolatedfrom draining LNs from both (B) CFA- and (C) Fx1A-immunized rats and cultured onPBS or conA for 40 hours. Qa-1 expression on CD4+ T cells was increased from 13%(PBS) to 18% (conA) for CFA-immunized rats and 11% to 27% for Fx1A-immunized ratsafter conA stimulation (P,0.05, n=6).
1060 Journal of the American Society of Nephrology J Am Soc Nephrol 23: 1058–1067, 2012
BASIC RESEARCH www.jasn.org
T cells in vitro (Figure 5C). This finding was compared with thecytotoxicity that was again found when CD8+ T cells isolatedfrom Fx1A-derived TCVrats were added to Qa-1 high CD4+ Tcells in coculture.
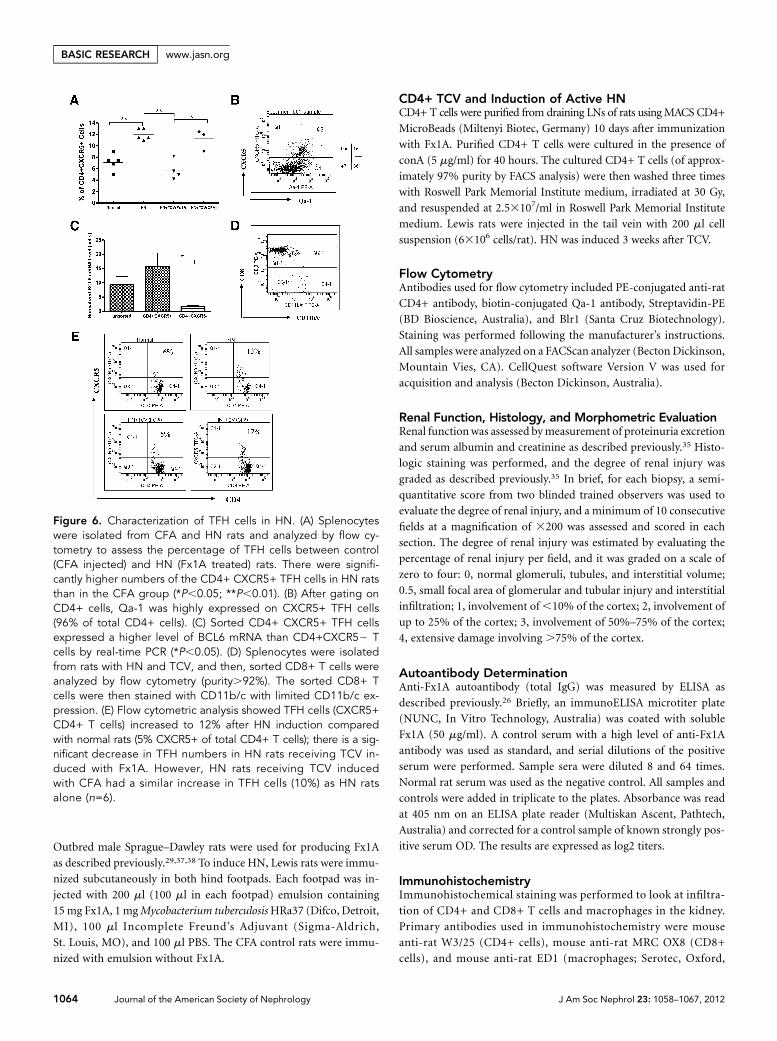
Increase in TFH Cells (CD4+ CXCR5+)in HN and Strong Expression of Qa-1on TFH CellsWe compared the percentage of TFH cellsbetween normal rats (CFA injected) andHN rats (Fx1A treated). There were signif-icantly higher numbers of CD4+ CXCR5+TFH cells in HN rats than the CFA group(Figure 6, A and E, P,0.01). There was asignificant decrease in TFH cells after treat-ment with Fx1A-derived TCV in HN rats.However, TFH cells were not reduced inHN rats receiving CFA-derived TCV (Fig-ure 6A, P,0.05). When gated on CD4+cells, Qa-1 was highly expressed onCXCR5+ TFH cells (96% of TFH cells)(Figure 6B). Confirmation of the CXCR5+CD4 T cells as TFH cells was done by assess-ing the level of theTFH-specific transcriptionfactor B cell lymphoma (BCL)6 in CD4+/CXCR5+ and CD4+/CXCR52 cells fromTCV rats. CD4+/CXCR5+ cells hadgreatly increased expression of BCL6compared with CD4+/CXCR52 cells byreal-time PCR consistent with a TFH phe-notype (Figure 6C, P,0.05, n=6). To en-sure that our CD8 population did notinclude DCs, we costained the sortedCD8 T cells with CD11b/c and confirmedthat they were not of myeloid/dendriticcell lineage (Figure 6D).
TCV Downregulates InflammatoryCytokines and Upregulates TGF-bTo determine the mechanisms by whichTCV protects renal function, we examinedsplenic and kidney cytokine levels at 12weeks post-Fx1A/CFA immunization. Weobserved a significant reduction in IFN-gand IL-6 and increase in TGF-b mRNAlevels in the HN + TCV group comparedwith the HN group in spleen (Figure 7A,P,0.05). There was a similar expressionpattern in the kidney, which showed asignificant reduction in IFN-g mRNA(P,0.05), but there was no significant dif-ference for TGF-b in kidney (Figure 7C).However, there was a significant increase inIL-10 expression in the spleen of HN ratsreceiving TCV compared with HN ratsalone (Figure 7B, P,0.05). Thus, TCV lim-
its inflammatory cytokines, including IL-6 and IFN-g, andinduces the anti-inflammatory cytokines, such as TGF-b andIL-10. IL-21, a major TFH cytokine, was increased (but notsignificantly) in the spleens of HN rats (Figure 7B).
Figure 2. TCV reduced proteinuria, inflammation, and protected renal function in HN,and the protection was induced by antigen-specific TCV. HN + TCV rats had signifi-cantly reduced (A) proteinuria excretion at weeks 10 and 12 and (B) serum creatininecompared with HN rats. (C) Semiquantitative scores of morphologic changes at week12 showed significantly less damage in glomeruli and tubules and reduced interstitialmonocyte infiltration in HN + TCV rats compared with HN rats. (D) Interstitial infiltrationscores of macrophage and CD8+ and CD4+ cells on immunohistochemical sectionsshowed reduced kidney infiltration in HN + TCV rats. (E) Comparison of Fx1A IgGresponses in the three groups of rats; anti-Fx1A antibody levels expressed as per-centage binding of known positive serum (anti-Fx1A titer; 1:200) measured by ELISAwere significantly reduced by TCV in the course of HN. (F) Proteinuria and (G) serumcreatinine were significantly reduced in Fx1A-primed TCV rats compared with CFA-primed TCV (n=6 for each group, mean 6 SD). *P,0.05; **P,0.01.
J Am Soc Nephrol 23: 1058–1067, 2012 TCV Protects HN through CD8+ Tregs 1061
www.jasn.org BASIC RESEARCH
Foxp3+ CD4+ Tregs Remained Unchanged with orwithout TCV in Both Spleen and CirculationTo investigate whether TCV-induced protection was because ofthe expansion of Foxp3+CD4+Tregs, we examined Foxp3mRNAlevel in the spleen and the number of Foxp3+ Tregs in circulation.As shown in Figure 7, D and E, the Foxp3mRNA level remainedunchanged with or without TCV (Figure 7D), and there was nosignificant difference of the numbers of CD4+ Foxp3+ T cells incirculation between HNand HN + TCVrats (Figure 7, D and E).Thus, our results suggest that Foxp3+ CD4+ Tregs are not in-volved in protecting TCV rats from renal injury, because Tregnumbers and Foxp3 expression were not altered by TCV.
DISCUSSION
TCV has been shown to induce protection in a variety ofexperimental autoimmune diseases.32–34 We described the
important role of TCV in limiting autoim-mune nephritis induced by Fx1A immuniza-tion. Rats were protected from renal injury byTCV, with normal renal function and signif-icantly reduced proteinuria and autoantibod-ies.Histologically, glomerulosclerosis, tubulardamage, and tubulointerstitial infiltrates werealso significantly ameliorated by TCV.
Previous studies have suggested that Tcells play an important role in the HN ratmodel and that they are associated withglomerular immune deposits and infiltra-tion of glomeruli and interstitium bymononuclear cells.23 T cells activated withFx1A seem crucial to the pathogenesis ofHN.24,25,31 Treatment with depletingmonoclonal antibodies to CD4 beforeFx1A administration has been shown tocompletely prevent proteinuria in thismodel.26 The existence of T cells in the glo-meruli at 8 weeks, coincident with the de-velopment of proteinuria, also implies arole for glomerular T cells in the pathogen-esis of this disease model.25 Furthermore,the disease can be induced even in rats lack-ing components of the complement path-way, suggesting a more limited role ofantibody comparedwith T cells inmediatinginjury.25,26 Although the potential epitopesexpressed in the context of Qa-1 in thismodel have not been determined, Fx1A,which contains a mixture of renal tubularprotein extract, may contain appropriateepitopes that canbe expressedby autoreactiveT cells in the context of Qa-1. The reduc-tions in serum Ig level and immune depo-sition in the glomeruli of TCV-treated rats
suggest either a loss of T cell help or a direct effect on antibodygeneration in the TCV-treated HN rats.
We have evaluated specificity using CFA TCV derived fromCFA-treated rats and compared the effect with Fx1A-derivedTCVin invivo experiments.Wehave found that theprotective ef-fects of TCV (Fx1A primed) were not achieved by CD4+ T cells(CFAprimed), with higher proteinuria, higher serumcreatinine,and histologic renal injury that was equivalent to standardHN. We again found an increase in TFH cells in HN, and thisincrease was limited by TCV but only from TCV derived fromFx1A-immunized rats.We then assessed specificity with in vitrocytotoxicity assays; CD8+ T cells immunized by CFA did noteliminateQa-1–expressingCD4+T cells, whereas CD8+T cellsfrom Fx1A-immunized rats were effective in eliminating theseQa-1–expressing CD4 T cells.
Although we showed the elimination of Qa-1–expressingpotentially autoreactive CD4 T cells by CD8 T cells from vac-cinated rats in vitro, there is still a possibility that TCV can
Figure 3. TCV reduced renal damage in HN. (A) Histology of renal representativesections under PAS staining showed reduced renal damage and cellular infiltration inrats that received TCV. Magnification, 3200. (B) Representative sections of kidneysshowed that TCV reduced subepithelial glomerular immune deposits. Magnification,3400. (C) In the TCV (CFA) control group, the renal injury was equivalent to thestandard HN rats group, whereas TCV (Fx1A) showed less injury. Magnification, 3200.
1062 Journal of the American Society of Nephrology J Am Soc Nephrol 23: 1058–1067, 2012
BASIC RESEARCH www.jasn.org
induce CD4 Tregs. However, the Foxp3mRNA level remainedunchanged with or without TCV, and there was no significantdifference of the numbers of CD4+ Foxp3+ T cells in circula-tion between HN and HN + TCV rats. These results suggestthat Foxp3+ CD4+ Tregs are not involved in the protectionof TCV rats. Although we and others have found a role forCD4 Tregs in limiting renal injury by either inhibiting patho-genic T cells or possibly limiting innate immune injury in theadriamycin nephropathy model, these Tregs are not involvedin the mechanism of TCV protection.35 In HN treated withTCV, CD8+ Tregs limit autoreactive CD4 T cells potentiallythrough their actions on TFH cells.33,36
Our studies show that CD8+ Tregs induced by TCV areeffective in limiting T cell-mediated renal disease. This protectiveeffect is mediated through CD8 effector mechanisms that areQa-1–restricted rather than through theMHC, potentially target-ing TFH cells expressing high levels of Qa-1 but also potentiallythrough other Qa-1–expressing target cells. We conclude that therange of autoimmune diseases that can be treated with TCVshould be extended to autoimmune renal disease, and this findingalso raises the intriguing question about the role of TFH cells inautoimmune disease such as membranous nephritis.
CONCISE METHODS
Rats and Induction of Active HNMale Lewis rats at the age of 8 weeks (weighing 180–200 g) were
purchased from the Animal Resources Centre in Perth, Australia.
Figure 4. Photomicrographs of immunohistochemical sectionsfrom normal, CFA, HN, and HN + TCV groups of rats showing re-duction in infiltration with macrophages, CD4+ T cells, and CD8+T cells in the HN + TCV group. Magnification, 3400.
Figure 5. In vitro cytotoxicity analysis showed the induction ofcytotoxic CD8+ T cells by antigen-specific TCV. (A) CD4+ cellsisolated from TCV or nonimmunized rats were stained with highor low concentrations of CFSE, sorted depending on Qa-1 highor low by flow cytometry, and cocultured alone or with TCV CD8+T cells. (B) Qa-1 high CD4+ cells were significantly eliminated whencocultured with TCV CD8+ cells (**P,0.01), but CD4+ Qa-1 lowcells were not eliminated. Anti–Qa-1 antibody inhibited the cyto-toxic effects of TCV-derived CD8+ T cells on Qa-1 high CD4 T cells.(C) In vitro cytotoxicity assays were performed using CD4 targetcells and CD8 effector cells from rats that had received eitherFx1A TCV or CFA TCV using the same method as in A and B.There was no effect for CFA-derived CD8+ T cells to eliminateQa-1–expressing CD4+ T cells in vitro compared with the cyto-toxicity of CD8+ T cells isolated from Fx1A-derived TCV rats thatwere added to Qa-1 high CD4+ T cells in coculture. These resultsare representative of three independent experiments.
J Am Soc Nephrol 23: 1058–1067, 2012 TCV Protects HN through CD8+ Tregs 1063
www.jasn.org BASIC RESEARCH
Outbred male Sprague–Dawley rats were used for producing Fx1A
as described previously.29,37,38 To induce HN, Lewis rats were immu-
nized subcutaneously in both hind footpads. Each footpad was in-
jected with 200 ml (100 ml in each footpad) emulsion containing
15mg Fx1A, 1 mgMycobacterium tuberculosisHRa37 (Difco, Detroit,
MI), 100 ml Incomplete Freund’s Adjuvant (Sigma-Aldrich,
St. Louis, MO), and 100 ml PBS. The CFA control rats were immu-
nized with emulsion without Fx1A.
CD4+ TCV and Induction of Active HNCD4+ T cells were purified from draining LNs of rats usingMACS CD4+
MicroBeads (Miltenyi Biotec, Germany) 10 days after immunization
with Fx1A. Purified CD4+ T cells were cultured in the presence of
conA (5 mg/ml) for 40 hours. The cultured CD4+ T cells (of approx-
imately 97% purity by FACS analysis) were then washed three times
with Roswell Park Memorial Institute medium, irradiated at 30 Gy,
and resuspended at 2.53107/ml in Roswell Park Memorial Institute
medium. Lewis rats were injected in the tail vein with 200 ml cell
suspension (63106 cells/rat). HN was induced 3 weeks after TCV.
Flow CytometryAntibodies used for flow cytometry included PE-conjugated anti-rat
CD4+ antibody, biotin-conjugated Qa-1 antibody, Streptavidin-PE
(BD Bioscience, Australia), and Blr1 (Santa Cruz Biotechnology).
Staining was performed following the manufacturer’s instructions.
All samples were analyzed on a FACScan analyzer (Becton Dickinson,
Mountain Vies, CA). CellQuest software Version V was used for
acquisition and analysis (Becton Dickinson, Australia).
Renal Function, Histology, and Morphometric EvaluationRenal functionwas assessed bymeasurement of proteinuria excretion
and serum albumin and creatinine as described previously.35 Histo-
logic staining was performed, and the degree of renal injury was
graded as described previously.35 In brief, for each biopsy, a semi-
quantitative score from two blinded trained observers was used to
evaluate the degree of renal injury, and a minimum of 10 consecutive
fields at a magnification of 3200 was assessed and scored in each
section. The degree of renal injury was estimated by evaluating the
percentage of renal injury per field, and it was graded on a scale of
zero to four: 0, normal glomeruli, tubules, and interstitial volume;
0.5, small focal area of glomerular and tubular injury and interstitial
infiltration; 1, involvement of,10% of the cortex; 2, involvement of
up to 25% of the cortex; 3, involvement of 50%–75% of the cortex;
4, extensive damage involving .75% of the cortex.
Autoantibody DeterminationAnti-Fx1A autoantibody (total IgG) was measured by ELISA as
described previously.26 Briefly, an immunoELISA microtiter plate
(NUNC, In Vitro Technology, Australia) was coated with soluble
Fx1A (50 mg/ml). A control serum with a high level of anti-Fx1A
antibody was used as standard, and serial dilutions of the positive
serum were performed. Sample sera were diluted 8 and 64 times.
Normal rat serum was used as the negative control. All samples and
controls were added in triplicate to the plates. Absorbance was read
at 405 nm on an ELISA plate reader (Multiskan Ascent, Pathtech,
Australia) and corrected for a control sample of known strongly pos-
itive serum OD. The results are expressed as log2 titers.
ImmunohistochemistryImmunohistochemical staining was performed to look at infiltra-
tion of CD4+ and CD8+ T cells and macrophages in the kidney.
Primary antibodies used in immunohistochemistry were mouse
anti-rat W3/25 (CD4+ cells), mouse anti-rat MRC OX8 (CD8+
cells), and mouse anti-rat ED1 (macrophages; Serotec, Oxford,
Figure 6. Characterization of TFH cells in HN. (A) Splenocyteswere isolated from CFA and HN rats and analyzed by flow cy-tometry to assess the percentage of TFH cells between control(CFA injected) and HN (Fx1A treated) rats. There were signifi-cantly higher numbers of the CD4+ CXCR5+ TFH cells in HN ratsthan in the CFA group (*P,0.05; **P,0.01). (B) After gating onCD4+ cells, Qa-1 was highly expressed on CXCR5+ TFH cells(96% of total CD4+ cells). (C) Sorted CD4+ CXCR5+ TFH cellsexpressed a higher level of BCL6 mRNA than CD4+CXCR52 Tcells by real-time PCR (*P,0.05). (D) Splenocytes were isolatedfrom rats with HN and TCV, and then, sorted CD8+ T cells wereanalyzed by flow cytometry (purity.92%). The sorted CD8+ Tcells were then stained with CD11b/c with limited CD11b/c ex-pression. (E) Flow cytometric analysis showed TFH cells (CXCR5+CD4+ T cells) increased to 12% after HN induction comparedwith normal rats (5% CXCR5+ of total CD4+ T cells); there is a sig-nificant decrease in TFH numbers in HN rats receiving TCV in-duced with Fx1A. However, HN rats receiving TCV inducedwith CFA had a similar increase in TFH cells (10%) as HN ratsalone (n=6).
1064 Journal of the American Society of Nephrology J Am Soc Nephrol 23: 1058–1067, 2012
BASIC RESEARCH www.jasn.org
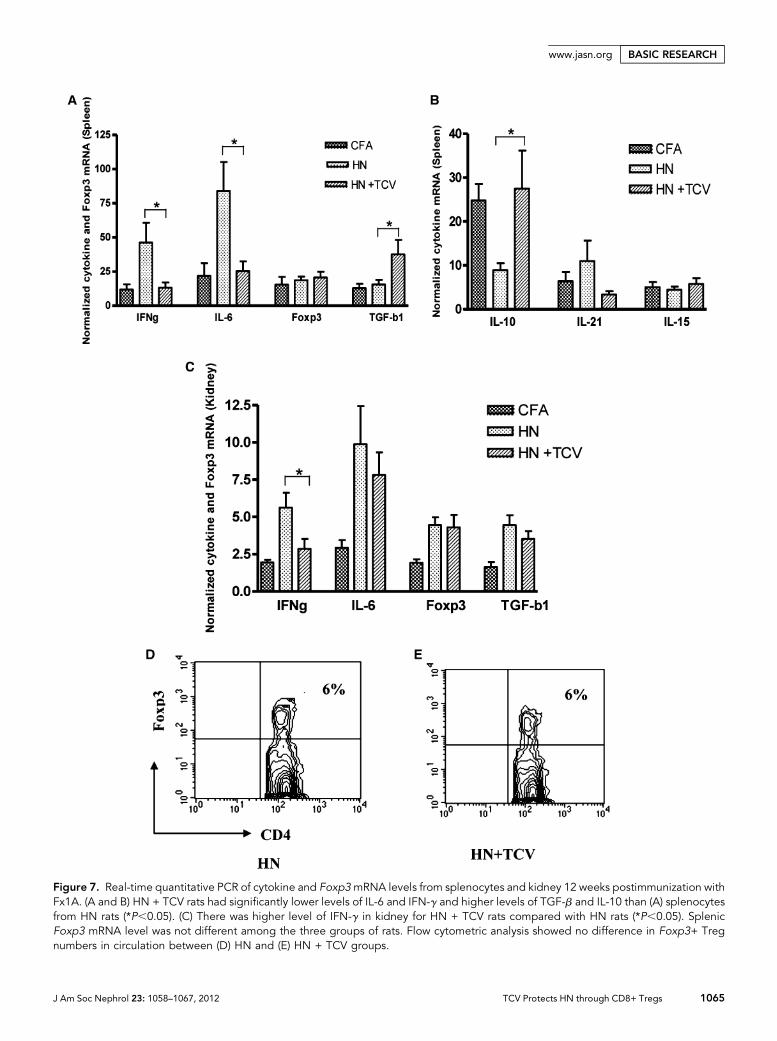
Figure 7. Real-time quantitative PCR of cytokine and Foxp3mRNA levels from splenocytes and kidney 12 weeks postimmunization withFx1A. (A and B) HN + TCV rats had significantly lower levels of IL-6 and IFN-g and higher levels of TGF-b and IL-10 than (A) splenocytesfrom HN rats (*P,0.05). (C) There was higher level of IFN-g in kidney for HN + TCV rats compared with HN rats (*P,0.05). SplenicFoxp3 mRNA level was not different among the three groups of rats. Flow cytometric analysis showed no difference in Foxp3+ Tregnumbers in circulation between (D) HN and (E) HN + TCV groups.
J Am Soc Nephrol 23: 1058–1067, 2012 TCV Protects HN through CD8+ Tregs 1065
www.jasn.org BASIC RESEARCH

UK). The secondary antibody was biotinylated goat anti-mouse
Ig (BD Biosciences, Australia). Immunohistochemical staining was
performed, and interstitial infiltration was assessed as described
previously.35
In Vitro Cytotoxicity Assay of CD8 T Cells againstCD4 T Cell TargetsCD4+ and CD8+ cells were purified from normal, TCV, or HN rats as
indicated. The target cells were either Qa-1–expressing or Qa-1–non-
expressing CD4+ T cells. CD8+ T cells isolated from both TCV and
normal rats were tested in two different groups. Specificity for Qa-1
was tested using blocking antibodies. CD4+ T cells isolated from TCV
were stained with biotin-conjugated Qa-1 monoclonal antibody fol-
lowed by Streptavidin-PE, and Qa-1–positive CD4+ T cells were then
sorted by flow cytometry according to the level of Qa-1expression on
their surface. Qa-1 high and low CD4+ T cells labeled with CFSE low
or normal CD4+ T cells labeled with CFSE high were cultured alone
or cocultured with TCVCD8+ T cells for 4 days. In blocking experi-
ments, coculture was performed in the presence of mouse anti-rat
Qa-1 antibody. CD4+ cells were measured with FACS for their Qa-1
expression or sorted according to the level of Qa-1 expression on
their surface.
Real-Time PCR for IL-6, Foxp3, IFN-g, and TGF-bTotal RNA was isolated from rat spleen and was reverse-transcribed.35
cDNAwas subjected to quantitative PCR analysis using commercial pri-
mers and Taqman probes specific for IL-6, Foxp3, IFN-g, TGF-b,
and glyceraldehyde-3-phosphate dehydrogenase (Applied Biosystem,
Australia)35;n=6 fromeach group, andeach samplewas run in triplicate.
Statistical AnalysesStatistical analyses were performed using one-way ANOVA for
multiple comparisons. Results are expressed as the group mean 6
SD. Two group differences were analyzed by t test, with a P value
(two-tailed) ,0.05 considered statistically significant.
ACKNOWLEDGMENTS
We thank Prof. Carola Vinuesa of ANU for advice.We thank the animal
house staff at the Children’s Hospital Research Institute for care of the
animals. We thank Mary Sartor and Sanda Lum for help in cell sorting.
This workwas supported byNationalHealth andMedical Research
Council of Australia Grants 249414 and 457408.
DISCLOSURESNone.
REFERENCES
1. Akbar AN, Vukmanovic-Stejic M, Taams LS, Macallan DC: The dynamicco-evolution of memory and regulatory CD4+ T cells in the periphery.Nat Rev Immunol 7: 231–237, 2007
2. Cantor H, Shen FW, Boyse EA: Separation of helper T cells from sup-pressor T cells expressing different Ly components. II. Activation byantigen: After immunization, antigen-specific suppressor and helperactivities are mediated by distinct T-cell subclasses. J Exp Med 143:1391–1440, 1976
3. Wang YM, Alexander SI: CD8 regulatory T cells: What’s old is now new.Immunol Cell Biol 87: 192–193, 2009
4. Jiang H, Kashleva H, Xu LX, Forman J, Flaherty L, Pernis B, BraunsteinNS, Chess L: T cell vaccination induces T cell receptor Vbeta-specificQa-1-restricted regulatory CD8(+) T cells. Proc Natl Acad Sci USA 95:4533–4537, 1998
5. Panoutsakopoulou V, Huster KM, McCarty N, Feinberg E, Wang R,Wucherpfennig KW, Cantor H: Suppression of autoimmune diseaseafter vaccination with autoreactive T cells that express Qa-1 peptidecomplexes. J Clin Invest 113: 1218–1224, 2004
6. Kim HJ, Verbinnen B, Tang X, Lu L, Cantor H: Inhibition of follicularT-helper cells by CD8(+) regulatory T cells is essential for self tolerance.Nature 467: 328–332, 2010
7. Lu L, Werneck MB, Cantor H: The immunoregulatory effects of Qa-1.Immunol Rev 212: 51–59, 2006
8. Kumar V: Homeostatic control of immunity by TCR peptide-specificTregs. J Clin Invest 114: 1222–1226, 2004
9. Cantor H, Hugenberger J, McVay-Boudreau L, Eardley DD, Kemp J,Shen FW, Gershon RK: Immunoregulatory circuits among T-cell sets.Identification of a subpopulation of T-helper cells that induces feed-back inhibition. J Exp Med 148: 871–877, 1978
10. Hu D, Ikizawa K, Lu L, Sanchirico ME, Shinohara ML, Cantor H: Analysisof regulatory CD8 T cells in Qa-1-deficient mice. Nat Immunol 5: 516–523, 2004
11. Correale J, Farez M, Gilmore W: Vaccines for multiple sclerosis: Prog-ress to date. CNS Drugs 22: 175–198, 2008
12. Fujii T, OkadaM, Fujita Y, Sato T, TanakaM, Usui T, Umehara H,MimoriT: Vaccination with autoreactive CD4(+)Th1 clones in lupus-proneMRL/Mp-Fas(lpr/lpr) mice. J Autoimmun 33: 125–134, 2009
13. Tang X, Maricic I, Purohit N, Bakamjian B, Reed-Loisel LM, Beeston T,Jensen P, Kumar V: Regulation of immunity by a novel population ofQa-1-restricted CD8alphaalpha+TCRalphabeta+ T cells. J Immunol
177: 7645–7655, 200614. Wu Y, Zheng Z, Jiang Y, Chess L, Jiang H: The specificity of T cell
regulation that enables self-nonself discrimination in the periphery.Proc Natl Acad Sci USA 106: 534–539, 2009
15. ChenW, Zhang L, Liang B, Saenger Y, Li J, Chess L, Jiang H: Perceivingthe avidity of T cell activation can be translated into peripheral T cellregulation. Proc Natl Acad Sci USA 104: 20472–20477, 2007
16. Loftus B, Newsom B, Montgomery M, Von Gynz-Rekowski K, Riser M,Inman S, Garces P, Rill D, Zhang J, Williams JC: Autologous attenuatedT-cell vaccine (Tovaxin) dose escalation in multiple sclerosis relapsing-remitting and secondary progressive patients nonresponsive to ap-proved immunomodulatory therapies.Clin Immunol131: 202–215, 2009
17. Correale J, Lund B, McMillan M, Ko DY, McCarthy K, Weiner LP: T cellvaccination in secondary progressive multiple sclerosis. J Neuro-
immunol 107: 130–139, 200018. Medaer R, Stinissen P, Truyen L, Raus J, Zhang J: Depletion of myelin-
basic-protein autoreactive T cells by T-cell vaccination: Pilot trial inmultiple sclerosis. Lancet 346: 807–808, 1995
19. Trivedi S, Zang Y, Culpepper S, Rosenbaum E, Fernandez I, Martinez L,Hoffman RW, Greidinger EL: T cell vaccination therapy in an inducedmodel of anti-RNP autoimmune glomerulonephritis.Clin Immunol 137:281–287, 2010
20. Cattran D: Management of membranous nephropathy: When andwhat for treatment. J Am Soc Nephrol 16: 1188–1194, 2005
21. Beck LH Jr, Bonegio RG, LambeauG, BeckDM, Powell DW, Cummins TD,Klein JB, Salant DJ: M-type phospholipase A2 receptor as target an-tigen in idiopathicmembranous nephropathy.NEngl JMed 361: 11–21,2009
1066 Journal of the American Society of Nephrology J Am Soc Nephrol 23: 1058–1067, 2012
BASIC RESEARCH www.jasn.org

22. Lee G: End-stage renal disease in the Asian-Pacific region. SeminNephrol 23: 107–114, 2003
23. HeymannW, Hackel DB, Harwood S, Wilson SG, Hunter JL: Productionof nephrotic syndrome in rats by Freund’s adjuvants and rat kidneysuspensions. Proc Soc Exp Biol Med 100: 660–664, 1959
24. Penny MJ, Boyd RA, Hall BM: Permanent CD8(+) T cell depletion pre-vents proteinuria in active Heymann nephritis. J Exp Med 188: 1775–1784, 1998
25. Penny MJ, Boyd RA, Hall BM: Role of T cells in the mediation ofHeymann nephritis. ii. Identification of Th1 and cytotoxic cells in glo-meruli. Kidney Int 51: 1059–1068, 1997
26. Quiza CG, Leenaerts PL, Hall BM: The role of T cells in the mediation ofglomerular injury in Heymann’s nephritis in the rat. Int Immunol 4: 423–432, 1992
27. Schiller B, He C, Salant DJ, Lim A, Alexander JJ, Quigg RJ: Inhibition ofcomplement regulation is key to the pathogenesis of active Heymannnephritis. J Exp Med 188: 1353–1358, 1998
28. Spicer ST, TranGT, KillingsworthMC, Carter N, Power DA, Paizis K, BoydR, Hodgkinson SJ, Hall BM: Induction of passive Heymann nephritis incomplement component 6-deficient PVG rats. J Immunol 179: 172–178,2007
29. WuH,WaltersG,Knight JF,AlexanderSI:DNAvaccination against specificpathogenic TCRs reduces proteinuria in active Heymann nephritis by in-ducing specific autoantibodies. J Immunol 171: 4824–4829, 2003
30. Wu H, ZhangGY, Knight JF: T cell receptor BV gene usage in interstitialcellular infiltrates in active Heymann nephritis.Nephrol Dial Transplant16: 1374–1381, 2001
31. Walters G, Wu H, Knight JF: Glomerular T cells in Heymann nephritis.Clin Exp Immunol 126: 319–325, 2001
32. Friese MA, Fugger L: Pathogenic CD8(+) T cells in multiple sclerosis.Ann Neurol 66: 132–141, 2009
33. Smith TR, Kumar V: Revival of CD8+ Treg-mediated suppression.Trends Immunol 29: 337–342, 2008
34. Lu L, Cantor H: Generation and regulation of CD8(+) regulatory T cells.Cell Mol Immunol 5: 401–406, 2008
35. Wang YM, Zhang GY, Wang Y, Hu M, Wu H, Watson D, Hori S,Alexander IE, Harris DC, Alexander SI: Foxp3-transduced polyclonalregulatory T cells protect against chronic renal injury from adriamycin.J Am Soc Nephrol 17: 697–706, 2006
36. Madakamutil LT, Maricic I, Sercarz E, Kumar V: Regulatory T cells con-trol autoimmunity in vivo by inducing apoptotic depletion of activatedpathogenic lymphocytes. J Immunol 170: 2985–2992, 2003
37. de Heer E, Daha MR, van Es LA: Lymph node cells from rats withHeymann’s nephritis produce in vitro autoantibodies directed againstpurified renal tubular antigen. Immunology 52: 743–752, 1984
38. Edgington TS, Glassock RJ, Dixon FJ: Autologous immune complexnephritis induced with renal tubular antigen. I. Identification and isolationof the pathogenetic antigen. J Exp Med 127: 555–572, 1968
J Am Soc Nephrol 23: 1058–1067, 2012 TCV Protects HN through CD8+ Tregs 1067
www.jasn.org BASIC RESEARCH




















