
Beyond immunosuppression - challenges in the clinical management of lupus nephritis
Upload
independentCategory
view
1download
0
Autoimmunity Reviews xxx (2015) xxx–xxx
AUTREV-01704; No of Pages 11
Contents lists available at ScienceDirect
Autoimmunity Reviews
j ourna l homepage: www.e lsev ie r .com/ locate /aut rev
Review
Multi-antibody composition in lupus nephritis: Isotype and antigenspecificity make the difference
Alice Bonanni a, Augusto Vaglio c, Maurizio Bruschi b, Renato Alberto Sinico d, Lorenzo Cavagna e,Gabriella Moroni f, Franco Franceschini g, Landino Allegri c, Federico Pratesi h, Paola Migliorini h,Giovanni Candiano b, Giampaola Pesce i, Angelo Ravelli k, Francesco Puppo j, Alberto Martini k,⁎,Angela Tincani g, Gian Marco Ghiggeri a,b,⁎⁎a Division of Nephrology, Dialysis, and Transplantation, Scientific Institute for Research and Health Care (IRCCS), Istituto Giannina Gaslini, Genoa, Italyb Laboratory on Pathophysiology of Uremia, Scientific Institute for Research and Health Care (IRCCS), Istituto Giannina Gaslini, Genoa, Italyc Department of Clinical and Experimental Medicine, University of Parma, Parma, Italyd Division of Nephrology and Section of Clinical Immunology, San Carlo Hospital, Milan, Italye Division of Rheumatology, University and IRCCS Policlinico S. Matteo, Pavia, Italyf Division of Nephrology and Dialysis, Scientific Institute for Research and Health Care (IRCCS), Regina Elena, Milan, Italyg Rheumatology and Clinical Immunology, Spedali Civili and University of Brescia, Italyh Clinical Immunology Unit, Department of Internal Medicine, University of Pisa, Italyi Medical and Radiometabolic Therapy Unit, Department of Internal Medicine, University of Genoa, Italyj Department of Internal Medicine, University of Genoa, Italyk Division of Paediatric Rheumatology, Scientific Institute for Research and Health Care (IRCCS), Istituto Giannina Gaslini, Genoa, Italy
⁎ Correspondence to: Alberto Martini, Pediatrics, UnivePediatrics IRCCS G. Gaslini, Largo G. Gaslini 5, 16147 Geno⁎⁎ Correspondence to: GianMarco Ghiggeri, Division of NItaly. Tel.: +39 10 380742; fax: +39 10 395214.
E-mail addresses: [email protected].
http://dx.doi.org/10.1016/j.autrev.2015.04.0041568-9972/© 2015 Published by Elsevier B.V.
Please cite this article as: Bonanni A, et al, MAutoimmun Rev (2015), http://dx.doi.org/10
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 March 2015Accepted 2 April 2015Available online xxxx
Keywords:AutoimmunitySystemic lupus erythematosusLupus nephritisAnti-DNA antibodiesAnti-αenolase antibodiesAnti-annexin AI antibodies
Research on autoimmune processes involved in glomerulonephritis has been for years based on experimentalmodels. Recent progress in proteomics has radicallymodified perspectives: lasermicrodissection and proteomicswere crucial for an in vivo analysis of autoantibodies eluted from human biopsies. Lupus nephritis has been thesubject of recent independent researches.Main topics have been the definition of renal autoimmune components in human lupus biopsies; methods werelaser capture of glomeruli and/or of single cells (CD38+ or Ki-67+) from tubulointerstitial areas as starting stepfollowed by elution and characterization of renal antibodies by proteomics.The innovative approach highlighted different panels of autoantibodies deposited in glomeruli and in tubulo-interstitial areas that actually represented the unique autoimmune components in these patients. IgG2 was themajor isotype; new podocyte proteins (αenolase, annexin AI) and already known implanted molecules (DNA,histone 3, C1q) were their target antigens in glomeruli. Vimentin was the antigen in tubulo-interstitial areas.Matching renal autoantibodies with serum allowed the definition of a typical autoantibody serum map that in-cluded the same anti-αenolase, anti-annexin AI, anti-DNA, and anti-histone 3 IgG2 already detected in renal tis-sue. Serum levels of specific autoantibodies were tenfold increased in patients with lupus nephritis allowing aclear differentiation from both rheumatoid arthritis and other glomerulonephritis. In all cases, targeted antigenswere characterized as components of lupus NETosis.Matching renal/serum autoantibody composition in vivo furnishes new insights on human lupus nephritis andallows to refine composition of circulating antibodies in patients with lupus. A thoughtful passage from benchto bedside of new knowledge would expand our clinical and therapeutic opportunities.
© 2015 Published by Elsevier B.V.
rsity of Genova, Head Pediatria II, Reumatologia, EULAR Centre of Excellence in Rheumatology 2008-18, Head Department ofva, Italy. Tel.: +39 10 5636386; fax: +39 10 5636211.ephrology, Dialysis and Transplantation, Laboratory on Pathophysiology of Uremia, Istituto G. Gaslini, Largo G. Gaslini 5, Genoa,
it (A. Martini), [email protected] (G.M. Ghiggeri).
ulti-antibody composition in lupus nephritis: Isotype and antigen specificity make the difference,.1016/j.autrev.2015.04.004

2 A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02. ‘In vivo’ renal antibody characterization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
2.1. General consideration and methodology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02.2. Fields of application . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
3. Lupus nephritis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03.1. Anti-vimentin antibodies predominate in the tubulointerstitial compartment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03.2. Antigen targets and isotype specificity match glomerular and serum antibodies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03.3. Isotype preponderance and immunoglobulin shift . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03.4. Epitope spreading determines αenolase IgG2 reactivity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
4. Alpha-enolase and annexin AI are functionally linked with autoimmunity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04.1. Alpha-enolase and annexin AI are components of NETosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04.2. Functional consequences of alpha-enolase block . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04.3. Functional consequences of annexin AI block . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
5. Animal models support multi-antibodies and isotype specificity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06. Clinical and pathological impact . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
6.1. Mechanisms of autoantibody formation and deposition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.2. Hierarchy and time of appearance of antibodies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.3. How this modifies the future of clinical research on lupus auto-antibodies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.4. Prospective studies: the Zeus project . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
7. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Take-home messages . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Author contribution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Conflict of interest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
1. Introduction
Autoimmune renal diseases represent a relevant source of morbidityand mortality in human beings [1]. They include several conditions withdifferent clinical expressions, at the basis of which there is a commonmechanism linked to the formation of auto-antibodies versus renal pro-teins. Renal basement membrane and podocytes are the principal targetsbut also tubular cells may become the site for autoantibody deposition.Secondary events are complement deposition, mainly C5b-9 [2–6], andactivation of podocyte intracellular mediators such as phosphorylatedprotein kinase C and free oxygen radicals [7]. The imbalance between in-flammatorymediators and their inhibitors determines podocyte injury [8,9]. Lupus nephritis and membranous nephritis are the two autoimmuneconditions of major interest due to similarities in pathologic pictures(bothmembranous and class V lupus nephritis present sub-epithelial im-mune deposits) and potentially they share mechanisms. Anti-GBM is an-other rare autoimmune glomerulonephritis that is completely entailed bydeposition of antibodies versus the NC1 domain of collagen IV causing re-cruitment of neutrophils and monocytes, destruction of the glomerularcapillary wall and crescent formation [10]. While the target antigen andmechanisms of anti-GBMhave been elucidated in the past,most of the as-pects linking autoimmunity and renal damage in membranous and inlupus nephritis are of only recent evolution and are here reviewed inthe light of most recent discoveries. New advances in the field of tissuemicro-dissection and protein characterization by mass spectrometryrepresented the technology keys that allowed a direct in vivo approachto human biopsies. The starting consideration was that only auto-antibodies present in the kidney could be considered pathogenic andmatching the auto-antigen/antibody renal map with serum antibodieswas crucial for developing new concepts on mechanisms and make newbiomarkers available for patients with lupus nephritis.
2. ‘In vivo’ renal antibody characterization
2.1. General consideration and methodology
A simplified interpretation of the events leading to autoantibody for-mation and deposition in the kidney considers that renal proteins may
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
become immunogenic. In the case of components normally present incytoplasm of cells, their dislocation to membrane may also contribute.Cellular mechanisms, potentially involved in human pathologies, arebarely evaluated with traditional techniques of analysis and manyrenal diseases, different in pathogenesis, prognosis and treatmenthave similar, often undistinguishable, microscopical patterns. Experi-mental models are helpful but there are cases, such asmembranous ne-phropathy, in which mechanisms in animals do not reproduce humanpathology [11–13]. Studies on Heymann nephritis (the experimentalmodel of human membranous nephropathy) were critical to show[11,12,14,15] IgG4 prevalence and complement participation to im-mune deposit formation, as well as podocyte activation and productionof pathogenetic mediators such as reactive oxygen species [2,4,16,17].However, they did not result in the identification of the human targetantigen of membranous nephropathy since megalin, the podocyte anti-gen in mice, is not expressed in human podocytes. After some decadesof substantial no progress, developments of methods for a direct analy-sis of human tissue [18–22] allowed a direct definition of autoantibodiesin human biopsies. Glomeruli (or tubule, or any other structure) canbe precisely isolated by laser capture microdissection, a techniquethat allows the collection of sufficient amount of tissue from thesame patient thus permitting a personalized quantitative character-ization of different auto-antibodies/antigens. Mass spectrometryevolution is basic to this approach. In this way, the composition ofglomerular proteins can be determined, creating a sort of “glomeru-lar proteome map” for different diseases. Antibodies can be micro-eluted as well, allowing the definition of their target antigens, class,isotype and epitope spreading [22].
Evaluation of a potential renal auto-antigen/antibody requiresthe fulfillment of the following criteria: 1) the antigen must beexpressed in the kidney (glomeruli, tubule) in concomitancewith the pathological condition; 2) the antibody must be elutedfrom microdissected tissues/cells in vivo; 3) co-localization of an-tigen and antibody in immune deposits must be documented byimmunohistochemistry and confocal microscopy and, possibly,by immune-electron microscopy; and 4) elevated circulating anti-bodies of the same class and isotype should be documented inserum.
lupus nephritis: Isotype and antigen specificity make the difference,

3A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
2.2. Fields of application
Membranous nephritis and systemic lupus erythematosus are thetwo conditions investigated with the use of new technology ap-proaches. Clinical impact and potential similarities in pathogenesis be-tween the two conditions constituted the main reason for the choice.In fact, typical of membranous nephropathy, that is considered the‘model’ of any autoimmune nephritis [23], is the presence of sub-epithelial glomerular electron-dense deposits constituted mainly byIgG4 and C5b-9 [24] plus clusterin that potentially inhibits the formertwo [8,9]. The same pathology pattern characterized by sub-epithelialimmune deposits of membranous nephropathy is the hallmark of classV lupus nephritis, which creates a direct link between the twopathologies.
In primary membranous nephropathy, definition of glomerularauto-antigens implicated in the formation of sub-epithelial deposits“in vivo” represented a real break-through considering that for almost40 years no hypothesis on antibody composition of human cases hadbeen formulated. Several antigens/antibodies have been now recog-nized (i.e. anti-PLA2r, anti-THSD7A, anti-NEP, anti-cationic BSA, anti-AR, anti-SOD2 and anti-αenolase) and the topics are rapidly evolvingsince, in just 4 years, we passed from the complete lack of idea to 5 can-didate auto-antigens [18–21,25,26]. Research is now progressing in thefield of prediction of the clinical outcome and includes the control oftherapeutic approaches [27].
Organ specific antigens potentially determine the site of target insultin systemic lupus erythematosus. Lupus nephritis, the topic of this re-view, has been investigated in details only recently by two independentgroups [28–30]. Kinloch and colleagues [28] utilized laser capture to iso-late inflammatory cells from tubulo-interstitial area in 14 biopsy speci-mens that were clonally expanded; immunoglobulins recognizedspecific antigens. Bruschi and colleagues [29,30] microdissected glo-meruli from renal biopsies and eluted new antibodies. The objectivewas to create an antibody-map specific for human lupus nephritis utiliz-ing the in vivo approach as indicated above.
4 9non linear pH
1 2 3
4
A
BC E
G
I
200
10
MrkDa
4 9non linear pH
12
3
4
AB
CE
G
H
b)
I
a)serum
antibodiesglomerularantibodies
H
Fig. 1.Characterization of auto-antibodies eluted fromglomeruli of patientswith lupus ne-phritis and in corresponding serum. (a) Western blot analysis antibodies eluted frommi-crodissected glomeruli obtained from patients with lupus nephritis; equal amount ofantibodies deriving from different lupus nephritis classes III–V were pooled and utilizedfor western-blot. The reacting antibodies were then developed with anti-IgG (total) anti-bodies. Incubations with antibodies deriving from separate lupus nephritis were per-formed as well [29]. Microeluted antibodies from normal kidneys did not react with anyprotein. Several spots were recognized by lupus nephritis elutaes and were identified as11 different proteins (A, B, C, D, E, F, G, H, K, I, J) (for corresponding identification seeTable 1). One protein was characterized by liquid mass spectrometry (spot I, αenolase),the remaining spots were characterized by MALDI. (b) The same podocyte cell extractsseparated by two-dimensional electrophoresis were incubated with normal sera (notshown) andwith pooled lupus nephritis sera (classes III–V). Several proteinswere detect-ed, most corresponding to proteins recognized in glomerular eluates (A, B, C, E, G, H, I);three proteins were recognized only in serum or in few glomerular elutaes (K, L). Allspots were characterized by liquid mass spectrometry or MALDI as above.
3. Lupus nephritis
Lupus nephritis is one of themost severe manifestations of systemiclupus erythematosus that may evolve to end-stage renal disease if notrecognized and treated [31]; subtle clinical signs at the onset limitchances of early recognition that are, however, crucial to block, intime, disease progression. It is a prototype of any autoimmune condi-tion [32,33] with notable clinical importance due to a high incidence[34]. The development of biomarkers of activity of lupus nephritis hasbeen so far disappointing mainly because it was based on tentative ap-proaches performed without knowing the exact composition of depos-ited auto-antibodies in glomeruli. Moreover, most of our currentknowledge is based on experimental models of systemic lupus erythe-matosus and of lupus nephritis [35–37] that are only partially relatedto human beings. Actually, animal models determine single auto-antibody deposition, whereas there is consensus to consider lupusnephritis as a multi-antibody autoimmune condition. So far, studiesbased on pathological analysis of renal tissues showed the presenceof antibodies versus implanted antigens (such as DNA/histones/nucleosome) deriving from an early cell breakdown and C1q that isnot commonly detected in glomeruli [38,37]. A second class of antibod-ies that are directed to podocyte antigens [39–41] has been hypothe-sized on the basis of their presence in serum but have never beendocumented in glomeruli. It has been estimated that anti-DNA deposi-tion in lupus nephritis accounts for no more than 10–20% of overalleluted IgG overall [42] implying that IgG not recognizing DNA representthe vast majority of antibodies in glomeruli. Less emphasis hashistorically been paid to the tubulo-interstitial compartment that repre-sents, instead, an important area for ‘immunologic activity’ in most
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
glomerulonephritis often correlating with fibrosis and worsening ofrenal function [43,44].
The in vivo approaches to human lupus nephritis were aimed to fillup these gaps [28–30]. Renal biopsies were analyzed utilizing differentstrategies for glomeruli and tubulo-interstitium; results highlightingnew important concepts in both cases.
3.1. Anti-vimentin antibodies predominate in the tubulointerstitialcompartment
As already pointed out, in the study by Kinloch and colleagues[28] single cells (CD38+ or Ki-67+) were laser-captured fromtubulointerstitial areas of 15 renal biopsies of patients with lupus ne-phritis and were cloned. Overall, 25 immunoglobulins were expand-ed and characterized utilizing an array of techniques (confocalmicroscopy, ELISA, screening protoarrays, mass spectrometry).Vimentin was the antigen identified in most cases and anti-vimentinIgGs preferentially stained inflamed areas in tubulo-interstitium ofmany biopsies. Circulating levels of anti-vimentin IgGs were high in pa-tients with tubulointerstitial inflammation. The study is of interest andnew; results furnish an attractive option to separate patients with dif-ferent renal lesions on the basis of a circulating biomarkers that couldbe also utilized to monitor progression of renal lesions in patients un-dergoing therapies.
3.2. Antigen targets and isotype specificity match glomerular and serumantibodies
The approach to glomerular antibodies was different from above:frozen renal fragments were processed with laser-capture micro-dissection and antibodies eluted from micro-dissected glomeruli werecharacterized with western-blot and mass spectrometry [29,30]. Anti-bodies versus both implanted antigens (DNA, histones, C1q) andpodocyte antigens (11 antigens detected) were characterized; serumlevels of the same antibodies found in glomeruli were determinedwith specific ELISA in large cohorts of lupus nephritis, systemic lupus er-ythematosus patients and compared to rheumatoid arthritis and otherglomerulonephritis, overall considering more than 500 sera (Fig. 1 and
lupus nephritis: Isotype and antigen specificity make the difference,
ntibod
yleve
lswerede
term
ined
forev
eryisotyp
ewithspecificEL
ISAsan
dwereex
pressedas
mgpe
rliter.
RAn50
CTRn10
0
vs.LN
Med
ian
mg/l
Interqua
rtile
N95
°Pvs.LN
Med
ian
mg/l
Interqua
rtile
95°
Pvs.LN
0.03
0.01
00.44
0.37
0.79
60.00
13.67
8.42
8b0.00
15.43
10.57
15.82
b0.00
10.13
0.01
0b0.00
12.05
2.14
3.80
–
0.56
0.01
0b0.00
12.51
2.14
4.30
–
0.00
12.08
0.90
9b0.01
1.31
0.36
2.71
b0.00
10.01
050.00
41
b0.01
0.01
360.00
40.02
–
0.04
50.22
14–
0.03
870.05
30.15
b0.05
0.01
10.04
020
–0.01
20.00
10.01
3–
0.01
0.00
30.04
615
(30%
)–
0.00
30.00
10.00
4–
0.00
10.19
20.42
27(5
2%)
b0.01
0.03
60.05
0.14
b0.00
10.01
10.08
19(3
8%)
b0.05
0.01
10.00
40.01
2b0.00
10.00
10.25
00.45
31–
0.05
40.08
0.11
5b0.00
10.05
0.00
70.00
10
b0.05
0.00
80.00
10.00
8–
0.00
10.13
30.04
30(6
0%)
b0.05
0.11
30.01
0.12
6b0.00
10.00
10.52
80.10
31(6
2%)
b0.05
0.46
90.04
0.50
8b0.00
10.00
10.05
40.01
2b0.00
10.04
70.01
0.06
4b0.00
10.00
10.23
30.02
8b0.00
10.20
80.04
0.24
9b0.00
1
4 A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
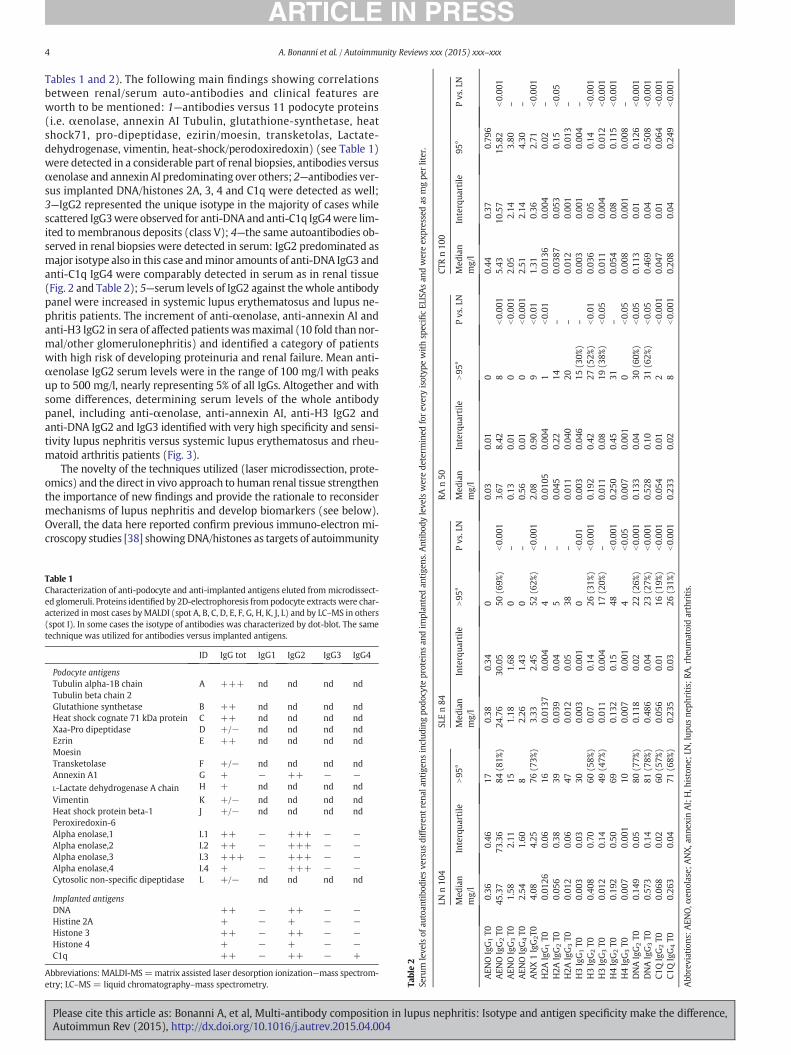
Tables 1 and 2). The following main findings showing correlationsbetween renal/serum auto-antibodies and clinical features areworth to be mentioned: 1—antibodies versus 11 podocyte proteins(i.e. αenolase, annexin AI Tubulin, glutathione-synthetase, heatshock71, pro-dipeptidase, ezirin/moesin, transketolas, Lactate-dehydrogenase, vimentin, heat-shock/perodoxiredoxin) (see Table 1)were detected in a considerable part of renal biopsies, antibodies versusαenolase and annexin AI predominating over others; 2—antibodies ver-sus implanted DNA/histones 2A, 3, 4 and C1q were detected as well;3—IgG2 represented the unique isotype in the majority of cases whilescattered IgG3were observed for anti-DNA and anti-C1q IgG4were lim-ited to membranous deposits (class V); 4—the same autoantibodies ob-served in renal biopsies were detected in serum: IgG2 predominated asmajor isotype also in this case andminor amounts of anti-DNA IgG3 andanti-C1q IgG4 were comparably detected in serum as in renal tissue(Fig. 2 and Table 2); 5—serum levels of IgG2 against thewhole antibodypanel were increased in systemic lupus erythematosus and lupus ne-phritis patients. The increment of anti-αenolase, anti-annexin AI andanti-H3 IgG2 in sera of affected patientswasmaximal (10 fold thannor-mal/other glomerulonephritis) and identified a category of patientswith high risk of developing proteinuria and renal failure. Mean anti-αenolase IgG2 serum levels were in the range of 100 mg/l with peaksup to 500 mg/l, nearly representing 5% of all IgGs. Altogether and withsome differences, determining serum levels of the whole antibodypanel, including anti-αenolase, anti-annexin AI, anti-H3 IgG2 andanti-DNA IgG2 and IgG3 identified with very high specificity and sensi-tivity lupus nephritis versus systemic lupus erythematosus and rheu-matoid arthritis patients (Fig. 3).
The novelty of the techniques utilized (laser microdissection, prote-omics) and the direct in vivo approach to human renal tissue strengthenthe importance of new findings and provide the rationale to reconsidermechanisms of lupus nephritis and develop biomarkers (see below).Overall, the data here reported confirm previous immuno-electron mi-croscopy studies [38] showingDNA/histones as targets of autoimmunity
Table 1Characterization of anti-podocyte and anti-implanted antigens eluted from microdissect-ed glomeruli. Proteins identified by 2D-electrophoresis frompodocyte extracts were char-acterized inmost cases byMALDI (spot A, B, C, D, E, F, G, H, K, J, L) and by LC–MS in others(spot I). In some cases the isotype of antibodies was characterized by dot-blot. The sametechnique was utilized for antibodies versus implanted antigens.
ID IgG tot IgG1 IgG2 IgG3 IgG4
Podocyte antigensTubulin alpha-1B chain A +++ nd nd nd ndTubulin beta chain 2Glutathione synthetase B ++ nd nd nd ndHeat shock cognate 71 kDa protein C ++ nd nd nd ndXaa-Pro dipeptidase D +/− nd nd nd ndEzrin E ++ nd nd nd ndMoesinTransketolase F +/− nd nd nd ndAnnexin A1 G + − ++ − −L-Lactate dehydrogenase A chain H + nd nd nd nd
Vimentin K +/− nd nd nd ndHeat shock protein beta-1 J +/− nd nd nd ndPeroxiredoxin-6Alpha enolase,1 I.1 ++ − +++ − −Alpha enolase,2 I.2 ++ − +++ − −Alpha enolase,3 I.3 +++ − +++ − −Alpha enolase,4 I.4 + − +++ − −Cytosolic non-specific dipeptidase L +/− nd nd nd nd
Implanted antigensDNA ++ − ++ − −Histine 2A + − + − −Histone 3 ++ − ++ − −Histone 4 + − + − −C1q ++ − ++ − +
Abbreviations: MALDI-MS=matrix assisted laser desorption ionization—mass spectrom-etry; LC–MS = liquid chromatography–mass spectrometry. Ta
ble2
Serum
leve
lsof
autoan
tibo
dies
versus
diffe
rent
rena
lantigen
sinclud
ingpo
docyte
proteins
andim
plan
tedan
tige
ns.A
LNn10
4SL
En84
Med
ian
mg/l
Interqua
rtile
N95
°Med
ian
mg/l
Interqua
rtile
N95
°P
AEN
OIgG1T0
0.36
0.46
170.38
0.34
0AEN
OIgG2T0
45.37
73.36
84(8
1%)
24.76
30.05
50(6
9%)
b
AEN
OIgG3T0
1.58
2.11
151.18
1.68
0–
AEN
OIgG4T0
2.54
1.60
82.26
1.43
0–
ANX1IgG2T0
4.08
4.25
76(7
3%)
3.33
2.45
52(6
2%)
b
H2A
IgG1T0
0.01
260.06
160.01
370.00
44
–
H2A
IgG2T0
0.05
60.38
390.03
90.04
5–
H2A
IgG3T0
0.01
20.06
470.01
20.05
38–
H3IgG1T0
0.00
30.03
300.00
30.00
10
b
H3IgG2T0
0.40
80.70
60(5
8%)
0.07
0.14
26(3
1%)
b
H3IgG3T0
0.01
20.14
49(4
7%)
0.01
10.00
417
(20%
)–
H4IgG2T0
0.19
20.50
690.13
20.15
48b
H4IgG3T0
0.00
70.00
110
0.00
70.00
14
b
DNAIgG2T0
0.14
90.05
80(7
7%)
0.11
80.02
22(2
6%)
b
DNAIgG3T0
0.57
30.14
81(7
8%)
0.48
60.04
23(2
7%)
b
C1QIgG2T0
0.06
80.02
60(5
7%)
0.05
60.01
16(1
9%)
b
C1QIgG4T0
0.26
30.04
71(6
8%)
0.23
50.03
26(3
1%)
b
Abb
reviations
:AEN
O,α
enolase;
ANX,ann
exin
AI;H,h
istone
;LN,lup
usne
phritis;
RA,rhe
umatoidarthritis.
Please cite this article as: Bonanni A, et al, Multi-antibody composition in lupus nephritis: Isotype and antigen specificity make the difference,Autoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004

Fig. 2. (a) Characterization of isotypes of serumantibodies versus both implanted antigens(histones, DNA, C1q) and two major podocyte antigens (anti-αenolase and anti-annexinAI). Isotypes were characterized with dot-blot analysis utilizing recombinant proteins/DNA linked to nitrocellulose as antigen; results were evaluated as signal intensity ofchemiluminescence detected by VersaDoc and computed with QuantyOne software,Bio-Rad. These results were preparative for specific ELISA for determination of serumlevels (see Table 2).
5A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
in human lupus nephritis. The implication of annexin AI was reportedfor peculiar autoimmune conditions such as discoid lupus [45] andanti-phospholipid syndrome. Results on αenolase and in particular thedefinition of the IgG2 isotype as unique for anti-αenolase antibodies inlupus nephritis are a real evolution since this antigen has been generical-ly associatedwith different auto-immune and inflammatory pathologiessuch as systemic lupus erythematosus (data on lupus nephritis not re-ported),mixed cryoglobulinemia, systemic sclerosis andANCAvasculitis[46–50]. Specificity of αenolase antibodies for selected conditions has
CTR MN AR SLE LN
anti-H3 IgG3
anti-DNA IgG2
anti-DNA IgG3
anti-H3 IgG2
anti-C1Q IgG2
anti-C1Q IgG4
anti-Annexin1 IgG2
anti- enolase IgG2
+_
Fig. 3.Hierarchical cluster analysis for levels of serum autoantibodies in various categoriesthat include patients with systemic lupus erythematosuswith (84) andwithout (104) ne-phritis, with rheumatoid arthritis (50), membranous nephritis (186) and normal controls(100). This test combines and compares simultaneously serum levels of each specific an-tibody (and includes different isotypes); the resulting heat map where colors from red(maximum) to green (minimum) indicate the relative abundances, gives an estimate ofhow different parameters separate patient on the basis of the presence of nephritis. It ap-pears that serum anti-αenolase and anti-annexin AI IgG2 separate lupus nephritis andrheumatoid arthritis better than other antibodies whereas anti-H3 IgG2/IgG3 and anti-DNA do it for lupus nephritis versus systemic lupus erythematosus.
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
never been addressed with the exception of anti-αenolase IgG4 in kid-neys of patients with MN [21,22]
A special issue was the presence of anti-C1q IgG4 in subepithelialdeposits of class V lupus nephritis since IgG4 deposition is the typicalpicture of idiopathic membranous nephropathy [2,18,20,21]. In thelatter case, intraglomerular IgG4 are directed versus a different panelof antigens, i.e. PLA2R, aldose reductase, superoxide dismutase andαenolase [18,20,21] and does not involve C1q that clearly makes a dif-ference with lupus nephritis. However, the presence of IgG4 and thecharacteristic sub-epithelial deposits constitute a pathogenic link be-tween membranous nephropathy and lupus nephritis that awaits fur-ther investigation.
3.3. Isotype preponderance and immunoglobulin shift
Preponderance of antibodies of IgG2 isotype in most lupus nephritisbiopsies was in part unexpected mainly because these antibodies areweak complement activators [51]. A report from a Chinese populationof patients with systemic lupus erythematosus indicated, however andfor the first time, an association of high serum levels of anti-C1q IgG2with both lupus flares and worsening of renal function [52]. On theother hand, immunofluorescence on the same biopsies utilized for mi-crodissection and antibody elution showed IgG2 prevalence (that con-forms the consolidated concept that lupus nephritis is an IgG2-mediated pathology) but did not exclude the presence of IgG1 andIgG3 thus confirming what was already reported by pathologists [53].Since the panel of anti-podocyte antigens here described includessome minor components (Table 1) we cannot exclude that antibodiesversus other antigens are of different isotypes. A second possibility is re-lated to the timing of renal biopsies that are performed when urinaryabnormalities are advanced; at this time, IgG1 and IgG3 that are earlyantibodies, could have already been cleared.
Experiments in animal models provide the molecular explanationfor the IgG2 shift as main mechanism for lupus related autoimmunity.In fact, immunoglobulin shift towards IgG2 inmice is determined by ac-tivation of Toll-Like receptor 9 (TLR9) signals that are known to be spe-cific for DNA and for proteins [54,55]. The same activation occurs byvirus-like particles loaded with CpGs [56]. In particular, the TLR9–MyD88 pathway is required for the generation of anti-DNA IgG2in vivo in mice with spontaneous systemic lupus erythematosus [57].Therefore, isotype switching and antibody response to TLR9–My88 aresufficient to explain preponderance of IgG2 as major auto-antibody insystemic lupus erythematosus and lupus nephritis and, consistentwith this idea, the absence of TLR9 abrogate anti-DNA formation inmice prone to systemic lupus erythematosus [58]. In systemic lupus er-ythematosus, activation of other TLRs may account for accumulation ofother IgGs, since accumulation of IgG1 was observed in mice lackingTLR9 [59] and probably compensate for the loss of IgG2.
3.4. Epitope spreading determines αenolase IgG2 reactivity
Several autoimmune conditions have been generically associatedwithcirculating anti-αenolase antibodies [46–50]; due to the methodologylimitations that characterized the pre-proteomics era, a pathogeneticrole has never been definitely demonstrated until now. Previous workson anti-αenolase in systemic lupus erythematosus (data on lupus nephri-tis not reported), mixed cryoglobulinemia, systemic sclerosis and ANCAvasculitis [46] did not define the isotype of antibodies and reported vari-able results, overall suggesting a limited specificity for various clinicalconditions.
The first observation of anti-αenolase antibody deposition in thekidney has been done in our laboratory in patients with MN in whichcase IgG4 was the unique isotype observed, well in accordance withthe knowledge thatMN is an IgG4-mediated disease [21]. Lupus nephri-tis is the second autoimmune condition associated with anti-αenolaseantibodies in glomeruli and, as discussed above, IgG2 was the unique
lupus nephritis: Isotype and antigen specificity make the difference,

6 A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
isotype amongdetected auto-antibodies.Mechanisms for IgG2 and IgG4specificity in MN and lupus nephritis require a molecular explanation.Bruschi and colleagues [29] characterized the epitopes recognized byanti-αenolase antibodies, probingαenolase CNBr digests with IgG2 pu-rified from glomeruli of patients with lupus nephritis or with IgG4 elut-ed from glomeruli of patients with membranous nephropathy [20,21].The results showed that IgG2 and IgG4 recognize different peptides de-rived from CNBr fragmentation with 1.3 kDa and 6.8 kDa respectivelythat identify different epitopes of the molecules (see the structuralmodel in Fig. 4). The 1.3 kDa epitope recognized by IgG2 is, in particular,the site for acetylation of themolecule. There is noother isotype-specificepitope for anti-αenolase antibodies reported in the literature. Howev-er, two other works recognized specificαenolase epitopes (without de-fining antibody isotype) in patients with cancer-associated retinopathysyndrome, a progressive blinding condition associated with cancer [60],and in Hashimoto encephalopathy [61] two conditions that are evident-ly different from systemic lupus erythematosus. Overall, data on epitopedifference of anti-αenolase antibodies based on disease and isotypespecificity are of interest and support a role of molecular modificationin inducing antibodieswith different characteristics andpotentially spe-cific for different diseases.
4. Alpha-enolase and annexin AI are functionally linkedwith autoimmunity
Alpha-enolase and annexin AI are ubiquitous proteins; their crucialrole for cell life being linked to different functions that have developedduring evolution. This concept implies that formation of antibodies ver-sus these two proteins may occur outside the kidney and be linked togeneralmechanisms correlated with lupus autoimmunity (for potentialmechanisms see scheme in Fig. 5). In fact, both proteins have been iden-tified in the complex extracellular trapping that composeNETosis that isconsidered a major trigger of autoimmunity in lupus [62–64]. On theother hand, their functions may be modified upon binding with anti-bodies and potentially represent themselves a mechanism leading toproteinuria and inflammation. The sub-cellular localization of both pro-teins is another key factor that may amplify their antigenic potential.
4.1. Alpha-enolase and annexin AI are components of NETosis
NETosis is a form of cell death that has the ability to modify the im-mune response. It specifically involves neutrophils which produce and
anti -enoepitope ma
MN L N
Fig. 4. Anti-αenolase IgG isotypes interact with specific epitopes of the protein. Characterizatioanti-αenolase and were then immunoblotted with IgG2 and IgG4 eluted respectively from glomfigure the different epitopes recognized by the two antibodies are indicated in a three-dimensiotibodies characterized in ‘cancer associated retinopathy’ a progressive condition leading to blin
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
extrude material that may entrap bacteria. Actually, NETosis has abasic antimicrobial function in normal cells. DNA, histones and a net-work of 23 proteins form NETosis and include αenolase and annexinAI. C1q was described as a component of NETosis as well [62–65]. Re-cent studies indicate that most of NETosis components function asautoantigens in autoimmunity and there is consensus about the possi-bility that proteins extruded in NETosis may undergo specific post-translational modification including acetylation, deimination, etc. [66].This mechanism seems crucial to lupus since abnormality of dead cellclearance has been proposed at the basis of aberrant DNA exposure tophagocytes and antibody formation in systemic lupus erythematosus[67–69]. Moreover, there is consensus about the possibility that geneticrisk factors associated with systemic lupus erythematosus alter lym-phocyte apoptosis [69,70]. Most studies on lupus autoimmunity linkedto NETosis have focused on DNA. In fact, due to the delay in removalfrom circulation, free DNAmay escape normal processing (methylationand degradation [71]) leading to activation of innate and adaptive im-munity via specific toll like receptors for viral RNA/DNA (i.e. TLR3,TLR7, TLR8 and TLR9 respectively) and release of pro-inflammatory cy-tokines/IFN [72–75]. This is a basic mechanism that in normal condi-tions leads to recognition of viral DNA and activates innate immunitythus explaining why the pathogenesis of systemic lupus erythematosusis also described as pseudoviral immunity. It is of interest to stress herethe concept that activation of TLR8/TLR9 by DNA causes immunoglobu-lin shift with production of antibodies of IgG2 isotype as already de-scribed in the previous section [57].
4.2. Functional consequences of alpha-enolase block
Alpha-enolase is a cytoplasmic enzyme that participates in gly-colysis as catalyst of the dehydration of 2-phosphoglycerate tophosphoenolpiruvate. When expressed at the surface of cells,αenolase also functions as a plasminogen receptor involved in extracel-lular fibrinolytic activity [76,77]. The binding ofαenolase with plasmin-ogen, in the presence of urokinase-type plasminogen activator,determines the conversion of plasminogen into plasmin, a proteolyticregulator of extracellular matrix. It is worth to report here that alsoannexin AII [78] and histone 2B (two antigens of the panel involved inlupus nephritis) [79] play similar functions. Externalization ofαenolaseis a commonmechanism for achieving apoptosis [80–82]; in fact, the in-teraction of αenolase and plasminogen with formation of plasmin isnecessary for proteolysis of extracellular matrix deriving from dead
lase pping
Cancer-Associated
Retinopathy syndrome
n ofαenolase epitopes target of anti-αenolase IgG2 and IgG4 was done on CNBr digests oferuli and serum of patients with lupus nephritis, with membranous nephropathy. In thisnal model. As comparison, it is also indicated the epitope recognized by anti-αenolase an-dness [60].
lupus nephritis: Isotype and antigen specificity make the difference,

1- Neutrophil netosis/apoptosis with DNA and alpha-enolase/Annexin AI exposure
2- Interception of the Ag (TLR9 for DNA) by DCs and its presentation to cells for adaptive immunity
3- Production of Abs (IgG2) against native DNA/Ags
4- Interaction of circulating antibodies with both podocyte (alpha-enolase/Annexin AI)aand implanted antigens (DNA, Histones) in glomeruli with formation of immunedeposits.
5- ROS, InflammationActivation of complement (MAC)
6- Anti-C1q antibody deposition
7- Damage of podocyteAltered actin cytoskeletonLoss of glomerular barrier permselectivity
Potential mechanisms and steps for DNA,alpha-enolase/Annexin AI antibodies formation and renal effects
Fig. 5. Proposed mechanisms for auto-antibody formation in patients with systemic lupus erythematosus. DNA deriving from breakdown of circulating cells undergoing apoptosis is rec-ognized by specialized TLR 9 and exposed to specialized cells for antibody production. Soluble proteins such as αenolase and annexin AI that participate to apoptosis or to netosis of cir-culating neutrophils may undergo a similar outcome, with formation of specific auto-antibodies in circulation. The successive steps are deposition of auto-antibodies within glomeruli andinduction of ancillarymechanisms (inflammatory, ROS) and complement activation leading to podocyte damage, altering the glomerular barrier and endingwith proteinuria, glomerularcell proliferation and sclerosis.Modification of cytoskeleton functionsmay represent an intermediate step for renal lesions. Anti-C1q deposition represents a secondary phenomenon clear-ly occurring after that C1q is deposited in glomeruli.
7A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
cells that, once digested, should be cleared. As pointed out above,αenolase is a component of NETosis supporting the concept thatexternalization of the protein from neutrophils during lupus flares is amechanism that leads to formation of circulating anti-αenolase anti-bodies in circulation that seems also logical in view of the very highlevels of serum antibodies (accounting for 5% of all IgGs in somecases) [30]. This implies that their glomerular deposition is a secondaryphenomenon.
The binding of specific antibodies toαenolase in glomeruli may alsobe critical for the development of nephritis following other mecha-nisms. One possibility is that this interaction modifies adhesion ofpodocytes to integrin and cytoskeleton [83] that makes sense in viewof the analogies betweenαenolase and the urokinase plasminogen acti-vator receptor (uPAR).Modification of the uPAR–αvβ3 integrin interac-tion in podocyte has been proposed as the cause of proteinuria inexperimental and human focal segmental glomerulosclerosis (FSGS), acondition characterized by sclerotic degeneration of the glomerulartuft [84,85]. Blocking one of the two pathways for plasmin may alterthe turn-over of extracellular matrix and makes the glomerulus moreprone to matrix accumulation typical of classes III and IV of lupusnephritis.
4.3. Functional consequences of annexin AI block
Annexin AI is a downstream mediator of anti-inflammatory effectsof glucocorticoids that are programmed to reduce local inflammatoryevents [86]. It is the first characterized member of a superfamily of 13proteins all sharing phospholipid binding properties acting as inhibitorsof phospholipase A2 and thus leading to reduction of eicosanoid synthe-sis [87]. Annexins are cell markers of apoptosis and similar to αenolaseis present in the NETosis matrix [64].
Several in vitro and in vivo observations support an anti-inflammatory role of annexin AI and its strong correlation with gluco-corticoid anti-inflammatory activity: 1—glucocorticoids increase thelevels of annexin AI in the neutrophils and mobilize it to the plasmamembrane [86]; 2—annexin AI inhibits neutrophil adhesion and trans-migration through endothelial monolayers in vitro [88,89]; the samepro-inflammatory activities of neutrophils that are stimulated to reachthe site of inflammation are blocked or reduced in cell lacking annexin
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
AI; 3—knock-out mice for this protein (Anxa1−/−) develop joint inflam-matory symptoms such as cartilage destruction in experimental arthri-tis [90] and are more prone to the toxic effect of LPS [91].
It is currently unknown how anti-annexin AI antibodies form in sys-temic lupus erythematosus and whether they modify glomerular in-flammation. One reasonable possibility mimics what was alreadyprospected for αenolase according to which high anti-annexin AI anti-bodies in circulation reflect peripheral annexin AI activation in neutro-phils with renal deposition as a direct consequence of serum levels. Inglomeruli, anti-annexin antibodiesmay reduce thephysiological cellularcounter-regulatory response to inflammation and thus represent a pro-inflammatory factor. Clarifying this aspect is crucial before concludingthat anti-annexin AI antibodies reduce the anti-inflammatory responseto steroids in patients with active systemic lupus erythematosus thatwould represent ‘per se’ a specific factor in the therapeutic approachto these patients [92].
5. Animal models support multi-antibodies and isotype specificity
Data from experimental models of lupus nephritis are generated bytwo different kinds of approaches that give different information. A firstmodel involves lupus-prone mice (MRL-lpr/lpr; NZBxNZB F1) [36,37,93]. In this case, several cell clones are derived from mice that developspontaneously lupus and are re-infused in normal mice. A second ap-proach is infusion in mice of antibodies potentially implicated in lupusnephritis.
Vlahakos and colleagues [94,95] reproduced lupus nephritis infusingthe clone H147 (an IgG2 encoded by 7183/81X VHgene) derived fromMRL-lpr/lpr showing the formation of immune deposits within the glo-merular and tubular basement membrane and the mesangium, as wellas proliferative glomerulonephritis after passive transfer to normalmice. Besides the reported findings that only antibodies of IgG2 isotypederiving from theH147 clone determine renal lesions, it is of great inter-est that the target antigen of H147 antibodies was found to beαenolase[29,93] showing important similarities between lupus nephritis inhumans and mice.
Krishnan and colleagues [37] utilized NZB prone mice and injectedIgG2 deriving from hybridomas with DNA specificity reproducingsome features of lupus nephritis. It is of interest that not all anti-DNA
lupus nephritis: Isotype and antigen specificity make the difference,

8 A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
mAbs injected reproduced lupus nephritis but only those that were ableto bind basement membrane. Their conclusion was that anti-DNA anti-bodies determine renal lesions after binding directly with glomerularantigens that remained unfortunately unknown [37].
In the second approach that considers the direct nephritogenic po-tential of specific antibodies, studies have been performed by direct in-fusion of antibodies that recognized soluble antigens [29]. BALB/c andSCID mice were injected intra peritoneum with hybridomas producinganti-enolase IgGs or anti-dsDNA IgM antibody as negative control. Inthis case, proteinuria (1–3 g) and renal histology of diffuse glomerularproliferative lesions and basement membrane thickening werereproduced in only a part of mice infused with anti-enolase IgGs show-ing a partial nephrotoxic potential of these antibodies but also suggest-ing that a multi-antibody composition is necessary to develop a fullblown picture of lupus nephritis.
It is of interest that in all experimental models that successfullyreproduced lupus nephritis, IgG2 were the unique isotype and soluble(non DNA) antigens their target. Overall, these data support the generalconcept of a multi-antibody pathogenesis of lupus nephritis in whichthe binding of specific antibodies to soluble antigens and to DNA havea role; IgG2 is the major if not unique isotype of implicated antibodiessupporting the findings deriving from human beings.
6. Clinical and pathological impact
Many unresolved issues related to renal auto-antibodies seem rele-vant to understand the pathogenesis of autoimmune conditions. Lupuserythematosus is the clinical conditionwith the highest number of anti-bodies potentially involved [96]. The coexistence of multiple antibodiesin lupus erythematosus is a key point that may explain the multi-organinvolvement [42,97–100]. Relatively to lupus nephritis, the finding of aconcomitant and parallel increase of the same antibodies in serum andin the kidney strengthened this concept. The very high levels of autoan-tibodies in serum that exceed renal levels by a factor of 103 and thefind-ing of target antigens in the NETosis matrix support the basic idea thatin lupus nephritis anti-podocyte and anti-implanted antigen antibodiesform in circulation and then deposit in the glomeruli. This possibilityconstitutes a notable difference with other autoimmune conditions,principally versus membranous nephropathy, in which it is consideredthat most antibodies form in the kidney. On the other hand, it is alsopossible that there is a hierarchy of antibody formation inwhich, follow-ing an initial hit due to deposition of a first class of antibodies (anti-DNAor anti-histone is a possibility), podocytes are perturbed and cytoplas-mic proteins are externalized this representing a secondary event(Fig. 5).
6.1. Mechanisms of autoantibody formation and deposition
Results coming from the in vivo approach to human lupus nephritisconfirm a multi-antibody panel in serum and kidneys; a few are newlydescribed antibodies (anti-αenolase, multiple anti-podocyte) whileothers were already known (i.e. anti-dsDNA/histone, anti-C1q) [59,94,95,101,102]. Confirmatory data and new results should constitute asound start to approach the pathogenesis of lupus nephritis. The basicnew point is that all renal auto-antibodies are very high in serum ofthe same patients; this finding excludes mimicry as mechanism for de-position of circulating autoantibodies in glomeruli [103–106]. Mimicrywas, in particular, popular in the past to explain interactivity betweenanti-dsDNA/histone antibodies and podocyte antigens; this seemsnow obsolete on the basis of the new findings. On the other hand, ourresults excluding an interaction of αenolase with anti-dsDNA antibod-ies [29,30] confirm the above conclusion and modify the prevailingview of anti-dsDNA as main players in lupus nephritis.
In the section dedicated to αenolase, we already discussed the pos-sibility that high levels of αenolase in serum may come from apoptoticcells in the same way as DNA. Hierarchy and time of appearance of
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
different antibodies are possible explanations for specific roles of differ-ent autoantibodies (see below) and possibly for the formulation of atwo-hit theory.
6.2. Hierarchy and time of appearance of antibodies
Membranous nephropathy is a useful model to speculate about thepossibility of a first and a second auto-immune hit in auto-immune ne-phritis. In this case, the formation of auto-antibodies against membraneproteins, such as anti-PLA2r is probably thefirst event,whereas proteinstypically localized inside the cell, such as superoxide dismutase 2 (mito-chondrial) and aldose reductase (cytoplasm) should become immuno-genic after externalization [18,20,107,108]. Hybrid proteins such asαenolase localized both on the membrane and in the cytosol may func-tion in both ways.
For systemic lupus erythematosus, the hierarchy can be guessed onthe basis of the dynamic of circulating antibody formation that con-siders antibodies versus DNA as the first step. However, we cannot ex-clude that the timing of anti-αenolase and anti-annexin AI formationis concomitant with anti-DNA, since it is now clear that all proteins par-ticipate in NETosis. Also C1q is a part of NETosis and may follow thesame mechanism.
Hierarchy in terms of antibody formation and organ specificityshould put the basis for new concepts on lupus pathogenesis since tem-poral activation of different antibody clusters could be a key forinterpreting organ-specificity of lupus subgroups. Clinical implicationsare clear. We hope that the road between bench and bedside will beshort.Membranous nephropathy is an example of how rapid clinical ap-plications of data deriving from human studies in vivo may be [26,107–109]. The attention of scientists shouldmove quickly to test predic-tion and disease prognosis and/or help clinician in therapy planning.
6.3. How this modifies the future of clinical research on lupus auto-antibodies
A final point of interest is to define whether and how the presentfindings would modify the clinical approach to systemic lupus erythe-matosus patients. Indeed, treating lupus nephritis is a possible task giv-ing that this condition is rapidly recognized and, if possible, anticipated.A number of tests have been already proposed as surrogate biomarkersof disease activity in lupus nephritis [110] and we are probably at thestep that the ‘renal auto-antibody panel’ that is also detected in serumcan be proposed as a reliable possibility.
It is clear that part of the newdata are confirmatory of previous stud-ies and should be considered on this basis. Indeed, several clinical re-ports proposed anti-dsDNA antibodies as serological markers of lupusnephritis [111] and anti-histones for identifying patients at risk of pro-liferative glomerulonephritis. However, data on specific isotypes werelacking so far [112,113]. In the most recent report on these topics, Suiand colleagues [114] showed concomitant positivity of anti-histone,anti-dsDNA and anti-nucleosome total IgG in the 50–60% of patientswith lupus nephritis compared to 35% of the general population of sys-temic lupus erythematosus. Studies in mid eighties showed high levelsof anti-histone antibodies (in particular anti H2A and 2B) that correlat-ed with lupus activity in drug-induced lupus [115] but they seem notspecific for systemic lupus erythematosus [116]. Moreover, anti-histone 2B antibodies were here excluded from the renal panel. Anti-C1q antibodies have also received wide attention: Moroni and col-leagues [117] suggested that they may help in diagnosis of lupus renalflare a finding confirmed by Mosca and colleagues [118]. Fang and col-leagues studied a large cohort of Chinese patients with lupus nephritisand found anti-C1q antibodies of IgG2 isotype prevalent among otherisotypes [52]. Isotype specificity of anti-C1q, anti-DNA and anti-histone antibodies playing a crucial role in renal deposition must beconsidered in future studies.
lupus nephritis: Isotype and antigen specificity make the difference,

9A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
Few studies considered anti-podocyte antibodies as biomarkers ofsystemic lupus erythematosus and lupus nephritis. Anti-actinin hasbeen independently analyzed in a consistent series of patients (overall91 patients with lupus nephritis and 301 without lupus nephritis) witha positivity index of 39% and 20% respectively [40,104,119,120]. Thepresence of these antibodies in serum and more important, in kidneys,has not been confirmed by proteomic studies. Anti-αenolase total IgGswere determined in a small Italian cohort of patients with systemiclupus erythematosus [118] andwere not found to be associated with ac-tive renal disease. However, technology issues (and now we can sayisotype specificity) were recognized representing a limitation factor.
We are therefore at the point to reconsider circulating auto-antibodies in lupus nephritis on the basis of results coming from prote-omics. IgG2 renal isotype and antibodies versus renal antigens such asanti-αenolase, anti-annexin AI and DNA/histone 3 represent a goodstart to design new studies. Recruitment of adequately large cohorts ofpatients that include lupus nephritis, systemic lupus erythematosusand other rheumatologic diseases could be programmed. The Zeus pro-ject is just an example (see below) but other consortiamay be built thatinvolve rheumatologists and nephrologists worldwide.
6.4. Prospective studies: the Zeus project
Prospective studies are now required. The Zeus project seeks to en-roll 1000 patients with systemic lupus erythematosus/lupus nephritis;in particular the program is to follow for 36–60 months 250 incidentsystemic lupus erythematosus patients without and 250 incident sys-temic lupus erythematosus patients with nephropathy; other 500 prev-alent systemic lupus erythematosus patients have been alreadyrecruited for antibody testing (see SLE1 ClinicalTrials.gov). Newly devel-oped antibodies and classical assays based on dsDNA IgGs will be uti-lized and compared for sensitivity and specificity. Other cohortsinclude 250 patients with rheumatoid arthritis and 600 patients affect-ed by primary glomerulonephritis, e.g. 200 with IgA, 200 with MN and200 with focal segmental glomerulonephritis. The Zeus study panel in-cludes all the authors of the present review. The final objective is to de-velop a panel of antibodies to be tested in clinical practice for a sensitiveand specific diagnosis of systemic lupus erythematosus and lupusnephritis.
7. Conclusions
Correspondence of target antigens and antibody isotype of renal andcirculating auto-antibodies has generated an immediate rebound for po-tential clinical applications. In fact, the definition of IgG2 as majorisotype and of αenolase, annexin AI, histone 3 and DNA as target of au-toimmunity has prompted the definition of an assay that was proved tobe highly specific for a diagnosis of systemic lupus erythematosus andlupus nephritis. In particular, the presence in serum of IgG2 versusαenolase and annexin AI differentiates systemic lupus erythematosusand lupus nephritis patients from patients with rheumatoid arthritisand/or with other glomerulonephritis; the presence of anti-DNA andanti-H3 IgG2 in serum is specific for systemic lupus erythematosus.The presence of anti-C1q IgG4 in class V lupus nephritis can be utilizedin differentiating primary MN from lupus lesions. Overall, antibodiesversus the same antigens but with different isotype are less specific.We are therefore moving towards a new panel of serum antibodiesthat can be assembled in an assay in which only antibodies specific forsystemic lupus erythematosus and lupus nephritis are determined, thisrepresenting an easy and more powerful tool for a direct diagnosis ofsystemic lupus erythematosus even in patientswith glomerulonephritis.
Take-home messages
• Autoantibodies potentially implicated in pathogenesis of lupus ne-phritis in human beings have been directly eluted from micro-
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
dissected glomeruli obtained from renal biopsies and characterizedby proteomics.
• Anti-vimentin antibodies are implicated in tubulointerstitial lesionsand correlate with progression of renal lesions.
• Immunoglobulins directed towards 11 podocyte proteins and 5 im-planted antigens were detected and identified; antibodies of IgG2isotype versus αenolase, annexin AI, histone 3 and DNA were themajor components; anti-C1q Ig4 were also detected in sub-epithelialdeposits of class V lupus nephritis.
• A high homology between renal and serum autoantibodies was dem-onstrated; antibodies of IgG2 isotype versus the whole panel of renalantigens were also detected in serum of systemic lupus erythemato-sus and lupus nephritis patients. In some cases, serum levels increasedby 10 times and reached levels in the range of hundred milligrams/l(e.g. 500 mg/l for anti-αenolase IgG2).
• Concomitant positivity of all IgG2 antibodies allows to differentiatesystemic lupus erythematosus patients from lupus nephritis andfrom other rheumatologic conditions and lupus nephritis from otherprimary glomerulonephritis.
• TLR9 activation by free DNA following NETosis may explain IgG2 pre-ponderance in lupus. Epitope spreading explains IgG2 reactivity insome cases.
Author contribution
AB, study design, patient recruitment, manuscript discussion, coordi-nation of the study; MB, study design, proteomics, data managing, man-uscript discussion; RAS, study design, patient recruitment, manuscriptdiscussion; GM, study design, patient recruitment, manuscript discus-sion; GP, study design, patient recruitment, manuscript discussion; LA,study design, patient recruitment, manuscript discussion; FP, study de-sign, patient recruitment, manuscript discussion; PM, study design, pa-tient recruitment, manuscript discussion; FF, study design, patientrecruitment,manuscript discussion; GC, study design, biochemistry, pro-teomics, manuscript discussion; AM, study design, patient recruitment,manuscript discussion; LC, study design, patient recruitment,manuscriptdiscussion; CM, study design, patient recruitment, manuscript discus-sion; AV, study supervision, patient recruitment, manuscript discussion;AT, study supervision, patient recruitment, manuscript discussion; GMG,study design and coordination, data managing and supervision, patientsrecruitment, manuscript writing and discussion.
Conflict of interest
None.
Acknowledgments
Istituto Giannina Gaslini provided logistic and financial support. Thispaper was also supported by the Italian Ministry of Health “RicercaCorrente, contributo per la ricerca intramuraria 2013.”, Renal ChildFoundation, and Fondazione La Nuova Speranza (“Progetto integratoper la definizione dei meccanismi implicati nella glomerulo sclerosifocale”). A patent on the potential use of anti-alpha enolase/anti-H3/anti-DNA IgG2 antibodies as surrogate biomarker of lupus nephritis ispending.
References
[1] Segelmark M, Hellmark T. Autoimmune kidney diseases. Autoimmun Rev 2010;9:A366–71.
[2] Kerjaschki D. Pathomechanisms and molecular basis of membranous glomerulop-athy. Lancet 2004;364:1194–6.
[3] Couser WG, Steinmuller DR, Stilmant MM, Salant DJ, Lowenstein LM. Experimentalglomerulonephritis in the isolated perfused rat kidney. J Clin Invest 1978;62:1275–87.
[4] Salant DJ, Belok S, Madaio MP, Couser WG. A new role for complement in experi-mental membranous nephropathy in rats. J Clin Invest 1980;66:1339–50.
lupus nephritis: Isotype and antigen specificity make the difference,

10 A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
[5] Van Damme BJ, Fleuren GJ, BakkerWW, Vernier RL, Hoedemaeker PJ. Experimentalglomerulonephritis in the rat induced by antibodies directed against tubular anti-gens. V. Fixed glomerular antigens in the pathogenesis of heterologous immunecomplex glomerulonephritis. Lab Invest 1978;38:502–10.
[6] Mihai S, Nimmerjahn F. The role of Fc receptors and complement in autoimmunity.Autoimmun Rev 2013;12:657–60.
[7] Cybulsky AV, Takano T, Papillon J, McTavish AJ. Complement-induced phospholi-pase A2 activation in experimental membranous nephropathy. Kidney Int 2000;57:1052–62.
[8] Ghiggeri GM, Bruschi M, Candiano G, Rastaldi MP, Scolari F, Passerini P, et al. Deple-tion of clusterin in renal diseases causing nephrotic syndrome. Kidney Int 2002;62:2184–94.
[9] Rastaldi MP, Armelloni S, Berra S, Calvaresi N, Corbelli A, Giardino LA, et al. Glomer-ular podocytes contain neuron-like functional synaptic vesicles. FASEB J 2006;20:976–8.
[10] Greco A, Rizzo MI, De Virgilio A, Gallo A, Fusconi M, Pagliuca G, et al. Goodpasture'ssyndrome: a clinical update. Autoimmun Rev 2015;14:246–53.
[11] Kerjaschki D, Farquhar MG. Immunocytochemical localization of the Heymann ne-phritis antigen (GP330) in glomerular epithelial cells of normal Lewis rats. J ExpMed 1983;157:667–86.
[12] Kerjaschki D, Horvat R, Binder S, Susani M, Dekan G, Ojha PP, et al. Identification ofa 400-kd protein in the brush borders of human kidney tubules that is similar togp330, the nephritogenic antigen of rat Heymann nephritis. Am J Pathol 1987;129:183–91.
[13] Allegri L. Antigens in experimental models of membranous nephropathy: are theyinvolved in human disease? Nephrol Dial Transplant 1997;12:1801–4.
[14] Saito A, Pietromonaco S, Loo AK, Farquhar MG. Complete cloning and sequencing ofrat gp330/“megalin,” a distinctive member of the low density lipoprotein receptorgene family. Proc Natl Acad Sci U S A 1994;91:9725–9.
[15] Farquhar MG, Saito A, Kerjaschki D, Orlando RA. The Heymann nephritis antigeniccomplex: megalin (gp330) and RAP. J Am Soc Nephrol 1995;6:35–47.
[16] Adler S, Baker PJ, Pritzl P, Couser WG. Detection of terminal complement compo-nents in experimental immune glomerular injury. Kidney Int 1984;26:830–7.
[17] Adler S, Huang H. Oxidant stress in kidneys of spontaneously hypertensive rats in-volves both oxidase overexpression and loss of extracellular superoxide dismutase.Am J Physiol Renal Physiol 2004;287:F907–13.
[18] Beck Jr LH, Bonegio RG, Lambeau G, Beck DM, Powell DW, Cummins TD, et al. M-type phospholipase A2 receptor as target antigen in idiopathic membranous ne-phropathy. N Engl J Med 2009;361:11–21.
[19] Tomas NM, Beck Jr LH, Meyer-Schwesinger C, Seitz-Polski B, Ma H, Zahner G, et al.Thrombospondin type-1 domain-containing 7A in idiopathic membranous ne-phropathy. N Engl J Med 2014;371:2277–87.
[20] Prunotto M, Carnevali ML, Candiano G, Murtas C, Bruschi M, Corradini E, et al. Au-toimmunity in membranous nephropathy targets aldose reductase and SOD2. J AmSoc Nephrol 2010;21:507–19.
[21] Bruschi M, Carnevali ML, Murtas C, Candiano G, Petretto A, Prunotto M, et al. Directcharacterization of target podocyte antigens and auto-antibodies in human mem-branous glomerulonephritis: alfa-enolase and borderline antigens. J Proteomics2011;74:2008–17.
[22] Murtas C, Bruschi M, Carnevali ML, Petretto A, Corradini E, Prunotto M, et al. In vivocharacterization of renal auto-antigens involved in human auto-immune diseases:the case of membranous glomerulonephritis. Proteomics Clin Appl 2011;5:90–7.
[23] Makker SP, Tramontano A. Differential capacity of anti-RAP and anti-megalin anti-bodies to produce progressive passive Heymann nephritis—implications for thepathogenesis of idiopathic human membranous glomerulonephritis. J Pathol2006;210:282–7.
[24] Makker SP, Tramontano A. Idiopathic membranous nephropathy: an autoimmunedisease. Semin Nephrol 2011;31:333–40.
[25] Debiec H, Guigonis V, Mougenot B, Decobert F, Haymann JP, Bensman A, et al. An-tenatal membranous glomerulonephritis due to anti-neutral endopeptidase anti-bodies. N Engl J Med 2002;346:2053–60.
[26] Murtas C, Bruschi M, Candiano G, Moroni G, Magistroni R, Magnano A, et al. Coex-istence of different circulating anti-podocyte antibodies in membranous nephrop-athy. Clin J Am Soc Nephrol 2012;7:1394–400.
[27] Schlumberger W, Hornig N, Lange S, Probst C, Komorowski L, Fechner K, et al. Dif-ferential diagnosis of membranous nephropathy with autoantibodies to phospho-lipase A2 receptor 1. Autoimmun Rev 2014;13:108–13.
[28] Kinloch AJ, Chang A, Ko K, Henry Dunand CJ, Henderson S, Maienschein-Cline M,et al. Vimentin is a dominant target of in situ humoral immunity in human lupustubulointerstitial nephritis. Arthritis Rheum 2014;66:3359–70.
[29] Bruschi M, Sinico RA, Moroni G, Pratesi F, Migliorini P, Galetti M, et al. Glomerularautoimmune multi-components of human lupus nephritis in vivo (1): alpha-enolase and annexin AI. J Am Soc Nephrol 2014;25:2483–98.
[30] Bruschi M, Galetti M, Sinico RA, Moroni G, Bonanni A, Radice A, et al. Glomerularautoimmune multicomponents of human lupus nephritis in vivo (2): planted anti-gens. J Am Soc Nephrol 2014;26.
[31] Borchers AT, Leibushor N, Naguwa SM, Cheema GS, Shoenfeld Y, Gershwin ME.Lupus nephritis: a critical review. Autoimmun Rev 2012;12:174–94.
[32] Doria A, Zen M, Bettio S, Gatto M, Bassi N, Nalotto L, et al. Autoinflammation andautoimmunity: bridging the divide. Autoimmun Rev 2012;12:22–30.
[33] Singh RP, Waldron RT, Hahn BH. Genes, tolerance and systemic autoimmunity.Autoimmun Rev 2012;11:664–9.
[34] Borchers AT, Naguwa SM, Shoenfeld Y, Gershwin ME. The geoepidemiology of sys-temic lupus erythematosus. Autoimmun Rev 2010;9:A277–87.
[35] Madaio MP. The relevance of antigen binding to the pathogenicity of lupus autoan-tibodies. Kidney Int 1982;82:125–7.
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
[36] Vargas MT, Gustilo K, D'Andrea DM, Kalluri R, Foster MH, Madaio MP. Structuralfeatures of nephritogenic lupus autoantibodies. Methods 1997;11:62–9.
[37] Krishnan MR, Wang C, Marion TN. Anti-DNA autoantibodies initiate experimentallupus nephritis by binding directly to the glomerular basement membrane inmice. Kidney Int 2012;82:184–92.
[38] Kalaaji M, Fenton KA, Mortensen ES, Olsen R, Sturfelt G, Alm P, et al. Glomerular ap-optotic nucleosomes are central target structures for nephritogenic antibodies inhuman SLE nephritis. Kidney Int 2007;71:664–72.
[39] Mjelle JE, Rekvig OP, Van Der Vlag J, Fenton KA. Nephritogenic antibodies bind inglomeruli through interaction with exposed chromatin fragments and not withrenal cross-reactive antigens. Autoimmunity 2011;44:373–83.
[40] Renaudineau Y, Croquefer S, Jousse S, Renaudineau E, Devauchelle V, Gueguen P,et al. Association of alpha-actinin-binding anti-double-stranded DNA antibodieswith lupus nephritis. Arthritis Rheum 2006;54:2523–32.
[41] Deocharan B, Zhou Z, Antar K, Siconolfi-Baez L, Angeletti RH, Hardin J, et al. Alpha-actinin immunization elicits anti-chromatin autoimmunity in nonautoimmunemice. J Immunol 2007;179:1313–21.
[42] Mannik M, Merrill CE, Stamps LD, Wener MH. Multiple autoantibodies form theglomerular immune deposits in patients with systemic lupus erythematosus. JRheumatol 2003;30:1495–504.
[43] Ortega LM, Schultz DR, Lenz O, Pardo V, Contreras GN. Review: lupus nephritis:pathologic features, epidemiology and a guide to therapeutic decisions. Lupus2010;19:557–74.
[44] Davidson A. Editorial: autoimmunity to vimentin and lupus nephritis. ArthritisRheum 2014;66:3251–4.
[45] Kretz CC, NorpoM, Abeler-Dorner L, Linke B, Haust M, Edler L, et al. Anti-annexin 1antibodies: a new diagnostic marker in the serum of patients with discoid lupus er-ythematosus. Exp Dermatol 2010;19:919–21.
[46] Moodie FD, Leaker B, Cambridge G, Totty NF, Segal AW. Alpha-enolase: a novel cy-tosolic autoantigen in ANCA positive vasculitis. Kidney Int 1993;43:675–81.
[47] Migliorini P, Pratesi F, Bongiorni F, Moscato S, Scavuzzo M, Bombardieri S. The tar-gets of nephritogenic antibodies in systemic autoimmune disorders. AutoimmunRev 2002;1:168–73.
[48] Sabbatini A, Dolcher MP, Marchini B, Chimenti D, Moscato S, Pratesi F, et al. Alpha-enolase is a renal-specific antigen associated with kidney involvement in mixedcryoglobulinemia. Clin Exp Rheumatol 1997;15:655–8.
[49] Wakui H, Imai H, Komatsuda A, Miura AB. Circulating antibodies against alpha-enolase in patients with primary membranous nephropathy (MN). Clin ExpImmunol 1999;118:445–50.
[50] Terrier B, Degand N, Guilpain P, Servettaz A, Guillevin L, Mouthon L. Alpha-enolase:a target of antibodies in infectious and autoimmune diseases. Autoimmun Rev2007;6:176–82.
[51] Bindon CI, Hale G, Bruggemann M, Waldmann H. Human monoclonal IgG isotypesdiffer in complement activating function at the level of C4 aswell as C1q. J ExpMed1988;168:127–42.
[52] Fang QY, Yu F, Tan Y, Xu LX, Wu LH, Liu G, et al. Anti-C1q antibodies and IgG sub-class distribution in sera from Chinese patients with lupus nephritis. Nephrol DialTransplant 2009;24:172–8.
[53] Song YS, Min KW, Kim JH, Kim GH, ParkMH. Differential diagnosis of lupus and pri-mary membranous nephropathies by IgG subclass analysis. Clin J Am Soc Nephrol2012;7:1947–55.
[54] Christensen SR, Shupe J, Nickerson K, Kashgarian M, Flavell RA, Shlomchik MJ. Toll-like receptor 7 and TLR9 dictate autoantibody specificity and have opposing inflam-matory and regulatory roles in amurinemodel of lupus. Immunity 2006;25:417–28.
[55] Tian J, Avalos AM, Mao SY, Chen B, Senthil K, Wu H, et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated byHMGB1 and RAGE. Nat Immunol 2007;8:487–96.
[56] Jegerlehner A, Maurer P, Bessa J, Hinton HJ, Kopf M, Bachmann MF. TLR9 signalingin B cells determines class switch recombination to IgG2a. J Immunol 2007;178:2415–20.
[57] Ehlers M, Fukuyama H, McGaha TL, AderemA, Ravetch JV. TLR9/MyD88 signaling isrequired for class switching to pathogenic IgG2a and 2b autoantibodies in SLE. JExp Med 2006;203:553–61.
[58] Christensen SR, Kashgarian M, Alexopoulou L, Flavell RA, Akira S, Shlomchik MJ.Toll-like receptor 9 controls anti-DNA autoantibody production in murine lupus. JExp Med 2005;202:321–31.
[59] Lau CM, Broughton C, Tabor AS, Akira S, Flavell RA, Mamula MJ, et al. RNA-associated autoantigens activate B cells by combined B cell antigen receptor/Toll-like receptor 7 engagement. J Exp Med 2005;202:1171–7.
[60] Adamus G, Amundson D, Seigel GM, Machnicki M. Anti-enolase-alpha autoanti-bodies in cancer-associated retinopathy: epitope mapping and cytotoxicity on ret-inal cells. J Autoimmun 1998;11:671–7.
[61] Fujii A, Yoneda M, Ito T, Yamamura O, Satomi S, Higa H, et al. Autoantibodiesagainst the amino terminal of alpha-enolase are a useful diagnostic marker ofHashimoto's encephalopathy. J Neuroimmunol 2005;162:130–6.
[62] Saffarzadeh M, Juenemann C, Queisser MA, Lochnit G, Barreto G, Galuska SP, et al.Neutrophil extracellular traps directly induce epithelial and endothelial celldeath: a predominant role of histones. PLoS One 2012;7:e32366.
[63] Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, et al.Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex in-volved in host defense against Candida albicans. PLoS Pathog 2009;5:e1000639.
[64] Darrah E, Andrade F. NETs: the missing link between cell death and systemic auto-immune diseases? Front Immunol 2013;3:428.
[65] Lande R, Ganguly D, Facchinetti V, Frasca L, Conrad C, Gregorio J, et al. Neutrophilsactivate plasmacytoid dendritic cells by releasing self–DNA–peptide complexes insystemic lupus erythematosus. Sci Transl Med 2011;3:73ra19.
lupus nephritis: Isotype and antigen specificity make the difference,

11A. Bonanni et al. / Autoimmunity Reviews xxx (2015) xxx–xxx
[66] Knight JS, Carmona-Rivera C, Kaplan MJ. Proteins derived from neutrophil extracel-lular traps may serve as self-antigens and mediate organ damage in autoimmunediseases. Front Immunol 2012;3:380.
[67] Herrmann M, Voll RE, Zoller OM, Hagenhofer M, Ponner BB, Kalden JR. Impairedphagocytosis of apoptotic cell material by monocyte-derived macrophages frompatients with systemic lupus erythematosus. Arthritis Rheum 1998;41:1241–50.
[68] Cohen PL, Caricchio R, Abraham V, Camenisch TD, Jennette JC, Roubey RA, et al. De-layed apoptotic cell clearance and lupus-like autoimmunity in mice lacking the c-mer membrane tyrosine kinase. J Exp Med 2002;196:135–40.
[69] Hakkim A, Furnrohr BG, Amann K, Laube B, Abed UA, Brinkmann V, et al. Impair-ment of neutrophil extracellular trap degradation is associated with lupus nephri-tis. Proc Natl Acad Sci U S A 2010;107:9813–8.
[70] Anders HJ. Pseudoviral immunity—a novel concept for lupus. Trends Mol Med2009;15:553–61.
[71] Huck S, Deveaud E, Namane A, Zouali M. Abnormal DNA methylation anddeoxycytosine–deoxyguanine content in nucleosomes from lymphocytes under-going apoptosis. FASEB J 1999;13:1415–22.
[72] Savarese E, Chae OW, Trowitzsch S, Weber G, Kastner B, Akira S, et al. U1 small nu-clear ribonucleoprotein immune complexes induce type I interferon inplasmacytoid dendritic cells through TLR7. Blood 2006;107:3229–34.
[73] Pawar RD, Ramanjaneyulu A, Kulkarni OP, Lech M, Segerer S, Anders HJ. Inhibitionof toll-like receptor-7 (TLR-7) or TLR-7 plus TLR-9 attenuates glomerulonephritisand lung injury in experimental lupus. J Am Soc Nephrol 2007;18:1721–31.
[74] Theofilopoulos AN, Baccala R, Beutler B, Kono DH. Type I interferons (alpha/beta) inimmunity and autoimmunity. Annu Rev Immunol 2005;23:307–36.
[75] Garcia-Romo GS, Caielli S, Vega B, Connolly J, Allantaz F, Xu Z, et al. Netting neutro-phils are major inducers of type I IFN production in pediatric systemic lupus ery-thematosus. Sci Transl Med 2011;3:73ra20.
[76] Ossowski L, Aguirre-Ghiso JA. Urokinase receptor and integrin partnership: coordi-nation of signaling for cell adhesion, migration and growth. Curr Opin Cell Biol2000;12:613–20.
[77] Lopez-Alemany R, Correc P, Camoin L, Burtin P. Purification of the plasmin receptorfrom human carcinoma cells and comparison to alpha-enolase. Thromb Res 1994;75:371–81.
[78] Hajjar KA, Jacovina AT, Chacko J. An endothelial cell receptor for plasminogen/tissue plasminogen activator I. Identity with annexin II. J Biol Chem 1994;269:21191–7.
[79] Herren T, Burke TA, Das R, Plow EF. Identification of histone H2B as a regulatedplasminogen receptor. Biochemistry 2006;45:9463–74.
[80] Meilhac O, Ho-Tin-Noe B, Houard X, Philippe M, Michel JB, Angles-Cano E.Pericellular plasmin induces smooth muscle cell anoikis. FASEB J 2003;17:1301–3.
[81] Kochtebane N, Choqueux C, Passefort S, Nataf P, Messika-Zeitoun D, Bartagi A, et al.Plasmin induces apoptosis of aortic valvular myofibroblasts. J Pathol 2010;221:37–48.
[82] Ucker DS, Jain MR, Pattabiraman G, Palasiewicz K, Birge RB, Li H. Externalized gly-colytic enzymes are novel, conserved, and early biomarkers of apoptosis. J BiolChem 2012;287:10325–43.
[83] Blasi F, Carmeliet P. uPAR: a versatile signalling orchestrator. Nat Rev Mol Cell Biol2002;3:932–43.
[84] Wei C, Moller CC, Altintas MM, Li J, Schwarz K, Zacchigna S, et al. Modification ofkidney barrier function by the urokinase receptor. Nat Med 2008;14:55–63.
[85] Wei C, El Hindi S, Li J, Fornoni A, Goes N, Sageshima J, et al. Circulating urokinasereceptor as a cause of focal segmental glomerulosclerosis. Nat Med 2011;17:952–60.
[86] Perretti M, D'Acquisto F. Annexin A1 and glucocorticoids as effectors of the resolu-tion of inflammation. Nat Rev Immunol 2009;9:62–70.
[87] Blackwell GJ, Carnuccio R, Di Rosa M, Flower RJ, Parente L, Persico P. Macrocortin: apolypeptide causing the anti-phospholipase effect of glucocorticoids. Nature 1980;287:147–9.
[88] Walther A, Riehemann K, Gerke V. A novel ligand of the formyl peptide receptor:annexin I regulates neutrophil extravasation by interacting with the FPR. Mol Cell2000;5:831–40.
[89] Zouki C, Ouellet S, Filep JG. The anti-inflammatory peptides, antiflammins, regulatethe expression of adhesionmolecules on human leukocytes and prevent neutrophiladhesion to endothelial cells. FASEB J 2000;14:572–80.
[90] Frey O, Petrow PK, GajdaM, Siegmund K, Huehn J, Scheffold A, et al. The role of reg-ulatory T cells in antigen-induced arthritis: aggravation of arthritis after depletionand amelioration after transfer of CD4+CD25+ T cells. Arthritis Res Ther 2005;7:R291–301.
[91] Damazo AS, Yona S, D'Acquisto F, Flower RJ, Oliani SM, Perretti M. Critical protec-tive role for annexin 1 gene expression in the endotoxemic murine microcircula-tion. Am J Pathol 2005;166:1607–17.
[92] Bazso A, Szappanos A, Patocs A, Poor G, Shoenfeld Y, Kiss E. The importance of glu-cocorticoid receptors in systemic lupus erythaematosus. A systematic review.Autoimmun Rev 2015;14:349–51.
[93] D'Andrea DM, Coupaye-Gerard B, Kleyman TR, Foster MH, Madaio MP. Lupus auto-antibodies interact directly with distinct glomerular and vascular cell surface anti-gens. Kidney Int 1996;49:1214–21.
[94] Vlahakos D, Foster MH, Ucci AA, Barrett KJ, Datta SK, Madaio MP. Murine monoclo-nal anti-DNA antibodies penetrate cells, bind to nuclei, and induce glomerular pro-liferation and proteinuria in vivo. J Am Soc Nephrol 1992;2:1345–54.
Please cite this article as: Bonanni A, et al, Multi-antibody composition inAutoimmun Rev (2015), http://dx.doi.org/10.1016/j.autrev.2015.04.004
[95] Vlahakos DV, Foster MH, Adams S, Katz M, Ucci AA, Barrett KJ, et al. Anti-DNA an-tibodies form immune deposits at distinct glomerular and vascular sites. Kidney Int1992;41:1690–700.
[96] Yaniv G, Twig G, Shor DB, Furer A, Sherer Y, Mozes O, et al. A volcanic explosion ofautoantibodies in systemic lupus erythematosus: a diversity of 180 different anti-bodies found in SLE patients. Autoimmun Rev 2015;14:75–9.
[97] Pankewycz OG, Migliorini P, Madaio MP. Polyreactive autoantibodies arenephritogenic in murine lupus nephritis. J Immunol 1987;139:3287–94.
[98] Sabbaga J, Pankewycz OG, Lufft V, Schwartz RS, Madaio MP. Cross-reactivity distin-guishes serum and nephritogenic anti-DNA antibodies in human lupus from theirnatural counterparts in normal serum. J Autoimmun 1990;3:215–35.
[99] Zhang J, Jacobi AM, Wang T, Berlin R, Volpe BT, Diamond B. Polyreactive autoanti-bodies in systemic lupus erythematosus have pathogenic potential. J Autoimmun2009;33:270–4.
[100] Li QZ, Xie C, Wu T, Mackay M, Aranow C, Putterman C, et al. Identification of auto-antibody clusters that best predict lupus disease activity using glomerular prote-ome arrays. J Clin Invest 2005;115:3428–39.
[101] Waters ST, McDuffie M, Bagavant H, Deshmukh US, Gaskin F, Jiang C, et al. Breakingtolerance to double stranded DNA, nucleosome, and other nuclear antigens is notrequired for the pathogenesis of lupus glomerulonephritis. J Exp Med 2004;199:255–64.
[102] Madaio MP, Carlson J, Cataldo J, Ucci A, Migliorini P, Pankewycz O. Murine mono-clonal anti-DNA antibodies bind directly to glomerular antigens and form immunedeposits. J Immunol 1987;138:2883–9.
[103] Kalaaji M, Mortensen E, Jorgensen L, Olsen R, Rekvig OP. Nephritogenic lupus anti-bodies recognize glomerular basement membrane-associated chromatin frag-ments released from apoptotic intraglomerular cells. Am J Pathol 2006;168:1779–92.
[104] Kalaaji M, Sturfelt G, Mjelle JE, Nossent H, Rekvig OP. Critical comparative analysesof anti-alpha-actinin and glomerulus-bound antibodies in human and murinelupus nephritis. Arthritis Rheum 2006;54:914–26.
[105] S. Yung, K.F. Cheung, Q. Zhang and T.M. Chan, Anti-dsDNA antibodies bind tomesangial annexin II in lupus nephritis, J Am Soc Nephrol 21 1912–27.
[106] Foster MH, Sabbaga J, Line SR, Thompson KS, Barrett KJ, Madaio MP. Molecularanalysis of spontaneous nephrotropic anti-laminin antibodies in an autoimmuneMRL-lpr/lpr mouse. J Immunol 1993;151:814–24.
[107] Hofstra JM, Debiec H, Short CD, Pelle T, Kleta R, Mathieson PW, et al.Antiphospholipase A2 receptor antibody titer and subclass in idiopathic membra-nous nephropathy. J Am Soc Nephrol 2012;23:1735–43.
[108] Hoxha E, Kneissler U, Stege G, Zahner G, Thiele I, Panzer U, et al. Enhanced expres-sion of the M-type phospholipase A2 receptor in glomeruli correlates with serumreceptor antibodies in primary membranous nephropathy. Kidney Int 2012;82:797–804.
[109] L.H. Beck, Jr., F.C. Fervenza, D.M. Beck, R.G. Bonegio, F.A. Malik, S.B. Erickson, F.G.Cosio, D.C. Cattran and D.J. Salant, Rituximab-induced depletion of anti-PLA2R au-toantibodies predicts response in membranous nephropathy, J Am Soc Nephrol 221543–50.
[110] Moroni G, Radice A, Giammarresi G, Quaglini S, Gallelli B, Leoni A, et al. Are labora-tory tests useful for monitoring the activity of lupus nephritis? A 6-year prospec-tive study in a cohort of 228 patients with lupus nephritis. Ann Rheum Dis 2009;68:234–7.
[111] Kiss E, Lakos G, Szegedi G, Poor G, Szodoray P. Anti-nuscleosome antibody, a reli-able indicator for lupus nephritis. Autoimmunity 2009;42:393–8.
[112] Gomez-Puerta JA, Burlingame RW, Cervera R. Anti-chromatin (anti-nucleosome)antibodies: diagnostic and clinical value. Autoimmun Rev 2008;7:606–11.
[113] HungWT, Chen YM, Lan JL, Chen HH, Chen YH, Chen DY, et al. Antinucleosome an-tibodies as a potential biomarker for the evaluation of renal pathological activity inpatients with proliferative lupus nephritis. Lupus 2011;20:1404–10.
[114] Sui M, Lin Q, Xu Z, Han X, Xie R, Jia X, et al. Simultaneous positivity for anti-DNA,anti-nucleosome and anti-histone antibodies is a marker for more severe lupus ne-phritis. J Clin Immunol 2013;33:378–87.
[115] Rubin RL, Bell SA, Burlingame RW. Autoantibodies associated with lupus inducedby diverse drugs target a similar epitope in the (H2A–H2B)–DNA complex. J ClinInvest 1992;90:165–73.
[116] Costa O, Monier JC. Antihistone antibodies detected by ELISA and immunoblottingin systemic lupus erythematosus and rheumatoid arthritis. J Rheumatol 1986;13:722–5.
[117] Moroni G, Trendelenburg M, Del Papa N, Quaglini S, Raschi E, Panzeri P, et al. Anti-C1q antibodies may help in diagnosing a renal flare in lupus nephritis. Am J KidneyDis 2001;37:490–8.
[118] Mosca M, Chimenti D, Pratesi F, Baldini C, Anzilotti C, Bombardieri S, et al. Preva-lence and clinico-serological correlations of anti-alpha-enolase, anti-C1q, andanti-dsDNA antibodies in patients with systemic lupus erythematosus. J Rheumatol2006;33:695–7.
[119] Croquefer S, Renaudineau Y, Jousse S, Gueguen P, Ansart S, Saraux A, et al. Theananti-alpha-actinin test completes ananti-DNA determination in systemic lupuserythematosus. Ann N Y Acad Sci 2005;1050:170–5.
[120] Becker-Merok A, Kalaaji M, Haugbro K, Nikolaisen C, Nilsen K, Rekvig OP, et al.Alpha-actinin-binding antibodies in relation to systemic lupus erythematosusand lupus nephritis. Arthritis Res Ther 2006;8:R162.
lupus nephritis: Isotype and antigen specificity make the difference,
Copyright © 2022 FDOKUMEN




















![[Proliferative lupus nephritis treatment: practice survey in nephrology and internal medicine in France]](https://static.fdokumen.com/doc/165x107/6336db7d20d9c9602f0b0be8/proliferative-lupus-nephritis-treatment-practice-survey-in-nephrology-and-internal.jpg)