
beta-Arrestin1 Mediates the Endocytosis and Functions of Macrophage Migration Inhibitory Factor

Calcium-Sensing Receptor Endocytosis LinksExtracellular Calcium Signaling to ParathyroidHormone-Related Peptide Secretion via a Rab11a-Dependent and AMSH-Sensitive Mechanism
Alma P. Reyes-Ibarra, Alejandro Garcıa-Regalado, Iliana Ramırez-Rangel, Ana L. Esparza-Silva,Margarita Valadez-Sanchez, Jose Vazquez-Prado, and Guadalupe Reyes-Cruz
Departments of Cell Biology (A.P.R.-I., A.G.-R., M.V.-S., G.R.-C.) and Pharmacology (I.R.-R., A.L.E.-S.,J.V.-P.), Centro de Investigacion y de Estudios Avanzados-Instituto Politecnico Nacional, 07000Mexico D.F., Mexico
The calcium-sensing receptor (CaR) helps to main-tain the homeostasis of extracellular calcium bycontrolling the secretion of hormones associatedwith this process. The mechanism of agonist-in-duced endocytosis and down-regulation of CaRand the influence of this event on the secretion ofCaR-regulated hormones is not fully understood.In this study, we show that CaR is constitutivelyendocytosed and recycled to the plasma mem-brane by a Rab11a-dependent mechanism; duringthis process, the level of total cellular CaR is main-tained. This trafficking of CaR promotes the secre-tion of PTH-related peptide (PTHrP), as evidencedby a decrease on PTHrP secretion in the presence
of a dominant-negative mutant of Rab11a. Interest-ingly, this Rab11a dominant-negative mutant doesnot interfere with CaR-dependent activation ofERK 1/2, suggesting that ERK signaling is not suf-ficient to promote PTHrP secretion downstream ofCaR. In addition, AMSH (associated molecule withthe SH3 domain of STAM), a CaR carboxyl-terminalbinding protein, redirects CaR from slow recyclingto down-regulation, reducing CaR expression anddecreasing PTHrP secretion. Our results indicatethat endocytosis and trafficking of CaR modulatePTHrP secretion. (Molecular Endocrinology 21:1394–1407, 2007)
THE CALCIUM-SENSING RECEPTOR (CaR) is acentral regulator of extracellular calcium (Ca2�
o)homeostasis by controlling the secretory properties ofthe parathyroid gland. CaR-modulated PTH secretionleads to a decrease in calcium excretion in kidney andits removal from bone by activated osteoclasts (1, 2).CaR gene mutations are frequently associated withinherited disorders involving calcium homeostasis (3,4). CaR signaling is mediated by heterotrimeric G pro-teins, including Gq, Gi, and G12/13, promoting intracel-lular calcium mobilization, adenylyl cyclase inhibition,and Rho GTPase activation; CaR also transactivatesepidermal growth factor receptor (EGFR) signaling andits downstream ERK cascade (3, 5–9). In addition, CaRmediates G protein-independent signals through inter-actions of its carboxyl-terminal tail with intracellularproteins, including filamin A and AMSH (associatedmolecule with the SH3 domain of STAM) to modulate
the receptor desensitization and internalization rate(10–13).
In addition to the parathyroid gland, CaR is ex-pressed in diverse cellular settings in which it regu-lates PTHrP secretion (14). The mechanism by whichCaR regulates the trafficking and secretion of PTH- orPTHrP-containing vesicles is not known but might in-volve the endocytosis and trafficking of CaR itself.Endocytosis and trafficking of G protein-coupled re-ceptors (GPCRs) are considered late events of theirdesensitization (15, 16); however, GPCR internaliza-tion may promote MAPK cascade activation and re-lease of secretory vesicles (17). In the latter case, aninterplay between endocytic vesicles and secretoryvesicles has been described. Trafficking of intracellu-lar vesicles is coordinated by a variety of Rab GT-Pases. Rab11, one of the 40 known Rab GTPases,controls the transport of membrane receptors, includ-ing some GPCRs from early endosomes to perinuclearslow-recycling endosomes (18–20). The potential ef-fect of CaR internalization, trafficking, and down-regulation on hormone secretion that is known tobe regulated by this receptor has not been fullyelucidated.
To investigate the role of CaR endocytosis in itsability to regulate PTHrP secretion, we studied theeffect of mutant Rab GTPases and sorting proteins onCaR trafficking, stability, and ability to control PTHrP
First Published Online April 10, 2007Abbreviations: ADD, Anti-CaR ADD antibody; Ca2�
o, ex-tracellular calcium; CaR, calcium-sensing receptor; EGFR,epidermal growth factor receptor; GASP, G protein-coupledreceptor-associated sorting protein; GFP, green fluorescentprotein; GPCR, G protein-coupled receptor; HEK-293, hu-man embryonic kidney 293; shRNA, short hairpin RNA.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/07/$15.00/0 Molecular Endocrinology 21(6):1394–1407Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/me.2006-0523
1394
secretion. Our results indicate that CaR is constitu-tively endocytosed and recycled to the plasma mem-brane via a Rab11a-dependent mechanism that has apositive influence on PTHrP secretion but is not re-quired for the activation of ERK. The protein level ofCaR remains constant for up to 4 h of ligand-inducedactivation; nonetheless, CaR interaction with AMSHmodifies the trafficking of this receptor by redirectingit to degradation. The functional impact of CaR deg-radation is reflected in a decrease of PTHrP secretion.Our studies suggest that CaR-regulated hormonal se-cretion is influenced by the vesicular trafficking of thisreceptor. In particular, PTHrP secretion requires CaRconstitutive intracellular trafficking, and this process isattenuated by AMSH, a CaR-interacting protein thatdirects the receptor to degradation.
RESULTS
CaR Endocytosis and Trafficking Occurs inEndosomes Containing Rab11a
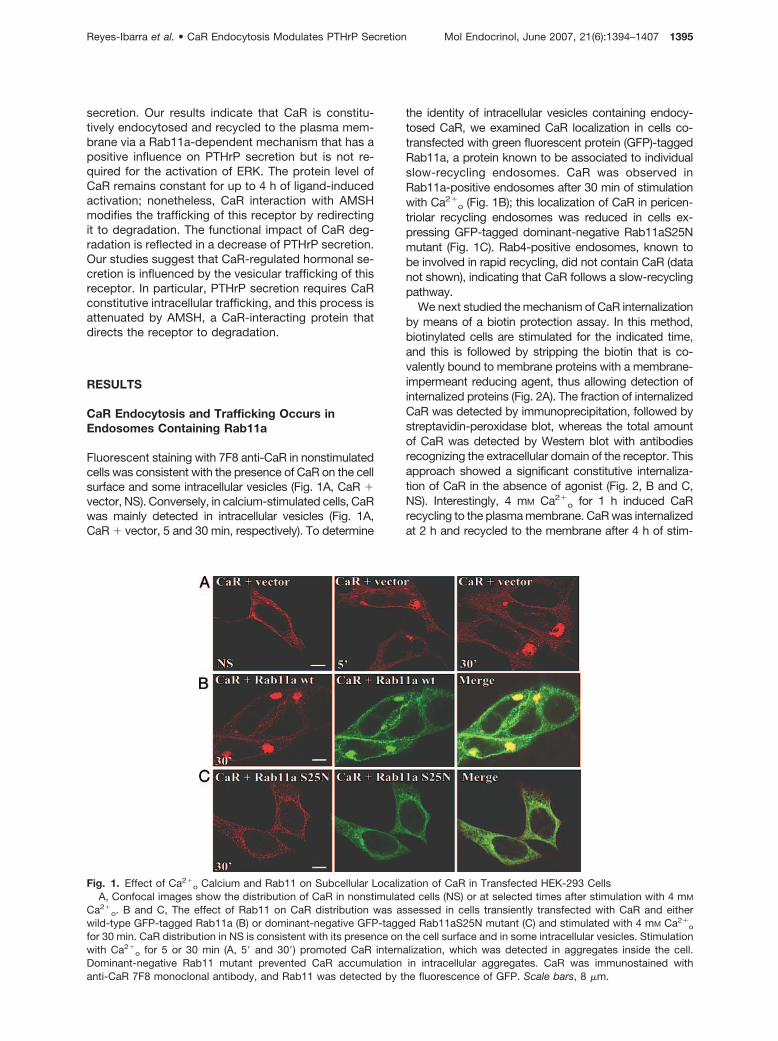
Fluorescent staining with 7F8 anti-CaR in nonstimulatedcells was consistent with the presence of CaR on the cellsurface and some intracellular vesicles (Fig. 1A, CaR �vector, NS). Conversely, in calcium-stimulated cells, CaRwas mainly detected in intracellular vesicles (Fig. 1A,CaR � vector, 5 and 30 min, respectively). To determine
the identity of intracellular vesicles containing endocy-tosed CaR, we examined CaR localization in cells co-transfected with green fluorescent protein (GFP)-taggedRab11a, a protein known to be associated to individualslow-recycling endosomes. CaR was observed inRab11a-positive endosomes after 30 min of stimulationwith Ca2�
o (Fig. 1B); this localization of CaR in pericen-triolar recycling endosomes was reduced in cells ex-pressing GFP-tagged dominant-negative Rab11aS25Nmutant (Fig. 1C). Rab4-positive endosomes, known tobe involved in rapid recycling, did not contain CaR (datanot shown), indicating that CaR follows a slow-recyclingpathway.
We next studied the mechanism of CaR internalizationby means of a biotin protection assay. In this method,biotinylated cells are stimulated for the indicated time,and this is followed by stripping the biotin that is co-valently bound to membrane proteins with a membrane-impermeant reducing agent, thus allowing detection ofinternalized proteins (Fig. 2A). The fraction of internalizedCaR was detected by immunoprecipitation, followed bystreptavidin-peroxidase blot, whereas the total amountof CaR was detected by Western blot with antibodiesrecognizing the extracellular domain of the receptor. Thisapproach showed a significant constitutive internaliza-tion of CaR in the absence of agonist (Fig. 2, B and C,NS). Interestingly, 4 mM Ca2�
o for 1 h induced CaRrecycling to the plasma membrane. CaR was internalizedat 2 h and recycled to the membrane after 4 h of stim-
Fig. 1. Effect of Ca2�o Calcium and Rab11 on Subcellular Localization of CaR in Transfected HEK-293 Cells
A, Confocal images show the distribution of CaR in nonstimulated cells (NS) or at selected times after stimulation with 4 mM
Ca2�o. B and C, The effect of Rab11 on CaR distribution was assessed in cells transiently transfected with CaR and either
wild-type GFP-tagged Rab11a (B) or dominant-negative GFP-tagged Rab11aS25N mutant (C) and stimulated with 4 mM Ca2�o
for 30 min. CaR distribution in NS is consistent with its presence on the cell surface and in some intracellular vesicles. Stimulationwith Ca2�
o for 5 or 30 min (A, 5� and 30�) promoted CaR internalization, which was detected in aggregates inside the cell.Dominant-negative Rab11 mutant prevented CaR accumulation in intracellular aggregates. CaR was immunostained withanti-CaR 7F8 monoclonal antibody, and Rab11 was detected by the fluorescence of GFP. Scale bars, 8 �m.
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1395
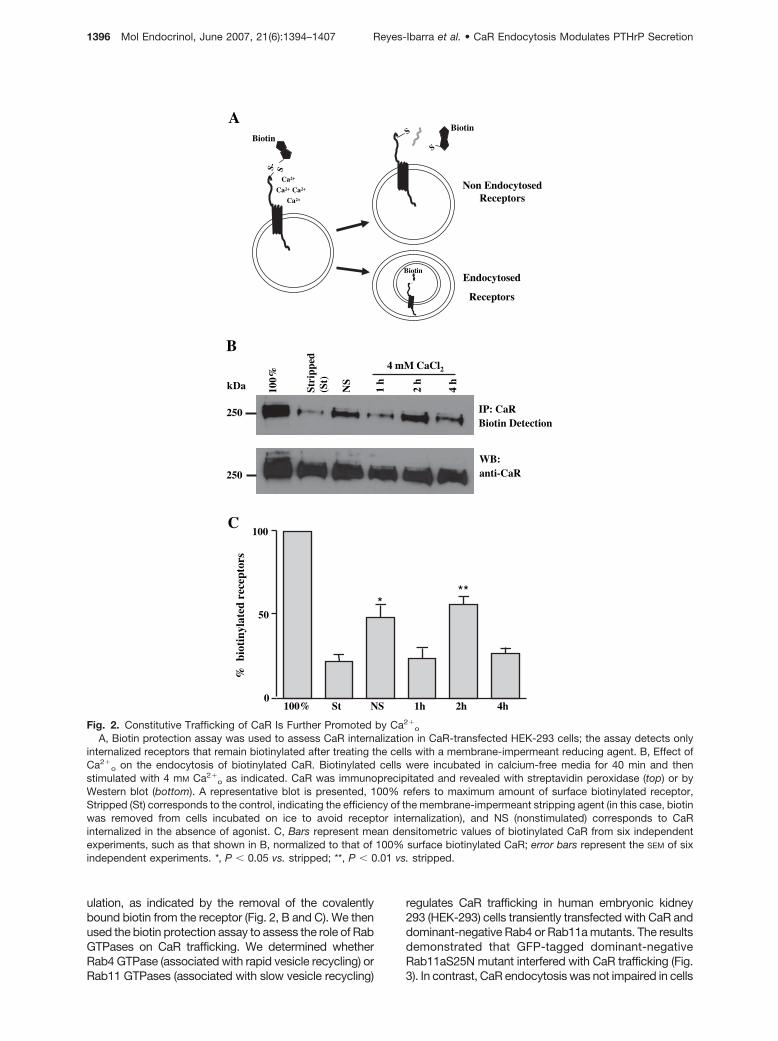
ulation, as indicated by the removal of the covalentlybound biotin from the receptor (Fig. 2, B and C). We thenused the biotin protection assay to assess the role of RabGTPases on CaR trafficking. We determined whetherRab4 GTPase (associated with rapid vesicle recycling) orRab11 GTPases (associated with slow vesicle recycling)
regulates CaR trafficking in human embryonic kidney293 (HEK-293) cells transiently transfected with CaR anddominant-negative Rab4 or Rab11a mutants. The resultsdemonstrated that GFP-tagged dominant-negativeRab11aS25N mutant interfered with CaR trafficking (Fig.3). In contrast, CaR endocytosis was not impaired in cells
A
B
250
250
C
100% St NS 1h 2h 4h0
50
100
% b
ioti
nyla
ted
rece
ptor
s
S- S
S
SBiotin
Biotin
Biotin
Non Endocytosed Receptors
Endocytosed
Receptors
Ca2+
Ca2+
Ca2+
Ca2+
***
100%
Stri
pped
(St)
NS
1 h
2 h
4 h
4 mM CaCl2
IP: CaRBiotin Detection
WB: anti-CaR
kDa
Fig. 2. Constitutive Trafficking of CaR Is Further Promoted by Ca2�o
A, Biotin protection assay was used to assess CaR internalization in CaR-transfected HEK-293 cells; the assay detects onlyinternalized receptors that remain biotinylated after treating the cells with a membrane-impermeant reducing agent. B, Effect ofCa2�
o on the endocytosis of biotinylated CaR. Biotinylated cells were incubated in calcium-free media for 40 min and thenstimulated with 4 mM Ca2�
o as indicated. CaR was immunoprecipitated and revealed with streptavidin peroxidase (top) or byWestern blot (bottom). A representative blot is presented, 100% refers to maximum amount of surface biotinylated receptor,Stripped (St) corresponds to the control, indicating the efficiency of the membrane-impermeant stripping agent (in this case, biotinwas removed from cells incubated on ice to avoid receptor internalization), and NS (nonstimulated) corresponds to CaRinternalized in the absence of agonist. C, Bars represent mean densitometric values of biotinylated CaR from six independentexperiments, such as that shown in B, normalized to that of 100% surface biotinylated CaR; error bars represent the SEM of sixindependent experiments. *, P � 0.05 vs. stripped; **, P � 0.01 vs. stripped.
1396 Mol Endocrinol, June 2007, 21(6):1394–1407 Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion
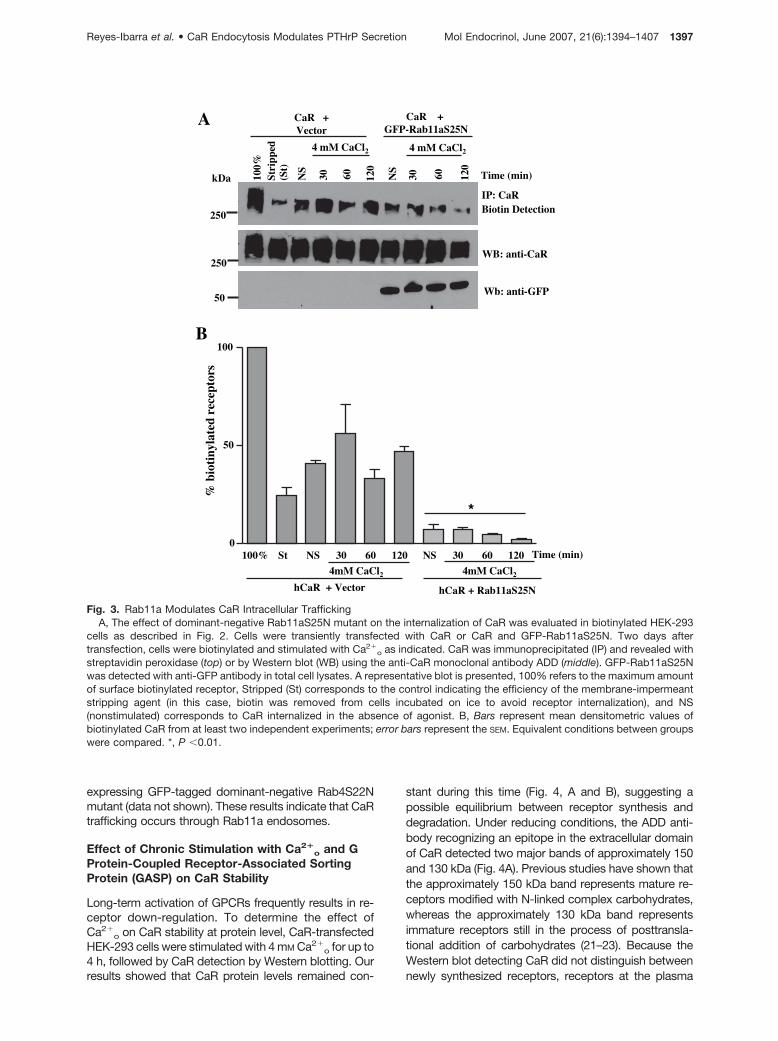
expressing GFP-tagged dominant-negative Rab4S22Nmutant (data not shown). These results indicate that CaRtrafficking occurs through Rab11a endosomes.
Effect of Chronic Stimulation with Ca2�o and G
Protein-Coupled Receptor-Associated SortingProtein (GASP) on CaR Stability
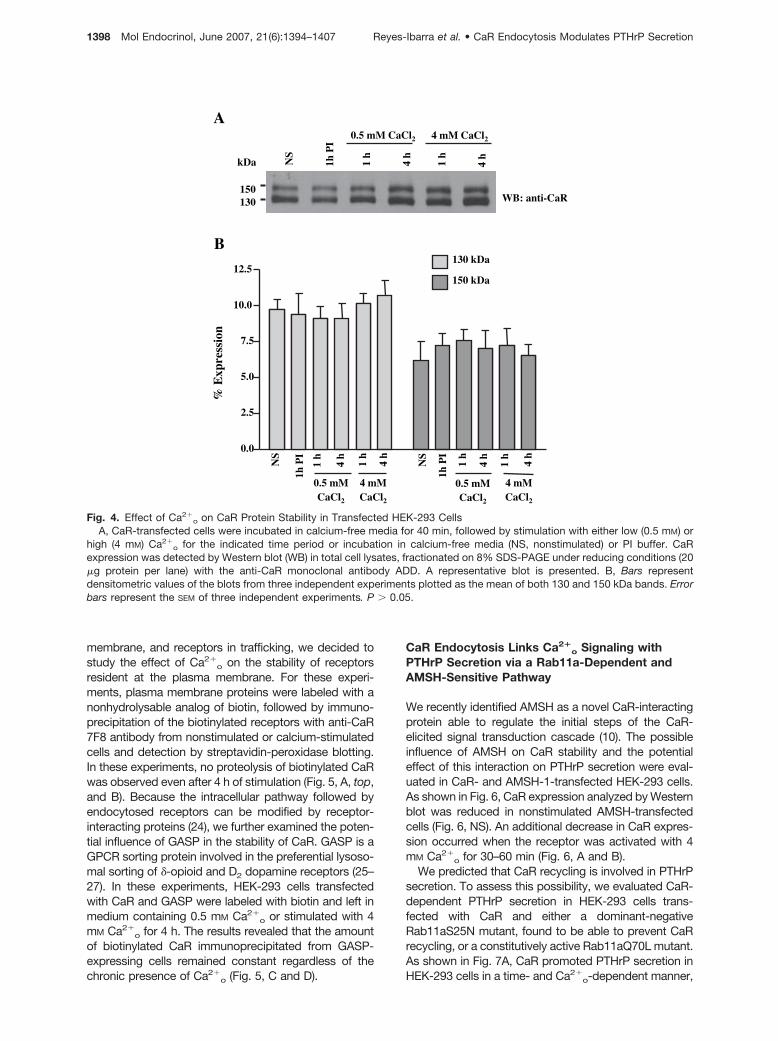
Long-term activation of GPCRs frequently results in re-ceptor down-regulation. To determine the effect ofCa2�
o on CaR stability at protein level, CaR-transfectedHEK-293 cells were stimulated with 4 mM Ca2�
o for up to4 h, followed by CaR detection by Western blotting. Ourresults showed that CaR protein levels remained con-
stant during this time (Fig. 4, A and B), suggesting apossible equilibrium between receptor synthesis anddegradation. Under reducing conditions, the ADD anti-body recognizing an epitope in the extracellular domainof CaR detected two major bands of approximately 150and 130 kDa (Fig. 4A). Previous studies have shown thatthe approximately 150 kDa band represents mature re-ceptors modified with N-linked complex carbohydrates,whereas the approximately 130 kDa band representsimmature receptors still in the process of posttransla-tional addition of carbohydrates (21–23). Because theWestern blot detecting CaR did not distinguish betweennewly synthesized receptors, receptors at the plasma
A
B
100%
NS
30 60 120
4 mM CaCl2
NS
30 60 120
4 mM CaCl2
CaR + Vector
CaR +GFP-Rab11aS25N
Wb: anti-GFP
WB: anti-CaR
250
Time (min)
250
50
Stri
pped
(St)
kDa
IP: CaRBiotin Detection
100% St NS 30 60 120 NS 30 60 120 0
50
100
hCaR + Vector
4mM CaCl2 4mM CaCl2
hCaR + Rab11aS25N
Time (min)
% b
ioti
nyla
ted
rece
ptor
s
*
Fig. 3. Rab11a Modulates CaR Intracellular TraffickingA, The effect of dominant-negative Rab11aS25N mutant on the internalization of CaR was evaluated in biotinylated HEK-293
cells as described in Fig. 2. Cells were transiently transfected with CaR or CaR and GFP-Rab11aS25N. Two days aftertransfection, cells were biotinylated and stimulated with Ca2�
o as indicated. CaR was immunoprecipitated (IP) and revealed withstreptavidin peroxidase (top) or by Western blot (WB) using the anti-CaR monoclonal antibody ADD (middle). GFP-Rab11aS25Nwas detected with anti-GFP antibody in total cell lysates. A representative blot is presented, 100% refers to the maximum amountof surface biotinylated receptor, Stripped (St) corresponds to the control indicating the efficiency of the membrane-impermeantstripping agent (in this case, biotin was removed from cells incubated on ice to avoid receptor internalization), and NS(nonstimulated) corresponds to CaR internalized in the absence of agonist. B, Bars represent mean densitometric values ofbiotinylated CaR from at least two independent experiments; error bars represent the SEM. Equivalent conditions between groupswere compared. *, P �0.01.
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1397
membrane, and receptors in trafficking, we decided tostudy the effect of Ca2�
o on the stability of receptorsresident at the plasma membrane. For these experi-ments, plasma membrane proteins were labeled with anonhydrolysable analog of biotin, followed by immuno-precipitation of the biotinylated receptors with anti-CaR7F8 antibody from nonstimulated or calcium-stimulatedcells and detection by streptavidin-peroxidase blotting.In these experiments, no proteolysis of biotinylated CaRwas observed even after 4 h of stimulation (Fig. 5, A, top,and B). Because the intracellular pathway followed byendocytosed receptors can be modified by receptor-interacting proteins (24), we further examined the poten-tial influence of GASP in the stability of CaR. GASP is aGPCR sorting protein involved in the preferential lysoso-mal sorting of �-opioid and D2 dopamine receptors (25–27). In these experiments, HEK-293 cells transfectedwith CaR and GASP were labeled with biotin and left inmedium containing 0.5 mM Ca2�
o or stimulated with 4mM Ca2�
o for 4 h. The results revealed that the amountof biotinylated CaR immunoprecipitated from GASP-expressing cells remained constant regardless of thechronic presence of Ca2�
o (Fig. 5, C and D).
CaR Endocytosis Links Ca2�o Signaling with
PTHrP Secretion via a Rab11a-Dependent andAMSH-Sensitive Pathway
We recently identified AMSH as a novel CaR-interactingprotein able to regulate the initial steps of the CaR-elicited signal transduction cascade (10). The possibleinfluence of AMSH on CaR stability and the potentialeffect of this interaction on PTHrP secretion were eval-uated in CaR- and AMSH-1-transfected HEK-293 cells.As shown in Fig. 6, CaR expression analyzed by Westernblot was reduced in nonstimulated AMSH-transfectedcells (Fig. 6, NS). An additional decrease in CaR expres-sion occurred when the receptor was activated with 4mM Ca2�
o for 30–60 min (Fig. 6, A and B).We predicted that CaR recycling is involved in PTHrP
secretion. To assess this possibility, we evaluated CaR-dependent PTHrP secretion in HEK-293 cells trans-fected with CaR and either a dominant-negativeRab11aS25N mutant, found to be able to prevent CaRrecycling, or a constitutively active Rab11aQ70L mutant.As shown in Fig. 7A, CaR promoted PTHrP secretion inHEK-293 cells in a time- and Ca2�
o-dependent manner,
A
B130 kDa
150 kDa
0.0
2.5
5.0
7.5
10.0
12.5
% E
xpre
ssio
n
NS
1h P
I
1 h
1 h
4 h
4 h
0.5 mM CaCl2
4 mM CaCl2
NS
1h P
I
1 h
1 h
4 h
4 h
0.5 mM CaCl2
4 mM CaCl2
NS
1h P
I
1 h
1 h
4 h
4 h
0.5 mM CaCl2 4 mM CaCl2
150 130
kDa
WB: anti-CaR
Fig. 4. Effect of Ca2�o on CaR Protein Stability in Transfected HEK-293 Cells
A, CaR-transfected cells were incubated in calcium-free media for 40 min, followed by stimulation with either low (0.5 mM) orhigh (4 mM) Ca2�
o for the indicated time period or incubation in calcium-free media (NS, nonstimulated) or PI buffer. CaRexpression was detected by Western blot (WB) in total cell lysates, fractionated on 8% SDS-PAGE under reducing conditions (20�g protein per lane) with the anti-CaR monoclonal antibody ADD. A representative blot is presented. B, Bars representdensitometric values of the blots from three independent experiments plotted as the mean of both 130 and 150 kDa bands. Errorbars represent the SEM of three independent experiments. P � 0.05.
1398 Mol Endocrinol, June 2007, 21(6):1394–1407 Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion
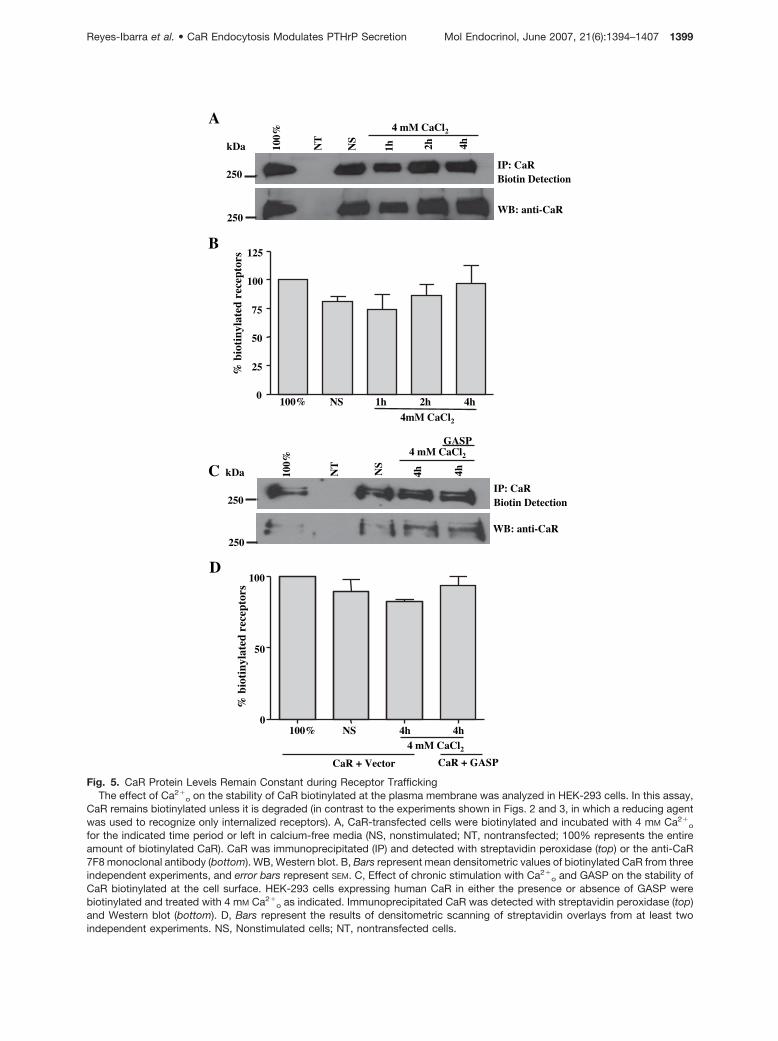
A
4h
4 mM CaCl2
100%
250
NS
1h 2hNT
kDa
IP: CaRBiotin Detection
WB: anti-CaR250
B
GASP
100%
NT
NS
4h 4h
4 mM CaCl2
IP: CaRBiotin Detection
WB: anti-CaR
kDa
250
250
C
D
4mM CaCl2
100% NS 1h 2h 4h0
25
50
75
100
125
% b
ioti
nyla
ted
rece
ptor
s
100% NS 4h 4h0
50
100
4 mM CaCl2
CaR + GASPCaR + Vector
% b
ioti
nyla
ted
rece
ptor
s
Fig. 5. CaR Protein Levels Remain Constant during Receptor TraffickingThe effect of Ca2�
o on the stability of CaR biotinylated at the plasma membrane was analyzed in HEK-293 cells. In this assay,CaR remains biotinylated unless it is degraded (in contrast to the experiments shown in Figs. 2 and 3, in which a reducing agentwas used to recognize only internalized receptors). A, CaR-transfected cells were biotinylated and incubated with 4 mM Ca2�
o
for the indicated time period or left in calcium-free media (NS, nonstimulated; NT, nontransfected; 100% represents the entireamount of biotinylated CaR). CaR was immunoprecipitated (IP) and detected with streptavidin peroxidase (top) or the anti-CaR7F8 monoclonal antibody (bottom). WB, Western blot. B, Bars represent mean densitometric values of biotinylated CaR from threeindependent experiments, and error bars represent SEM. C, Effect of chronic stimulation with Ca2�
o and GASP on the stability ofCaR biotinylated at the cell surface. HEK-293 cells expressing human CaR in either the presence or absence of GASP werebiotinylated and treated with 4 mM Ca2�
o as indicated. Immunoprecipitated CaR was detected with streptavidin peroxidase (top)and Western blot (bottom). D, Bars represent the results of densitometric scanning of streptavidin overlays from at least twoindependent experiments. NS, Nonstimulated cells; NT, nontransfected cells.
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1399
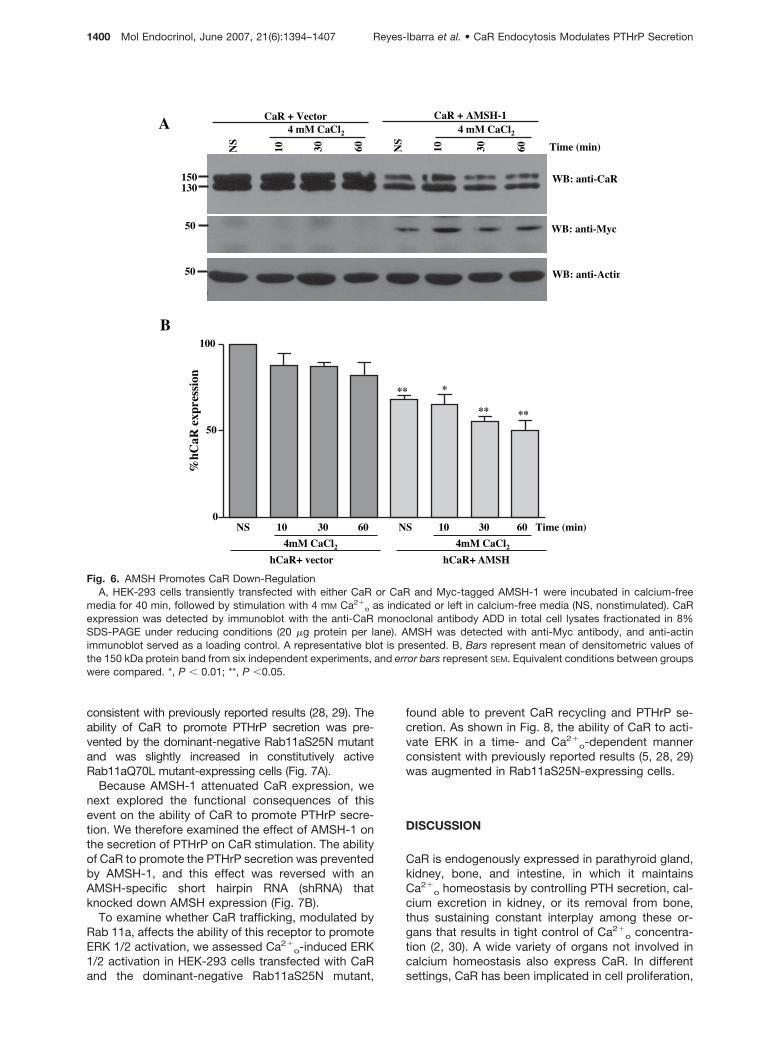
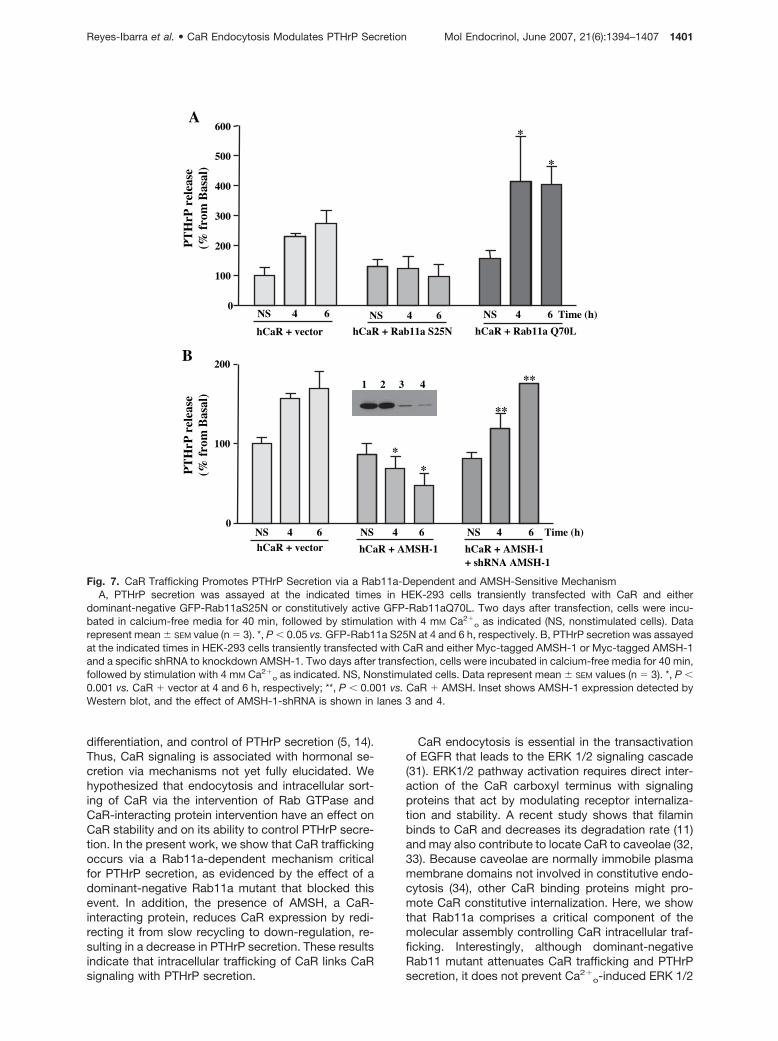
consistent with previously reported results (28, 29). Theability of CaR to promote PTHrP secretion was pre-vented by the dominant-negative Rab11aS25N mutantand was slightly increased in constitutively activeRab11aQ70L mutant-expressing cells (Fig. 7A).
Because AMSH-1 attenuated CaR expression, wenext explored the functional consequences of thisevent on the ability of CaR to promote PTHrP secre-tion. We therefore examined the effect of AMSH-1 onthe secretion of PTHrP on CaR stimulation. The abilityof CaR to promote the PTHrP secretion was preventedby AMSH-1, and this effect was reversed with anAMSH-specific short hairpin RNA (shRNA) thatknocked down AMSH expression (Fig. 7B).
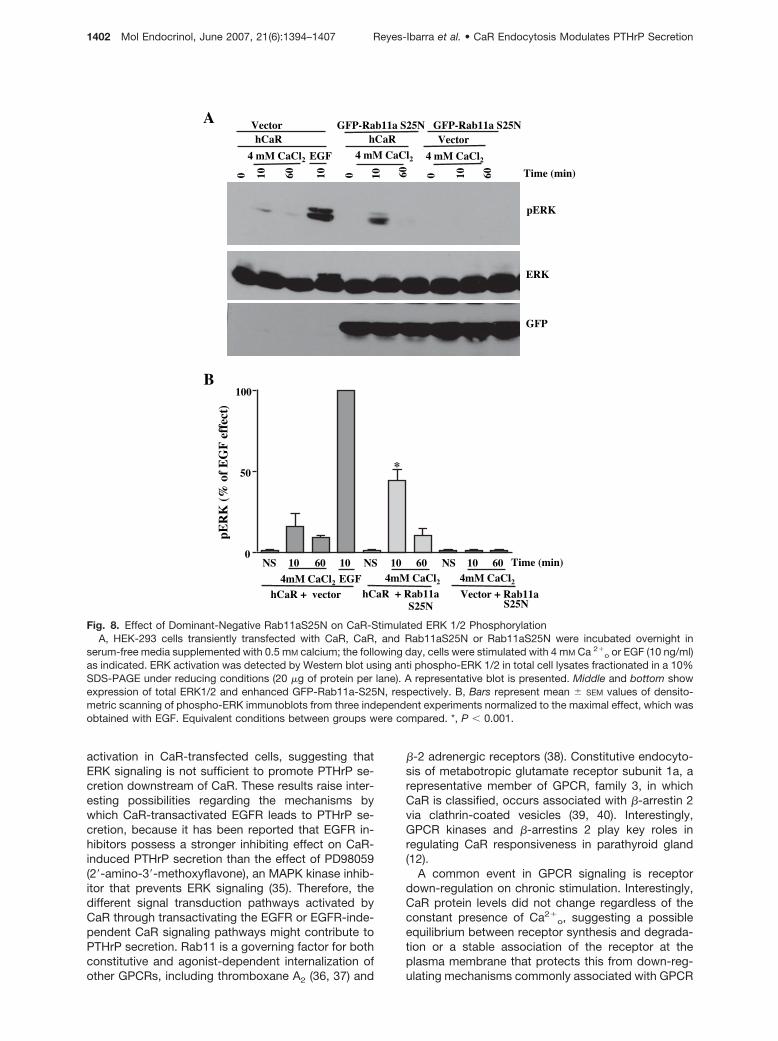
To examine whether CaR trafficking, modulated byRab 11a, affects the ability of this receptor to promoteERK 1/2 activation, we assessed Ca2�
o-induced ERK1/2 activation in HEK-293 cells transfected with CaRand the dominant-negative Rab11aS25N mutant,
found able to prevent CaR recycling and PTHrP se-cretion. As shown in Fig. 8, the ability of CaR to acti-vate ERK in a time- and Ca2�
o-dependent mannerconsistent with previously reported results (5, 28, 29)was augmented in Rab11aS25N-expressing cells.
DISCUSSION
CaR is endogenously expressed in parathyroid gland,kidney, bone, and intestine, in which it maintainsCa2�
o homeostasis by controlling PTH secretion, cal-cium excretion in kidney, or its removal from bone,thus sustaining constant interplay among these or-gans that results in tight control of Ca2�
o concentra-tion (2, 30). A wide variety of organs not involved incalcium homeostasis also express CaR. In differentsettings, CaR has been implicated in cell proliferation,
A
150130
NS
10 30 60
4 mM CaCl2
NS
10 30 60
4 mM CaCl2
CaR + Vector CaR + AMSH-1
WB: anti-CaR
WB: anti-Myc
WB: anti-Actin
Time (min)
50
50
B
NS 10 30 60 NS 10 30 60 Time (min)0
50
100
4mM CaCl2 4mM CaCl2
****
** *
hCaR+ vector hCaR+ AMSH
%hC
aR e
xpre
ssio
n
Fig. 6. AMSH Promotes CaR Down-RegulationA, HEK-293 cells transiently transfected with either CaR or CaR and Myc-tagged AMSH-1 were incubated in calcium-free
media for 40 min, followed by stimulation with 4 mM Ca2�o as indicated or left in calcium-free media (NS, nonstimulated). CaR
expression was detected by immunoblot with the anti-CaR monoclonal antibody ADD in total cell lysates fractionated in 8%SDS-PAGE under reducing conditions (20 �g protein per lane). AMSH was detected with anti-Myc antibody, and anti-actinimmunoblot served as a loading control. A representative blot is presented. B, Bars represent mean of densitometric values ofthe 150 kDa protein band from six independent experiments, and error bars represent SEM. Equivalent conditions between groupswere compared. *, P � 0.01; **, P �0.05.
1400 Mol Endocrinol, June 2007, 21(6):1394–1407 Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion
differentiation, and control of PTHrP secretion (5, 14).Thus, CaR signaling is associated with hormonal se-cretion via mechanisms not yet fully elucidated. Wehypothesized that endocytosis and intracellular sort-ing of CaR via the intervention of Rab GTPase andCaR-interacting protein intervention have an effect onCaR stability and on its ability to control PTHrP secre-tion. In the present work, we show that CaR traffickingoccurs via a Rab11a-dependent mechanism criticalfor PTHrP secretion, as evidenced by the effect of adominant-negative Rab11a mutant that blocked thisevent. In addition, the presence of AMSH, a CaR-interacting protein, reduces CaR expression by redi-recting it from slow recycling to down-regulation, re-sulting in a decrease in PTHrP secretion. These resultsindicate that intracellular trafficking of CaR links CaRsignaling with PTHrP secretion.
CaR endocytosis is essential in the transactivationof EGFR that leads to the ERK 1/2 signaling cascade(31). ERK1/2 pathway activation requires direct inter-action of the CaR carboxyl terminus with signalingproteins that act by modulating receptor internaliza-tion and stability. A recent study shows that filaminbinds to CaR and decreases its degradation rate (11)and may also contribute to locate CaR to caveolae (32,33). Because caveolae are normally immobile plasmamembrane domains not involved in constitutive endo-cytosis (34), other CaR binding proteins might pro-mote CaR constitutive internalization. Here, we showthat Rab11a comprises a critical component of themolecular assembly controlling CaR intracellular traf-ficking. Interestingly, although dominant-negativeRab11 mutant attenuates CaR trafficking and PTHrPsecretion, it does not prevent Ca2�
o-induced ERK 1/2
A
B
1 2 3 4
0
100
200
NS 4 6 NS 4 6 NS 4 6 Time (h)hCaR + vector hCaR + AMSH-1 hCaR + AMSH-1
+ shRNA AMSH-1
PT
HrP
rel
ease
(% f
rom
Bas
al)
*
*
**
**
0
100
200
300
400
500
600
NS 4 6NS 4 6 NS 4 6 Time (h)
hCaR + vector hCaR + Rab11a S25N hCaR + Rab11a Q70L
PT
HrP
rel
ease
(%fr
om B
asal
)
*
*
Fig. 7. CaR Trafficking Promotes PTHrP Secretion via a Rab11a-Dependent and AMSH-Sensitive MechanismA, PTHrP secretion was assayed at the indicated times in HEK-293 cells transiently transfected with CaR and either
dominant-negative GFP-Rab11aS25N or constitutively active GFP-Rab11aQ70L. Two days after transfection, cells were incu-bated in calcium-free media for 40 min, followed by stimulation with 4 mM Ca2�
o as indicated (NS, nonstimulated cells). Datarepresent mean � SEM value (n � 3). *, P � 0.05 vs. GFP-Rab11a S25N at 4 and 6 h, respectively. B, PTHrP secretion was assayedat the indicated times in HEK-293 cells transiently transfected with CaR and either Myc-tagged AMSH-1 or Myc-tagged AMSH-1and a specific shRNA to knockdown AMSH-1. Two days after transfection, cells were incubated in calcium-free media for 40 min,followed by stimulation with 4 mM Ca2�
o as indicated. NS, Nonstimulated cells. Data represent mean � SEM values (n � 3). *, P �0.001 vs. CaR � vector at 4 and 6 h, respectively; **, P � 0.001 vs. CaR � AMSH. Inset shows AMSH-1 expression detected byWestern blot, and the effect of AMSH-1-shRNA is shown in lanes 3 and 4.
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1401
activation in CaR-transfected cells, suggesting thatERK signaling is not sufficient to promote PTHrP se-cretion downstream of CaR. These results raise inter-esting possibilities regarding the mechanisms bywhich CaR-transactivated EGFR leads to PTHrP se-cretion, because it has been reported that EGFR in-hibitors possess a stronger inhibiting effect on CaR-induced PTHrP secretion than the effect of PD98059(2�-amino-3�-methoxyflavone), an MAPK kinase inhib-itor that prevents ERK signaling (35). Therefore, thedifferent signal transduction pathways activated byCaR through transactivating the EGFR or EGFR-inde-pendent CaR signaling pathways might contribute toPTHrP secretion. Rab11 is a governing factor for bothconstitutive and agonist-dependent internalization ofother GPCRs, including thromboxane A2 (36, 37) and
�-2 adrenergic receptors (38). Constitutive endocyto-sis of metabotropic glutamate receptor subunit 1a, arepresentative member of GPCR, family 3, in whichCaR is classified, occurs associated with �-arrestin 2via clathrin-coated vesicles (39, 40). Interestingly,GPCR kinases and �-arrestins 2 play key roles inregulating CaR responsiveness in parathyroid gland(12).
A common event in GPCR signaling is receptordown-regulation on chronic stimulation. Interestingly,CaR protein levels did not change regardless of theconstant presence of Ca2�
o, suggesting a possibleequilibrium between receptor synthesis and degrada-tion or a stable association of the receptor at theplasma membrane that protects this from down-reg-ulating mechanisms commonly associated with GPCR
NS 10 60 10 NS 10 60 NS 10 600
50
100
4mM CaCl2 4mM CaCl2 4mM CaCl2
hCaR + vector hCaR + Rab11a S25N
Vector + Rab11a S25N
EGF
Time (min)
A
B
pER
K(%
of
EG
F e
ffec
t)pERK
GFP
ERK
4 mM CaCl2
hCaR
4 mM CaCl2
hCaR
10 60
EGF
10 Time (min)0 10 60
Vector GFP-Rab11a S25NVector
0 10 60
GFP-Rab11a S25N
4 mM CaCl2
0
*
Fig. 8. Effect of Dominant-Negative Rab11aS25N on CaR-Stimulated ERK 1/2 PhosphorylationA, HEK-293 cells transiently transfected with CaR, CaR, and Rab11aS25N or Rab11aS25N were incubated overnight in
serum-free media supplemented with 0.5 mM calcium; the following day, cells were stimulated with 4 mM Ca 2�o or EGF (10 ng/ml)
as indicated. ERK activation was detected by Western blot using anti phospho-ERK 1/2 in total cell lysates fractionated in a 10%SDS-PAGE under reducing conditions (20 �g of protein per lane). A representative blot is presented. Middle and bottom showexpression of total ERK1/2 and enhanced GFP-Rab11a-S25N, respectively. B, Bars represent mean � SEM values of densito-metric scanning of phospho-ERK immunoblots from three independent experiments normalized to the maximal effect, which wasobtained with EGF. Equivalent conditions between groups were compared. *, P � 0.001.
1402 Mol Endocrinol, June 2007, 21(6):1394–1407 Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion

internalization. These possibilities were assessed inexperiments using biotinylated cells with either reduc-ing agent-sensitive biotin that remains bound to re-ceptors if these are internalized or with noncleavablebiotin to assess the fate of internalized receptors. Ourresults indicate that CaR is subject to constant intra-cellular trafficking. Our data also indicate that CaR isnot degraded after ligand-induced activation. Contraryto our findings, it has been reported that m-calpainparticipates in CaR degradation in caveolae (41). Weevaluated whether intracellular sorting is critical forCaR degradation. We first evaluated the effect ofGASP, a well-characterized GPCR sorting protein crit-ical for �-opioid and D2 dopamine receptor degrada-tion (26, 27). GASP shows affinity for a broad variety ofGPCRs, including members of the metabotropic glu-tamate receptor family in which CaR is included (42).Our results indicate that GASP does not promote CaRdegradation, suggesting that additional proteins arerequired to promote CaR down-regulation. In this re-gard, AMSH, a CaR carboxyl-terminal interacting pro-tein, was able to promote CaR degradation. Interest-ingly, CaR ubiquitination and degradation are linkedwith the activity of E3 ubiquitin ligases such as dorfin(43), which might link CaR intracellular sorting withdegradation, affecting its role as a controller of hor-monal secretion. Together, these findings suggest thatAMSH redefines the sorting of endocytosed CaR fromslow recycling to degradative pathways, perhapsthrough an interaction with the multiprotein complexformed by AMSH and RNF11 (ring finger protein 11)-Smurf2 (SMAD-specific E3 ubiquitin protein ligase 2)(44) that may lead CaR to degradation.
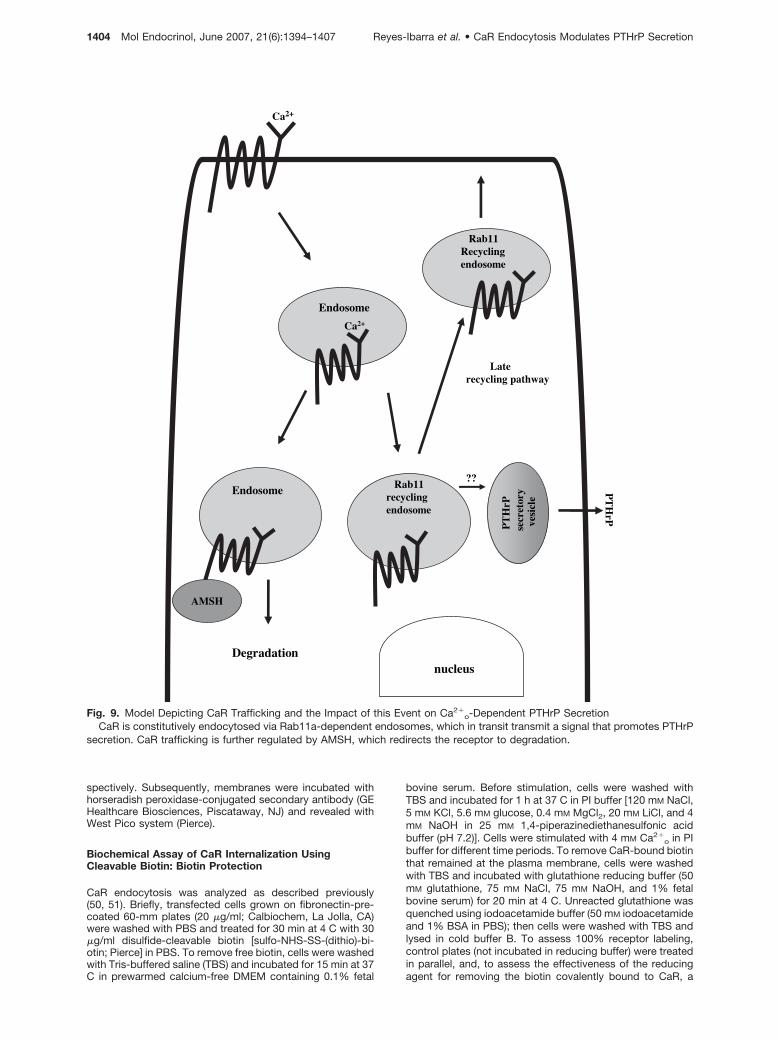
Finally, our results showing the effect of the domi-nant-negative Rab11a mutant that attenuated CaR-dependent PTHrP secretion and the role of AMSHdecreasing PTHrP secretion suggest that CaR modu-lates PTHrP secretion by a mechanism described inthe model illustrated in Fig. 9. Accordingly, CaR isconstitutively subjected to intracellular trafficking viaRab11a-dependent endosomes, which stimulatePTHrP secretion by transmitting a signal that pro-motes the trafficking of secretory vesicles releasingPTHrP outside the cell (45–47). This process is furtherregulated by AMSH, which redirects CaR to degrada-tive pathways. This model, based on results obtainedin transfected HEK-293 (which are frequently used asa working model for studies oriented toward definingthe molecular basis of CaR signaling because these donot express endogenous CaR) sets forth a workinghypothesis to be assessed in cells endogenously ex-pressing this receptor for further defining the physio-logic consequences of the findings described herein.
In conclusion, our results support a role for CaRendocytosis and recycling via Rab11a-dependent en-dosomes as a critical step in signal transmission topromote PTHrP secretion. This process is interferedby AMSH, a CaR-interacting protein that redirects CaRfrom recycling to degradation pathways, thus attenu-ating its ability to control PTHrP secretion.
MATERIALS AND METHODS
DNA Constructs and Antibodies
Wild-type human CaR cDNA was kindly provided by Dr. AllenSpiegel (National Institute of Diabetes and Digestive and KidneyDiseases/National Institutes of Health, Bethesda, MD). GASPcDNA and full-length human AMSH-1 were obtained from Dr.Mark von Zastrow (University of California, San Francisco, SanFrancisco, CA) and Dr. Itoh Susumu (Tohoku University of Med-icine, Sendai, Japan), respectively. Rab11 and Rab4 constructswere donated by Dr. Tamas Balla (National Institute of ChildHealth and Human Development/National Institutes of Health,Bethesda, MD). Anti-CaR ADD antibody raised against a syn-thetic peptide corresponding to residues 214–235 of humanCaR and anti-CaR 7F8 monoclonal antibody were donated byDr. Allen Spiegel (48). shRNA for AMSH-1, which targets theAMSH-1 sequence reported previously (49), was generatedbased on the following hairpin sequence, TGCTGTTGACAGTGAGCGCGTTACAAATCTGCTGTCATTTTTTAGTGAAGCCACAGATGTAAAATGACAGCAGATTTGTAATTCATGCCTACTGCCTCGGA, and cloned into the pSM2 vector (http://codex.cshl.org/scripts/newdisplayhairpin.cgi?row�4&query_id�24529_no_one_0_0_0_0klokos.cshl.edu).
Cell Culture
HEK-293 cells were routinely cultured in DMEM (Sigma, St.Louis, MO) supplemented with 10% fetal bovine serum, 1%glutamine, penicillin, and streptomycin at 37 C in a 5% CO2atmosphere.
Transient Transfections
HEK-293 cells were transfected with 4 �g plasmid DNA di-luted in DMEM (Sigma) using Lipofectamine reagent accord-ing to the instructions of the manufacturer (Invitrogen, Carls-bad, CA). The mixture was incubated at room temperature for30 min. The DNA-Lipofectamine complex was further dilutedin 6 ml of serum-free DMEM and was added to 80% confluentcells plated in 100-mm Petri dishes. After 5 h of incubation,10 ml DMEM containing 10% fetal bovine serum was added.Twenty-four hours after transfection, cells were split into60-mm Petri dishes, cultured in completed DMEM, and cul-tured for another 24 h.
Immunoblot Analysis
Confluent cells in 60-mm plates were placed on ice, rinsedwith ice-cold PBS, and scraped with buffer B [20 mM Tris-HCl(pH 7.5), 1 mM EDTA, 150 mM NaCl, 1% Triton X-100, 1 mM
sodium vanadate, 1 mM NaF, 10 mM �-glycerophosphate, 1mM phenylmethylsulfonyl fluoride, 10 �g/ml leupeptin, and 10�g/ml aprotinin]. The extracts were forced through a 22-gauge needle five to eight times and centrifuged for 15 min at14,000 rpm at 4 C. The protein concentration was determinedby the bicinchoninic acid BCA Protein Assay (Pierce, Rock-ford, IL), and approximately 30 �g of each sample was sep-arated on 6 or 7.5% SDS-PAGE for CaR or AMSH blots,respectively, and electrotransferred to nitrocellulose mem-branes. CaR was detected with 0.1 �g/ml ADD anti-CaRantibody, and Myc-tagged AMSH was detected with anti-Myc antibody (MMS-150R; Covance, Berkeley, CA). ERK 1/2activation was detected by Western blot using a rabbit poly-clonal anti-phospho-ERK 1/2 antibody (9101L; Cell SignalingTechnology, Beverly, MA), and expression of endogenousERK 1/2 or enhanced GFP-tagged transfected proteins weredetected with a rabbit polyclonal anti-ERK antibody (SC-154;Santa Cruz Biotechnology, Santa Cruz, CA) or a mousemonoclonal anti-GFP antibody (MMS-118R; Covance), re-
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1403
spectively. Subsequently, membranes were incubated withhorseradish peroxidase-conjugated secondary antibody (GEHealthcare Biosciences, Piscataway, NJ) and revealed withWest Pico system (Pierce).
Biochemical Assay of CaR Internalization UsingCleavable Biotin: Biotin Protection
CaR endocytosis was analyzed as described previously(50, 51). Briefly, transfected cells grown on fibronectin-pre-coated 60-mm plates (20 �g/ml; Calbiochem, La Jolla, CA)were washed with PBS and treated for 30 min at 4 C with 30�g/ml disulfide-cleavable biotin [sulfo-NHS-SS-(dithio)-bi-otin; Pierce] in PBS. To remove free biotin, cells were washedwith Tris-buffered saline (TBS) and incubated for 15 min at 37C in prewarmed calcium-free DMEM containing 0.1% fetal
bovine serum. Before stimulation, cells were washed withTBS and incubated for 1 h at 37 C in PI buffer [120 mM NaCl,5 mM KCl, 5.6 mM glucose, 0.4 mM MgCl2, 20 mM LiCl, and 4mM NaOH in 25 mM 1,4-piperazinediethanesulfonic acidbuffer (pH 7.2)]. Cells were stimulated with 4 mM Ca2�
o in PIbuffer for different time periods. To remove CaR-bound biotinthat remained at the plasma membrane, cells were washedwith TBS and incubated with glutathione reducing buffer (50mM glutathione, 75 mM NaCl, 75 mM NaOH, and 1% fetalbovine serum) for 20 min at 4 C. Unreacted glutathione wasquenched using iodoacetamide buffer (50 mM iodoacetamideand 1% BSA in PBS); then cells were washed with TBS andlysed in cold buffer B. To assess 100% receptor labeling,control plates (not incubated in reducing buffer) were treatedin parallel, and, to assess the effectiveness of the reducingagent for removing the biotin covalently bound to CaR, a
Endosome
Rab11recycling endosome
nucleus
PT
HrP
PT
HrP
vesi
cle
secr
etor
y
Ca2+
Ca2+
Endosome
AMSH
Rab11Recycling endosome
Laterecycling pathway
Degradation
??
Fig. 9. Model Depicting CaR Trafficking and the Impact of this Event on Ca2�o-Dependent PTHrP Secretion
CaR is constitutively endocytosed via Rab11a-dependent endosomes, which in transit transmit a signal that promotes PTHrPsecretion. CaR trafficking is further regulated by AMSH, which redirects the receptor to degradation.
1404 Mol Endocrinol, June 2007, 21(6):1394–1407 Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion

plate designated as stripped remained at 4 C throughout theexperiment. CaR was immunoprecipitated with anti-CaR 7F8monoclonal antibody and protein A/G agarose (Santa CruzBiotechnology). Samples were denatured in SDS samplebuffer under nonreducing conditions, resolved by 6% SDS-PAGE, and transferred to nitrocellulose. Biotinylated CaRwas visualized using Vectastain ABC immunoperoxidase re-agent (Vector Laboratories, Burlingame, CA) and the WestPico system (Pierce).
Biochemical Assay of CaR Degradation UsingNoncleavable Biotin
Degradation of surface-biotinylated CaR was analyzed asdescribed previously (50, 52). Briefly, transfected cells grownon fibronectin-precoated 60-mm plates were biotinylatedwith 100 �g/ml noncleavable sulfo-NHS-biotin (Pierce) for 30min at 4 C in PBS. Unreacted biotin was quenched andremoved by three successive washes with TBS. The platelabeled as 100% was chilled on ice to stop additional mem-brane trafficking. The plates for Ca2�
o stimulation were incu-bated with PI buffer for 1 h. Nonstimulated cells were thenplaced on ice. Cells were stimulated with 4 mM Ca2�
o in PIbuffer for different periods of time. Cells were lysed in coldbuffer B, and CaR was immunoprecipitated with 7F8 mono-clonal antibody. Immunoprecipitated CaR was denatured inSDS sample buffer under nonreducing conditions, resolvedby 6% SDS-PAGE, and transferred to nitrocellulose. Biotin-ylated CaR was visualized using Vectastain ABC immuno-peroxidase reagent (Vector Laboratories). For a set of exper-iments in which GASP was transfected, the Vectastain ABCimmunoperoxidase reagent was diluted at a ratio of at least1:2 to detect small differences in receptor degradation. CaRdegradation was detected by a decrease of biotinylated CaRin the immunoprecipitates.
Immunofluorescence
HEK-293 cells grown on fibronectin-precoated coverslipswere transfected with either CaR or CaR and GFP-taggedRab proteins (800 ng total DNA). After 48 h, cells werewashed with PBS and fixed with paraformaldehyde (4%) inPBS for 20 min; then coverslips were permeabilized withmethanol (100%) and washed with PBS. Fixed and perme-abilized cells were incubated with anti-CaR 7F8 monoclonalantibody for 1 h at 37 C, followed by incubation with asecondary rhodamine-conjugated antimouse IgG antibody(Jackson ImmunoResearch, West Grove, PA) for 45 min atroom temperature. After washing, cells were mounted onglass slides using ProLong Antifade (Invitrogen). Images wereacquired with a Leica (Nussloch, Germany) DMIRE2 confocallaser microscope.
PTHrP Secretion Immunoassay
To detect secreted PTHrP 1–34, HEK-293 cells in 10-cmdishes were transiently transfected with the indicated cDNAsor shRNA and were distributed the following day into 6-cmdishes. After 24 h, media was replaced with Ca2�-free DMEMsupplemented with 0.5 mM CaCl2 and incubated for 16 h at37 C, followed by stimulation with 4 mM Ca2�
o for 4 or 6 h.The medium was recovered and centrifuged for 5 min at13,000 rpm, and PTHrP was measured by the use of com-petitive enzyme immunoassay according to the instructionsof the manufacturer (Peninsula Laboratories, Belmont, CA).
Statistics
Statistical analyses were performed by one-way ANOVAwhen appropriate. P � 0.05 was considered significant using
GraphPad Prism version 2.0 software (GraphPad Software,San Diego, CA).
Acknowledgments
We thank Drs. Brent Miller and Alicia Santiago for criticalreading of this manuscript and Estanislao Escobar-Islas, Ad-rian Trejo-Carmona, Oscar Casas-Mejıa, and Omar Hernan-dez-Garcıa for technical assistance.
Received December 5, 2006. Accepted April 6, 2007.Address all correspondence and requests for reprints to:
Guadalupe Reyes-Cruz, Ph.D., Centro de Investigacion y deEstudios Avanzados-Instituto Politecnico Nacional, Cell Biol-ogy Department, Avenida Instituto Politecnico Nacional2508, Col. San Pedro Zacatenco, 07360 Mexico D.F., Mex-ico. E-mail: [email protected].
This work was supported by Consejo Nacional de Cienciay Technologıa (CONACyT) Mexico Grants 45957 (to G.R.-C.)and 43970-Q (to J.V.-P.), The Academy of Sciences for De-veloping World (TWAS) Grant 03-139 RG/BIO/LA (to G.R.-C.),National Institutes of Health Grant R01TW006664 (to J.V.-P.),and the Fundacion Miguel Aleman. A.P.R.-I., A.G.-R., I.R.-R.,and A.L.E.-S. are graduate students supported by fellow-ships from CONACyT Mexico.
Disclosure Statement: The authors have nothing todisclose.
REFERENCES
1. Brown EM, Gamba G, Riccardi D, Lombardi M, Butters R,Kifor O, Sun A, Hediger MA, Lytton J, Hebert SC 1993Cloning and characterization of an extracellular Ca2�-sensing receptor from bovine parathyroid. Nature 366:575–580
2. Tfelt-Hansen J, Brown EM 2005 The calcium-sensingreceptor in normal physiology and pathophysiology: areview. Crit Rev Clin Lab Sci 42:35–70
3. Hu J, Spiegel AM 2003 Naturally occurring mutations ofthe extracellular Ca2�-sensing receptor: implications forits structure and function. Trends Endocrinol Metab 14:282–288
4. Hendy GN, D’Souza-Li L, Yang B, Canaff L, Cole DE2000 Mutations of the calcium-sensing receptor (CASR)in familial hypocalciuric hypercalcemia, neonatal severehyperparathyroidism, and autosomal dominant hypocal-cemia. Hum Mutat 16:281–296
5. Brown EM, MacLeod RJ 2001 Extracellular calciumsensing and extracellular calcium signaling. Physiol Rev81:239–297
6. Kifor O, Dıaz R, Butters R, Brown EM 1997 The Ca2�-sensing receptor (CaR) activates phospholipases C, A2,and D in bovine parathyroid and CaR-transfected, humanembryonic kidney (HEK293) cells. J Bone Miner Res 12:715–725
7. Chang W, Pratt S, Chen TH, Bourguignon L, Shoback D2001 Amino acids in the cytoplasmic C terminus of theparathyroid Ca2�-sensing receptor mediate efficient cell-surface expression and phospholipase C activation.J Biol Chem 276:44129–44136
8. Chang W, Chen TH, Pratt S, Shoback D 2000 Aminoacids in the second and third intracellular loops of theparathyroid Ca2�-sensing receptor mediate efficientcoupling to phospholipase C. J Biol Chem 275:19955–19963
9. Ward DT 2004 Calcium receptor-mediated intracellularsignalling. Cell Calcium 35:217–228
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1405

10. Herrera-Vigenor F, Hernandez-Garcıa R, Valadez-Sanchez M, Vazquez-Prado J, Reyes-Cruz G 2006AMSH regulates calcium-sensing receptor signalingthrough direct interactions. Biochem Biophys Res Com-mun 347:924–930
11. Zhang M, Breitwieser GE 2005 High affinity interactionwith filamin A protects against calcium-sensing receptordegradation. J Biol Chem 280:11140–11146
12. Pi M, Oakley RH, Gesty-Palmer D, Cruickshank RD,Spurney RF, Luttrell LM, Quarles LD 2005 �-Arrestin-and G protein receptor kinase-mediated calcium-sens-ing receptor desensitization. Mol Endocrinol 19:1078–1087
13. Gama L, Breitwieser GE 1998 A carboxyl-terminal do-main controls the cooperativity for extracellular Ca2�
activation of the human calcium sensing receptor. Astudy with receptor-green fluorescent protein fusions.J Biol Chem 273:29712–29718
14. Chattopadhyay N 2006 Effects of calcium-sensing re-ceptor on the secretion of parathyroid hormone-relatedpeptide and its impact on humoral hypercalcemia ofmalignancy. Am J Physiol Endocrinol Metab 290:E761–E770
15. Ferguson SSG 2001 Evolving concepts in G protein-coupled receptor endocytosis: the role in receptor de-sensitization and signaling. Pharmacol Rev 53:1–24
16. Zastrow V 2003 Mechanisms regulating membrane traf-ficking of G protein-coupled receptors in the endocyticpathway. Life Sci 74:217–224
17. Lefkowitz RJ, Shenoy SK 2005 Transduction of receptorsignals by �-arrestins. Science 308:512–517
18. Miaczynska M, Zerial M 2002 Mosaic organization of theendocytic pathway. Exp Cell Res 272:8–14
19. Seachrist JL, Ferguson S 2003 Regulation of G protein-coupled receptor endocytosis and trafficking by RabGTPases. Life Sci 74:225–235
20. Rosenfeld JL, Knoll BJ, Moore RH 2002 Regulation ofG-protein-coupled receptor activity by rab GTPases. Re-ceptors Channels 8:87–97
21. Ray K, Fan GF, Goldsmith PK, Spiegel AM 1997 Thecarboxyl terminus of the human calcium receptor. Re-quirements for cell-surface expression and signal trans-duction. J Biol Chem 272:31355–31361
22. Bai M, Quinn S, Trivedi S, Kifor O, Pearce S H, Pollak MR,Krapcho K, Hebert SC, Brown EM 1996 Expression andcharacterization of inactivating and activating mutationsin the human Ca2�
o-sensing receptor. J Biol Chem 271:19537–19545
23. Ray K, Clapp P, Goldsmith PK, Spiegel AM 1998 Iden-tification of the sites of N-linked glycosylation on thehuman calcium receptor and assessment of their role incell surface expression and signal transduction. J BiolChem 273:34558–34567
24. Bockaert J, Fagni L, Dumuis A, Marin P 2004 GPCRinteracting proteins (GIP). Pharmacol Ther 103:203–221
25. Simonin F, Karcher P, Boeuf JJ, Matifas A, Kieffer BL2004 Identification of a novel family of G protein-coupledreceptor associated sorting proteins. J Neurochem 89:766–775
26. Whistler JL, Enquist J, Marley A, Fong J, Gladher F,Tsuruda P, Murray SR, Von Zastrow M 2002 Modulationof postendocytic sorting of G protein-coupled receptors.Science 297:615–620
27. Bartlett SE, Enquist J, Hopf FW, Lee JH, Gladher F,Kharazia V, Waldhoer M, Mailliard WS, Armstrong R,Bonci A, Whistler JL 2005 Dopamine responsiveness isregulated by targeted sorting of D2 receptors. Proc NatlAcad Sci USA 102:11521–11526
28. MacLeod RJ, Yano S, Chattopadhyay N, Brown EM 2004Extracellular calcium-sensing receptor transactivates theepidermal growth factor receptor by a triple-membrane-spanning signaling mechanism. Biochem Biophys ResCommun 320:455–460
29. MacLeod RJ, Chattopadhyay N, Brown EM 2003 PTHrPstimulated by the calcium-sensing receptor requiresMAP kinase activation. Am J Physiol Endocrinol Metab284:E435–E442
30. Chen RA, Goodman WG 2004 Role of the calcium-sens-ing receptor in parathyroid gland physiology. Am JPhysiol Renal Physiol 286:F1005–F1011
31. Holstein DM, Berg KA, Leeb-Lundberg LM, Olson MS,Saunders C 2004 Calcium-sensing receptor-mediatedERK1/2 activation requires G�i2 coupling and dynamin-independent receptor internalization. J Biol Chem 279:10060–10069
32. Hjalm G, MacLeod RJ, Kifor O, Chattopadhyay N, BrownEM 2001 Filamin-A binds to the carboxyl-terminal tail ofthe calcium-sensing receptor, an interaction that partic-ipates in CaR-mediated activation of mitogen-activatedprotein kinase. J Biol Chem 276:34880–34887
33. Kifor O, Dıaz R, Butters R, Kifor I, Brown EM 1998 Thecalcium-sensing receptor is localized in caveolin-richplasma membrane domains of bovine parathyroid cells.J Biol Chem 273:21708–21713
34. Thomsen P, Roepstorff K, Stahlhut M, van Deurs B 2002Caveolae are highly immobile plasma membrane mi-crodomains, which are not involved in constitutive endo-cytic trafficking. Mol Biol Cell 13:238–250
35. Yano S, Macleod RJ, Chattopadhyay N, Tfelt-Hansen J,Kifor O, Butters RR, Brown EM 2004 Calcium-sensingreceptor activation stimulates parathyroid hormone-re-lated protein secretion in prostate cancer cells: role ofepidermal growth factor receptor transactivation. Bone35:664–672
36. Hamelin E, Theriault C, Laroche G, Parent JL 2005 Theintracellular trafficking of the G protein-coupled receptorTP� depends on a direct interaction with Rab11. J BiolChem 280:36195–36205
37. Theriault C, Rochdi MD, Parent JL 2004 Role of theRab11-associated intracellular pool of receptorsformed by constitutive endocytosis of the � isoform ofthe thromboxane A2 receptor (TP�). Biochemistry 43:5600–5607
38. Moore RH, Millman EE, Alpizar-Foster E, Dai W, Knoll BJ2004 Rab11 regulates the recycling and lysosome tar-geting of �2-adrenergic receptors. J Cell Sci 117:3107–3117
39. Pula G, Mundell SJ, Roberts PJ, Kelly E 2004 Agonist-independent internalization of metabotropic glutamatereceptor 1a is arrestin-and clathrin-dependent and issuppressed by receptor inverse agonists. J Neurochem89:1009–1020
40. Dale LB, Bhattacharya M, Seachrist JL, Anborgh PH,Ferguson SS 2001 Agonist-stimulated and tonic internal-ization of metabotropic glutamate receptor 1a in humanembryonic kidney 293 cells: agonist-stimulated endocy-tosis is �-arrestin1 isoform-specific. Mol Pharmacol 60:1243–1253
41. Kifor O, Kifor I, Moore Jr FD, Butters Jr RR, Brown EM2003 m-Calpain colocalizes with the calcium-sensing re-ceptor (CaR) in caveolae in parathyroid cells and partic-ipates in degradation of the CaR. J Biol Chem 278:31167–31176
42. Heydorn A, Sondergaard BP, Ersboll B, Holst B, NielsenFC, Haft CR, Whistler J, Schwartz TW 2004 A library of7TM receptor C-terminal tails. Interactions with the pro-posed post-endocytic sorting proteins ERM-bindingphosphoprotein 50 (EBP50), N-ethylmaleimide-sensitivefactor (NSF), sorting nexin 1 (SNX1), and G protein-cou-pled receptor-associated sorting protein (GASP). J BiolChem 279:54291–54303
43. Huang Y, Niwa J, Sobue G, Breitwieser GE 2006 Calci-um-sensing receptor ubiquitination and degradation me-diated by the E3 ubiquitin ligase dorfin. J Biol Chem281:11610–11617
1406 Mol Endocrinol, June 2007, 21(6):1394–1407 Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion

44. Li H, Seth A 2004 An RNF11:Smurf2 complex mediatesubiquitination of the AMSH protein. Oncogene 23:1801–1808
45. Kavalali ET 2006 Synaptic vesicle reuse and its implica-tions. Neuroscientist 12:57–66
46. Sara Y, Virmani T, Deak F, Liu X, Kavalali ET 2005 Anisolated pool of vesicles recycles at rest and drives spon-taneous neurotransmission. Neuron 45:563–573
47. Sun JY, Wu XS, Wu LG 2002 Single and multiple vesiclefusion induce different rates of endocytosis at a centralsynapse. Nature 417:555–559
48. Goldsmith P, Fan GF, Ray K, Shiloach J, McPhie P,Rogers KV, Spiegel AM 1999 Expression, purification,and biochemical characterization of the amino-terminalextracellular domain of the human calcium receptor.J Biol Chem 274:11303–11309
49. McCullough J CM, Urbe S 2004 AMSH is an endosome-associated ubiquitin isopeptidase. J Cell Biol 166:487–492
50. Whistler JL, Tsao P, von Zastrow M 2001 A phosphory-lation-regulated brake mechanism controls the initial en-docytosis of opioid receptors but is not required forpost-endocytic sorting to lysosomes. J Biol Chem 276:34331–34338
51. Kim KA, von Zastrow M 2003 Neurotrophin-regulatedsorting of opioid receptors in the biosyntheticpathway of neurosecretory cells. J Neurosci 23:2075–2085
52. Vickery RG, von Zastrow M 1999 Distinct dynamin-dependent and -independent mechanisms target structur-ally homologous dopamine receptors to different endocyticmembranes. J Cell Biol 144:31–43
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
Reyes-Ibarra et al. • CaR Endocytosis Modulates PTHrP Secretion Mol Endocrinol, June 2007, 21(6):1394–1407 1407
Copyright © 2022 FDOKUMEN




















