
Stretch-regulated Exocytosis/Endocytosis in Bladder Umbrella Cells
Upload
independentCategory
view
1download
0
original article© The American Society of Gene & Cell Therapy
Molecular Therapy vol. 17 no. 8, 1411–1417 aug. 2009 1411
The understanding of internalization pathways of lipo- or polyplexes is crucial for engineering successful reagents for nonviral gene transfection. A known inhibitor of fluid phase endocytosis (FPE), rottlerin, was used to quantify the contribution of this pathway by flow cytometric and fluorescence assays. Rottlerin was shown to be a specific inhibitor of transfection by polyethylene imine (PEI-25)/DNA complexes, leading to a decrease in the amount of transfected HeLa and CHO-K1 cells and a decrease in the expression of enhanced green fluorescent protein (EGFP) reporter gene by up to 50%. Experiments using fluorescently labeled polyplexes result in a decrease of uptake by up to 40%. Additionally, rottlerin does not cross-inhibit clathrin- and caveolin-mediated endo cytotic pathways of internalization, consistent with direct uptake inhibition by rottlerin. Nonspecific effects as a result of toxicity were ruled out by control experiments at concentrations where rottlerin inhibition was specific. These findings suggest that for CHO-K1 and HeLa cells, internalization of PEI-25/DNA complexes by FPE plays a decisive role in gene transfection. The establishment of an additional pathway that is independent of clathrin- and caveolin-mediated endocytotic uptake may have an impact on the design of future reagents of nonviral gene therapy and investigations of the uptake pathways and intracellular trafficking involved.
Received 14 August 2008; accepted 4 May 2009; published online 16 June 2009. doi:10.1038/mt.2009.121
IntroductIonThe ability to manipulate mammalian cells by altering their genetic makeup is crucial for the treatment of genetic diseases, such as cancer or cystic fibrosis.1 It is also relevant for applica-tions in biotechnology and cell biology that require expression of a desired protein.2–4
Currently, viral gene transfer remains the most efficient method to transport DNA into cells and nuclei,5 but due to dis-advantages such as toxicity, possible immune responses against viral coat proteins, and residual infectivity of carrier viruses, the development of nonviral vectors is desired.6 Such nonviral vectors include nanoparticles,7,8 synthetic transfection reagents,9–12 cationic lipids, including the commercial Lipofectamine 2000 (refs. 3,4), or
polymers, such as poly-l-lysine,13 polyamidoamine,14 poly(β-amino esters),15 and polyethylene imine (PEI).2,16,17
However, nonviral gene vectors are currently limited by poor transfection efficiency as a consequence of several cellular barriers18 between the extracellular environment and the nucleus. The uptake mechanisms of complexes, solutes, and particles are still not well understood. Uptake by endocytosis includes a number of distinctly different uptake routes. Pinocytosis is responsible for the uptake of macromolecules and encompasses a variety of routes that are followed as a function of the cell-type and the size of the cargo. These include macropinocytosis (for molecules up to a diameter >1 µm), clathrin-mediated endo-cytosis (≤120 nm), and caveolin- mediated endocytosis (≤90 nm) (ref. 19). In all these processes, complexes of DNA and cationic polymers, known as polyplexes, are transferred into intracellu-lar organelles such as endosomes or caveosomes,20 before their release into the cytosol and further intracellular trafficking to the nucleus.21,22
We aim to resolve the processes involved in import of the widely used transfection reagent of branched PEI (Mw 25 kd; PEI-25). PEI-25 and DNA form polyplexes with sizes ranging from 150 nm (refs. 23,24) to >1,000 nm (refs. 25–27). Involvement of clathrin- and caveolin-mediated endocytosis in the uptake of PEI polyplexes has been shown,21,28,29 but does not explain uptake of particles bigger than the upper limit described for these path-ways (150–200 nm) (ref. 19).
In this work, a pathway distinct from clathrin- and caveolin-mediated endocytosis that contributes to the uptake of PEI-25/DNA polyplexes in CHO-K1 and HeLa cells was identified by rot-tlerin inhibition. This selective inhibitor of the fluid phase endo-cytosis (FPE),30 interacts with protein kinase C31 to inhibit FPE. FPE is considered a mannose receptor–independent and unregu-lated macropinocytotic pathway.30,32,33 It was found that rottlerin decreases both the amount of cells expressing enhanced green fluorescent protein (EGFP) and the amount of expressed EGFP after transfection with PEI-25/DNA polyplexes by up to 50%, as a consequence of obtained decreased uptake of polyplexes. Our findings suggest that FPE contributes substantially to PEI-25-mediated transfection processes in CHO-K1 and HeLa cells without interfering with clathrin- and caveolin-mediated path-ways. We also conclude that FPE is independent of clathrin- and caveolin-mediated endocytotic uptake pathways regardless of the exact size cutoffs of these endocytotic pathways.
Correspondence: Florian Hollfelder, Department of Biochemistry, University of Cambridge, 80 Tennis Court Road, Cambridge CB2 1GA, UK. E-mail: [email protected]
Fluid Phase Endocytosis Contributes to Transfection of DNA by PEI-25Hansjörg Hufnagel1, Parvez Hakim1, Aline Lima1 and Florian Hollfelder1
1Department of Biochemistry, University of Cambridge, Cambridge, UK
1412 www.moleculartherapy.org vol. 17 no. 8 aug. 2009
© The American Society of Gene & Cell TherapyRottlerin Inhibits Transfection by PEI-25/DNA
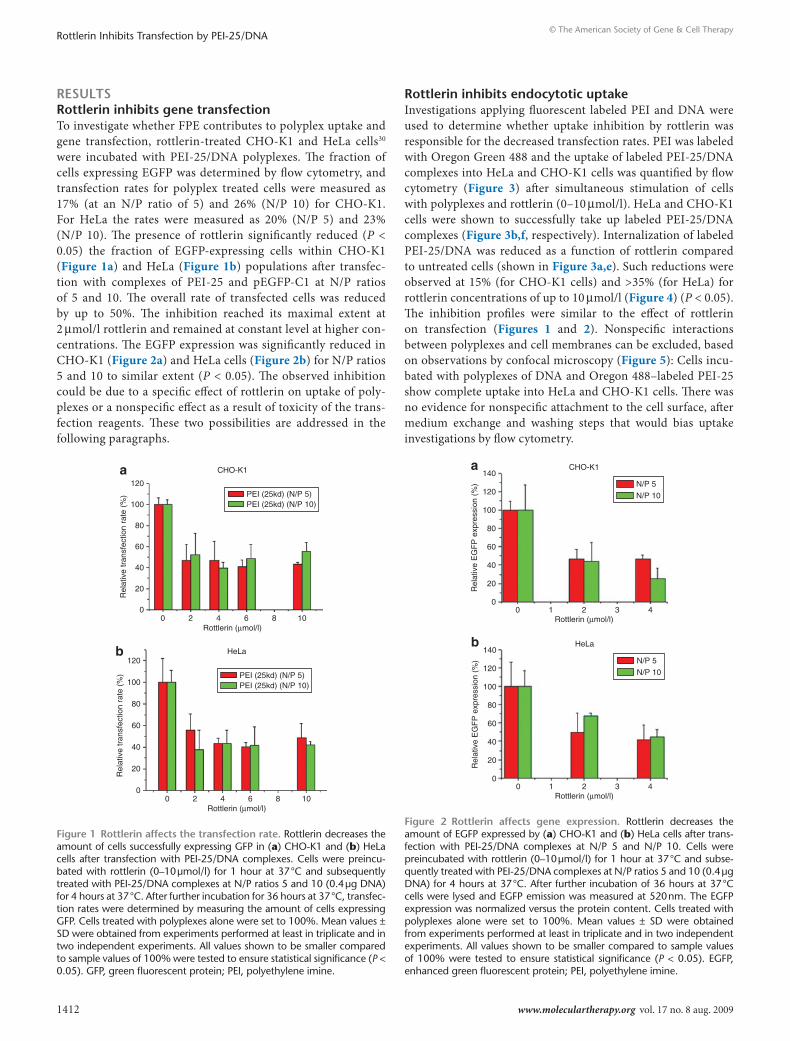
resultsrottlerin inhibits gene transfectionTo investigate whether FPE contributes to polyplex uptake and gene transfection, rottlerin-treated CHO-K1 and HeLa cells30 were incubated with PEI-25/DNA polyplexes. The fraction of cells expressing EGFP was determined by flow cytometry, and transfection rates for polyplex treated cells were measured as 17% (at an N/P ratio of 5) and 26% (N/P 10) for CHO-K1. For HeLa the rates were measured as 20% (N/P 5) and 23% (N/P 10). The presence of rottlerin significantly reduced (P < 0.05) the fraction of EGFP-expressing cells within CHO-K1 (Figure 1a) and HeLa (Figure 1b) populations after transfec-tion with complexes of PEI-25 and pEGFP-C1 at N/P ratios of 5 and 10. The overall rate of transfected cells was reduced by up to 50%. The inhibition reached its maximal extent at 2 μmol/l rottlerin and remained at constant level at higher con-centrations. The EGFP expression was significantly reduced in CHO-K1 (Figure 2a) and HeLa cells (Figure 2b) for N/P ratios 5 and 10 to similar extent (P < 0.05). The observed inhibition could be due to a specific effect of rottlerin on uptake of poly-plexes or a nonspecific effect as a result of toxicity of the trans-fection reagents. These two possibilities are addressed in the following paragraphs.
rottlerin inhibits endocytotic uptakeInvestigations applying fluorescent labeled PEI and DNA were used to determine whether uptake inhibition by rottlerin was responsible for the decreased transfection rates. PEI was labeled with Oregon Green 488 and the uptake of labeled PEI-25/DNA complexes into HeLa and CHO-K1 cells was quantified by flow cytometry (Figure 3) after simultaneous stimulation of cells with polyplexes and rottlerin (0–10 μmol/l). HeLa and CHO-K1 cells were shown to successfully take up labeled PEI-25/DNA complexes (Figure 3b,f, respectively). Internalization of labeled PEI-25/DNA was reduced as a function of rottlerin compared to untreated cells (shown in Figure 3a,e). Such reductions were observed at 15% (for CHO-K1 cells) and >35% (for HeLa) for rottlerin concentrations of up to 10 μmol/l (Figure 4) (P < 0.05). The inhibition profiles were similar to the effect of rottlerin on transfection (Figures 1 and 2). Nonspecific interactions between polyplexes and cell membranes can be excluded, based on observations by confocal microscopy (Figure 5): Cells incu-bated with polyplexes of DNA and Oregon 488–labeled PEI-25 show complete uptake into HeLa and CHO-K1 cells. There was no evidence for nonspecific attachment to the cell surface, after medium exchange and washing steps that would bias uptake investigations by flow cytometry.
00 2 4 6
Rottlerin (µmol/l)8 10
20
40
60
80
100
CHO-K1
Rel
ativ
e tr
ansf
ectio
n ra
te (
%)
120PEI (25kd) (N/P 5)PEI (25kd) (N/P 10)
a
PEI (25kd) (N/P 5)PEI (25kd) (N/P 10)
00 2 4 6
Rottlerin (µmol/l)
HeLa
8 10
20
40
60
80
100
Rel
ativ
e tr
ansf
ectio
n ra
te (
%)
120b
Figure 1 rottlerin affects the transfection rate. Rottlerin decreases the amount of cells successfully expressing GFP in (a) CHO-K1 and (b) HeLa cells after transfection with PEI-25/DNA complexes. Cells were preincu-bated with rottlerin (0–10 μmol/l) for 1 hour at 37 °C and subsequently treated with PEI-25/DNA complexes at N/P ratios 5 and 10 (0.4 μg DNA) for 4 hours at 37 °C. After further incubation for 36 hours at 37 °C, transfec-tion rates were determined by measuring the amount of cells expressing GFP. Cells treated with polyplexes alone were set to 100%. Mean values ± SD were obtained from experiments performed at least in triplicate and in two independent experiments. All values shown to be smaller compared to sample values of 100% were tested to ensure statistical significance (P < 0.05). GFP, green fluorescent protein; PEI, polyethylene imine.
00 1 2
Rottlerin (µmol/l)
CHO-K1a
3 4
20
40
60
80
100
120
Rel
ativ
e E
GF
P e
xpre
ssio
n (%
)140
N/P 5
N/P 10
00 1 2
Rottlerin (µmol/l)
HeLab
3 4
20
40
60
80
100
120
Rel
ativ
e E
GF
P e
xpre
ssio
n (%
)
140N/P 5
N/P 10
Figure 2 rottlerin affects gene expression. Rottlerin decreases the amount of EGFP expressed by (a) CHO-K1 and (b) HeLa cells after trans-fection with PEI-25/DNA complexes at N/P 5 and N/P 10. Cells were preincubated with rottlerin (0–10 μmol/l) for 1 hour at 37 °C and subse-quently treated with PEI-25/DNA complexes at N/P ratios 5 and 10 (0.4 μg DNA) for 4 hours at 37 °C. After further incubation of 36 hours at 37 °C cells were lysed and EGFP emission was measured at 520 nm. The EGFP expression was normalized versus the protein content. Cells treated with polyplexes alone were set to 100%. Mean values ± SD were obtained from experiments performed at least in triplicate and in two independent experiments. All values shown to be smaller compared to sample values of 100% were tested to ensure statistical significance (P < 0.05). EGFP, enhanced green fluorescent protein; PEI, polyethylene imine.

Molecular Therapy vol. 17 no. 8 aug. 2009 1413
© The American Society of Gene & Cell TherapyRottlerin Inhibits Transfection by PEI-25/DNA
After the application of rottlerin (2 and 4 μmol/l), polyplexes using fluorescently labeled DNA complexed to PEI-25 show a significant reduction of uptake by 40% in CHO-K1 (Figure 6a) and HeLa cells (Figure 6b) at N/P ratios 5 and 10, respectively (P < 0.05).
These observations contrast with the analogous experiments for transferrin (Figure 7a) and cholera toxin B subunit (CTB) (Figure 7b), which were labeled with fluorescein isothiocya-nate (FITC). These compounds are known to be taken up by clathrin- and caveolin-mediated endocytosis, respectively. Their uptake was analyzed by flow cytometry using medians of the distributions of populations of cells treated with fluorescent
markers. No inhibitory effect for up to 6 μmol/l rottlerin was observed with FITC-transferrin (Figure 7a) and FITC-CTB (Figure 7b).
Given that CTB has been shown to be internalized by caveolin-mediated endocytosis and transferrin by clathrin-mediated endocytosis, the lack of inhibition of the uptake of FITC-CTB and FITC-transferrin, respectively, by rottlerin at concentrations where inhibition of transfection was at a maximum (≥2 µmol/l), provides evidence that PEI-25 poly-plexes are not only taken up by these pathways, but instead internalized by an additional route. Inhibition of the uptake of CTB or transferrin would be expected, if rottlerin cross- inhibited these pathways.
In case of HeLa cells, increased slight marker uptake was observed while treatment of CHO-K1 cells with rottlerin resulted in enhanced internalization by even up to 20% of FITC-transferrin.
cell viabilityTo rule out that the treatment of cells with inhibitors influences the transfection assays, we investigated the viability of HeLa and CHO-K1 cells after treatment with inhibitors and polyplexes (Figure 8).
Cell viability of CHO-K1 cells prepared with PEI-25/DNA polyplexes decreased with increasing rottlerin concentration, but still remains at about 80–90% for concentrations up to 4 µmol/l and ~70% at 10 µmol/l rottlerin. HeLa cells were prone cytotoxic-ity to a slightly greater extent. Nevertheless, ~70% still survive at a rottlerin concentration of 4 μmol/l. For both cell lines, the viability was decreasing constantly with increasing amounts of rottlerin, in contrast to the inhibition profile of uptake and transfection by rottlerin. These findings suggest that cytotoxicity can be ruled out as an explanation of the effects of rottlerin.
100 101 102
FITC log
0
57
114
171Cells only
Cou
nts
228
103 104100 101 102
FITC log
0
35
71
106Polyplex
Cou
nts
142
103 104
a
100 101 102
FITC log
0
23
47
70CTB
Cou
nts
94
103 104
c
100 101 102
FITC log
0
67
135
203Cells only
Cou
nts
271
103 104
e
100 101 102
FITC log
0
25
51
76Polyplex
Cou
nts
102
103 104
f
100 101 102
FITC log
0
36
72
108Transferrin
HeLa
CH
O-K
1
Cou
nts
144
103 104
d
b
Figure 3 Flow cytometry of markers of uptake. (a–d) HeLa and (e–f) CHO-K1 cells take up markers of endocytosis and PEI/DNA polyplexes. a and e show the intrinsic fluorescence of a cell population. Fluorescence is enhanced by uptake of Oregon Green 488–labeled PEI-25/DNA com-plexes (0.4 μg DNA, N/P 5) (b and f). For comparison, cells were also incubated with fluorescently labeled molecules that are markers for endocytotic pathways (c: FITC-cholera toxin B subunit 1 μg/ml; d: FITC-transferrin, 0.5 mg/ml). Cells were incubated with markers of endocytosis for 4 hours at 37 °C. CTB, cholera toxin B subunit; FITC, fluorescein isothiocyanate.
40−1 0 1 2 3 4 5
Rottlerin (µmol/l)6 7 8 9 10 11
60
80
100
Rel
ativ
e up
take
of P
EI-
25 (
%)
HelaCHO-K1
Figure 4 rottlerin reduces the internalization of PeI-25/dnA in Hela and cHo-K1 cells. Cells were preincubated with rottlerin (0–10 μmol/l) for 1 hour at 37 °C and subsequently treated with Oregon 488–labeled PEI-25/DNA complexes (0.4 μg DNA, N/P ratio: 5) for 4 hours at 37 °C before measuring uptake using flow cytometry. Values for cells treated with polyplexes alone were set as 100%. Mean values ± SD were obtained from experiments performed at least in triplicate and in two independent experiments. All values shown to be smaller compared to sample values of 100% were tested to ensure statistical significance (P < 0.05).

1414 www.moleculartherapy.org vol. 17 no. 8 aug. 2009
© The American Society of Gene & Cell TherapyRottlerin Inhibits Transfection by PEI-25/DNA
dIscussIonIn earlier studies, inhibition of transfection of mammalian cells by the macropinocytosis inhibitor wortmannin had failed21,28 and involvement of macropinocytotic uptake of PEI/DNA poly-plexes had been ruled out. Furthermore, complete abolishment of polyplex uptake by inhibitors of caveolae-mediated endocytosis, genistein, and fillipin III, was seen in A549 cells.21,29 These find-ings emphasized caveolae-mediated endocytosis of PEI/DNA complexes at the expense of a macropinocytotic pathway,27 even for particles with a diameter as large as 1,000 nm.
Despite the lack of precise size limits of clathrin- and caveolin-mediated endocytosis in the literature (from 120 up to 500 nm) (refs. 19,22), a large fraction of PEI-25/DNA polyplexes have a diameter exceeding the upper limit allowed for uptake by these routes.26
In our study, inhibition of transfection by rottlerin was detect-able from concentrations of 2 µmol/l, where an inhibition of EGFP expression and decrease of transfected cells of up to 50% was reached. This corresponds well to an IC50 of 3 µmol/l previously measured for rottlerin inhibition of protein kinase C.32 Cell viability was only significantly lowered from 4 µmol/l onwards (Figure 8), suggesting that genuine rottlerin inhibition is monitored.
The observation of rottlerin inhibiting uptake (Figures 4–6) and transfection (Figures 1 and 2) leads to an extension of this scenario for the relatively large complexes of PEI-25/DNA (>150 nm diameter).25,26 These polyplexes are taken up by a path-way that is not sensitive to wortmannin and therefore independent
of a mannose receptor–mediated mechanism.30 Our data lead to the conclusion that polyplexes of branched PEI-25 and DNA use an unregulated and unselective macropinocytotic pathway for successful transfection of mammalian cells. Bigger particles appear to resort to this rottlerin-sensitive pathway and the remain-ing level of transfection in the presence of rottlerin may represent transfection by polyplexes of sizes smaller than the actual cutoff, due to the polydispersity of PEI-25.
Our results are consistent with microscopic studies in which PEI-25/DNA complexes were observed to colocalize with macropinosomes.34
Nonspecific inhibition as a cause for reduced transfection can be safely excluded, as rottlerin does not decrease uptake of markers of clathrin- or caveolin-mediated endocytosis in CHO-K1 and HeLa cells. The slight increase in observed uptake for these markers (by 20%) in the presence of rottlerin may be caused by upregulation of other pathways than FPE, in this case clathrin- and caveolae-mediated endocytosis. Such overcompensation after inhibition of specific pathways by alternative routes of endocytosis has been described previously.35
We conclude that transfection is decreased to a larger extent (>50%) than uptake of polyplexes by rottlerin, as the transfection is more likely to happen by polyplexes taken up by this particular
Untreated
Rottlerin
N/P
5N
/P 10
CH
O-K
1
N/P
5N
/P 10
0 µmol/l
30 µm
2 µmol/l 4 µmol/l
HeLa
30 µm 30 µm 30 µm
Figure 5 cHo-K1 and Hela cells show complete uptake of PeI-25/dnA polyplexes. CHO-K1 and HeLa cells were seeded into 8-well chamber slides and preincubated with rottlerin (0–4 μmol/l) for 1 hours at 37 °C and subsequently treated with Oregon 488–labeled PEI-25/DNA polyplexes (0.4 μg DNA, N/P ratios 5 and 10) for 4 hours at 37 °C. Despite extensive washing (three times with PBS) cells showed intra-cellular distribution of fluorescence, ruling out nonspecific attachment of polyplexes to cell membranes after application of rottlerin. For visu-alization chamber slides were mounted onto the stage of a Leica 6000 CS inverted microscope. Images were acquired after excitation with a 488 nm argon laser by monitoring emission at 520 nm.
0−1 0 1 2
Rottlerin (µmol/l)
CHO-K1a
3 4 5
20
40
60
80
100
Rel
ativ
e up
take
of D
NA
(%
)
120 N/P 5N/P 10
0−1 0 1 2
Rottlerin (µmol/l)
HeLab
3 4 5
20
40
60
80
100
Rel
ativ
e up
take
of D
NA
(%
)
120 N/P 5N/P 10
Figure 6 rottlerin affects dnA uptake. Application of rottlerin reduces the internalization of fluorescently labeled DNA in PEI-25/DNA in (a) CHO-K1 and (b) HeLa cells. Cells were preincubated with rottlerin (0–4 μmol/l) for 1 hour at 37 °C and subsequently treated with PEI-25/DNA complexes (for 4 hours at 37 °C; 0.4 μg DNA, N/P ratio: 5 and 10) in which the DNA was fluorescently labeled. Cells were lysed and fluorescence was measured at 420 nm. The fluorescence was normal-ized against protein content in wells. Fluorescence values of cells treated with polyplexes alone were set as 100%. Mean values ± SD were obtained from experiments performed at least in triplicate and in two independent experiments. All values shown to be smaller compared to sample values of 100% were tested to ensure statistical significance (P < 0.05).

Molecular Therapy vol. 17 no. 8 aug. 2009 1415
© The American Society of Gene & Cell TherapyRottlerin Inhibits Transfection by PEI-25/DNA
pathway. This suggests that macropinosomes have a higher propensity to deliver PEI-25/DNA cargo than endosomes.36 This observation is consistent with the availability of FPE as an efficient alternative pathway for transfection by larger particles. Ogris et al. reported an imbalance between transfection and uptake12 and noted that larger aggregates of >500 nm diameter transfect more efficiently than small polyplexes. An uptake pathway using macropinosomes is also consistent with earlier work on involvement of processes that do not lead directly to an endosome, but instead use the detour of caveosomes.21,28 This more complex scenario for internalization of PEI polyplexes is similar to observations for lipoplexes.37
Taken together, our data suggest that multiple pathways of intracellular trafficking—including macropinocytosis—should be considered in the analysis of DNA transfection reagents and may explain why consistent structure–activity relationships for transfec-tion reagents have been difficult to obtain.3 It is possible that in some respects the entry pathways have similar features. For example, osmolytic rupture due to pH changes in endosomes16,38 is considered an important part of the route of DNA polyplexes into the nucleus. The proton sponge character of PEI has been frequently used to explain the efficiency of PEI in transfection by endosomal buffer-ing.16 However, pH-buffering by PEI may not only be beneficial for DNA release from the endosome, but also from related compart-ments, such as macropinosomes arising from alternative uptake routes. Although some characteristics of the alternative pathways
may coincide, others will differ and can be exploited by new transfection reagents. For example, the tolerance of FPE may allow a wider variety of suitably sized compounds to penetrate the cell and variation of polyplex diameter may allow channeling of polyplexes into specific pathways. Further definition of the limits of alterna-tive pathways will clarify this picture, and whether this mechanism of uptake is cell specific. However, we expect additional degrees of freedom with larger particles (>500 nm), as they are at least partly taken up by FPE. In the current absence of a monocausal explana-tion for possible structural improvements of transfection reagents, a combinatorial approach (for example by PEI modification17,39) may be better suited to take advantage of multiple entry pathways15,37 to optimize the efficiency of gene transfer.
MAterIAls And MetHodsAll reagents were purchased from Sigma (Gilingham, UK) unless otherwise stated. pEGFP-C1 encoding EGFP under the control of a human cytomegalovirus promoter was used as an reporter gene (Plasmid Factory, Bielefeld, Germany).
Cell lines, media, and reagents. CHO-K1 cells were grown in F12-Ham medium (Invitrogen, Paisley, UK) supplemented with penicillin (100 units/ml), streptomycin (100 μg/ml), and 10% heat inactivated fetal bovine serum (Invitrogen). HeLa cells were grown in RPMI 1640 medium (Invitrogen) supplemented with penicillin (100 units/ml), streptomycin (100 μg/ml), and 10% heat inactivated fetal bovine serum (Invitrogen). Cells were maintained at 37 °C in 5% CO2 humidified air atmosphere and passaged every other day.
PEI labeling with Oregon Green 488. PEI-25 was labeled according to Beber et al.40 with Oregon Green 488 (Invitrogen). The solution of fluores-cent product was stored at 4 °C in the dark.
DNA labeling with Mirus labeling kit. The pEGFP-C1 plasmid was labeled with a Mirus fluorescein nucleic acid labeling kit (Cambridge Biosciences, Cambridge, UK) according to the manufacturer’s instructions.
Uptake studies with Oregon 488–labeled polyplexes, labeled DNA, and markers of endocytosis. For uptake studies, cells were seeded 12 hours before the experiment in medium (200 μl) containing 10% fetal
60
−1 0 1 2 3 4 5Rottlerin (µmol/l)
6 7 8 9 10 11
80
100
120
140
160
FIT
C-t
rans
ferr
in u
ptak
e (%
)180
HeLaCHO-K1
a
60
0
20
40
−1 0 1 2 3 4 5Rottlerin (µmol/l)
6 7 8 9 10 11
80
100
120
140
160
FIT
C-C
TB
upt
ake
(%)
200
180HeLaCHO-K1
b
Figure 7 Impact of rottlerin on uptake of markers of endocytosis. Rottlerin does not inhibit the uptake of transferrin and cholera toxin subunit B (CTB) as marker of (a) clathrin- and (b) caveolin-mediated endocytosis, respectively, in HeLa and CHO-K1 cells up to a concentration of 6 μmol/l. Cells were preincubated with rottlerin (0–10 μmol/l) for 1 hour at 37 °C and subsequently treated with FITC-cholera-toxin subunit B (1.0 mg/ml) and FITC-transferrin (0.5 mg/ ml) for 4 hours at 37 °C. Values of cells treated with markers alone were set to 100%. Mean values ± SD were obtained from experiments performed in triplicate and two independent experi-ments. CTB, cholera toxin B subunit; FITC, fluorescein isothiocyanate.
0−1 0 1 2 3 4 5
Rottlerin (µmol/l)6 7 8 9 10 11
20
40
60
80
100
Cel
l via
bilit
y (%
)
120 HeLaCHO-K1
Figure 8 cell viability of cHo-K1 and Hela cells after treatment with rottlerin and PeI/dnA. Cells were preincubated with rottlerin (0–10 μmol/l) for 1 hour at 37 °C and subsequently treated with PEI-25/DNA complexes (0.4 μg DNA, N/P ratio: 5) for 4 hours at 37 °C. After further incubation (36 hours at 37 °C) cells were washed and their viabil-ity measured using crystal violet staining. Values of cell viability of cells neither treated with polyplexes nor inhibitor were set to 100%. Mean values ± SD were obtained from experiments performed in triplicate.

1416 www.moleculartherapy.org vol. 17 no. 8 aug. 2009
© The American Society of Gene & Cell TherapyRottlerin Inhibits Transfection by PEI-25/DNA
bovine serum. Cells were preincubated with inhibitors for 1 hour at 37 °C before application of polyplexes for transfection or markers of endo-cytosis. Fluorescently labeled polyplexes for uptake studies were prepared in OptiMem (Invitrogen) solutions with 0.4 µg pEGFP-C1 per well at an N/P ratio of 5 for PEI-25. After 4 hours of incubation cells were washed three times with phosphate-buffered saline (PBS) (pH 7.4, 200 µl). Cells were harvested by treatment with trypsin (0.5 mg/ml)/EDTA (0.2 mg/ml) solution and uptake of fluorescent markers was assessed by flow cytometry using a Cyan ADP-MLE cytometer with Summit Software 4.3. (Dako, Ely, UK). The median fluorescence intensity data for a minimum of 5,000 cells were recorded.
For uptake studies with labeled DNA, cells were seeded 12 hours before the experiment in medium (200 μl) containing 10% fetal bovine serum. Cells were preincubated with inhibitors for 1 hour at 37 °C before application of polyplexes for transfection. Polyplexes for uptake studies were prepared in OptiMem solutions with fluorescently labeled pEGFP-C1 (0.4 µg/well) at N/P ratios of 5 and 10 for PEI-25. After 4 hours of incubation, cells were washed three times with PBS (pH 7.4, 200 µl) and cells were lysed with GLO-Lysis Buffer (Promega, Madison, WI) (200 μl) for 10 minutes before fluorescence was read in 96-well Spectramax M5 fluorescence reader (Wokingham, UK) at 520 nm. To normalize EGFP expression, the amount of protein in the lysate was measured. Lysis solution (70 μl) was incubated for 30 minutes at 37 °C with the Pierce BCA-Protein Assay Kit (Rockford, IL) (200 μl) before absorption was measured at 562 nm and protein content determined according to a standard curve.
Transfection studies and gene expression studies. For transfection stud-ies cells polyplexes were prepared in OptiMem solutions with 0.4 µg DNA (pEGFP-C1) per well at N/P ratios 5 and 10. After 4 hours of incubation at 37 °C, medium was changed and cells were washed three times with medium, and incubated for another 36 hours. For determination of the fraction of cells that were successfully transfected and expressing EGFP, cells were washed three times (PBS, pH 7.4, 200 μl) before being treated with a trypsin/EDTA solution in PBS (0.5 mg/ml trypsin, 0.2 mg/ml EDTA; 150 μl) to harvest cells. The fraction of GFP-expressing cells was determined using flow cytometry.
To determine the amount of EGFP expression, cells were washed with PBS (pH 7.4, 200 µl) three times 36 hours after transfection and incubated with GLO-Lysis Buffer (Promega) (200 μl) for 10 minutes before fluorescence was read in a 96-well Spectramax M5 fluorescence reader at 520 nm. To normalize EGFP expression, the amount of protein in the lysate was determined. Lysis solution (70 μl) was incubated for 30 minutes at 37 °C with the Pierce BCA-Protein Assay Kit (200 μl) before absorption was measured at 562 nm and protein content determined according to a standard curve.
Microscopic studies with CHO-K1 and HeLa. CHO-K1 and HeLa cells were seeded 18 hours before start of the experiment into 8-well chamber slides (Labtek, Napperville, IL) at 20,000 cells per well. Cells were preincubated with rottlerin (0–4 μmol/l) for 1 hour at 37 °C, subsequently treated with Oregon 488–labeled PEI-25/DNA complexes (0.4 μg DNA, N/P ratio: 5 and 10) for 4 hours at 37 °C, and washed with PBS (pH 7.4, 200 µl) three times. Chambers with cells were mounted onto the stage of a Leica TCS SP5 confo-cal scanning laser microscopy system using a ×63 water immersion objective lens (numerical aperture 1.2; confocal aperture 600 μm) on a Leica 6000 CS inverted microscope (Leica, Heidelberg, Germany). Images of fluorescent samples after excitation at a 488 nm by using an argon laser and the emission at 520 nm were acquired and analyzed using Leica AF software (Leica).
Cell viability assay. Cells were treated as described above omitting trypsin treatment. Dead cells were discarded with the supernatant followed by washing of plates with PBS and incubated with crystal violet solution (50 µl; Sigma-Aldrich, Gillingham, UK) for 15 minutes. Stained cells were
subsequently washed with PBS for three times (200 µl) and solution was dissolved in MeOH (250 µl per well). Measured absorption correlates to the amount of living cells and was read at 550 nm using a Molecular Devices Spectramax M5 photometer.
AcKnowledgMentsH.H. acknowledges the support of Stiftung der Deutschen Wirtschaft (Foundation of German Business), the EPRSC, and the Cambridge Trusts. A.L. thanks CAPES for a short-term fellowship. We thank Nigel Miller for technical support and Stefanie Jonas, Tony Kirby, Viktor Stein, and Tony Jackson for critical reading. F.H. is an ERC Starting Investigator.
reFerences1. Edelstein, ML, Abedi, MR and Wixon, J (2007). Gene therapy clinical trials worldwide
to 2007--an update. J Gene Med 9: 833–842.2. Neu, M, Fischer, D and Kissel, T (2005). Recent advances in rational gene transfer
vector design based on poly(ethylene imine) and its derivatives. J Gene Med 7: 992–1009.
3. Kirby, AJ, Camilleri, P, Engberts, JB, Feiters, MC, Nolte, RJ, Söderman, O et al. (2003). Gemini surfactants: new synthetic vectors for gene transfection. Angew Chem Int Ed Engl 42: 1448–1457.
4. Scarzello, M, Smisterová, J, Wagenaar, A, Stuart, MC, Hoekstra, D, Engberts, JB et al. (2005). Sunfish cationic amphiphiles: toward an adaptative lipoplex morphology. J Am Chem Soc 127: 10420–10429.
5. Verma, IM and Weitzman, MD (2005). Gene therapy: twenty-first century medicine. Annu Rev Biochem 74: 711–738.
6. Mastrobattista, E, van der Aa, MA, Hennink, WE and Crommelin, DJ (2006). Artificial viruses: a nanotechnological approach to gene delivery. Nat Rev Drug Discov 5: 115–121.
7. Chittimalla, C, Zammut-Italiano, L, Zuber, G and Behr, JP (2005). Monomolecular DNA nanoparticles for intravenous delivery of genes. J Am Chem Soc 127: 11436–11441.
8. Na, K, Kim, S, Sun, BK, Woo, DG, Chung, HM and Park, KH (2007). Blended construct consisting of thermo-reversible hydrogels and heparinized nanoparticles for increasing the proliferation activity of the rabbit chondrocyte in vivo test. Biotechnol Lett 29: 1447–1452.
9. Pack, DW, Hoffman, AS, Pun, S and Stayton, PS (2005). Design and development of polymers for gene delivery. Nat Rev Drug Discov 4: 581–593.
10. Davis, ME (2002). Non-viral gene delivery systems. Curr Opin Biotechnol 13: 128–131.11. Wolff, JA and Rozema, DB (2008). Breaking the bonds: non-viral vectors become
chemically dynamic. Mol Ther 16: 8–15.12. Ogris, M, Steinlein, P, Carotta, S, Brunner, S and Wagner, E (2001). DNA/
polyethylenimine transfection particles: influence of ligands, polymer size, and PEGylation on internalization and gene expression. AAPS PharmSci 3: E21.
13. Pouton, CW, Lucas, P, Thomas, BJ, Uduehi, AN, Milroy, DA and Moss, SH (1998). Polycation-DNA complexes for gene delivery: a comparison of the biopharmaceutical properties of cationic polypeptides and cationic lipids. J Control Release 53: 289–299.
14. Radu, DR, Lai, CY, Jeftinija, K, Rowe, EW, Jeftinija, S and Lin, VS (2004). A polyamidoamine dendrimer-capped mesoporous silica nanosphere-based gene transfection reagent. J Am Chem Soc 126: 13216–13217.
15. Green, JJ, Langer, R and Anderson, DG (2008). A combinatorial polymer library approach yields insight into nonviral gene delivery. Acc Chem 41: 749–759.
16. Boussif, O, Lezoualc’h, F, Zanta, MA, Mergny, MD, Scherman, D, Demeneix, B et al. (1995). A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: polyethylenimine. Proc Natl Acad Sci USA 92: 7297–7301.
17. Van Vliet, LD, Chapman, MR, Avenier, F, Kitson, CZ and Hollfelder, F (2008). Relating chemical and biological diversity space: a tunable system for efficient gene transfection. Chembiochem 9: 1960–1967.
18. Wiethoff, CM and Middaugh, CR (2003). Barriers to nonviral gene delivery. J Pharm Sci 92: 203–217.
19. Conner, SD and Schmid, SL (2003). Regulated portals of entry into the cell. Nature 422: 37–44.
20. Nichols, B (2003). Caveosomes and endocytosis of lipid rafts. J Cell Sci 116(Pt 23): 4707–4714.
21. Rejman, J, Bragonzi, A and Conese, M (2005). Role of clathrin- and caveolae-mediated endocytosis in gene transfer mediated by lipo- and polyplexes. Mol Ther 12: 468–474.
22. Rejman, J, Oberle, V, Zuhorn, IS and Hoekstra, D (2004). Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem J 377(Pt 1): 159–169.
23. Neu, M, Sitterberg, J, Bakowsky, U and Kissel, T (2006). Stabilized nanocarriers for plasmids based upon cross-linked poly(ethylene imine). Biomacromolecules 7: 3428–3438.
24. Mao, S, Neu, M, Germershaus, O, Merkel, O, Sitterberg, J, Bakowsky, U et al. (2006). Influence of polyethylene glycol chain length on the physicochemical and biological properties of poly(ethylene imine)-graft-poly(ethylene glycol) block copolymer/SiRNA polyplexes. Bioconjug Chem 17: 1209–1218.
25. Jeong, GJ, Byun, HM, Kim, JM, Yoon, H, Choi, HG, Kim, WK et al. (2007). Biodistribution and tissue expression kinetics of plasmid DNA complexed with polyethylenimines of different molecular weight and structure. J Control Release 118: 118–125.
26. Turk, M, Dincer, S, Yulug, IG and Piskin, E (2004). In vitro transfection of HeLa cells with temperature sensitive polycationic copolymers. J Control Release 96: 325–340.

Molecular Therapy vol. 17 no. 8 aug. 2009 1417
© The American Society of Gene & Cell TherapyRottlerin Inhibits Transfection by PEI-25/DNA
27. van der Aa, MAEM, Huth, US, Hafele, SY, Schubert, R, Oosting, RS, Mastrobattista, E et al. (2007). Cellular uptake of cationic polymer-DNA complexes via caveolae plays a pivotal role in gene transfection in COS-7 Cells. Pharm Res 24: 1590–1598.
28. von Gersdorff, K, Sanders, NN, Vandenbroucke, R, De Smedt, SC, Wagner, E and Ogris, M (2006). The internalization route resulting in successful gene expression depends on both cell line and polyethylenimine polyplex type. Mol Ther 14: 745–753.
29. Rejman, J, Conese, M and Hoekstra, D (2006). Gene transfer by means of lipo- and polyplexes: role of clathrin and caveolae-mediated endocytosis. J Liposome Res 16: 237–247.
30. Sarkar, K, Kruhlak, MJ, Erlandsen, SL and Shaw, S (2005). Selective inhibition by rottlerin of macropinocytosis in monocyte-derived dendritic cells. Immunology 116: 513–524.
31. Grimmer, S, van Deurs, B and Sandvig, K (2002). Membrane ruffling and macropinocytosis in A431 cells require cholesterol. J Cell Sci 115(Pt 14): 2953–2962.
32. Gschwendt, M, Müller, HJ, Kielbassa, K, Zang, R, Kittstein, W, Rincke, G et al. (1994). Rottlerin, a novel protein kinase inhibitor. Biochem Biophys Res Commun 199: 93–98.
33. Sallusto, F, Cella, M, Danieli, C and Lanzavecchia, A (1995). Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the
major histocompatibility complex class II compartment: downregulation by cytokines and bacterial products. J Exp Med 182: 389–400.
34. Grosse, S, Aron, Y, Thévenot, G, François, D, Monsigny, M and Fajac, I (2005). Potocytosis and cellular exit of complexes as cellular pathways for gene delivery by polycations. J Gene Med 7: 1275–1286.
35. Duchardt, F, Fotin-Mleczek, M, Schwarz, H, Fischer, R and Brock, R (2007). A comprehensive model for the cellular uptake of cationic cell-penetrating peptides. Traffic 8: 848–866.
36. Hoekstra, D, Rejman, J, Wasungu, L, Shi, F and Zuhorn, I (2007). Gene delivery by cationic lipids: in and out of an endosome. Biochem Soc Trans 35(Pt 1): 68–71.
37. Khalil, IA, Kogure, K, Futaki, S and Harashima, H (2006). High density of octaarginine stimulates macropinocytosis leading to efficient intracellular trafficking for gene expression. J Biol Chem 281: 3544–3551.
38. Jones, AT (2007). Macropinocytosis: searching for an endocytic identity and role in the uptake of cell penetrating peptides. J Cell Mol Med 11: 670–684.
39. Schaffert, D and Wagner, E (2008). Gene therapy progress and prospects: synthetic polymer-based systems. Gene Ther 15: 1131–1138.
40. Bieber, T, Meissner, W, Kostin, S, Niemann, A and Elsasser, HP (2002). Intracellular route and transcriptional competence of polyethylenimine-DNA complexes. J Control Release 82: 441–454.
Copyright © 2022 FDOKUMEN




















