
Development of bacterial inoculums based on biodegradable ...
Upload
khangminh22Category
view
0download
0
Linköping University Medical Dissertations No. 1147
Biodegradable gelatin microcarriers in tissue engineering
In vitro studies on cartilage and bone
Sofia Pettersson
Division of Surgery
Department of Clinical and Experimental Medicine Faculty of Health Sciences
SE-581 85 Linköping, Sweden
Linköping 2009

© Sofia Pettersson, 2009, unless otherwise stated. ISBN 978-91-7393-557-9 ISSN 0345-0082
During the course of the research underlying this thesis, the author was enrolled in Forum
Scientium, a multidisciplinary doctoral program at Linköping University, Sweden.
Published articles have been reprinted with the permission of the respective copyright holder:
Paper I: John Wiley & Sons, Ltd. 2009. Paper II: British Association of Plastic, Reconstructive
and Aesthetic Surgeons. Published by Elsevier, Ltd. 2009.
Cover: Macroporous gelatin microcarrier
Printed by LiU-Tryck, Linköping, Sweden, 2009

Till pappa.

Supervisor Gunnar Kratz, Professor
Laboratory for Reconstructive Plastic Surgery
Department of Clinical and Experimental Medicine
Linköping University, Sweden
Co-Supervisors Pentti Tengvall, Professor
Institute of Clinical Sciences
Department of Biomaterials
The Sahlgrenska Academy at University of Gothenburg
Göteborg, Sweden
Jonas Wetterö, Ph.D.
Rheumatology/AIR
Department of Clinical and Experimental Medicine
Linköping University, Sweden
Opponent Julie Gold, Associate Professor
Division of Biological Physics
Department of Applied Physics
Chalmers University of Technology
Göteborg, Sweden

ABSTRACT
Tissue engineering is a multidisciplinary field that combines cells, biomaterial scaf-
folds and environmental factors to achieve functional tissue repair. This thesis focuses
on the use of macroporous gelatin microcarriers as scaffolds in tissue engineering
applications, with a special focus on cartilage and bone formation by human adult
cells in vitro.
In our first study, human articular chondrocytes were seeded on macroporous
gelatin microcarriers. The microcarriers were subsequently encapsulated in coagu-
lated blood-derived biological glues and cultured under free-swelling conditions for
up to 17 weeks. Even in the absence of recombinant chondrogenic growth factors,
the chondrocytes remained viable and metabolically active for the duration of the
culture period, as indicated by an increased amount of cell nuclei and extracellular
matrix (ECM). The ECM showed several cartilage characteristics, but lacked the
cartilage specific collagen type II. Furthermore, ECM formation was seen primarily
in a capsule surrounding the tissue-engineered constructs, leading to the conclusion
that the used in vitro models were unable to support true cartilage formation.
The capacity of human dermal fibroblasts to produce cartilage- and bone-like tissue
in the previously mentioned model was also investigated. Under the influence of
chondrogenic induction factors, including TGF-β1 and insulin, the fibroblasts pro-
duced cartilage specific molecules, as confirmed by indirect immunohistochemistry,
however not collagen type II. Under osteogenic induction, by dexamethasone,
ascorbate-2-phosphate and β–glycerophosphate, the fibroblasts formed a calcified
matrix with bone specific markers, and an alkaline phosphatase assay corroborated a
shift towards an osteoblast like phenotype. The osteogenic induction was enhanced
by flow-induced shear stress in a spinner flask system.
In addition, four different types of gelatin microcarriers, differing by their inter-
nal pore diameter and their degree of gelatin cross-linking, were evaluated for their
ability to support chondrocyte expansion. Chondrocyte densities on the microcarriers
were monitored every other day over a two-week period, and chondrocyte growth
was analyzed by piecewise linear regression and analysis of variance (ANOVA). No

ii
differences were seen between the different microcarriers during the first week. How-
ever, during the second week of culture both microcarrier pore diameter and gelatin
cross-linking had significant impacts on chondrocyte density.
Lastly, a dynamic centrifugation regime (f=12.5 mHz for 16 minutes every other
day) was administered to chondrocyte-seeded microcarriers, with or without encap-
sulation in platelet rich plasma (PRP), to study the possible effect of dynamic stimuli
on cartilage formation. Presence of PRP enhanced the structural stability of the
tissue-engineered constructs, but we were not able to confirm any dose-response
pattern between ECM formation and the applied forces. After 12 weeks, distinct
gelatin degradation had occurred independent of both dynamic stimuli and presence
of PRP.
In summary, this thesis supports a plausible use for gelatin microcarriers in tissue
engineering of cartilage and bone. Microcarrier characteristics, specifically gelatin
cross-linking and pore diameter, have been shown to affect chondrocyte expansion.
In addition, the use of human dermal fibroblasts as an alternative cell source for car-
tilage and bone formation in vitro was addressed.


Biodegradable gelatin microcarriers in tissue engineering
iv
TABLE OF CONTENTS
Abstract .................................................................................................. iTable of contents .................................................................................. ivAbbreviations.........................................................................................6List of papers..........................................................................................7Introduction...........................................................................................9
Articular cartilage .......................................................................................................... 9Tissue Engineering....................................................................................................... 13Cells ............................................................................................................................. 15Biomaterials ................................................................................................................. 17Environmental factors .................................................................................................. 21
Aims of the study .................................................................................25Paper I ......................................................................................................................... 25Paper II ........................................................................................................................ 25Paper III ....................................................................................................................... 25Paper IV ....................................................................................................................... 25
Comments on methods........................................................................27Cell culture .................................................................................................................. 27Analysis methods ......................................................................................................... 33
Results and discussion..........................................................................39Gelatin microcarriers support adhesion and expansion of human articular chondrocytes (HACs) and human dermal fibroblasts (HDFs) ............................................................. 39Blood-derived glues can be used for microcarrier encapsulation in vitro...................... 39PRP encapsulation enhances pellet stability and generates pellets with more uniform morphologies............................................................................................................... 40Expanded HACs produce cartilage-like tissue components but do not generate cartilage tissue in free-swelling conditions in the absence of recombinant chondrogenic growth factors.......................................................................................................................... 41Dynamic centrifugation every other day does not dramatically alter ECM synthesis of microcarrier-expanded HACs but may contribute to collagen fiber organization ........ 42HDFs produce cartilage-like tissue components on gelatin microcarriers under chondrogenic induction .............................................................................................. 43

Table of contents
v
HDFs produce bone-like tissue components and upregulate ALP activity on gelatin microcarriers under osteogenic induction ...................................................................43Flow-induced shear stress enhances ECM mineralization and affects ALP activity in HDFs.....................................................................................................................................44Gelatin cross-linking affects HAC proliferation during microcarrier expansion ............44Pore diameter affects HAC proliferation during microcarrier expansion ......................45Summary......................................................................................................................46
General discussion and future perspectives ......................................... 47On the efficacy of microcarriers for tissue engineering of cartilage..............................47On the use of in vitro models ........................................................................................48On the influence of microcarrier characteristics ...........................................................48On the use of dermal fibroblasts in tissue engineering ................................................49
Conclusions ......................................................................................... 51Acknowledgements ............................................................................. 53References ........................................................................................... 59

ABBREVIATIONS
2D/3D two/three-dimensional ACI/ACT autologous chondrocyte
implantation/transplantation ALP alkaline phosphatase ANOVA analysis of variance BMP bone morphogenetic protein BMSC bone marrow stem cells CS CultiSpher (Paper III) DAPI 4’6-diamino-2-phenylindole DMEM Dulbecco's modified Eagle's medium ECM extracellular matrix EDTA ethylenediaminetetraacetic acid FCS fetal calf serum FGF fibroblast growth factor g gram (unit) g acceleration (unit) G, GL and GLS CultiSpher microcarrier types GAG glucoseaminoglycan H&E hematoxylin & eosin HAC human articular chondrocytes HDF human dermal fibroblasts IGF insulin-like growth factor IHC immunohistochemistry LM light microscopy MEM minimal essential medium MSC mesenchymal stem cells MTT 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide NCS newborn calf serum OD optical density PBS phosphate buffered saline PCA personal cell analysis (Guava PCA) PLM polarized light microscopy PPP platelet poor plasma PRP platelet rich plasma PSR picrosirius red RCF relative centrifugal force RT room temperature SEM scanning electron microscopy SZP superficial zone protein, also known as PRG4 or lubricin TGF-β transforming growth factor beta WB whole blood

LIST OF PAPERS
Paper I Sofia Pettersson, Jonas Wetterö, Pentti Tengvall, and Gunnar Kratz
Human articular chondrocytes on macroporous gelatin microcarriers
form structurally stable constructs with blood-derived biological glues in
vitro
Journal of Tissue Engineering and Regenerative Medicine. 2009; 3: 450 – 460.
Paper II Pehr Sommar, Sofia Pettersson, Charlotte Ness, Hans Johnson,
Gunnar Kratz, and Johan P.E. Junker
Engineering three-dimensional cartilage- and bonelike tissues using
human dermal fibroblasts and macroporous gelatine microcarriers
Journal of Plastic, Reconstructive & Aesthetic Surgery. 2009
In press
Paper III Sofia Pettersson, Jonas Wetterö, Pentti Tengvall, and Gunnar Kratz
Cell expansion of human articular chondrocytes on macroporous gela-
tin microcarriers – impact of pore diameter and degree of gelatin cross-
linking on cell proliferation
Manuscript
Paper IV Sofia Pettersson, Jonas Wetterö, and Gunnar Kratz
The role of platelet rich plasma and dynamic centrifugation on
extracellular matrix formation of human articular chondrocytes on
macroporous gelatin microcarriers in pellet culture
Manuscript


INTRODUCTION
A frequent problem in modern medicine involves the loss or failure of tissues and
organs. The consequences can be devastating for the individual patients, and treat-
ment options are often limited by the lack of suitable donor tissue for transplantation
or of appropriate prosthetic alternatives. The field of tissue engineering aims at re-
storing lost tissue by combining engineering and medical sciences, to develop func-
tional biological substitutes [Langer and Vacanti 1993].
This thesis covers the use of gelatin microcarriers in tissue engineering applica-
tions, with focus on cartilage and bone formation in vitro. The basics and issues in-
volved with tissue engineering will be discussed using examples from articular carti-
lage, as this presents relevant background for all four papers included in this thesis.
Hence, the general function and structure of cartilage will be outlined in the intro-
duction. For bone tissue other issues arise, most notably the need for vascularization,
but this tissue will only be discussed briefly.
Articular cartilage Articular cartilage serves as a load bearing, shock absorbing, and lubricating tissue in
diarthrodial joints, where it covers the contact ends of long bones (Figure 1a). Besides
being anchored to the underlying bone, the tissue is encased with synovial fluid that is
enclosed by a synovial membrane surrounding the joint. The cartilage is neither vas-
cularized nor innervated, and relies on the movement of synovial fluid to maintain
the function and metabolism of the residing cells. Despite its relatively simple com-
position, the tissue has remarkable material properties, withstanding contact pres-
sures in the megapascal range in human hip joints [Hodge et al. 1986].

Biodegradable gelatin microcarriers in tissue engineering
10
Figure 1: Articular cartilage covers the ends of long bones in diarthrodial joints (a). Several topographical zones can be distinguished between the synovial cavity and the subchondral bone (b). These zones differ in terms of collagen fiber organization (c) and chondrocyte mor-phology (d).
Articular cartilage structure Cartilage has a fairly basic structure and consists of one unique cell type, known as
chondrocytes, the extracellular matrix (ECM) that these cells produce and the syno-
vial fluid. The chondrocytes are responsible for the synthesis and turnover of the tis-
sue, yet they are sparsely distributed and make up a mere 1 % of the tissue volume.
The ECM is a complex network of collagens, proteoglycans, non-collagenous pro-
teins and glycoproteins that provide the tissue with its unique material properties.
The water content is high, accounting for 75–80 % of the wet tissue mass, while the
remaining matrix is composed of collagens (10–30 %), proteoglycans (3–10 %) and
non-collagenous proteins and glycoproteins [Schulz and Bader 2007]. The collagen
content is dominated by collagen type II (90–95 %) and these fibers provide the tissue
with tensile strength. Other collagen types, including collagen type IX and VI, are
found mainly in the pericellular regions surrounding the chondrocytes [Poole 1997].
The major proteoglycan in articular cartilage tissue is aggrecan. It consists of a pro-
tein backbone, to which several glycoseaminoglycans (GAG), such as keratan sulfate
and chondroitin sulfate, are covalently attached. The highly negative charge of these
side molecules gives the biomolecule a high swelling capacity that in turn enables the
tissue to absorb high compressive forces [Schulz and Bader 2007].
Articular cartilage can be categorized into different topographical zones – the
superficial zone, the middle zone, the deep zone, the tidemark zone and the calcified
zone (Figure 1b). In the superficial zone, chondrocyte morphology is flattened, and
the ECM is densely packed with collagen fibers running parallel to the surface. In the

Introduction
11
middle zone, the cell density is reduced, and the chondrocytes display a more spheri-
cal morphology. Only few direct cell–cell interactions occur between the chondro-
cytes as the cells are relatively isolated, either individually or in small aggregates, in
small cavities within the ECM called lacunae. The organization of the collagen fibers
becomes more stochastic in the middle zone, and the proteoglycan content in the
ECM increases. Even further down, in the deep zone, the chondrocytes are arranged
in columns perpendicular to the surface and underlying bone (Figure 1d). Collagen
fibers in the deep zone are also organized in this manner. The specific molecular
composition of the ECM varies between the zones, with ECM molecules residing
uniquely in some zones. For example, the superficial zone protein (SZP, also known
as PRG4 or lubricin) acts as a lubricant at cartilage surfaces and is not found in the
deeper zones [Schumacher et al. 1994]. Among the collagens, collagen type X is cou-
pled with chondrocyte hypertrophy and is considered unique for the calcified zone
that borders on the underlying bone [Schulz and Bader 2007]. The cellular activity
also differs between the topographical zones, and the metabolic potential of the cells
can vary depending on their topographic origin [Lee et al. 1998; Darling and
Athanasiou 2005; Stenhamre et al. 2008].
Articular cartilage homeostasis relies on biomechanical conditioning. Joint
movement leads to compression of the cartilage tissue and the displacement of syno-
vial fluid, resulting in complex patterns of direct compression, hydrostatic pressure,
tensile and shear forces throughout the tissue. Chondrocytes respond to these forces
and the biomechanical stimuli is converted into intracellular signals through a process
known as mechanotransduction. The detailed mechanisms of the mechanotransduc-
tion pathways, or the causal relationships between the applied forces and cellular re-
sponses, are not yet fully understood, but the current view has recently been reviewed
[Ramage et al. 2009].
Articular cartilage pathology Under normal conditions, articular cartilage is subject to little or no wear. Once
damaged through trauma or degenerative joint disease, for example osteoarthritis,
articular cartilage has a limited capacity for self-repair. The avascular nature of the
tissue inhibits progenitor cells to be introduced from the blood flow. Also, unlike
other forms of hyaline cartilage, articular cartilage is not surrounded by a perichon-
drium from which chondroprogenitor cells can migrate and subsequently differenti-
ate into chondrocytes that contribute to cartilage healing. Instead, chondrogenesis

Biodegradable gelatin microcarriers in tissue engineering
12
must be accomplished by residing cells, through interstitial growth. The intrinsic
ability of chondrocytes to do so is however limited. There have been recent reports
on mesenchymal progenitor cells residing in articular cartilage, but these findings also
indicate a possible correlation with osteoarthritis [Alsalameh et al. 2004].
There is a vast demand for clinically applicable cartilage regeneration strategies.
In Sweden alone, 9600 total knee replacements and 14000 hip replacements were
performed in 2007 [Höftprotesregistret 2008; Knäprotesregistret 2008]. The clinical
approaches in practice today for articular cartilage repair often rely on the introduc-
tion of progenitor cells to the wound site. Microdrilling and microfracture are both
subchondral techniques that induce bleeding and clot formation in the cartilage de-
fect [Redman et al. 2005]. Progenitor cells from the blood flow and the bone marrow
are then introduced to the void where they differentiate towards a chondrogenic
phenotype. Soft tissue grafts, such as perichondrium and periosteum grafts, also in-
troduce progenitor cells.
In 1994, Brittberg et al. described a procedure where a single cell suspension of
expanded autologous chondrocytes is introduced underneath a periosteum flap that
has been firmly sutured over the debrided cartilage defect [Brittberg et al. 1994]. The
autologous chondrocyte implantation (ACI) method, sometimes referred to as auto-
logous chondrocyte transplantation (ACT), thus offers direct delivery of differentiated
chondrocytes as well as the possibility of migrating progenitor cells from the perio-
steum to assist during cartilage regeneration. Long-term outcomes demonstrate im-
provement for several patients [Peterson et al. 2000; Brittberg et al. 2003]. However,
the original method is rather invasive, as it requires open knee surgery to attach the
periosteum securely, and subsequent research has focused on finding arthroscopic
methods to shorten patient recovery periods [Erggelet et al. 2003; Zheng et al. 2007].
The ACI method from 1994 is one of the most referenced methods in the field of
cartilage tissue engineering.
Though several cartilage repair strategies improve joint function, most yield a
cartilage tissue that is immature and fails to meet the mechanical properties of arti-
cular cartilage [Redman et al. 2005]. In addition, the repair tissue often lacks the
zonal organization of articular cartilage. Combined, these shortcomings may lead to
insufficient tissue integration and degradation of the repair tissue. The field of carti-
lage tissue engineering aims to identify materials and methods that will improve car-
tilage tissue repair.

Introduction
13
Tissue Engineering According to a recent editorial, the term tissue engineering was first described in the
1980s, and the first official definition was agreed upon in 1987 [Lysaght and Crager
2009]. There have been many subsequent attempts to redefine the term. In addition,
it has been coupled with the related term regenerative medicine.
Regenerativemedicine/tissue engineering is a rapidly grow‐
ing multidisciplinary field involving the life, physical, and
engineeringsciencesthatseekstodevelopfunctionalcell,tis‐
sue,andorgansubstitutestorepair,replace,orenhancebio‐
logical function that has been lost due to congenital abnor‐
malities,injury,disease,oraging.
NationalInstituteofBiomedicalImagingand
Bioengineering,2004
In practice, a common approach in tissue engineering is to seed cells on a biomaterial
scaffold that serves as a substrate onto which anchorage dependent cells can adhere.
Scaffolds can be designed and formed into practically any desired shape to suit the
intended application. The cell–biomaterial construct is then cultured in vitro and/or
implanted in vivo until tissue formation occurs (Figure 2). A widespread way to illus-
trate the field is thus to use a triangle, where each corner represent cells,
biomaterials and environmental factors respectively (Figure 3). Below,
environmental factors is used as a collective term for biomolecules, engineering methods,
and in vitro designs that are used with the purpose of initiating, stimulating, guiding or
enhancing tissue formation. The distinction between the research areas depicted in
the triangle is not always obvious. Biomaterials can for example, by their design
alone, stimulate cellular activities and tissue formation. This highly co-dependent
pattern between parameters illustrates the multidisciplinarity of the research field.

Biodegradable gelatin microcarriers in tissue engineering
14
Figure 2: A commonly used approach in tissue engineering involves an initial harvest of donor tissue (i), from which cells are isolated and subsequently expanded (ii). Expanded cells are seeded onto a biomaterial scaffold (iii) in an in vitro setting (iv). The resulting cell–bioma-terial construct can be cultured in vitro or directly implanted in vivo (v).
Figure 3: The triangle representing the tissue engi-neering paradigm where cells, biomaterials and en-vironmental factors are combined to engineer tissues.
The previously mentioned approach, to
seed cells on a biomaterial scaffold, has been named the open matrix strategy by Langer
and Vacanti and is frequently used [Langer and Vacanti 1993]. However, not all
regenerative strategies employ all of the three elements in the triangle. In guided regen-
eration, biomaterial scaffolds can be implanted without previous cell seeding. The
biomaterial then acts as a scaffold into which cells from adjacent tissues can migrate.
If instead the biomaterial is omitted, the term cell-based or scaffold-free tissue engineering
is often seen in the literature. The environmental factors can never be fully removed,
as cells are always affected by their surroundings, physiological or artificial. Current
research in this area is consequently focused on identifying the environmental cues
that will optimize tissue formation.

Introduction
15
Cells An ideal cell type for tissue engineering must be easy to harvest, isolate and expand
into great numbers in a rapid and cost-effective manner. Additionally, it should not
cause harm to or elicit an immune response in the intended host. In reality, this is not
always so easily accomplished. The supply of donor tissue is often limited and donor
site morbidity must be considered, especially for tissues where the capacity of self-
repair is limited, such as cartilage. For some cell types, the isolation of the desired cell
type can also be challenging and expansion techniques complex and costly.
Cell sources in tissue engineering The long-term clinical goal for most tissue engineering approaches involves the use of
autologous cells, that is, cells that have been isolated from the intended recipient. As
discussed above, the supply of these cells is often limited. For research purposes,
many researchers instead opt for animal cells or commercially available cell lines.
Animal cells are often easier to obtain and well-established cell lines offer the advan-
tage of reproducibility. The relevance of such findings is however difficult to put into
perspective, as the metabolic activity and phenotypic stability of the cells may vary
significantly between different species [Akens and Hurtig 2005; Giannoni et al. 2005].
This makes it difficult to compare and translate results obtained with such cells to
applications where primary human cells are eventually meant to be used. Even
within a species, results can vary depending on donor age and biopsy location
[Barbero et al. 2004; Akens and Hurtig 2005; Stenhamre et al. 2008]. The donor-to-
donor variability is one of the general difficulties with experimental designs using
human cells, as the reproducibility of obtained results can, and should, be questioned.
Another issue concerns differences between cells from unaffected donors and the in-
tended patient groups. For example, studies have shown discrepancies between
chondrocytes from osteoarthritic patients and healthy controls [Tallheden et al.
2005a; Yang et al. 2006].
Human cell sources in tissue engineering can be divided into i) differentiated, or
tissue specific, cells and ii) adult stem cells.
Differentiated cells Differentiated cells are specialized cells that have been isolated from the tissue of in-
terest. Intuitively, these cells are a logical choice, as they build and maintain the tis-
sues in vivo. However, there are some problems associated with their use. Apart from
the issues involving the limited supply of donor tissue and donor site morbidity that

Biodegradable gelatin microcarriers in tissue engineering
16
have been previously discussed, some differentiated cell types require time-consuming
and costly methods, such as for example co-culture with other cells, to adhere and
proliferate in vitro. The stability of the intrinsic phenotype is another issue. In the case
of cartilage, chondrocytes rapidly lose their phenotype when cultured in monolayer
culture in a process known as dedifferentiation [Holtzer et al. 1960; von der Mark et al.
1977]. The most crucial consequence of this process is that these cells no longer pro-
duce collagen type II, a cartilage-specific collagen that provides tensile strength.
Though these cells have been shown to regain some of their phenotypic characteris-
tics when transferred into a three-dimensional environment, a process consequently
known as redifferentiation, the redifferentiation potential is related to the expansion pe-
riod in vitro [Benya and Shaffer 1982; Darling and Athanasiou 2005; Kang et al.
2007]. This poses a powerful dilemma as high initial cell seeding densities signifi-
cantly improves the outcome [Mauck et al. 2003; Eyrich et al. 2007; Hayes et al.
2007]. Different strategies have been evaluated to protect the chondrogenic pheno-
type during in vitro expansion [Malda et al. 2003a; Malda et al. 2003b; Gigout et al.
2005; Hendriks et al. 2006].
Adult stem cells In recent years, researchers have begun to investigate alternative cell sources for re-
generative medicine, such as stem cells. Stem cells are undifferentiated cells that are
defined by their ability to self-renew and to differentiate along certain developmental
pathways. They are classified according to their differentiation potential. Totipotent
stem cells can give rise to any cell lineage, pluri- and multipotent stem cells are more
limited in this capacity, and monopotent cells are considered to be tissue-committed.
In vivo, these cells aid in the repair and renewal of human tissue, while maintaining
the adult stem cell population by self-renewal. Embryonic stem cells are by definition
totipotent, but the issues associated with the difficulty to control their differentiation
in vivo, are discouraging [Blum and Benvenisty 2008].
In regenerative medicine, the controlled differentiation of adult stem cells in vitro
has gained momentous attention in later years. Bone marrow stem cells, BMSCs, or
mesenchymal stem cells, MSCs, are able to proliferate in vitro while maintaining their
ability to differentiate towards chondrocyte, adipocyte and osteoblast phenotypes in
response to biochemical factors [Jaiswal et al. 1997; Yoo et al. 1998; Pittenger et al.
1999; Jaiswal et al. 2000]. Cells isolated from various other human tissues, including
adipose, muscle and dermal tissue, have also been reported to possess a certain level

Introduction
17
of stem cell plasticity in response to environmental factors [Young et al. 1995;
Warejcka et al. 1996; Young et al. 2001; Zuk et al. 2001; Jahoda et al. 2003; Bartsch et
al. 2005]. The underlying mechanisms, including the identification of possible cellular
sub-populations responsible for this multipotency, are subject to an ongoing debate
[Eisenberg and Eisenberg 2003].
The ability of adult stem cells to differentiate into phenotypes and produce tis-
sues distinct from their tissue of origin provides an exciting alternative to differenti-
ated cells. For these strategies to become clinically feasible, the challenges regarding
isolation, long-term expansion and classification of adult stem cells must be solved
[Gardner 2007; Seeger et al. 2007]. Yet, the fact that cells isolated from several di-
verse tissues possess the ability to alter their phenotype, in effect, challenges the con-
cept of adult somatic cell monopotency.
Biomaterials Numerous classes of biomaterials are used in medicine today, including metals, ce-
ramics, glasses, polymers and various composites. The field of biomaterial science is
vast, and covers areas well beyond tissue engineering. In order to understand the role
of biomaterials in tissue engineering, a brief introduction to the field, highlighting
some of the definitions and principles that are most relevant to tissue engineering and
this thesis, is given below.
In 1986, the European Society for Biomaterials agreed on the following defini-
tion [Williams 1987].
A biomaterial is a nonviablematerial used in amedical de‐
vice,intendedtointeractwithbiologicalsystems.
WilliamsDF,1987
With the emergence of several biomaterial applications in later years some alterations
have been suggested to the definition, especially regarding the words non-viable and
medical, to include the wide range of pre-clinical, analytical and regenerative uses for
these materials. The intent to interact with a biological system, specifically to a host,
requires the material not to elicit an inappropriate host response. This crucial point is
addressed with the term biocompatibility.

Biodegradable gelatin microcarriers in tissue engineering
18
Biocompatibilityistheabilityofamaterialtoperformwithan
appropriatehostresponseinaspecificapplication.
WilliamsDF,1987
The phrasing is deliberately vague, as both the material performance and the appro-
priate response may differ considerably between different applications. In fact, the
definitions have recently been revisited and rephrased to mirror this more clearly
[Williams 2008]. Finally, the scaffolds are often designed to act as an intermediate
support for tissue regeneration and are consequently meant to gradually degrade and
disappear when new tissue is formed. The term biodegradable is used with the following
definition.
Biodegradation is the chemical breakdown of materials by
the action of living organisms, which leads to changes in
physicalproperties.
WilliamsDF,1987
Though this is a wide definition, a host-induced deterioration of the biomaterial is
implied. Needless to say, the degradation products of a biocompatible material must
be biocompatible as well.
Biomaterial design in tissue engineering The role of a biomaterial scaffold in tissue engineering can be fundamentally different
from that of many biomaterials in various long-term medical devices, in that it should
typically more profoundly encourage and elicit a cellular response. The chemical and
architectural properties of a scaffold are utilized to trigger and optimize a response
rather than to minimize it, as would be the case for any biomaterial to be used in for
example coronary stents. The following definition for biocompatibility has been sug-
gested for scaffolds to be used in tissue engineering.
Thebiocompatibilityofascaffoldormatrixforatissueengi‐
neeringproductreferstotheabilitytoperformasasubstrate
that will support the appropriate cellular activity, including
the facilitation of molecular and mechanical signaling sys‐

Introduction
19
tems, inorder tooptimize tissue regeneration,without elic‐
itinganyundesirablelocalorsystemicresponsesintheeven‐
tualhost.
WilliamsDF,2008
This definition summarizes the crucial requirements for any biomaterial to be used in
tissue engineering. In short, it should support and optimize the formation of new tis-
sue without causing harm to the intended host. The challenge in biomaterial design
thus lies in the identification and optimization of the scaffold that will optimize the
tissue formation for each specific tissue and cell type.
A primary requirement is that the scaffold must support cellular adhesion. Not
all materials allow for direct cell–material interactions, and initial protein adsorption
is often a pre-requisite for such connections [Wilson et al. 2005]. Alternatively, the
surfaces can be chemically modified to allow for cell adhesion [Chen et al. 2006]. In
addition to surface chemistry, the topography and porosity of biomaterial surfaces
can affect cellular adhesion and morphology [Lee et al. 1994; Curtis and Wilkinson
1997; Mukherjee et al. 2008]. Porous structures enhance surface areas, facilitate ho-
mogenous cell distribution and enable cell–cell interactions, provided that pores are
interconnected [Spiteri et al. 2006; Chung et al. 2008; Kang et al. 2009]. The three-
dimensional structure of the bulk material, such as porosity and pore diameters, can
also affect cellular activities, including ECM synthesis, and should thus be identified
and chosen to favor a rapid and tissue-specific metabolism [Karageorgiou and
Kaplan 2005; Griffon et al. 2006; Yamane et al. 2007]. The material properties of the
scaffold should be considered to promote regeneration [Kelly and Prendergast 2006].
When biodegradable scaffolds are used, the degradation rate should be adjusted to
match the formation of new tissue, in order to avoid loss of integrity or inhibition of
ECM synthesis during tissue regeneration. The stability of the material can be con-
trolled by altering the chemical composition and degree of cross-linking [Zeugolis et
al. 2009]. Once implanted, the newly formed tissue should preferably integrate seam-
lessly with the surrounding tissue, to avoid the risk of wear at the interface. This can
be accomplished with a biological glue fixative, when the tissue has been developed in
vitro, or by encouraging adjacent cells to partake in tissue formation in situ.

Biodegradable gelatin microcarriers in tissue engineering
20
Polymer scaffolds in cartilage and bone tissue engineering A common approach in cartilage engineering, and the most relevant for this thesis, is
the use of porous polymer scaffolds. Both natural and synthetic polymers are being
investigated in musculoskeletal tissue engineering. Natural polymers are derived from
naturally occurring polymers. These include polymers derived from ECM proteins
and polysaccharides, such as collagen, gelatin and chondroitin sulfate, as well as
polymers derived from diverse parts of nature, including starch-based polymers, chi-
tosan and alginate [Malafaya et al. 2007]. The advantages of these polymers include
their intrinsic ability to support biological processes. However, it can be difficult to
retain control between different batches, resulting in material variability that may
affect reproducibility. In contrast, the chemistry and material properties of synthetic
polymers can be well controlled. Synthetic polymers include polyesters such as
poly(glycolic acid), poly(lactic acid) and their copolymers, polylactones, most notably
polycaprolactone, as well as various polyanhydrides and polyurethanes [Gunatillake
and Adhikari 2003]. These polymers offer extensive possibilities to tailor material
properties and also allow longer shelf life than natural polymers. On the other hand,
they lack the inherent ability to support biological processes and may therefore be
more prone to elicit foreign body responses from the immune system. A common
approach is to combine different polymers to optimize material properties and gain
some of the advantages from each type used [Gunatillake and Adhikari 2003; Hong
et al. 2005; Thissen et al. 2006; Malafaya et al. 2007; Lao et al. 2008].
The three-dimensional architecture of these materials can be varied in numer-
ous ways. Polymers can be woven, spun, freeze-dried, solvent-cast or processed with
gas foam into porous three-dimensional structures. With the ACI method from 1994
in mind, scaffold materials are often designed to allow for arthroscopic delivery.
Apart from development of deformable sponges, that can also be administered
arthroscopically, two strategies distinguish themselves – the use of hydrogels and
microcarriers. Hydrogels are defined as water-based colloidal gels, whereas microcarriers
often come in the form of sphere-shaped scaffolds, with diameters in the micrometer
range. A wide range of microcarriers, manufactured from synthetic and/or natural
polymers, with porous or non-porous structures, has been investigated as scaffolds for
cell expansion and tissue engineering [Frondoza et al. 1996; Malda et al. 2003b;
Chung et al. 2008; Lao et al. 2008]. The microcarriers that are the focus of this thesis
are manufactured from gelatin, derived from collagen type I, and have a macro-
porous structure (Figure 4).

Introduction
21
Figure 4: CultiSpher microcarriers are macroporous particles manufactured from gelatin, de-rived from collagen type I.
For bone, several different types of biomaterials, including ceramics, glasses and
various composites, are investigated in addition to the polymer approach
[Karageorgiou and Kaplan 2005]. Considering the natural link between the two tis-
sues, some efforts towards developing biphasic scaffolds for osteochondral constructs
have also been made, as has been recently reviewed elsewhere [Martin et al. 2007;
Keeney and Pandit 2009].
Environmental factors The most elusive discovery in tissue engineering concerns the identification of the
environmental factors required to stimulate optimal tissue formation in vitro and in
vivo. Several different strategies, all with the common goal of enhancing the quantity
and quality of the engineered tissue, are currently being investigated. Combinations
of two or more stimulating factors often yield synergistic effects, illustrating the need

Biodegradable gelatin microcarriers in tissue engineering
22
to identify the correct types, doses, combinations and sequences of these factors for
each tissue and cell type. Some of the most commonly used principles in tissue engi-
neering are discussed below.
Biochemical factors A potent and commonly used approach to stimulate differentiation and tissue forma-
tion is by adding biochemical factors to the cell culture media. These include growth
factors, hormones and cytokines that guide the proliferation and differentiation of
cells. Their use is especially crucial for guiding adult stem cells through induction of
differentiation pathways, but the strategy can also be used to preserve the intrinsic
phenotype, or trigger the redifferentiation of chondrocytes after the expansion pe-
riod. A large number of growth factors have been shown to have chondrogenic and
osteogenic properties. For cartilage, the transforming growth factor beta (TGF-β)
superfamily, including the bone morphogenetic protein (BMP) subclass, has proven
especially important, but several other factors, such as fibroblast growth factor (FGF),
insulin-like growth factor (IGF) as well as insulin itself have positive effects on
chondrogenesis [Leboy et al. 1997; Jakob et al. 2001; Blunk et al. 2002; Awad et al.
2003; Malda et al. 2003a]. In addition, the presence and concentrations of other me-
dia additives, such as ascorbic acid and calcium, affect cellular mechanisms [Sullivan
et al. 1994; Leboy et al. 1997; Gigout et al. 2005]. The chondrogenic and osteogenic
differentiation pathways are closely related, and the BMPs in the TGF-β superfamily
are consequently highly significant for bone formation as well [Cheng et al. 2003].
Other widely used osteogenic differentiation factors include ascorbate-2-phosphate,
dexamethasone and β-glycerophosphate [Grigoriadis et al. 1988; Cheng et al. 1994;
Zuk et al. 2001].
As an alternative to recombinant growth factors, autologous sources for chon-
drogenic growth factors have recently gained interest. For cartilage, human platelet
supernatants have been used as cell culture media additives during chondrocyte ex-
pansion as well as three-dimensional culture [Gaissmaier et al. 2005; Tallheden et al.
2005b; Akeda et al. 2006]. In these cases, platelets from human plasma have been
activated and the growth factors released from platelet alpha granules subsequently
extracted in liquid form. Alternatively, platelet rich plasma (PRP) can be used as a
scaffold or as part of the study design for cell adhesion purposes [Malicev et al. 2007;
Wu et al. 2007; Haberhauer et al. 2008]. Regardless of method, the concentration of
growth factors can differ as a result of donor-to-donor variability as well as differences

Introduction
23
in the methods used during plasma coagulation and platelet activation [Weibrich et
al. 2002; Tallheden et al. 2005b]. A strategy that merges with biomaterial surface
modification involves recombinant growth factors to be chemically bound to or en-
trapped within biomaterial scaffolds to allow for controlled release during in vivo or in
vitro culture [Fan et al. 2006].
Biomechanical factors In many cases, the quantity and quality of the engineered tissue is enhanced when
tissues are cultured under the influence of biomechanical conditioning. This is espe-
cially true when engineering tissues that experience biomechanical cues in vivo, such
as bone, cartilage and blood vessels.
A bioreactor can be regarded as a well-defined in vitro system that controls or sup-
ports a biologically active environment. As such, the definition can apply to an ordi-
nary incubator. For tissue engineering purposes however, the optimal bioreactor sys-
tem should enable a controlled environment that mimics the physiological conditions
that appear in vivo [Schulz and Bader 2007]. In reality, this task is difficult to achieve
and most bioreactors mainly aim to minimize the difference between the complex,
fine-tuned in vivo environment and the comparatively crude in vitro system. This can
be achieved by maintaining conditions, for example by introducing one or more pa-
rameters, that stimulate tissue formation in vitro. In the literature, the term is often
used for any device that improves cell seeding, cell specific behavior or tissue forma-
tion in vitro.
Several different principles for biomechanical stimulation are used in bioreactors
for cartilage and bone engineering, including compression, hydrostatic pressure,
perfusion, shear stress as well as combinations thereof. The rationale for using bio-
reactors in cartilage engineering, along with detailed descriptions on the different
principles and systems used, has recently been extensively reviewed [Schulz and
Bader 2007]. In addition to the underlying principle of the applied forces, the dura-
tion, frequency and amplitude of the administered regimes matter [Lee and Bader
1997; Maeda et al. 2001; Davisson et al. 2002; Waldman et al. 2003; Waldman et al.
2004].
Oxygen tension In addition to biochemical and biomechanical factors, other environmental factors
have also gained recent interest. In vivo, articular cartilage is subjected to low oxygen
levels of approximately 1–6 % [Silver 1975; Fermor et al. 2007]. The importance of

Biodegradable gelatin microcarriers in tissue engineering
24
these hypoxic conditions has been investigated recently, and reduced oxygen levels
during three-dimensional in vitro culture elevate tissue formation in comparison to
normoxic conditions [Domm et al. 2002; Malda et al. 2004; Wernike et al. 2008]. Hy-
poxia also induces chondrogenic differentiation of MSC cell lines [Robins et al. 2005].
3D environment The three-dimensional environment experienced by the cells can also have stimulat-
ing qualities. For example, the accumulation or presence of bonelike ECM molecules
enhance the osteogenic differentiation of both rat and human MSCs [Salasznyk et al.
2004; Datta et al. 2006]. This again highlights the significance of cell–material
interactions.

AIMS OF THE STUDY
The overall aim of this thesis has been to investigate the use of biodegradable macro-
porous gelatin microcarriers in tissue engineering applications with focus on in vitro
formation of cartilage and bone.
Paper I To investigate the use of chondrocyte-seeded gelatin microcarriers for in vitro forma-
tion of cartilage. To evaluate the potential of blood-derived glues for encapsulation of
cell-seeded microcarriers. To appraise the performance of two free-swelling culture
models for in vitro formation of cartilaginous tissue in the absence of recombinant
growth factors.
Paper II To study in vitro formation of bone- and cartilage-like tissue by human dermal fibro-
blasts on gelatin microcarriers under osteogenic and chondrogenic induction. To
investigate the influence of flow-induced shear stress on osteogenic differentiation of
human dermal fibroblasts adhering to gelatin microcarriers in a spinner flask system.
Paper III To analyze the impact of microcarrier characteristics, specifically pore diameters and
degrees of gelatin cross-linking, on chondrocyte expansion in a spinner flask system.
Paper IV To explore the effect of dynamic centrifugation every other day on ECM synthesis by
human articular chondrocytes on gelatin microcarriers. To examine the influence of
platelet rich plasma (PRP) in this model.


COMMENTS ON METHODS
This chapter features the methods used during this thesis. For further specific details
regarding the materials and methods used, see the Materials and methods section of
Papers I–IV.
Cell culture
Chondrocyte isolation In Papers I, III and IV, human articular chondrocytes (HACs) were isolated from
tissue obtained from total knee arthroplasties. The discarded tissue was covered with
sterile saline solution during transportation, and, unless processed immediately, cov-
ered with low serum cell culture media and kept at 4 °C over night. Visibly damaged,
i.e. coarse and yellowed, tissue was removed and the remaining cartilage tissue was
dissected and enzymatically digested according to the details in each respective pa-
per. Chondrocytes were isolated from both the femoral and tibial condyles, from all
topographical zones. When the cell yield following chondrocyte isolation was deter-
mined, it reached approximately 8 × 106 chondrocytes per knee.
Fibroblast isolation In Paper II, human dermal fibroblasts (HDFs) were isolated from dermal tissue fol-
lowing routine plastic surgery. The tissue was stored briefly in sterile saline and proc-
essed within 24 hours. The dermal layer was dissected in smaller fragments and fi-
broblasts were isolated by enzymatic digestion according to Paper II.
Cell culture media Initially, all chondrocyte expansion and tissue construct cultures were conducted us-
ing a basic growth media. This cell culture media was later changed to a frequently

Biodegradable gelatin microcarriers in tissue engineering
28
used chondrocyte growth media [Freed et al. 1997; Malda et al. 2003a; Chung et al.
2008]. In Paper I, both cell culture media are represented in the included experi-
ments. In Papers III–IV, only the chondrocyte growth media was used.
With the exception of Paper II, where TGF-β1 plays a crucial part in the study
design, no recombinant chondrogenic growth factors were added to the cell culture
media. Several chondrogenic growth factors, TGF-β1 included, are however known
to be released from platelet alpha granules [Weibrich et al. 2002; Gaissmaier et al.
2005; Akeda et al. 2006]. This concerns all groups using PRP and whole blood
encapsulation, in Papers I, II and IV respectively. Recombinant growth factors were
excluded to minimize the risk of masking the effects of the included materials. For
example, the relevance of any comparison between the platelet rich plasma (PRP)
and platelet poor plasma (PPP) groups in Paper I would have been limited if a pow-
erful chondrogenic growth factor had been continuously added.
The differentiation factors used in Paper II for chondrogenic and osteogenic
differentiation have been described previously [Grigoriadis et al. 1988; Pittenger et al.
1999; Zuk et al. 2002]. In short, the chondrogenic induction media contained TGF-β1
and insulin, whereas the osteogenic induction media contained dexamethasone and
β-glycerophosphate. In addition, both media contained ascorbate-2-phosphate.
These induction media have previously been used for differentiation of HDFs in
monolayer culture [Junker et al. 2009].
Expansion methods In Papers I and III, HACs were expanded by traditional monolayer culture tech-
niques using polystyrene flasks. This cell culture technique yields great expansion
numbers for many cell types, including chondrocytes. The disadvantages of using this
type of plastic have been mentioned previously [Holtzer et al. 1960; von der Mark et
al. 1977]. In Paper I (Experiment VI) and Paper IV, HACs were seeded directly onto
the gelatin microcarriers once isolated. These changes were made to enhance the
chondrocyte redifferentiation potential [Malda et al. 2003a; Melero-Martin et al.
2006]. In Paper II, HDFs were expanded in monolayer using polystyrene flasks. The
cells were then seeded onto microcarriers and expanded for an additional two weeks
in spinner flasks before induction started.
Microcarrier selection In Papers I, II and IV, CultiSpher GL microcarriers were used. Compared to the
more commonly used CultiSpher G and CultiSpher S microcarriers, this microcar-

Comments on methods
29
rier has a slightly larger average internal pore diameter, 30 µm compared to 20 µm,
when hydrated in PBS (for details, refer to Paper III). The microcarrier choice for
these papers was made before the results of Paper III were analyzed, and was based
on visual evaluation of sectioned and histologically stained material, indicating that
cells did not occupy the interior of the microcarriers. The rationale for using a micro-
carrier with a larger pore size was thus to encourage cells to migrate into these pores
and populate the entire microcarrier.
Microcarrier encapsulation In Paper I, microcarriers were encapsulated in whole blood, re-calcified citrated
PRP, re-calcified citrated PPP and diluted Tisseel [Goessl and Redl 2005]. Non-di-
luted Tisseel has also been evaluated, but our results indicated that it was too dense to
support diffusion of nutrients in free-swelling conditions. In Papers II and IV, gelatin
microcarriers were encapsulated in re-calcified citrated PRP. The basis for using PRP
for encapsulation originates from previous research indicating that PRP, when used
in combination with bone grafts, accelerated bone healing [Marx et al. 1998]. We
hypothesized that PRP could be used to encapsulate pre-seeded microcarriers in a
similar way. The whole blood and PPP groups were added to the design following a
discussion regarding the role of platelets and other blood and plasma constituents in
this regard. As these glues are readily available from the patient’s own blood, they
present an appealing autologous alternative to the off the shelf commercial fibrin
glues. It is however important to note that the PRP used in this study was not pre-
pared from blood from the chondrocyte donors. There was no microcarrier encap-
sulation in Paper III.
In vitro models for tissue engineering of cartilage In Paper I, cell-seeded microcarriers were encapsulated in PRP in two models. The
first model utilizes cell culture inserts with permeable membranes. This model allows
media perfusion from the top as well as from the bottom of the tissue-engineered con-
structs. This approach was also used in Paper II. The second experimental model in-
volves forming of the clots as pellets in polypropylene tubes. As this strategy facilitates
centrifugation of tissue-engineered constructs, the pellet model was used in Paper IV.
Centrifugation of constructs in the cell culture insert model, by using microwell plate
holders, has also been performed according to an acceleration curve similar to those
used in Paper IV (see Biomechanical stimulation below). For both models, calcium
was added to initiate clotting of the citrated blood-derived glues. The most straight-

Biodegradable gelatin microcarriers in tissue engineering
30
forward way of differentiating between the two models is by comparing Experiments
V and VI in Paper I, where HACs from the same donor were used.
In vitro models for tissue engineering of bone Two different models were used for osteogenic induced fibroblasts in Paper II. In
addition to the cell culture insert model described above, cell-seeded microcarriers
were kept in a spinner flask system for the duration of the experiment, to investigate if
fluid flow-induced shear stress increases the level of osteogenic induction in HDFs.
Biomechanical stimulation There was no biomechanical stimulation in Paper I, as the study was designed to in-
vestigate cartilage formation under free-swelling conditions.
In Paper II, cell-seeded microcarriers were kept in a spinner flask system in fi-
broblast and osteoinductive cell culture media. Microcarrier aggregates were allowed
to form, as the primary objective in this study was to study 3D formation of bone
tissue. In Paper III, using the same spinner flask system, these aggregates were dis-
rupted to ensure a homogeneous distribution of microcarriers in the flask at each
sampling point.
In Paper IV, dynamic centrifugation was administered to the microcarrier pel-
lets according to three different acceleration curves, differing by their top relative
centrifugal force gmax (Figure 5). The centrifugation regime was administered for a
total of 16 min with a frequency of 12.5 mHz every other day. This is not within
physiological range, but dynamic stimuli have been proven efficacious at these fre-
quencies previously [Sah et al. 1989; Buschmann et al. 1995; Davisson et al. 2002].
The acceleration curves also incorporated a period of total rest, to allow the con-
structs to regain their shape after compression [Barker and Seedhom 1997].

Comments on methods
31
Figure 5: In Paper IV, dynamic centrifugation was administered with a Sigma K415C centrifuge with swing out rotors (a). Dynamic acceleration curves (f=12.5 mHz) were pro-grammed with differing top speeds, corresponding to 500 g, 1500 g, and 3000 g respec-tively (b).
In an unpublished study, direct unconfined compression was tested using a cus-
tom-build compression device (Figure 6) designed for use with cell culture inserts in a
12-well culture plate. The dynamic compression regime included loaded and non-
loaded periods and was run for a total of 15.5 min with a frequency of 13 mHz.
Again, this frequency is outside the physiological range but inside the effective range
[Sah et al. 1989; Buschmann et al. 1995; Davisson et al. 2002]. The same study in-
cluded cell culture insert plates subjected to dynamic centrifugation (gmax=500 g) with
the same frequency as the dynamic compression. Control constructs were kept under
free-swelling conditions. Due to the lack of quantitative control of the forces applied
to the constructs with the compression device, these results have not been published.

Biodegradable gelatin microcarriers in tissue engineering
32
Figure 6: The custom-built compression device used during supplemental experiments to this thesis.
Control groups In Paper I, containing six separate experiments, several different types of control
groups were included (see Paper I for details). In Experiment I and II, microcarrier-
free clots with monolayer-expanded chondrocytes were used to assess the role of the
gelatin scaffold. In Experiment V, cell-free microcarriers were embedded in PRP to
investigate the stability of the microcarrier–PRP clots. In Experiment VI, pellets were
formed with cell-seeded microcarriers in the absence of biological glue to study ma-
trix formation without PRP. In Paper II, controls included cell-free microcarriers
kept under identical conditions. To study the effects of the different induction media,
control fibroblasts were kept in equivalent models.
In Paper III, the CultiSpher G microcarrier served as a reference in the statisti-
cal analysis. The cell counts were converted to cell densities and normalized for gram
gelatin dry weight. For further details regarding the gelatin effect on cell counting, see
the details for the Guava PCA analysis below. In Paper IV, unstimulated groups
acted as control groups for the centrifugation regime, while PRP-free pellets served as
controls for each PRP-containing equivalent.
For controls related to analysis methods, see details for each method in the fol-
lowing section.

Comments on methods
33
Time points In Paper I, time points varied between 4 hours and 17 weeks. The earlier time points
were chosen to investigate when cell migration or cell death occurred at sample cen-
ters. Obtained results indicate that little or no matrix formed prior to 4 weeks. As a
result, samples were only taken after 4, 8 and 12 weeks of culture in Papers II and IV.
In Paper III, samples were withdrawn from the culture flasks every other day to
monitor cell growth.
Analysis methods This section highlights some aspects of the analysis methods that have been employed
to evaluate scaffold characteristics, cell densities as well as ECM formation.
Microcarrier characterization In Paper III, scanning electron microscopy (SEM) was used to visualize microcarrier
structures. Dehydrated microcarriers were sputtered with a 15-µm layer of platinum
prior to microscopy.
Viability assay In Papers I–IV, the viability of cells seeded on the microcarriers was evaluated with
the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
[Mosmann 1983; Denizot and Lang 1986]. The method is based on the reduction of
MTT and the formation of violet MTT formazan crystals in the cytoplasm of viable
cells. There has been no indication of non-specific staining of MTT associated with
the gelatin matrices used in this thesis. The Guava ViaCount assay can distinguish
between viable and non-viable cells, but this data has been used sparsely. In Paper
III, the viable cell count is used to determine the number of viable cells that were
seeded onto the gelatin microcarriers; however, the viable cell count was not used
when determining cell densities on the microcarriers. For details, see Cell counting
below and Paper III.
Cell counting In Papers I–IV, cell counts were established using a Guava Personal Cell Analysis
(PCA) flow cytometer with the Guava ViaCount assay. This method uses two DNA-
binding dyes with different permeabilities to distinguish between nucleated and dying
cells [Phi-Wilson et al. 2001]. When seeded on microcarriers, cells were detached by
dissolving the gelatin in a mixture of EDTA and trypsin. A representative plot from

Biodegradable gelatin microcarriers in tissue engineering
34
the ViaCount assay following microcarrier-based cell expansion can be seen in Figure
7. For well-populated microcarriers, prolonged trypsin incubations and vigorous
micropipette mixing were required to ensure a homogeneous cell solution. This may
well have affected the accuracy of the cell viability assay (for details, refer to Paper
III). This method was validated using i) dissolved cell-free microcarriers in
EDTA/trypsin, ii) EDTA/Trypsin and iii) ViaCount reagent alone. The results show
that the accuracy of the measurements is acceptable up to the third significant figure.
Figure 7: Representative plot from the Guava PCA flow cytometer ViaCount analysis used in Paper III.
Sample Processing In Papers I-IV, sample constructs were fixed in buffered formaldehyde, for up to 24
hours, and paraffin embedded prior to sectioning. Samples were uniformly positioned
when possible, yet differences in pellet morphologies and the influence of the sec-
tioning plane may have had an affect on the outcome of these analyses. In addition,
artifacts such as ripping, tearing and parallel structures derived from the movement
of the blade over the sample surface cannot be excluded in the sectioned material.
For constructs where modest ECM formation had occurred, the risk of microtome
tearing, in turn collapsing the section morphology, was evident.
Histology Routine histology methods were used for evaluation of the composition and mor-
phology of tissue-engineered constructs. In this thesis, the gelatin microcarriers
stained intensely for Mayer’s and Weigert’s hematoxylin, obscuring individual cell

Comments on methods
35
morphology on the scaffolds. For Alcian Blue, we occasionally observed a partial col-
oration of the scaffold. As an alternative to hematoxylin, cell nuclei were counter-
stained with DNA-binding 4’6-diamino-2-phenylindole (DAPI) and examined under
UV light.
Specialized stains were used in Papers II and IV. In Paper II, the von Kossa
stain was used to visualize mineralized extracellular matrix in the osteogenic induced
groups. This method is based on the binding of silver to carbonate and phosphate
groups, predominantly calcium phosphate and calcium carbonate, and positive
staining is indicative of mineralized matrix [Bills et al. 1971]. In Paper IV, a combina-
tion of picrosirius red and polarized light microscopy was used to investigate the oc-
currence of collagen fibers in centrifuged pellets [Junqueira et al. 1979]. The method
uses the birefringence of collagen fibers stained with picrosirius red to distinguish
between collagen fibers and randomly orientated collagen.
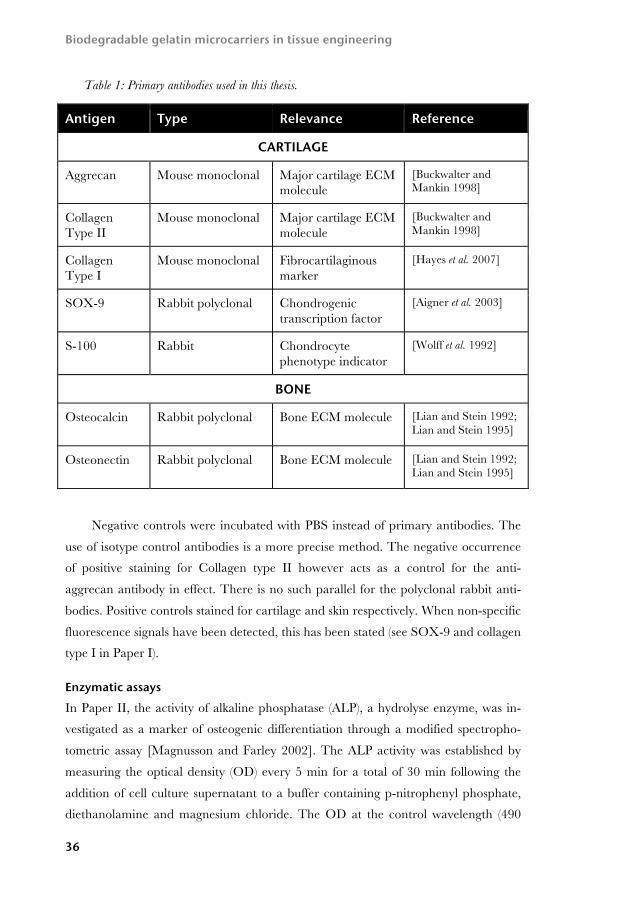
Immunohistochemistry For more specific determination of the ECM components, indirect immunohisto-
chemical analysis was performed for a number of antigens. These are described in
Table 1. In Papers I and IV, identical sets of antibodies were used, including aggre-
can, collagen types I and II, SOX-9 and S-100. In Paper III, SOX-9, S-100 and ag-
grecan were used. In Paper II, chondrogenic differentiation was evaluated with ag-
grecan and collagen type II, while osteogenic differentiation was evaluated immuno-
histochemically with anti-osteocalcin and osteonectin. The other antibodies of chon-
drogenic interest were taken into use after Paper II was submitted.
For most immunohistochemical applications, the signal to noise ratio was highly
improved by additional antigen retrieval and autofluorescence quenching. The anti-
gen retrieval and autofluorescence techniques used in this thesis are derived from the
individual data sheets of each antibody or modified from published data after evalu-
ating different techniques and sequences with each antibody. Primary antibody con-
centrations and incubation times have also been developed from the initial data sheet
recommendations or through systematic experiments evaluating different techniques
and sequences with each antibody. For context, cell nuclei were counterstained with
a DAPI-containing mounting media.
Biodegradable gelatin microcarriers in tissue engineering
36
Table 1: Primary antibodies used in this thesis.
Negative controls were incubated with PBS instead of primary antibodies. The
use of isotype control antibodies is a more precise method. The negative occurrence
of positive staining for Collagen type II however acts as a control for the anti-
aggrecan antibody in effect. There is no such parallel for the polyclonal rabbit anti-
bodies. Positive controls stained for cartilage and skin respectively. When non-specific
fluorescence signals have been detected, this has been stated (see SOX-9 and collagen
type I in Paper I).
Enzymatic assays In Paper II, the activity of alkaline phosphatase (ALP), a hydrolyse enzyme, was in-
vestigated as a marker of osteogenic differentiation through a modified spectropho-
tometric assay [Magnusson and Farley 2002]. The ALP activity was established by
measuring the optical density (OD) every 5 min for a total of 30 min following the
addition of cell culture supernatant to a buffer containing p-nitrophenyl phosphate,
diethanolamine and magnesium chloride. The OD at the control wavelength (490
Antigen Type Relevance Reference
CARTILAGE
Aggrecan Mouse monoclonal Major cartilage ECM molecule
[Buckwalter and Mankin 1998]
Collagen Type II
Mouse monoclonal Major cartilage ECM molecule
[Buckwalter and Mankin 1998]
Collagen Type I
Mouse monoclonal Fibrocartilaginous marker
[Hayes et al. 2007]
SOX-9 Rabbit polyclonal Chondrogenic transcription factor
[Aigner et al. 2003]
S-100 Rabbit Chondrocyte phenotype indicator
[Wolff et al. 1992]
BONE
Osteocalcin
Rabbit polyclonal Bone ECM molecule [Lian and Stein 1992; Lian and Stein 1995]
Osteonectin
Rabbit polyclonal Bone ECM molecule [Lian and Stein 1992; Lian and Stein 1995]

Comments on methods
37
nm) was subtracted from the OD at the maximum absorbance wavelength of p-
nitrophenyl phosphate (405 nm) to eliminate unspecific absorbance.
Statistical methods In Paper II, the ALP activity from osteogenic induced experimental groups and cor-
responding control groups was analyzed with linear regression. Regression line slopes
were compared to detect differences between experimental groups. In Paper III,
piecewise linear regression and analysis of variance (ANOVA) was used to determine
differences between growth characteristics on the four different microcarriers. A
break point was introduced at 7 days to allow for time dependent changes in the
growth characteristics. This time point was chosen with support from earlier studies
on chondrocyte proliferation, and is discussed in Paper III [Grad et al. 2003; Melero-
Martin et al. 2006; Wu et al. 2008]. Tukey pairwise comparison was used for further
comparison when significant differences were detected, using the CultiSpher G
microcarrier as a reference.


RESULTS AND DISCUSSION
This section highlights the major findings of this thesis. For detailed descriptions of
the results, please refer to each respective paper.
Gelatin microcarriers support adhesion and expansion of human articular chondrocytes (HACs) and human dermal fibroblasts (HDFs) In this thesis, four gelatin microcarriers – CultiSpher G, CultiSpher S, CultiSpher
GL and CultiSpher GLS – have been verified to support adhesion and expansion of
HACs (Papers I, III and IV). The CultiSpher GL microcarrier was shown to support
adhesion and expansion of HDFs (Paper II). These results cohere with observations
by others. The CultiSpher G microcarrier has previously been used for expansion of
human nasal chondrocytes and chondroprogenitor cells, while the CultiSpher S
microcarrier has been seeded with fibroblasts [Malda et al. 2003a; Melero-Martin et
al. 2006; Huss et al. 2007]. Once transferred to the gelatin microcarriers, the increase
in both cell number and accumulated ECM with time suggested an ongoing prolif-
eration and synthesis for both cell types (Papers I–IV).
Blood-derived glues can be used for microcarrier encapsulation in vitro Three re-calcified citrated blood-derived glues – whole blood (WB), platelet rich
plasma (PRP) and platelet poor plasma (PPP) – were all able to encapsulate the
microcarriers, and maintain an ongoing cell proliferation and synthesis of cartilage-
like tissue components for up to 16 weeks in vitro (Paper I). These results were not
dependent on the presence of platelets or other whole blood constituents, as all
blood-derived glues, as well as the commercially available fibrin glue, gave relatively

Biodegradable gelatin microcarriers in tissue engineering
40
similar results. When HACs were seeded in similar numbers in biological glue alone,
without the microcarriers, the clots dissolved before the chondrocytes were able to
produce sufficient amounts of ECM to uphold the structural integrity of the con-
structs. This correlates with earlier reports of rapid onset of fibrin glue disintegration
in vitro [Homminga et al. 1993]. Our results are also corroborated by findings that
presence of chitosan–GP polymer stabilizes blood clots from lysis [Marchand et al.
2009]. Others have evaluated PRP, blood plasma and fibrin glue as vehicles for ex-
panded chondrocytes without supporting microcarriers, and reported that porcine
chondrocytes accumulated more GAGs in blood plasma clots compared to whole
blood clots [Homminga et al. 1993; Wu et al. 2007; Haberhauer et al. 2008].
Results from Paper IV show that HAC-seeded microcarriers also form cartilage-
like tissue components in the absence of encapsulating glue, however at the expense
of some structural stability (see below).
PRP encapsulation enhances pellet stability and generates pellets with more uniform morphologies In Paper IV, the influence of PRP was examined by forming pellets of chondrocyte-
seeded microcarriers with or without plasma encapsulation. When compared to PRP-
free pellets, the data suggests that the engineered constructs are more stable and ob-
tain a more uniform morphology when the microcarriers are sealed within the
plasma. However, the immunohistochemical analysis did not disclose distinct differ-
ences between the two groups regarding the ECM components or the cellular mark-
ers. Nor did the plasma prove to be crucial for microcarrier degradation (Paper IV).
There is however a possibility that the presence of PRP may have accelerated the
chain of events, and that this could have been missed due to the choice of time points
and analysis methods. It is unclear what, if any, influence the single exposure of
chondrogenic growth factors from platelet alpha-granules may have had on the long-
term outcome, especially considering the longevity and efficiency of its stimulating
factors [Yoo et al. 1998; Landesberg et al. 2000; Jakob et al. 2001; Blunk et al. 2002;
Weibrich et al. 2002; McCarrel and Fortier 2009]. When used in combination with
porous collagen fleece scaffolds, both fibrin glue and blood plasma enhanced the col-
lagen type II gene levels of HACs [Malicev et al. 2007]. The uses of PRP for cell cul-
ture media supplementation, rather than for scaffold encapsulation, is discussed in
Paper I.

Results and discussion
41
Expanded HACs produce cartilage-like tissue components but do not generate cartilage tissue in free-swelling conditions in the absence of recombinant chondrogenic growth factors In each long-term culture using expanded HACs in this thesis, the amount of cells
and accumulated ECM increased with time, indicating an ongoing proliferation and
metabolism (Papers I and IV). Immunohistochemical analysis revealed presence of
the cartilage proteoglycan aggrecan in the newly formed ECM. In addition, cells
stained positive for the chondrogenic transcription factor SOX-9 and the calcium
binding protein S-100 that has been associated with the chondrogenic phenotype
[Wolff et al. 1992; Wehrli et al. 2003; Zheng et al. 2007]. However, the collagen con-
tent revealed a fibrocartilaginous ECM. None of the in vitro models were capable of
supporting full cartilage regeneration, either in terms of homogeny or quality, leading
to the conclusion that free-swelling in vitro culture conditions do not sustain 3D carti-
lage formation of expanded HACs on macroporous gelatin microcarriers in the ab-
sence of recombinant chondrogenic growth factors. These results may have been
affected by several factors. The chondrocytes used throughout this thesis has been
isolated from patients undergoing total knee arthroplasties. HACs from OA patients
have been shown to produce limited amounts of type II collagen in micromass pellet
culture, even in the presence of TGF-β1, and significantly less so than chondrocytes
isolated from unaffected donors [Tallheden et al. 2005a]. Though cartilage formation
can be achieved in high-density micromass pellet culture under free-swelling condi-
tions it is generally accepted that the homeostasis of articular cartilage is highly de-
pendent on mechanical conditioning [Schulz and Bader 2007]. For tissue engineer-
ing of larger cartilage constructs, where diffusion can no longer be relied on to
maintain the subsurface chondrocytes, several bioreactor approaches have been em-
ployed. These are thought not only to facilitate circulation of nutrients but also to
activate the mechanotransduction pathways via integrin-mediated cell–matrix inter-
actions [Millward-Sadler and Salter 2004; Ramage et al. 2009]. Several other
parameters can have significant effects on cartilage formation in vitro. These include,
but are not limited to, topographic origin of chondrocytes, cell expansion method,
cell culture passage number and initial seeding density on scaffolds [Lee et al. 1998;
Mauck et al. 2000; Malda et al. 2003a; Darling and Athanasiou 2005; Yang et al.
2006; Eyrich et al. 2007; Kang et al. 2007; Stenhamre et al. 2008].

Biodegradable gelatin microcarriers in tissue engineering
42
Dynamic centrifugation every other day does not dramatically alter ECM synthesis of microcarrier-expanded HACs but may con-tribute to collagen fiber organization In Paper IV, we evaluated dynamic centrifugation as a means of stimulating tissue
formation. Acquired results demonstrate that chondrocytes seeded on gelatin micro-
carriers remain viable and metabolically active when subjected to relative centrifugal
forces of up to 3000 g. There was no dramatic effect on matrix accumulation between
the centrifuged groups and the unstimulated controls. These results are corroborated
with observations from a parallel experiment, using the cell culture insert model from
Paper I, where tissue engineered constructs were subjected to dynamic centrifugation
with a peak relative centrifugal force of 500 g. Also, results obtained after dynamic
compression via a custom-built compression device, using equivalent frequencies,
closely resemble those seen with centrifuged and free-swelling constructs in the cell
culture insert model. In all these experiments, histological analysis revealed clear
signs of microcarrier degradation. The immunohistochemical analysis of the pellets
also generated similar results for all groups, regardless of exposure to centrifugation,
or presence of PRP (Paper IV). Previous studies have reported that centrifugation can
stimulate proteoglycan synthesis as well as DNA content [Inoue et al. 1990; Maeda et
al. 2001]. Others have used centrifugation every other day when studying different
aspects of chondrocytes in pellet culture [Nakagawa et al. 1993; Oyajobi et al. 1998].
Whether the lack of detectable beneficial effect has to do with the absence of soluble
factors described previously, shortages in the models themselves, or a combination of
both, cannot be distinguished from the experimental data. Considering the non-
quantitative nature of immunohistochemistry, potential subtle differences may have
been too modest to discern. Model shortages could involve inadequate duration, fre-
quencies or amplitudes of the administered regimes [Lee and Bader 1997; Lee et al.
1998; Maeda et al. 2001]. Furthermore, the reoccurrence of the biomechanical stim-
uli may affect the outcome [Maeda et al. 2001]. Centrifugal pressure may lead to
alignment of collagen fibers [Maeda et al. 2005], and histological analysis of the
centrifuged pellets indicated a higher level of fiber organization in the groups sub-
jected to high relative centrifugal forces, prompting further investigation. While the
picrosirius red analysis in Paper IV indeed revealed a distinct presence of collagen
fibers, there is a risk that any observations on fiber organization are dependent on
sectioning artifacts or discrepancies manifested during centrifugation.

Results and discussion
43
HDFs produce cartilage-like tissue components on gelatin micro-carriers under chondrogenic induction In Paper II, we investigated the ability of HDFs to produce cartilage-like tissue in a
3D cell culture insert model, analogous to the one described in Paper I. In accor-
dance with our earlier findings, the amount of cells and ECM increased with time,
indicating an ongoing proliferation and metabolism. The newly formed ECM stained
rich for aggrecan, indicating a cartilage proteoglycan synthesis and a differentiation
towards a chondrocyte like phenotype. However, no positive staining for collagen
type II could be detected. This may be attributed to the difficulty of retaining, or
rather in this case inducing, the collagen type II phenotype in vitro. However, HDFs
have been shown to stain positively for type II collagen when subjected to the same
chondrogenic induction media in monolayer culture, a result that we were not able to
reproduce here [Junker et al. 2009]. In addition, there have been reports of collagen
type II formation by mesenchymal and adipose derived adult stem cells under free-
swelling in vitro conditions [Awad et al. 2003; Li et al. 2005].
As has been discussed previously, the in vitro model used in Paper II proved in-
capable of supporting cartilage formation when used with HACs. The discussion re-
garding the in vitro model limitations also applies here. Furthermore, positive syner-
gistic effects with biomechanical stimuli, such as cyclic dynamic compression and
hydrostatic pressure, have been reported for chondrogenic differentiation of mesen-
chymal progenitor cells [Angele et al. 2003; Angele et al. 2004].
HDFs produce bone-like tissue components and upregulate ALP activity on gelatin microcarriers under osteogenic induction In Paper II, we also investigated the ability of HDFs to produce bone-like tissue when
seeded on gelatin microcarriers. Two models were used, the cell culture insert model
with PRP encapsulation, and a spinner flask model. Results regarding cell prolifera-
tion and ECM formation were analogous to the chondrogenic induced fibroblasts.
The total amount of matrix was greater in the spinner flask system than in the cell
culture inserts, arguably due to the mechanically active environment. Presence of the
bone ECM markers osteonectin and osteocalcin was confirmed by indirect immuno-
histochemistry in all osteogenic induced cultures. In addition, the von Kossa stain
revealed presence of mineralized matrix for both osteogenic induced models. These
results corroborate with earlier findings on similar induction of osteogenic phenotype
by HDFs [Lorenz et al. 2008]. The differentiation towards an osteoblast phenotype

Biodegradable gelatin microcarriers in tissue engineering
44
was also observed through an upregulated ALP activity of the HDFs under os-
teogenic induction. Importantly, fibroblast controls in the cell culture insert model
showed negative results for all these analysis, demonstrating that the osteogenic in-
duction indeed resulted in a phenotypical shift of the HDFs. These results concur
with recently published data concerning single cell HDF clones in monolayer culture
[Junker et al. 2009]. Results from the spinner flask model, highlighting the influence
of flow-induced shear stress, are discussed below.
Flow-induced shear stress enhances ECM mineralization and af-fects ALP activity in HDFs To investigate the influence of flow-induced shear stress on osteogenic induction,
HDFs were kept in a spinner flask system as well as in cell culture inserts. Matrix
mineralization was confirmed for osteogenic induced fibroblasts in both culture mod-
els, but the degree of mineralization was markedly higher in the spinner flask model
for the osteogenic induced fibroblasts. A similar pattern has been reported for human
MSCs on collagen scaffolds in spinner flasks [Meinel et al. 2004]. Interestingly, we
detected some extent of mineralization in the matrix produced by un-induced fibro-
blasts in the spinner flask model as well, indicating that shear stress can induce or
enhance matrix mineralization. The influence of shear stress on osteogenic differen-
tiation in the absence of soluble osteogenic factors has been reported previously,
though not with HDFs [Holtorf et al. 2005; Datta et al. 2006; Kreke et al. 2008]. We
also found an increase in ALP activity for the un-induced cells in spinner flasks com-
pared to control fibroblasts in cell culture inserts. However, there was no corre-
sponding elevation in ALP activity between the culture models for the osteogenic
induced fibroblasts. The results indicate that the key factor for increased ALP activity
is the soluble factors in the osteogenic induction media rather than shear stress. Fur-
thermore there was no formation of osteonectin or osteocalcin by control fibroblasts
in the spinner flask model, illustrating that shear stress alone is not enough to cause
the phenotypic shift discussed above.
Gelatin cross-linking affects HAC proliferation during micro-carrier expansion In Paper III, we investigated if the degree of gelatin cross-linking affects chondrocyte
proliferation on gelatin microcarriers over a two-week expansion period. We found a

Results and discussion
45
statistically significant impact of gelatin cross-linking on chondrocyte density during
the second week of culture. The less tightly cross-linked gelatin supported higher cell
densities compared to microcarriers with a stronger cross-link. In a recent study,
Sisson et al. demonstrated that viability and proliferation of MG63 cells, human
osteosarcoma cells with osteoblastic characteristics, on electrospun gelatin scaffolds is
significantly affected by the cross-linking method [Sisson et al. 2009]. They reported
that gelatin fiber morphology was influenced by the cross-linking agents, arguably
due to differences in reaction times and the amount of gelatin dissolved before com-
plete cross-linking. For the CultiSpher microcarriers investigated in Paper III, the
same cross-linking agent was used during manufacturing, but it is possible that differ-
ences in the manufacturing process (see Paper III for details) may have led to a dis-
similar release of gelatin during the cross-linking stage that in turn may have affected
scaffold morphology, and consecutively the proliferative behavior of the chondrocytes
in this study.
When compared to the others, the CultiSpher G microcarrier yielded the high-
est chondrocyte density after two weeks. This is in contrast to earlier findings, using
Chinese hamster ovary cells, where instead the CultiSpher S microcarrier gave the
highest cell fold increase compared to both the CultiSpher G and the GL counter-
parts [Kong et al. 1999]. The CultiSpher GLS microcarrier was not part of that
study. This discrepancy again highlights the importance of identifying suitable scaf-
folds for individual cell types. The overall cell yield in Paper III was modest, and the
cell culture parameters, such as stirring speed and serum concentrations, may be op-
timized further to encourage faster cell growth and limit aggregate formation [Malda
et al. 2003a; Gigout et al. 2005; Melero-Martin et al. 2006].
Pore diameter affects HAC proliferation during microcarrier ex-pansion In Paper III, we also found a statistically significant impact of microcarrier pore di-
ameter on chondrocyte density, again manifested during the second week of culture.
Gelatin matrices with an average internal pore diameter of 20 µm had higher cell
densities than matrices with a 30 µm pore diameter. We have found no comparable
experiments on human chondrocytes cultured on porous gelatin scaffolds with pore
diameters in the 20–30 µm range. Kong et al. however reported that the CultiSpher
G microcarrier yielded higher numbers of Chinese hamster ovary cells compared to
the CultiSpher GL counterpart [Kong et al. 1999]. Again, no such comparison for the

Biodegradable gelatin microcarriers in tissue engineering
46
tighter cross-linked S-type microcarriers, CultiSpher S and CultiSpher GLS, was
made by these authors. For other gelatin- or collagen-based matrices, rat chondro-
cyte densities were higher in scaffolds with pore diameters ranging between 50–
200 µm compared to 250–500 µm pore scaffolds [Lien et al. 2008], while osteogenic
MC3T3 cells adhere better to collagen–GAG based scaffolds with decreasing pore
diameters [O'Brien et al. 2005]. For chitosan scaffolds, porcine chondrocytes reached
higher densities in scaffolds with pore diameters of up to 120 µm [Griffon et al. 2006],
but pore diameter did not significantly influence rabbit chondrocyte proliferation on
chitosan hybrid materials [Yamane et al. 2007]. It has also been reported that porous
PLGA microcarriers with analogous dimensions to the CultiSpher GL microcarrier
supported greater DNA content with bovine chondrocytes compared to nonporous
microcarriers [Chung et al. 2008].
Summary The plausibility of using cell-seeded gelatin microcarriers in combination with PRP
has been addressed in three different in vitro studies (Papers I, II and IV), each yield-
ing results that demonstrate a capacity to support ECM formation in vitro. Still, the
proposed system has yet to be proven efficacious for tissue engineering of cartilage
and bone in vivo. Microcarrier characteristics, specifically internal pore diameter and
degree of cross-linking, have been shown to influence HAC expansion in terms of cell
density per gram dry weight gelatin over a two-week culture period in spinner flasks
(Paper III). In addition, the use of HDFs for cartilage and bone formation presents a
highly interesting cell source for parallel investigation (Paper II).

GENERAL DISCUSSION AND FUTURE PERSPECTIVES
The long-term clinical goal of this thesis is to enable cell delivery of human cells via
gelatin microcarriers for the purpose of regenerating cartilage and bone tissue. Sev-
eral experimental studies have been conducted, and the main results were discussed
in the previous chapter. The efforts presented here do not give any definitive answers.
Rather, it has raised additional scientific questions. This section addresses some of the
topics that encourage further examination.
On the efficacy of microcarriers for tissue engineering of cartilage Papers I, II and IV describe the plausibility of using macroporous gelatin microcarri-
ers for tissue engineering of cartilage. Though several important cartilage molecules
were detected in the synthesized ECM, true cartilage formation was not achieved.
The lack of uniform tissue formation, in combination with the absence of collagen
type II, may undeniably be an indication that the biomaterial scaffolds are not capa-
ble of supporting full cartilage generation, but it may also be attributed to inadequate
in vitro culture systems that lack important environmental cues, according to the ear-
lier discussion. Thus, the results from these in vitro studies are difficult to put into a
clinical perspective to date. It is not until evaluated in situ that the true efficacy of the
microcarrier and blood-derived glue system can be confirmed or rejected. Before
such in vivo studies are launched, a number of in vitro experiments are needed, some of
which will be described below.

Biodegradable gelatin microcarriers in tissue engineering
48
On the use of in vitro models There is a great need for a reliable, straightforward in vitro model for initial evalua-
tions of biomaterial characteristics, including biomaterial modifications, on chondro-
cyte metabolism. The dynamic centrifugation, as administered in Paper IV, did not
dramatically enhance chondrogenesis in vitro. Still, centrifugation at other intervals or
with other frequencies may have a positive influence on ECM synthesis in vitro. Fur-
ther development of the amplitude and frequency parameters could enable an easy
and affordable system for this type of screening, thus efforts towards establishing a
better centrifugation regime would be beneficial. Preferably, such regimes would run
with frequencies in the physiological range, 0.5–1 Hz [Lee and Bader 1997]. A fully
programmable centrifuge able to withstand the humidity and temperature in an in-
cubator would be highly advantageous for this purpose. Also, it is important not to
overestimate the dynamic nature of such regimes, and consider the reoccurrence in-
terval of such stimuli with utmost care. Dynamic compression can be administered
with the custom-built compression device mentioned previously in this thesis. If this
device is to become more useful for scientific purposes, further modifications are
needed, such as introducing sensors above and below the tissue-engineered constructs
to measure the applied forces. It is also possible that the cell culture media used dur-
ing expansion and three-dimensional culture could be optimized further [Malda et al.
2003a; Tallheden et al. 2005b; Akeda et al. 2006]. Finally, the influence of hypoxic
conditions on chondrocyte ECM production in a gelatin microcarrier model should
be investigated.
On the influence of microcarrier characteristics In Paper III, we showed that gelatin pore diameter and degree of cross-linking influ-
ence chondrocyte proliferation in vitro. The direct clinical relevance of this result is
perhaps limited. Rather, we have identified the microcarrier that will optimize the
chondrocyte expansion in vitro. Given that the CultiSpher G microcarrier yielded the
highest cell count during expansion, this microcarrier is the expected frontrunner for
future use. However, in vitro experiments studying ECM formation, rather than
chondrocyte proliferation, on these four microcarriers are needed. In order for such
an approach to be successful, quantitative analysis methods evaluating sulfate and
proline incorporation, GAG and collagen accumulation, and gelatin degradation

General discussion and future perspectives
49
rates in the respective microcarrier groups, should be added to the study design to
allow possible differences to be statistically confirmed.
By studying the SEM images in Paper III (Figure 1A), it is evident that the sur-
faces of individual microcarriers differ in terms of porosity and pore diameters. Simi-
larly, it is clear from the MTT pictures that not all microcarriers are populated dur-
ing the initial days (Figure 2A, Paper III). This is in agreement with data from Kong
et al., where only 14–47 % of the CultiSpher microcarriers used in that study were
populated by cells after 24 hours [Kong et al. 1999]. There may be a link between
these two observations and further investigations into this relationship, by means of
scanning electron microscopy on cell-seeded microcarriers during the first days post-
seeding, may elicit a deeper understanding of cell–biomaterial interactions during
adhesion, potentially enabling optimally designed scaffolds.
The experiments in Paper II were conducted on the CultiSpher GL micro-
carrier. As we have shown that microcarrier characteristics can influence chondro-
cyte behavior, an analogue screening of gelatin matrices for HDFs may be useful.
Furthermore, the introduction of growth factors to the gelatin scaffold may prove
beneficial to maintain or enhance the differentiated phenotype in situ. Previous non-
published efforts from our lab addressed the possibility of coupling chondrogenic
growth factors from the TGF-β super family to the gelatin matrices. Such strategies
should be verified in vitro to ensure that the growth factors are chemically active post-
binding and once released.
On the use of dermal fibroblasts in tissue engineering The fact that HDFs, a cell type that can be isolated with minimal discomfort and
expanded into great numbers, can change their phenotype offers a promising and
novel alternative cell source for tissue regeneration. Yet, one crucial question remains
regarding the stability of the induced phenotype – can these cells contribute to intrin-
sic tissue regeneration in situ? This outcome is difficult to predict. There have been
reports that the prolonged exposure to osteogenic factors may suppress bone growth
in vivo [Cheng et al. 2000]. Other studies have shown that HDFs are induced towards
a chondrocyte phenotype in the presence of demineralized bone in vitro [Mizuno and
Glowacki 1996]. Furthermore, the adhesion to bone ECM proteins such as
vitronectin and collagen type I, is enough to promote osteogenic differentiation of
BMSCs [Salasznyk et al. 2004]. These results could indicate that the in situ environ-
ment can, at least to some extent, help to establish and maintain a tissue specific

Biodegradable gelatin microcarriers in tissue engineering
50
phenotype. For cartilage the question is even more intriguing. As most cartilage re-
pair strategies today yield fibrocartilaginous tissues, even with all the physiological
cues in place, it would be truly groundbreaking if HDFs could regenerate intrinsic
cartilage formation in situ. The influence of environmental factors on chondrogenic
differentiation is interesting. The importance of these factors, including the synergis-
tic effects between chondrogenic induction media and biomechanical stimuli and/or
hypoxia, have been well established for adult stem cells [Angele et al. 2003; Angele et
al. 2004; Robins et al. 2005]. The incorporation of such strategies may enhance the
chondrogenic differentiation of HDFs as well [Mizuno and Glowacki 2005]. Regard-
less of the outcome, data from such experiments may prove helpful in the under-
standing of the mechanisms responsible for phenotype plasticity in human adult cells.
A major challenge in tissue engineering concerns the vascularization of large tis-
sue engineered constructs [Rouwkema et al. 2008]. Recent efforts by members of our
lab have investigated the ability of HDFs to differentiate towards an endothelial cell
like phenotype [Karlsson et al. 2009]. In addition, the differentiation of HDFs to-
wards chondrocyte, osteoblast, and adipocyte like phenotypes has been assessed
[Junker et al. 2009]. As has been discussed earlier, the efficacy of these cells to re-
generate tissue in vivo remains to be proven. Yet, the ease with which these cells can
be isolated and expanded in comparison offers a huge advantage and the use of
HDFs for tissue regeneration may revolutionize cell-based therapies as we know them
today.
As a final remark, the long-term clinical success of regenerative strategies does not
rely solely on the identification of the cells, scaffolds and environmental factors
needed to support tissue formation in vitro. The mechanisms behind any underlying
pathogenesis, such as osteoarthritis, must also be identified and addressed, in order to
restore permanent tissue function for the individual patients.

CONCLUSIONS
Based on the results from Papers I–IV, we conclude the following
• Gelatin microcarriers support adhesion, expansion and matrix formation of
HACs and HDFs
• Biological blood-derived glues can be used to encapsulate cell-seeded micro-
carriers in vitro
• PRP encapsulation enhances structural integrity of microcarrier constructs and
does not affect cartilage formation negatively
• Free-swelling conditions do not support true cartilage formation by HACs or
HDFs in vitro
• Dynamic centrifugation (for 16 min at 0.0125 Hz) every other day does not dra-
matically enhance or alter cartilage formation in vitro
• PRP encapsulation or dynamic centrifugation are not crucial for microcarrier
degradation in vitro
• HDFs seeded on gelatin microcarriers can produce cartilage-like tissue compo-
nents in free-swelling conditions under chondrogenic induction
• HDFs seeded on gelatin microcarriers can produce bone-like tissue components
in free-swelling conditions as well as in spinner flask systems under osteogenic
induction
• Flow-induced shear stress enhances matrix formation and mineralization and
leads to an upregulation of ALP activity in un-induced HDFs seeded on gelatin
microcarriers
• The degree of gelatin cross-linking affects microcarrier-based expansion of
HACS in a spinner flask system
• The average pore diameter affects microcarrier-based expansion of HACs in a
spinner flask system


ACKNOWLEDGEMENTS
Experimental research can be a tricky business, and I would not have been able to
complete this thesis without the support of my colleagues, friends and family.
I am deeply indebted to you all.
TO MY SUPERVISORS Gunnar Kratz – for giving me the opportunity to write this book. Your unique
ability to stay optimistic has been inspiring, especially when faced with the “norrländ-
ska svårmod” I possess. Thank you for your advice and not least your valuable time.
It’s been a privilege to discuss research ideas with you during these past years. I’ll
always remember the ever-weird anecdotes and words of wisdom – they were always
entertaining and, eventually, some of them actually made sense. I should have
known.
Pentti Tengvall – for introducing me to the field of biomaterials in the first place.
Thank you for your support and encouragement along the way, despite the change in
direction.
Jonas Wetterö – for always making time for me when I needed help. I am infinitely
grateful for your invaluable suggestions and comments – your efforts have undoubt-
edly improved my work. Thanks for the extensive discussions, whether research-re-
lated or not, and for all the laughs during the years. You have been a true mentor!
Keep on drumming.

Biodegradable gelatin microcarriers in tissue engineering
54
TO MY COLLEAGUES
The laboratory for reconstructive surgery To Anita Lönn for drilling me in the fine art of cell culture and keeping me on my
toes by asking continuous questions.
To Kristina Briheim for being so patient and supportive, and for always being
there with a contagious smile.
To my dear friend Camilla Fredriksson. You intimidated me quite a bit at first!
Since then you’ve become one of the most important people in my life. Thank you
for all our sessions (a lot of insight came out of them), the licorice tips, the great times,
the not so great times, and everything in between. You inspire me through your
courage, talent and resourcefulness. Thank you for always believing in me and for
giving me a solid push in the right direction when needed.
To Lisa Karlsson. You are one special lady! Life is never dull around you – it sel-
dom is around strong-headed and outspoken people like you – and it’s been a privi-
lege to share office space with you. Thank you for always lending me your ear, and
for deflating any irrational thought that reared its ugly head with laughter. I’m really
going to miss having you around.
To Johan Junker. Thanks for the everyday coffees and crosswords, the laughs and
the beers (foreign and domestic) and the hilarious movie nights (Taste the tea!). You
are a talented and fearless researcher and there is no doubt in my mind that you will
reach every academic goal you set for yourself. Also, you made me a hugging person
– I never thought I’d see the day..
To Fredrik Huss for helping me get started and offering valuable help along the
way.
To Pehr Sommar and Charlotte Ness for fruitful collaborations.
To the students and researchers that have crossed paths with me at the lab during the
years, Jonathan, Susanna, Mårten, Christopher, Erika, and Carin just to
name a few. I wish you all the best!
The immediate surroundings To my fellow PhD students – Tina Bendrik, Ulrika Nilsson, Git Johansson,
Caroline Skoglund, Karin Wermelin, Sa’ad Salim, Femke Lutgendorff,
Pernilla Eliasson, Sebastian Schultz, Henrik Åberg, Joakim Henriksson,
Calle Jonson, and the people I may have forgotten – for being there with a smile, a

Acknowledgements
55
listening ear, a helpful tip, or a coffee break when they were needed the most. You
are all brilliant!
To the entire staff at KEF – especially Kerstin Hagersten, Håkan Wiktander
and Pia Karlsson – for making the everyday life at work run so smoothly.
The various networks To Forum Scientium, especially Stefan Klintström, for providing a network
where help is only a phone call away and where opportunities for personal growth
and educational meetings are plentiful.
To the DOMFIL board of 2005-2006 – Ulrika, Git, Karin, Pernilla and Calle –
for a memorable year. I miss our discussions greatly.
To Alexandra Karström for getting me started and providing useful advice.
To Jonas Wetterö and Kajsa Holmgren Peterson for bringing me aboard the
CDIO-project – it’s been a great experience and I’m grateful for the opportunity.
Also, to the Engineering Biology students I’ve encountered during the years, for all
the fruitful discussions and blindsiding questions during the CDIO-projects – you’ve
kept me on my toes and taught me a lot of valuable lessons as well.
To the Materials in Medicine strategic research group for providing a rewarding
network. A special thank you to Agneta Askendal and Per Aspenberg for valu-
able help and support during the initial years.

Biodegradable gelatin microcarriers in tissue engineering
56
TO FAMILY AND FRIENDS To my beloved family for their never-ending love and support.
To mom and dad for always believing in me. I hope you know how much I value
your words of encouragement and advice. Also, on the occasion of your 50th
birthday – this, my first book, is dedicated to you, dad!
To my sister Anna for being so generous and giving and unbelievably cool.
To my incredibly strong and brave sister Maria for being my life-long inspiration.
To my brother Pär for showing me that the path ahead is what you make of it.
I love you all dearly!
To the Nilsson family for being wonderful relatives, I’ve seen far too little of you
during this thesis work.
To the Blom family for being so welcoming and generous. Thank you for your hos-
pitality and the occasional discussions on jazz and the family tree.
To Niklas Odelholm – the great love of my life! There is no way I could have
done this without you. Your unconditional encouragement and faith in me means
more to me than I can put in writing. I’m lucky to have found someone as wonderful
as you to share my life with. I love you. Hrrrumphhh..
To the rest of the Odelholm clan for welcoming me into your family (especially to
Hans and Maja for being such great critics).
To former and present members of Lusen Big Band – my second family. I don’t
think I can find the words to express how much you’ve meant to me, but I know for
certain that I wouldn’t be the same person without you. Thanks to you, my child-
hood dream of becoming a jazz vocalist came true. I also know that life would have
been a lot less fun if I hadn’t spent Thursdays, and countless late nights, with you
guys learning the trade. Who knew hauling PA systems and struggling with feedback
could be so life defining. Thank you for all the great times, including all the unbe-
lievably cool concerts we pulled off, but most of all for being great friends whom I’ll
cherish forever. I love you all. A special thanks to Andreas Johansson, Jonathan
Svensson, Simon Åström, Niklas Odelholm, Johan and Isabel

Acknowledgements
57
Fredriksson, Mattias Danielsson, Lina Vallin and David Westin – your
friendship means the world to me.
While I’m on the subject, a huge thank you and lots of love to the old friends that
helped me drum up the courage to get up on stage in the first place – the original
Soul Kitchen crew of 1998 – I almost didn’t make it up there that first time and now
I can’t imagine my life any other way. I owe you. Big time.
Also, to the miscellaneous crew of jazz musicians I’ve been fortunate to play with at
various venues – especially Johan Nilsson, Glenn Fransson, Daniel Forsell,
Jonathan Svensson, Simon Åström and Niklas Odelholm. Thanks for all the
fun and laughs during rehearsals and gigs (and for your patience when I couldn’t
keep up). You inspire me to constantly improve my skills so that they may one day
(though I doubt it) match yours.. “I’m not worthy, I’m not worthy..”
To Shintai Kempo Linköping for literally kicking my ass and pushing me harder than
I thought possible – the lesson actually stuck with me after a while, and it couldn’t
have happened at a better time. I’m a more harmonious person because of you guys,
and for that I thank you.
To my dear friends – Niklas Fridolf, Anna Rahmqvist, Emma Wester-
Johansson, Jan Bergström et al.. You contributed to this thesis by being great
friends! I have not been keeping in touch with you as much as I should have these
past years, and for that I am truly sorry. Time to make amends. I love you dearly!
FUNDING AND SUPPORT The work presented in this thesis has been carried out at the Laboratory for Recon-
structive Plastic Surgery, Division of Surgery, Department of Clinical and Experi-
mental Medicine, Faculty of Health Sciences, and in part at the Division of Applied
Physics, Department of Physics and Measurement Technology, Linköping Univer-
sity.
The Materials in Medicine strategic research area (Östergötland County Council and
Linköping University) and the Swedish Foundation for Strategic Research are ac-
knowledged for funding.


REFERENCES
Aigner T, Gebhard PM, Schmid E, Bau B, Harley V, Poschl E. 2003. SOX9 expression does not correlate with type II collagen expression in adult articular chondrocytes. Matrix Biol 22(4):363-72.
Akeda K, An HS, Okuma M, Attawia M, Miyamoto K, Thonar EJ, Lenz ME, Sah RL, Masuda K. 2006. Platelet-rich plasma stimulates porcine articular chondrocyte proliferation and matrix biosynthesis. Osteoarthritis Cartilage 14(12):1272-80.
Akens MK, Hurtig MB. 2005. Influence of species and anatomical location on chondrocyte expansion. BMC Musculoskelet Disord 6:23.
Alsalameh S, Amin R, Gemba T, Lotz M. 2004. Identification of mesenchymal progenitor cells in normal and osteoarthritic human articular cartilage. Arthritis Rheum 50(5):1522-32.
Angele P, Schumann D, Angele M, Kinner B, Englert C, Hente R, Fuchtmeier B, Nerlich M, Neumann C, Kujat R. 2004. Cyclic, mechanical compression enhances chondrogenesis of mesenchymal progenitor cells in tissue engineering scaffolds. Biorheology 41(3-4):335-46.
Angele P, Yoo JU, Smith C, Mansour J, Jepsen KJ, Nerlich M, Johnstone B. 2003. Cyclic hydrostatic pressure enhances the chondrogenic phenotype of human mesenchymal progenitor cells differentiated in vitro. J Orthop Res 21(3):451-7.
Awad HA, Halvorsen YD, Gimble JM, Guilak F. 2003. Effects of transforming growth factor beta1 and dexamethasone on the growth and chondrogenic differentiation of adipose-derived stromal cells. Tissue Eng 9(6):1301-12.
Barbero A, Grogan S, Schafer D, Heberer M, Mainil-Varlet P, Martin I. 2004. Age related changes in human articular chondrocyte yield, proliferation and post-expansion chondrogenic capacity. Osteoarthritis Cartilage 12(6):476-84.
Barker MK, Seedhom BB. 1997. Articular cartilage deformation under physiological cyclic loading--apparatus and measurement technique. J Biomech 30(4):377-81.
Bartsch G, Yoo JJ, De Coppi P, Siddiqui MM, Schuch G, Pohl HG, Fuhr J, Perin L, Soker S, Atala A. 2005. Propagation, expansion, and multilineage differentiation of human somatic stem cells from dermal progenitors. Stem Cells Dev 14(3):337-48.

Biodegradable gelatin microcarriers in tissue engineering
60
Benya PD, Shaffer JD. 1982. Dedifferentiated chondrocytes reexpress the differentiated collagen phenotype when cultured in agarose gels. Cell 30(1):215-24.
Bills CE, Eisenberg H, Pallante SL. 1971. Complexes of organic acids with calcium phosphate: the von Kossa stain as a clue to the composition of bone mineral. Johns Hopkins Med J 128(4):194-207.
Blum B, Benvenisty N. 2008. The tumorigenicity of human embryonic stem cells. Adv Cancer Res 100:133-58.
Blunk T, Sieminski AL, Gooch KJ, Courter DL, Hollander AP, Nahir AM, Langer R, Vunjak-Novakovic G, Freed LE. 2002. Differential effects of growth factors on tissue-engineered cartilage. Tissue Eng 8(1):73-84.
Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson O, Peterson L. 1994. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N Engl J Med 331(14):889-95.
Brittberg M, Peterson L, Sjogren-Jansson E, Tallheden T, Lindahl A. 2003. Articular cartilage engineering with autologous chondrocyte transplantation. A review of recent developments. J Bone Joint Surg Am 85-A Suppl 3:109-15.
Buckwalter JA, Mankin HJ. 1998. Articular cartilage: tissue design and chondrocyte-matrix interactions. Instr Course Lect 47:477-86.
Buschmann MD, Gluzband YA, Grodzinsky AJ, Hunziker EB. 1995. Mechanical compression modulates matrix biosynthesis in chondrocyte/agarose culture. J Cell Sci 108 ( Pt 4):1497-508.
Chen R, Curran SJ, Curran JM, Hunt JA. 2006. The use of poly(l-lactide) and RGD modified microspheres as cell carriers in a flow intermittency bioreactor for tissue engineering cartilage. Biomaterials 27(25):4453-60.
Cheng H, Jiang W, Phillips FM, Haydon RC, Peng Y, Zhou L, Luu HH, An N, Breyer B, Vanichakarn P and others. 2003. Osteogenic activity of the fourteen types of human bone morphogenetic proteins (BMPs). J Bone Joint Surg Am 85-A(8):1544-52.
Cheng SL, Lai CF, Fausto A, Chellaiah M, Feng X, McHugh KP, Teitelbaum SL, Civitelli R, Hruska KA, Ross FP and others. 2000. Regulation of alphaVbeta3 and alphaVbeta5 integrins by dexamethasone in normal human osteoblastic cells. J Cell Biochem 77(2):265-76.
Cheng SL, Yang JW, Rifas L, Zhang SF, Avioli LV. 1994. Differentiation of human bone marrow osteogenic stromal cells in vitro: induction of the osteoblast phenotype by dexamethasone. Endocrinology 134(1):277-86.
Chung HJ, Kim IK, Kim TG, Park TG. 2008. Highly open porous biodegradable microcarriers: in vitro cultivation of chondrocytes for injectable delivery. Tissue Eng Part A 14(5):607-15.
Curtis A, Wilkinson C. 1997. Topographical control of cells. Biomaterials 18(24):1573-83.
Darling EM, Athanasiou KA. 2005. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. J Orthop Res 23(2):425-32.
Datta N, Pham QP, Sharma U, Sikavitsas VI, Jansen JA, Mikos AG. 2006. In vitro generated extracellular matrix and fluid shear stress synergistically enhance 3D osteoblastic differentiation. Proc Natl Acad Sci U S A 103(8):2488-93.
Davisson T, Kunig S, Chen A, Sah R, Ratcliffe A. 2002. Static and dynamic compression modulate matrix metabolism in tissue engineered cartilage. J Orthop Res 20(4):842-8.

References
61
Denizot F, Lang R. 1986. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J Immunol Methods 89(2):271-7.
Denker AE, Nicoll SB, Tuan RS. 1995. Formation of cartilage-like spheroids by micromass cultures of murine C3H10T1/2 cells upon treatment with transforming growth factor-beta 1. Differentiation 59(1):25-34.
Domm C, Schunke M, Christesen K, Kurz B. 2002. Redifferentiation of dedifferentiated bovine articular chondrocytes in alginate culture under low oxygen tension. Osteoarthritis Cartilage 10(1):13-22.
Eisenberg LM, Eisenberg CA. 2003. Stem cell plasticity, cell fusion, and transdifferentiation. Birth Defects Res C Embryo Today 69(3):209-18.
Erggelet C, Sittinger M, Lahm A. 2003. The arthroscopic implantation of autologous chondrocytes for the treatment of full-thickness cartilage defects of the knee joint. Arthroscopy 19(1):108-10.
Eyrich D, Brandl F, Appel B, Wiese H, Maier G, Wenzel M, Staudenmaier R, Goepferich A, Blunk T. 2007. Long-term stable fibrin gels for cartilage engineering. Biomaterials 28(1):55-65.
Fan H, Hu Y, Qin L, Li X, Wu H, Lv R. 2006. Porous gelatin-chondroitin-hyaluronate tri-copolymer scaffold containing microspheres loaded with TGF-beta1 induces differentiation of mesenchymal stem cells in vivo for enhancing cartilage repair. J Biomed Mater Res A 77(4):785-94.
Fermor B, Christensen SE, Youn I, Cernanec JM, Davies CM, Weinberg JB. 2007. Oxygen, nitric oxide and articular cartilage. Eur Cell Mater 13:56-65; discussion 65.
Freed LE, Langer R, Martin I, Pellis NR, Vunjak-Novakovic G. 1997. Tissue engineering of cartilage in space. Proc Natl Acad Sci U S A 94(25):13885-90.
Frondoza C, Sohrabi A, Hungerford D. 1996. Human chondrocytes proliferate and produce matrix components in microcarrier suspension culture. Biomaterials 17(9):879-88.
Gaissmaier C, Fritz J, Krackhardt T, Flesch I, Aicher WK, Ashammakhi N. 2005. Effect of human platelet supernatant on proliferation and matrix synthesis of human articular chondrocytes in monolayer and three-dimensional alginate cultures. Biomaterials 26(14):1953-60.
Gardner RL. 2007. Stem cells and regenerative medicine: principles, prospects and problems. C R Biol 330(6-7):465-73.
Giannoni P, Crovace A, Malpeli M, Maggi E, Arbico R, Cancedda R, Dozin B. 2005. Species variability in the differentiation potential of in vitro-expanded articular chondrocytes restricts predictive studies on cartilage repair using animal models. Tissue Eng 11(1-2):237-48.
Gigout A, Jolicoeur M, Buschmann MD. 2005. Low calcium levels in serum-free media maintain chondrocyte phenotype in monolayer culture and reduce chondrocyte aggregation in suspension culture. Osteoarthritis Cartilage 13(11):1012-24.
Goessl A, Redl H. 2005. Optimized thrombin dilution protocol for a slowly setting fibrin sealant in surgery. Eur Surg 37(1):43-51.
Grad S, Zhou L, Gogolewski S, Alini M. 2003. Chondrocytes seeded onto poly (L/DL-lactide) 80%/20% porous scaffolds: a biochemical evaluation. J Biomed Mater Res A 66(3):571-9.

Biodegradable gelatin microcarriers in tissue engineering
62
Griffon DJ, Sedighi MR, Schaeffer DV, Eurell JA, Johnson AL. 2006. Chitosan scaffolds: interconnective pore size and cartilage engineering. Acta Biomater 2(3):313-20.
Grigoriadis AE, Heersche JN, Aubin JE. 1988. Differentiation of muscle, fat, cartilage, and bone from progenitor cells present in a bone-derived clonal cell population: effect of dexamethasone. J Cell Biol 106(6):2139-51.
Gunatillake PA, Adhikari R. 2003. Biodegradable synthetic polymers for tissue engineering. Eur Cell Mater 5:1-16; discussion 16.
Haberhauer M, Zernia G, Deiwick A, Pösel C, Bader A, Huster D, Schulz R. 2008. Cartilage Tissue Engineering in Plasma and Whole Blood Scaffolds. Adv Mater Deerfield 20(11):2061-2067.
Hayes AJ, Hall A, Brown L, Tubo R, Caterson B. 2007. Macromolecular organization and in vitro growth characteristics of scaffold-free neocartilage grafts. J Histochem Cytochem 55(8):853-66.
Hendriks J, Riesle J, Vanblitterswijk CA. 2006. Effect of stratified culture compared to confluent culture in monolayer on proliferation and differentiation of human articular chondrocytes. Tissue Eng 12(9):2397-405.
Hodge WA, Fijan RS, Carlson KL, Burgess RG, Harris WH, Mann RW. 1986. Contact pressures in the human hip joint measured in vivo. Proc Natl Acad Sci U S A 83(9):2879-83.
Höftprotesregistret. 2008. Svenska Höftprotesregistret Årsrapport 2007. www.jru.orthop.gu.se ISSN 1654-5982.
Holtorf HL, Jansen JA, Mikos AG. 2005. Flow perfusion culture induces the osteoblastic differentiation of marrow stroma cell-scaffold constructs in the absence of dexamethasone. J Biomed Mater Res A 72(3):326-34.
Holtzer H, Abbott J, Lash J, Holtzer S. 1960. The Loss of Phenotypic Traits by Differentiated Cells in Vitro, I. Dedifferentiation of Cartilage Cells. Proc Natl Acad Sci U S A 46(12):1533-42.
Homminga GN, Buma P, Koot HW, van der Kraan PM, van den Berg WB. 1993. Chondrocyte behavior in fibrin glue in vitro. Acta Orthop Scand 64(4):441-5.
Hong Y, Gao C, Xie Y, Gong Y, Shen J. 2005. Collagen-coated polylactide microspheres as chondrocyte microcarriers. Biomaterials 26(32):6305-13.
Huss FR, Junker JP, Johnson H, Kratz G. 2007. Macroporous gelatine spheres as culture substrate, transplantation vehicle, and biodegradable scaffold for guided regeneration of soft tissues. In vivo study in nude mice. J Plast Reconstr Aesthet Surg 60(5):543-55.
Inoue H, Hiasa K, Samma Y, Nakamura O, Sakuda M, Iwamoto M, Suzuki F, Kato Y. 1990. Stimulation of proteoglycan and DNA syntheses in chondrocytes by centrifugation. J Dent Res 69(9):1560-3.
Jahoda CA, Whitehouse J, Reynolds AJ, Hole N. 2003. Hair follicle dermal cells differentiate into adipogenic and osteogenic lineages. Exp Dermatol 12(6):849-59.
Jaiswal N, Haynesworth SE, Caplan AI, Bruder SP. 1997. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J Cell Biochem 64(2):295-312.
Jaiswal RK, Jaiswal N, Bruder SP, Mbalaviele G, Marshak DR, Pittenger MF. 2000. Adult human mesenchymal stem cell differentiation to the osteogenic or

References
63
adipogenic lineage is regulated by mitogen-activated protein kinase. J Biol Chem 275(13):9645-52.
Jakob M, Demarteau O, Schafer D, Hintermann B, Dick W, Heberer M, Martin I. 2001. Specific growth factors during the expansion and redifferentiation of adult human articular chondrocytes enhance chondrogenesis and cartilaginous tissue formation in vitro. J Cell Biochem 81(2):368-77.
Junker JP, Sommar P, Skog M, Johnson H, Kratz G. 2009. Adipogenic, Chondrogenic and Osteogenic Differentiation of Clonally Derived Human Dermal Fibroblasts. Cells Tissues Organs.
Junqueira LC, Bignolas G, Brentani RR. 1979. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem J 11(4):447-55.
Kang SW, La WG, Kim BS. 2009. Open Macroporous Poly(lactic-co-glycolic Acid) Microspheres as an Injectable Scaffold for Cartilage Tissue Engineering. J Biomater Sci Polym Ed 20(3):399-409.
Kang SW, Yoo SP, Kim BS. 2007. Effect of chondrocyte passage number on histological aspects of tissue-engineered cartilage. Biomed Mater Eng 17(5):269-76.
Karageorgiou V, Kaplan D. 2005. Porosity of 3D biomaterial scaffolds and osteogenesis. Biomaterials 26(27):5474-91.
Karlsson LK, Junker JP, Grenegard M, Kratz G. 2009. Human Dermal Fibroblasts: A Potential Cell Source for Endothelialization of Vascular Grafts. Ann Vasc Surg.
Keeney M, Pandit A. 2009. The osteochondral junction and its repair via bi-phasic tissue engineering scaffolds. Tissue Eng Part B Rev 15(1):55-73.
Kelly DJ, Prendergast PJ. 2006. Prediction of the optimal mechanical properties for a scaffold used in osteochondral defect repair. Tissue Eng 12(9):2509-19.
Knäprotesregistret. 2008. Svenska Knäprotesregistret Årsrapport 2008 www.knee.se ISBN 978-91-976019-9-3.
Kong D, Chen M, Gentz R, Zhang J. 1999. Cell growth and protein formation on various microcarriers. Cytotechnology 29(2):151-158.
Kreke MR, Sharp LA, Lee YW, Goldstein AS. 2008. Effect of intermittent shear stress on mechanotransductive signaling and osteoblastic differentiation of bone marrow stromal cells. Tissue Eng Part A 14(4):529-37.
Landesberg R, Roy M, Glickman RS. 2000. Quantification of growth factor levels using a simplified method of platelet-rich plasma gel preparation. J Oral Maxillofac Surg 58(3):297-300; discussion 300-1.
Langer R, Vacanti JP. 1993. Tissue engineering. Science 260(5110):920-6. Lao L, Tan H, Wang Y, Gao C. 2008. Chitosan modified poly(L-lactide)
microspheres as cell microcarriers for cartilage tissue engineering. Colloids Surf B Biointerfaces 66(2):218-25.
Leboy PS, Sullivan TA, Nooreyazdan M, Venezian RA. 1997. Rapid chondrocyte maturation by serum-free culture with BMP-2 and ascorbic acid. J Cell Biochem 66(3):394-403.
Lee DA, Bader DL. 1997. Compressive strains at physiological frequencies influence the metabolism of chondrocytes seeded in agarose. J Orthop Res 15(2):181-8.

Biodegradable gelatin microcarriers in tissue engineering
64
Lee DA, Noguchi T, Knight MM, O'Donnell L, Bentley G, Bader DL. 1998. Response of chondrocyte subpopulations cultured within unloaded and loaded agarose. J Orthop Res 16(6):726-33.
Lee SJ, Lee YM, Han CW, Lee HB, Khang G. 1994. Response of human chondrocytes on polymer surfaces with diffferent micropore sizes for tissue-engineered cartilage. J Appl Polym Sci 92(5):2784-2790.
Li WJ, Tuli R, Okafor C, Derfoul A, Danielson KG, Hall DJ, Tuan RS. 2005. A three-dimensional nanofibrous scaffold for cartilage tissue engineering using human mesenchymal stem cells. Biomaterials 26(6):599-609.
Lian JB, Stein GS. 1992. Concepts of osteoblast growth and differentiation: basis for modulation of bone cell development and tissue formation. Crit Rev Oral Biol Med 3(3):269-305.
Lian JB, Stein GS. 1995. Development of the osteoblast phenotype: molecular mechanisms mediating osteoblast growth and differentiation. Iowa Orthop J 15:118-40.
Lien SM, Ko LY, Huang TJ. 2008. Effect of pore size on ECM secretion and cell growth in gelatin scaffold for articular cartilage tissue engineering. Acta Biomater.
Lorenz K, Sicker M, Schmelzer E, Rupf T, Salvetter J, Schulz-Siegmund M, Bader A. 2008. Multilineage differentiation potential of human dermal skin-derived fibroblasts. Exp Dermatol 17(11):925-32.
Lysaght MJ, Crager J. 2009. Origins. Tissue Eng Part A 15(7):1449-50. Maeda S, Nishida J, Sato T, Inomata Y, Shimamura T, Horiuchi S. 2005. Changes
in microstructure and gene expression of articular chondrocytes cultured in a tube under mechanical stress. Osteoarthritis Cartilage 13(2):154-61.
Maeda S, Yoshida M, Hirano H, Horiuchi S. 2001. Effects of mechanical stimulation on gene expression of articular chondrocytes in polylayer culture. Tohoku J Exp Med 193(4):301-10.
Magnusson P, Farley JR. 2002. Differences in sialic acid residues among bone alkaline phosphatase isoforms: a physical, biochemical, and immunological characterization. Calcif Tissue Int 71(6):508-18.
Malafaya PB, Silva GA, Reis RL. 2007. Natural-origin polymers as carriers and scaffolds for biomolecules and cell delivery in tissue engineering applications. Adv Drug Deliv Rev 59(4-5):207-33.
Malda J, Kreijveld E, Temenoff JS, van Blitterswijk CA, Riesle J. 2003a. Expansion of human nasal chondrocytes on macroporous microcarriers enhances redifferentiation. Biomaterials 24(28):5153-61.
Malda J, van Blitterswijk CA, Grojec M, Martens DE, Tramper J, Riesle J. 2003b. Expansion of bovine chondrocytes on microcarriers enhances redifferentiation. Tissue Eng 9(5):939-48.
Malda J, van Blitterswijk CA, van Geffen M, Martens DE, Tramper J, Riesle J. 2004. Low oxygen tension stimulates the redifferentiation of dedifferentiated adult human nasal chondrocytes. Osteoarthritis Cartilage 12(4):306-13.
Malicev E, Radosavljevic D, Velikonja NK. 2007. Fibrin gel improved the spatial uniformity and phenotype of human chondrocytes seeded on collagen scaffolds. Biotechnol Bioeng 96(2):364-70.
Marchand C, Rivard GE, Sun J, Hoemann CD. 2009. Solidification mechanisms of chitosan-glycerol phosphate/blood implant for articular cartilage repair. Osteoarthritis Cartilage 17(7):950-7.

References
65
Martin I, Miot S, Barbero A, Jakob M, Wendt D. 2007. Osteochondral tissue engineering. J Biomech 40(4):750-65.
Marx RE, Carlson ER, Eichstaedt RM, Schimmele SR, Strauss JE, Georgeff KR. 1998. Platelet-rich plasma: Growth factor enhancement for bone grafts. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 85(6):638-46.
Mauck RL, Nicoll SB, Seyhan SL, Ateshian GA, Hung CT. 2003. Synergistic action of growth factors and dynamic loading for articular cartilage tissue engineering. Tissue Eng 9(4):597-611.
Mauck RL, Soltz MA, Wang CC, Wong DD, Chao PH, Valhmu WB, Hung CT, Ateshian GA. 2000. Functional tissue engineering of articular cartilage through dynamic loading of chondrocyte-seeded agarose gels. J Biomech Eng 122(3):252-60.
McCarrel T, Fortier L. 2009. Temporal growth factor release from platelet-rich plasma, trehalose lyophilized platelets, and bone marrow aspirate and their effect on tendon and ligament gene expression. J Orthop Res.
Meinel L, Karageorgiou V, Fajardo R, Snyder B, Shinde-Patil V, Zichner L, Kaplan D, Langer R, Vunjak-Novakovic G. 2004. Bone tissue engineering using human mesenchymal stem cells: effects of scaffold material and medium flow. Ann Biomed Eng 32(1):112-22.
Melero-Martin JM, Dowling MA, Smith M, Al-Rubeai M. 2006. Expansion of chondroprogenitor cells on macroporous microcarriers as an alternative to conventional monolayer systems. Biomaterials 27(15):2970-9.
Millward-Sadler SJ, Salter DM. 2004. Integrin-dependent signal cascades in chondrocyte mechanotransduction. Ann Biomed Eng 32(3):435-46.
Mizuno S, Glowacki J. 1996. Chondroinduction of human dermal fibroblasts by demineralized bone in three-dimensional culture. Exp Cell Res 227(1):89-97.
Mizuno S, Glowacki J. 2005. Low oxygen tension enhances chondroinduction by demineralized bone matrix in human dermal fibroblasts in vitro. Cells Tissues Organs 180(3):151-8.
Mosmann T. 1983. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 65(1-2):55-63.
Mukherjee DP, Smith DF, Rogers SH, Emmanual JE, Jadin KD, Hayes BK. 2008. Effect of 3D-microstructure of bioabsorbable PGA:TMC scaffolds on the growth of chondrogenic cells. J Biomed Mater Res B Appl Biomater.
Nakagawa Y, Shimizu K, Hamamoto T, Kotani S, Yamamuro T. 1993. Electron microscopy of calcification during high-density suspension culture of chondrocytes. Calcif Tissue Int 53(2):127-34.
O'Brien FJ, Harley BA, Yannas IV, Gibson LJ. 2005. The effect of pore size on cell adhesion in collagen-GAG scaffolds. Biomaterials 26(4):433-41.
Oyajobi BO, Frazer A, Hollander AP, Graveley RM, Xu C, Houghton A, Hatton PV, Russell RG, Stringer BM. 1998. Expression of type X collagen and matrix calcification in three-dimensional cultures of immortalized temperature-sensitive chondrocytes derived from adult human articular cartilage. J Bone Miner Res 13(3):432-42.
Peterson L, Minas T, Brittberg M, Nilsson A, Sjögren-Jansson E, Lindahl A. 2000. Two- to 9-year outcome after autologous chondrocyte transplantation of the knee. Clin Orthop Relat Res(374):212-34.

Biodegradable gelatin microcarriers in tissue engineering
66
Phi-Wilson J, Harvey J, Goix P, O'Neill R. 2001. A technology for the rapid acquisition of cell number and viability. Am Biotechnol Lab 19(6):34-36.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. 1999. Multilineage potential of adult human mesenchymal stem cells. Science 284(5411):143-7.
Poole CA. 1997. Articular cartilage chondrons: form, function and failure. J Anat 191 ( Pt 1):1-13.
Ramage L, Nuki G, Salter DM. 2009. Signalling cascades in mechanotransduction: cell-matrix interactions and mechanical loading. Scand J Med Sci Sports.
Redman SN, Oldfield SF, Archer CW. 2005. Current strategies for articular cartilage repair. Eur Cell Mater 9:23-32; discussion 23-32.
Robins JC, Akeno N, Mukherjee A, Dalal RR, Aronow BJ, Koopman P, Clemens TL. 2005. Hypoxia induces chondrocyte-specific gene expression in mesenchymal cells in association with transcriptional activation of Sox9. Bone 37(3):313-22.
Rouwkema J, Rivron NC, van Blitterswijk CA. 2008. Vascularization in tissue engineering. Trends Biotechnol 26(8):434-41.
Sah RL, Kim YJ, Doong JY, Grodzinsky AJ, Plaas AH, Sandy JD. 1989. Biosynthetic response of cartilage explants to dynamic compression. J Orthop Res 7(5):619-36.
Salasznyk RM, Williams WA, Boskey A, Batorsky A, Plopper GE. 2004. Adhesion to Vitronectin and Collagen I Promotes Osteogenic Differentiation of Human Mesenchymal Stem Cells. J Biomed Biotechnol 2004(1):24-34.
Schulz RM, Bader A. 2007. Cartilage tissue engineering and bioreactor systems for the cultivation and stimulation of chondrocytes. Eur Biophys J 36(4-5):539-68.
Schumacher BL, Block JA, Schmid TM, Aydelotte MB, Kuettner KE. 1994. A novel proteoglycan synthesized and secreted by chondrocytes of the superficial zone of articular cartilage. Arch Biochem Biophys 311(1):144-52.
Seeger FH, Tonn T, Krzossok N, Zeiher AM, Dimmeler S. 2007. Cell isolation procedures matter: a comparison of different isolation protocols of bone marrow mononuclear cells used for cell therapy in patients with acute myocardial infarction. Eur Heart J 28(6):766-72.
Silver IA. 1975. Measurement of pH and ionic composition of pericellular sites. Philos Trans R Soc Lond B Biol Sci 271(912):261-72.
Sisson K, Zhang C, Farach-Carson MC, Chase DB, Rabolt JF. 2009. Evaluation of Cross-Linking Methods for Electrospun Gelatin on Cell Growth and Viability. Biomacromolecules.
Spiteri CG, Pilliar RM, Kandel RA. 2006. Substrate porosity enhances chondrocyte attachment, spreading, and cartilage tissue formation in vitro. J Biomed Mater Res A 78(4):676-83.
Stenhamre H, Slynarski K, Petren C, Tallheden T, Lindahl A. 2008. Topographic variation in redifferentiation capacity of chondrocytes in the adult human knee joint. Osteoarthritis Cartilage 16(11):1356-62.
Sullivan TA, Uschmann B, Hough R, Leboy PS. 1994. Ascorbate modulation of chondrocyte gene expression is independent of its role in collagen secretion. J Biol Chem 269(36):22500-6.

References
67
Tallheden T, Bengtsson C, Brantsing C, Sjogren-Jansson E, Carlsson L, Peterson L, Brittberg M, Lindahl A. 2005a. Proliferation and differentiation potential of chondrocytes from osteoarthritic patients. Arthritis Res Ther 7(3):R560-8.
Tallheden T, van der Lee J, Brantsing C, Mansson JE, Sjogren-Jansson E, Lindahl A. 2005b. Human serum for culture of articular chondrocytes. Cell Transplant 14(7):469-79.
Thissen H, Chang KY, Tebb TA, Tsai WB, Glattauer V, Ramshaw JA, Werkmeister JA. 2006. Synthetic biodegradable microparticles for articular cartilage tissue engineering. J Biomed Mater Res A 77(3):590-8.
von der Mark K, Gauss V, von der Mark H, Muller P. 1977. Relationship between cell shape and type of collagen synthesised as chondrocytes lose their cartilage phenotype in culture. Nature 267(5611):531-2.
Waldman SD, Spiteri CG, Grynpas MD, Pilliar RM, Kandel RA. 2003. Long-term intermittent shear deformation improves the quality of cartilaginous tissue formed in vitro. J Orthop Res 21(4):590-6.
Waldman SD, Spiteri CG, Grynpas MD, Pilliar RM, Kandel RA. 2004. Long-term intermittent compressive stimulation improves the composition and mechanical properties of tissue-engineered cartilage. Tissue Eng 10(9-10):1323-31.
Warejcka DJ, Harvey R, Taylor BJ, Young HE, Lucas PA. 1996. A population of cells isolated from rat heart capable of differentiating into several mesodermal phenotypes. J Surg Res 62(2):233-42.
Wehrli BM, Huang W, De Crombrugghe B, Ayala AG, Czerniak B. 2003. Sox9, a master regulator of chondrogenesis, distinguishes mesenchymal chondrosarcoma from other small blue round cell tumors. Hum Pathol 34(3):263-9.
Weibrich G, Kleis WK, Hafner G, Hitzler WE. 2002. Growth factor levels in platelet-rich plasma and correlations with donor age, sex, and platelet count. J Craniomaxillofac Surg 30(2):97-102.
Wernike E, Li Z, Alini M, Grad S. 2008. Effect of reduced oxygen tension and long-term mechanical stimulation on chondrocyte-polymer constructs. Cell Tissue Res 331(2):473-83.
Williams DF. 1987. Definitions in Biomaterials. Amsterdam. Elsevier. Williams DF. 2008. On the mechanisms of biocompatibility. Biomaterials
29(20):2941-53. Wilson CJ, Clegg RE, Leavesley DI, Pearcy MJ. 2005. Mediation of biomaterial-cell
interactions by adsorbed proteins: a review. Tissue Eng 11(1-2):1-18. Wolff DA, Stevenson S, Goldberg VM. 1992. S-100 protein immunostaining
identifies cells expressing a chondrocytic phenotype during articular cartilage repair. J Orthop Res 10(1):49-57.
Wu H, Wan Y, Cao X, Wu Q. 2008. Proliferation of chondrocytes on porous poly(DL-lactide)/chitosan scaffolds. Acta Biomater 4(1):76-87.
Wu W, Chen F, Liu Y, Ma Q, Mao T. 2007. Autologous injectable tissue-engineered cartilage by using platelet-rich plasma: experimental study in a rabbit model. J Oral Maxillofac Surg 65(10):1951-7.
Yamane S, Iwasaki N, Kasahara Y, Harada K, Majima T, Monde K, Nishimura S, Minami A. 2007. Effect of pore size on in vitro cartilage formation using chitosan-based hyaluronic acid hybrid polymer fibers. J Biomed Mater Res A 81(3):586-93.

Biodegradable gelatin microcarriers in tissue engineering
68
Yang KG, Saris DB, Geuze RE, van Rijen MH, van der Helm YJ, Verbout AJ, Creemers LB, Dhert WJ. 2006. Altered in vitro chondrogenic properties of chondrocytes harvested from unaffected cartilage in osteoarthritic joints. Osteoarthritis Cartilage 14(6):561-70.
Yoo JU, Barthel TS, Nishimura K, Solchaga L, Caplan AI, Goldberg VM, Johnstone B. 1998. The chondrogenic potential of human bone-marrow-derived mesenchymal progenitor cells. J Bone Joint Surg Am 80(12):1745-57.
Young HE, Mancini ML, Wright RP, Smith JC, Black AC, Jr., Reagan CR, Lucas PA. 1995. Mesenchymal stem cells reside within the connective tissues of many organs. Dev Dyn 202(2):137-44.
Young HE, Steele TA, Bray RA, Hudson J, Floyd JA, Hawkins K, Thomas K, Austin T, Edwards C, Cuzzourt J and others. 2001. Human reserve pluripotent mesenchymal stem cells are present in the connective tissues of skeletal muscle and dermis derived from fetal, adult, and geriatric donors. Anat Rec 264(1):51-62.
Zeugolis DI, Paul GR, Attenburrow G. 2009. Cross-linking of extruded collagen fibers--a biomimetic three-dimensional scaffold for tissue engineering applications. J Biomed Mater Res A 89(4):895-908.
Zheng MH, Willers C, Kirilak L, Yates P, Xu J, Wood D, Shimmin A. 2007. Matrix-induced autologous chondrocyte implantation (MACI): biological and histological assessment. Tissue Eng 13(4):737-46.
Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P, Hedrick MH. 2002. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell 13(12):4279-95.
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP, Hedrick MH. 2001. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng 7(2):211-28.
Copyright © 2022 FDOKUMEN




















