
Rapid development of colitis in NSAID-treated IL10–deficient mice
Upload
independentCategory
view
0download
0
CELLULAR IMMUNOLOGY 170, 41–53 (1996)ARTICLE NO. 0132
Antigen-Specific IL-4- and IL-10-Secreting CD4/ LymphocytesIncrease in Vivo Susceptibility to Trypanosoma cruzi Infection
LAURA CARDELLINI BARBOSA DE OLIVEIRA, MARIA ALICIA CUROTTO DE LAFAILLE,GLORIA M. COLLET DE ARAUJO LIMA, AND ISES DE ALMEIDA ABRAHAMSOHN1
Departamento de Imunologia, Instituto de Ciencias Biomedicas, Universidade de Sao Paulo, Sao Paulo, 05508-900 SP, Brazil
Received January 25, 1996; accepted February 1, 1996
of parasite-specific host cells toward a Th2 pheno-type. q 1996 Academic Press, Inc.Control of macrophage parasiticidal function by
treatment with recombinant cytokines or their neu-tralizing antibodies modifies the severity of experi-
INTRODUCTIONmental Trypanosoma cruzi infections. However, sofar, no direct in vivo evidence has demonstrated
Trypanosoma cruzi is a dygenetic protozoan whichchanges in disease outcome after altering the initialinfects several kinds of mammals and is the etiologicratios of parasite-specific IFN-g and IL-10/IL-4-se-agent of Chagas’ disease in humans. Multiple genescretor cells in secondary lymphoid organs. To thisoutside the H-2 locus determine the outcome of infec-end, a population of predominantly CD4/ parasite-tion in the mouse and yet a spectrum of resistanceAg-reactive, IL-4- and IL-10-secreting T lymphocytespatterns is found among inbred strains (1). C3H andderived from T. cruzi-immunized mice was adop-A mice generally rank as the most susceptible, whereastively transferred to naive recipients. ComparedC57BL/6 and SJL mice are more resistant, althoughwith cell responses from normal mice, spleen cells of
uninfected recipients proliferated significantly to T. such distinctions may eventually vary depending oncruzi Ag and produced much greater amounts of IL- the strain of T. cruzi studied (2). Efficient control of4 and IL-10; lower IFN-g levels and increased IL-4/ parasite load and host survival rely on T-cell-mediatedIL-10 levels were induced by Con A stimulation. Re- immunity via T-helper-cell-dependent protective Ab re-cipient mice challenged with T. cruzi presented over- sponses and macrophage activation for intracellularwhelming tissue and blood parasitemia and early killing of the protozoan. In addition, class I-dependentdeath, contrasting with typically resistant controls. effector mechanisms have been recently shown to playUninfected recipients did not exhibit tissue damage an important role in host defense against T. cruzi (3, 4).following cell transfer. No disease exacerbation oc- Nevertheless, the mounting of an immune responsecurred in recipients of OVA-reactive CD4/, IL-4/IL- does not ensure total elimination of the parasites,10-secreting T lymphocytes stimulated with OVA at which persist in low numbers in the vertebrate hostthe start of infection. On Day 6 postinfection, not throughout life.only spleen cells but also LN cells from infected re- Available data have demonstrated that several mac-cipients showed decreased production of IFN-g and rophage-activating or -deactivating cytokines deter-augmented secretion of IL-4/IL-10 compared to cells mine the fate of intracellular T. cruzi amastigotes infrom untransferred infected mice. The results indi-
vitro. The addition of IFN-g (5–8), GM-CSF (9), IL-4cate that an imbalance of Th cell populations leading(10), or TNF-a (8, 11, 12) to cultures of T. cruzi-infectedto the predominance of secreted IL-4 and IL-10 at themacrophages results in more efficient killing of amasti-start of infection and the concomitant down-regula-gotes by the phagocytes, whereas the addition of TGF-tion of IFN-g secretion reversed the host’s resistanceb or IL-10 inhibits the trypanocidal action of INF-g-to T. cruzi. Moreover, transfer of anti-T. cruzi Th2-activated macrophages (13–15).type cells most likely favored the in vivo expansion
Most importantly, the in vivo administration of IFN-g (16) early during infection effectively reduces bloodparasitism and mortality in mice, whereas in vivo1 To whom correspondence should be addressed at Departamento treatment with anti-IFN-g Ab (17, 18) or with TGF-bde Imunologia, Instituto de Ciencias Biomedicas, Universidade de(13) has the opposite effect. Conversely, mice treatedSao Paulo, EdifıB cio Biomedicas III, Cidade Universitaria, 05508-900,
Sao Paulo, SP, Brazil. Fax: 55-11-813-0845 or 55-11-818-7224. with anti-IL-4 (18) or with anti-IL-10 (19) Ab become
41 0008-8749/96 $18.00Copyright q 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.
AID CI 8135 / 6c0f$$$181 04-30-96 09:19:34 cias AP: Cell Immuno

42 BARBOSA DE OLIVEIRA ET AL.
more resistant to challenge with a reticulotropic strain companied by increased IL-4 and IL-10 and reducedIFN-g secretion by spleen and lymph node (LN)2 cellsof T. cruzi.
Results obtained to date have thus established that during the course of infection. This imbalance of cyto-kine secretion at the onset of infection resulted inthe severity of experimental Chagas’ disease can be
modified by manipulating in vivo the availability of higher parasite loads in both tissues and blood.cytokines which control macrophage parasiticidal func-tion. It must be emphasized, however, that this manip- MATERIALS AND METHODSulation has been accomplished by treatment with re-
Mice. Ten- to twelve-week-old C57BL/6 male micecombinant cytokines (IFN-g, TGF-b) or with Aband (BALB/c 1 A/J)F1 male mice were obtained fromagainst the cytokines (anti-IFN-g, anti-IL-4, anti-IL-the Breeding Facilities of the Department of Immunol-10). Nonetheless, some experimental data indicate thatogy, Institute of Biomedical Sciences, University of Saoin vitro responses to recombinant cytokines not alwaysPaulo.match those observed for the secreted species, particu-
larly when a given cytokine under study is acting in Parasite infection and parasite Ag preparation. T.the presence of other cytokines simultaneously pro- cruzi (Y strain) was maintained by weekly ip passageduced by T cells (7). into male (BALB/c 1 A/J) F1 mice. Infected blood
So far, there is no direct in vivo experimental evi- drawn by cardiac puncture from mice killed by CO2dence that the course of a T. cruzi infection can be inhalation at the peak of parasitemia was appropri-changed by altering the initial ratios of IFN-g and IL- ately diluted with PBS (0.01 M phosphate-buffered sa-10/IL-4 parasite-specific secretor cells in secondary line–0.14 M NaCl, pH 7.2). C56BL/6 mice were infectedlymphoid organs. In this respect, the adoptive transfer by ip injection of 21 105 blood trypomastigotes. T. cruziof Th cells with known cytokine secretory profile and tissue-culture trypomastigote Ag (Tc-Ag) was preparedtheir in vivo tracing by specific proliferative and secre- from parasites harvested from infected monolayers oftory responses prior to infection represents a particu- LLC-MK2 cells (American Type Culture Collection,larly useful model of investigation. Cytokine secretion ATCC CCL7.1) by six to eight cycles of freezing andby transferred cells will take place in the lymphoid thawing (FT), as previously described (24). Plasmo-organs where the immune response against the infec- dium chabaudi chabaudi maintained by weekly ip in-tion is stimulated. fection of BALB/c mice with schizont-infected erythro-
In the murine experimental model for Leishmania cytes was a kind gift from Dr. Thereza L. Kipnis (De-major infection, resistant and susceptible mouse partment of Immunology, Institute of Biomedicalstrains exhibit, respectively, an overwhelming domi- Sciences, University of Sao Paulo). P. chabaudi Ag wasnance of either Th1 or Th2 parasite-specific subsets prepared from the intracellular stages of the parasite(20). Yet the situation does not appear to be so clear- after lysis of erythrocytes and purification by centrifu-cut for T. cruzi infections, where both resistant and gation in a discontinuous Percoll gradient (Pharmaciasusceptible mice show increased IFN-g secretion by Fine Chemicals, Upsala, Sweden), as described by Fa-spleen cells (21). On the other hand, production of MAF langa et al. (25); P. chabaudi parasites were disruptedby spleen cells is delayed (22) and IL-4 production is by eight cycles of freezing and thawing. Leishmaniadetectable during the acute phase of infection in sus- major, Leishmania amazonensis, and Leishmania don-ceptible strains as compared to resistant strains (23). ovani chagasi Ags were obtained from FT promasti-
In view of this, we became interested in investigating gotes grown in Grace’s culture medium (Sigma Chemi-whether, by expanding a Th parasite-specific subpopu- cal Co., St. Louis, MO) according to Sacks et al. (26).lation, we might selectively alter the predominance of
Immunization and cutaneous hypersensitivity test-secreted regulatory cytokines in vivo and thereby inter-ing. C57BL/6 mice were immunized with 5 1 107 Tc-fere with a genetically preestablished pattern of resis-Ag and 100 mg saponin (Sigma Chemical Co.) as adju-tance to T. cruzi in mice.vant by sc injection at the base of the tail (final volume:In this paper, we show that spleen cells of C57BL/6100 ml). Mice were tested 10 days later by injecting 2mice transferred with a predominantly T CD4/, IL-4-1 107 Tc-Ag (30 ml) intradermically into one of the hindand IL-10-secreting, parasite-Ag-reactive lymphocytefootpads. Footpad swelling was monitored with a met-population, proliferate significantly in response to T.ric caliper (precision 0.01 mm) and expressed as thecruzi Ag, presenting a reduced capacity to secrete IFN-increase in thickness relative to the uninjected contra-g and increased production of IL-4 and IL-10 with re-lateral paw. Results are presented as the arithmeticspect to control values. These recipient mice, infected
soon after cell transfer, become very susceptible to T.cruzi infection, contrasting with their typically resis- 2 Abbreviations used: Tc-Ag, T. cruzi trypomastigote antigen; pi,tant untransferred counterparts. The shift toward sus- postinfection; FT, frozen and thawed; LN, lymph node; OVA, oval-
bumin.ceptibility observed in the infected recipients was ac-
AID CI 8135 / 6c0f$$$182 04-30-96 09:19:34 cias AP: Cell Immuno

43Th2 CELLS IN EXPERIMENTAL CHAGAS’ DISEASE
mean { SEM (n Å 5). Immunization with ovalbumin fected ip with 2 1 105 T. cruzi blood forms. The courseof infection was monitored by counting (optical micro-(OVA) was carried out in the same manner using 200
mg of 21 crystallized OVA (Sigma Chemical Co.) and scope, 401 objective) the number of trypomastigotes in5-ml samples of citrated blood drawn from the tail veins,100 mg saponin. Eight days later, OVA-sensitized mice
were tested in the footpad with 30 ml of 2% heat-aggre- as described by Brener (31). A group of mice transferredwith OVA-specific lymphoblasts and infected with T.gated OVA (27) and the delayed cutaneous reaction
was measured as described. cruzi was boostered ip with 2 mg OVA at selected timepoints, beginning on the second day of infection. ThePreparation of Ag-reactive lymphoblasts. Cell sus- significance of differences in parasitemia between dis-pensions were prepared from the draining (inguinal and tinct experimental groups was examined by analysisperiaortic) lymph nodes of Tc-Ag or OVA-sensitized mice of variance with repeated measurements followed bysacrificed on Day 10 or 8 postimmunization, respectively. Tukey’s test for multiple comparisons. Differences ofCells were cultured in 24-well flat-bottomed plates at 6 0.5 log10 or over were significant at P õ 0.05.1 106 cells/well in 1.5 ml DMEM supplemented with 10
Cell cultures and proliferation assays. Inguinal andmM Hepes, 1.5 mM L-glutamine, 0.05 mM 2-ME, 0.24periaortic lymph nodes were removed from mice 10mM L-asparagine, 0.55 mM arginine, 1 mM sodium piru-days after immunization with Tc-Ag and saponin. Cellvate, 43 mM NaHCO3, penicillin, and streptomycin (100suspensions of the organs were cultured for 86 hr inU/ml and 100 mg/ml, respectively) and 7% endotoxin-free96-well flat-bottomed plates at 4 1 105 cells/well inFCS (Cultilab, Campinas, SP, Brazil). Cell cultures weremedium containing 7% FCS (final volume: 200 ml) andspecifically stimulated with either Tc-Ag (1.51 106 para-stimulated with Tc-Ag (2 1 106/ml), Leishmania Agssites/well) or OVA (150 mg/well) for 72 hr at 377C in a(2.5 1 106/ml), P. chabaudi Ag (12.5 mg/ml), FT LLC-humidified incubator (5% CO2). Large-size/low-densityMK2 cells (1.5 1 104/ml), OVA (100 mg/ml). Prolifera-lymphoblasts were isolated by centrifugation (1200g, 15tion was measured by adding 1 mCi [3H]TdR/well (Am-min, 227C) on a discontinuous Percoll gradient (25, 40,ersham, Arlington Heights, IL, sp act 5 Ci/mmol) 18 hr50, and 70%) and obtained from the 40–50% interface,before termination of the cultures. Radioactivity wasas previously described (28). Lymphoblasts were addi-measured by scintillation counting and the data weretionally cultured for 72 hr in the absence of further Agpresented as the arithmetic mean (cpm) of triplicatestimulation, in conditioned medium supplemented withcultures and the SD. Evaluation of cytokine production8% (v/v) TCGF-containing EL-4 supernatant obtainedby either normal mice or T. cruzi-infected mice trans-as described by Hilfiker et al. (29). The expandedferred or not with Tc-Ag-specific lymphoblasts prior tolymphoblasts were then washed twice (200g, 5 min atinfection (and sacrificed 6 days after challenge with187C) in medium and used in cell transfer experiments.the parasite) was carried out using supernatants fromThese lymphoblasts were also stained for cell surfacespleen and lymph node (inguinal, periaortic, and medi-markers by immunofluorescence, or further stimulatedastinal) cell cultures. Cell suspensions were distributedin vitro with Con A and Ags in the presence of irradiatedinto 24-well plates at 2 1 106 cells/ml in medium withspleen APC from normal syngeneic mice (2700 rad) to7% FCS and stimulated with Con A (5 mg/ml) or Tc-Agdetermine their pattern of cytokine secretion. Tc-Ag-spe-(1 1 106/ml) or OVA (100 mg/ml). Supernatants werecific lymphoblasts consisted of 86–90% CD4/ (GK 1.5),harvested by centrifugation (200g, 10 min, 47C) after4–6% CD8/ (53–6.7) (Amersham, Arlington Heights, IL)24 hr (for IL-2 and IL-4 measurements) or 66–72 hr (forand 4–8% surface Ig/ (FITC-goat anti-mouse Ig, SigmaIFN-g and IL-10 measurements) of culture. SimilarlyChemical Co.). In some experiments, the cells wereproduced supernatants from blasts cells (1 1 106 cells/treated (30 min, 47C) with the 31 M IgM anti-CD8 mAbwell) cocultured with g-irradiated (2700 rad) normal(30), washed, and incubated (30 min, 377C) with normalspleen APC (5 1 106 cells/well) were used to determinefresh rabbit serum to eliminate CD8/ cells; dead cellscytokine secretion by lymphoblasts.and debris were then removed by centrifugation on a
cushion of lympholyte M (Cedarlane, Hornby, Ontario, Detection of cytokines. Concentrations of IL-2 in theCanada). The final cell suspensions consisted of 98% via- supernatants were quantified in a biological assay us-ble cells with minimal contamination by CD8/ cells (less ing the HT-2 cell line as indicator cell in the presencethan 0.1%). of the neutralizing anti-IL-2 mAb (S4B6), and anti-IL-
2 mAb plus anti-IL-4 mAb (11B11) (32). Two-site sand-Transfer of lymphoblasts and infection of recipientwich ELISA was used to measure IFN-g (33) and alsomice. Tc-Ag- or OVA-specific lymphoblasts were in-IL-10 and IL-4 (34). Duplicate samples were assayedjected iv (lateral tail vein) into normal C57BL/6 mice.in a range of serial dilutions; the SD of mean valuesFootpad testing was performed 24 hr after cell transferdid not exceed 20% of the mean.using either Tc-Ag or aggregated OVA and the reaction
was measured 24 hr later. Groups of these transferred Evaluation of parasite load in spleen macrophagesand histopathology. Spleen cell suspensions from in-mice and of naive mice were then simultaneously in-
AID CI 8135 / 6c0f$$$182 04-30-96 09:19:34 cias AP: Cell Immuno

44 BARBOSA DE OLIVEIRA ET AL.
fected mice were incubated for 72 hr at 2 1 106/ml in further expanded by isolating lymphoblasts from spe-cifically Tc-Ag-stimulated LN cell cultures on a Percolltissue culture wells containing individual round glass
coverslips. At the end of the incubation period, cov- gradient and growing the blast cells for 72 hr in me-dium enriched with EL-4 supernatant.erslips were gently flushed with 0.01 M PBS and the
adherent cells were fixed with 1% paraformaldehydeIn Vitro Expanded T. cruzi Ag-Reactive Lymphoblastsin PBS. Staining was done with Giemsa stain (Sigma
Produce High Levels of IL-4 and IL-10 and LowChemical Co.). The coverslips were finally mounted onLevels of IFN-g and IL-2 after Ag-Specifichistological slides and the number of amastigotes perRestimulation in Cultureinfected cell and of parasitized macrophages per 500
cells were counted on a light microscope using a 1001 The T. cruzi Ag-Reactive lymphoblasts were predom-objective. Organs were removed for histopathological inantly CD4/ cells producing high levels of IL-4 andevaluation from both naive and transferred mice on IL-10 and very low levels of IFN-g and IL-2 when re-Day 12 of infection, from uninfected naive (control) stimulated with Tc-Ag in the presence of irradiatedmice and from uninfected recipient mice at different normal spleen APC (Table 1). The levels of IL-4 pro-times after cell transfer, and fixed for 24 hr in Bouin’s duced and the ratios of IL-4/IL-2 and of IL-4/IFN-gfixative. Five-millimeter-thick paraffin-embedded sec- production by Tc-Ag-reactive lymphoblasts were com-tions were cut and stained with hematoxylin and eosin. parable to those observed upon antigen stimulation ofSeveral nonserial sections of each organ were exam- BCG-specific Th cell lines exhibiting a dominant Th2ined; organs were collected from three to five mice for pattern of lymphokine secretion (35). However, theeach experimental group. Photomicrographs were lymphoblasts make up a mixed population that in-taken on a Nikon Labophot photomicroscope (Nikon, cludes cells capable of producing IL-2 and IFN-g, sinceTokyo, Japan). an increased detection of these cytokines was observedafter polyclonal stimulation with Con A. Having estab-
RESULTS lished the phenotypic characteristics of anti-Tc-Aglymphoblasts, these cells were next used in adoptive
Induction of Cell-Mediated Immune Responses by transfer experiments to study the outcome of T. cruziImmunization with T. cruzi Trypomastigote Ag and infection in recipient mice. In order to compare theSaponin infection in such animals with that of mice transferred
with T lymphoblasts specific to an unrelated antigen,To obtain Ag-specific T cells for studying the immu- OVA-reactive blast cells were obtained from OVA-im-noregulation of T. cruzi infections, immunization proto- munized mice following the same protocol used to pro-cols were developed that would stimulate intense duce Tc-Ag-reactive blasts. Those cells also secretedparasite-specific cell-mediated immune responses. In a predominantly IL-4 and IL-10 and very low levels ofseries of preliminary kinetic and dose–response experi- IFN-g upon restimulation with OVA and increased lev-ments, we established a subcutaneous immunization els of IFN-g after stimulation with Con A in vitro (Ta-protocol using freeze–thawed trypomastigotes (Tc-Ag) ble 1).in saponin as an adjuvant. Mice sensitized in this waygave a maximal delayed cutaneous response 10 days Systemic Transfer of T. cruzi Ag-Reactive CD4/after immunization when rechallenged with Tc-Ag in Lymphoblasts but Not of OVA-Reactivethe footpad (Fig. 1). Anti-T. cruzi DTH could be locally Lymphoblasts Aggravates T. cruzi Infection inand systemically transferred to naive syngeneic recipi- Recipient Mice Leading to Overwhelmingents by sensitized LN cells from immune donors but not Parasitemia and Early Deathby immune serum (inset, Fig. 1). On histopathologicalexamination of the reaction site, the inflammatory in- Blood parasitemia of both infected control mice and
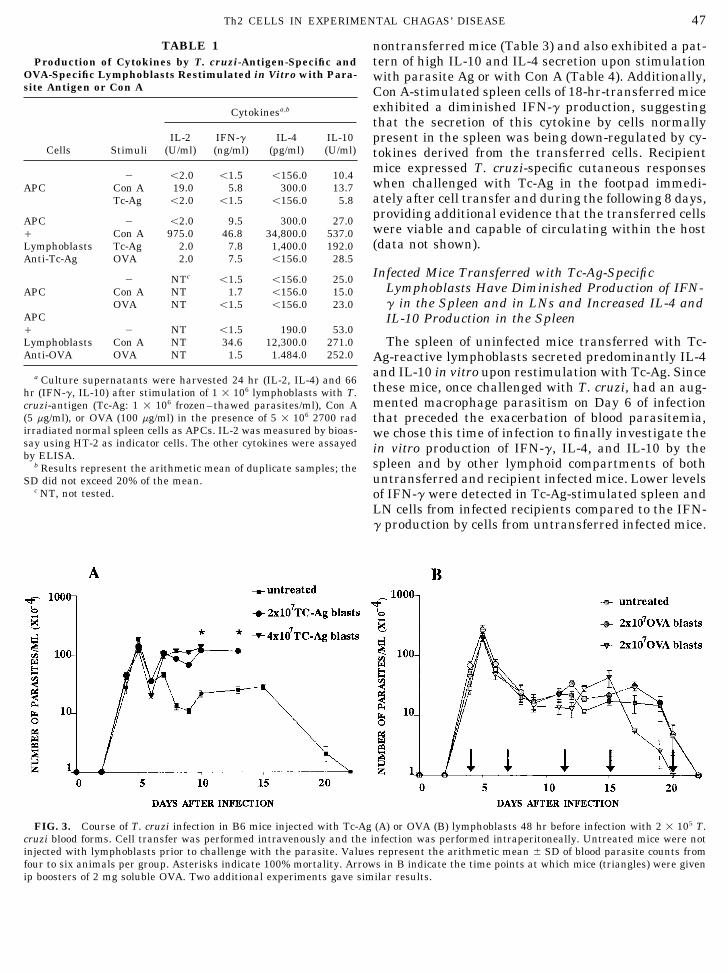
mice transferred with Tc-Ag-reactive lymphoblastsfiltrate at the peak of the cutaneous response (24 hr)was shown to be predominantly composed of mononu- 48 hr prior to challenge with T. cruzi were very simi-
lar during the first week of infection. However, overclear cells, indicative of an ongoing delayed type hyper-sensitivity reaction. the following weeks, the control group gradually
cleared blood trypomastigotes from the circulation toCells from the draining LN of immunized mice prolif-erated in vitro when stimulated with Tc-Ag. The re- undetectable levels, whereas the transferred group
developed higher and sustained parasitemias fromsponse was specific for T. cruzi antigens since FT Leish-mania promastigotes (L. major, L. amazonensis, and Day 8 of infection onward, exhibiting statistically sig-
nificant differences with respect to values from theL. chagasi), FT Plasmodium chabaudi, ovalbumin, orFT LLC-MK2 cells used to grow T. cruzi trypomasti- controls (Põ 0.05) (Fig. 3A). Therefore, naive C57BL/
6 mice were able to restrain the initial load of para-gotes did not stimulate significant proliferation (Fig.2). The population of T. cruzi-specific T cells was then sites and typically remained alive when reaching
AID CI 8135 / 6c0f$$$182 04-30-96 09:19:34 cias AP: Cell Immuno

45Th2 CELLS IN EXPERIMENTAL CHAGAS’ DISEASE
FIG. 1. Cutaneous hypersensitivity reaction of mice immunized at the base of the tail with Tc-Ag and saponin and challenged 10 dayslater with Tc-Ag in the footpad. Values represent the arithmetic mean { SEM (n Å 5). Open bars, immunized mice; solid bars, normalmice. Two further experiments gave similar results. The inset shows the 24-hr footpad reactions of naive recipients adoptively transferredwith lymph node cells (iv or local) or serum (iv) obtained from donors previously immunized with Tc-Ag / saponin. Results are expressedas the percentage value of 24-hr cutaneous reactions exhibited by donor mice.
subpatent levels of parasitemia (Day 22 of infection); ering that the anti-Tc-Ag blast population containedCD8/ cells (4–6%), mice were injected with Tc-Ag-reac-furthermore, they still presented very high survival
rates (of 80 to 100%) 40 days after infection (data not tive lymphoblasts depleted or not of CD8/ cells andthen challenged with T. cruzi. No significant differ-shown). By contrast, C57BL/6 mice that received Tc-
Ag-specific lymphoblasts prior to challenge with T. ences in the numbers of circulating parasites or in mor-tality rates were observed between these two groups ofcruzi appeared unable to control parasite replication
and exhibited mortality rates of 100% between Days transferred animals (data not shown).10 and 20 of infection. Increasing the number oftransferred cells to up to 4 1 107 was not paralleled Exacerbation of the Disease in T. cruzi-Infectedby further significant increases in the number of Recipient Mice Is Accompanied by Intensecirculating parasites but did accelerate death, as Parasitemia of Muscle Fibers and Macrophages100% mortality rates were observed on Day 20 post-infection in mice receiving the lowest number of Because the differences in blood parasitemias be-
tween nontransferred mice and recipient mice mightlymphoblasts (1 1 107 cells/animal; data not in-cluded), still on Days 13 and 10 in mice receiving 2 reflect differences in their clearance abilities but not
necessarily in the replication of T. cruzi within tissue1 107 and 4 1 107 cells, respectively. As shown inFig. 3B, no exacerbation of the infection occurred in cells, we next investigated the extent of parasite in-
fection and tissue damage in several organs exam-the recipients of OVA-reactive lymphoblasts trans-ferred prior to infection. Additional ip boosters of sol- ined on Day 12 postinfection. The analysis revealed
a more severe parasite load and inflammation (mono-uble OVA were given to a group of these recipientsin order to specifically stimulate the OVA-reactive nuclear and polymorphonuclear infiltrates) in the
heart, smooth muscle layers of the intestine and uri-cells along the infection; however, the dose of Ag em-ployed and/or frequency of boostering did not result nary bladder, skeletal muscle, and spleen of mice
transferred with Tc-Ag-reactive lymphoblasts priorin aggravation of the concomitant T. cruzi infection.Since CD8/ lymphocytes have been shown to partici- to infection (Table 2; Fig. 4). Most importantly (also
described in Table 2), as early as 6 days postinfection,pate in host resistance to T. cruzi (3, 4, 36) and consid-
AID CI 8135 / 6c0f$$$183 04-30-96 09:19:34 cias AP: Cell Immuno

46 BARBOSA DE OLIVEIRA ET AL.
FIG. 2. Specificity of in vitro proliferative responses of lymph node cells 10 days after immunization with Tc-Ag and saponin. Cells fromnormal (solid bars) or immunized mice (open bars) were stimulated in culture for 86 hr with Tc-Ag, ovalbumin (OVA), frozen–thawed purifiedprotozoa (Leishmania major, Leishmania amazonensis, Leishmania chagasi, Plasmodium chabaudi) or with frozen–thawed uninfected LLC-MK2 cells that were routinely used to grow T. cruzi trypomastigotes in vitro. [3H]TdR was added 18 hr prior to the termination of cultures.Values are shown as the arithmetic mean of cpm from triplicate cultures { SD from a representative experiment. The experiment wasrepeated once with all of the antigens and 15 times with Tc-Ag, OVA, and LLC-MK2 with similar results.
the number of intracellular parasites in cultured cal alterations (of the organs listed in Table 2) forboth groups of mice (data not shown).spleen macrophages was much higher in cells from
recipient mice compared to cells from nontransferredProduction of IFN-g Is Decreased and That of IL-4mice. Since on Day 6 the numbers of circulating blood
and IL-10 Is Increased in the Spleen and LNs ofparasites did not differ significantly between the twoNormal Mice Transferred with Tc-Ag-Specificgroups (see Fig. 3), this latter observation shows thatLymphoblaststhe initial load of T. cruzi had already expanded to
a greater extent in the splenic compartment of the Depending on their state of activation, macrophagesrecipients before differences in blood parasitemia be- harboring T. cruzi may either eliminate the parasitecame apparent. or allow it to multiply. The spleen is particularly rich
There are several reports on epitopes common to in macrophages and we found that it was parasitizedboth T. cruzi and normal host tissues antigens (37). above control levels in mice transferred with Tc-Ag-Thus, it was of interest to evaluate whether the adop- reactive lymphoblasts already on Day 6 postinfection.tively transferred parasite-Ag-specific lymphoblasts Since the observed in vitro secretion of lymphokines bywere able to bring about inflammation or tissue dam- these blasts would certainly establish conditions forage in the absence of infection and therefore contrib- macrophage deactivation (14, 15, 38), their early pres-ute through autoreactive mechanisms to the ob- ence in the splenic compartment of recipient miceserved exacerbation of infection. To investigate this might determine an environment favorable to T. cruzipossibility, mice were transferred with Tc-Ag-specific replication in vivo. To examine this possibility, we nextlymphoblasts: one group was autopsied at different investigated the splenic distribution of Tc-Ag-specifictime intervals after cell transfer and a second group lymphoblasts in the absence of infection. Tc-Ag-reac-received intravenous booster injections of 3 1 107 Tc- tive lymphoblasts were apparent in the spleens of re-Ag every 7 days before autopsy, which was performed cipient mice 18 hr after iv cell transfer, since culturesafter a total period of 47 days. Histopathology did not of spleen cells from these animals proliferated signifi-
cantly in the presence of Tc-Ag, unlike those of normalreveal any inflammation or conspicuous morphologi-
AID CI 8135 / 6c0f$$$183 04-30-96 09:19:34 cias AP: Cell Immuno
47Th2 CELLS IN EXPERIMENTAL CHAGAS’ DISEASE
TABLE 1 nontransferred mice (Table 3) and also exhibited a pat-tern of high IL-10 and IL-4 secretion upon stimulationProduction of Cytokines by T. cruzi-Antigen-Specific and
OVA-Specific Lymphoblasts Restimulated in Vitro with Para- with parasite Ag or with Con A (Table 4). Additionally,site Antigen or Con A Con A-stimulated spleen cells of 18-hr-transferred mice
exhibited a diminished IFN-g production, suggestingCytokinesa,b
that the secretion of this cytokine by cells normallyIL-2 IFN-g IL-4 IL-10 present in the spleen was being down-regulated by cy-
Cells Stimuli (U/ml) (ng/ml) (pg/ml) (U/ml) tokines derived from the transferred cells. Recipientmice expressed T. cruzi-specific cutaneous responses0 õ2.0 õ1.5 õ156.0 10.4when challenged with Tc-Ag in the footpad immedi-APC Con A 19.0 5.8 300.0 13.7ately after cell transfer and during the following 8 days,Tc-Ag õ2.0 õ1.5 õ156.0 5.8providing additional evidence that the transferred cells
APC 0 õ2.0 9.5 300.0 27.0were viable and capable of circulating within the host/ Con A 975.0 46.8 34,800.0 537.0(data not shown).Lymphoblasts Tc-Ag 2.0 7.8 1,400.0 192.0
Anti-Tc-Ag OVA 2.0 7.5 õ156.0 28.5Infected Mice Transferred with Tc-Ag-Specific0 NTc õ1.5 õ156.0 25.0
Lymphoblasts Have Diminished Production of IFN-APC Con A NT 1.7 õ156.0 15.0g in the Spleen and in LNs and Increased IL-4 andOVA NT õ1.5 õ156.0 23.0
APC IL-10 Production in the Spleen/ 0 NT õ1.5 190.0 53.0Lymphoblasts Con A NT 34.6 12,300.0 271.0 The spleen of uninfected mice transferred with Tc-Anti-OVA OVA NT 1.5 1.484.0 252.0 Ag-reactive lymphoblasts secreted predominantly IL-4
and IL-10 in vitro upon restimulation with Tc-Ag. Sincea Culture supernatants were harvested 24 hr (IL-2, IL-4) and 66these mice, once challenged with T. cruzi, had an aug-hr (IFN-g, IL-10) after stimulation of 1 1 106 lymphoblasts with T.mented macrophage parasitism on Day 6 of infectioncruzi-antigen (Tc-Ag: 1 1 106 frozen–thawed parasites/ml), Con A
(5 mg/ml), or OVA (100 mg/ml) in the presence of 5 1 106 2700 rad that preceded the exacerbation of blood parasitemia,irradiated normal spleen cells as APCs. IL-2 was measured by bioas- we chose this time of infection to finally investigate thesay using HT-2 as indicator cells. The other cytokines were assayed in vitro production of IFN-g, IL-4, and IL-10 by theby ELISA.
spleen and by other lymphoid compartments of bothb Results represent the arithmetic mean of duplicate samples; theuntransferred and recipient infected mice. Lower levelsSD did not exceed 20% of the mean.
c NT, not tested. of IFN-g were detected in Tc-Ag-stimulated spleen andLN cells from infected recipients compared to the IFN-g production by cells from untransferred infected mice.
FIG. 3. Course of T. cruzi infection in B6 mice injected with Tc-Ag (A) or OVA (B) lymphoblasts 48 hr before infection with 2 1 105 T.cruzi blood forms. Cell transfer was performed intravenously and the infection was performed intraperitoneally. Untreated mice were notinjected with lymphoblasts prior to challenge with the parasite. Values represent the arithmetic mean { SD of blood parasite counts fromfour to six animals per group. Asterisks indicate 100% mortality. Arrows in B indicate the time points at which mice (triangles) were givenip boosters of 2 mg soluble OVA. Two additional experiments gave similar results.
AID CI 8135 / 6c0f$$$183 04-30-96 09:19:34 cias AP: Cell Immuno

48 BARBOSA DE OLIVEIRA ET AL.
TABLE 2Increased Organ Parasitism in T. cruzi-Infected Mice Transferred with Tc-Ag-Reactive Lymphoblasts
Experimental group Organ or tissuea Description of findings
Skeletal muscle Focal mononuclear (MN) cell infiltrates and few parasites withinmacrophages.
Infection with 2 1 105 T. cruzi Smooth muscle Focal MN cell infiltrates in the urinary bladder and in both smalland large intestine. Few parasites mostly located withinmacrophages; occasional parasitism of muscle fibers.
Spleen Follicular hyperplasia and few parasitized macrophages.Quantification of Percentage of infected cells in 72-hr adherent spleen cell cultures:
spleen parasitismb 17 { 2. Number of parasites/cell in the same cultures: 90 { 10.Heart Moderate MN cell infiltrate; occasional polymorphonuclear (PMN)
foci. Parasite nests: 1–2/section
Transfer of Tc-Ag lymphoblasts Skeletal muscle Moderate to intense MN and focal PMN infiltrates. Moderateparasitism of fibers.
/Smooth muscle Very intense parasitism of muscle fibers in the large intestine
with scanty inflammatory infiltrate. Moderate parasitism of thefibers in the urinary bladder and small intestine. Foci of MNand PMN infiltrates.
Infection with 2 1 105 T. cruzi Spleen Loss of follicular architecture, marked increase in the number ofPMN, large mononuclear cells and macrophages.
Quantification of Percentage of infected cells in 72-hr adherent spleen cell cultures:spleen parasitismb 45 { 5. Number of parasites/cell in the same cultures: 201 { 23.
Heart Moderate to intense MN and PMN infiltrates. Markedparasitism. Parasite nests: 15–20/section.
a Tissue parasitism of autopsied organs was quantified on Day 12 postinfection.b Spleen cell parasitism was quantified on Day 6 postinfection.
A diminished production of this cytokine by polyclon- DISCUSSIONally (Con A) stimulated cells from the spleen and peri-aortic/inguinal LN of the recipients was also observed
Control of T. cruzi parasitemia is critically depen-(Table 5). It should be noted that cells grown in medium dent on the host’s ability to mount an effective cellularalone or medium with OVA contain T. cruzi-infected immune response during the first weeks of infectionmacrophages which constitute an endogenous source (2, 39). The data presented in this paper show thatof parasite-specific stimulation; the reduced in vitro se- resistant C57BL/6 mice could be turned into suscepti-cretion of IFN-g by the recipients with respect to the ble animals by the transfer of a predominantly CD4/controls was also evident in these cell cultures. By con- parasite-specific IL-4- and IL-10-secreting lymphocytetrast, Tc-Ag- or Con A-stimulated spleen cell cultures subpopulation. The production of IFN-g by spleen andfrom the transferred-infected mice (as well as their Con LN cells was decreased, whereas that of IL-4 and IL-A-stimulated LN cells) produced markedly higher lev- 10 was augmented in recipient mice, whose failure toels of IL-4 than similarly cultured spleen and LN cells restrain the extent of T. cruzi replication within spleenof the infected controls (Table 6). IL-10 production in macrophages was already apparent by Day 6 of infec-Tc-Ag-stimulated spleen and LN cell cultures from tion. Two days later, the inability to control T. cruzitransferred–infected mice was also increased (Table parasitemia was also reflected in the persistently ele-7), but to lesser extent than IL-4 levels. vated numbers of blood parasites and premature death
Taken together, these results show that very early of the animals.during the course of a T. cruzi infection in the resistant Following the iv transfer of Tc-Ag lymphoblasts,C57BL/6 mouse, spleen and LN cells secrete predomi- their presence in the spleen of recipient mice was de-nantly IFN-g, low or moderate amounts of IL-10, and tected by proliferative and IL-4/IL-10-secretory re-very little IL-4 upon in vitro stimulation. Conversely, sponses to in vitro stimulation with parasite antigenspleen and LN cells from C57BL/6 mice transferred (Tc-Ag). Thus, after introducing a specific antigenicwith a high IL-4- and IL-10-secreting parasite-specific stimulus in vivo in the form of a challenge T. cruziT cell population prior to infection reverse such pattern infection, transferred cells would be expected to secreteof secretion to one of low IFN-g and high IL-10 and IL- IL-4 and IL-10. This, in turn, might down-regulate the
drive toward a dominant IFN-g response normally ob-4 production.
AID CI 8135 / 6c0f$$$183 04-30-96 09:19:34 cias AP: Cell Immuno

49Th2 CELLS IN EXPERIMENTAL CHAGAS’ DISEASE
FIG. 4. Increased tissue parasitism of myocardial and colonic smooth muscle fibers in 12-day T. cruzi-infected mice transferred withTc-Ag lymphoblasts 48 hr before infection. Photomicrographs (1350) A (heart) and B (medial colon) of 12-day infected lymphoblast-trans-ferred mice; C (heart) and D (medial colon) from 12-day infected mice not transferred with Tc-Ag-specific lymphoblasts. The arrows pointto intracellular amastigotes.
AID CI 8135 / 6c0f$$813504-30-96 09:19:34 cias AP: Cell Immuno

50 BARBOSA DE OLIVEIRA ET AL.
TABLE 3 TABLE 5Production of IFN-g by Cells from Different Lymphoid Com-Proliferative Responses of Spleen Cells from Recipient
Mice Transferred with Tc-Ag-Reactive Lymphoblasts and partments of Infected Mice Tranferred with T. cruzi Antigen-Specific Lymphoblasts Prior to InfectionSacrificed 18 hr after Cell Transfer
cpm (11003) [3H]TdR StimuliLymphoblastsincorporation in cultures
stimulateda with Cell sourceb tranferreda None Con A Tc-Ag OVA
Spleen 0 18.0 70.0 15.0 7.8Treatment of mice FCS OVA Tc-Ag/ 5.0 32.5 4.7 3.5
Periaortic andTransference of Tc-Ag-reactivelymphoblasts 3.0 { 0.5 2.9 { 0.1 46.9 { 1.4 inguinal 0 8.5 90.0 30.0 3.0
LNs / õ1.5 21.7 9.8 õ1.5None 1.4 { 0.1 2.5 { 0.4 2.3 { 0.3Mediastinal 0 3.0 70.0 38.0 2.6LNs / 1.6 66.0 15.2 õ1.5a Spleen cells were cultured for 72 hr with Tc-Ag, OVA or no anti-
genic stimulus (FCS). [3H]TdR was added 18 hr prior to terminationof the cultures. Values are shown as the arithmetic mean of tripicate a Lymphoblasts (2 1 107) were transferred by iv injection; 48 hr
later, recipient mice and a group of normal mice were infected ipcultures { SD.with 2 1 105 T. cruzi blood trypomastigotes.
b The organs were removed for culture on Day 6 postinfection andthe supernatants were harvested after 72 hr of culture. IFN-g levelswere determined by ELISA and expressed in ng/ml. Results repre-
served in infected untransferred mice. In fact, the pat- sent the arithmetic mean of duplicate experiments; the SD did notexceed 20% of the mean.tern of high IFN-g and low IL-10/IL-4 in vitro cytokine
secretion of infected resistant C57BL/6 mice wasmodified to one of higher IL-4/IL-10 and diminishedIFN-g levels of secretion as a consequence of cell trans-
emphasized that, upon in vitro stimulation, rela-fer. Significant in vivo activation of Tc-Ag-reactivetively low amounts of IL-4 are required for drivinglymphoblasts by T. cruzi challenge is indicated by thethe development of Th2 cells (40–42) and that thisfact that the transfer of IL-4- and IL-10-producingeffect of IL-4 occurs in the presence of concentrationsOVA-reactive lymphoblasts prior to infection failed toof IFN-g as high as 2500 U/ml (42).modify its course.
Thus, irrespective of Tc-Ag-reactive lymphoblastWhether the elevated IL-4 and IL-10 productionexpansion occurring or not early during infection, theseen in the spleen and other lymphoid cell compart-amount of endogenous IL-4 secreted during primaryments of recipient mice on Day 6 of infection wasT. cruzi stimulation of naive host T cells is likely toaccompanied by proliferation of the transferred cellscounterbalance the strong signals that normallyor whether only their activation took place cannotdrive these T cell responses toward a high IFN-g se-be inferred from our data. Nonetheless, it must be
TABLE 6TABLE 4Production of IL-4 by Cells from Different Lymphoid Com-Production of Cytokines by Spleen Cells of Uninfected Mice
partments of Infected Mice Transferred with T. cruzi Anti-Transferred with T. cruzi Antigen-Specific Lymphoblastsgen-Specific Lymphoblasts Prior to Infection
StimuliStimuliLymphoblasts
LymphoblastsCytokinesb transferreda None Con A Tc-Ag OVACell sourceb transferreda None Con A Tc-AG OVA
IFN-g 0 õ1.5 34.4 õ1.5 õ1.5Spleen 0 271.0 808.0 410.0 250.0(ng/ml) / 2.0 9.5 õ1.5 õ1.5
/ 300.0 2875.0 1349.0 245.0IL-4 0 õ156.0 õ156.0 õ156.0 õ156.0 Periaortic and(pg/ml) / õ156.0 1984.0 1389.0 õ156.0 inguinal 0 õ156.0 õ156.0 õ156.0 õ156.0IL-10 0 7.1 35.6 6.9 6.8 LNs / õ156.0 1023.0 õ156.0 õ156.0(U/ml) / 6.7 189.9 109.0 6.3 Mediastinal 0 õ156.0 õ156.0 õ156.0 õ156.0
LNs / õ156.0 580.0 õ156.0 õ156.0a Lymphoblasts (2 1 107) were transferred by iv injection into nor-
mal syngeneic mice; 18 hr later the spleens of the recipients and of a The conditions of cell transfer, infection, and organ removal wereas described in Table 5.a group and normal mice were removed for culture.
b Supernatants were harvested after 24 hr (IL-4) and 72 hr (IFN-g b The supernatants were harvested after 24 hr of culture and IL-4 levels were determined by ELISA and expressed in pg/ml. Resultsand IL-10) of culture and cytokines were assayed by ELISA. Results
represent the arithmetic mean of duplicate samples; the SD did not represent the arithmetic mean of duplicate samples; the SD did notexceed 20% of the mean.exceed 20% of the mean.
AID CI 8135 / 6c0f$$$183 04-30-96 09:19:34 cias AP: Cell Immuno

51Th2 CELLS IN EXPERIMENTAL CHAGAS’ DISEASE
TABLE 7 site-Ag-stimulated IL-4 production by spleen and LNcells was found to be detectable in Balb/c (susceptible)Production of IL-10 by Cells from Different Lymphoid Com-
partments of Infected Mice Transferred with T. cruzi Anti- but not in C57BL/6 (resistant) mice 6 to 12 weeks aftergen-Specific Lymphoblasts Prior to Infection T. cruzi infection (23).
Inhibition of T CD4/ IFN-g production by alreadyStimuliprimed T cells has been shown to occur in LN culturesLymphoblasts
Cell sourceb transferreda None Con A Tc-Ag OVA of L. major-primed CD4/ cells simultaneously secre-ting IFN-g, IL-4, and IL-10 and in spleen cell cultures
Spleen 0 18.6 58.0 56.4 14.6 from Schistosoma mansoni-infected mice secreting/ 23.8 61.0 102.0 29.0IFN-g and IL-10 (46, 47). Thus, inasmuch as the im-Periaortic andportance of IFN-g for the control of in vitro and ininguinal 0 6.2 72.0 38.5 6.4
LNs / 3.9 126.0 51.4 10.7 vivo T. cruzi parasitism is now well established (5–Mediastinal 0 6.4 107.9 25.4 7.2 8, 14–16, 48), our results indicate that the outcomeLNS / 7.4 128.4 42.3 12.9 of acute-phase T. cruzi infection is dictated by the
initial ratios of interacting IFN-g and IL-4/IL-10-se-a The conditions of cell transfer, infection and organ removal werecretory cells.as described in Table 5.
b The supernatants were harvested after 72 hr of culture and IL-10 The generation of virus-specific CD8/ CTL re-levels were determined by ELISA and expressed in units/ml. Results sponses has been shown to be impaired in S. man-represent the arithmetic means of duplicate samples; the SD did not soni-infected mice at a stage when Th2 cytokine pro-exceed 20% of the mean.
duction is dominant (49). Since CD8/ cells partici-pate in the resistance to T. cruzi infection (3, 4), adiminished CD8/ response could also contribute tothe marked susceptibility to infection of the trans-cretion pattern. This would prevent further expan-
sion of IFN-g-producing cells, reduce IFN-g produc- ferred mice.In spite of its predominant IL-4- and IL-10-secre-tion, and allow priming of host CD4/ cells to IL-4
secretion (40–42). tory profile, we demonstrated that the population ofTc-Ag-reactive lymphoblasts was able to mediateIn contrast with IL-4, IL-10 is not required for Th2
phenotype development (41, 42); yet the shift toward parasite-specific DTH reaction after iv transfer tosyngeneic naive recipients. Yet this lymphoblast pop-a Th1 phenotype reported for Listeria monocytogenes
has been shown to be inhibited by the presence of ulation contained cells capable of producing IFN-gupon mitogenic stimulation (Table 1). Thus, the factendogenous levels of IL-10 as well as IL-4 during
priming (43, 44). T. cruzi infection also appears to that the lymphoblast population was not cloned andthat very few sensitized CD4/ cells are needed toinduce predominantly Th1 cytokine responses early
during infection (21, 23, 45, and our data). In our sys- trigger DTH at a skin site challenged with specificantigen (50) can explain this result. Partial inhibi-tem, when transferred cells were activated by T. cruzi
challenge in the recipient mice, both IL-4 and IL- tion of DTH expression was reported to occur in miceafter systemic administration of rIL-4 and rIL10 (51)10 prevailed in the splenic environment. Thus, by
analogy to the L. monocytogenes model, it is reason- or of rIL-10 (52). However, in our system, anti-T.cruzi DTH expression by the transferred Th1able to speculate that both IL-4 and IL-10 partially
inhibited Th1 development of anti-T. cruzi host T lymphoblast population was not suppressed by thesimultaneously transfer of IL-4/IL-10-secreting Th2cells and led to an in vivo inbalance toward a Th2
phenotype. lymphoblasts.Previous studies reported the occurrence of pre-At the effector cell level, recombinant IL-10 inhibits
IFN-g-mediated macrophage activation for intracellu- dominantly mononuclear inflammatory lesions in theliver as well as demyelination and mononuclear in-lar killing of T. cruzi in vitro (14, 15) and treatment of
mice with anti-IL-10 (19) results in decreased parasite filtration of the sciatic nerves following the systemicor local transfer of class II autoreactive CD4/ T cellcounts and reduced mortality, confirming a regulatory
role for IL-10 in T. cruzi–host interactions. The func- lines, derived from chronically T. cruzi-infected mice,to naive uninfected recipients (53). Expression of aut-tion of IL-4 is less clear. In vitro, this cytokine has been
reported to either enhance (alone or in synergysm with oreactivity was associated in one of these cell linesto a high IL-4- and IL-6-secretory pattern and to itsIFN-g) the elimination of T. cruzi by macrophages
(7, 10) or, conversely, to promote intracellular parasit- ability to induce IgG2 polyclonal B cell activation,although the significance of this association is notism (7). However, it has also been reported that in vivo
treatment with anti-IL-4 mAb decreases parasitemia clear (54). Local injection of CD4/ cells from chroni-cally infected mice into skin-grafted heart tissue pro-and mortality by the Tulahuen strain of T. cruzi,
though not by the CL strain (18). Furthermore, para- moted its rejection (55). We did not observe inflam-
AID CI 8135 / 6c0f$$$184 04-30-96 09:19:34 cias AP: Cell Immuno

52 BARBOSA DE OLIVEIRA ET AL.
17. Torrico, F., Heremans, H., Rivera, M. T., Van-Marck, E., Billiau,mation in any of the organs listed in Table 2 nor inA., and Carlier, Y., J. Immunol. 146, 3626, 1991.the liver of uninfected recipient mice, suggesting that
18. Petray, P. B., Rottenberg, M. E., Bertot, G., Corral, R. S., Diaz,infection with the live parasite rather than immuni-A., Orn, A., and Grinstein, S., Immunol. Lett. 35, 77, 1993.zation with parasite antigens is required for the pos-
19. Reed, S. G., Brownell, C. E., Russo, D. M., Silva, J. S., Grabstein,sible expansion of autoreactive T cells (56). K. H., and Morrissey, P. J., J. Immunol. 153, 3135, 1994.In conclusion, our results indicate that activation 20. Sher, A., and Coffman, R. L., Annu. Rev. Immunol. 10, 385,
of Th2 cells early in the course of T. cruzi infection 1992.can drastically increase the severity of the disease in 21. Nabors, G. S., and Tarleton, R. L., J. Immunol. 146, 3591, 1991.a normally resistant host. Inasmuch as the initial 22. Nogueira, N., Ellis, J., Chaplan, S., and Cohn, Z., Exp. Parasitol.control of the parasite load is critically dependent on 51, 325, 1981.efficient parasite killing by macrophages, it is con- 23. Hoft, D. F., Lynch, R. G., and Kirchhoff, L. V., J. Immunol. 151,
7038, 1993.ceivable that even moderate imbalances of the ratio24. Curotto-de-Lafaille, M. A., Barbosa-de-Oliveira, L. C., Lima,of IFN-g and IL-4/IL-10 production occurring during
G. C., and Abrahamsohn, I. A., Exp. Parasitol. 70, 164, 1990.the initial phase of the infection might lead to in-25. Falanga, P. B., Franco-da-Silveira, J. F., and Pereira-da-Silva,creased or longer persisting tissue parasitism and
L., Parasite Immunol. 6, 529, 1984.worsening of inflammation and tissue damage.26. Sacks, D. L., Hieny, S., and Sher, A., J. Immunol. 135, 564,
1985.ACKNOWLEDGMENTS
27. Titus, R. G., and Chiller, J. M., J. Immunol. Methods 45, 65,1981.This work was supported by grants from FAPESP, CNPq, and
28. Kurnick, J. T., Gronvik, K. O., Kimura, A. K., Lindblom, J. B.,the UNDP/World Bank/WHO Special Programme for Research andSkoog, V. T., Sjoberg, O., and Wigzell, H., J. Immunol. 122,Training in Tropical Diseases. Laura C. Barbosa de Oliveira and1255, 1979.M. A. Curotto de Lafaille were supported by FAPESP and CNPq
Ph.D fellowships. We gratefully acknowledge Dr. Robert L. Coff- 29. Hilfiker, M. L., Moore, R. N., and Farrar, J. J., J. Immunol.man (DNAX Research Institute, Palo Alto, CA) for encouragement, 127, 1983, 1981.initial cytokine determinations, and the gift of cytokine reagents; 30. Sarmiento, M., Glasebrook, A. L., and Fitch, F. W., J. Immunol.Ademir V. da Silva and Ulisses R. da Silva for technical assistance; 125, 2665, 1980.Dr. Mahasti S. de Macedo and Dr. Anne O’Garra for critical read-
31. Brener, Z., Rev. Inst. Med. Trop. S. Paulo 4, 389, 1962.ing of the manuscript.32. Mosmann, T. R., and Fong, T. A., J. Immunol. Methods 116,
151, 1989.REFERENCES
33. Cherwinski, H. M., Schumacher, J. H., Brown, K. D., and Mos-mann, T. R., J. Exp. Med. 166, 1229, 1987.
1. Wrightsman, R., Krassner, S., and Watson, J., Infect. Immun.34. Abrams, J. S., Roncarolo, M. G., Yssel, H., Andersson, U.,36, 637, 1982.
Gleich, G. J., and Silver, J. E., Immunol. Rev. 127, 5, 1992.2. Trischmann, T. M., Exp. Parasitol. 62, 194, 1986.35. Abehsira-Amar, O., Gibert, M., Joliy, M., Theze, J., and Jan-3. Tarleton, R. L., J. Immunol. 144, 717, 1990.
kovic, D. L., J. Immunol. 148, 3820, 1992.4. Tarleton, R. L., Koller, B. H., Latour, A., and Postan, M., Nature
36. Nickell, S. P., Stryker, G. A., and Arevalo, C., J. Immunol. 150,356, 338, 1992.1446, 1993.
5. Plata, F., Wietzerbin, J., Pons, F. G., Falcoff, E., and Eisen, H.,37. Petry, K., and Eisen, H., Parasitol. Today 5, 1989.Eur. J. Immunol. 14, 930, 1984.38. Liew, F. Y., Li, Y., Severn, A., Millott, S., Schmidt, J., Salter,6. Wirth, J. J., Kierszenbaum, F., Sonnenfeld, G., and Zlotnik, A.,
M., and Moncada, S., Eur. J. Immunol. 21, 2489, 1991.Infect. Immun. 49, 61, 1985.39. Trischmann, T. M., J. Immunol. 130, 1953, 1983.7. Golden, J. M., and Tarleton, R. L., Exp. Parasitol. 72, 391, 1991.40. Swain, S. L., Weinberg, A. D., English, M., and Huston, G., J.8. Munoz-Fernandez, M. A., Fernandez, M. A., and Fresno, M.,
Immunol. 145, 3796, 1990.Eur. J. Immunol. 22, 301, 1992.41. Hsieh, C. S., Heimberger, A. B., Gold, J. S., O’Garra, A., and9. Reed, S. G., Nathan, C. F., Pihl, D. L., Rodricks, P., Shanebeck,
Murphy, K. M., Proc. Natl. Acad. Sci. USA 89, 6065, 1992.K., Conlon, P. J., and Grabstein, K. H., J. Exp. Med. 166, 1734,1987. 42. Seder, R. A., Paul, W. E., Davis, M. M., and Fazekas-de-St.
Groth, B., J. Exp. Med. 176, 1091, 1992.10. Wirth, J. J., Kierszenbaum, F., and Zlotnik, A., Immunology66, 296, 1989. 43. Hsieh, C. S., Macatonia, S. E., O’Garra, A., and Murphy, K. M.,
Int. Immunol. 5, 371, 1993.11. De-Titto, E. H., Catterall, J. R., and Remington, J. S., J. Immu-nol. 137, 1342, 1986. 44. Hsieh, C. S., Macatonia, S. E., Tripp, C. S., Wolf, S. F., O’Garra,
A., and Murphy, K. M., Science 260, 547, 1993.12. Wirth, J. J., and Kierszenbaum, F., J. Immunol. 141, 286, 1988.45. Abrahamsohn, I. A., and Coffman, R. L., J. Immunol. 155, 3955,13. Silva, J. S., Twardzik, D. R., and Reed, S. G., J. Exp. Med. 174,
1995.539, 1991.46. Powrie, F., Correa-Oliveira, R., Mauze, S., and Coffman, R. L.,14. Silva, J. S., Morrissey, P. J., Grabstein, K. H., Mohler, K. M.,
J. Exp. Med. 179, 589, 1994.Anderson, D., and Reed, S. G., J. Exp. Med. 175, 169, 1992.47. Sher, A., Fiorentino, D., Caspar, P., Pearce, E., and Mosmann,15. Gazzinelli, R. T., Oswald, I. P., Hieny, S., James, S. L., and
T., J. Immunol. 147, 2713, 1991.Sher, A., Eur. J. Immunol. 22, 2501, 1992.16. Reed, S. G., J. Immunol. 140, 4342, 1988. 48. Minoprio, P., el-Cheikh, M. C., Murphy, E., Hontebeyrie-
AID CI 8135 / 6c0f$$$184 04-30-96 09:19:34 cias AP: Cell Immuno

53Th2 CELLS IN EXPERIMENTAL CHAGAS’ DISEASE
Joskowicz, M., Coffman, R., Coutinho, A., and O’Garra, A., J. 52. Li, L., Elliott, J. F., and Mosmann, T. R., J. Immunol. 153,3967, 1994.Immunol. 151, 4200, 1993.
53. Hontebeyrie-Joskowicz, M., Said, G., Milon, G., Marchal, G.,49. Actor, J. K., Shirai, M., Kullberg, M. C., Buller, R. M., Sher,and Eisen, H., Eur. J. Immunol. 17, 1027, 1987.A., and Berzofsky, J. A., Proc. Natl. Acad. Sci. USA 90, 948,
54. Spinella, S., Milon, G., and Hontebeyrie-Joskowicz, M., Eur. J.1993.Immunol. 20, 1045, 1990.
50. Marchal, G., Seman, M., Milon, G., Truffa-Bachi, P., and Zilb- 55. Ribeiro-dos-Santos, R., Rossi, M. A., Laus, J. L., Silva, J. S.,erfarb, V., J. Immunol. 129, 954, 1982. Savino, W., and Mengel, J., J. Exp. Med. 175, 29, 1992.
51. Powrie, F., Menon, S., and Coffman, R. L., Eur. J. Immunol. 56. Gatass, C. R., Lima, M. T., Nobrega, A. F., Barcinski, M. A.,and Dos-Reis, G. A., Infect. Immun. 56, 1402, 1988.23, 3043, 1993.
AID CI 8135 / 6c0f$$$184 04-30-96 09:19:34 cias AP: Cell Immuno
Copyright © 2022 FDOKUMEN




















