
Maintenance of long-term immunological memory by low avidity IgM-secreting cells in bone marrow...
11
Vaccine 22 (2004) 1553–1563 Maintenance of long-term immunological memory by low avidity IgM-secreting cells in bone marrow after mucosal immunizations with cholera toxin adjuvant Elawati Soenawan a , Indresh Srivastava a , Soumi Gupta a , Elaine Kan a , Ramesh Janani a , Jina Kazzaz a , Manmohan Singh a , Vijay Shreedhar b , Michael Vajdy a,b,∗ a Chiron Corporation, Immunology and Infectious Diseases, Emeryville, CA, USA b Children’s Hospital and Harvard Medical School, Boston, MA, USA Received 20 March 2003; received in revised form 1 October 2003; accepted 2 October 2003 Abstract To understand the mechanisms involved in maintaining long-term immunological memory following mucosal immunizations, we determined the quality of serum hapten-specific immunoglobulins (Ig) and localized Ig-secreting cells (SC) of various isotypes in acute, persistent/resting memory and effector memory phases following oral versus intra-muscular (IM) immunizations. In the acute phase, both oral and IM immunizations induced high avidity Ig. However, in the persistent/resting memory phase, oral immunizations induced low avidity Ig while IM immunizations induced high avidity Ig. Following oral immunizations, in the persistent/resting memory phase, hapten-specific IgM titers in serum and IgM-SC in bone marrow (BM) dominated the immune response, suggesting an important role for IgM in the maintenance of memory. © 2003 Elsevier Ltd. All rights reserved. Keywords: Mucosa; Memory; Immunoglobulin isotypes 1. Introduction The hallmark of vaccination, through natural infection or immunization, is the induction of long-term immunologi- cal memory. The concept of immunological memory dic- tates that upon exposure to a recall antigen faster and better quality effector functions are induced. Several theories have been put forward to explain the mechanism of induction and maintenance of immunological memory, all of which have as their underlying basis the clonal selection theory, set forth by Jerne and further modified by Burnet [1,2]. B cell “memory” has thus been associated with antigen- encounter followed by Ig isotype switch from IgM to IgG or IgA and affinity maturation following extensive prolif- eration and selection of high affinity clones in germinal centers (GC) [3]. Therefore, by far, most studies have ad- dressed the issue of “memory” in the context of IgG or IgA-bearing/producing B cells in peripheral lymphoid tis- sues either a short time after priming immunizations (acute phase) or after boosting immunizations following a resting period (effector memory phase). Thus, studies pertaining to ∗ Corresponding author. Tel.: +1-510-923-8595; fax: +1-510-923-2586. E-mail address: michael [email protected] (M. Vajdy). the maintenance of memory in the resting memory phase, when the acute phase of the response (particularly against non-replicating antigens) has completely subsided and in the absence of boosting immunizations, remain scarce. Moreover, most studies regarding immunological mem- ory have involved systemic immunization and little data is available on the mechanisms of persistence of “memory” following mucosal immunizations. However, induction and maintenance of immunological memory at mucosal sur- faces is of particular importance since most pathogens enter the host through mucosal membranes and several stud- ies have demonstrated that local immunological responses are required for local protection in mice and primates [4–8]. Studies pertaining to memory at mucosal surfaces have been hampered by the fact that induction of immunity through mucosal immunizations with soluble protein anti- gens in the absence of adjuvants is less achievable compared to systemic immunization and induction of systemic immu- nity [9]. Indeed, in general, oral immunizations with most soluble protein antigens in the absence of adjuvant induce systemic tolerance rather than immunity [9]. Therefore, po- tent mucosally effective adjuvants in the form of bacterial toxins, their detoxified derivatives and mucosal delivery 0264-410X/$ – see front matter © 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2003.10.018
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of Maintenance of long-term immunological memory by low avidity IgM-secreting cells in bone marrow...

Vaccine 22 (2004) 1553–1563
Maintenance of long-term immunological memory by low avidityIgM-secreting cells in bone marrow after mucosal immunizations
with cholera toxin adjuvant
Elawati Soenawana, Indresh Srivastavaa, Soumi Guptaa, Elaine Kana, Ramesh Janania,Jina Kazzaza, Manmohan Singha, Vijay Shreedharb, Michael Vajdya,b,∗
a Chiron Corporation, Immunology and Infectious Diseases, Emeryville, CA, USAb Children’s Hospital and Harvard Medical School, Boston, MA, USA
Received 20 March 2003; received in revised form 1 October 2003; accepted 2 October 2003
Abstract
To understand the mechanisms involved in maintaining long-term immunological memory following mucosal immunizations, wedetermined the quality of serum hapten-specific immunoglobulins (Ig) and localized Ig-secreting cells (SC) of various isotypes in acute,persistent/resting memory and effector memory phases following oral versus intra-muscular (IM) immunizations. In the acute phase,both oral and IM immunizations induced high avidity Ig. However, in the persistent/resting memory phase, oral immunizations inducedlow avidity Ig while IM immunizations induced high avidity Ig. Following oral immunizations, in the persistent/resting memory phase,hapten-specific IgM titers in serum and IgM-SC in bone marrow (BM) dominated the immune response, suggesting an important role forIgM in the maintenance of memory.© 2003 Elsevier Ltd. All rights reserved.
Keywords: Mucosa; Memory; Immunoglobulin isotypes
1. Introduction
The hallmark of vaccination, through natural infection orimmunization, is the induction of long-term immunologi-cal memory. The concept of immunological memory dic-tates that upon exposure to a recall antigen faster and betterquality effector functions are induced. Several theories havebeen put forward to explain the mechanism of induction andmaintenance of immunological memory, all of which haveas their underlying basis the clonal selection theory, set forthby Jerne and further modified by Burnet[1,2].
B cell “memory” has thus been associated with antigen-encounter followed by Ig isotype switch from IgM to IgGor IgA and affinity maturation following extensive prolif-eration and selection of high affinity clones in germinalcenters (GC)[3]. Therefore, by far, most studies have ad-dressed the issue of “memory” in the context of IgG orIgA-bearing/producing B cells in peripheral lymphoid tis-sues either a short time after priming immunizations (acutephase) or after boosting immunizations following a restingperiod (effector memory phase). Thus, studies pertaining to
∗ Corresponding author. Tel.:+1-510-923-8595; fax:+1-510-923-2586.E-mail address: [email protected] (M. Vajdy).
the maintenance of memory in the resting memory phase,when the acute phase of the response (particularly againstnon-replicating antigens) has completely subsided and inthe absence of boosting immunizations, remain scarce.Moreover, most studies regarding immunological mem-ory have involved systemic immunization and little data isavailable on the mechanisms of persistence of “memory”following mucosal immunizations. However, induction andmaintenance of immunological memory at mucosal sur-faces is of particular importance since most pathogens enterthe host through mucosal membranes and several stud-ies have demonstrated that local immunological responsesare required for local protection in mice and primates[4–8].
Studies pertaining to memory at mucosal surfaces havebeen hampered by the fact that induction of immunitythrough mucosal immunizations with soluble protein anti-gens in the absence of adjuvants is less achievable comparedto systemic immunization and induction of systemic immu-nity [9]. Indeed, in general, oral immunizations with mostsoluble protein antigens in the absence of adjuvant inducesystemic tolerance rather than immunity[9]. Therefore, po-tent mucosally effective adjuvants in the form of bacterialtoxins, their detoxified derivatives and mucosal delivery
0264-410X/$ – see front matter © 2003 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2003.10.018

1554 E. Soenawan et al. / Vaccine 22 (2004) 1553–1563
systems have been used to induce local immune responsesto unrelated antigens[9–11].
The model used in this study was established severalyears ago when it was shown long-term immunologicalmemory to unrelated antigens can be induced and main-tained by cholera toxin as adjuvant[12]. Using this model itwas also found that long-term hapten-specific (DNP) mem-ory was maintained by IgM in serum and that it appearedthat this hapten-specific memory required carrier-specific Tcell memory[13,14]. The aim of this study was to furtheradvance our understanding on the mechanisms that underlieimmunological memory following oral immunizations com-pared to systemic immunizations. We investigated the pres-ence of serum IgM as well as IgM-secreting cells (SC) formaintenance of long-term immunological memory follow-ing mucosal compared to systemic immunizations, charac-terized the generated antibodies with regards to their avidityand localized the Ig-secreting cells of various isotypes. Tothis end, we immunized mice orally or intra-muscularly withthe hapten fluorescein isothiocyanate (FITC) conjugated tothe cholera toxin B subunit (CTB) plus trace amounts ofthe CT holotoxin. We then followed the kinetics of the im-mune responses in sera and in various tissues in the acute,persistent resting memory and effector memory phases.
2. Materials and methods
2.1. Mice and immunizations
Female C57bl/6 mice were purchased from Charles RiverBreeding Laboratories at the age of 6–8 weeks at the onset ofthe studies. Groups of three mice were used for each immu-nization and tissues (spleen (SP), bone marrow (BM), andPeyer’s patches (PP)) and sera were assayed individuallyunless stated otherwise inSection 3. Oral immunizationswere performed with 25�g CTB-FITC (Sigma/Aldrich)plus 2.5�g CT (List Laboratories) in 3% bicarbonate bufferin a volume of 0.5 ml. Intra-muscular (IM) immunizationswere performed by injection of 10�g CTB-FITC plus1�g CT in PBS in a volume of 50�l into the right legthigh muscle. Oral and intra-muscular immunizations wereperformed without anesthesia three times at 2–3 week in-tervals. The mice were sacrificed 1 week following the finalimmunization (acute phase), 9–12 months following thefinal immunization (persistent/resting memory phase), orgiven a single booster immunization with CTB-FITC plusCT 10–12 months following the final immunization andsacrificed 1 week later (effector/memory, delete/active). Forthe immunostaining experiments, the mice were given asingle dose of CTB-FITC plus CT and sacrificed 6 h later.
2.2. Sera and tissue collection
Mice were bled through the retro orbital plexus or tails 1day prior to sacrifice and the sera separated for ELISA and
avidity assays. PP were removed from the entire small in-testine (between 6 and 12 per mouse) and meshed through anylon mesh. BM was collected from femur and tibia bonesof each mouse by flushing with PBS and re-suspendingin complete RPMI-1640 medium. SP were harvested andmeshed through a nylon mesh. The tissues were assayedfrom individual mice with three mice per group. Themice were housed and treated either in the vivarium ofChildren’s Hospital in Boston, MA or at Chiron Corpora-tion according to guidelines set forth by Association forAssessment and Accreditation of Laboratory Animal Care(AAALAC) and the United States Department of Agriculture(USDA).
2.3. ELISPOT assay
Single-cell suspensions from pooled PP, SP and BM fromindividual mice were added at concentrations of 0.2–0.8million mononuclear cells per first row in 200�l andtwo-fold serial dilutions onto microtiter 96 well nitrocellu-lose or pvdf plates (Milipore) pre-coated with BSA-FITC(Sigma/Aldrich) (10�g/ml PBS) and blocked with com-plete RPMI-1640 medium at pH 7.2, containing 10% fetalcalf serum (FCS), 5 mM Hepes, and antibiotics. Followingovernight incubation of cells the plates were washed andpolyclonal biotinylated goat-anti mouse IgM, IgG or IgA(Southern Biotechnology Associates, SBA) was added at1:10,000 dilution in PBS/0.1% BSA/0.02% Tween and in-cubated at room temperature (RT) for 2 h. The plates werewashed with PBS/0.02% Tween-20 (P/T) and incubatedfor 1 h at 37◦C with Avidin–peroxidase (Pharmingen) at1:1000 dilution. The plates were washed with P/T and thespots were visualized by adding DAB substrate (Biorad)in Tris–HCl (pH 7.5) buffer for 30 min. The plates werewashed with de-ionized H2O and air-dried. The spots werecounted manually under low magnification. Two to 4 wellsper mouse per tissue were counted. The data are presentedas mean± S.D. of two or three independent experimentswith pools of three individual mice per group and expressedas the number of FITC-specific antibody secreting cells(ASC) per 107 mononuclear cells (MNC).
2.4. ELISA assays
FITC-specific serum IgM, IgG and IgA titers were quan-tified by a standard ELISA assay. Briefly, ELISA plates(96 well U bottom by Nunc Maxisorp) were coated withBSA-FITC (Sigma/Aldrich) protein at 10�g per well inPBS overnight (O/N) at 4◦C. After washing with 1× PBS+ 0.3% Tween-20 (Sigma), the wells were blocked withPBS/5% goat serum/0.3% Tween-20 for 30 min at 37◦Cand sera samples were added in an initial dilution of 1:500for all isotypes and 1.3-fold serial dilutions were performedin the blocking reagent. A standard serum was included ineach assay as positive control. The samples and standardsera were incubated at 4◦C O/N and washed with PBS/0.3%

E. Soenawan et al. / Vaccine 22 (2004) 1553–1563 1555
Tween. The plates were washed and added biotinylatedanti-mouse IgM, IgA or IgG (Southern Biotechnology As-sociates) at 1:10,000 dilution in PBS/2% FCS and incubatedfor 2 h room temperature. The plates were then washed andadded Avidin–horse radish peroxidase (HRP) (Pharmingen)at 1:1000 and incubated at 37◦C for 30 min. The plateswere washed and developed with tetramethylbenzidine(TMB-Kirkegaard and Perry) for 15 min and then stoppedwith 2N HCl. The optical density of each well was mea-sured using a Titertek optical density reader. The titers wereconsidered positive at reciprocal serum dilutions giving anabsorbance of the cut-off line at optical density (OD) of 0.5and above at 450 nm, the lowest detectable titer being 500for all isotypes. The concentrations of total serum IgM, IgGand IgA was also measured a standard ELISA assay. Briefly,ELISA plates (96 well U bottom by Nunc Maxisorp) werecoated with goat anti-mouse Ig (Heavy+ Light chains)(SBA) at 1:10,000 dilution to capture all serum Ig. Afterwashing with 1× PBS + 0.03% Tween-20 (Sigma), thewells were blocked with PBS/2% fetal calf serum for 30 minat 37◦C and sera samples were added in an initial dilutionof 1:50 for IgM, 1:250 for IgA and 1:500 for IgG followedby 1:3 serial dilutions performed in the blocking reagent. Asstandards mouse IgM, IgG and IgA were added at the initialconcentrations of 1, 1, and 5�g/ml respectively. The sam-ples and standards were incubated at 2 h at RT and washedwith PBS/0.3% Tween. Biotinylated anti-mouse IgM, IgAor IgG (Southern Biotechnology Associates (SBA)) wereadded to the plates at 1:10000 dilution in the blockingbuffer and incubated for 2 h room temperature. The plateswere then washed and Avidin–HRP (Pharmingen) at 1:1000dilution was added and incubated at 37◦C for 30 min. Theplates were washed and developed with tetramethylben-zidine (Kirkegaard and Perry) for 5 min and then stoppedwith 2N HCl. The optical density of each well was readby a Titertek optical density reader at 450 nm. The concen-trations were determined by the SOFTmax® PRO software(Sunnyvale, CA).
2.5. Avidity measurement
Antibody avidity index determination was performedusing an ammonium thiocyanate displacement (NH4SCN)ELISA as described elsewhere in detail[15–17], with somemodifications. Briefly, plates were coated with 10�g/mlBSA-FITC in sodium carbonate/bicarbonate buffer pH9.6 at 4◦C, then blocked with 3% BSA containing 0.3%Tween-20 in phosphate buffered saline (PBS pH7.4). Testsera were added at the initial dilution of 100 and incu-bated for 2 h at RT. After incubation with test sera, theplates were washed three times with 0.1% Tween-20 inPBS, pH 7.4. The plates were then added PBS containing0–8 M NH4SCN to rows A through H, respectively andincubated for 20 min. The plates were washed five timeswith 0.1% Tween-20 in PBS at pH 7.4. The wells wereincubated with species-specific secondary antibody coupled
to horse radish peroxidase (anti-mouse 1/50,000, BethylLaboratories, Montgomery, TX) for 1 h at RT. After wash-ing, the plate was developed with TMB and H2O2 (PierceChemical Co., Rockford, IL) for 30 min at RT in the dark.The reaction was stopped with 4N H2SO4 and absorbancevalues were recorded at 450 nm using Power Wave 340×ELISA reader (Bio-TEK Instrument Inc.) and the data wasanalyzed using KC junior software (Bio-Tek InstrumentInc.). All samples were tested in duplicate and results wereexpressed, as NH4SCN concentration required to displace50% of the bound antibodies.
2.6. Immunohistochemistry
PP were carefully removed with scissors from smallintestines of naıve mice or mice orally immunized withCTB-FITC plus CT 6 h earlier (three mice per group and2–3 PP per mouse), embedded in OCT compound (MilesInc., Elkhart, IN) and snap frozen in 2-methylbutane (J.T.Baker, Phillipsburg, NJ) chilled by dry ice. Thin sections(5�m) were cut using a cryostat, transferred onto glassslides, and kept at−80◦C until stained. Before staining,the sections were fixed in ice-cold acetone for 10 min atroom temperature. After blocking in PBS/2% BSA/1%goat serum (Dako, CA) for 1 h, the sections were washedin PBS and stained with the antibodies designated belowat 1:100 (goat anti-IgM-FITC, goat anti-IgA-FITC, ratanti-mouse CD24 (Southern Biotechnology Associates) or1:40 (rat anti-mouse CD11b, rat anti-mouse CD11c) di-luted in the blocking buffer for 1 h at room temperature.The slides were then washed 6–8 times with PBS. Thestained sections on slides were then mounted in Fluor-save (Calbiochem, CA) and photographed using a digitalcamera linked to a fluorescent microscope with FITCand PE filters. The immunostaining results shown arerepresentative of 1–2 PP per mouse and 2–3 mice pergroup.
2.7. Flow cytometric analysis
PP and SP cell suspensions (1 million per staining) wereprepared from pools of five mice as described above andstained for intracellular IgA and IgM as well as for surfaceexpression of IgA and IgM (goat anti-mouse), and for ratanti-mouse CD24 (Southern Biotechnology Associates),CD11b, CD11c and B220 (Pharmingen/BD). All antibodieswere used at a 1:100 dilution. The stained cells were ana-lyzed using a FACSCalibur (BD). The data are representedas average of two groups of five mice each plus standarddeviation.
2.8. Statistical analysis
For statistical analysis, Student’sT-test on paired twosample for means was performed using Microsoft Excel’sstatistical analysis software program.
1556 E. Soenawan et al. / Vaccine 22 (2004) 1553–1563
3. Results
3.1. Acute and memory serum responses following oralimmunizations
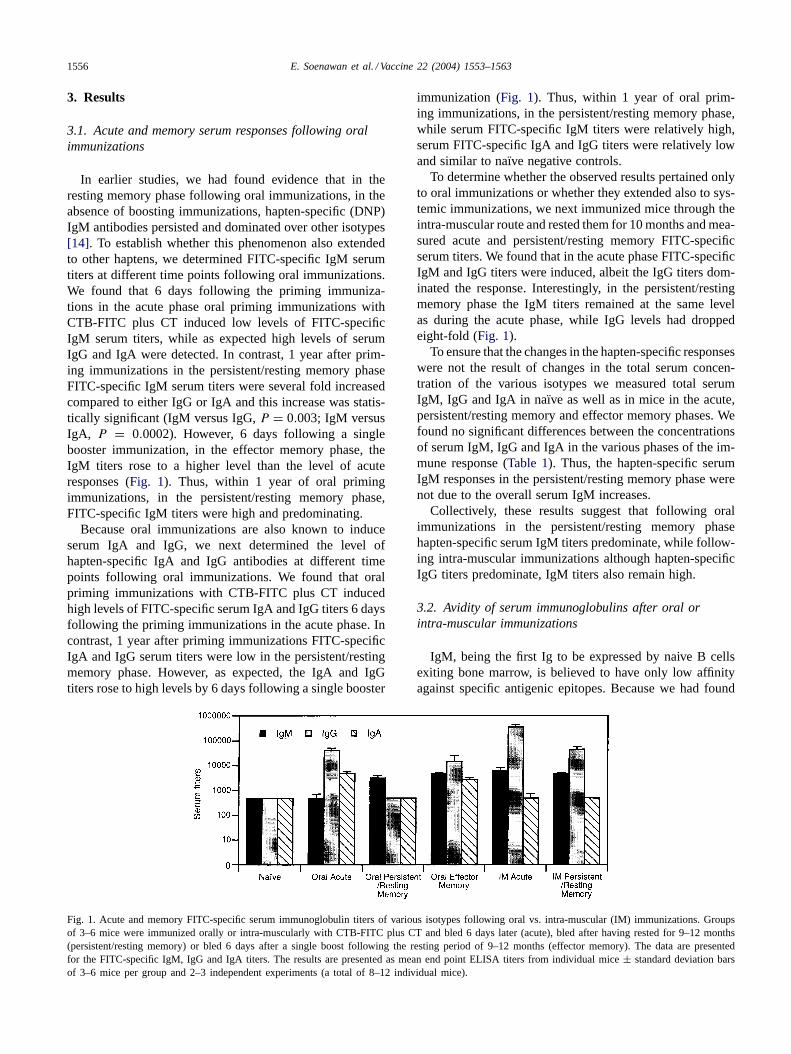
In earlier studies, we had found evidence that in theresting memory phase following oral immunizations, in theabsence of boosting immunizations, hapten-specific (DNP)IgM antibodies persisted and dominated over other isotypes[14]. To establish whether this phenomenon also extendedto other haptens, we determined FITC-specific IgM serumtiters at different time points following oral immunizations.We found that 6 days following the priming immuniza-tions in the acute phase oral priming immunizations withCTB-FITC plus CT induced low levels of FITC-specificIgM serum titers, while as expected high levels of serumIgG and IgA were detected. In contrast, 1 year after prim-ing immunizations in the persistent/resting memory phaseFITC-specific IgM serum titers were several fold increasedcompared to either IgG or IgA and this increase was statis-tically significant (IgM versus IgG,P = 0.003; IgM versusIgA, P = 0.0002). However, 6 days following a singlebooster immunization, in the effector memory phase, theIgM titers rose to a higher level than the level of acuteresponses (Fig. 1). Thus, within 1 year of oral primingimmunizations, in the persistent/resting memory phase,FITC-specific IgM titers were high and predominating.
Because oral immunizations are also known to induceserum IgA and IgG, we next determined the level ofhapten-specific IgA and IgG antibodies at different timepoints following oral immunizations. We found that oralpriming immunizations with CTB-FITC plus CT inducedhigh levels of FITC-specific serum IgA and IgG titers 6 daysfollowing the priming immunizations in the acute phase. Incontrast, 1 year after priming immunizations FITC-specificIgA and IgG serum titers were low in the persistent/restingmemory phase. However, as expected, the IgA and IgGtiters rose to high levels by 6 days following a single booster
Fig. 1. Acute and memory FITC-specific serum immunoglobulin titers of various isotypes following oral vs. intra-muscular (IM) immunizations. Groupsof 3–6 mice were immunized orally or intra-muscularly with CTB-FITC plus CT and bled 6 days later (acute), bled after having rested for 9–12 months(persistent/resting memory) or bled 6 days after a single boost following the resting period of 9–12 months (effector memory). The data are presentedfor the FITC-specific IgM, IgG and IgA titers. The results are presented as mean end point ELISA titers from individual mice± standard deviation barsof 3–6 mice per group and 2–3 independent experiments (a total of 8–12 individual mice).
immunization (Fig. 1). Thus, within 1 year of oral prim-ing immunizations, in the persistent/resting memory phase,while serum FITC-specific IgM titers were relatively high,serum FITC-specific IgA and IgG titers were relatively lowand similar to naıve negative controls.
To determine whether the observed results pertained onlyto oral immunizations or whether they extended also to sys-temic immunizations, we next immunized mice through theintra-muscular route and rested them for 10 months and mea-sured acute and persistent/resting memory FITC-specificserum titers. We found that in the acute phase FITC-specificIgM and IgG titers were induced, albeit the IgG titers dom-inated the response. Interestingly, in the persistent/restingmemory phase the IgM titers remained at the same levelas during the acute phase, while IgG levels had droppedeight-fold (Fig. 1).
To ensure that the changes in the hapten-specific responseswere not the result of changes in the total serum concen-tration of the various isotypes we measured total serumIgM, IgG and IgA in naıve as well as in mice in the acute,persistent/resting memory and effector memory phases. Wefound no significant differences between the concentrationsof serum IgM, IgG and IgA in the various phases of the im-mune response (Table 1). Thus, the hapten-specific serumIgM responses in the persistent/resting memory phase werenot due to the overall serum IgM increases.
Collectively, these results suggest that following oralimmunizations in the persistent/resting memory phasehapten-specific serum IgM titers predominate, while follow-ing intra-muscular immunizations although hapten-specificIgG titers predominate, IgM titers also remain high.
3.2. Avidity of serum immunoglobulins after oral orintra-muscular immunizations
IgM, being the first Ig to be expressed by naive B cellsexiting bone marrow, is believed to have only low affinityagainst specific antigenic epitopes. Because we had found
E. Soenawan et al. / Vaccine 22 (2004) 1553–1563 1557
Table 1Serum IgM, IgG and IgA concentrations during the acute, persis-tent/resting memory and effector memory phases of the immune response
Group name Ig isotype concentration (mg/ml)
IgM IgG IgA
Acute 0.22± 0.03 0.83± 0.2 0.09± 0.02Persistent/resting memory 0.20± 0.06 1.35± 0.3 0.08± 0.02Effector memory 0.28± 0.14 1.31± 0.67 0.085± 0.02Naive 0.29± 0.05 0.91± 0.27 0.08± 0.03
Naive and immunized mice were bled at the indicated immune responsephases as described in the legend toFig. 1. The data are presented inmg/ml and are the mean of three mice per group plus S.D.
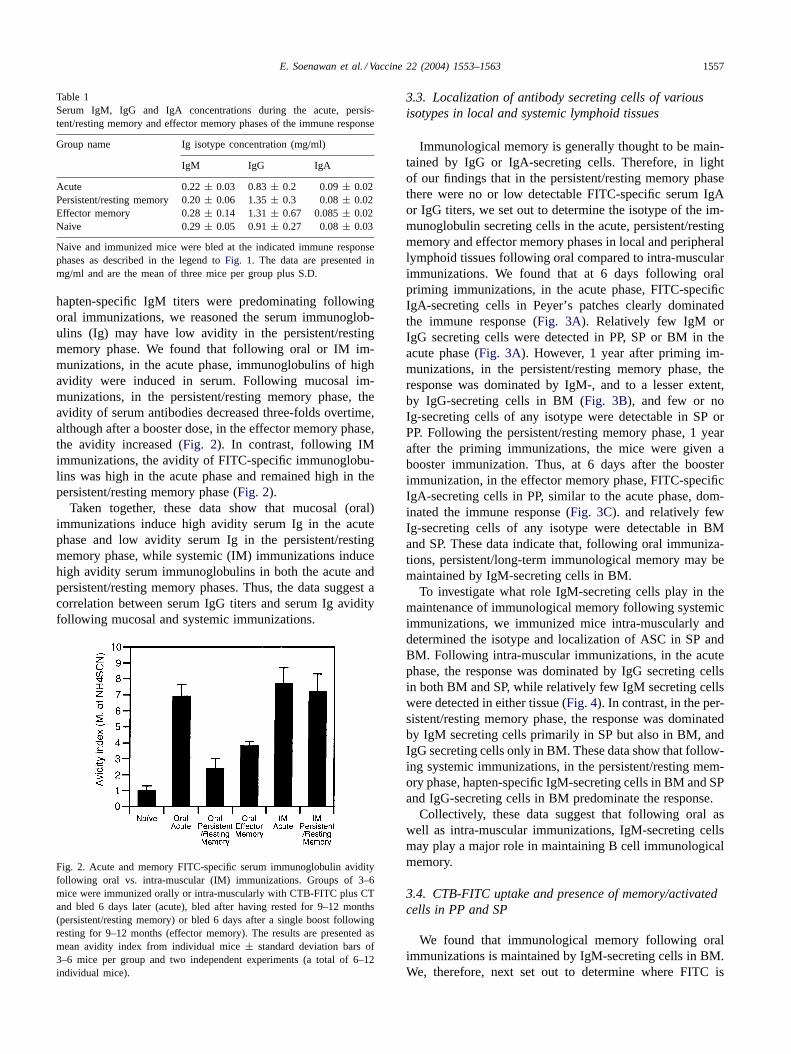
hapten-specific IgM titers were predominating followingoral immunizations, we reasoned the serum immunoglob-ulins (Ig) may have low avidity in the persistent/restingmemory phase. We found that following oral or IM im-munizations, in the acute phase, immunoglobulins of highavidity were induced in serum. Following mucosal im-munizations, in the persistent/resting memory phase, theavidity of serum antibodies decreased three-folds overtime,although after a booster dose, in the effector memory phase,the avidity increased (Fig. 2). In contrast, following IMimmunizations, the avidity of FITC-specific immunoglobu-lins was high in the acute phase and remained high in thepersistent/resting memory phase (Fig. 2).
Taken together, these data show that mucosal (oral)immunizations induce high avidity serum Ig in the acutephase and low avidity serum Ig in the persistent/restingmemory phase, while systemic (IM) immunizations inducehigh avidity serum immunoglobulins in both the acute andpersistent/resting memory phases. Thus, the data suggest acorrelation between serum IgG titers and serum Ig avidityfollowing mucosal and systemic immunizations.
Fig. 2. Acute and memory FITC-specific serum immunoglobulin avidityfollowing oral vs. intra-muscular (IM) immunizations. Groups of 3–6mice were immunized orally or intra-muscularly with CTB-FITC plus CTand bled 6 days later (acute), bled after having rested for 9–12 months(persistent/resting memory) or bled 6 days after a single boost followingresting for 9–12 months (effector memory). The results are presented asmean avidity index from individual mice± standard deviation bars of3–6 mice per group and two independent experiments (a total of 6–12individual mice).
3.3. Localization of antibody secreting cells of variousisotypes in local and systemic lymphoid tissues
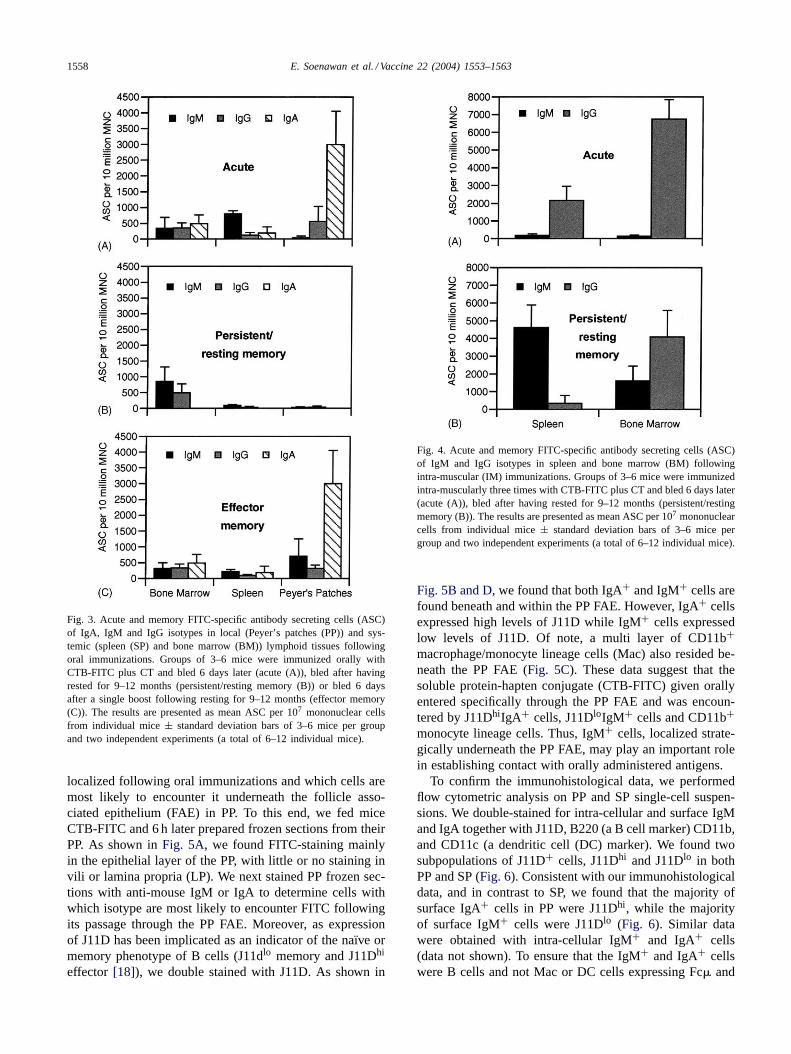
Immunological memory is generally thought to be main-tained by IgG or IgA-secreting cells. Therefore, in lightof our findings that in the persistent/resting memory phasethere were no or low detectable FITC-specific serum IgAor IgG titers, we set out to determine the isotype of the im-munoglobulin secreting cells in the acute, persistent/restingmemory and effector memory phases in local and peripherallymphoid tissues following oral compared to intra-muscularimmunizations. We found that at 6 days following oralpriming immunizations, in the acute phase, FITC-specificIgA-secreting cells in Peyer’s patches clearly dominatedthe immune response (Fig. 3A). Relatively few IgM orIgG secreting cells were detected in PP, SP or BM in theacute phase (Fig. 3A). However, 1 year after priming im-munizations, in the persistent/resting memory phase, theresponse was dominated by IgM-, and to a lesser extent,by IgG-secreting cells in BM (Fig. 3B), and few or noIg-secreting cells of any isotype were detectable in SP orPP. Following the persistent/resting memory phase, 1 yearafter the priming immunizations, the mice were given abooster immunization. Thus, at 6 days after the boosterimmunization, in the effector memory phase, FITC-specificIgA-secreting cells in PP, similar to the acute phase, dom-inated the immune response (Fig. 3C). and relatively fewIg-secreting cells of any isotype were detectable in BMand SP. These data indicate that, following oral immuniza-tions, persistent/long-term immunological memory may bemaintained by IgM-secreting cells in BM.
To investigate what role IgM-secreting cells play in themaintenance of immunological memory following systemicimmunizations, we immunized mice intra-muscularly anddetermined the isotype and localization of ASC in SP andBM. Following intra-muscular immunizations, in the acutephase, the response was dominated by IgG secreting cellsin both BM and SP, while relatively few IgM secreting cellswere detected in either tissue (Fig. 4). In contrast, in the per-sistent/resting memory phase, the response was dominatedby IgM secreting cells primarily in SP but also in BM, andIgG secreting cells only in BM. These data show that follow-ing systemic immunizations, in the persistent/resting mem-ory phase, hapten-specific IgM-secreting cells in BM and SPand IgG-secreting cells in BM predominate the response.
Collectively, these data suggest that following oral aswell as intra-muscular immunizations, IgM-secreting cellsmay play a major role in maintaining B cell immunologicalmemory.
3.4. CTB-FITC uptake and presence of memory/activatedcells in PP and SP
We found that immunological memory following oralimmunizations is maintained by IgM-secreting cells in BM.We, therefore, next set out to determine where FITC is
1558 E. Soenawan et al. / Vaccine 22 (2004) 1553–1563
Fig. 3. Acute and memory FITC-specific antibody secreting cells (ASC)of IgA, IgM and IgG isotypes in local (Peyer’s patches (PP)) and sys-temic (spleen (SP) and bone marrow (BM)) lymphoid tissues followingoral immunizations. Groups of 3–6 mice were immunized orally withCTB-FITC plus CT and bled 6 days later (acute (A)), bled after havingrested for 9–12 months (persistent/resting memory (B)) or bled 6 daysafter a single boost following resting for 9–12 months (effector memory(C)). The results are presented as mean ASC per 107 mononuclear cellsfrom individual mice± standard deviation bars of 3–6 mice per groupand two independent experiments (a total of 6–12 individual mice).
localized following oral immunizations and which cells aremost likely to encounter it underneath the follicle asso-ciated epithelium (FAE) in PP. To this end, we fed miceCTB-FITC and 6 h later prepared frozen sections from theirPP. As shown inFig. 5A, we found FITC-staining mainlyin the epithelial layer of the PP, with little or no staining invili or lamina propria (LP). We next stained PP frozen sec-tions with anti-mouse IgM or IgA to determine cells withwhich isotype are most likely to encounter FITC followingits passage through the PP FAE. Moreover, as expressionof J11D has been implicated as an indicator of the naıve ormemory phenotype of B cells (J11dlo memory and J11Dhi
effector [18]), we double stained with J11D. As shown in
Fig. 4. Acute and memory FITC-specific antibody secreting cells (ASC)of IgM and IgG isotypes in spleen and bone marrow (BM) followingintra-muscular (IM) immunizations. Groups of 3–6 mice were immunizedintra-muscularly three times with CTB-FITC plus CT and bled 6 days later(acute (A)), bled after having rested for 9–12 months (persistent/restingmemory (B)). The results are presented as mean ASC per 107 mononuclearcells from individual mice± standard deviation bars of 3–6 mice pergroup and two independent experiments (a total of 6–12 individual mice).
Fig. 5B and D, we found that both IgA+ and IgM+ cells arefound beneath and within the PP FAE. However, IgA+ cellsexpressed high levels of J11D while IgM+ cells expressedlow levels of J11D. Of note, a multi layer of CD11b+macrophage/monocyte lineage cells (Mac) also resided be-neath the PP FAE (Fig. 5C). These data suggest that thesoluble protein-hapten conjugate (CTB-FITC) given orallyentered specifically through the PP FAE and was encoun-tered by J11DhiIgA+ cells, J11DloIgM+ cells and CD11b+monocyte lineage cells. Thus, IgM+ cells, localized strate-gically underneath the PP FAE, may play an important rolein establishing contact with orally administered antigens.
To confirm the immunohistological data, we performedflow cytometric analysis on PP and SP single-cell suspen-sions. We double-stained for intra-cellular and surface IgMand IgA together with J11D, B220 (a B cell marker) CD11b,and CD11c (a dendritic cell (DC) marker). We found twosubpopulations of J11D+ cells, J11Dhi and J11Dlo in bothPP and SP (Fig. 6). Consistent with our immunohistologicaldata, and in contrast to SP, we found that the majority ofsurface IgA+ cells in PP were J11Dhi, while the majorityof surface IgM+ cells were J11Dlo (Fig. 6). Similar datawere obtained with intra-cellular IgM+ and IgA+ cells(data not shown). To ensure that the IgM+ and IgA+ cellswere B cells and not Mac or DC cells expressing Fc� and

E. Soenawan et al. / Vaccine 22 (2004) 1553–1563 1559
Fig. 5. Localization of FITC and cells beneath and within the follicle associated epithelium (FAE) overlying Peyer’s patches (PP). Mice were given asingle oral dose of CTB-FITC plus CT and 6 h later frozen sections were prepared to localize FITC staining (A). Frozen sections (5�m) were preparedfrom naive mice and double stained with anti-J11D (red) and anti-IgA (green) (B) or anti-J11D (red) and anti-IgM (green) (D) or anti-CD11b (red) (C).The shown results are representative of similar results from three mice and three independent staining experiments.
Fig. 6. The majority of IgM+ B cells in Peyer’s patches (PP) and spleen (SP) are J11Dlo. Single-cell suspensions were prepared from PP and SP andflow cytometric analysis was performed by double staining for surface IgM or IgA together with the J11D antibody. Two populations of J11Dlo andJ11Dhi cells are seen in both PP and SP. The histograms show the relative percentage in PP of IgM+ (A) and IgA+ (B) cells that are J11Dlo or J11Dhi,and the relative percentage in SP of IgM+ (C) and IgA+ (D) cells that are J11Dlo or J11Dhi. The data are representative of two experiments from twopools of five mice each with identical results.

1560 E. Soenawan et al. / Vaccine 22 (2004) 1553–1563
Fig. 7. The majority surface IgM+ and IgA+ cells in PP and SP are B200+ and not CD11b+ or CD11c+. Single-cell suspensions were prepared fromPP and SP and flow cytometric analysis was performed by double staining for surface IgM or IgA together with B220, CD11b or CD11c. The data arerepresented as means of relative percentage of double positive cells from two pools of five mice each. The S.D. between each pool of five mice eachwas less than 0.5% and is not shown.
Fc� receptors on their surfaces, we double stained for sur-face IgA and IgM together with B220, CD11b and CD11c.We found that by far the majority of surface IgA+ andIgM+ cells were B220+ and relatively very few CD11b+orCD11c+cells expressed IgA or IgM in both PP and SP(Fig. 7). These data show that the majority of J11Dlo cells inPP and SP are IgM+B220+ cells and that the IgM+J11Dlo
and IgA+J11Dhi populations in PP, and below FAE, aremost likely B cells and not macrophages or DC.
4. Discussion
The aims of this study were three-fold. Firstly, we soughtto determine serum Ig of various isotypes in the persis-tent/resting memory phase following oral versus systemicimmunizations using cholera toxin adjuvant. Secondly, inorder to determine whether the humoral response was main-tained by high or low affinity Ig we characterized the serumresponses with regards to their avidity. Thirdly, we aimedto localize the Ig-secreting cells of various istoypes to de-termine where they are generated and maintained and what
isotype they secrete. It is important to note that since dif-ferent reagents with different binding affinities were usedfor detection of antigen-specific Ig concentration of vari-ous isotypes in serum or tissues the differences in the IgM,IgG and IgA responses reflect relative and not absolutechanges.
We found that following oral immunizations hapten-specific IgM was the dominant isotype in the persis-tent/resting memory phase. This is a confirmation of pre-vious findings in which, following oral immunizationswith DNP-KLH plus CT adjuvant serum responses toanother hapten, DNP, was also dominated by IgM[14].Early studies also suggested a strong role of IgM in thegeneration or maintenance of memory following systemicimmunizations[19–21]. In general, relatively little infor-mation is available on the isotype distribution of serum Igor Ig-secreting cells in various tissues in long-term (over3 months) memory studies. Moreover, most of the studiesfocusing on the isotype distribution pattern of serum ortissue Ig have involved infection with replicating pathogensand therefore a multiplicity of antigenic epitopes[22,23].From these studies it has been deduced that isotypes other

E. Soenawan et al. / Vaccine 22 (2004) 1553–1563 1561
than IgM maintain the B cell memory response[22,23].However, studies involving immunization with solublehapten-carrier antigens, have suggested that memory typeresponses are maintained by dual expressing cells thatmay or may not express IgM (IgM+/IgG− or IgM−/IgG−or IgM+/IgG+) [24]. Recent strong evidence for a majorrole of IgM in memory type responses was provided byemploying RT-PCR to detect v-region mRNA transcriptsspecific for the hapten phenyl-oxazolone, following sys-temic immunizations[25]. Interestingly, it has been shownthat human splenic IgM transcripts are mutated at highfrequency[26] indicating that long-term memory can bemaintained by high affinity IgM. Thus, collectively, thesedata suggest that IgM is a major Ig isotype in memory typeresponses to non-replicating soluble haptenic antigens fol-lowing either mucosal, as we have shown here, or systemicimmunizations.
A major attribute of secondary/memory type Ig responsesis believed to be an increase in affinity through the pro-cess of somatic hypermutation during the proliferation ofB cells in germinal centers[27]. However, there have beenconflicting reports stating that somatic hypermutation maynot take place in primary antibody responses[28] or maytake place during secondary antibody responses but with-out further increase in the overall avidity[29]. Moreover, ithas been shown that somatic hypermutation may take placeindependent of presence of GC[29,30]. The above resultshave pertained to systemic immunizations and not to mu-cosal immunizations. In this study, we showed that in theacute phase both oral and systemic immunizations inducedhigh avidity serum Ig during the acute phase. However, inthe persistent/resting memory phase following oral immu-nizations only low avidity serum Ig was present, while fol-lowing systemic immunizations high avidity serum Ig waspresent. It may be hypothesized that in the acute phase, i.e. 1week after the third oral or systemic immunization (bearingin mind that there was only 10 days between each immu-nization) the overall avidity of the Ig was high. However,after oral immunizations in the persistent/resting memoryphase the predominant isotype was IgM and as indirectlyshown by us these IgM antibodies had low avidity. This sug-gests that the lack of a predominant IgA or IgG responsecorrelated with the low avidity of the response. In contrast,after systemic immunizations in the persistent/resting mem-ory phase, in the presence of high levels of both IgG andIgM the avidity was high again supporting the notion thatthe process of somatic hypermutation and isotype switch-ing are closely regulated. However, it should also be notedthat low titers of high avidity hapten-specific IgG will notbe detectable in the presence of high titers of high avidityhapten-specific IgM.
It is well known that mucosal (oral or intra-nasal) im-munizations with most soluble protein antigens can inducea state of systemic unresponsiveness known as oral toler-ance. From our data, it would appear that the avidity, whichin functional terms may determine neutralization of anti-
gens, differed following oral compared to systemic immu-nizations. Assuming the low avidity of the serum Ig afteroral immunizations was due to the absence of high levelsof IgG and IgA it may be deduced that the process of iso-type switching ceases rapidly after the acute phase. As theresponse to the hapten in the hapten-carrier model systemis dependent on cognate T helper responses following oralimmunizations[14] it may be deduced that the low levelsof IgG and IgA in the persistent/resting memory phase fol-lowing oral immunizations reflects presence of strong T cellsuppressor activity by CD8+ T cells in the gut mucosa asevidenced by two independent reports[31,32]. Due to thelow quantities of sera obtained from the mice we were un-able to address the issue of affinity or avidity residing inIgM, IgA and IgG fractions of sera independently. Thus, inthe persistent/resting memory phase, oral immunizations in-duced relatively low avidity serm Ig of which IgM is thepredominant isotype.
In this study, we found that BM was a major site of Igproduction in the persistent/resting memory phase follow-ing either mucosal or systemic immunizations. BM, whichis considered a primary lymphoid tissue producing naive Band T cells, has also been appreciated as a major site of Igproduction during acute phases. A role of murine bone mar-row in the maintenance of memory B cells following oralimmunizations was suggested some years ago[33] as wellas more recently[34]. Following systemic immunizations,it is believed that antigen-activation and proliferation of Bcells occurs in lymph nodes (LN) and once activated the Bcells migrate to BM as PNAhi IgG secreting blast cells about3 days after a secondary immunization[27]. However, themigration of IgM-secreting cells from LN to BM and theirphenotype has not been addressed and more importantly itis not clear why plasma cells would be found mainly in BMin the resting memory phase. If there was an active migra-tion of activated B cells to BM, we would expect to findthe activated B cells which secrete detectable Ig in SP/PPin the persistent/resting memory phase. This we indeedfound following systemic immunizations. However, sincefollowing oral immunizations, we detected hapten-specificIgM-secreting cells in BM and very low numbers ofhapten-specific IgA-secreting cells in PP this would meanthat in the persistent/resting memory phase there is produc-tion of hapten-specific IgM in BM independent of PP. Thus,long-term hapten specific responses in the persistent/restingmemory phase reside within the BM and SP following sys-temic immunizations and in BM following oral immuniza-tions and IgM is a major contributing isotype at this site.
Few markers have been found to define memory B cells.The J11D antibody which recognizes the heat stable antigen(HSA), CD24, has been proposed as a marker for memoryB cells in that putative memory B cells were found to ex-press low levels of this marker[18,35]. However, J11Dlo
cells become J11Dhi within the first week of secondaryimmunizations, and later they down regulate expression ofJ11D [18]. At any rate, assuming that low expression of

1562 E. Soenawan et al. / Vaccine 22 (2004) 1553–1563
J11D delineates putative memory B cells, we reasoned thatthis cell population would be strategically placed below thePP FAE to re-encounter the specific antigen. We found thatboth IgM+ and IgA+ cells were found within as well asbelow the FAE. However, IgM+ cells expressed low levelsof J11D, delineating a putative memory phenotype, whileIgA+ cells expressed high levels of J11D, delineating anon-memory B cell population. Moreover, the majority ofJ11Dlo cells in PP were IgM+ as found by our flow cyto-metric analysis. These finding are in contrast with previousreports where J11Dlo cells were IgG+, and not IgM+, cells[18], suggesting an inherent difference between mucosaland systemic routes of B cell activation. Taken together, inconjunction with the pre-dominanting presence of serumIgM and IgM-secreting cells in BM in the persistent/restingmemory phase, these data suggest an important role also forIgM+J11Dlo cells in the local mucosal inductive site of PP.
It may be suggested that the IgM-secreting B cell popu-lation that maintained memory in BM or PP were from theB-1 cell lineage. However, several lines of evidence wouldargue against this suggestion. B-1 cells are found primarilyin the peritoneal cavity and they are thought to contributeto the IgA-secreting pool of B cells in the gut lamina pro-pria [36,37]. However, there is no evidence to suggest thatin the adult animal B-1 cells are bone marrow derived orhome to BM. Indeed, it is believed that B-1 cells representa primordial B cell lineage, with a restricted antibody generepertoire that are self-regenerating in the peritoneal cavitydriven by self or environmental antigens[3,37,38]. More-over, B-1 cells are thought to participate in responses toT-independent antigens[39].
If long-term immunological memory is maintained byantigen-specific IgM-secreting cells these cells may be ei-ther long-lived or they may arise from new clones of Bcells exiting the BM. The former scenario would imply thata subset of antigen-specific IgM-secreting cells with highaffinity do not undergo isotype switch differentiation and re-main dormant until re-activated with the recall antigen. Thelatter scenario, however, would imply that the BM gener-ates new clones of antigen-specific IgM-secreting cells withhigh affinity that can presumably switch to downstream iso-types depending on the microenvironment. However, sincewe found low avidity serum anti-hapten responses in thepersistent/resting memory phase, we deem the presence ofhigh affinity new clones of BM-derived memory B cells un-likely. Thus, if immunological memory is maintained by lowaffinity new clones of BM-derived IgM-secreting B cellsthe question is whether specific new B cell clones or allnew B cells carry memory. In either case, the number andthe quality of contacts of IgM with an antigenic epitopeappears to dictate memory. To demonstrate the validity ofthese hypotheses, the affinity of hapten-specific IgM anti-bodies a long time after immunizations and the presence ofisotype-switched (IgG and IgA) memory responses in theabsence of IgM+ or IgM secreting cells should be deter-mined. Future studies will address these issues.
Acknowledgements
We thank Drs. John Donnelly and Nick Valiante for crit-ical reading of the manuscript, and Drs. Marian Neutra andDerek O’Hagan for their support of this work.
References
[1] Jerne N. The natural-selection theory of antibody formation. ProcNatl Acad Sci USA 1955;41:849–57.
[2] Burnet F. A modification of Jerne’s theory of antibody productionusing the concept of clonal selection. Aust J Sci 1957;20:67–9.
[3] Rajewsky K. Clonal selection and learning in the antibody system.Nature 1996;381(6585):751–8.
[4] Gallichan W, Rosenthal K. Long-lived cytotoxic T lymphocytememory in mucosal tissues after mucosal but not systemicimmunization. J Exp Med 1996;184(5):1879–90.
[5] Parr EL, Parr MB. Immunoglobulin G, plasma cells, and lymphocytesin the murine vagina after vaginal or parenteral immunization withattenuated herpes simplex virus type 2. J Virol 1998;72(6):5137–45.
[6] Kaul R, Trabattoni D, Bwayo JJ, Arienti D, Zagliani A, MwangiFM, et al. HIV-1-specific mucosal IgA in a cohort of HIV-1-resistantKenyan sex workers. AIDS 1999;13(1):23–9.
[7] Bernstein DI. Effect of route of vaccination with vaccinia virusexpressing HSV-2 glycoprotein D on protection from genital HSV-2infection. Vaccine 2000;18(14):1351–8.
[8] Vajdy M, Gardner J, Neidleman J, Cuadra L, Greer C, Perri S,et al. Human immunodeficiency virus type 1 Gag-specific vaginalimmunity and protection after local immunizations with sindbisvirus-based replicon particles. J Infect Dis 2001;184(12):1613–6.
[9] McGhee JR, Czerkinsky C, Mestecky J. Mucosal vaccines: anoverview. In: Ogra PL, Lamm ME, Bienenstock J, Mestecky J,Strober W, McGhee JR, editors. Mucosal immunology. San Diego:Academic Press; 1999. p. 741–57.
[10] Michalek SM, O’Hagan DT, Gould-Fogerite S, RimmelzwaanGF, Osterhaus ADME. Antigen delivery systems: nonlivingmicroparticles, liposomes, cochleates, and ISCOMS. In: Ogra PL,Mestecky J, Lamm ME, Strober W, Bienenstrock J, McGhee JR,editors. Mucosal immunology. San Diego: Academic Press; 1999.p. 759–78.
[11] Hantman MJ, Hohmann EL, Murphy CG, Knipe DM, Miller SI.Antigen delivery systems: development of recombinant live vaccinesusing viral or bacterial vectors. In: Ogra PL, Mestecky J, LammME, Strober W, Bienenstrock J, McGhee JR, editors. Mucosalimmunology. San Diego: Academic Press; 1999. p. 779–91.
[12] Vajdy M, Lycke NY. Cholera toxin adjuvant promotes long-termimmunological memory in the gut mucosa to unrelated immunogensafter oral immunization. Immunology 1992;75(3):488–92.
[13] Vajdy M, Lycke N. Stimulation of antigen-specific T- and B-cellmemory in local as well as systemic lymphoid tissues followingoral immunization with cholera toxin adjuvant. Immunology1993;80(2):197–203.
[14] Vajdy M, Lycke N. Mucosal memory B cells retain the ability toproduce IgM antibodies 2 years after oral immunization. Immunology1995;86(3):336–42.
[15] Charoenvit Y, Sedegah M, Yuan LF, Gross M, Cole C, Bechara R,et al. Active and passive immunization againstPlasmodium yoeliisporozoites. Bull World Health Organ 1990;68(Suppl):26–32.
[16] Luxton RW, Thompson EJ. Affinity distributions of antigen-specificIgG in patients with multiple sclerosis and in patients with viralencephalitis. J Immunol Methods 1990;131(2):277–82.
[17] Srivastava IK, Stamatatos L, Legg H, Kan E, Fong A, CoatesSR, et al. Purification and characterization of oligomeric envelopeglycoprotein from a primary r5 subtype B human immunodeficiencyvirus. J Virol 2002;76(6):2835–47.

E. Soenawan et al. / Vaccine 22 (2004) 1553–1563 1563
[18] Yin X-M, Lee WT, Vitetta ES. Changes in expression of J11d onmurine B cells during activation and generation of memory. CellImmunol 1991;137(2):448–60.
[19] Abney ER, Keeler KD, Parkhouse RM, Willcox HN. ImmunoglobulinM receptors on memory cells of immunoglobulin G antibody-formingcell clones. Eur J Immunol 1976;6(6):443–50.
[20] Strober S. Maturation of b lymphocytes in rates. III. Twosubpopulations of memory B cells in the thoracic duct lymph differby size, turnover rate, and surface immunoglobulin. J Immunol1976;117(4):1288–94.
[21] Zan-Bar I, Strober S, Vitetta ES. The relationship betweensurface immunoglobulin isotype and immune function of murine Blymphocytes. I. Surface immunoglobulin isotypes on primed B cellsin the spleen. J Exp Med 1977;45:1188–205.
[22] Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunitydue to long-lived plasma cells. Immunity 1998;8:363–72.
[23] Sangster M, Hyland L, Sealy R, Coleclough C. Distinctive kinetics ofthe antibody-forming cell response to Sendai virus infection of micein different anatomical compartments. Virology 1995;207:287–91.
[24] Wu CJ, Karttunen JT, Chin DH, Sen D, Gilbert W. Murine memoryB cells are multi-isotype expressors. Immunology 1991;72(1):48–55.
[25] White H, Gray D. Analysis of immunoglobulin (Ig) isotype diversityand IgM/D memory in the response to phenyl-oxazolone. J Exp Med2000;191(12):2209–19.
[26] Insel RA, Varade WS, Marin E. Human splenic IgM immunoglobulintranscripts are mutated at high frequency. Mol Immunol1994;31(5):383–92.
[27] Tew JG, di Losa RM, Burton GF, Kosco MH, Kupp LI, MasudaA, et al. Germinal centers and antibody production in bone marrow.Immunol Rev 1992;126:99–112.
[28] Kalinke U, Oxenius A, Lopez-Macias C, Zinkernagel RM,Hengartner H. Virus neutralization by germ-line vs. hypermutatedantibodies. Proc Natl Acad Sci USA 2000;97(18):10126–31.
[29] Kalinke U, Bucher EM, Ernst B, Oxenius A, Roost H-P, GeleyS, et al. The role of somatic mutation in the generation of the
protective humoral immune response against vesicular stomatitisvirus. Immunity 1996;5:639–52.
[30] Matsumoto M, Lo SF, Carruthers CJ, Min J, Mariathasan S,Huang G, et al. Affinity maturation without germinal centres inlymphotoxin-alpha-deficient mice. Nature 1996;382(6590):462–6.
[31] Hornquist E, Grdic D, Mak T, Lycke N. CD8-deficient mice exhibitaugmented mucosal immune responses and intact adjuvant effects tocholera toxin. Immunology 1996;87(2):220–9.
[32] Holland GP, Steward MW. Antibody affinity maturation: the role ofCD8+ cells. Clin Exp Immunol 1989;78(3):488–93.
[33] Alley CD, Kiyono H, McGhee JR. Murine bone marrow IgAresponses to orally administered sheep erythrocytes. J Immunol1986;136(12):4414–9.
[34] Benedetti R, Lev P, Massouh E, Flo J. Long-term antibodies afteran oral immunization with cholera toxin are synthesized in thebone marrow and may play a role in the regulation of memoryB-cell maintenance at systemic and mucosal sites. Res Immunol1998;149(2):107–18.
[35] Linton P-J, Decker DJ, Klinman NR. Primary antibody-forming cellsand secondary B cells are generated from separate precursor cellsubpopulations. Cell 1989;59:1049–59.
[36] Kroese FG, Butcher EC, Stall AM, Lalor PA, Adams S, HerzenbergLA. Many of the IgA producing plasma cells in murine gut arederived from self-replenishing precursors in the peritoneal cavity. IntImmunol 1989;1(1):75–84.
[37] Hardy RR, Carmack CE, Li YS, Hayakawa K. Distinctivedevelopmental origins and specificities of murine CD5+ B cells.Immunol Rev 1994;137:91–118.
[38] Hardy RR, Carmack CE, Shinton SA, Riblet RJ, Hayakawa K.A single VH gene is utilized predominantly in anti-BrMRBChybridomas derived from purified Ly-1 B cells: definition of theVH11 family. J Immunol 1989;142(10):3643–51.
[39] Forster I, Rajewsky K. Expansion and functional activity of Ly-1+ Bcells upon transfer of peritoneal cells into allotype-congenic, newbornmice. Eur J Immunol 1987;17(4):521–8.




















