
Low Avidity Recognition of Self-Antigen by T Cells Permits Escape from Central Tolerance
9
Immunity, Vol. 3, 407-415, October, 1995, Copyright 0 1995 by Cell Press Low Avidity Recognition of Self-Antigen by T Cells Permits Escape from Central Tolerance George Y. Liu,‘ll Paul J. Fairchild,*11 Richard M. Smith,” John R. Prowle,’ Dimitris Kioussis,f and David C. Wraith5 *Cambridge University Department of Pathology Tennis Court Road Cambridge CB2 1QP England fLaboratory of Molecular Immunology National Institute for Medical Research The Ridgeway, Mill Hill London NW7 1AA England Summary The immunodominant epitope of myelin basic protein, Ad -9, is encephalitogenic in H-2” mice. We have pre- viously demonstrated that this epitope displays low affinity for I-A” and have suggested that the avidity of T cell recognition in the thymus may be compromised, enabling autoreactive T cells to escape self-tolerance. We have addressed this hypothesis directly by con- structing transgenic mice expressing an encephalito- genie T cell receptor (TCR). Parenteral admininstration of Acl-9 had no discernable impact on developing thy- mocytes. In contrast, peptide analogs displaying far higher affinity for I-A”, provoked deletion of CD4 + CD8 + cells and transient down-regulation of the TCR by ma- ture CD4+CD8- thymocytes. The use of analogs of in- termediate affinity permitted a margin of error to be defined for the induction of tolerance and confirmed that the affinity of Acl-9 lies well below the critical threshold. Introduction The phenomenon of autoreactivity suggests that immuno- logical self-tolerance is far from absolute, sparing some T cells in spite of their anti-self mandate. Accordingly, T cells specific for various self-components may be readily detected in the periphery of otherwise healthy individuals and laboratory animals (Schluesener and Wekerle, 1985; Karachunski et al., 1994). Several strategies have been proposed to explain how such autoaggressive T cells pass undetected during repertoire selection in the thymus. Pre- sentation of an autoantigen at a concentration too low to promote self-tolerance has been invoked for some self- *Present address: Sir William Dunn School of Pathology, University of Oxford, South Parks Road, Oxford, OX1 3RE, England. SPresent address and address for correspondence: Department of Pathology and Microbiology, School of Medical Sciences, University of Bristol, Bristol, BSQ lTD, England. IfThese authors contributed equally to this work. proteins (Cibotti et al., 1992) and may contribute to the poor tolerogenicity of thyroglobulin, one of the autoanti- gens implicated in the aetiology of thyroiditis (Dawe et al., 1993). In contrast, the cryptic epitopes of an autoantigen may never be presented intrathymically due to limitations inherent in antigen processing (Mamula, 1993); the erro- neous induction of enzymes not normally involved in pro- cessing may, however, secure their presentation in the periphery, where T cell activation may ensue. Alterna- tively, sequestration of self-components in an immunologi- cally privileged site, such as the anterior chamber of the eye, may likewise prevent their presentation to developing thymocytes. It is commonly believed that the blood-brain barrier is responsible for rendering the CNS just such a secluded site from which potential autoantigens are denied access to the thymic microenvironment. The consequent inability of negative selection to purge the peripheral repertoire of T cells specific for self-components such as myelin basic protein (MBP) may be readily demonstrated in susceptible strains of experimental animals: administration of either spinal cord homogenate or purified MBP in an immuno- genie fashion results in the inflammatory condition, experi- mental autoimmune encephalomyelitis (EAE), bearing many of the hallmarks of multiple sclerosis in humans. Although the sequestration of MBP is widely believed to contribute to the lack of self-tolerance to this autoantigen, lymph draining the CNS is known to carry such proteins to the lymphoid tissue (Cserr and Knopf, 1992) where it may be presented to both the developing and peripheral Tcell repertoire. Furthermore, recent studies havedemon- strated that MBP is expressed in the fetal thymus from the earliest stages of ontogeny (Pribyl et al., 1993; Mathisen et al., 1993) consistent with its full participation in the induction of self-tolerance. Our recent characterization of an immunodominant epitopeof MBP hassuggested apos- sible solution to this paradox. Encephalitogenic T cells from H-2” mice recognize the acetylated N-terminal peptide of MBP, Acl-9 (Zamvil et al., 1986). We and others have recently demonstrated that this epitope displays very low affinity for its restriction ele- ment, I-A”, forming an interaction so transient as to prevent the measurement of its half-life in vitro (Fairchild et al., 1993; Mason and McConnell, 1994; Mason et al., 1995). This low affinity is a function of the principal anchor resi- due, Lys4, which, being nonhydrophobic and of limited size, fits poorly into the major hydrophobic pocket of I-A” (Fairchild et al., 1994). These findings suggest that the low affinity interaction between Acl-9 and class II MHC may undermine the efficacy of its presentation in vivo, enabling autoreactive T cells to escape self-tolerance. We have tested this hypothesis directly by constructing mice transgenic for an encephalitogenic TCR and assessing the impact on the developing T cell repertoire of either Acl-9 or peptide analogs designed to exhibit higher affin- ity for I-A”.
Transcript of Low Avidity Recognition of Self-Antigen by T Cells Permits Escape from Central Tolerance

Immunity, Vol. 3, 407-415, October, 1995, Copyright 0 1995 by Cell Press
Low Avidity Recognition of Self-Antigen by T Cells Permits Escape from Central Tolerance
George Y. Liu,‘ll Paul J. Fairchild,*11 Richard M. Smith,” John R. Prowle,’ Dimitris Kioussis,f and David C. Wraith5 *Cambridge University Department of Pathology Tennis Court Road Cambridge CB2 1QP England fLaboratory of Molecular Immunology National Institute for Medical Research The Ridgeway, Mill Hill London NW7 1AA
England
Summary
The immunodominant epitope of myelin basic protein, Ad -9, is encephalitogenic in H-2” mice. We have pre- viously demonstrated that this epitope displays low affinity for I-A” and have suggested that the avidity of T cell recognition in the thymus may be compromised, enabling autoreactive T cells to escape self-tolerance. We have addressed this hypothesis directly by con- structing transgenic mice expressing an encephalito- genie T cell receptor (TCR). Parenteral admininstration of Acl-9 had no discernable impact on developing thy- mocytes. In contrast, peptide analogs displaying far higher affinity for I-A”, provoked deletion of CD4 + CD8 + cells and transient down-regulation of the TCR by ma- ture CD4+CD8- thymocytes. The use of analogs of in- termediate affinity permitted a margin of error to be defined for the induction of tolerance and confirmed that the affinity of Acl-9 lies well below the critical threshold.
Introduction
The phenomenon of autoreactivity suggests that immuno- logical self-tolerance is far from absolute, sparing some T cells in spite of their anti-self mandate. Accordingly, T cells specific for various self-components may be readily detected in the periphery of otherwise healthy individuals and laboratory animals (Schluesener and Wekerle, 1985; Karachunski et al., 1994). Several strategies have been proposed to explain how such autoaggressive T cells pass undetected during repertoire selection in the thymus. Pre- sentation of an autoantigen at a concentration too low to promote self-tolerance has been invoked for some self-
*Present address: Sir William Dunn School of Pathology, University of Oxford, South Parks Road, Oxford, OX1 3RE, England.
SPresent address and address for correspondence: Department of Pathology and Microbiology, School of Medical Sciences, University of Bristol, Bristol, BSQ lTD, England.
IfThese authors contributed equally to this work.
proteins (Cibotti et al., 1992) and may contribute to the poor tolerogenicity of thyroglobulin, one of the autoanti- gens implicated in the aetiology of thyroiditis (Dawe et al., 1993). In contrast, the cryptic epitopes of an autoantigen may never be presented intrathymically due to limitations inherent in antigen processing (Mamula, 1993); the erro- neous induction of enzymes not normally involved in pro- cessing may, however, secure their presentation in the periphery, where T cell activation may ensue. Alterna- tively, sequestration of self-components in an immunologi- cally privileged site, such as the anterior chamber of the eye, may likewise prevent their presentation to developing thymocytes.
It is commonly believed that the blood-brain barrier is responsible for rendering the CNS just such a secluded site from which potential autoantigens are denied access to the thymic microenvironment. The consequent inability of negative selection to purge the peripheral repertoire of T cells specific for self-components such as myelin basic protein (MBP) may be readily demonstrated in susceptible strains of experimental animals: administration of either spinal cord homogenate or purified MBP in an immuno- genie fashion results in the inflammatory condition, experi- mental autoimmune encephalomyelitis (EAE), bearing many of the hallmarks of multiple sclerosis in humans.
Although the sequestration of MBP is widely believed to contribute to the lack of self-tolerance to this autoantigen, lymph draining the CNS is known to carry such proteins to the lymphoid tissue (Cserr and Knopf, 1992) where it may be presented to both the developing and peripheral Tcell repertoire. Furthermore, recent studies havedemon- strated that MBP is expressed in the fetal thymus from the earliest stages of ontogeny (Pribyl et al., 1993; Mathisen et al., 1993) consistent with its full participation in the induction of self-tolerance. Our recent characterization of an immunodominant epitopeof MBP hassuggested apos- sible solution to this paradox.
Encephalitogenic T cells from H-2” mice recognize the acetylated N-terminal peptide of MBP, Acl-9 (Zamvil et al., 1986). We and others have recently demonstrated that this epitope displays very low affinity for its restriction ele- ment, I-A”, forming an interaction so transient as to prevent the measurement of its half-life in vitro (Fairchild et al., 1993; Mason and McConnell, 1994; Mason et al., 1995). This low affinity is a function of the principal anchor resi- due, Lys4, which, being nonhydrophobic and of limited size, fits poorly into the major hydrophobic pocket of I-A” (Fairchild et al., 1994). These findings suggest that the low affinity interaction between Acl-9 and class II MHC may undermine the efficacy of its presentation in vivo, enabling autoreactive T cells to escape self-tolerance. We have tested this hypothesis directly by constructing mice transgenic for an encephalitogenic TCR and assessing the impact on the developing T cell repertoire of either Acl-9 or peptide analogs designed to exhibit higher affin- ity for I-A”.
Immunity 408
Results and Discussion
Pepttde Concentration (pgiml)
- AC,-9[4Y] - AC,-9[4V] - AC I-9[4A] - Acl-9
0 5 10 15 20
Competrtor : Probe
- Acl -9[4Y] - Acl-9[4A] - Ad-9
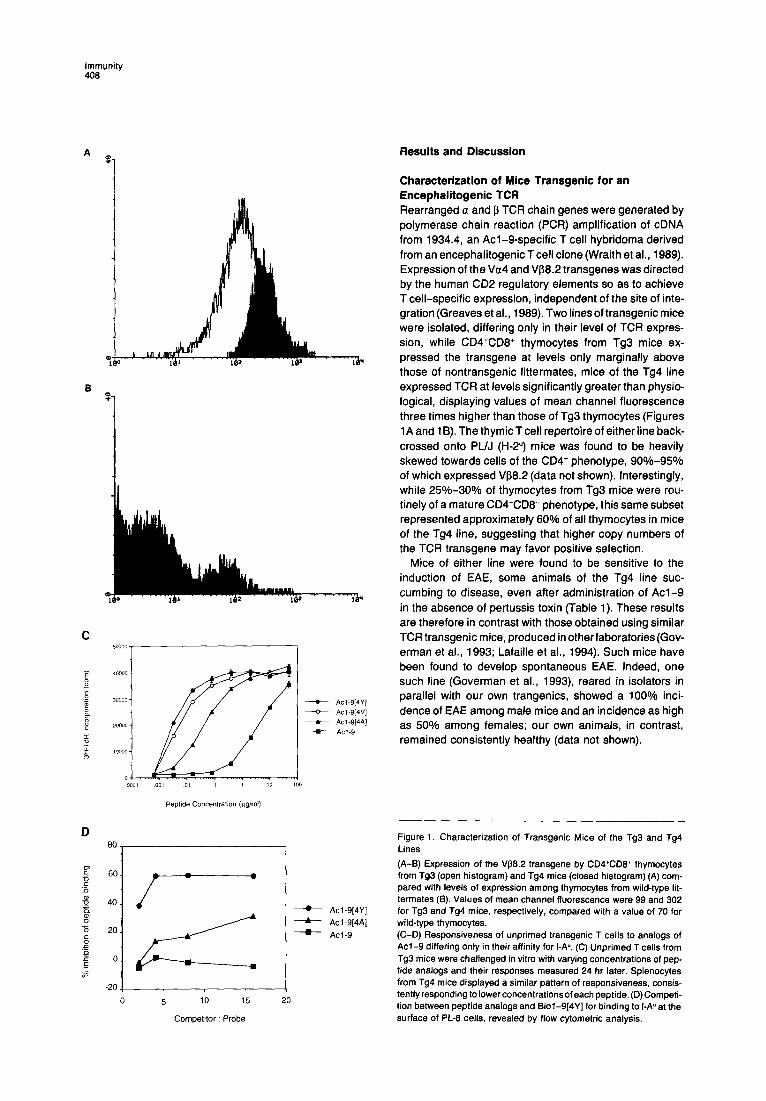
Characterization of Mice Transgenic for an Encephalitogenic TCR Rearranged a and p TCR chain genes were generated by polymerase chain reaction (PCR) amplification of cDNA from 1934.4, an Acl-S-specific T cell hybridoma derived from an encephalitogenic Tcell clone (Wraith et al., 1989). Expression of the Vu4 and Vf38.2 transgenes was directed by the human CD2 regulatory elements so as to achieve T cell-specific expression, independent of the site of inte- gration (Greaves et al., 1989). Two linesof transgenic mice were isolated, differing only in their level of TCR expres- sion, while CD4+CD8+ thymocytes from Tg3 mice ex- pressed the transgene at levels only marginally above those of nontransgenic littermates, mice of the Tg4 line expressed TCR at levels significantly greater than physio- logical, displaying values of mean channel fluorescence three times higher than those of Tg3 thymocytes (Figures 1A and 16). The thymicT cell repertoire of either line back- crossed onto PUJ (H-2”) mice was found to be heavily skewed towards cells of the CD4’ phenotype, SO%-95% of which expressed Vp8.2 (data not shown). Interestingly, while 25%-30% of thymocytes from Tg3 mice were rou- tinely of a mature CD4+CD8- phenotype, this same subset represented approximately 60% of all thymocytes in mice of the Tg4 line, suggesting that higher copy numbers of the TCR transgene may favor positive selection.
Mice of either line were found to be sensitive to the induction of EAE, some animals of the Tg4 line suc- cumbing to disease, even after administration of Acl-9 in the absence of pertussis toxin (Table 1). These results are therefore in contrast with those obtained using similar TCR transgenic mice, produced in other laboratories (Gov- erman et al., 1993; Lafaille et al., 1994). Such mice have been found to develop spontaneous EAE. Indeed, one such line (Goverman et al., 1993) reared in isolators in parallel with our own trangenics, showed a 100% inci- dence of EAE among male mice and an incidence as high as 50% among females; our own animals, in contrast, remained consistently healthy (data not shown).
Figure 1. Characterization of Transgenic Mice of the Tg3 and Tg4 Lines
(A-B) Expression of the V88.2 transgene by CD4+CD8+ thymocytes from Tg3 (open histogram) and Tg4 mice (closed histogram) (A) com- pared with levels of expression among thymocytes from wild-type lit- termates (B). Values of mean channel fluorescence were 99 and 302 for Tg3 and Tg4 mice, respectively, compared with a value of 70 for wild-type thymocytes. (C-D) Responsiveness of unprimed transgenic T cells to analogs of Acl-9 differing only in their affinity for I-A”‘. (C) Unprimed T cells from Tg3 mice were challenged in vitro with varying concentrations of pep- tide analogs and their responses measured 24 hr later. Splenocytes from Tg4 mice displayed a similar pattern of responsiveness, consis- tentlyresponding tolowerconcentrationsofeach peptide. (D)Competi- tion between peptide analogs and Biol-9[4Y] for binding to 1-A” at the surface of PL-8 cells, revealed by flow cytometric analysis.

Escape from Tolerance Induction 409
Table 1. Suscepitibility of Transgenic Mice to Induction of EAE
Non-transgenic littermates TN TM
Median Mean Median Mean Median Mean day of maximal day of maximal day of maximal
Treatment Incidence onset grade Incidence onset grade Incidence onset grade
Pertussis Toxin 015 015 015 -
Acl -9 015 - nt nt nt 2/5(1/5) 9, 27 1 .oo Acl-9 + Pertussis experiment 1 10110 11 2.2 8/8(1/E) 14 1.75 7/9(3/9) 11 2.78 Acl-9 + Pertussis experiment 2 9/9(1/9) 11 2.55 7/8(3/E) 10 2.88 8/8(616) 10 4.25
Age- and sex-matched mice were primed at the base of the tail with 200 ug of Acl-9 in CFA with or without the administration of pertussis toxin 1 and 3 days later. Control mice received pertussis toxin alone. EAE was monitored daily for a minimum of 35 days and the mean maximal grade of disease was calculated based on both paralyzed and unaffected mice. nt, not tested. ’ Numbers enclosed in parentheses denote the incidence of death.
Influence of Residue 4 on MHC Binding and T Cell Activation Both Tg3 and Tg4 mice were found to mount vigorous primary responses in vitro to Acl-9 (Figure 1C). We have shown previously that lysine at position 4 of this peptide actsasthe principal anchor residue, since analogs bearing substitutions at this position for residues of greater hydro- phobicity display demonstrably higher affinity for I-A”, while an analog bearing alanine at this position (Acl- 9[4A]) has been shown to exhibit an affinity for I-A” greater than ten times that of the wild-type peptide (Wraith et al., 1989), the tyrosine-substituted analog (Acl-9[4Y]) dis- plays a far higher affinity, rivaled only by Acl-9[4V] (Fair- child et al., 1993). Each of these analogs elicited enhanced T cell responses in an affinity-dependent manner, the mini- mal concentration of each peptide required to provoke a maximal response closely mirroring its affinity for class II determinants, measured in competition binding assays (Figure 1 D).
Previous studies of one of the two TCR contact residues of this peptide have shown that most substitutions at posi- tion 3 are nonpermissive, as few as 5 of the 20 possible analogs stimulating a cross-reactive response (Wraith et al., 1992). Interestingly, substitution of Lys with Tyr at posi- tion 4 of each of these analogs failed to modify the pattern of cross-reactivity observed, suggesting that residue 4 is solely concerned with class II binding, failing to contribute either directly or indirectly to interaction of the peptide with the encephalitogenic TCR.
Effect of Acl-9[4Y] on Immature CD4 + CD8 + T hymocytes To determine whether transgenic thymocytes are suscep- tible to the induction of tolerance centrally within the thy- mus, mice of the Tg3 line were treated intraperitoneally with 100 ug of Acl-9[4Y] in phosphate-buffered saline (PBS) and the impact on thymocyte subsets was assessed at 4 hr intervals. Three-color flow cytometry was employed to determine expression of CD4, CD8, and the Vp8.2 transgene. Figure 2 shows that the high affinity analog provoked marked deletion of CD4+CD8’ cells within 4 hr of treatment (Figure 28) the extent of clonal deletion be- coming most apparent by 16 hr when as few as 2.3% of
cells retained a CD4+CD8+ phenotype (Figure 2D). Abso- lute numbers of thymocytes were also dramatically af- fected by administration of the peptide, there being a de- crease from an average of 5.8 x lo7 cells in untreated animals, to as few as 0.7 x 10’ thymocytes, 24 hr after treatment. Injection of 10 pg of Acl-9[4Y] likewise had a profound impact, while even 1 ug of the peptide provoked detectable levels of clonal deletion of immature thymo- cytes (data not shown), results that are consistent with findings from various other transgenic models (Murphy et al., 1990; Vasquez et al., 1992; Spain and Berg, 1992; Mamalaki et al., 1992) and that confirm the peculiar sensi- tivity of CD4+CD8+ cells to negative selection pressures (Rothenberg, 1994). In accordance with these findings, the intraperitoneal administration of Acl-9[4Y] had no dis- cernable effect on numbers of mature T cells in the periph- ery (data not shown).
Effect of Acl-8[4Y] on Mature CD4+ CD8- Thymocytes Although mature CD4+CD8- thymocytes survived clonal deletion, we consistently observed the appearance of cells expressing a CD4r°CD8- phenotype (Figure 2D, arrow- head). Since this subset first appeared at 8 hr posttreat- ment, some time after the disappearance of CD4+CD8+ thymocytes (Figure 2C), such cells are most likely derived from the mature CD4+CD8- subset by the partial down- regulation of CD4. The mature status of these cells was, however, confirmed by an analysis of the early activation marker, CD69. Approximately 40% of CD4+CD8- cells from untreated mice expressed intermediate levels of CD69 (Figure 3A), consistent with their recent positive se- lection (Yamashita et al., 1993; Swat et al., 1993). Intra- peritoneal administration of 100 ug of Acl-9[4Y], how- ever, resulted in the rapid and universal up-regulation of CD69 to ten times its original level within 16 hr of treatment (Figure 38) reminiscent of the activation of mature T cells in the periphery.
Consistent with this apparent activation, flow cytometric analysis of V88.2 expression by CD4+CD8- thymocytes revealed up-regulation of the encephalitogenic TCR to twice its normal level for up to 8 hr after treatment (see Figures 2H-21) although these kinetics varied slightly be-

Immunity 410
Figure 2. Time Course of Clonal Deletion and TCR Modulation in Mice Treated with the High- est Affinity Peptide, Acl-9(4Y]
Tg3 mice were injected intraperitoneally with 100 pg of peptide and the thymus removed for phenotypic analysis either at time 0 (A and G). or at 4 hr (6 and H), 8 hr (C and I), 16 hr (D and J), 24 hr (E and K), or 4 days (F and L) posttreatment. Thymocytes were stained for three-color flow cytometry using anti-CD4-PE, anti-CDB-FITC, and biotinylated anti-V68.2 (clone F23.1). Binding of F23.1 was visualized by the addition of streptavidin-Tricolor and 10,000 events were analyzed. The CD4/CD8 profile of thymocytes was assessed (A-F) and used to set a live acquisition gate around the CD4+CD8- subset for analysis of V68.2 expres- sion (G-L). The arrowhead identifies the CD4%D8- population defined as having a mean channel fluorescence less than 70.
-CDB- - vpe - -CD8-
tween experiments. Within 12-16 hr, however, a rapid down-regulation was observed in all mice treated with the peptide. The majority revealed two distinct populations of mature thymocytes differing markedly in their surface density of both CD3 and TCR; while 40%-50% of mature thymocytes resumed pretreatment levels of TCR expres- sion, the remainder down-regulated their receptors a fur- ther 5-fold (Figure 2J). Interestingly, while cells of either phenotype were equally represented among thymocytes expressing the highest levels of CD4 (Figures 4A-46) CD41° cells were found to belong exclusively to the TCFP subset (Figure 4C), suggesting that those cells that had reduced their expression of CD4 had also maximally down- regulated theencephalitogenicTCR. Importantly, thissub- set began to re-express the TCR at levels approaching physiological within 24 hrof treatment, coincident with the expected clearance of peptide from the circulation (Figure 2K). Mature thymocytes were not uniquely susceptible to such fluxes in the expression of TCR, there being evidence
8 38.3 I B 1 96.9
- CD69 -
Figure 3. Expression of CD69 by CDQCDB- Thymocytes
CD69 expression was assessed either (A) before, or (6) 16 hr after treatment with 100 ug of Acl-9[4Y]. Thymocyies were stained for three-color flow cytometry as described in Figure 2 with the exception that biotinylated anti-CD69 (clone H1.2F3) was used in place of anti- V68.2. CD4’CD8- cells (5,000) were analyzed for surface expression of CD69 by applying a live acquisition gate around the appropriate subset.
- VP8 -
of similar phenotypic changes among peripheral T cells following administration of Acl-9[4Y] (data not shown).
In various transgenic models, the down-regulation of both CD8 and TCR ha8 been shown to maintain self- tolerance among peripheral class l-restricted T cells in response to continuous antigenic challenge (Schbnrich et al., 1991, 1992; Rocha and von Boehmer, 1991; Homer et al., 1993). Interestingly, the interaction of peripheral T cells with antigen has likewise been shown to result in a biphasic pattern of TCR expression (Ferber et al., 1994) the extent of down-regulation having been found to de- pend on the availability of antigen. This suggests that the
- VP8 -
Figure 4. Vf38.2 Expression by Mature Thymocytes Differing in their Levels of Expression of CD4
Live gates were placed around the CD41°CD8- and CD4”CD8- subsets (A) defined as exhibiting a fluorescence intensity less than 70 channels or greater than 200 channels, respectively. Events (5,000) were ac- quired and the expression of VR8.2 by CD4”’ cells (6) was compared with that of thymocytes expressing low levels of CD4 (C).

Escape from Tolerance Induction 411
TCRhi population, observed in our own experiments (see Figure 2J), may represent cells whose distribution within the thymus renders them inaccessible to peptide. Adminis- tration of Acl-9[4Y], however, results in activation of virtu- ally all CD4+CD8-thymocytesirrespective of their location, accompanied by the almost universal up-regulation of CD89 (see Figure 38) arguing strongly against such an explanation. Furthermore, the abundant expression of CD89 suggests that almost all CD4+CD8- thymocytes are antigen specific, and therefore express the clonotypic TCR, calling into question the possibility that the two popu- lationsof mature thymocytesmay bedistinguished bytheir expression of endogenous a chain in preference to the Vu4 transgene (Rocha and von Boehmer, 1991). Interest- ingly, the proportion of CD4+CD8- thymocytes down- regulating the encephalitogenicTCR toone-fifth its normal level was found to correlate approximately with the propor- tion of cells expressing CD89 prior to treatment (see Figure 3A). This may imply that the TCR’O population represents those cells that have most recently undergone positive selection and that have yet to be rendered refractory to clonal deletion (Swat et al., 1993).
Effect of Acl -9[4Y] on Reactivity of Mature CD4+ T Cells The state of reactivity of mature CD4’CD8- thymocytes was investigated in Tg3 mice following a single 100 ug dose of Acl-9[4Y]. Phenotypically mature thymocytes were purified using anti-CD8 plus complement and were cultured with mitomycin C-treated PUJ splenocytes in the presence of Acl-9. Figure 5A shows that such a regimen rendered mature thymocytes hyperresponsive to their cognate peptide compared with CD4+CD8- thymocytes from mice receiving PBS alone. A similar state of hyperres- ponsiveness was apparent among peripheral CD4+ T cells 24 hr after treatment with Acl-9[4Y], but was found to be transient, the levels of responsiveness observed falling below control levels within 72 hr of treatment (Figure 5B). These kinetics may reflect the initial up-regulation of the encephalitogenic TCR prior to its subsequent down- regulation, as seen in earlier experiments.
A profound state of nonresponsiveness was achieved among peripheral T cells by the chronic administration of Acl-9[4Y]; injection of 200 pg of peptide on 10 occasions 2-3 days apart rendered cells incapable of responding to Acl-9 in vitro. In contrast, control mice receiving the I-E”-restricted MBP epitope, 31-47, responded normally to Acl-9 (Figure 5C). Although this state of nonrespon- siveness may refect the induction of anergy, we are pres- ently unable to exclude the preferential expansion of T cell clones bearing an endogenous a chain under these experimental conditions.
Impact of the Wild-Type Peptide, Acl-9, on T Cell Repertoire Selection Having characterized the effects of Acl-9[4Y] on thymo- cytes and T cells at different stages of development, we next sought to determine whether analogs of lower affinity, and the wild-type peptide in particular, were capable of
Figure 5. Responsiveness of Mature CD4+ Thymocytes and Periph- eral T Cells to Acl-9 following In Vivo Treatment with Ad-9[4Y]
CD4’ thymocytes (A) and peripheral T cells (6 and C) were purified and cocultured with mitomycin C-treated PUJ splenocytes in the pres- ence of varying concentrations of Acl-9. Animals had been treated previously with either a single 100 pg dose of Acl-9[4Y] (A and S), or 10 successive doses of 200 ug at intervals of 2-3 days(C). Control mice were treated either with the I-E”-restricted epitope of LISP, 31- 47, or PBS alone.
provoking either clonal deletion or modulation of the TCR. Transgenic mice were therefore treated intraperitoneally with 100 ug of peptide analogs varying only in their affinity for I-A” and their impact on developing thymocytes was assessed 18 hr later when both clonal deletion and down- regulation of the TCR were expected to be most apparent (see Figures 2D and 2J).
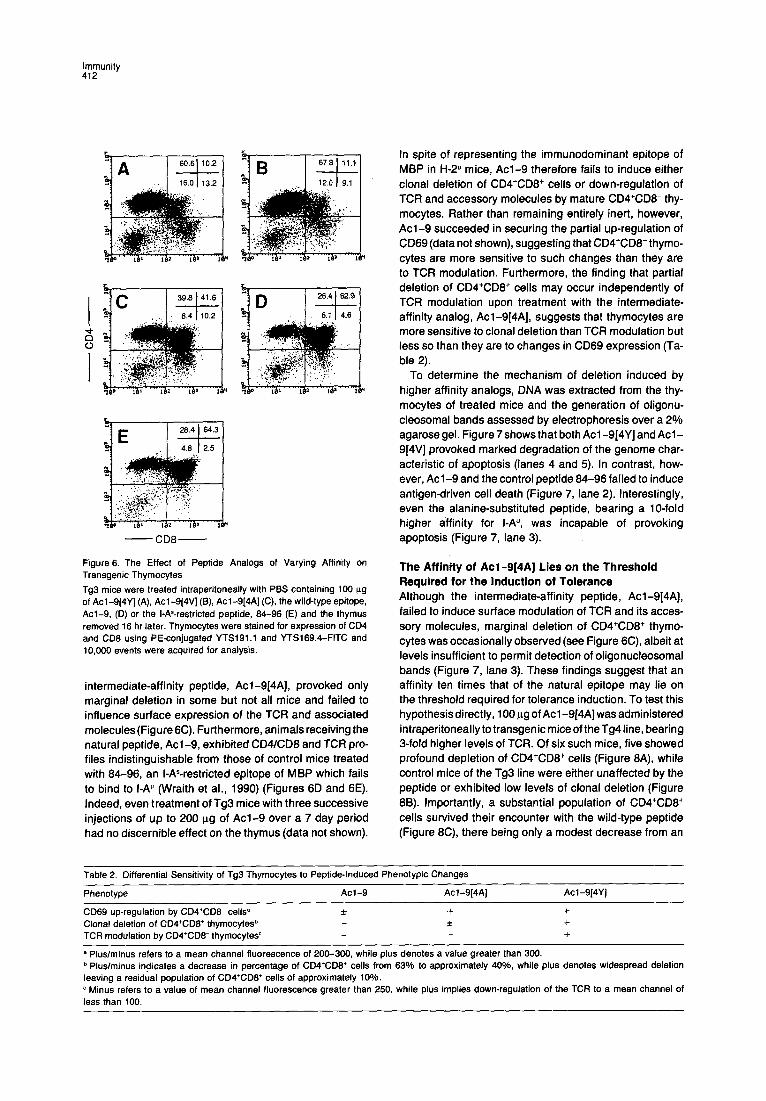
Administration of Acl-9[4Y] in PBS was again found to induce profound depletion of CD4+CD8+ thymocytes in mice of the Tg3 line, resulting in a decrease from 64.3% to 10.2% (Figure 6A). Acl-9[4V] likewise induced marked deletion of CD4+CD8+ cells (Figure 6B) in spite of dis- playing a relative affinity for I-A” lower than that of Acl- 9(4Y]. Like this higher affinity peptide, Acl-9[4V] was also responsible for the partial down-regulation of both CD4 (Figure 6B) and TCR (data not shown). In contrast, the
Immunity 412
- CD8-
Figure 6. The Effect of Peptide Analogs of Varying Affinity on Transgenic Thymocytes
Tg3 mice were treated intraperitoneally with PBS containing 100 ug of Acl-9(4Y] (A), Acl-9(4V] (B), Acl-9[4A] (C), the wild-type epitope, Acl-9, (D) or the I-As-restricted peptide, 84-96 (E) and the thymus removed 16 hr later. Thymocytes were stained for expression of CD4 and CD8 using PE-conjugated YTS191.1 and YTSl89.4-FITC and 10,000 events were acquired for analysis.
intermediate-affinity peptide, Acl-9[4A], provoked only marginal deletion in some but not all mice and failed to influence surface expression of the TCR and associated molecules(Figure6C). Furthermore, animals receiving the natural peptide, Acl-9, exhibited CD4/CD8 and TCR pro- files indistinguishable from those of control mice treated with 84-96, an I-AS-restricted epitope of MBP which fails to bind to I-AU (Wraith et al., 1990) (Figures 6D and 6E). Indeed, even treatment of Tg3 mice with three successive injections of up to 200 ug of Acl-9 over a 7 day period had no discernible effect on the thymus (data not shown).
Table 2. Differential Sensitivity of Tg3 Thymocytes to Peptide-Induced Phenotypic Changes
Phenotype Acl-9 Acl-9[4A] Acl -9[4Y]
CD69 up-regulation by CD4+CD8- cells* f + +
Clonal deletion of CD4+CD8’ thymocytesb + +
TCR modulation by CD4+CD8- thymocytes” +
a Plus/minus refers to a mean channel fluorescence of 200-300, while plus denotes a value greater than 300. b Plus/minus indicates a decrease in percentage of CD4+CD8+ cells from 63% to approximately 400% while plus denotes widespread deletion leaving a residual population of CD4+CD8’ cells of approximately 10%. c Minus refers to a value of mean channel fluorescence greater than 250, while plus implies down-regulation of the TCR to a mean channel of less than 100.
In spite of representing the immunodominant epitope of MBP in H-2” mice, Acl-9 therefore fails to induce either clonal deletion of CD4+CD8+ cells or down-regulation of TCR and accessory molecules by mature CD4+CD8- thy- mocytes. Rather than remaining entirely inert, however, Acl-9 succeeded in securing the partial up-regulation of CD69 (data not shown), suggesting that CD4+CD8- thymo- cytes are more sensitive to such changes than they are to TCR modulation. Furthermore, the finding that partial deletion of CD4+CD8+ cells may occur independently of TCR modulation upon treatment with the intermediate- affinity analog, Acl-9(4A], suggests that thymocytes are more sensitive to clonal deletion than TCR modulation but less so than they are to changes in CD69 expression (Ta- ble 2).
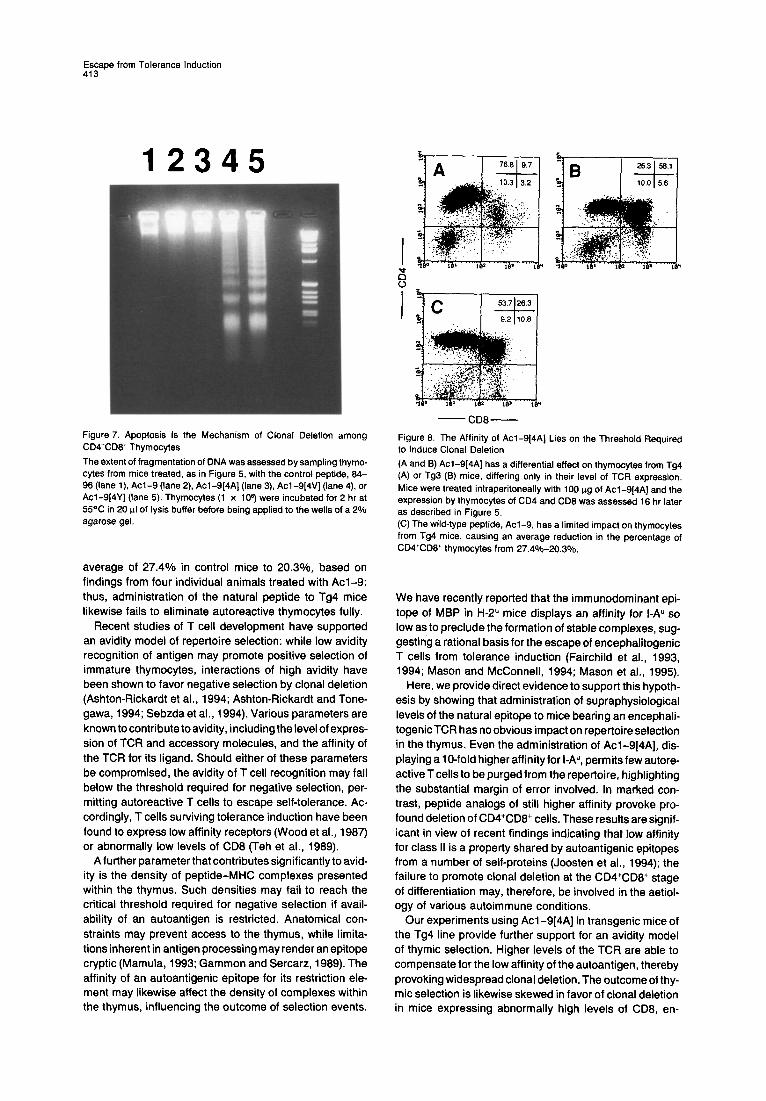
To determine the mechanism of deletion induced by higher affinity analogs, DNA was extracted from the thy- mocytes of treated mice and the generation of oligonu- cleosomal bands assessed by electrophoresis over a 2% agarose gel. Figure 7shows that both Acl-9[4Y] and Acl- 9[4V] provoked marked degradation of the genome char- acteristic of apoptosis (lanes 4 and 5). In contrast, how- ever, Acl-9 and the control peptide 84-96 failed to induce antigen-driven cell death (Figure 7, lane 2). Interestingly, even the alanine-substituted peptide, bearing a lo-fold higher affinity for I-A”, was incapable of provoking apoptosis (Figure 7, lane 3).
The Affinity of Acl-9[4A] Lies on the Threshold Required for the Induction of Tolerance Although the intermediate-affinity peptide, Acl-9[4A], failed to induce surface modulation of TCR and its acces- sory molecules, marginal deletion of CD4+CD8+ thymo- cytes was occasionally observed (see Figure 6C), albeit at levels insufficient to permit detection of oligonucleosomal bands (Figure 7, lane 3). These findings suggest that an affinity ten times that of the natural epitope may lie on the threshold required for tolerance induction. To test this hypothesis directly, 100 kg of Acl-9[4A] was administered intraperitoneally to transgenic mice of the Tg4 line, bearing 3-fold higher levels of TCR. Of six such mice, five showed profound depletion of CD4+CD8+ cells (Figure 8A), while control mice of the Tg3 line were either unaffected by the peptide or exhibited low levels of clonal deletion (Figure 8B). Importantly, a substantial population of CD4+CD8+ cells survived their encounter with the wild-type peptide (Figure 8C), there being only a modest decrease from an
Escape from Tolerance Induction 413
12345
- CD8- Figure 7. Apoptosis Is the Mechanism of Clonal Deletion among CDQCDB’ Thymocytes
The extent of fragmentation of DNA was assessed by sampling thymo- cytes from mice treated, as in Figure 5, with the control peptide, 84- 96 (lane l), Acl-9 (lane 2), Acl-9[4A] (lane 3) Acl-9[4V] (lane 4) or Acl-9[4Y] (lane 5). Thymocytes (1 x 10s) were incubated for 2 hr at 55% in 20 ul of lysis buffer before being applied to the wells of a 2% agarose gel.
average of 27.4% in control mice to 20.30/o, based on findings from four individual animals treated with Acl-9: thus, administration of the natural peptide to Tg4 mice likewise fails to eliminate autoreactive thymocytes fully.
Recent studies of T cell development have supported an avidity model of repertoire selection: while low avidity recognition of antigen may promote positive selection of immature thymocytes, interactions of high avidity have been shown to favor negative selection by clonal deletion (Ashton-Rickardt et al., 1994; Ashton-Rickardt and Tone- gawa, 1994; Sebzda et al., 1994). Various parameters are known to contribute to avidity, including the level of expres- sion of TCR and accessory molecules, and the affinity of the TCR for its ligand. Should either of these parameters be compromised, the avidity of T cell recognition may fall below the threshold required for negative selection, per- mitting autoreactive T cells to escape self-tolerance. Ac- cordingly, T cells surviving tolerance induction have been found to express low affinity receptors (Wood et al., 1987) or abnormally low levels of CD8 (Teh et al., 1989).
A further parameter that contributes significantly to avid- ity is the density of peptide-Ml-K complexes presented within the thymus. Such densities may fail to reach the critical threshold required for negative selection if avail- ability of an autoantigen is restricted. Anatomical con- straints may prevent access to the thymus, while limita- tions inherent in antigen processing may render an epitope cryptic (Mamula, 1993; Gammon and Sercarz, 1989). The affinity of an autoantigenic epitope for its restriction ele- ment may likewise affect the density of complexes within the thymus, influencing the outcome of selection events.
Figure 6. The Affinity of Acl-9[4A] Lies on the Threshold Required to Induce Clonal Deletion
(A and 9) Acl-9(4A] has a differential effect on thymocytes from Tg4 (A) or Tg3 (9) mice, differing only in their level of TCR expression. Mice were treated intraperitoneally with 100 ug of Acl-9]4A] and the expression by thymocytes of CD4 and CDB was assessed 16 hr later as described in Figure 5. (C)The wild-type peptide, Acl-9, has a limited impact on thymocytes from Tg4 mice, causing an average reduction in the percentage of CD4+CD8+ thymocytes from 27.4%20.3%.
We have recently reported that the immunodominant epi- tope of MBP in H-2” mice displays an affinity for I-A” so low as to preclude the formation of stable complexes, sug- gesting a rational basis for the escape of encephalitogenic T cells from tolerance induction (Fairchild et al., 1993, 1994; Mason and McConnell, 1994; Mason et al., 1995).
Here, we provide direct evidence to support this hypoth- esis by showing that administration of supraphysiological levels of the natural epitope to mice bearing an encephali- togenicTCR has no obvious impact on repertoire selection in the thymus. Even the administration of Acl-9[4A), dis- playing a lo-fold higher affinity for I-A”, permits few autore- active T cells to be purged from the repertoire, highlighting the substantial margin of error involved. In marked con- trast, peptide analogs of still higher affinity provoke pro- found deletion of CD4+CD8+ cells. These results are signif- icant in view of recent findings indicating that low affinity for class II is a property shared by autoantigenic epitopes from a number of self-proteins (Joosten et al., 1994); the failure to promote clonal deletion at the CD4’CDB’ stage of differentiation may, therefore, be involved in the aetiol- ogy of various autoimmune conditions.
Our experiments using Acl-9[4A] in transgenic mice of the Tg4 line provide further support for an avidity model of thymic selection. Higher levels of the TCR are able to compensate for the low affinity of the autoantigen, thereby provoking widespread clonal deletion. The outcome of thy- mic selection is likewise skewed in favor of clonal deletion in mice expressing abnormally high levels of CD8, en-

immunity 414
coded both endogenously as well as by means of a transgene (Robey et al., 1992). These findings suggest that the various parameters governing the avidity of T cell recognition determine the fate of autoreactive thymocytes, either ensuring their submission to self-tolerance or fa- voring their ultimate escape.
Experimental Procedures
Transgenic Mice TCR transgenic mice were obtained using standard approaches to transgenesis. In brief, the TCR a and 5 transgenic constructs were generated by inserting appropriate cDNAs containing the leader and coding sequences into the human CD2 minigene (Greaves et al., 1989). RNA splice sites and a poly(A) signal sequence were introduced by ligating a genomic segment of the human f3-globin gene 3’ to the TCR sequence. The final constructs were injected at equimolar con- centrations into (CBA x BlO)F2 fertilized eggs and the founder mice backcrossed onto a PUJ (H-2”) background. All animals were main- tained under specific pathogen-free conditions at Central Biomedical Services, University of Cambridge, in accordance with institutional guidelines.
Peptides All peptides were based on the acetylated N-terminal sequence of murine MBP (AC-ASQKRPSQR), with the exception of the 64-96 epi- tope, whose sequence is as follows: VHFFKNIVTPRTP. Analogs of Acl-9 displaying higher affinity for I-A” were constructed by the substi- tution of the wild-type Lys at position 4 for either Ala, Val, or Tyr. These peptides were synthesized using standard Fmoc chemistry on an LKB Biolynx 4175 synthesizer as described in detail elsewhere (Fairchild et al., 1993). The biotinylated probe, Biol-9[4Y], was likewise based on the l-9 sequence of MBP but with the addition of Gly and Tyr spacer residues at the N terminus. This extended peptide was biotinyl- ated prior to cleavage from the resin using a long chain derivative of NHS-biotin (Calbiochem), thereby inserting a 14 atom spacer between the biotin and the N-terminal amino group.
Induction of EAE Age- and sex-matched mice were primed subcutaneously at the base of the tail with 200 ug of Acl-9 in 0.1 ml of a sonicated emulsion consisting of an equal volume of complete Freund’s adjuvant (CFA) and PBS containing 4 mglml of heat-killed Mycobacterium tuberculo- sis, strain H37RA (Difco Laboratories). On days 1 and 3 postimmuniza- tion, 200 ng of pertussis toxin (Porton Products) were administered intraperitoneally in 0.5 ml of PBS. Alternatively, mice were injected with Acl-9 in CFA or pertussis toxin alone. EAE was monitored daily thereafter for 35-50 days, The severity of EAE was assessed as fol- lows: (1) flaccid tail; (2) partial hindlimb paralysis and impaired righting reflex; (3) total hindlimb paralysis; (4) fore- and hindlimb paralysis; moribund.
Proliferation Assays The responsiveness of unprimed transgenic T cells to Acl-9 and its analogs was determined by culturing unfractionated splenocytes from Tg3 mice at 3 x lo5 cells per well in IMDM medium (GIBCO) supple- mented with 1% fetal calf serum (Flow Laboratories), 2mM L-glutamine (GIBCO), 5 x 10m5 M 2-mercaptoethanol, 100 U/ml penicillin, and 100 uglml streptomycin. Peptides were titrated into cultures and individual wells were pulsed after 24 hr with 0.5 pCi of PHI-TdR (Amersham International); levels of incorporation were assessed a further 24 hr later and the results plotted as the arithmetic mean of triplicate cultures f standard deviation.
The reactivity of CD4+CD6- thymocytes to Ad-9 following in vivo treatment with Acl-9[4Y] was ascertained by preparing single cell suspensions of thymocytes and depleting residual CD4+CD6+ and CD4-CD6’ cells using YTS169.4 (antiCD8; rat y2b) and complement. The responsiveness of peripheral T cells was also investigated follow- ing their enrichment by passage over nylon wool as described (Julius et al., 1973). Thymocytes or T cells were washed and plated at 2 x lo5 cells per well of a microtiter plate in the presence of varying concentra-
tions of Acl-9 and 5 x IO5 mitomycin C-treated PUJ splenocytes. Proliferation was assessed by pulsing with PHETdR after 24 hr and measuring its incorporation a further 24 hr later.
Flow Cytometry Thymocytes were obtained by gently teasing thymi from mice treated intraperitoneally with peptide in PBS. Released thymocytes were washed in PBS containing 1% fetal calf serum and 2 mM NaN3 (wash- ing solution) and were stained for expression of CD4 and CD6 using phycoerythrin (PE)-conjugated YTS191 .I (rat y2b; Caltag) and fluores- cein isothiocyanate (FITC)-conjugated MS1 69.4 (rat y2b). Expression of a third marker was assessed by the addition of either biotinylated F23.1 (anti-V56.2; mousey2a)orbiotinylated H1.2F3(anti-CD69; Phar- Mingen), binding of either monoclonal antibody being visualized by the addition of streptavidin-Tricolor (Caltag). All incubations were of 40 min duration and were performed at 4OC on ice. Thymocytes were fixed in washing solution containing 1% formaldehyde (BDH) and were analysed on a Becton Dickinson FACSort using Lysys II software. Thymocytes were gated first according to their forward and side scat- ter characteristics and 10,000 events were acquired for phenotypic analysis,
Competition Peptide Binding Assay The I-Au-expressing B cell hybridoma, PL-6 (Wraith et al., 1991) was distributed into the wells of a 96-well round-bottomed plate and resus- pended in medium containing serial dilutions of the peptide probe, Biol-9(4Y] and a fixed concentration of competitor peptides. Cells were incubated at 37OC for 16 hr before being washed and incubated on ice for 40 min with ExtrAvidin-FITC (Sigma). Samples were ana- lyzed by flow cytometry and the percentage inhibition of peptide bind- ing calculated following subtraction of background fluorescence.
Studies of Apoptosis The extent of DNA fragmentation was assessed by sampling thymo- cytes from mice treated 16 hr earlier with peptide analogs of Acl-9 or the negative control peptide 64-96. Thymocytes (lob) were incu- bated for 2 hr at 55°C in 20 ul of lysis buffer containing 10 mM EDTA, 50 mM Tris-HCI (pH 7.5) 0.5% sodium lauryl sarkosinate, and 0.5 mg/ml of proteinase K (Stratagene). Samples were briefly warmed to 70°C, mixed with loading buffer containing 0.5% agarose, and loaded into dry wells of a 2% agarose gel.
Acknowledgments
We thank Drs. S. Anderton and M. MacEachern for critically reviewing this manuscript and Dr. M Tone for advice in preparation of transgenic constructs. Work in the authors’ laboratory was supported by grants from the Wellcome Trust and the Multiple Sclerosis Society of Great Britain and Northern Ireland. D. K. is supported by the Medical Re- search Council (United Kingdom) at the National Institute for Medical Research. G. Y. L. was supported by a Predoctoral Fellowship from the Howard Hughes Medical Institute and was the recipient of an Overseas Research Student Award from the University of Cambridge. D. C. W. was supported by a Wellcome Trust Senior Research Fellowship.
Received March 17, 1995; revised August 10, 1995.
References
Ashton-Rickardt, P.G., and Tonegawa, S. (1994). A differential avidity model of T cell selection. Immunol. Today 15, 362-366.
Ashton-Rickardt, P.G., Bandeira, A., Delaney, J.R., Van Kaer, L., Pircher, H.-P., Zinkernagel, R.M., and Tonegawa, S. (1994). Evidence for a differential avidity model of T cell selection in the thymus. Cell 76, 651-663.
Cibotti, R., Kanellopoulos, J.M., Cabaniols, J.-P., Halle-Panenko, 0.. Kosmatopoulos, K., Sercarz, E., and Kourilsky. P. (1992). Tolerance to a self-protein involves its immunodominant but does not involve its subdominant determinants. Proc. Natl. Acad. Sci. USA 89, 416-420.
Cserr, H.F., and Knopf, P.M. (1992). Cervical lymphatics, the blood- brain barrier and the immunoreactivity of the brain: a new view. Immu- not Today 73, 507-512.

Escape from Tolerance Induction 415
Dawe, K., Hutchings, P., Champion, B., Cooke, A., and Roitt, I. (1993). Autoantigens in thyroid diseases. Springer Sem. Immunopathol. 74. 285-307.
Fairchild, P.J.. Wildgoose, R., Atherton, E., Webb, S.. and Wraith, D.C. (1993). An auotantigenic T cell epitope forms unstable complexes with class II MHC: a novel route for escape from tolerance induction. Int. Immunol. 5, 1151-1158.
Fairchild, P.J., Thorpe, C.J., Travers, P.J., and Wraith, D.C. (1994). Modulation of the immune response with T-cell epitopes: the ultimate goal for specific immunotherapy of autoimmune disease. Immunology 81, 487-496.
Ferber, I., Schbnrich, G., Schenkel, J., Mellor, A.L., Hammerling, G.J.. and Arnold, 6. (1994). Levels of peripheral T cell tolerance induced by different doses of tolerogen. Science 283, 674-676.
Gammon, G., and Sercarz, E. (1989). How some T cells escape toler- ance induction. Nature 342, 183-185.
Goverman, J., Woods, A., Larson, L., Weiner, L.P., Hood, L., and Zaller, D.M. (1993). Transgenic mice that express a myelin basic pro- tein-specific T cell receptor develop spontaneous autoimmunity. Cell 72, 551-560.
Greaves, DR., Wilson, F.D.. Lang,G..andKioussis, D. (1989). Human CD2 3’.flanking sequences confer high-level, T cell-specific, position- independent gene expression in transgenic mice. Cell 56, 979-986.
Homer, R.J., Mamalaki, C., Kioussis, D., and Flavell, R.A. (1993).T-cell unresponsiveness correlates with quantitative TCR levels in a transgenic model. Int. Immunol. 5, 1495-1500.
Joosten, I., Wauben, M.H.M.. Holewijn, M.C., Reske, K., Pedersen, L.F., Roosenboom, C.F.P., Hensen, E.J., Van Eden, W.. and Buus, S. (1994). Direct binding of autoimmune disease related T cell epitopes to purified Lewis rat MHC class II molecules. Int. Immunol. 6, 751- 759.
Julius, M.H., Simpson, E.. and Herzenberg, L.A. (1973). A rapid method for the isolation of functional thymus-derived murine lympho- cytes Eur. J. Immunol. 3, 645-649.
Karachunski. P.I., Osttie. N., Conti-Tronconi, B.M., and Bellone, M. (1994). Residues within the a subunit of sequence 304-322 of muscle ACh-receptor forming autoimmune CD4+ epitopes in BALBlc mice. Immunology 82, 22-27.
Lafaille, J.J., Nagashima, K., Katsuki, M., and Tonegawa, S. (1994). High incidence of spontaneous autoimmune encephalomyelitis in im- munodeficient anti-myelin basic protein T cell receptor transgenic mice. Cell 78, 399-408.
Mamalaki, C., Norton, T., Tanaka, Y., Townsend, A.R., Chandler, P., Simpson, E., and Kioussis, D. (1992). Thymic deletion and peripheral activation of class I MHC-restricted T cells by soluble peptide in T-cell receptor transgenic mice. Proc. Natl. Acad. Sci. USA 89, 11342- 11346.
Mamula, M.J. (1993). The inability to process a self-peptide allows autoreactive T cells to escape self-tolerance. J. Exp. Med. 177, 567- 571.
Mason, K.. and McConnell, H.M. (1994). Short-lived complexes be- tween myelin basic protein peptides and IAL. Proc. Natl. Acad. Sci. USA 97, 12463-12466.
Mason, K., Denney, D.W., and McConnell, H.M. (1995). Myelin basic protein peptide complexes with the class II MHC molecules I-A” and I-A* form and dissociate rapidly at neutral pH. J. Immunol. 754, 5216- 5227.
Mathisen, P.M., Pease, S., Garvey, J., Hood, L., and Readhead, C. (1993). Identification of an embryonic isoform of myelin basic protein that is expressed widely in the mouse embryo. Proc. Natl. Acad. Sci. USA 90, 10125-l 0129.
Murphy, K.M., Heimberger, A.B., and Loh, D.Y. (1990). Induction by antigen of intrathymic apoptosisof CD4+CD8+TcR1”thymocytes in vivo. Science 250, 1720-l 723.
Pribyl, T.M., Campagnoni, C.W., Kampf, K., Kashima, T., Handley, V.W., McMahon, J., and Campagnoni, A.T. (1993). The human myelin basic protein gene is included within a 179-kitobase transcription unit:
expression in the immune and central nervous systems. Proc. Natl. Acad. Sci. USA 90, 10695-10699.
Robey, E.A.. Ramsdell, F., Kioussis, D.. Sha, W., Loh, D., Axel, R., and Fowlkes, E.J. (1992). The level of CD8 expression can determine the outcome of thymic selection. Cell 69, 1089-1096.
Rocha, B., and von Boehmer, H. (1991). Peripheral selection of the T cell repertoire. Science 251, 1225-l 228.
Rothenberg. E.V. (1994). Signaling mechanisms in thymocyte selec- tion. Curr. Opin. Immunol. 6, 257-265.
Schluesener, H.J., and Wekerle, H. (1985). Autoaggressive T lympho- cyte lines recognizing the encephalitogenic region of myelin basic protein: in vitro selection from unprimed rat T lymphocyte populations. J. Immunol. 135, 3128-3133.
Schbnrich. G.. Kalinke, U., Momburg, F., Malissen, M., Schmitt- Verhulst, A.-M., Malissen. B., Hlmmerling, G., and Arnold, B. (1991). Down-regulation of T cell receptors on self-reactive T cells as a normal mechanism for extrathymic tolerance induction. Cell 65, 293-304.
Schonrich, G., Momburg. F., Malissen, M., Schmitt-Verhulst, A.-M., Malissen, B., Hammerling, G.J., and Arnold, B. (1992). Distinct mecha- nisms of extrathymic T cell tolerance due to differential expression of self antigen. Int. Immunol. 4, 581-590.
Sebzda, E.. Wallace, V.A., Mayer, J., Yeung, R.S.M.. Mak, T.W., and Ohashi, P.S. (1994). Positive and negative thymocyte selection in- duced by different concentrations of a single peptide. Science 263, 1615-l 618.
Spain, L.M., and Berg, L.J. (1992). Developmental regulation of thymo- cyte susceptibility to deletion by’self-peptide. J. Exp. Med. 776,213- 223.
Swat, W., Dessing, M.. von Boehmer, H., and Kisielow, P. (1993). CD69 expression during selection and maturation of CD4+CD8+ thymocytes. Eur. J. Immunol. 23, 739-748.
Teh. H.-S.. Kishi, H., Scott, B.. and von Boehmer, H. (1989). Deletion of autospecific T cells in T cell receptor (TCR) transgenic mice spares cells with normal TCR levels and low levels of CD6 molecule. J. Exp. Med. 169,795-806.
Vasquez, N.J., Kaye, J.. and Hedrick, S.M. (1992). In vivo and in vitro clonal deletion of double-positive thymocytes. J. Exp. Med. 775,1307- 1316.
Wood, P.J., Socarras, S., and Steilein, J.W. (1987). Modification of the cytotoxic T cell repertoire in neonatal tolerance: evidence for the preferential survival of cells with low avidity for tolerogen. J. Immunol. 139, 3236-3244.
Wraith, D.C., Smilek, D.E., Mitchell, D.J.. Steinman, L., and McDevitt, H.O. (1969). Antigen recognition in autoimmune encephalomyelitis and the potential for peptide-mediated immunotherapy. Cell 59, 247- 255.
Wraith, DC., Smilek, D.E., Mitchell, D.J., Steinman, L., and McDevitt, H.O. (1990). Tcell recognition inexperimental auotimmuneencephalo- myelitis: prospects for immune intervention with synthetic peptides. Int. Rev. Immunol. 6, 37-47.
Wraith, DC., Smilek, D.E., and Webb, S. (1991). MHC-binding pep- tides for immunotherapy of experimental autoimmune disease. J. Au- toimmunol. 5, 103-113.
Wraith, DC., Bruun, B., and Fairchild, P.J. (1992). Cross-reactiveanti- gen recognition by an encephalitogenic T cell receptor: implications for T cell biology and autoimmunity. J. Immunol. 749, 3765-3770.
Yamashita, I., Nagata, T., Tada, T., and Nakayama, T. (1993). CD69 cell surface expression identifies developing thymocytes which audi- tion for T cell antigen receptor-mediated positive selection. Int. Immu- not 5. 1139-l 150.
Zamvil, S.S., Mitchell, D.J., Moore, A.C., Kitamura, K., Steinman, L., and Rothbard. J.E. (1986). T-cell epitope of the autoantigen myetin basic protein that induces encephalomyelitis. Nature 324, 258-260.




















