
Bioactivity of Flavonoids on Insulin-Secreting Cells
10
Bioactivity of Flavonoids on Insulin-Secreting Cells Montserrat Pinent, Anna Castell, Isabel Baiges, Gemma Montagut, Llu´ ıs Arola, and Anna Ard ´ evol ABSTRACT: Flavonoids are usually found in fruits and other plant organs and therefore widely consumed. They are antioxidants, anti-inflammatory, anticarcinogenic, and protective against coronary disease and metabolic disorders. These beneficial effects make them good candidates for the development of new functional foods with potential protective/preventive properties against several diseases. We must consider that this fact could lead to a higher intake of some of these flavonoids. Most of the studies concerning their beneficial effects showed peripheral activity of these molecules, but there is no clear information about their central effects on a key organ on metabolic control: the endocrine pancreas. The pancreas has an endocrine function of major importance to regulate nutrient metabolism, such as control of glucose homeostasis via insulin and glucagon secretion. Its importance in whole body nutrient equilibrium is highlighted by the fact that several pathologies, such as type 1 and/or 2 diabetes, are related at some point to a pancreatic cell deregulation. In this review, we compile the most relevant results concerning the effects of flavonoids on several aspects of pancreatic functionality. Studies using animals with drug-induced diabetes support the hypothesis that flavonoids can ameliorate this pathogenesis. The great diversity of flavonoid structures makes it difficult to establish common effects in the pancreas. Published data suggest that there might be direct effects of flavonoids on insulin secretion, as well as on prevention of beta-cell apoptosis, and they could even act via modulation of proliferation. The mechanisms of action involve mainly their antioxidant properties, but other pathways might also take place. Introduction It is now generally accepted that food can have health- promoting properties that go beyond their traditional nutritional value (Saris and others 1998). Recently, much attention has been paid to some food factors that may be beneficial for the preven- tion of body fat accumulation and possibly reduce the risk of dia- betes and heart diseases (Kovacs and Mela 2006). Flavonoids are a class of such bioactive compounds, usually found in fruits and other plant organs and therefore widely consumed. Their roles as antioxidants (Williams and others 2004; Scalbert and others 2005), anti-inflammatories (Li and others 2001), anticarcinogen- ics (Lu and others 2004), and protective agents against coronary disease (Bagchi and others 2003) and metabolic syndrome are widely accepted. These beneficial effects make them good can- didates for the development of new functional foods with po- tential protective/preventive properties against several diseases. We must consider that this fact could lead to a higher intake in some of these flavonoids. Most of the studies concerning the beneficial effects of flavonoids have shown peripheral activity of these molecules (Pinent and others 2006; Koo and Noh 2007; Vafeiadou and others 2007; Meeran and Katiyar 2008), but there is no clear information about their central effects on a key organ MS 20080177 Submitted 3/10/2008, Accepted 6/2/2008. Authors are with Dept. of Biochemistry and Biotechnology, Rovira i Virgili Univ., Tarragona, Spain. Direct inquiries to author Ard ´ evol (E-mail: [email protected] ). on metabolic control: the endocrine pancreas. The pancreas has an endocrine function of major importance to regulate nutrient metabolism, such as control of glucose homeostasis via insulin and glucagon secretion. Its importance in whole body nutrient equilibrium is highlighted by the fact that several pathologies, such as type 1 and/or 2 diabetes, involved in nutrient metabolism, are related at some point to a pancreatic cell deregulation. In ad- dition, the pancreas is located first in line after enteric absorption, so it could be reached by high concentrations of the bioactive- absorbed flavonoids. Therefore, it might be a target for the effects of flavonoids. Given the great importance of an optimal functionality of the pancreatic tissue and the abundance of flavonoids in the hu- man diet (including their potential use as bioactive molecules), this review discusses the most relevant results concerning the effects of flavonoids on several aspects of pancreatic functional- ity, including insulin secreting capacity, beta-cell apoptosis, and oxidative stress (also summarized in Table 1 and 2). We also revise the related effects of resveratrol because it is as well a phenolic compound and the literature describing its healthy ef- fects has increased enormously in recent years (summarized in Table 3). Flavonoid classification and metabolism Flavonoids are a class of phenolic compounds of plant origin. They are found in fruits, vegetables, and drinks such as wine, tea, and chocolate (Bhagwat and others 2003; Aron and Kennedy C 2008 Institute of Food Technologists R Vol. 7, 2008—COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY 299
Transcript of Bioactivity of Flavonoids on Insulin-Secreting Cells

Bioactivity ofFlavonoids on
Insulin-SecretingCells
Montserrat Pinent, Anna Castell, Isabel Baiges, GemmaMontagut, Lluıs Arola, and Anna Ardevol
ABSTRACT: Flavonoids are usually found in fruits and other plant organs and therefore widely consumed. They areantioxidants, anti-inflammatory, anticarcinogenic, and protective against coronary disease and metabolic disorders.These beneficial effects make them good candidates for the development of new functional foods with potentialprotective/preventive properties against several diseases. We must consider that this fact could lead to a higherintake of some of these flavonoids. Most of the studies concerning their beneficial effects showed peripheral activityof these molecules, but there is no clear information about their central effects on a key organ on metabolic control: theendocrine pancreas. The pancreas has an endocrine function of major importance to regulate nutrient metabolism,such as control of glucose homeostasis via insulin and glucagon secretion. Its importance in whole body nutrientequilibrium is highlighted by the fact that several pathologies, such as type 1 and/or 2 diabetes, are related at somepoint to a pancreatic cell deregulation. In this review, we compile the most relevant results concerning the effects offlavonoids on several aspects of pancreatic functionality. Studies using animals with drug-induced diabetes supportthe hypothesis that flavonoids can ameliorate this pathogenesis. The great diversity of flavonoid structures makesit difficult to establish common effects in the pancreas. Published data suggest that there might be direct effects offlavonoids on insulin secretion, as well as on prevention of beta-cell apoptosis, and they could even act via modulationof proliferation. The mechanisms of action involve mainly their antioxidant properties, but other pathways mightalso take place.
IntroductionIt is now generally accepted that food can have health-
promoting properties that go beyond their traditional nutritionalvalue (Saris and others 1998). Recently, much attention has beenpaid to some food factors that may be beneficial for the preven-tion of body fat accumulation and possibly reduce the risk of dia-betes and heart diseases (Kovacs and Mela 2006). Flavonoids area class of such bioactive compounds, usually found in fruits andother plant organs and therefore widely consumed. Their rolesas antioxidants (Williams and others 2004; Scalbert and others2005), anti-inflammatories (Li and others 2001), anticarcinogen-ics (Lu and others 2004), and protective agents against coronarydisease (Bagchi and others 2003) and metabolic syndrome arewidely accepted. These beneficial effects make them good can-didates for the development of new functional foods with po-tential protective/preventive properties against several diseases.We must consider that this fact could lead to a higher intakein some of these flavonoids. Most of the studies concerning thebeneficial effects of flavonoids have shown peripheral activity ofthese molecules (Pinent and others 2006; Koo and Noh 2007;Vafeiadou and others 2007; Meeran and Katiyar 2008), but thereis no clear information about their central effects on a key organ
MS 20080177 Submitted 3/10/2008, Accepted 6/2/2008. Authors are withDept. of Biochemistry and Biotechnology, Rovira i Virgili Univ., Tarragona,Spain. Direct inquiries to author Ardevol (E-mail: [email protected]).
on metabolic control: the endocrine pancreas. The pancreas hasan endocrine function of major importance to regulate nutrientmetabolism, such as control of glucose homeostasis via insulinand glucagon secretion. Its importance in whole body nutrientequilibrium is highlighted by the fact that several pathologies,such as type 1 and/or 2 diabetes, involved in nutrient metabolism,are related at some point to a pancreatic cell deregulation. In ad-dition, the pancreas is located first in line after enteric absorption,so it could be reached by high concentrations of the bioactive-absorbed flavonoids. Therefore, it might be a target for the effectsof flavonoids.
Given the great importance of an optimal functionality of thepancreatic tissue and the abundance of flavonoids in the hu-man diet (including their potential use as bioactive molecules),this review discusses the most relevant results concerning theeffects of flavonoids on several aspects of pancreatic functional-ity, including insulin secreting capacity, beta-cell apoptosis, andoxidative stress (also summarized in Table 1 and 2). We alsorevise the related effects of resveratrol because it is as well aphenolic compound and the literature describing its healthy ef-fects has increased enormously in recent years (summarized inTable 3).
Flavonoid classification and metabolismFlavonoids are a class of phenolic compounds of plant origin.
They are found in fruits, vegetables, and drinks such as wine, tea,and chocolate (Bhagwat and others 2003; Aron and Kennedy
C© 2008 Institute of Food Technologists R© Vol. 7, 2008—COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY 299

CRFSFS: Comprehensive Reviews in Food Science and Food Safety
Table 1 --- Summary of in vitro effects of flavonoids.
Flavonoid Concentration Cell type Effect Reference
Genistein 10 to 100 µmol/L Mouse islets Increase in glucose-stimulatedinsulin secretion (GSIS)
Jonas and others (1995)
100 µmol/L Rat islets Sorenson and others (1994)MIN6 Ohno and others (1993)
>100 µmol/L Rat islets Increase in GSIS Persaud and others (1999)50 µmol/L Rat islets Inhibition of GSIS0.01 to 10 µmol/L INS-1 Increase in GSIS Liu and others (2006)
MIN6Mouse islets
25 µmol/L Human islets Reduction of NaF-inducedapoptosis
Elliott and others (2002)
100 µmol/L Human and rat islets Increase in apoptosisIncrease in NaF-induced
apoptosis100 µmol/L RIN-m5F Increase in apoptosis
Reduction in NaF-inducedapoptosis
100 µmol/L Rat islets Inhibition proliferation Sorenson and others (1994)Anthocyanins and
anthocyanidins50 µmol/L INS-1 Increase in basal insulin
secretionJayaprakasam and others
(2005)(cyanidin-3-galactoside
and pelargonidin)Increase in GSIS
Puerarine 50100 µmol/L Rat islets Inhibition of the H2O2-inducedapoptosis
Xiong and others (2006)
Increase in the activities ofantioxidant enzymes
EGCG 0 to 436.3 µmol/L RIN-m5F Prevention of cytokine-induced cell death
Han (2003)
Mouse islets Song and others (2003)10 to 100 µmol/L Rat islets Reduction of the apoptosis
induced by hypoxia-reperfusion
Hara and others (2006)
Decrease in the markers ofoxidative damage
Sylimarin 100 µmol/L RIN-m5F Prevention of cytokine-induced cell death
Matsuda and others (2005)
Down-regulation of iNOSexpression
2008). Their basic structure consists of 3 phenolic rings: the ben-zene ring (A) is condensed with a 6-member ring (C), which inthe 2-position carries a phenyl benzene ring (B) as a substituent(Aherne and O’Brien 2002). Depending on the structure and ox-idation level of the ring C, flavonoids are further divided intoseveral subclasses. The main groups of flavonoids are anthocyani-dins, flavonols, flavones, flavanones, isoflavones, and flavanols(Table 4). In addition, the basic flavonoid structures canbe modified and have several kinds of substitutions (includ-ing glycosylation, hydrogenation, hydroxylation, malonylation,methylation, glucuronidation, and sulfatation) following differentpatterns, which confers them different physical properties. Fur-thermore, the degree of polymerization adds more variabilityto the flavonoids. So, the term flavonoid includes thousandsof structures with different chemical, physical, and biologicalproperties.
Some researchers support the view that the form in whichflavonoids are found in plants is not their bioactive form (Manachand others 2005; Stevenson and Hurst 2007). It has been sug-gested that most aglycone flavonoids are absorbed by passivediffusion (Crespy and others 2003). One hypothesis points outthat flavonoid glycosides must be modified (deglycosilated) by
intestinal enzymes or colonic microflora in the intestine priorto absorption (Williamson and others 2000; Crespy and others2003; Manach and others 2005). Next, absorbed flavonoids canbe further metabolized in the enterocyte and in the liver (Spencerand others 2004; Walle 2004; Silberberg and others 2006; Walle2007; Zhang and others 2007). All this high diversity of structuresmakes its detailed identification and plasma measurement verymuch complicated, but despite all these difficulties, several formsof modified flavonoids have been found in plasma (Manach andothers 2005; Lotito and Frei 2006).
Furthermore, some flavonoids have also been found within tis-sues: stomach, small intestine, colon, liver, pancreas, spleen, kid-ney, muscle, heart, endothelial cells, lung, brain, thyroid, bone,skin, bladder, prostate, testes, vagina, uterus, ovary, mammarygland, fat, adrenal gland, esophagus, eyes, lymph nodes, and pi-tuitary gland (Ueno and others 1983; Suganuma and others 1998;Chang and others 2000; Coldham and Sauer 2000; Kim and oth-ers 2000; Youdim and others 2000; Datla and others 2001; Abd ElMohsen and others 2002; Meng and others 2002; Mullen and oth-ers 2002; Swezey and others 2003; Vitrac and others 2003; Grafand others 2005). Concerning flavonoids in the pancreas, epigal-locatechin gallate (EGCG) was detected in this tissue as soon as
300 COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY—Vol. 7, 2008
Flavonoids on beta cells . . .
1 h after oral administration and reached about 3 times higherlevels after 24 h. A 2nd oral administration 6 h later resulted ina 4-fold increase of EGCG levels in the pancreas (Suganuma andothers 1998).
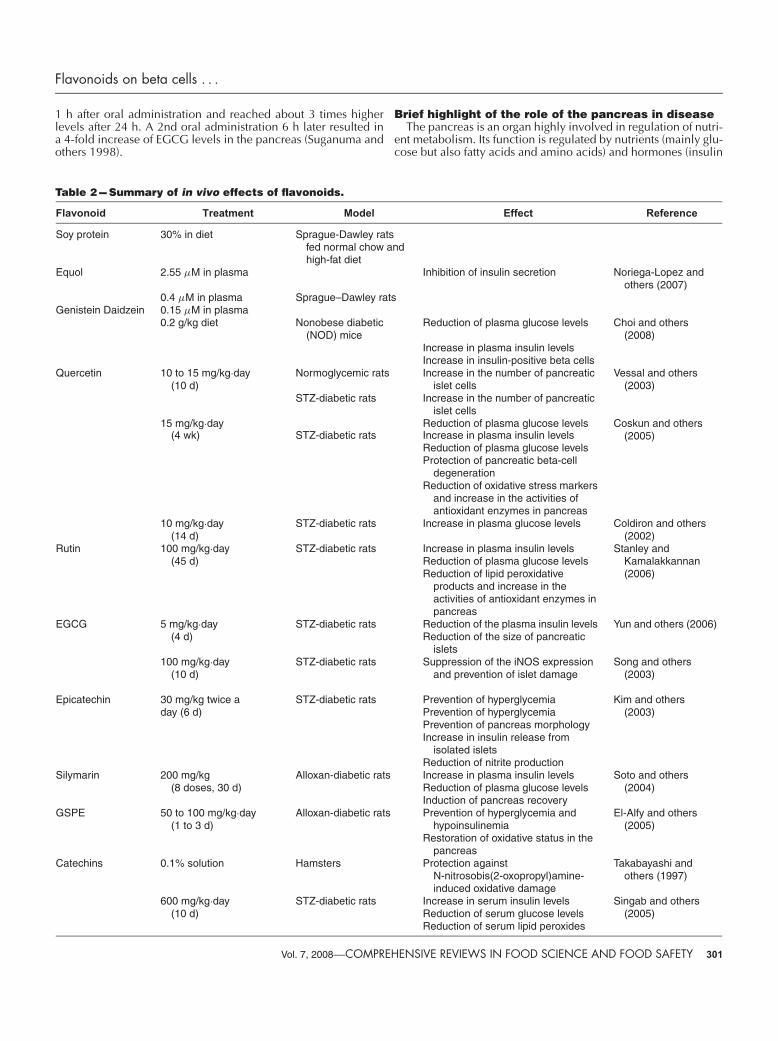
Table 2 --- Summary of in vivo effects of flavonoids.
Flavonoid Treatment Model Effect Reference
Soy protein 30% in diet Sprague-Dawley ratsfed normal chow andhigh-fat diet
Equol 2.55 µM in plasma Inhibition of insulin secretion Noriega-Lopez andothers (2007)
0.4 µM in plasma Sprague–Dawley ratsGenistein Daidzein 0.15 µM in plasma
0.2 g/kg diet Nonobese diabetic(NOD) mice
Reduction of plasma glucose levels Choi and others(2008)
Increase in plasma insulin levelsIncrease in insulin-positive beta cells
Quercetin 10 to 15 mg/kg·day(10 d)
Normoglycemic rats Increase in the number of pancreaticislet cells
Vessal and others(2003)
STZ-diabetic rats Increase in the number of pancreaticislet cells
15 mg/kg·day Reduction of plasma glucose levels Coskun and others(2005)(4 wk) STZ-diabetic rats Increase in plasma insulin levels
Reduction of plasma glucose levelsProtection of pancreatic beta-cell
degenerationReduction of oxidative stress markers
and increase in the activities ofantioxidant enzymes in pancreas
10 mg/kg·day STZ-diabetic rats Increase in plasma glucose levels Coldiron and others(2002)(14 d)
Rutin 100 mg/kg·day STZ-diabetic rats Increase in plasma insulin levels Stanley andKamalakkannan(2006)
(45 d) Reduction of plasma glucose levelsReduction of lipid peroxidative
products and increase in theactivities of antioxidant enzymes inpancreas
EGCG 5 mg/kg·day STZ-diabetic rats Reduction of the plasma insulin levels Yun and others (2006)(4 d) Reduction of the size of pancreatic
islets100 mg/kg·day
(10 d)STZ-diabetic rats Suppression of the iNOS expression
and prevention of islet damageSong and others
(2003)
Epicatechin 30 mg/kg twice aday (6 d)
STZ-diabetic rats Prevention of hyperglycemiaPrevention of hyperglycemia
Kim and others(2003)
Prevention of pancreas morphologyIncrease in insulin release from
isolated isletsReduction of nitrite production
Silymarin 200 mg/kg Alloxan-diabetic rats Increase in plasma insulin levels Soto and others(2004)(8 doses, 30 d) Reduction of plasma glucose levels
Induction of pancreas recoveryGSPE 50 to 100 mg/kg·day
(1 to 3 d)Alloxan-diabetic rats Prevention of hyperglycemia and
hypoinsulinemiaEl-Alfy and others
(2005)Restoration of oxidative status in the
pancreasCatechins 0.1% solution Hamsters Protection against
N-nitrosobis(2-oxopropyl)amine-induced oxidative damage
Takabayashi andothers (1997)
600 mg/kg·day STZ-diabetic rats Increase in serum insulin levels Singab and others(2005)(10 d) Reduction of serum glucose levels
Reduction of serum lipid peroxides
Brief highlight of the role of the pancreas in diseaseThe pancreas is an organ highly involved in regulation of nutri-
ent metabolism. Its function is regulated by nutrients (mainly glu-cose but also fatty acids and amino acids) and hormones (insulin
Vol. 7, 2008—COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY 301

CRFSFS: Comprehensive Reviews in Food Science and Food Safety
Table 3 --- Summary of resveratrol effects.
In vitro studies
Concentration Cell type Effect Reference
219.1 µmol/L INS-1 No effect on insulin secretion Zhang and others (2004)3, 10, 30, 100 µmol/L RIN-m5F10, 30, 100 µmol/L Hit-T15 Increase in insulin secretion Chen and others (2007)10, 30 µmol/L MIN61 to 100 µmol/L Rat islets Inhibition of GSIS (dose-dependent) Szkudelski (2006, 2007, 2008)
In vivo studies
Treatment Model Effect Reference
3 mg/kg Normal wistar rats Increase in plasma insulin levels Chen and others (2007)(1 dose) Reduction of plasma glucose levels
0.04% in diet HFD- fed mice Reduction of hyperinsulinemia Baur and others (2006)(18 mo)
400 mg/kg·day HFD- fed mice Reduction of fasting plasma insulin Lagouge and others (2006)50 mg/kg Normoglycemic rats Reduction of blood insulin Szkudelski (2008)
(1 dose)0.5 mg/ kg, 3 times a day STZ-nicotinamide-diabetic mice Reduction of plasma insulin Su and others (2006)
(14 d)3 and 10 mg/ kg, (1 dose) Normal mice Increase in plasma insulin levels, Chi and other (2007)
reduction of plasma glucose levelsSTZ-nicotinamide-diabetic mice Increase in plasma insulin levels,
reduction of plasma glucose levelsSTZ-diabetic mice No effect in plasma insulin levels,
reduction of plasma glucose levels
itself, as well as others such as incretins). In turn, pancreas hasan endocrine function of major importance to regulate nutrientmetabolism, that is, the control of glucose homeostasis via insulinand glucagon secretion (by beta and alpha cells, respectively). Inaddition, pancreas has an exocrine function, since it secretes di-gestive enzymes into the gut. Its importance in whole body nutri-ent equilibrium is highlighted by the fact that several pathologiesinvolved in nutrient metabolism are related at some point to apancreatic cell deregulation (Muoio and Newgard 2006). It hasbeen shown that during obesity there are changes in pancreaticbeta cells to adapt to the increased energy intake and thereforeglucose availability (Muoio and Newgard 2006). Obesity mightalso cause insulin resistance, which stimulates further insulin se-cretion by the pancreas. And this can lead to type 2 diabetes,where loss of effective beta-cell function is the key determinantof deteriorating glycemic control (Bagust and Beale 2003). Type1 diabetes results also from loss of beta cells due to an immuneassault (Ichinose and others 2007).
In fact, diabetes poses a major challenge to the present pop-ulation globally. It is a major threat to global public health thatis rapidly reaching epidemic scale, with the biggest impact onadults at working age in developing countries. According to theWorld Health Organization, at least 171 million people world-wide have diabetes. This figure is likely to be more than doubleby 2030 and reach 366 million (Wild and others 2004). It is postu-lated that most of this increase will occur as a result of a 150% risein developing countries (Wild and others 2004). This estimationis likely to be even higher taking into consideration populationgrowth, aging, urbanization, declining levels of physical activity,which—combined with unbalanced diet—increases the risk ofobesity linked to the onset of type 2 diabetes. These facts implythat the pancreas is an important tissue for nutrient regulationand therefore a clear target for the action of putative bioactivecompounds (Feick and others 2007).
Effects of Flavonoids on Insulin-Secreting Cells
Effects of flavonoids on insulin secretionAs already mentioned, a proper pancreatic function implies
a well-regulated insulin secretion. Several studies describe theeffects of flavonoids in insulin secretion.
In vitro studies demonstrate that some flavonoids modify theinsulin-secreting capacity of the cell. Genistein is the most studiedisoflavone. Early studies showed that this compound increasesglucose-stimulated insulin secretion in the beta-pancreatic cellline MIN6 (mouse-derived) (Ohno and others 1993) as well asin cultured islets from mice (Jonas and others 1995) and rats(Sorenson and others 1994) at concentrations up to 100 µmol/L.Higher concentration of genistein, on the other hand, inhibitedinsulin secretion in rat islets (Persaud and others 1999). Morerecently, acute genistein treatment at physiologically achievableconcentrations was shown to potentiate glucose-stimulated in-sulin secretion both in cell lines and isolated mouse islets (Liuand others 2006).
Several molecules from the class of anthocyanins and antho-cyanidins are also effective insulin secretagogues when tested inpancreatic cell lines, with different efficiencies depending on thestructure. It seems that the number of hydroxyl groups in ring B ofanthocyanins played an important role in their ability to secreteinsulin. Among the anthocyanidins tested, pelargonidin was themost active one (Jayaprakasam and others 2005).
Resveratrol, a plant phenolic compound with potential ben-eficial effects including prevention of diabetes and attenuationof some diabetic complications, has been shown to interferewith pancreatic beta-cell insulin secretion, although results arecontroversial. In 1 study where components of grape skin wereextracted, the fraction corresponding to the compound resver-atrol did not show any activity on insulin secretion by INS-1cells (Zhang and others 2004). The concentrations used for this
302 COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY—Vol. 7, 2008

Flavonoids on beta cells . . .
assay were high (50 µg/mL, which is about 219 µmol/L)(Zhangand others 2004). In fact, the concentration of resveratrol usedfor the experiments seems to be critical, since other experimentsshowed no effect of 100 µmol/L resveratrol on insulin secretionin MIN6 cells while lower concentrations (10 and 30 µmol/L) ef-fectively activated it (Chen and others 2007). Instead, in other celllines (Hamster-derived Hit-T15 and rat-derived RIN-m5F), resver-atrol was insulin-inducer at all 3 concentrations tested (10, 30,
Table 4 --- Classification, representative structure, and food source of main flavonoids.
100 µmol/L) (Chen and others 2007). In pancreatic islets isolatedfrom normal rats, incubation with resveratrol (1 to 100 µmol/L) in-hibited glucose-stimulated insulin secretion in a dose-dependentway (Szkudelski 2006, 2008), whereas the basal hormone releasewas not modified (Szkudelski 2008).
Concerning in vivo effects, long-term studies of the effects ofsoy protein containing genistein and daidzein were performedin models fed a normal and a high-fat diet (Noriega-Lopez and
Vol. 7, 2008—COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY 303

CRFSFS: Comprehensive Reviews in Food Science and Food Safety
others 2007). The study showed that chronic consumption ofsaturated fat increased insulin secretion associated with an in-crease in pancreatic islets, and soy protein ameliorated this sit-uation. Thus, soy protein (due to its amino acid pattern as wellas its isoflavones) reduces blood insulin. This was analyzed byhyperglycemic clamps, in which it was observed that rats fed soyprotein secreted less insulin to maintain glucose at normal lev-els. Interestingly, this effect was observed also in normal-fed rats.Furthermore, a hyperglycemic clamp of rats infused with phy-toestrogens (genistein, daidzein, and equol) showed that thesecompounds rapidly inhibit the release of insulin, suggesting ashort-term mechanism regulating this process involving down-regulation of peroxisome proliferator-activated receptor gamma(PPARγ ) and glucose transporter type 2 (Glut2) mRNA expression(Noriega-Lopez and others 2007). On the other hand, genisteinand daidzein have been shown to elevate plasma insulin levelsin nonobese diabetic (NOD) mice, an animal model that sponta-neously develops autoimmune diabetes (Choi and others 2008). A9-wk treatment with genistein or daidzein (0.2 g/kg diet, animalsfed freely) suppressed the blood glucose rising in NOD mice by el-evating plasma insulin levels. Such effects were accompanied byan increase in insulin-positive beta cells although it remained un-resolved whether there was more insulin secretion from remain-ing beta cells or increased beta-cell mass in isoflavone-treatedmice.
Regarding resveratrol, different results are found. A 3 mg/kgbody weight dose (intraperitoneally administrated) increased fast-ing plasma insulin in normal rats, in association with a glucosereduction (Chen and others 2007). Such effect was observed at20 min and it was sustained for 60 min. In contrast, blood in-sulin was reduced in normal rats by a 50 mg/kg body weightdose of resveratrol (Szkudelski 2008). Reduction in plasma insulinwas also found in hyperinsulinemic mice models. The hyperin-sulinemia induced by a high fat diet was reduced by resveratroltreatment (0.04% in diet, 18 mo) (Baur and others 2006). Simi-larly, another experiment with HFD-fed mice showed reductionin fasting plasma insulin by 400 mg/kg·day resveratrol (Lagougeand others 2006).
Many studies on diabetes make use of animal models in whichpancreatic function is impaired due to drug (streptozotocin, al-loxan) treatment and there are a few studies of the action offlavonoids using these models. Since in these animal models in-sulin secretion is impaired due to enhancement of beta-cell death,flavonoid effects will be discussed in another section.
Effects of flavonoids on beta-cell apoptosisComplex diseases such as diabetes lead to loss of functional
beta-cell mass. Actually, beta-cell apoptosis may be a commonfeature of type 1 and type 2 diabetes, although the mechanismactivating it in each case may be different (reviewed by Cnopand others (2005)). Thus, the potential antiapoptotic effects offlavonoids, already demonstrated in other cell types (Bagchi andothers 2002; Du and others 2007), must be carefully consideredwhen analyzing their potential beneficial effects against thesepathologies.
The flavonoid genistein modulates beta-cell apoptosis. Bothin pancreatic islets and in a beta-cell line, low (25 µmol/L)genistein doses reduced NaF-induced apoptosis, whereas high(100 µmol/L) genistein doses caused apoptosis (Elliott and others2002). The isoflavone puerarin was shown to significantly sup-press the apoptosis of H2O2-induced rat islet cells. This effect wasachieved by pretreating the cells with either 50 or 100 µmol/Lpuerarin, and was accompanied by an increase in the activitiesof antioxidant enzymes (Xiong and others 2006).
In type 1 diabetes; beta cells are destroyed by autoimmune re-sponses mediated by cytokines. The monomer epigallocatechingallate (EGCG) can prevent such cytokine-induced cell deathboth in insulinoma cell line (Han 2003) and in isolated mice isletcells (Song and others 2003). In both studies, the flavonoid effectswere dose-dependent (maximal tested dose: 200 µg/mL), andachieved by cotreatment of the EGCG together with the cytokinesfor 24 h. Such effects could be mediated by down-regulation ofnitric oxide synthase inducible (iNOS) expression through inhibi-tion of nuclear factor-kappa B (NF-κB) activation (Han 2003). Fur-thermore, apoptosis can also be induced by hypoxia-reperfusion,a condition that, for example, takes place after transplantation ofpancreatic islets. EGCG in rat pancreatic islets, reduced as wellthe apoptosis induced by hypoxia-reperfusion (Hara and oth-ers 2006). At the same time, EGCG decreased the markers ofoxidative damage that had been induced by hypoxia (Hara andothers 2006).
The cytokine-induced cell death is also prevented by theflavonoid silymarin (Matsuda and others 2005). In this case, theexperiment was performed in an insulinoma cell line (RINm5F).The effect was mediated by suppression of Jun N-terminal kinase(JNK) and signal transducer and activator of transcription (STAT),although it is not clear how this effect is achieved, and whetherit is only due to antioxidant activity (Matsuda and others 2005).
Thus, it seems that some flavonoids can exert antiapoptotic ef-fects in pancreatic beta cells. More experiments are, however,required to elucidate whether such antiapoptotic effects can alsotake place in vivo and if this could ameliorate or prevent pancre-atic dysfunctions linked to apoptosis of beta-cell mass.
Effects of flavonoids on beta-cell proliferationAnother important aspect of pancreatic integrity is regeneration
of the endocrine pancreas. Beta-cell mass expansion in adult an-imals is slow. However, under circumstances of insulin demandadult animals can adapt beta-cell mass. The inability of the en-docrine pancreas to undergo this adaptation can result in hyper-glycemia and diabetes (Dhawan and others 2007). Thus, actingon mechanisms that regulate plasticity of beta cells can be an im-portant step in developing effective strategies to treat the above-mentioned diseases. Furthermore, after beta-cell loss due to eithertype 1 or type 2 diabetes, beta-cell regeneration can occur (Tour-rel and others 2002); (Meier and others 2005). Some hypothe-ses point out to neogenesis from pancreatic ducts as a mech-anism for beta-cell regeneration, although others suggest thatthe regeneration of beta cells is mediated by replication of betacells rather than neogenesis (reviewed by Levine and Itkin-Ansari(2007).
Several flavonoids modulate cell proliferation. Procyanidins(Pinent and others 2005; Lizarraga and others 2007), monomerssuch as quercetin (Notoya and others 2004) and gallic acid (Veluriand others 2006), and other flavonoids such as genistein (Harmonand Harp 2001) modulate cell proliferation in many cell types,including among others fibroblasts (Harmon and Harp 2001), os-teoblasts (Notoya and others 2004), and several carcinoma celllines (Veluri and others 2006; Lizarraga and others 2007). Studiesperformed in pancreatic beta cells are scarce, but the fact thatthere are key cell cycle regulators common in all these cell typessuggests that flavonoids might have effects on beta-cell prolifera-tion. In fact, genistein is a potent inhibitor of proliferation of cul-tured islet cell, when added at high concentrations (100 µmol/L)for 4 d (Sorenson and others 1994).
Oxidative stress diminishes beta-cell proliferation, and the pro-tective effect of some phenolics on proliferation after oxidativestress in pancreatic beta cells has been tested. Some of such com-pounds, chrysin, quercetin, catechin, and caffeic acid, protected
304 COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY—Vol. 7, 2008

Flavonoids on beta cells . . .
the cells significantly, whereas gallic acid and hesperitin did not(Lapidot and others 2002).
Concerning in vivo effects on beta-cell regeneration, themonomer quercetin (10 to 15 mg/kg per day for 10 d) increasedthe number of pancreatic islet cells in streptozotocin (STZ)-diabetic rats. Curiously, an increased number of beta cells wasalso found in normal (nondiabetic) rats (Vessal and others 2003).In any case, the rest of the studies concerning the effects offlavonoids as a potential preventive against pancreas destructionin drug-treated animals (discussed subsequently) did not showeffects on nontreated animals.
Thus, although some evidence points out the potential roleof flavonoids as modulators of beta-cell proliferation, their ex-act effects and the implications that this could have against thedevelopment of diabetes are still quite unexplored.
Mechanisms of Action
Flavonoid antioxidant effects on insulin secretionSome of the most used models for diabetes are the animals
with drug-induced diabetes, either by STZ treatment or by alloxantreatment. Both diabetogens lead to beta-cell loss, and thus low-ered insulin secretion, mimicking human type 1 diabetes. Themechanism of action of such drugs is, however, not well under-stood. In alloxan diabetes, reactive oxygen mediates the selectivenecrosis of beta cells (Lenzen 2008). In STZ diabetes, DNA alky-lation might mediate the toxic action of this drug, although pro-duction of reactive oxygen species may also be involved (Lenzen2008). Since flavonoids can act as potent antioxidant molecules,some studies have focused on their effects in drug-induced dia-betes.
Quercetin is a well-studied flavonoid. Its antioxidant beneficialeffects are extended to protection of pancreatic beta-cell destruc-tion by STZ (Coskun and others 2005). In this study, quercetin(15 mg/kg/d) was injected intraperitoneally 3 d prior to induc-tion of diabetes with STZ. The effects of STZ treatment on low-ering plasma insulin levels as well as in increasing the glucoselevels were partly prevented by quercetin treatment. Immunohis-tochemical examination showed that quercetin ameliorates thedegeneration of beta cells and decreases oxidative stress markers.However, in a previous study, the same flavonoid quercetin didnot show modification of oxidative stress status in STZ-diabeticrats (Coldiron and others 2002). Rutin, a glycosidic form ofquercetin, ameliorated the decrease in fasting insulin plasma lev-els induced by STZ-treatment in rats, while it had no effect in nor-mal rats when administered at 100 mg/kg for 45 d. The same treat-ment also showed amelioration of STZ-induced oxidative stressmarkers, by lowering pancreatic concentrations of thiobarbituricacid-reactive substances (TBARS) and lipid hydroperoxides andincreasing the activities of antioxidant enzymes (Stanley andKamalakkannan 2006).
The monomer EGCG in islet cells obtained after multiple lowdoses of STZ-treated mice suppressed the expression of iNOS, theenzyme involved in cytokine-induced pancreatic beta-cell dam-age. This antioxidant effect was accompanied by a reduction ofthe STZ-induced hyperglycemia (Song and others 2003). On theother hand, EGCG showed also prooxidant effects in an STZ-diabetic model. When administered for 4 d at a concentration(5 mg/kg·day), which leads to nanomolar plasma levels, EGCGdid not ameliorate but further reduced the lowering insulinsecretion provoked by the STZ treatment. In normal rats,EGCG showed no effect on insulin secretion (Yun and others2006).
Green tea (–)-epicatechin administered in rats at 30 mg/kgtwice a day for 6 d protected them from the hyperglycemia in-
duced by STZ, prevented pancreas morphology, and increasedinsulin release of isolated pancreas of such STZ-treated rats, con-comitant with a reduction in nitrite production (Kim and others2003).
The flavonoid silymarin was shown to induce pancreas recov-ery against alloxan-induced diabetes, both when administeredduring alloxan treatment or after it. The reduction in serum in-sulin levels induced by alloxan was inhibited by simultaneousadministration with silymarin and pancreas tissue morphologywas normal (Soto and others 2004). Considering previous litera-ture, the researchers suggest that this effect could be due to theantioxidant properties of silymarin, although no markers of ox-idative stress were analyzed in this study (Soto and others 2004).
In addition to pure monomers, plant extracts that contain amixture of flavonoids have also been studied. Grape seed pro-cyanidins are effective in preventing the hyperglycemia and hy-poinsulinemia in alloxan-induced diabetic rats. The concomitantpancreatic damage induced by alloxan in terms of lipid peroxi-dation was also prevented by a grape seed procyanidin extract(GSPE), suggesting therefore a protective effect of GSPE againstalloxan action via restoration of oxidative status in the pancreas(El-Alfy and others 2005).
Tea catechins might also be protective against induced-oxidative damage, as suggested by experiments with hamsterswhere catechins reduced the amount of oxidative markers (per-oxides and 8-hydroxydeoxyguanosine) (Takabayashi and others1997).
The flavonoid-rich fraction of 70% alcoholic extract of Egyptianmoru alba root has also been claimed to be protective againstSTZ-induced damage. At a high concentration, (600 mg/kg/d, for10 d) this fraction reduced serum glucose levels and increasedserum insulin levels in STZ-diabetic rats. At the same time, itimproved cellular oxidative damage, decreasing lipid peroxides(Singab and others 2005).
Finally, resveratrol is another phenolic compound with antiox-idant activity (Jia and others 2008), whose antioxidant capacitycould also contribute to its effects on modifying insulin secretion.Reduction of glucose plasma levels due to resveratrol treatmenthas been shown in different diabetic models, although the effectson insulin secretion are diverse. STZ-nicotinamide diabetic miceis an animal model with moderate hyperglycemia, reduced pan-creatic insulin stores but significant glucose-stimulated insulin se-cretion (Masiello and others 1998). In such model, resveratrol wasshown to reduce plasma insulin levels, likely due to the reduc-tion of hyperglycemia (Su and others 2006). Thus the researcherspointed out that the hypoglycemic effects of resveratrol were notdue to modulation of insulin secretion by the pancreas but dueto its effects on other tissues, and, although the study did not in-vestigate whether such effects were due to resveratrol antioxidantproperties, they suggested the possibility that resveratrol changesthe oxidative stress of the diabetic tissues. On the contrary, otherresearchers found out opposite effects on insulin plasma levelsin the STZ-nicotinamide diabetic model (Chi and others 2007).In that study, reduction of glycemia was at least in part due to in-creased insulin secretion. The same study was also carried out inSTZ-diabetic mice (where the pancreas insulin-secreting capacityis impaired) but in that case resveratrol treatment did not modifyinsulin levels while it reduced hyperglycemia. Thus, resveratrolshowed no antioxidant effects on pancreas but enhancement ofglucose uptake in periferial tissues. Resveratrol showed as well noeffects on insulin secretion in STZ-diabetic rats (Chen and others2007), in opposite to the above-mentioned effects in normal rats.
Therefore, some flavonoids might prevent beta-cell destructionin models of drug-induced diabetes (mimicking human type 1diabetes), and this could be attributed, at least in part, to theirantioxidant properties. The concrete effects, however, depend
Vol. 7, 2008—COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY 305

CRFSFS: Comprehensive Reviews in Food Science and Food Safety
on the flavonoid and the concentration and/or experimental con-ditions. Since oxidative stress is also suggested to take part intype 2 diabetes, the antioxidant properties of flavonoids could bea general mechanism of preventing this pathology.
Other mechanisms of action of flavonoids on insulinsecretion
Although flavonoids have been widely studied because of theirantioxidant properties, they can also act through other mech-anisms, by interacting with protein function, modulating intra-cellular cascades, and modulating gene expression (Del Bas andothers 2005; Pinent and others 2006; Stevenson and Hurst 2007).Concerning the mechanisms that may lead to beta-cell dysfunc-tion, these are not well understood. The insulin and insulin-likegrowth factor (IGF) signaling pathways are critical for propermaintenance and functioning of beta cells (Dhawan and others2007). Mice models with inactivation of the insulin receptor inbeta cells exhibit a selective loss of insulin secretion and a pro-gressive impairment of glucose tolerance (Kulkarni and others1999). Flavonoids such as procyanidins have been shown to in-teract with insulin signaling pathways in other cell types (Pinentand others 2004). Therefore, they might modulate beta-cell func-tion, insulin secretion, and proliferation in part via interactionwith insulin signaling cascade.
In fact, other flavonoids have been demonstrated to interferewith insulin signaling cascade in pancreas. In pancreatic cell linesand mouse islets, the insulin-secreting activity of genistein is me-diated at least in part via cAMP accumulation and protein kinaseA (PKA) activation (Liu and others 2006), and is independent ofestrogen receptor mechanisms protein tyrosine kinases, and nitricoxide (NO)-signaling pathways.
Some flavonoids interfere with the glucose-induced depolar-ization of the cell membrane that initiates firing of action poten-tials that result on insulin secretion. Of green tea catechins, (–)-epigallocatechin-3-gallate (EGCG) and (–)-epicatechin-3-gallate(ECG), but not (–)-epicatechin and (–)-epigallocatechin, inhibitthe activity of ATP-sensitive potassium KATP channels at tens ofmicromolar concentrations, ECG being 3 times more effectivethan EGCG. Further, by using cloned beta-cell-type KATP chan-nels, these researchers showed that only EGCG at 1 µmol/L, areadily achievable plasma concentration by oral intake in hu-mans, but not other epicatechins, significantly blocked chan-nel reactivation after ATP washout, suggesting that interactionof phosphatidylinositol polyphosphates (PIP) with the channelwas impaired by EGCG (Jin and others 2007). Previously, Baekand others (2005), also reported that the gallate-ester moietyof epicatechins may be critical for inhibiting the KATP channelactivity via the pore-forming subunit Kir6.2 and this may be apossible mechanism by which green tea extracts or EGCG maycause unexpected side effects at the micromolar plasma level.Less agreement exists around resveratrol effects. Chen and oth-ers (2007) stated that resveratrol increases insulin secretion byblocking KATP and Kv channels in mouse beta-cell lines, and thiscould contribute to the increased plasma insulin in normal rats.Opposite effects were found by Szkudelski (2006, 2007, 2008)in pancreatic islets of normal rats, where resveratrol inhibitedinsulin release. The researchers found that interaction of resvera-trol with KATP was not involved in such inhibitory effects. Further,the researchers suggested that the impairment of mitochondrialmetabolism is involved in resveratrol based on their findings: (1)resveratrol inhibits as well insulin-secretion stimulated with thesecretagogues leucine, glutamine, and succinate (2) resveratrolreduced glucose oxidation, enhanced lactate release, decreasedATP and attenuated glucose-induced hyperpolarisation of the in-ner mitochondrial membrane.
The above-mentioned mechanism would be in addition to theantioxidant effects described for plant compounds, which wouldmodify the oxidative stress and/or modulate the action of reactivespecies that are in part responsible for the pancreatic dysfunction(Soto and others 2004; Coskun and others 2005; El-Alfy and oth-ers 2005; Yun and others 2006; Feick and others 2007).
ConclusionsThe great diversity of flavonoid structures makes it difficult to
establish their common effects in the pancreas. In any case, it isclear that the effects depend not only on the flavonoid structurebut also on the experimental design, and remarkably so on theconcentrations tested. Studies using animals with drug-induceddiabetes support the hypothesis that flavonoids can amelioratethis pathogenesis. Published data suggest that there might be di-rect effects of flavonoids on insulin secretion, as well as pre-venting beta-cell apoptosis, and they could even act throughmodulation of proliferation. The mechanisms of action involvemainly their antioxidant properties, although other pathwaysmight also be involved. To date, the contribution of all theseflavonoid effects to the overall improvement of experimentaldiabetes remains obscure.
AcknowledgmentsThis study was supported by grant nr AGL2005-04889/ALI from
the Comision Interministerial de Ciencia y Tecnologıa (CICYT) ofthe Spanish Government. Anna Castell is the recipient of a fellow-ship from the autonomous government of Catalonia (Generalitatde Catalunya). Gemma Montagut is the recipient of a fellow-ship from the Rovira i Virgili Univ. in Tarragona. We are gratefulto Montserrat Vaque for kindly providing the flavonoid structurefigures.
ReferencesAbd El Mohsen M, Kuhnle G, Rechner A, Schroeter H, Rose S, Jenner P, Rice-Evans C. 2002.
Uptake and metabolism of epicatechin and its access to the brain after oral ingestion. FreeRadic Biol Med 33(12):1693–702.
Aherne S, O’Brien N. 2002. Dietary flavonols: chemistry, food content, and metabolism.Nutrition 18(1):75–81.
Aron P, Kennedy J. 2008. Flavan-3-ols: nature, occurrence and biological activity. Mol NutrFood Res 52(1):79–104.
Baek W, Jang B, Lim J, Kwon T, Lee H, Cho C, Kim D, Shin D, Park J, Lim J and others.2005. Inhibitory modulation of ATP-sensitive potassium channels by gallate-ester moietyof (–)-epigallocatechin-3-gallate. Biochem Pharmacol 70(11):1560–7.
Bagchi D, Bagchi M, Stohs S, Ray S, Sen C, Preuss H. 2002. Cellular protection with proan-thocyanidins derived from grape seeds. Ann N Y Acad Sci 957:260–70.
Bagchi D, Sen C, Ray S, Das D, Bagchi M, Preuss H, Vinson J. 2003. Molecular mechanisms ofcardioprotection by a novel grape seed proanthocyanidin extract. Mutat Res 523–4:87–97.
Bagust A, Beale S. 2003. Deteriorating beta-cell function in type 2 diabetes: a long-termmodel. QJM 96(4):281–8.
Baur J, Pearson K, Price N, Jamieson H, Lerin C, Kalra A, Prabhu V, Allard J, Lopez-Lluch G,Lewis K and others. 2006. Resveratrol improves health and survival of mice on a high-caloriediet. Nature 444(7117):337–42.
Bhagwat SA, Holden JM, Haytowitz DB, Gebhardt SE, Eldridge A, Dwyer J, PetersonJ. 2003. USDA database for the flavonoid content of selected foods. Available from:http://www.nal.usda.gov/fnic/foodcomp. Accessed Aug. 2008.
Chang H, Churchwell M, Delclos K, Newbold R, Doerge D. 2000. Mass spectrometric de-termination of Genistein tissue distribution in diet-exposed Sprague-Dawley rats. J Nutr130(8):1963–70.
Chen W-P, Chi T-C, Chuang L-M, Su M-J. 2007. Resveratrol enhances insulin secretion byblocking K(ATP) and K(V) channels of beta cells. Eur J Pharmacol 568(1–3):269–77.
Chi T, Chen W, Chi T, Kuo T, Lee S, Cheng J, Su M. 2007. Phosphatidylinositol-3-kinase isinvolved in the antihyperglycemic effect induced by resveratrol in streptozotocin-induceddiabetic rats. Life Sci 80(18):1713–20.
Choi M, Jung U, Yeo J, Kim M, Lee M. 2008. Genistein and daidzein prevent diabetes onset byelevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activitiesin non-obese diabetic (NOD) mice. Diabetes Metab Res Rev 24(1):74–81.
Cnop M, Welsh N, Jonas J, Jorns A, Lenzen S, Eizirik D. 2005. Mechanisms of pancreaticbeta-cell death in type 1 and type 2 diabetes: many differences, few similarities. Diabetes54(Suppl 2):S97–107.
Coldham N, Sauer M. 2000. Pharmacokinetics of [(14)C]Genistein in the rat: gender-relateddifferences, potential mechanisms of biological action, and implications for human health.Toxicol Appl Pharmacol 164(2):206–15.
306 COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY—Vol. 7, 2008

Flavonoids on beta cells . . .
Coldiron AJ, Sanders R, Watkins J. 2002. Effects of combined quercetin and coenzyme Q(10)treatment on oxidative stress in normal and diabetic rats. J Biochem Mol Toxicol 16(4):197–202.
Coskun O, Kanter M, Korkmaz A, Oter S. 2005. Quercetin, a flavonoid antioxidant, preventsand protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas.Pharmacol Res 51(2):117–23.
Crespy V, Morand C, Besson C, Cotelle N, Vezin H, Demigne C, Remesy C. 2003. Thesplanchnic metabolism of flavonoids highly differed according to the nature of the com-pound. Am J Physiol Gastrointest Liver Physiol 284(6):G980–8.
Datla K, Christidou M, Widmer W, Rooprai H, Dexter D. 2001. Tissue distribution andneuroprotective effects of citrus flavonoid tangeretin in a rat model of Parkinson’s disease.Neuroreport 12(17):3871–5.
Del Bas J, Fernandez-Larrea J, Blay M, Ardevol A, Salvado M, Arola L, Blade C. 2005. Grapeseed procyanidins improve atherosclerotic risk index and induce liver CYP7A1 and SHPexpression in healthy rats. FASEB J 19(3):479–81.
Dhawan S, Georgia S, Bhushan A. 2007. Formation and regeneration of the endocrine pan-creas. Curr Opin Cell Biol 19(6):634–45.
Du Y, Guo H, Lou H. 2007. Grape seed polyphenols protect cardiac cells from apoptosis viainduction of endogenous antioxidant enzymes. J Agric Food Chem 55(5):1695–701.
El-Alfy AT, Ahmed AAE, Fatani AJ. 2005. Protective effect of red grape seeds proanthocyani-dins against induction of diabetes by alloxan in rats. Pharmacol Res 52(3):264–70.
Elliott J, Scarpello J, Morgan N. 2002. Differential effects of genistein on apoptosis inducedby fluoride and pertussis toxin in human and rat pancreatic islets and RINm5F cells. JEndocrinol 172(1):137–43.
Feick P, Gerloff A, Singer M. 2007. Effect of non-alcoholic compounds of alcoholic drinkson the pancreas. Pancreatology 7(2–3):124–30.
Graf B, Mullen W, Caldwell S, Hartley R, Duthie G, Lean M, Crozier A, Edwards C. 2005.Disposition and metabolism of [2-14C]quercetin-4’-glucoside in rats. Drug Metab Dispos33(7):1036–43.
Han M. 2003. Epigallocatechin gallate, a constituent of green tea, suppresses cytokine-induced pancreatic beta-cell damage. Exp Mol Med 35(2):136–9.
Hara Y, Fujino M, Adachi K, Li X. 2006. The reduction of hypoxia-induced and reoxygenation-induced apoptosis in rat islets by epigallocatechin gallate. Transplant Proc 38(8):2722–5.
Harmon A, Harp J. 2001. Differential effects of flavonoids on 3T3-L1 adipogenesis and lipol-ysis. Am J Physiol Cell Physiol 280(4):C807–13.
Ichinose K, Kawasaki E, Eguchi K. 2007. Recent advancement of understanding pathogenesisof type 1 diabetes and potential relevance to diabetic nephropathy. Am J Nephrol 27(6):554–64.
Jayaprakasam B, Vareed SK, Olson LK, Nair MG. 2005. Insulin secretion by bioactive antho-cyanins and anthocyanidins present in fruits. J Agric Food Chem 53(1):28–31.
Jia Z, Zhu H, Misra B, Mahaney J, Li Y, Misra H. 2008. EPR studies on the superoxide-scavenging capacity of the nutraceutical resveratrol. Mol Cell Biochem 313(1–2):187–94.
Jin J, Park S, Bae J, Cho H, Lim J, Park W, Han J, Lee J, Song D. 2007. Uncoupling by (–)-epigallocatechin-3-gallate of ATP-sensitive potassium channels from phosphatidylinositolpolyphosphates and ATP. Pharmacol Res 56(3):237–47.
Jonas J, Plant T, Gilon P, Detimary P, Nenquin M, Henquin J. 1995. Multiple effects andstimulation of insulin secretion by the tyrosine kinase inhibitor genistein in normal mouseislets. Br J Pharmacol 114(4):872–80.
Kim M-J, Ryu GR, Chung J-S, Sim SS, Min DS, Rhie D-J, Yoon SH, Hahn SJ, Kim M-S, JoY-H. 2003. Protective effects of epicatechin against the toxic effects of streptozotocin onrat pancreatic islets: in vivo and in vitro. Pancreas 26(3):292–9.
Kim S, Lee M, Hong J, Li C, Smith T, Yang G, Seril D, Yang C. 2000. Plasma and tissue levelsof tea catechins in rats and mice during chronic consumption of green tea polyphenols.Nutr Cancer 37(1):41–8.
Koo S, Noh S. 2007. Green tea as inhibitor of the intestinal absorption of lipids: potentialmechanism for its lipid-lowering effect. J Nutr Biochem 18(3):179–83.
Kovacs EMR, Mela DJ. 2006. Metabolically active functional food ingredients for weightcontrol. Obes Rev 7(1):59–78.
Kulkarni R, Bruning J, Winnay J, Postic C, Magnuson M, Kahn C. 1999. Tissue-specific knock-out of the insulin receptor in pancreatic beta cells creates an insulin secretory defect similarto that in type 2 diabetes. Cell 96(3):329–39.
Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, Messadeq N, MilneJ, Lambert P, Elliott P and others. 2006. Resveratrol improves mitochondrial function andprotects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 127(6):1109–22.
Lapidot T, Walker M, Kanner J. 2002. Antioxidant and prooxidant effects of phenolics onpancreatic beta-cells in vitro. J Agric Food Chem 50(25):7220–5.
Lenzen S. 2008. The mechanisms of alloxan- and streptozotocin-induced diabetes. Dia-betologia 51(2):216–26.
Levine F, Itkin-Ansari P. 2007. beta-cell regeneration: Neogenesis, replication or both? J MolMed 86(3):247–58.
Li W, Zhang X, Wu Y, Tian X. 2001. Anti-inflammatory effect and mechanism of proantho-cyanidins from grape seeds. Acta Pharmacol Sin 22(12):1117–20.
Liu D, Zhen W, Yang Z, Carter JD, Si H, Reynolds KA. 2006. Genistein acutely stimulates in-sulin secretion in pancreatic beta-cells through a cAMP-dependent protein kinase pathway.Diabetes 55(4):1043–50.
Lizarraga D, Lozano C, Briede J, van Delft J, Tourino S, Centelles J, Torres J, Cascante M. 2007.The importance of polymerization and galloylation for the antiproliferative properties ofprocyanidin-rich natural extracts. FEBS J 274(18):4802–11.
Lotito S, Frei B. 2006. Consumption of flavonoid-rich foods and increased plasma antioxi-dant capacity in humans: cause, consequence, or epiphenomenon? Free Radic Biol Med41(12):1727–46.
Lu Y, Zhao W, Chang Z, Chen W, Li L. 2004. Procyanidins from grape seeds protectagainst phorbol ester-induced oxidative cellular and genotoxic damage. Acta PharmacolSin 25(8):1083–9.
Manach C, Williamson G, Morand C, Scalbert A, Remesy C. 2005. Bioavailability and bio-efficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am J Clin Nutr81(1 Suppl):230S–42S.
Masiello P, Broca C, Gross R, Roye M, Manteghetti M, Hillaire-Buys D, Novelli M, RibesG. 1998. Experimental NIDDM: development of a new model in adult rats administeredstreptozotocin and nicotinamide. Diabetes 47(2):224–9.
Matsuda T, Ferreri K, Todorov I, Kuroda Y, Smith C, Kandeel F, Mullen Y. 2005. Silymarinprotects pancreatic beta-cells against cytokine-mediated toxicity: implication of c-Jun NH2-terminal kinase and janus kinase/signal transducer and activator of transcription pathways.Endocrinology 146(1):175–85.
Meeran S, Katiyar S. 2008. Cell cycle control as a basis for cancer chemoprevention throughdietary agents. Front Biosci 13:2191–202.
Meier J, Bhushan A, Butler A, Rizza R, Butler P. 2005. Sustained beta cell apoptosis in patientswith long-standing type 1 diabetes: indirect evidence for islet regeneration? Diabetologia48(11):2221–8.
Meng X, Sang S, Zhu N, Lu H, Sheng S, Lee M, Ho C, Yang C. 2002. Identification and char-acterization of methylated and ring-fission metabolites of tea catechins formed in humans,mice, and rats. Chem Res Toxicol 15(8):1042–50.
Mullen W, Graf B, Caldwell S, Hartley R, Duthie G, Edwards C, Lean M, Crozier A. 2002.Determination of flavonol metabolites in plasma and tissues of rats by HPLC-radiocountingand tandem mass spectrometry following oral ingestion of [2-(14)C]quercetin-4’-glucoside.J Agric Food Chem 50(23):6902–9.
Muoio DM, Newgard CB. 2006. Obesity-related derangements in metabolic regulation. AnnuRev Biochem 75:367–401.
Noriega-Lopez L, Tovar AR, Gonzalez-Granillo M, Hernandez-Pando R, Escalante B,Santillan-Doherty P, Torres N. 2007. Pancreatic insulin secretion in rats fed a soy proteinhigh fat diet depends on the interaction between the amino acid pattern and isoflavones. JBiol Chem 282(28):20657–66.
Notoya M, Tsukamoto Y, Nishimura H, Woo J, Nagai K, Lee I, Hagiwara H. 2004. Quercetin,a flavonoid, inhibits the proliferation, differentiation, and mineralization of osteoblasts invitro. Eur J Pharmacol 485(1–3):89–96.
Ohno T, Kato N, Ishii C, Shimizu M, Ito Y, Tomono S, Kawazu S. 1993. Genistein augmentscyclic adenosine 3’5’-monophosphate(cAMP) accumulation and insulin release in MIN6cells. Endocr Res 19(4):273–85.
Persaud S, Harris T, Burns C, Jones P. 1999. Tyrosine kinases play a permissive role in glucose-induced insulin secretion from adult rat islets. J Mol Endocrinol 22(1):19–28.
Pinent M, Blay M, Blade MC, Salvado MJ, Arola L, Ardevol A. 2004. Grape seed-derivedprocyanidins have an antihyperglycemic effect in streptozotocin-induced diabetic rats andinsulinomimetic activity in insulin-sensitive cell lines. Endocrinology 145(11):4985–90.
Pinent M, Blade MC, Salvado MJ, Arola L, Hackl H, Quackenbush J, Trajanoski Z, Ardevol A.2005. Grape-seed derived procyanidins interfere with adipogenesis of 3T3-L1 cells at theonset of differentiation. Int J Obes (Lond) 29(8):934–41.
Pinent M, Blade C, Salvado MJ, Blay M, Pujadas G, Fernandez-Larrea J, Arola L, ArdevolA. 2006. Procyanidin effects on adipocyte-related pathologies. Crit Rev Food Sci Nutr46(7):543–50.
Saris WH, Asp NG, Bjorck I, Blaak E, Bornet F, Brouns F, Frayn KN, Furst P, Riccardi G,Roberfroid M and others. 1998. Functional food science and substrate metabolism. Br JNutr 80(Suppl 1):S47–75.
Scalbert A, Johnson I, Saltmarsh M. 2005. Polyphenols: antioxidants and beyond. Am J ClinNutr 81(1 Suppl):215S–217S.
Silberberg M, Morand C, Mathevon T, Besson C, Manach C, Scalbert A, Remesy C. 2006.The bioavailability of polyphenols is highly governed by the capacity of the intestine andof the liver to secrete conjugated metabolites. Eur J Nutr 45(2):88–96.
Singab ANB, El-Beshbishy HA, Yonekawa M, Nomura T, Fukai T. 2005. Hypoglycemic ef-fect of Egyptian Morus alba root bark extract: effect on diabetes and lipid peroxidation ofstreptozotocin-induced diabetic rats. J Ethnopharmacol 100(3):333–8.
Song E, Hur H, Han M. 2003. Epigallocatechin gallate prevents autoimmune diabetes inducedby multiple low doses of streptozotocin in mice. Arch Pharm Res 26(7):559–63.
Sorenson R, Brelje T, Roth C. 1994. Effect of tyrosine kinase inhibitors on islets of Langer-hans: evidence for tyrosine kinases in the regulation of insulin secretion. Endocrinology134(4):1975–8.
Soto C, Mena R, Luna J, Cerbon M, Larrieta E, Vital P, Urıa E, Sanchez M, Recoba R, Barron Hand others. 2004. Silymarin induces recovery of pancreatic function after alloxan damagein rats. Life Sci 75(18):2167–80.
Spencer J, Abd-el-Mohsen M, Rice-Evans C. 2004. Cellular uptake and metabolism offlavonoids and their metabolites: implications for their bioactivity. Arch Biochem Biophys423(1):148–61.
Stanley MPP, Kamalakkannan N. 2006. Rutin improves glucose homeostasis in streptozotocindiabetic tissues by altering glycolytic and gluconeogenic enzymes. J Biochem Mol Toxicol20(2):96–102.
Stevenson D, Hurst R. 2007. Polyphenolic phytochemicals—just antioxidants or much more?Cell Mol Life Sci 64(22):2900–16.
Su H, Hung L, Chen J. 2006. Resveratrol, a red wine antioxidant, possesses an insulin-like ef-fect in streptozotocin-induced diabetic rats. Am J Physiol Endocrinol Metab 290(6):E1339–46.
Suganuma M, Okabe S, Oniyama M, Tada Y, Ito H, Fujiki H. 1998. Wide distribution of[3H](-)-epigallocatechin gallate, a cancer preventive tea polyphenol, in mouse tissue. Car-cinogenesis 19(10):1771–6.
Swezey R, Aldridge D, LeValley S, Crowell J, Hara Y, Green C. 2003. Absorption, tissuedistribution and elimination of 4-[(3)h]-epigallocatechin gallate in beagle dogs. Int J Toxicol22(3):187–93.
Szkudelski T. 2006. Resveratrol inhibits insulin secretion from rat pancreatic islets. Eur JPharmacol 552(1–3):176–81.
Szkudelski T. 2007. Resveratrol-induced inhibition of insulin secretion from rat pancreaticislets: evidence for pivotal role of metabolic disturbances. Am J Physiol Endocrinol Metab293(4):E901–7.
Szkudelski T. 2008. The insulin-suppressive effect of resveratrol—an in vitro and in vivophenomenon. Life Sci 82(7–8):430–5.
Takabayashi F, Harada N, Tahara S, Kaneko T, Hara Y. 1997. Effect of green tea catechinson the amount of 8-hydroxydeoxyguanosine (8-OHdG) in pancreatic and hepatic DNAafter a single administration of N-nitrosobis(2-oxopropyl)amine (BOP). Pancreas 15(2):109–12.
Vol. 7, 2008—COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY 307

CRFSFS: Comprehensive Reviews in Food Science and Food Safety
Tourrel C, Bailbe D, Lacorne M, Meile M, Kergoat M, Portha B. 2002. Persistent improvementof type 2 diabetes in the Goto-Kakizaki rat model by expansion of the beta-cell mass duringthe prediabetic period with glucagon-like peptide-1 or exendin-4. Diabetes 51(5):1443–52.
Ueno I, Nakano N, Hirono I. 1983. Metabolic fate of [14C] quercetin in the ACI rat. Jpn J ExpMed 53(1):41–50.
Vafeiadou K, Vauzour D, Spencer J. 2007. Neuroinflammation and its modulation byflavonoids. Endocr Metab Immune Disord Drug Targets 7(3):211–24.
Veluri R, Singh R, Liu Z, Thompson J, Agarwal R, Agarwal C. 2006. Fractionation of grapeseed extract and identification of gallic acid as one of the major active constituents causinggrowth inhibition and apoptotic death of DU145 human prostate carcinoma cells. Carcino-genesis 27(7):1445–53.
Vessal M, Hemmati M, Vasei M. 2003. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp Biochem Physiol C Toxicol Pharmacol 135C(3):357–64.
Vitrac X, Desmouliere A, Brouillaud B, Krisa S, Deffieux G, Barthe N, Rosenbaum J, MerillonJ. 2003. Distribution of [14C]-trans-resveratrol, a cancer chemopreventive polyphenol, inmouse tissues after oral administration. Life Sci 72(20):2219–33.
Walle T. 2004. Absorption and metabolism of flavonoids. Free Radic Biol Med 36(7):829–37.Walle T. 2007. Methylation of dietary flavones greatly improves their hepatic metabolic
stability and intestinal absorption. Mol Pharm 4(6):826–32.
Wild S, Roglic G, Green A, Sicree R, King H. 2004. Global prevalence of diabetes: estimatesfor the year 2000 and projections for 2030. Diabetes Care 27(5):1047–53.
Williams R, Spencer J, Rice-Evans C. 2004. Flavonoids: antioxidants or signalling molecules?Free Radic Biol Med 36(7):838–49.
Williamson G, Day A, Plumb G, Couteau D. 2000. Human metabolic pathways of dietaryflavonoids and cinnamates. Biochem Soc Trans 28(2):16–22.
Xiong F, Sun X, Gan L, Yang X, Xu H. 2006. Puerarin protects rat pancreatic islets fromdamage by hydrogen peroxide. Eur J Pharmacol 529(1–3):1–7.
Youdim K, Martin A, Joseph J. 2000. Incorporation of the elderberry anthocyanins by en-dothelial cells increases protection against oxidative stress. Free Radic Biol Med 29(1):51–60.
Yun S, Kim S, Song D. 2006. Effects of (–)-epigallocatechin-3-gallate on pancreatic beta-cell damage in streptozotocin-induced diabetic rats. Eur J Pharmacol 541(1–2):115–21.
Zhang L, Zuo Z, Lin G. 2007. Intestinal and hepatic glucuronidation of flavonoids. Mol Pharm4(6):833–45.
Zhang Y, Jayaprakasam B, Seeram NP, Olson LK, DeWitt D, Nair MG. 2004. Insulin secretionand cyclooxygenase enzyme inhibition by cabernet sauvignon grape skin compounds. JAgric Food Chem 52(2):228–33.
308 COMPREHENSIVE REVIEWS IN FOOD SCIENCE AND FOOD SAFETY—Vol. 7, 2008




















