
Bioactivity of anti-angiogenic ribozymes targeting Flt1 and KDR mRNA
9
© 1999 Oxford University Press Nucleic Acids Research, 1999, Vol. 27, No. 13 2569–2577 Bioactivity of anti-angiogenic ribozymes targeting Flt-1 and KDR mRNA Tom J. Parry, Cynthia Cushman 1 , Anna M. Gallegos, Arun B. Agrawal, Michele Richardson, Lori E. Andrews, Lara Maloney, Victor R. Mokler, Francine E. Wincott and Pamela A. Pavco* Ribozyme Pharmaceuticals, Inc., 2950 Wilderness Place, Boulder, CO 80301, USA and 1 Chiron Corporation, 4560 Horton Street, Emeryville, CA 94608-2916, USA Received as resubmission May 12, 1999; Accepted May 14, 1999 ABSTRACT Vascular endothelial growth factor (VEGF) and its receptors Flt-1 and KDR play important roles in phys- iological and pathological angiogenesis. Ribozymes that target the VEGF receptor mRNAs were devel- oped and their biological activities in cell culture and an animal model were assessed. Ribozymes targeting Flt-1 or KDR mRNA sites reduced VEGF-induced pro- liferation of cultured human vascular endothelial cells and specifically lowered the level of Flt-1 or KDR mRNA present in the cells. Anti-Flt-1 and KDR ribozymes also exhibited anti-angiogenic activity in a rat corneal pocket assay of VEGF-induced angio- genesis. This report illustrates the anti-angiogenic potential of these ribozymes as well as their value in studying VEGF receptor function in normal and pathophysiologic states. INTRODUCTION Physiologic processes such as vascular development, wound healing and the endometrial cycle depend upon angiogenesis (1–4). Angiogenesis also plays an important role in the patho- genesis of several diseases such as diabetic retinopathy, neovascular glaucoma, solid tumor growth and metastasis, endometriosis, rheumatoid arthritis and psoriasis (2,5,6). Because few effective therapies exist for these diseases, the development of anti-angiogenic agents could have a substantial impact on their treatment. Vascular endothelial growth factor (VEGF) has been shown to be an important mediator of vascular endothelial cell proliferation and migration (4,7). As a consequence, VEGF-induced angio- genesis has been shown to support solid tumor growth (6,8) and to underlie the pathology of certain neovascular diseases of the eye (9,10). In these diseases, ischemia-induced local hypoxia is thought to enhance VEGF secretion and thereby stimulate angiogenesis (4,11–13). A causative role for VEGF has been established in diabetic retinopathy (9,10,14–16) and is implicated in age-related macular degeneration (17,18). VEGF production is also elevated in synovial explants from patients with rheumatoid arthritis (19) and in psoriatic skin (20). Endothelial cells are the primary cell type known to express VEGF receptors (21). The two human VEGF receptor subtypes are known as Flt-1 and KDR. Although both are tyrosine kinases, the precise function of each VEGF receptor subtype in the process of normal angiogenesis is not known. In addition, their roles in pathologic angiogenesis may be obscured by changes in their relative expression as a function of disease (20,22) and by the presence of other cytokine and inflammatory mediators. The reduction of VEGF receptor signaling represents a possible anti- angiogenic strategy as well as an avenue to explore the relative role of each receptor in pathologic neovascularization. Ribozymes are RNA-based enzymes that bind to and cleave RNA molecules in a sequence-specific manner. This results in the destruction of the message and decreased expression of the encoded protein. Because ribozymes can be designed to target practically any RNA, the potential use of ribozymes to reduce gene expression is quite broad. Therapeutic applications of ribozymes have been facilitated by the development of cata- lytically active ribozymes that are chemically stabilized to resist nuclease degradation (23) and maintain activity in vivo (24). Here we describe the use of hammerhead ribozymes targeting Flt-1 and KDR mRNA as potential inhibitors of angiogenesis. Ribozymes targeting each receptor message were screened in cultured human microvascular endothelial cells (HMVEC-d) to determine their effect on VEGF-induced cell proliferation. Ribozymes that inhibited cell proliferation were also shown to decrease the abundance of targeted mRNA present in the cell. In addition, angiogenesis was significantly reduced when either an Flt-1 or a KDR targeted ribozyme was tested in a rat corneal model of VEGF-induced angiogenesis. Our results suggest that anti-Flt-1 and/or anti-KDR ribozymes may be effective therapeutic agents as well as useful tools for identifying the roles of Flt-1 and KDR receptors in angiogenesis. MATERIALS AND METHODS Ribozymes Hammerhead ribozymes and controls designed to have attenuated activity (attenuated controls) were synthesized and purified as previously described (25). The attenuated ribozyme controls maintain the binding arm sequence of the parent ribozyme and thus are still capable of binding to the mRNA target. However, *To whom correspondence should be addressed. Tel: +1 303 449 6500; Fax: +1 303 449 6995; Email: [email protected] Present address: Cynthia Cushman, Elan Corporation, 800 Gateway Boulevard, South San Francisco, CA 94080, USA
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Bioactivity of anti-angiogenic ribozymes targeting Flt1 and KDR mRNA

© 1999 Oxford University Press Nucleic Acids Research, 1999, Vol. 27, No. 132569–2577
sses
es,essinheirthehenti-tive
avein
f therget
ceof
ta-ist
ting.d in).
tocell.entlts
ng
atedas
olsndr,
Bioactivity of anti-angiogenic ribozymes targeting Flt-1and KDR mRNATom J. Parry, Cynthia Cushman 1, Anna M. Gallegos, Arun B. Agrawal, Michele Richardson,Lori E. Andrews, Lara Maloney, Victor R. Mokler, Francine E. Wincott and Pamela A. Pavco*
Ribozyme Pharmaceuticals, Inc., 2950 Wilderness Place, Boulder, CO 80301, USA and 1Chiron Corporation,4560 Horton Street, Emeryville, CA 94608-2916, USA
Received as resubmission May 12, 1999; Accepted May 14, 1999
ABSTRACT
Vascular endothelial growth factor (VEGF) and itsreceptors Flt-1 and KDR play important roles in phys-iological and pathological angiogenesis. Ribozymesthat target the VEGF receptor mRNAs were devel-oped and their biological activities in cell culture andan animal model were assessed. Ribozymes targetingFlt-1 or KDR mRNA sites reduced VEGF-induced pro-liferation of cultured human vascular endothelialcells and specifically lowered the level of Flt-1 orKDR mRNA present in the cells. Anti- Flt-1 and KDRribozymes also exhibited anti-angiogenic activity in arat corneal pocket assay of VEGF-induced angio-genesis. This report illustrates the anti-angiogenicpotential of these ribozymes as well as their value instudying VEGF receptor function in normal andpathophysiologic states.
INTRODUCTION
Physiologic processes such as vascular development, woundhealing and the endometrial cycle depend upon angiogenesis(1–4). Angiogenesis also plays an important role in the patho-genesis of several diseases such as diabetic retinopathy,neovascular glaucoma, solid tumor growth and metastasis,endometriosis, rheumatoid arthritis and psoriasis (2,5,6).Because few effective therapies exist for these diseases, thedevelopment of anti-angiogenic agents could have a substantialimpact on their treatment.
Vascular endothelial growth factor (VEGF) has been shown tobe an important mediator of vascular endothelial cell proliferationand migration (4,7). As a consequence, VEGF-induced angio-genesis has been shown to support solid tumor growth (6,8)and to underlie the pathology of certain neovascular diseasesof the eye (9,10). In these diseases, ischemia-induced localhypoxia is thought to enhance VEGF secretion and therebystimulate angiogenesis (4,11–13). A causative role for VEGFhas been established in diabetic retinopathy (9,10,14–16) andis implicated in age-related macular degeneration (17,18).VEGF production is also elevated in synovial explants frompatients with rheumatoid arthritis (19) and in psoriatic skin (20).
Endothelial cells are the primary cell type known to expreVEGF receptors (21). The two human VEGF receptor subtypare known as Flt-1 and KDR. Although both are tyrosine kinasthe precise function of each VEGF receptor subtype in the procof normal angiogenesis is not known. In addition, their rolespathologic angiogenesis may be obscured by changes in trelative expression as a function of disease (20,22) and bypresence of other cytokine and inflammatory mediators. Treduction of VEGF receptor signaling represents a possible aangiogenic strategy as well as an avenue to explore the relarole of each receptor in pathologic neovascularization.
Ribozymes are RNA-based enzymes that bind to and cleRNA molecules in a sequence-specific manner. This resultsthe destruction of the message and decreased expression oencoded protein. Because ribozymes can be designed to tapractically any RNA, the potential use of ribozymes to redugene expression is quite broad. Therapeutic applicationsribozymes have been facilitated by the development of calytically active ribozymes that are chemically stabilized to resnuclease degradation (23) and maintain activityin vivo(24).
Here we describe the use of hammerhead ribozymes targeFlt-1 andKDR mRNA as potential inhibitors of angiogenesisRibozymes targeting each receptor message were screenecultured human microvascular endothelial cells (HMVEC-dto determine their effect on VEGF-induced cell proliferationRibozymes that inhibited cell proliferation were also showndecrease the abundance of targeted mRNA present in theIn addition, angiogenesis was significantly reduced wheither anFlt-1 or aKDR targeted ribozyme was tested in a racorneal model of VEGF-induced angiogenesis. Our resusuggest that anti-Flt-1 and/or anti-KDR ribozymes may beeffective therapeutic agents as well as useful tools for identifyithe roles of Flt-1 and KDR receptors in angiogenesis.
MATERIALS AND METHODS
Ribozymes
Hammerhead ribozymes and controls designed to have attenuactivity (attenuated controls) were synthesized and purifiedpreviously described (25). The attenuated ribozyme contrmaintain the binding arm sequence of the parent ribozyme athus are still capable of binding to the mRNA target. Howeve
*To whom correspondence should be addressed. Tel: +1 303 449 6500; Fax: +1 303 449 6995; Email: [email protected] address:Cynthia Cushman, Elan Corporation, 800 Gateway Boulevard, South San Francisco, CA 94080, USA

2570 Nucleic Acids Research, 1999, Vol. 27, No. 13
atd
r/re
dd
nted
msasth
g thet
en.
velar
of6eris
forrchized
-e
ngl.nt
e
s
neras
they have two nucleotide changes in the core sequence thatsubstantially reduce their ability to carry out the cleavage reaction.Ribozymes were designed to targetFlt-1 or KDR mRNA sitesconserved in human, mouse and rat. In general, ribozymes withbinding arms of 7 nucleotides (nt) were designed and tested. If,however, only 6 nt surrounding the cleavage site were conservedin all three species, 6-nt binding arms were used. A subset ofribozyme and attenuated control sequences and modificationsare listed in Table 1. Other sequences are available uponrequest. In studies reported here, two previously describedribozyme motifs (23) were tested. While both motifs yieldedsimilar results in all test systems, data are presented for 2'-NH2uridine modified ribozymes in cell proliferation studies and for2'-C-allyl uridine modified ribozymes in RNase protection,invitro cleavage and corneal studies.
In vitro ribozyme cleavage assays
In vitro RNA cleavage rates on a 15-nt synthetic RNA substratewere measured as previously described (23). Briefly, theribozymes or attenuated controls (500 nM) were incubatedwith stoichiometric amounts of a short complementary RNAsubstrate (and trace amounts of 5'-32P-end-labeled substrate) in50 mM Tris–HCl buffer, pH 8.0, 40 mM MgCl2 at 37°C. Themagnesium-dependent cleavage reactions were quenched atspecified timepoints with EDTA-containing buffer. Followingpolyacrylamide gel electrophoresis, full-length and product bandswere quantified using a Molecular Dynamics PhosphorImager.
Cell culture
Human dermal microvascular endothelial cells (HMVEC-d,Clonetics Corp.) were maintained at 37°C in flasks or platescoated with 1.5% porcine skin gelatin (300 bloom, Sigma) inGrowth medium (Clonetics Corp.) supplemented with 10–20%fetal bovine serum (FBS, Hyclone). Cells were grown to confluencyand used up to the seventh passage. Stimulation mediumconsisted of 50% Sigma 99 media and 50% RPMI 1640 withL-glutamine and additional supplementation with 10µg/ml insulin–transferrin–selenium (Gibco BRL) and 10% FBS. Cell growth wasstimulated by incubation in Stimulation medium supplementedwith 20 ng/ml of either VEGF165 or bFGF. VEGF165(165 aminoacids) was selected for cell culture and animal studies becauseit is the predominant form of the four native forms of VEGFgenerated by alternative mRNA splicing (4). Cell cultureassays were carried out in triplicate.
Ribozyme and ribozyme/LipofectAMINE™ formulation
Cell culture.Ribozymes or attenuated controls (50–200 nM)were formulated for cell culture studies and used immediately.Formulations were carried out with LipofectAMINE™ (GibcoBRL) at a 3:1 lipid to phosphate charge ratio in serum-freemedium (Opti-MEM™, Gibco BRL) by mixing for 20 min atroom temperature. For example, a 3:1 lipid to phosphatecharge ratio was established by complexing 200 nM ribozymewith 10.8µg/µl LipofectAMINE™ (13.5µM DOSPA).
In vivo. For corneal studies, lyophilized ribozyme or attenuatedcontrols were resuspended in sterile water at a final stockconcentration of 170µg/µl (highest dose). Lower doses (1.7–50 µg/µl) were prepared by serial dilution in sterile water.
Proliferation assay
HMVEC-d were seeded (5× 103 cells per well) in 48-wellplates (Costar) and incubated for 24–30 h in Growth medium37°C. After removal of the Growth medium, cells were treatewith 50–200 nM LipofectAMINE™ complexes of ribozyme oattenuated controls for 2 h in Opti-MEM™. The ribozymecontrol-containing medium was removed and the cells wewashed extensively in 1× PBS. The medium was then replacewith Stimulation medium or Stimulation medium supplementewith 20 ng/ml VEGF165 or bFGF. After 48 h, the cell numberwas determined using a Coulter™ cell counter. Data are preseas cell number per well following 48 h of VEGF stimulation.
RNase protection assay
HMVEC-d were seeded (2× 105 cells per well) in 6-well plates(Costar) and allowed to grow for 32–36 h in Growth mediuat 37°C. Cells were treated with LipofectAMINE™ complexecontaining 200 nM ribozyme or attenuated control for 2 hdescribed in Proliferation Assay and then incubated in Growmedium containing 20 ng/ml VEGF165for 24 h. Cells were har-vested and an RNase protection assay was carried out usinAmbion Direct Protect kit and protocol with the exception tha50 mM EDTA was added to the lysis buffer to eliminate thpossibility of ribozyme cleavage during sample preparatioAntisense RNA probes targeting portions ofFlt-1 and KDRwere prepared by transcription in the presence of [32P]UTP.Samples were analyzed on polyacrylamide gels and the leof protected RNA fragments was quantified using a MoleculDynamics PhosphorImager. The levels ofFlt-1 andKDR werenormalized to the level of cyclophilin (human cyclophilinprobe template, Ambion) in each sample. The coefficientvariation for cyclophilin levels was 11% [265 940 c.p.m. ± 29 38(SD)] for all conditions tested here (i.e. in the presence of eithactive ribozymes or attenuated controls). Thus, cyclophilinuseful as an internal standard in these studies.
Rat corneal pocket assay of VEGF-induced angiogenesis
Animal guidelines and anesthesia. Animal housing and exper-imentation adhered to standards outlined in the 1996 Guidethe Care and Use of Laboratory Animals (National ReseaCouncil). Male Sprague Dawley rats (250–300 g) were anesthetwith ketamine (50 mg/kg), xylazine (10 mg/kg) and acepromazine (0.5 mg/kg) administered intramuscularly (im). Thlevel of anesthesia was monitored every 2–3 min by applyihind limb paw pressure and examining for limb withdrawaAtropine (0.4 mg/kg, im) was also administered to prevepotential corneal reflex-induced bradycardia.
Preparation of VEGF soaked disk. For corneal implantation,0.57 mm diameter nitrocellulose disks, prepared from 0.45µmpore diameter nitrocellulose filter membranes (MilliporCorporation), were soaked for 30 min in 1µl of 30 µM VEGF165in 82 mM Tris–HCl (pH 6.9) in covered Petri dishes on ice.
Corneal surgery. The rat corneal model used in this study wamodified from Kochet al. (26) and Pandeyet al. (27). Briefly,corneas were irrigated with 0.5% povidone iodine solutiofollowed by normal saline and two drops of 2% lidocaine. Unda dissecting microscope (Leica MZ-6), a stromal pocket w

Nucleic Acids Research, 1999, Vol. 27, No. 132571
lus,s
iniskhe
toed.F/as
io-hisgr
ical.
hiseree
edesyd
esre
created and a presoaked filter disk (see above) was inserted intothe pocket such that its edge was 1 mm from the corneal limbus.
Intraconjunctival injection of test solutions. Immediately afterdisk insertion, the tip of a 40–50µm OD injector (constructedin our laboratory) was inserted within the conjunctival tissue1 mm away from the edge of the corneal limbus that wasdirectly adjacent to the VEGF-soaked filter disk. 600 nl of testsolution (ribozyme, attenuated control or sterile water vehicle)were dispensed at a rate of 1.2µl/min using a syringe pump(Kd Scientific). The injector was then removed, serially rinsedin 70% ethanol and sterile water and immersed in sterile waterbetween each injection. Once the test solution was injected,closure of the eyelid was maintained using microaneurismclips until the animal began to recover gross motor activity.Following treatment, animals were warmed on a heating pad at37°C.
Animal treatment groups/experimental protocol. RibozymestargetingFlt-1 site 4229 andKDR mRNA site 726 were testedin the corneal model along with their attenuated controls. Fivetreatment groups were assigned to examine the effects of fivedoses of each test substance over a dose range of 1–100µg onVEGF-stimulated angiogenesis. Negative (30µM VEGFsoaked filter disk and intraconjunctival injection of 600 nl sterilewater) and no stimulus (Tris-soaked filter disk and intra-conjunctival injection of sterile water) control groups werealso included. Each group consisted of five animals (10 eyes)receiving the same treatment.
Quantitation of angiogenic response. Five days after diskimplantation, animals were euthanized following im adminis-tration of 0.4 mg/kg atropine and corneas were digitallyimaged. The neovascular surface area (NSA, expressed inpixels) was measured post-mortem from blood-filled corneal
vessels using computerized morphometry (Image Pro PMedia Cybernetics, v.2.0). The individual mean NSA wadetermined in triplicate from three regions of identical sizethe area of maximal neovascularization between the filter dand the limbus. The number of pixels corresponding to tblood-filled corneal vessels in these regions was summatedproduce an index of NSA. A group mean NSA was then calculatData from each treatment group were normalized to VEGribozyme vehicle-treated control NSA and finally expressedpercent inhibition of VEGF-induced angiogenesis.
Statistics. After determining the normality of treatment groupmeans, group mean percent inhibition of VEGF-induced anggenesis was subjected to a one-way analysis of variance. Twas followed by two post-hoc tests for significance includinDunnett’s (comparison to VEGF control) and Tukey-Krame(all other group mean comparisons) at alpha = 0.05. Statistanalyses were performed using JMP v.3.1.6 (SAS Institute)
RESULTS
Ribozyme-mediated reduction of VEGF-induced cellproliferation
Ribozyme cleavage ofFlt-1 or KDR mRNA should result in adecrease in the density of cell surface VEGF receptors. Tdecrease should limit VEGF binding and consequently interfwith the mitogenic signaling induced by VEGF. To determinif cell proliferation was impacted by anti-Flt-1 and/or anti-KDR ribozyme treatment, proliferation assays using culturhuman microvascular cells were carried out. Ribozymincluded in the proliferation assays were initially chosen btheir ability to decrease the level of VEGF binding to treatecells (data not shown). In these initial studies, ribozymtargeting 20 sites in the coding region of each mRNA we
Table 1.Ribozyme and attenuated control sequences and locations of modified nucleotides
*Ribozyme target sites are named according to the cleavage position in humanFlt-1 (DDBJ/EMBL/GenBank accession no. X51602) or humanKDR mRNA (DDBJ/EMBL/GenBank accession no. L04947).†Modifications are indicated as follows: 2'-O-methyl nucleotides, lowercase; ribonucleotides, uppercase G,A; inverted 3'-3' deoxyabasic,B. Two ribozyme motifs were tested (23). For the NH2-modified ribozymes,U indicates the two core positions where 2'-NH2 uridines are present. For theC-allyl-modified ribozyme,one 2'-C-allyl uridine is present at the 5'-mostU, while the second or 3'-mostU is 2'-O-methyl uridine. Thepositions of four phosphorothioate linkages at the 5' end are indicated by a subscript s. Changes in the attenuatedcontrols are underlined. Nucleotides comprising the base-paired region of stem II are in italics. A 3 bp stem II isshown above. Ribozymes or attenuated controls referred to in the text with a 4 bp stem II have one additional basepair such that the stem II/loop sequence isggccgaaaggcc.

2572 Nucleic Acids Research, 1999, Vol. 27, No. 13
nts.
ere,
ificted
aof
eoftrols
eirteatee
NE™wasae
ed
p ster
h
screened. The most effective ribozymes against two sites ineach target (Table 1),Flt-1 sites 1358 and 4229 andKDRsites726 and 3950, were included in the proliferation assaysreported here (Fig. 1). In addition, attenuated analogs of eachribozyme were used as controls (Table 1). These attenuatedcontrols are still capable of binding to the mRNA target sincethe binding arm sequence is maintained. However, these controlshave two nucleotide changes in the core sequence that substan-tially reduce their ability to carry out the cleavage reaction.
The anti-proliferative effect of active ribozymes targetingtwo lead sites on each VEGF receptor mRNA is shown inFigure 1. The active ribozymes tested decreased the relativeproliferation of HMVEC-d after VEGF stimulation, an effectthat increased with ribozyme concentration. This concentrationdependency was not observed following treatment with theattenuated controls designed for these sites. In fact, little or nochange in cell growth was noted following treatment with theattenuated controls, even though these controls can still bind tothe specific target sequences. At 200 nM, there was a distinct‘window’ between the anti-proliferative effects of eachribozyme and its attenuated control; a trend also observed atlower doses. This window of inhibition of proliferation (56–77%based on total cells per well) reflects the contribution ofribozyme-mediated activity. In comparison, no effect of anti-Flt-1 or anti-KDR ribozymes was noted on bFGF-stimulatedcell proliferation (Fig. 1C and F). Moreover, an irrelevant, butactive, ribozyme whose binding sequence is not found in eitherFlt-1 or KDR mRNA had no effect in this assay (Fig. 1B).These data are consistent with the basic ribozyme mechanism
in which binding and cleavage are necessary componeAlthough the relative surface distribution of Flt-1 and KDRreceptors in this cell type is not known, the anti-proliferativeffects of these ribozymes indicate that, at least in cell cultuboth receptors are functionally coupled to proliferation.
Specific reduction ofFlt-1 or KDR mRNA by ribozymetreatment
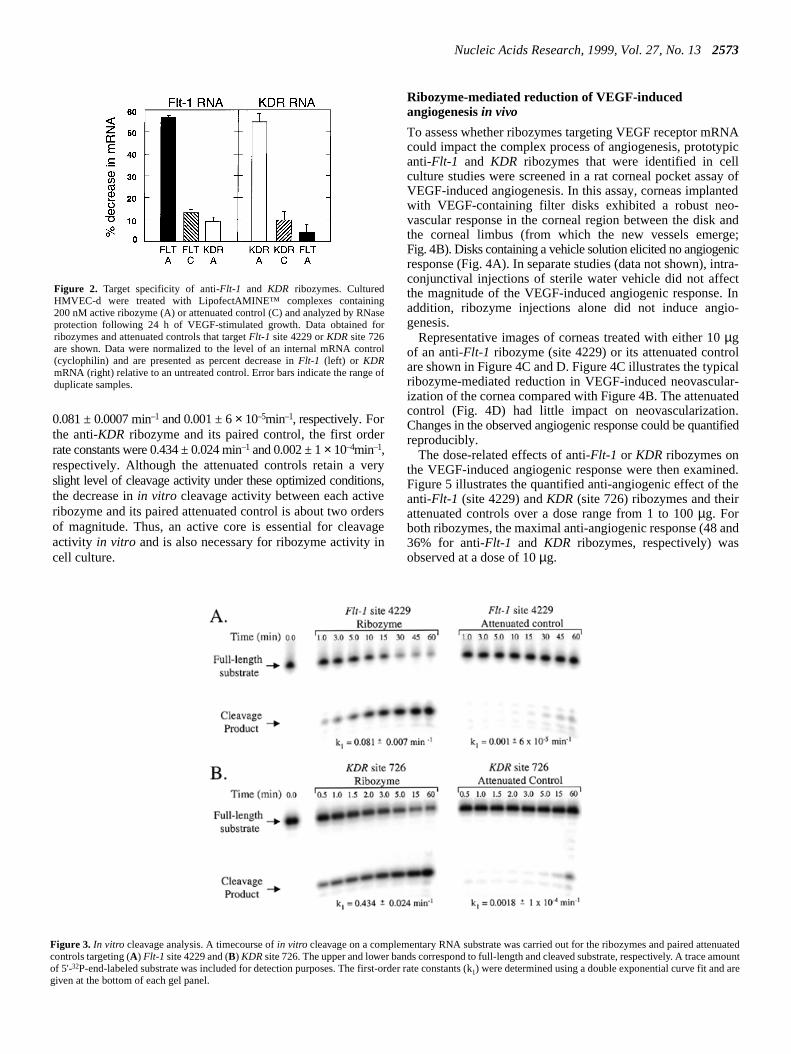
To confirm that anti-Flt-1 and anti-KDR ribozymes reducetheir respective mRNA targets, cellular levels ofFlt-1 or KDRwere quantified using an RNase protection assay with specFlt-1 or KDR probes. For each target, one ribozyme/attenuacontrol pair was chosen for continued study. Data fromrepresentative experiment are shown in Figure 2. ExposureHMVEC-d to active ribozyme targetingFlt-1 site 4229decreasedFlt-1 mRNA, but notKDRmRNA. Likewise, treatmentwith the active ribozyme targetingKDR site 726 decreasedKDR, but not Flt-1 mRNA. Both ribozymes decreased thlevel of their respective target RNA by >50%. The degreereduction associated with the corresponding attenuated conwas not >13%.
In vitro activity of anti- Flt and anti-KDR ribozymes
To confirm further the necessity of an active ribozyme core,invitro cleavage activities were determined for theFlt-1 site4229 ribozyme and the KDR site 726 ribozyme as well as thpaired attenuated controls (Fig. 3). The first order raconstants calculated from the timecourse of short substrcleavage for the anti-Flt-1 ribozyme and its attenuated control wer
Figure 1. Ribozyme-mediated inhibition of cell proliferation. Cultured HMVEC-d were treated with ribozyme or attenuated controls as LipofectAMIcomplexes. After treatment, cells were stimulated with VEGF165or bFGF and allowed to grow for 48 h prior to determining the cell number. Each ribozymetested in triplicate at three concentrations and data are presented as mean cell number per well + SD. The data obtained following ribozyme treatmentnd VEGFstimulation are presented in (A) and (B) for anti-Flt-1 ribozymes and (D) and (E) for anti-KDR ribozymes. Representative data obtained following ribozymtreatment and bFGF stimulation are shown in (C) for one anti-Flt-1 ribozyme and in (F) for one anti-KDR ribozyme. In all panels, active ribozymes are representwith filled symbols; attenuated controls with open symbols. In addition to the ribozymes and attenuated controls listed in Table 1, a second set having the samesequences but with an additional basepair in the ‘stem II’ region of the ribozyme are also shown for VEGF-induced proliferation studies. These 4 bm IIribozymes and attenuated controls have one additional base pair such that the stem II/loop sequence isggccgaaaggcc. Therefore, ribozymes and controls with 3 o4 bp stem IIs are denoted with circles and squares, respectively. The data for one irrelevant ribozyme [filled triangle, (B)] are also shown. This irrelevant ribozymecontains an active core sequence but has no binding site in eitherFlt-1 or KDRmRNA. Its sequence is 5'-gsasasgsgaacUGAuGaggccgaaaggccGaaAgauggcT-3' witmodifications as in Table 1 except that T indicates a 3'-3' inverted deoxythymidine. For reference, the average number of cells in control wells after48 h in theabsence of VEGF or bFGF for each of the panels are as follows: (A), (B) and (C), 12 477 ± 617; (D), (E) and (F), 17 182 ± 1053.
Nucleic Acids Research, 1999, Vol. 27, No. 132573
NApic
y ofted-ande;ic
tra-tIn
o-
lal
ar-ted.
tified
ed.e
nd
nuatedamountre
0.081 ± 0.0007 min–1 and 0.001 ± 6× 10–5min–1, respectively. Forthe anti-KDR ribozyme and its paired control, the first orderrate constants were 0.434 ± 0.024 min–1 and 0.002 ± 1× 10–4min–1,respectively. Although the attenuated controls retain a veryslight level of cleavage activity under these optimized conditions,the decrease inin vitro cleavage activity between each activeribozyme and its paired attenuated control is about two ordersof magnitude. Thus, an active core is essential for cleavageactivity in vitro and is also necessary for ribozyme activity incell culture.
Ribozyme-mediated reduction of VEGF-inducedangiogenesisin vivo
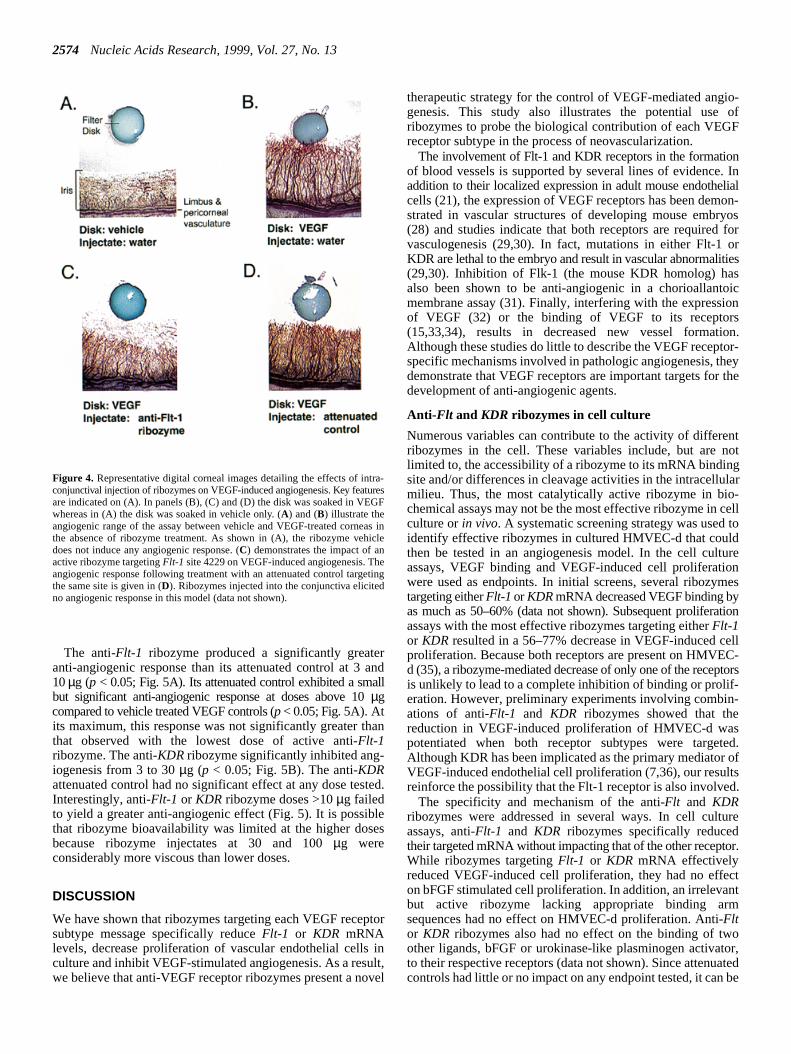
To assess whether ribozymes targeting VEGF receptor mRcould impact the complex process of angiogenesis, prototyanti-Flt-1 and KDR ribozymes that were identified in cellculture studies were screened in a rat corneal pocket assaVEGF-induced angiogenesis. In this assay, corneas implanwith VEGF-containing filter disks exhibited a robust neovascular response in the corneal region between the diskthe corneal limbus (from which the new vessels emergFig. 4B). Disks containing a vehicle solution elicited no angiogenresponse (Fig. 4A). In separate studies (data not shown), inconjunctival injections of sterile water vehicle did not affecthe magnitude of the VEGF-induced angiogenic response.addition, ribozyme injections alone did not induce angigenesis.
Representative images of corneas treated with either 10µgof an anti-Flt-1 ribozyme (site 4229) or its attenuated controare shown in Figure 4C and D. Figure 4C illustrates the typicribozyme-mediated reduction in VEGF-induced neovasculization of the cornea compared with Figure 4B. The attenuacontrol (Fig. 4D) had little impact on neovascularizationChanges in the observed angiogenic response could be quanreproducibly.
The dose-related effects of anti-Flt-1 or KDR ribozymes onthe VEGF-induced angiogenic response were then examinFigure 5 illustrates the quantified anti-angiogenic effect of thanti-Flt-1 (site 4229) andKDR (site 726) ribozymes and theirattenuated controls over a dose range from 1 to 100µg. Forboth ribozymes, the maximal anti-angiogenic response (48 a36% for anti-Flt-1 and KDR ribozymes, respectively) wasobserved at a dose of 10µg.
Figure 2. Target specificity of anti-Flt-1 and KDR ribozymes. CulturedHMVEC-d were treated with LipofectAMINE™ complexes containing200 nM active ribozyme (A) or attenuated control (C) and analyzed by RNaseprotection following 24 h of VEGF-stimulated growth. Data obtained forribozymes and attenuated controls that targetFlt-1 site 4229 orKDR site 726are shown. Data were normalized to the level of an internal mRNA control(cyclophilin) and are presented as percent decrease inFlt-1 (left) or KDRmRNA (right) relative to an untreated control. Error bars indicate the range ofduplicate samples.
Figure 3. In vitro cleavage analysis. A timecourse ofin vitro cleavage on a complementary RNA substrate was carried out for the ribozymes and paired attecontrols targeting (A) Flt-1 site 4229 and (B) KDRsite 726. The upper and lower bands correspond to full-length and cleaved substrate, respectively. A traceof 5'-32P-end-labeled substrate was included for detection purposes. The first-order rate constants (k1) were determined using a double exponential curve fit and agiven at the bottom of each gel panel.
2574 Nucleic Acids Research, 1999, Vol. 27, No. 13
io-ofF
n. Inlialon-
yosfororiessoiconson.or-eythe
ntot
glar-ellto
urenes
tion
ellC-tors
-
ed.oftsd.
re
or.
ctnt
oor,ted
be
The anti-Flt-1 ribozyme produced a significantly greateranti-angiogenic response than its attenuated control at 3 and10µg (p < 0.05; Fig. 5A). Its attenuated control exhibited a smallbut significant anti-angiogenic response at doses above 10µgcompared to vehicle treated VEGF controls (p < 0.05; Fig. 5A). Atits maximum, this response was not significantly greater thanthat observed with the lowest dose of active anti-Flt-1ribozyme. The anti-KDR ribozyme significantly inhibited ang-iogenesis from 3 to 30µg (p < 0.05; Fig. 5B). The anti-KDRattenuated control had no significant effect at any dose tested.Interestingly, anti-Flt-1 or KDR ribozyme doses >10µg failedto yield a greater anti-angiogenic effect (Fig. 5). It is possiblethat ribozyme bioavailability was limited at the higher dosesbecause ribozyme injectates at 30 and 100µg wereconsiderably more viscous than lower doses.
DISCUSSION
We have shown that ribozymes targeting each VEGF receptorsubtype message specifically reduceFlt-1 or KDR mRNAlevels, decrease proliferation of vascular endothelial cells inculture and inhibit VEGF-stimulated angiogenesis. As a result,we believe that anti-VEGF receptor ribozymes present a novel
therapeutic strategy for the control of VEGF-mediated anggenesis. This study also illustrates the potential useribozymes to probe the biological contribution of each VEGreceptor subtype in the process of neovascularization.
The involvement of Flt-1 and KDR receptors in the formatioof blood vessels is supported by several lines of evidenceaddition to their localized expression in adult mouse endothecells (21), the expression of VEGF receptors has been demstrated in vascular structures of developing mouse embr(28) and studies indicate that both receptors are requiredvasculogenesis (29,30). In fact, mutations in either Flt-1KDR are lethal to the embryo and result in vascular abnormalit(29,30). Inhibition of Flk-1 (the mouse KDR homolog) haalso been shown to be anti-angiogenic in a chorioallantmembrane assay (31). Finally, interfering with the expressiof VEGF (32) or the binding of VEGF to its receptor(15,33,34), results in decreased new vessel formatiAlthough these studies do little to describe the VEGF receptspecific mechanisms involved in pathologic angiogenesis, thdemonstrate that VEGF receptors are important targets fordevelopment of anti-angiogenic agents.
Anti- Flt and KDR ribozymes in cell culture
Numerous variables can contribute to the activity of differeribozymes in the cell. These variables include, but are nlimited to, the accessibility of a ribozyme to its mRNA bindinsite and/or differences in cleavage activities in the intracellumilieu. Thus, the most catalytically active ribozyme in biochemical assays may not be the most effective ribozyme in cculture orin vivo. A systematic screening strategy was usedidentify effective ribozymes in cultured HMVEC-d that couldthen be tested in an angiogenesis model. In the cell cultassays, VEGF binding and VEGF-induced cell proliferatiowere used as endpoints. In initial screens, several ribozymtargeting eitherFlt-1 or KDRmRNA decreased VEGF binding byas much as 50–60% (data not shown). Subsequent proliferaassays with the most effective ribozymes targeting eitherFlt-1or KDR resulted in a 56–77% decrease in VEGF-induced cproliferation. Because both receptors are present on HMVEd (35), a ribozyme-mediated decrease of only one of the recepis unlikely to lead to a complete inhibition of binding or prolif-eration. However, preliminary experiments involving combinations of anti-Flt-1 and KDR ribozymes showed that thereduction in VEGF-induced proliferation of HMVEC-d waspotentiated when both receptor subtypes were targetAlthough KDR has been implicated as the primary mediatorVEGF-induced endothelial cell proliferation (7,36), our resulreinforce the possibility that the Flt-1 receptor is also involve
The specificity and mechanism of the anti-Flt and KDRribozymes were addressed in several ways. In cell cultuassays, anti-Flt-1 and KDR ribozymes specifically reducedtheir targeted mRNA without impacting that of the other receptWhile ribozymes targetingFlt-1 or KDR mRNA effectivelyreduced VEGF-induced cell proliferation, they had no effeon bFGF stimulated cell proliferation. In addition, an irrelevabut active ribozyme lacking appropriate binding armsequences had no effect on HMVEC-d proliferation. Anti-Fltor KDR ribozymes also had no effect on the binding of twother ligands, bFGF or urokinase-like plasminogen activatto their respective receptors (data not shown). Since attenuacontrols had little or no impact on any endpoint tested, it can
Figure 4. Representative digital corneal images detailing the effects of intra-conjunctival injection of ribozymes on VEGF-induced angiogenesis. Key featuresare indicated on (A). In panels (B), (C) and (D) the disk was soaked in VEGFwhereas in (A) the disk was soaked in vehicle only. (A) and (B) illustrate theangiogenic range of the assay between vehicle and VEGF-treated corneas inthe absence of ribozyme treatment. As shown in (A), the ribozyme vehicledoes not induce any angiogenic response. (C) demonstrates the impact of anactive ribozyme targetingFlt-1 site 4229 on VEGF-induced angiogenesis. Theangiogenic response following treatment with an attenuated control targetingthe same site is given in (D). Ribozymes injected into the conjunctiva elicitedno angiogenic response in this model (data not shown).

Nucleic Acids Research, 1999, Vol. 27, No. 132575
peinly aed
ivegenti-
e-
ityth
nti-
otee
nti-
alo-innotllgen-
dedherlay
o-ysby
ayr
ic-1redo-
5).ven-ringe-nalor
the
stheionthat
ted
concluded that the effects of the active ribozymes are due to acleavage mechanism. Taken together, these results indicatethat anti-Flt-1 and KDR ribozymes reduce the level of theirrespective molecular targets and impact VEGF-mediated cellsignaling through a sequence-specific cleavage mechanism.
Anti- Flt and KDR ribozymes in vivo
Corneal pocket assays are widely used to study the complexbiology of angiogenesis and to identify potential therapeuticagents which impinge upon this process (26,37–43). Theseassays are reproducible, adaptable to specific pharmacologicstimulation (i.e. VEGF), exhibit low background and are amenableto quantitation. In the present study, we used a rat cornealpocket assay to examine whether anti-Flt-1 and KDRribozymes (targeting conserved sites between human, rat andmouse) impact VEGF-induced angiogenesis.
In general, ribozymes that target either VEGF receptor subtymRNA significantly reduced angiogenesis in the rat corneaa dose-dependent manner. Attenuated controls produced onlow level of anti-angiogenic activity at the highest doses test(30 or 100µg). This low level of activity was never significantlygreater than that observed with the lowest dose of actribozyme, and could be due to the slight amount of cleavaactivity retained by these controls. Therefore, the greater aangiogenic effect of the active anti-Flt-1 andKDR ribozymesversus the attenuated controls strongly supports a ribozymspecific mechanism.
The possibility exists that a greater anti-angiogenic activmay be achieved by combining ribozymes targeting boreceptor mRNAs. Although initial studies in culturedHMVEC-d suggest a potentiated response with combined aFlt-1 and KDR ribozyme treatment, preliminary anti-Flt-1/KDR ribozyme combination studies in the rat cornea did nenhance the anti-angiogenic activity over either ribozymalone (data not shown). We have not yet explored ribozymdosing combinations to assess their enhancement of aangiogenic efficacy.
Anti- Flt and KDR ribozymes and VEGF receptor function
Many factors complicate our understanding of the functioncontributions of the two VEGF receptor subtypes to nevascularization, particularly when considering their roledisease. For instance, there is some evidence that KDR butFlt-1 stimulation mediates VEGF-induced endothelial ceproliferation as well as enhanced motility, cell shape chanand actin reorganization (7,36). Our results indicate that dowregulation ofKDR or Flt-1 receptor mRNA leads to decreaseendothelial cell proliferation and decreased VEGF-inducangiogenesis. It is possible that the Flt-1 receptor can eitdirectly mediate these processes or that there may be interpbetween receptors (7,44,45).
Mutations in either receptor lead to abnormalities in nevascularization through presumably different pathwa(29,30). For KDR, vasculogenesis may be mediated directlychanges in endothelial cell development (30) while Flt-1 mbe important in cellular interactions with the extracellulamatrix (29). It appears that this effect of Flt-1 on embryologneovascularization is mediated by the binding domain of Fltrather than the kinase domain (46). In addition, there adifferences in the relative abilities of isoforms of VEGF anplacenta growth factor (and dimers thereof) to induce homand heterodimerization of KDR and Flt-1 receptors (7,44,4Further complicating this picture is the existence of a natisoluble form of the Flt-1 receptor that may serve to dowregulate VEGF-mediated angiogenesis (47,48) by sequesteVEGF and/or forming inactive heterodimers with membranspanning VEGF receptors. Thus, changes in post-transcriptioreceptor processing (47,48) as well as the effect of diseasehypoxia on receptor expression (49,50) may also impactrelative role of VEGF receptor subtypes in angiogenesis.
By virtue of their high degree of selectivity, ribozymetargeting VEGF receptor mRNA can be used to determinerelative role of each receptor subtypes in signal transductand neovascularization. The current study demonstratesthese ribozymes are effective in both cell culture andin vivoconditions. Thus, it is possible to use anti-Flt-1 and KDRribozymes to address receptor-specific concerns in isola
Figure 5. Anti-angiogenic efficacy of ribozymes in the rat corneal model ofVEGF-induced angiogenesis. The percent inhibition of VEGF-induced angio-genesis for locally administered (A) anti-Flt-1 (site 4229) and (B) anti-KDR(site 726) ribozymes (filled circles) and their attenuated controls (open circles)are plotted over the dose range tested. Pixels associated with background structuresincluding the iris (Fig. 3A) were subtracted from all treatment groups. Data areexpressed as mean percent reduction in VEGF-induced angiogenesis ± SEM.* p < 0.05 relative to VEGF/vehicle treated controls by Dunnett’s, **p < 0.05relative to attenuated dose-matched controls by Tukey-Kramer.

2576 Nucleic Acids Research, 1999, Vol. 27, No. 13
.,
,
.,
,
.
.,
.
G.,
cells (vascular endothelial, pericytes, macrophages, etc.) andtissue explants as well as in animal systems.
Anti- Flt-1 and KDR ribozymes as therapeutics
In addition to their utility in differentiating the role of VEGFreceptors, ribozymes that reduce VEGF receptor expressionhave broad therapeutic potential, especially where VEGF maymediate angiogenesis in solid tumors (22,33,34,51–55) andneovascular diseases of the eye (9,10,14–16,34,56). Severalstudies have shown that reducing VEGF signaling and sub-sequent angiogenesis through a variety of mechanisms canhave therapeutic effects. Treatment with neutralizing VEGFantibodies reduces tumor growth (33,34), metastasis (34) andtumor neovascularization (33,49) in a variety of human tumorxenograft models. In a VEGF-dependent model of retinalneovascularization (56,57), soluble VEGF-receptor chimericproteins (15) or antisense oligonucleotides targeting VEGFmessage (32) inhibited retinal neovascularization in responseto hypoxia. Also, retinal vein occlusion-induced neovascular-ization of the non-human primate iris was reduced followingtreatment with an anti-VEGF monoclonal antibody (16).
Summary
In conclusion, we have identified ribozymes directed againstFlt-1 andKDR mRNA that specifically reduce VEGF bindingand endothelial cell proliferation in cell culture as well asVEGF-induced angiogenesisin vivo. The therapeutic potentialof these ribozymes is currently being evaluated in a variety ofdisease models involving VEGF-mediated angiogenesis.Results from these studies will not only support the develop-ment of a potential therapeutic agent, but will also help clarifythe relative importance and functional contribution of eachVEGF receptor subtype in pathophysiological angiogenesis.
ACKNOWLEDGEMENTS
We wish to thank Dr Denis Gospodarowicz (Chiron Corp.) forsupplying VEGF165 and bFGF. Dr Lori Stephenson (ChironCorp.) provided the templates for RNase protection probes.
REFERENCES
1. Drake,C.J. and Little,C.D. (1995)Nature, 376, 70–74.2. Folkman,J. and Shing,Y. (1992)J. Biol. Chem., 267, 10931–10934.3. Jakeman,L., Armanini,M., Phillips,H.S. and Ferrara,N. (1993)
Endocrinology, 133, 848–859.4. Thomas,K.A. (1996)J. Biol. Chem., 271, 603–606.5. Folkman,J. (1990)J. Natl Cancer Inst., 82, 4–6.6. Folkman,J. and Klagsbrun,M. (1987)Science, 235, 442–447.7. Waltenberger,J., Claesson-Welsh,L., Siegbahn,A., Shibuya,M. and
Heldin,C.-H. (1994)J. Biol. Chem., 269, 26988–26995.8. Martiny-Baron,G. and Marmé,D. (1995)Curr. Opin. Biotechnol., 6, 675–680.9. Adamis,A.P., Shima,D.T., Yeo,K.T., Yeo,T.K., Brown,L.F., Berse,B.,
D’Amore,P.A. and Folkman,J. (1993)Biochem. Biophys. Res. Commun.,193, 631–638.
10. Aiello,L.P., Avery,R., Arrigg,P., Keyt,B., Jampel,H., Shah,S.,Pasquale,L., Thieme,H., Iwamoto,M., Park,J., Nguyen,H., Aiello,L.M.,Ferrara,N. and King,G. (1994)New Eng. J. Med., 331, 1480–1487.
11. Levy,A.P., Levy,N.S. and Goldberg,M.A. (1996)J. Biol. Chem.271,2746–2753.
12. Minchenko,A., Salceda,S., Bauer,T. and Caro,J. (1994)Cell. Mol. Biol. Res.,40, 35–39.
13. Shweiki,D., Itin,A., Soffer,D. and Keshet,E. (1992)Nature, 359, 843–845.14. Adamis,A., Miller,J., Bernal,M.-T., D’Amico,D., Folkman,J., Yeo,T.-K.
and Yeo,K.-T. (1994)Am. J. Ophthal., 118, 445–450.
15. Aiello,L., Pierce,E., Foley,E., Takagi,H., Chen,H., Riddle,L., Ferrara,NKing,G. and Smith,L. (1995)Proc. Natl Acad. Sci. USA, 92, 10457–10461.
16. Adamis,A.P., Shima,D.T., Tolentino,M.J., Gragoudas,E.S., Ferrara,N.D’Amore,P.A. and Miller,J.W. (1996)Arch. Ophthal., 114, 66–71.
17. Kvanta,A., Algvere,P.V., Berglin,L. and Seregard,S. (1996)Invest. Ophthal.Vis. Sci., 37, 1929–1934.
18. Lopez,P.F., Sippy,B.D., Lambert,H.M., Thach,A.B. and Hinton,D.R.(1996)Invest. Ophthal. Vis. Sci., 37, 855–868.
19. Koch,A.E., Harlow,L.A., Haines,G.K., Amento,E.P., Unemori,E.N.,Wong,W.L., Pope,R.M. and Ferrara,N. (1994)J. Immunol., 152, 4149–4156.
20. Detmar,M., Brown,L.F., Claffey,K.P., Yeo,K.-T., Kocher,O., Jackman,R.WBerse,B. and Dvorak,H.F. (1994)J. Exp. Med.180, 1141–1146.
21. Jakeman,L., Winer,J., Bennet,G.L., Altar,A. and Ferrara,N. (1992)J. Clin. Invest., 89, 244–253.
22. Plate,K.H., Breier,G., Millauer,B., Ullrich,A. and Risau,W. (1993)Cancer Res., 53, 5822–5827.
23. Beigelman,L., McSwiggen,J.A., Draper,K.G., Gonzalez,C., Jensen,K.Karpeisky,A.M., Modak,A.S., Matulic-Adamic,J., DiRenzo,A.B.,Haeberli,P., Sweedler,D., Tracz,D., Grimm,S., Wincott,F., Thackray,V.Gand Usman,N. (1995)J. Biol. Chem., 270, 25702–25708.
24. Flory,C.M., Pavco,P.A., Jarvis,T.C., Lesch,M.E., Wincott,F.E., Beigelman,LHunt,S.W.,III and Schrier,D.J. (1996)Proc. Natl Acad. Sci. USA, 93,754–758.
25. Wincott,F., DiRenzo,A., Shaffer,C., Grimm,S., Tracz,D., Workman,C.,Sweedler,D., Gonzalez,C., Scaringe,S. and Usman,N. (1995)Nucleic Acids Res., 23, 2677–2684.
26. Koch,A.E., Halloran,M.M., Haskell,C.J., Shah,M.R. and Polverini,P.J.(1995)Nature, 376, 517–519.
27. Pandey,A., Shao,H., Marks,R.M., Polverini,P.J. and Dixit,V.M. (1995)Science, 268, 567–599.
28. Millauer,B., Wizigmann-Voos,S., Schnürch,H., Martinez,R.,Moller,N.P.H., Risau,W. and Ullrich,A. (1993)Cell, 72, 835–846.
29. Fong,G.-H., Rossant,J., Gertsenstein,M. and Breitman,M.L. (1995)Nature, 376, 66–69.
30. Shalaby,F., Rossant,J., Yamaguchi,T.P., Gertsenstein,M., Wu,X.-F.,Breitman,M.L. and Schuh,A.C. (1995)Nature, 376, 62–66.
31. Strawn,L., McMahon,G., App,H., Schreck,R., Kuchler,W., Longhi,M.,Hui,T., Tang,C., Levitzki,A., Gazit,A., Chen,I., Keri,G., Orfi,L.,Risau,W., Flamme,I., Ullrich,A., Hirth,P. and Shawyer,L. (1996)Cancer Res., 56, 3540–3545.
32. Robinson,G.S., Pierce,E.A., Rook,S.L., Foley,E., Webb,R. andSmith,L.E.H. (1996)Proc. Natl Acad. Sci. USA, 93, 4851–4856.
33. Kim,K.J., Winer,J., Armanini,M., Gillett,N., Phillips,H.S. and Ferrara,N(1993)Nature, 362, 841–844.
34. Warren,R.S., Yuan,H., Matli,M.R., Gillett,N.A. and Ferrara,N. (1995)J. Clin. Invest., 95, 1789–1797.
35. Hewett,P.W. and Murray,J.C. (1996)Biochem. Biophys. Res. Commun.,221, 697–702.
36. Keyt,B.A., Nguyen,H.V., Berleau,L.T., Duarte,C.M., Park,J., Chen,H.and Ferrara,N. (1996)J. Biol. Chem., 271, 5638–5646.
37. BenEzra,D., Griffen,B.W., Maftzir,G. and Aharonov,O. (1993)Invest. Ophthal. Vis. Sci., 34, 3601–3608.
38. Burger,P.C., Chandler,D.B. and Klintworth,G.K. (1985/1986)Cornea, 4,35–41.
39. Gimbrone,M.A.J., Cotran,R.S., Leapman,S.B. and Folkman,J. (1974)J. Natl Cancer Inst., 52, 413–427.
40. Glatt,H.J. and Klintworth,G.K. (1986)Microvasc. Res., 31, 104.41. O’Reilly,M.S., Holmgren,L., Shing,Y., Chen,C., Rosenthal,R.A.,
Moses,M., Lane,W.S., Cao,Y., Sage,E.H. and Folkman,J. (1994)Cell, 79,315–328.
42. Strieter,R.M., Kunkel,S.L., Elner,V.M., Martonyi,C.L., Koch,A.E.,Polverini,P.J. and Elner,S.G. (1992)Am. J. Pathol., 141, 1279–1284.
43. Sunderkötter,C., Beil,W., Roth,J. and Sorg,C. (1991)Am. J. Pathol., 138,931–939.
44. Barleon,B., Totzke,F., Herzog,C., Blanke,S., Kremmer,E., Siemeister,Marmé,D. and Martiny-Baron,G. (1997)J. Biol. Chem., 272, 10382–10388.
45. Maglione,D., Guerriero,V., Viglietto,G., Ferraro,M.G., Aprelikova,O.,Alitalo,K., del Vecchio,S., Lei,K.J., Chon,J.Y. and Persico,M.G. (1993)Oncogene, 8, 925–931.
46. Hirasuka,S., Minowa,O., Kuno,J., Noda,T. and Shibuga,M. (1998)Proc. Natl Acad. Sci. USA, 95, 9349–9354.
47. Kendall,R.L. and Thomas,K.A. (1993)Proc. Natl Acad. Sci. USA, 90,10705–10709.

Nucleic Acids Research, 1999, Vol. 27, No. 132577
)
.
48. Goldman,C.K., Kendall,R.L., Cabrera,G., Soroceanu,L., Heike,Y.,Gillespie,G.Y., Siegal,G.P., Mao,X., Bett,A.J., Huckle,W.R.,Thomas,K.A. and Curiel,D.T. (1998)Proc. Natl Acad. Sci. USA,95, 8795–8800.
49. Takagi,H., King,G.L. and Aiello,L.P. (1996)Diabetes45, 1016–1023.50. Marti,H.H. and Risau,W. (1998)Proc. Natl Acad. Sci. USA, 95,
15809–15814.51. Yuan,F., Chen,Y., Dellian,M., Safabakhsh,N., Ferrara,N. and Jain,R.K.
(1996)Proc. Natl Acad. Sci. USA, 93, 14765–14770.52. Cohen,T., Gitay-Goren,H., Sharon,R., Shibuya,M., Halaban,R.,
Levi,B.-Z. and Neufeld,G. (1995)J. Biol. Chem., 270, 11322–11326.
53. Liu,B., Earl,H.M., Baban,D., Shoaibi,M., Fabra,A., Kerr,D.J. andSeymour,L.W. (1995)Biochem. Biophys. Res. Commun., 217, 721–727.
54. Brown,L.F., Berse,B., Jackman,R.W., Tognazzi,K., Guidi,A.J.,Dvorak,H.F., Senger,D.R., Connolly,J.L. and Schnitt,S.J. (1995)Human Pathol., 26, 86–91.
55. O’Brien,T., Cranston,D., Fuggle,S., Bicknell,R. and Harris,A.L. (1995)Cancer Res., 55, 510–513.
56. Pierce,E.A., Avery,R.L., Foley,E.D., Aiello,L.P. and Smith,L.E.H. (1995Proc. Natl Acad. Sci. USA, 92, 905–909.
57. Smith,L., Wesolowski,E., McLellan,A., Kostyk,S., Amato,R., Sullivan,Rand D’Amore,P. (1994)Invest. Ophthal. Vis. Sci., 35, 101–111.




















