
Genetic polymorphisms of matrix metalloproteinases in lung, breast and colorectal cancer

Clinical Immunology
A Peptide Based on an Anti-DNA Autoantibody Downregulates MatrixMetalloproteinases in Murine Models of Lupus
Anat Faber-Elmann, Eran Eilat, Heidy Zinger, and Edna Mozes1
Systemic lupus erythematosus (SLE) is an autoim-mune disease characterized by the increased produc-tion of autoantibodies and by T cell dysfunction asso-ciated with general clinical manifestations. A model ofinduced experimental SLE by the immunization withthe human monoclonal anti-DNA 16/6Id� autoantibodyand a model of the SLE-prone mice (NZB � NZW)F1were used in the present study. Two peptides based onthe complementarity determining regions (CDR) 1 and3 of a murine monoclonal anti-DNA 16/6Id� autoanti-body were shown to ameliorate spontaneous and in-duced SLE in mice. We demonstrate here that levels ofmatrix metalloproteinase (MMP)-3 and MMP-9 wereelevated in plasma and kidneys of SLE-afflicted mice.Levels of both MMP-3 and MMP-9 were elevated inkidneys of mice with the 16/6Id induced experimentalSLE already in the early phases of disease develop-ment. However, increased levels of only MMP-3 weredetected in the plasma at the early stages of disease,while MMP-9 activity was elevated later, when clinicalmanifestations were already observed. Treatment ofSLE-afflicted mice, with the CDR1-based peptide thatameliorates disease manifestations in mice, led to areduction in MMP-9 activity and in MMP-3 proteinlevels both in plasma and in kidneys. We thus suggestthat these enzymes may play a pathogenic role in thedisease and may serve as markers for the determina-tion of disease progression or amelioration. © 2002
Elsevier Science (USA)Elsevier Science (USA)
Key Words: involvement of MMPs in lupus; immuno-modulation; CDR-based peptide; MMPs in affectedkidneys.
INTRODUCTION
Systemic lupus erythematosus (SLE)2 is an autoim-mune disease characterized by the increased produc-tion of antibodies against several self antigens and
1 To whom correspondence should be addressed. Telephone: �972 89343646. Fax: �972 8 9344141. E-mail: [email protected].
2 Abbreviations used: CDR, complementarity determining region;CFA, complete Freund’s adjuvant; Id, idiotype; IL, interleukin; i.v.,intravenous; mAb, monoclonal antibody; MMP, matrix metallopro-
223
defective T-cell-mediated responses. The latter are as-sociated with various clinical manifestations includingimmune complex depositions in the kidneys and otherorgans (1). (NZB � NZW)F1 female mice that developSLE spontaneously have been used extensively as amodel of lupus nephritis (2). The induction of experi-mental SLE has been previously reported by our labo-ratory, and was achieved by immunization with mono-clonal anti-DNA antibodies that express the majoridiotype (Id), designated 16/6 of either human or mouseorigin (3, 4). The disease can be induced in mice ofdifferent susceptible strains (5). The 16/6Id-induceddisease resembles SLE in human and is manifested byhigh levels of autoantibodies that include anti-DNAand anti-nuclear protein antibodies, as well as 16/6Idand anti-16/6Id-specific antibodies (3). The 16/6Id im-munized mice also develop SLE-associated clinicalsymptoms (e.g., leukopenia, proteinuria, and kidneydamage). Two peptides based on the sequences of thecomplementarity determining regions (CDR) 1 and 3 ofa murine anti-DNA, 16/6Id designated mAb 5G12 (6),were capable of either preventing (7, 8) or treating analready established SLE-like disease (8; Zinger, H.,Eilat, E., Meshorer, A., Mozes, E., unpublished re-sults). The beneficial effects of the peptides were man-ifested in the reduction of autoantibodies and in thedownregulation of clinical symptoms including kidneydamage (7, 8). The amelioration of disease manifesta-tions was associated with the immunomodulation ofcytokine production (8; Zinger, H., Eilat, E., Meshorer,A., Mozes, E., unpublished results).
Matrix metalloproteinases (MMPs) (9–11) constitutea family of zinc containing endoproteinases that playan important role in the remodeling of extracellularmatrix in normal tissues, and also contribute to patho-logical processes. They share structural domains butdiffer in substrate specificity, cellular sources, and in-ducibility. The MMPs are synthesized as zymogen-likelatent precursors and are converted subsequently to anactive form. In terms of protein and domain structure,MMP-9 is the largest and most complex member iden-tified so far. MMP-9 expression is characterized by a
Department of Immunology, The Weizman
Vol. 105, No. 2, November, pp. 223–232, 2002doi:10.1006/clim.2002.5279
nstitute of Science, Rehovot 76100, Israel
n Icomplex regulation with tight control at the levels ofteinase; s.c., subcutaneous; SLE, systemic lupus erythematosus.
1521-6616/02 $35.00© 2002 Elsevier Science (USA)
All rights reserved.

gene transcription and protein secretion by cytokines,chemokines, inflammatory mediators, tissue inhibitorsof metalloproteinases (TIMPs), and proenzyme activa-tion by components of the plasminogen activation sys-tem or other MMPs (12) The involvement of MMPs inautoimmune diseases was demonstrated in multiplesclerosis (13)and experimental autoimmune encepha-lomyelitis (14), rheumatoid arthritis (15), Guillain-Barre syndrome (16), experimental bullous pemphi-goid (17), and experimental autoimmune neuritis (18).In patients with lupus nephritis the plasma levels ofMMP-3 (15, 19, 20) and TIMP-2 were reported to besignificantly higher than those of healthy controls.
The potential importance of the many activities ofMMPs in inflammatory responses has been suggestedby the inhibitory effects of MMP inhibitors in severalanimal models of autoimmune diseases. Specific inhi-bition of MMPs in vivo suppresses edema, pathologictissue proliferation, and damage to specialized tissuestructures in several inflammatory and autoimmunediseases (14, 21, 22).
In the present study we show that levels of MMP-3and MMP-9 are elevated both in plasma and in kidneysof mice afflicted with either the spontaneous or theinduced experimental SLE. We also demonstrate thattreatment of SLE-afflicted mice with the CDR1-basedpeptide, which ameliorates manifestations of lupus inmice, diminished the levels of MMP-3 and MMP-9 inthe plasma and kidneys of the treated mice.
MATERIALS AND METHODS
Mice
Female (NZB � NZW)F1 mice were obtained fromthe Jackson Laboratory (Bar Harbor, ME). Femalemice of the BALB/c inbred strain at the age of 6–8weeks were obtained from the Experimental AnimalUnit, The Weizmann Institute of Science. The studyhas been approved by the Animal Care and Use Com-mittee of the Weizmann Institute of Science.
Anti DNA Monoclonal Antibodies
The human anti-DNA monoclonal antibody (mAb)that expresses the 16/6Id (IgG1/�) (23, 24) was secretedby hybridoma cells that were grown in culture and waspurified using a protein G–Sepharose column (Phar-macia).
Synthetic Peptides
The peptide based on CDR1 of the murine anti-DNAmonoclonal antibody 5G12 was used. The CDR1-basedpeptide (pCDR1; TGYYMQWVKQSPEKSLEWIG; theCDR is underlined) was prepared with an automated
synthesizer (Applied Biosystems, Model 430A, Germany)using the company’s protocols for t -butyloxycarbonyltechnique (25). A peptide synthesized in the reversedorder of pCDR1 (GIWELSKEPSQKVWQMYYGT) wasused as a control peptide for pCDR1.
Induction and Treatment of Experimental SLE
In order to induce experimental SLE, BALB/c micewere immunized with 2 �g of the human mAb 16/6Idand boosted 3 weeks later. For the prevention of exper-imental SLE mice were given pCDR1 or the reversedpCDR1 (control peptide) intravenously (i.v.) or subcu-taneously (s.c.) concomitant with the immunizationand were injected weekly thereafter for 5 weeks. Treat-ment of an established disease had started three and ahalf months following disease induction with the 16/6Id when clinical manifestations were already ob-served. Mice received 10 weekly injections (s.c. or i.v.)of the CDR1-based peptide, or the reversed pCDR1. Inall cases the above peptides were administered in adose of 100 �g/mouse.
Prevention and Treatment of SLE-like Disease in(NZB � NZW)F1 Mice with the CDR1-Based Peptide
For prevention of SLE, mice at the age of 2 months,before disease manifestations are observed, were in-jected s.c. with pCDR1 (250 �g/mouse) once a week for10 weeks. To treat an established disease, mice at theage of 5–7 months were injected with pCDR1 (250�g/mouse) s.c., once a week for 10 weeks.
Measurement of Activities of MMP-2 and MMP-9
MMP activity was tested by gelatin zymography,which measures the activity of gelatinases MMP-2 andMMP-9. Pooled plasma samples of 8–10 individualmice per group of the different in vivo experimentswere analyzed. Samples were separated by an 8%SDS–PAGE polymerized with 1 mg/ml gelatin. Follow-ing electrophoresis, gels were washed once for 30 minin 2.5% Triton X-100 to remove the SDS, and once for30 min in the reaction buffer containing 50 mM Tris–HCl, 200 mM NaCl, 10 mM CaCl2, and 0.02% (w/v) Brij35 (pH 7.5). The reaction buffer was changed to a freshone, and the gels were incubated at 37°C for 24 h.Gelatinolytic activity was visualized by staining thegels with 0.5% Coomassie brilliant blue.
Western Blot Analysis of MMP-3 in Plasma
Samples of 5 �l of each serum were loaded on 12%SDS–PAGE, separated under reducing conditions, andtransferred to nitrocellulose. The blots were probed
224 FABER-ELMANN ET AL.

(0.5 �g/ml, 1 h, room temperature) with anti-MMP-3antibodies (Oncogene research products, MA; CatalogNo. IM 36L, crossreactive with mouse) and developedusing chemiluminescence. Intensity of the bands wasperformed with a Densitometer Kodak Image Station440CF with the Kodak Digital Science 1D computerprogram.
Immunostaining of Kidney Sections for MMP-3or MMP-9
For immunostaining of MMP-3 or MMP-9, kidneysections (5 �M) were fixed with cold acetone (5 min atroom temperature), washed twice in PBS, permeabil-ized (1 min at room temperature) in 0.05% TritonX-100 (diluted in PBS), and washed twice in PBS. Inorder to get a specific anti-MMP staining and to avoidthe staining of immune complex deposits by the FITC-labeled goat anti-mouse, kidney sections were blocked(1 h at room temperature) with unlabeled goat anti-mouse IgG�IgM (diluted 1:1 in 1% BSA/PBS; JacksonImmunoResearch Laboratories) and washed threetimes with PBS containing 0.05% Tween. Monoclonalanti-MMP-9 (1:100; Chemicon International, Inc.; Cat-alog No. MAB13415, crossreactive with mouse) or anti-
MMP-3 (1:50; Oncogene research products) antibodydiluted in 1% BSA/PBS were added for 30 min, at roomtemperature. For all immunostaining proceduresFITC-labeled goat anti-mouse IgG�IgM (Jackson Im-munoResearch Laboratories), diluted 1:30 in 1% BSA/PBS (30 min at room temperature) was used.
Statistical Analysis
To evaluate the significance of the difference be-tween untreated and treated groups, the nonparamet-ric Mann–Whitney test was used. Values of P � 0.05were considered significant.
RESULTS
Kinetics of Appearance of MMP-3 and MMP-9 inPlasma of (NZB � NZW)F1 Mice
We first examined if the development of spontaneousSLE in (NZB � NZW)F1 mice is associated withchanges in the levels of MMP-3 and MMP-9 in theirplasma. We thus followed the levels of the latter start-ing at the age of 2 months, before disease manifesta-tions are observed, until the age of 8 months when themice suffer from a full-blown disease. The results areshown in Fig. 1. As can be seen in Fig. 1A, the 50-kDaform of MMP-3 which may represent the MMP-3 zy-mogen is mildly elevated with time (37 � 105 at the ageof 2 months as compared to 66 � 105 densitometricunits at the age of 8 months). Both the 34- and the40-kDa forms of MMP-3 in plasma of 2-months-oldmice are very low (undetectable by densitometry) andare gradually elevated toward the age of 8 months(12 � 105,densitometric units for both time points
FIG. 1. Kinetics of appearance of MMP-3 and MMP-9 in theplasma of (NZB � NZW)F1 mice. (NZB � NZW)F1 mice (10 mice/group in each experiment) were bled at the indicated time points.Pooled plasma samples (4 �l) of mice from each group were tested forMMP-3 expression levels using Western blot (A) or for MMP-9 ac-tivity using gel zymography (B). The results represent four indepen-dent in vivo experiments.
FIG. 2. Kinetics of appearance of MMP-3 and MMP-9 in theplasma of BALB/c mice. Nonimmunized BALB/c mice (10 mice/group) or mice that were immunized with the 16/6Id (in CFA; 10mice/group) were bled at the indicated time points. Pooled plasmasamples of mice (4 �l) from each group were tested for MMP-3expression levels using Western blot (A) or for MMP-9 activity usinggel zymography (B). The results represent three independent in vivoexperiments.
225DOWNREGULATION OF MMPs BY A CDR-BASED PEPTIDE

tested). The high-molecular-weight bands (which mayrepresent the complex of MMP-3 with other proteins)are also elevated with time by 6- to 12-fold. Similarly,Fig. 1B shows that MMP-9 activity, assessed by gelzymography, is low at the age of 2 months and isgradually elevated in plasma of mice with the progres-sion of disease until the age of 8 months.
Kinetics of Appearance of MMP-3 and MMP-9in Plasma a of BALB/c mice that WereImmunized with 16/6Id
Previous results from our laboratory have shownthat experimental SLE can be induced in BALB/c micefollowing immunization with the 16/6Id (3, 4). There-fore, it was of interest to test if the induced model ofSLE resembles the (NZB � NZW)F1 model with re-spect to the changes in MMP-3 and MMP-9. We thuslooked at levels of MMP-3 and MMP-9 in this experi-mental model of SLE. The results demonstrated in Fig.2 indicate that the levels of MMP-3 (all isoforms) wereelevated 10 days after the boost (4.5 weeks postimmuni-
zation), and were higher than in control unimmunizedBALB/c mice of the same age (Fig. 2A). The levels ofMMP-3 increased with aging in all groups; however, thelatter were always higher in the 16/6Id-immunized micethan in the control group. In contrast, no inducedchanges in MMP-9 activity could be detected during thefirst few months after the boost. As can be seen in Fig. 2B,higher activity of MMP-9 in the plasma of 16/6Id-immu-nized mice than in plasma of nonimmunized mice couldbe detected at 3.5 months after the boost.
Specific Upregulation of MMP-3 and MMP-9 in KidneySections of BALB/c Mice that Were Immunizedwith the 16/6Id
Since immunization of BALB/c mice with the 16/6Idleads to clinical manifestations characteristic of SLEincluding kidney damage (3, 4) and because kidneydamage in (NZB � NZW)F1 mice was reported to beassociated with elevation in the levels of MMP-3 andMMP-9 (26), we looked at the expression of these en-zymes in kidneys of BALB/c mice that were immunized
FIG. 3. Immunostaining of kidney sections of immunized BALB/c mice for MMP-3 and MMP-9. Nonimmunized BALB/c mice or mice thatwere immunized with PBS/CFA or with the 16/6Id (in CFA) (3 mice/group) were sacrificed 5.5 months following their boost with the 16/6Id.Kidneys were removed, and their 5-�m cryostat sections were immunostained for MMP-3 (A) and MMP-9 (B). A control staining to theefficiency of blocking was performed (see Materials and Methods) (C). Magnification, �200. The results represent three independent in vivoexperiments.
226 FABER-ELMANN ET AL.
with 16/6Id. As controls we used mice that were immu-nized with CFA and age-matched unimmunized mice.Both control groups were injected with PBS at the timeof the boost. Two months and 5 months after the boostwith 16/6Id, kidney sections of mice were immuno-stained for the presence of MMP-3 or MMP-9. Figure 3represents immunohistology of kidneys taken 5months after the boost. As can be seen in the figure,immunization of mice with CFA upregulated the ex-pression of both MMP-3 (Fig. 3A) and MMP-9 (Fig. 3B)in the kidneys, in the glomeruli and in the surroundingtissue. Nevertheless, immunization with 16/6Id (inCFA) further elevated significantly the expression lev-els of these two MMPs. This elevation was observedalready 2 months after the boost with the 16/6Id (datanot shown). The nonspecific background levels of thestaining are also demonstrated (Fig. 3C).
The CDR1-Based Peptide Downregulates Levelsof MMP-3 and Activity of MMP-9 in Plasmaof (NZB � NZW)F1 Mice
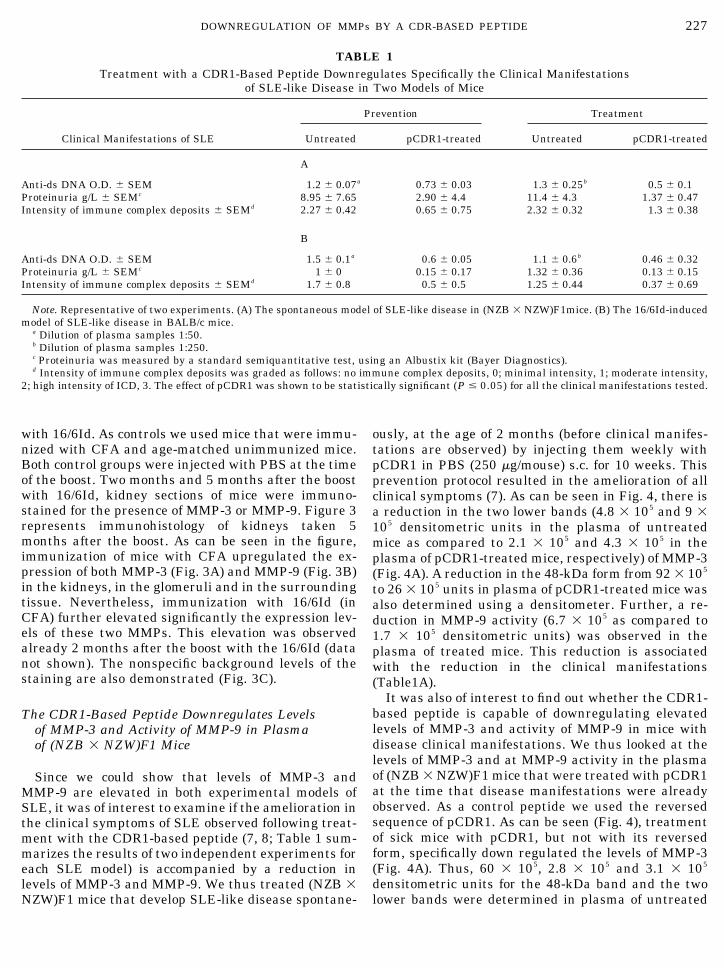
Since we could show that levels of MMP-3 andMMP-9 are elevated in both experimental models ofSLE, it was of interest to examine if the amelioration inthe clinical symptoms of SLE observed following treat-ment with the CDR1-based peptide (7, 8; Table 1 sum-marizes the results of two independent experiments foreach SLE model) is accompanied by a reduction inlevels of MMP-3 and MMP-9. We thus treated (NZB �NZW)F1 mice that develop SLE-like disease spontane-
ously, at the age of 2 months (before clinical manifes-tations are observed) by injecting them weekly withpCDR1 in PBS (250 �g/mouse) s.c. for 10 weeks. Thisprevention protocol resulted in the amelioration of allclinical symptoms (7). As can be seen in Fig. 4, there isa reduction in the two lower bands (4.8 � 105 and 9 �105 densitometric units in the plasma of untreatedmice as compared to 2.1 � 105 and 4.3 � 105 in theplasma of pCDR1-treated mice, respectively) of MMP-3(Fig. 4A). A reduction in the 48-kDa form from 92 � 105
to 26 � 105 units in plasma of pCDR1-treated mice wasalso determined using a densitometer. Further, a re-duction in MMP-9 activity (6.7 � 105 as compared to1.7 � 105 densitometric units) was observed in theplasma of treated mice. This reduction is associatedwith the reduction in the clinical manifestations(Table1A).
It was also of interest to find out whether the CDR1-based peptide is capable of downregulating elevatedlevels of MMP-3 and activity of MMP-9 in mice withdisease clinical manifestations. We thus looked at thelevels of MMP-3 and at MMP-9 activity in the plasmaof (NZB � NZW)F1 mice that were treated with pCDR1at the time that disease manifestations were alreadyobserved. As a control peptide we used the reversedsequence of pCDR1. As can be seen (Fig. 4), treatmentof sick mice with pCDR1, but not with its reversedform, specifically down regulated the levels of MMP-3(Fig. 4A). Thus, 60 � 105, 2.8 � 105 and 3.1 � 105
densitometric units for the 48-kDa band and the twolower bands were determined in plasma of untreated
TABLE 1Treatment with a CDR1-Based Peptide Downregulates Specifically the Clinical Manifestations
of SLE-like Disease in Two Models of Mice
Clinical Manifestations of SLE
Prevention Treatment
Untreated pCDR1-treated Untreated pCDR1-treated
A
Anti-ds DNA O.D. � SEM 1.2 � 0.07a 0.73 � 0.03 1.3 � 0.25b 0.5 � 0.1Proteinuria g/L � SEMc 8.95 � 7.65 2.90 � 4.4 11.4 � 4.3 1.37 � 0.47Intensity of immune complex deposits � SEMd 2.27 � 0.42 0.65 � 0.75 2.32 � 0.32 1.3 � 0.38
B
Anti-ds DNA O.D. � SEM 1.5 � 0.1a 0.6 � 0.05 1.1 � 0.6b 0.46 � 0.32Proteinuria g/L � SEMc 1 � 0 0.15 � 0.17 1.32 � 0.36 0.13 � 0.15Intensity of immune complex deposits � SEMd 1.7 � 0.8 0.5 � 0.5 1.25 � 0.44 0.37 � 0.69
Note. Representative of two experiments. (A) The spontaneous model of SLE-like disease in (NZB � NZW)F1mice. (B) The 16/6Id-inducedmodel of SLE-like disease in BALB/c mice.
a Dilution of plasma samples 1:50.b Dilution of plasma samples 1:250.c Proteinuria was measured by a standard semiquantitative test, using an Albustix kit (Bayer Diagnostics).d Intensity of immune complex deposits was graded as follows: no immune complex deposits, 0; minimal intensity, 1; moderate intensity,
2; high intensity of ICD, 3. The effect of pCDR1 was shown to be statistically significant (P � 0.05) for all the clinical manifestations tested.
227DOWNREGULATION OF MMPs BY A CDR-BASED PEPTIDE

mice as compared to 27 � 105, 0.4 � 105 and 0.9 � 105
for pCDR1-treated and 31 � 105,5.6 � 105 and 9.6 �105 for the reversed form of pCDR1-treated mice. Theactivity of MMP-9 (Fig. 4B) in the plasma was affectedsimilarly. Thus, there were 6.4 � 105 densitometricunits in the plasma of untreated mice and 8.8 � 105 inthe plasma of the control reversed pCDR1 as comparedto 1.2 � 105 densitometric units measured in theplasma of pCDR1-treated mice.
The CDR1-Based Peptide Downregulates Levels ofMMP-3 and Activity of MMP-9 in Plasmaof 16/6Id-Immunized Mice
As we have demonstrated, activity of MMP-9 is ele-vated in both experimental models [(NZB � NZW)F1and 16/6Id-induced] of SLE. Because the CDR1-basedpeptide was shown to ameliorate the disease in bothmodels, and to downregulate MMP-3 levels andMMP-9 activity in plasma of treated (NZB � NZW)F1mice, we further looked for the effect of treatment onthese enzymes in plasma of 16/6Id-immunized BALB/cmice. Figure 5 represents results of a prevention and ofa treatment experiment. The figure shows that pCDR1could downregulate MMP-3 and MMP-9 in this exper-
imental model as well. Downregulation of levels ofMMP-3 by two- to fourfold (measured by a densitome-ter) was observed for both the prevention and the treat-ment protocols. Activity of MMP-9 was also reduced bytreatment with pCDR1. Thus, for the prevention pro-tocol there were 4.6 � 105 densitometric units (4.8 �105 units were determined for normal mice) as com-pared to 8.1 � 105 densitometric units in the plasma ofuntreated mice. In plasma samples taken from mice ofthe treatment protocol (Fig. 5B), densitometric unitsmeasured for MMP-9 activity were 3.9 � 105, 5 � 105,and 1.6 � 105 for normal nonimmunized, 16/6Id-immu-nized and 16/6Id-immunized and pCDR1-treated mice,respectively. Thus, in vivo administration of pCDR1,either at the stage of disease induction or at the stageof a full-blown disease, could reduce the levels ofMMP-3 (Fig. 5A) and MMP-9 (Fig. 5B).
Effect of Treatment with the CDR1-Based Peptideon the Levels of MMP-3 and MMP-9 in KidneySections of 16/6Id-Immunized BALB/c Mice
Because MMP-3 and MMP-9 were shown to be ele-vated in kidneys of mice with induced experimentalSLE it was of interest to look for the effect of treatmentwith the CDR1-based peptide on the expression of thelatter MMPs in the kidneys of the treated mice. Table2 summarizes the results of immunostaining with anti-
FIG. 4. Levels of MMP-3 and MMP-9 in plasma of (NZB �NZW)F1 mice treated with pCDR1. In prevention experiments, mice(10/group) were given weekly s.c. injections with pCDR1 starting atthe age of 2 months during 10 weeks as described under Materialsand Methods. The results represent plasma samples taken 4 monthsafter the end of the treatment. In treatment experiments, mice(10/group) were injected s.c. with either PBS or 250 �g/mouse ofpCDR1 starting at the age of 5 months. The results represent plasmasamples taken 3 weeks after the end of the treatment. Pooled plasmasamples of each experimental group were tested for MMP-3 levels byWestern blot analysis and for MMP-9 activity using gel zymography.The results represent two similar experiments.
FIG. 5. Levels of MMP-3 and MMP-9 in 16/6Id-immunizedBALB/c mice treated with pCDR1. In prevention experiments, mice(8/group) were treated with pCDR1 as described under Materialsand Methods. The results shown are of plasma samples taken 4.5months after the end of the treatment. In treatment experiments,mice (8/group) were treated with pCDR1 (see Materials and Meth-ods.). The results are of plasma samples obtained at the end oftreatment. Pools of plasma samples of each experimental group weretested for MMP-3 by Western blot analysis (A) and for MMP-9activity by gel zymography (B). The results are representative of twosimilar experiments.
228 FABER-ELMANN ET AL.

MMP-3 and anti-MMP-9. The results demonstrate thatin both prevention and treatment experiments, pCDR1downregulated the expression levels of both MMP-3and MMP-9 in the kidneys of the treated mice. Figure6 represents immunostaining of sections of mice of bothprevention (Fig. 6A) and treatment (Fig. 6B) experi-ments. The reduction in the levels of MMP was ob-served in the glomeruli and interstitium. The observedreduction in MMP expression correlated with the re-duction in staining for immune complex deposits usinganti-Ig (Fig. 6B).
DISCUSSION
The present report shows that both MMP-3 andMMP-9 are elevated in plasma and in kidneys of mu-rine models of SLE. The CDR1-based peptide coulddownregulate the levels of MMP-9 and MMP-3 in bothplasma and kidneys of SLE-afflicted mice. We demon-strate that in the spontaneous model of SLE, upregu-lation of both MMP-9 and MMP-3 in the plasma occursat the first 3.5 months. In the induced model the ele-vation in the levels of MMP-3 in plasma is very early(10 days after the boost with the 16/6Id), while theelevation of MMP-9 activity occurs 3–4 months later.The results with the 16/6Id-induced model that allowsus to follow processes at the first stages of the diseasemay suggest that MMP-3 is upregulated at earlystages of experimental SLE while MMP-9 is elevated ina later stage. Different roles for MMP-3 and MMP-9were also suggested by Wang et al. (27), who reportedthat MMP-3 is required for initiation of contact hyper-sensitivity, whereas MMP-9 plays a critical role in itsresolution. Since MMP-3, in addition to plasmin (28),can activate pro MMP-9 (29), its early induction maycontribute to the elevation in levels of MMP-9 in theplasma and in kidneys.
The origin of the elevated MMPs in plasma and inkidneys is not known. An enhanced expression of MMP-9
by glomerular epithelial cells (30) or by mesangial cellshas been demonstrated in a model of immune complex-mediated glomerulonephritis (31). In polymorphonucle-ar-dependent glomerulonephritis proteolysis of the glo-merular basement membrane that cause glomerularinjury was shown to be mediated by neutrophil-derivedproteinases (32). Another source of the enhanced MMP-9might be mononuclear cells that infiltrate the kidneys of(NZB � NZW)F1 mice as the disease progresses.
Our results showing the elevation of MMP-3 in themouse models of SLE are in agreement with resultsdemonstrating that MMP-3 is significantly increasedin plasma of patients with SLE (33) and that MMP-3transcript is significantly increased with the progres-sion of nephritis in (NZB � NZW)F1 mice (26). Takentogether, these results suggest that MMP-3 may con-tribute to the development of glomerular injury in lu-pus nephritis.
Our results show that activity levels of MMP-2 inplasma were not significantly elevated with diseaseprogression in both the induced and the spontaneousmodels of SLE (data not shown). These results arecompatible with those reported by Zucker (20) thatMMP-2 levels were not increased in SLE. Accordingly,this enzyme was not modulated by treatment with theCDR1-based peptide (data not shown). Levels ofMMP-2 were secreted constitutively and did notchange in other pathological conditions (like optic neu-ritis and multiple sclerosis) as well, whereas levels ofMMP-9 were elevated relative to the healthy controls(34, 35).
Interestingly, the CDR1-based peptide that specifi-cally and directly affects SLE-associated T-cell re-sponses could downregulate levels of MMP-3 andMMP-9 that are already upregulated (treatment pro-tocol) as well as prevent their elevation (preventionprotocol). The latter might suggest that MMP levels inplasma and in kidney are controlled either directly or
TABLE 2The Immunostaining with Anti-MMP-3 and Anti-MMP-9 of Kidney Sections of BALB/c Mice
BALB/c mice
Preventiona Treatmenta
MMP-3 MMP-9 MMP3 MMP-9
16/6Id immunized 1.65 � 0.22 2.89 � 0.11 2.9 � 0.18 2.17 � 0.3016/6Id immunized and pCDR1 treated 0.45 � 0.12 0.71 � 0.21 0.25 � 0.09 0.21 � 0.10
P � 0.0001 b P � 0.0001 P � 0.0002 b P � 0.0001P � 0.395 c P � 0.5 P � 0.245 c P � 0.085
Normal 0.5 � 0.14 0.71 � 0.14 0.16 � 0.16 0.71 � 0.28
Note. Representative of 2–3 experiments.a The intensity of MMP-3- and MMP-9-specific staining was graded as follows: no staining, 0; traces, 0.5; weak staining, 1; moderate
staining, 2; intensive staining, 3. P values were calculated by the Mann–Whitney test.b P values as compared with the 16/6Id-immunized group.c P values as compared with normal BALB/c mice.
229DOWNREGULATION OF MMPs BY A CDR-BASED PEPTIDE

FIG. 6. Immunostaining for MMP-3 and MMP-9 of kidney sections of BALB/c mice that were treated with pCDR1 for prevention (A) ortreatment (B) of experimental SLE. Mice were sacrificed 8 months following disease induction, and their kidneys were removed. Cryostatsections (5 �m) were prepared and immunostained for MMP-3, MMP-9, and the presence of immune complex deposits (Magnification, �200).W/O, control staining to the efficiency of blocking, without the first antibody (see Materials and Methods). The results represent two similarexperiments.
230 FABER-ELMANN ET AL.

indirectly (e.g., via shift in cytokines) by T cells.MMP-9 has been shown to be secreted by T cells, neu-trophils, and macrophages, and its secretion is affectedby the interactions between the latter (12). For exam-ple, mononuclear phagocytes, which play a role in mod-ulating the turnover of extracellular matrix, produceMMP when stimulated with activated T cells (36). Fur-ther, cytokines of both Th1- and Th2-type cells canmodulate MMP secretion by macrophages (37).
TNF-� and IL-1 were shown to play an importantrole in the pathogenesis of SLE both in the humandisease (38) and in murine models (2, 8, 39). It has beenshown in several systems that the above cytokinesinduce MMP-9 (12, 40) and MMP-3 (41), and thus it ispossible that the induction of the latter MMPs is partof the pathogenic effect of these cytokines in SLE.Since the CDR1-based peptide was shown to down-regulate the levels of TNF-� (8), the latter may ac-count, at least in part, for the inhibitory effect of CDR1on MMP-3 and MMP-9 levels in the experimental mod-els of SLE in the present study.
MMP-9 activity in plasma was neither studied inSLE patients nor in animal models of SLE. In thepresent report we demonstrate, for the first time, cor-relation between SLE and MMP-9. This report demon-strates that MMP-9 is elevated in plasma and kidneysof the SLE-afflicted mice. Although the elevation inplasma appears relatively late (around 4 months afterthe boost), the elevation of MMP-9 in the kidneys isobserved at early stages after disease induction (2months, data not shown). We have further shown thata peptide that ameliorates SLE manifestations (7, 8,Table 1) in mice (pCDR1), also downregulates MMP-9both in plasma and in kidneys. The latter data werealso supported by preliminary results, using theMMP-9 activity assay system, that showed that immu-nization with the 16/6Id elevated MMP-9 activity,whereas treatment with the CDR1-based peptidedownregulated it to the activity levels of unimmunizedmice. A causal relationship was also suggested be-tween expression of MMP-9 by glomerular epithelialcells and changes in glomerular capillary permeabilityand proteinuria in Heyman nephritis (30).
We also demonstrate here for the first time a differ-ent kinetic of induction of MMP-3 and MMP-9 in ex-perimental models of autoimmune diseases as shownhere specifically for SLE. This may indicate the differ-ent roles of the above MMPs in the pathogenesis of thedisease. Our results demonstrate that a peptide thatimmunomodulates SLE-associated T-cell responsesand downregulates disease manifestations can controlthe secretion (although not necessarily by the T cellsthemselves) of those MMPs in plasma and kidneys. Wethus suggest that MMP-3 and MMP-9 play a role in thepathogenesis of SLE and may serve as surrogate mark-
ers of disease progression on one hand, and of diseaseamelioration by a given treatment on the other hand.
ACKNOWLEDGMENT
This research was supported by Teva Pharmaceutical Industries,Netanya, Israel.
REFERENCES
1. Hahn, B. H., An overview of the pathogenesis of systemic lupuserythematosus. In “Dubois’ Lupus Erythematosus” (D. J. Wal-lace, and B. H. Hahn, Eds.),. 5th ed. Williams and Wilkins;Philadelphia,1993.
2. Theofilopoulos, A. N., and Lawson, B. R., Tumor necrosis factorand other cytokines in murine lupus. Ann. Rheum. Dis.58(Suppl.), 149–155, 1999.
3. Mendlovic, S., Brocke, S., Shoenfeld, Y., Ben-Bassat, M., Me-shorer, A., Bakimer, R., and Mozes, E., Induction of a SLE-likedisease in mice by a common human anti-DNA idiotype. Proc.Natl. Acad. Sci. USA 85, 2260–2264, 1988.
4. Waisman, A., Mendelovic, S., Ruiz, J. P., Zinger, H., Meshorer,A., and Mozes, E., The role of the 16/6 idiotype network in theinduction and manifestation of systemic lupus erythematosus.Int. Immunol. 5,1293–1300, 1993.
5. Mendlovic, S., Brocke, S., Fricke, H., Shoenfeld, Y., Bakimer, R.,and Mozes, E., The genetic regulation of the induction of exper-imental SLE. Immunology 69, 228–236, 1990.
6. Waisman, A., Ruiz, P. J., Israeli, E., Eilat, E., Konen-Waisman,S., Zinger, H., Dayan, M., and Mozes, E., Modulation of murinesystemic lupus erythematosus with peptides based on comple-mentary determining regions of a pathogenic anti-DNA mono-clonal antibody. Proc. Natl. Acad. Sci. USA 94, 4620–4625,1997.
7. Eilat, E., Zinger, H., Nyska, A., and Mozes, E., Prevention ofsystemic lupus erythematosus-like disease in (NZBxNZW)F1mice by treating with CDR1- and CDR3-based peptides of apathogenic autoantibody. J. Clin. Immunol. 20, 268–278, 2000.
8. Eilat, E., Dayan, M., Zinger, H., and Mozes, E., The mechanismby which a peptide based on the complementarity determiningregion-1 of a pathogenic anti-DNA autoantibody ameliorates ex-perimental SLE. Proc. Natl. Acad. Sci. USA 98, 1148–1153,2001.
9. Shingleton, W. D. Hodges, D. J., Brick, P., and Cawston, T. E.,Collagenases: A key enzyme in collagen turnover. Biochem. Cell.Biol. 74, 759–775, 1996.
10. Goetzl, E. J., Banda, M. J., and Leppert, D., Matrix metallopro-teinases in immunity. J. Immunol. 156, 1–4, 1996.
11. Massova, I., Kotra, L. P., Fridman, R., and Mobashery, S., Ma-trix metalloproteinases: structures, evolution and diversifica-tion. FASEB. J. 12, 1075–1095, 1998.
12. Guedez, L., Lim, M. S., and Stetler-Stevenson, W. G., The role ofmetalloproteinases and their inhibitors in hematological disor-ders. Crit. Rev. Oncog. 7, 205–225, 1996.
13. Ozenci, V., Rinaldi, L., Teleshova, N., Matusevicius, D., Kivisak,P., Kouwenhoven, M., and Link, H., Metalloproteinases andtheir tissue inhibitors in multiple sclerosis. J. Autoimmun. 12,297–303, 1999.
14. Gijbels, K., Galardy, R. E., and Steinman, L., Reversal of exper-imental autoimmune encephalomyelitis with a hydroxamate in-hibitor of matrix metalloproteinases. J. Clin. Invest. 94, 2177–2189, 1994.
15. Keyszer, G., Lambiri, I., Nagel, R., Keysser, C., Keysser, M.,Gromnica-Ihle, E., Franz, J., Burmester, G. R., and Jung, K.,
231DOWNREGULATION OF MMPs BY A CDR-BASED PEPTIDE

Circulating levels of matrix metalloproteinases MMP-3 andMMP-1, tissue inhibitor of metalloproteinases 1 (TIMP-1), andMMP-1/TIMP-1 complex in rheumatic disease. Correlation withclinical activity of rheumatoid arthritis versus other surrogatemarkers. J. Rheumatol. 26, 251–258, 1999.
16. Creange, A., Sharshar,T., Planchenault, T., Christov, C., Poron,F., Raphael, J-C., and Gherardi, R. K., Matrix metalloprotein-ase-9 is increased and correlates with severity in Guillain-Barresyndrome. Neurology 53, 1683–1691, 1999.
17. Liu, Z., Shipley, J. M., Vu, T. H., Zhou, X., Diaz, L. A., Werb, Z.,and Senior, R. M., Gelatinase B-deficient mice are resistant toexperimental bullous Pemphigoid. J. Exp. Med. 188, 475–482,1998.
18. Hughes, P. M., Wells, G. M. A., Clements, J. M., Gearing, A. J.,Redford, E. J., Davies, M., Smith, K. J., Hughes, R. A., Brown,M. C., and Miller, K. M., Matrix metalloproteinase expressionduring experimental neuritis. Brain 121, 481–494, 1998.
19. Ichikawa, Y., Yamada, C., Horiki, T., Hoshina, Y., and Uch-iyama, M., Serum matrix metalloproteinase-3 and fibrin degra-dation product levels correlate with clinical disease activity inrheumatoid arthritis. Clin. Exp. Rheumatol. 16, 533–540, 1998.
20. Zucker, S., Increased serum stromelysin-1 levels in systemiclupus erythematosus: Lack of correlation with disease activity.J. Rheumatol. 26, 78–80, 1999.
21. Wallace, G. R., Whiston, R. A., Stanford, M. R., Wells, G. M. A.,Gearing, A. J. H., and Clements, J. M., The matrix metallopro-teinase inhibitor BB-1101 prevents experimental autoimmuneuveoretinitis (EAU). Clin. Exp. Immunol. 118, 364–370, 1999.
22. Conway, J. G., Wakefield, J. A., Brown, R. H., Marron, B. E.,Sekut, L., Clark, R. L., McGeechan, G. M., and Connolly, K. M.,Inhibition of cartilage and bone destruction in adjuvant arthritisin the rat by a matrix metalloproteinase inhibitor. J. Exp. Med.182, 449–457, 1995.
23. Waisman, A., Shoenfeld, Y., Blank, M., Ruiz, P. J., and Mozes,E., The pathogenic human monoclonal anti-DNA that inducesexperimental systemic lupus erythematosus in mice is encodedby a VH4 gene segment. Int. Immunol. 7, 689–696, 1995.
24. Shoenfeld, Y., Hsu-Lin, S. C., Gabriels, J. E., Silberstein, L. E.,Furie, B. C., Furie, B., Stollar, B. D., and Schwartz, R. S.,Production of autoantibodies by human-human hybridomas.J. Clin. Invest. 7, 205–208, 1982.
25. Schnolzer, M., Alewood, P. F., and Kent, S. B. H., In situ neu-tralization in Boc-chemistry solid phase peptide synthesis.Rapid, high yield assembly of difficult sequences. Int. J. Pept.Protein Res. 40, 180–193, 1992.
26. Nakamura, T., Ebihara, I., Osada, S., Takahashi, T., Yamamoto,M., Tomino, Y., and Koide, H., Gene expression of metallopro-teinases and their inhibitor in renal tissue of New ZealandBlack/White F1 mice. Clin. Sci. 85, 295–301, 1993.
27. Wang, M., Qin, X., Mudgett, J. S., Ferguson, T. A., Senior, R. M.,and Welgus, H. G., Matrix metalloproteinase deficiencies affectcontact hypersensitivity: Stromelysin-1 deficiency prevents theresponse and gelatinase B deficiency prolongs the response.Proc. Natl. Acad. Sci. USA 96, 6885–6889, 1999.
28. Lijnen, H. R., Silence, J., Van Hoef, B., and Collen, D., Strome-lysin-1 (MMP-3)-independent gelatinase expression and activa-tion in mice. Blood 91; 2045–2053, 1998.
29. Ogata, Y., Enghild, J. J., and Nagase, H., Matrix metalloprotein-ase 3 (stromelysin) activates the precursor for the human matrixmetalloproteinase 9. J. Biol. Chem. 267, 3581–3584, 1992.
30. McMillan, J. I., Riordan, J. W., Couser, W. G., Polock, A. S., andLovett, D., Characterization of a glomerular epithelial cell met-alloproteinase as matrix metalloproteinase-9 with enhanced ex-pression in a model of membranous nephropathy. J. Clin. Invest.97, 1094–1101, 1996.
31. Lovett, D. H., Johnson, R. J., Marti, H. P., Martin, J., Davis, M.,and Couser, W. G., Structural characterization of the mesangialcell type IV collagenase and enhanced expression in a model ofimmune complex mediated glomerulonephritis. Am. J. Pathol.141, 85–98, 1992.
32. Hawkins, D., and Cochrane, C. G., Glomerular basement mem-brane in immunologic glomerulonephritis. Immunology 14, 665–681, 1968.
33. Kotajima, L., Aotsuka, S., Fujimani, M., Okawa-Takatsuji, M.,Kinoshita, M., Sumiya, M., and Obata, K., Increased levels ofmatrix metalloproteinase-3 in sera from patients with activelupus nephritis. Clin. Exp. Rheum. 16, 409–415, 1998.
34. Gijbels, K., Masure, S., Carton, H., and Opdenakker, G., Gela-tinase in the cerebrospinal fluid of patients with multiple scle-rosis and other inflammatory neurological disorders. J. Neuro-immunol. 41, 29–34, 1992.
35. Paemen, L., Olsson, T., Soderstrom, M., Van Damme, J., andOpdenakker, G., Evaluation of gelatinases and IL-6 in the cere-brospinal fluid of patients with optic neuritis, multiple sclerosisand other inflammatory neurological disease. Eur. J. Neurol.. 1,55–63, 1994.
36. Miltenburg, A. M., Lacraz, S., Welgus, H. G., and Dayer, J. M.,Immobilized anti-CD3 antibody activates T cell clones to inducethe production of interstitial collagenase, but not tissue inhibitorof metalloproteinases, in monocytic THP-1 cells and dermal fi-broblasts. J. Immunol. 154, 2655–2667, 1995.
37. Chizzolini, C., Rezzonico, R., De Luca, C., Burger, D., and Dayer,J-M., Th2 cell membrane factors in association with IL-4 en-hance matrix metalloproteinase-1 (MMP-1) while decreasingMMP-9 production by granulocyte-macrophage colony-stimulat-ing factor-differentiated human monocytes. J. Immunol. 164,5952–5960, 2000.
38. Dean, G. S., Tyrrell-Price, J., Crawley, E., and Isenberg, D. A.,Cytokines and systemic lupus erythematosus. Ann. Rheum. Dis.59, 243–251, 2000.
39. Segal, R., Bermas, B. L., Dayan, M., Kalush, F., Shearer, G. M.,and Mozes, E., Kinetics of cytokine production in experimentalsystemic lupus erythematosus: Involvement of T helper cell 1/Thelper cell 2-type cytokines in disease. J. Immunol. 158, 3009–3016, 1997.
40. Saren, P., Welgus, H. G., and Kovanen, P. T., TNF-� and IL-1�selectively induce expression of 92-kDa gelatinase by humanmacrophages. J. Immunol. 157, 4159–4165, 1996.
41. MacNaul, K. L., Chartrain, N., Lark, M., Tocci, M. J., andHutchinson, N. I., Discoordinate expression of stromelysin, col-lagenase, and tissue inhibitor of metalloproteinase-1 in rheuma-toid synovial fibroblasts. Synergistic effects of interleukin-1 andtumor necrosis factor-alpha on stromelysin expression. J. Biol.Chem. 265, 17238–17245, 1990.
Received April 29, 2002; accepted with revision July 24, 2002
232 FABER-ELMANN ET AL.
Copyright © 2022 FDOKUMEN




















