
Trypanosoma cruzi surface molecule gp90 downregulates invasion of gastric mucosal epithelium in...
9
Original article Trypanosoma cruzi surface molecule gp90 downregulates invasion of gastric mucosal epithelium in orally infected mice Mauro Cortez a , Marcelo R. Silva a , Ivan Neira a , Daniele Ferreira a , Gisela R.S. Sasso b , Alejandro O. Luquetti c , Anis Rassi c , Nobuko Yoshida a, * a Departamento de Microbiologia, Imunologia e Parasitologia, Escola Paulista de Medicina, Universidade Federal de São Paulo, R. Botucatu, 862- 6° andar, 04023-062 São Paulo, SP, Brazil b Departamento de Morfologia, Universidade Federal de São Paulo, São Paulo, Brazil c Laboratório de Doença de Chagas, Hospital das Clínicas, Faculdade de Medicina, Universidade Federal de Goiás, Brazil Received 10 January 2005; received in revised form 6 May 2005; accepted 23 May 2005 Available online 02 August 2005 Abstract Experiments were performed to elucidate why Trypanosoma cruzi isolates 573 and 587 differ widely in their efficiency to infect gastric mucosal epithelium when administered orally to mice. These isolates have the same surface profile and a similar capacity to enter host cells in vitro. Metacyclic forms of isolates 573 and 587 and the control CL isolate expressed similar levels of gp82, which is a cell invasion-promoting molecule. Expression of gp90, a molecule that downregulates cell invasion, was lower in the CL isolate. Consistent with this profile, ~three- fold fewer parasites of isolates 573 and 587 entered epithelial HeLa cells, as compared to the CL isolate. No difference in the rate of intrac- ellular parasite replication was observed between isolates. When given orally to mice, metacyclic forms of isolate 573, like the CL isolate, produced high parasitemia (> 10 6 parasites per ml at the peak), killing ~40% of animals, whereas infection with isolate 587 resulted in low parasitemia (< 10 5 parasites per ml), with zero mortality. On the fourth day post-inoculation, tissue sections of the mouse stomach stained with hematoxylin and eosin showed a four to sixfold higher number of epithelial cells infected with isolate 573 or CL than with isolate 587. The rate of intracellular parasite development was similar in all isolates. Mimicking in vivo infection, parasites were treated with pepsin at acidic pH and then assayed for their ability to enter HeLa cells or explanted gastric epithelial cells. Pepsin extensively digested gp90 from isolate 573 and significantly increased invasion of both cells, but had minor effect on gp90 or infectivity of isolates 587 and CL. The profile of g82 digestion was similar in isolates 573 and 587, with partial degradation to a ~70 kDa fragment, which preserved the target cell binding domain as well as the region involved in gastric mucin adhesion. Gp82 from CL isolate was resistant to pepsin. Assays with parasites recov- ered from the mouse stomach 2 h after oral infection showed an extensive digestion of gp90 and increased infectivity of isolate 573, but not of isolate 587 or CL. Our data indicate that T. cruzi infection in vitro does not always correlate with in vivo infection because host factors may act on parasites, modulating their infectivity, as is the case of pepsin digestion of isolate 573 gp90. © 2005 Elsevier SAS. All rights reserved. Keywords: Trypanosoma cruzi; Cell invasion; Metacyclic trypomastigote; Oral infection; Gp90 surface molecule 1. Introduction Studies aimed at clarifying the molecular mechanisms of Trypanosoma cruzi infection have been performed in vitro, using various types of mammalian cells. To what extent the phenomena observed in cultured cells are relevant in T. cruzi infection in vivo remains to be determined. Using tissue culture-derived trypomastigotes, which are equivalent to the parasite forms circulating in the blood, Toma et al. [1] found no correlation between infectivity in vitro and the para- sitemia produced in mice. As multiple factors, not present in culture conditions, are expected to act in vivo, in many instances the results obtained in vitro may not be predictive of infection in mammalian hosts. A number of surface and/or secreted parasite molecules, implicated in the crucial step of host cell invasion, have been identified and characterized. In tissue culture trypomastigotes, the laminin-binding surface glycoprotein Tc-85 [2,3] and enzymes that are expressed at the parasite surface and/or secreted, such as trans-sialidase * Corresponding author. Tel.: +55 11 5576 4532; fax: +55 11 5571 1095. E-mail address: [email protected] (N.Yoshida). Microbes and Infection 8 (2006) 36–44 www.elsevier.com/locate/micinf 1286-4579/$ - see front matter © 2005 Elsevier SAS. All rights reserved. doi:10.1016/j.micinf.2005.05.016
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Trypanosoma cruzi surface molecule gp90 downregulates invasion of gastric mucosal epithelium in...

Original article
Trypanosoma cruzi surface molecule gp90 downregulates invasionof gastric mucosal epithelium in orally infected mice
Mauro Cortez a, Marcelo R. Silva a, Ivan Neira a, Daniele Ferreira a, Gisela R.S. Sasso b,Alejandro O. Luquetti c, Anis Rassi c, Nobuko Yoshida a,*
a Departamento de Microbiologia, Imunologia e Parasitologia, Escola Paulista de Medicina, Universidade Federal de São Paulo, R. Botucatu,862- 6° andar, 04023-062 São Paulo, SP, Brazil
b Departamento de Morfologia, Universidade Federal de São Paulo, São Paulo, Brazilc Laboratório de Doença de Chagas, Hospital das Clínicas, Faculdade de Medicina, Universidade Federal de Goiás, Brazil
Received 10 January 2005; received in revised form 6 May 2005; accepted 23 May 2005
Available online 02 August 2005
Abstract
Experiments were performed to elucidate why Trypanosoma cruzi isolates 573 and 587 differ widely in their efficiency to infect gastricmucosal epithelium when administered orally to mice. These isolates have the same surface profile and a similar capacity to enter host cells invitro. Metacyclic forms of isolates 573 and 587 and the control CL isolate expressed similar levels of gp82, which is a cell invasion-promotingmolecule. Expression of gp90, a molecule that downregulates cell invasion, was lower in the CL isolate. Consistent with this profile, ~three-fold fewer parasites of isolates 573 and 587 entered epithelial HeLa cells, as compared to the CL isolate. No difference in the rate of intrac-ellular parasite replication was observed between isolates. When given orally to mice, metacyclic forms of isolate 573, like the CL isolate,produced high parasitemia (> 106 parasites per ml at the peak), killing ~40% of animals, whereas infection with isolate 587 resulted in lowparasitemia (< 105 parasites per ml), with zero mortality. On the fourth day post-inoculation, tissue sections of the mouse stomach stained withhematoxylin and eosin showed a four to sixfold higher number of epithelial cells infected with isolate 573 or CL than with isolate 587. The rateof intracellular parasite development was similar in all isolates. Mimicking in vivo infection, parasites were treated with pepsin at acidic pHand then assayed for their ability to enter HeLa cells or explanted gastric epithelial cells. Pepsin extensively digested gp90 from isolate573 and significantly increased invasion of both cells, but had minor effect on gp90 or infectivity of isolates 587 and CL. The profile ofg82 digestion was similar in isolates 573 and 587, with partial degradation to a ~70 kDa fragment, which preserved the target cell bindingdomain as well as the region involved in gastric mucin adhesion. Gp82 from CL isolate was resistant to pepsin. Assays with parasites recov-ered from the mouse stomach 2 h after oral infection showed an extensive digestion of gp90 and increased infectivity of isolate 573, but not ofisolate 587 or CL. Our data indicate that T. cruzi infection in vitro does not always correlate with in vivo infection because host factors may acton parasites, modulating their infectivity, as is the case of pepsin digestion of isolate 573 gp90.© 2005 Elsevier SAS. All rights reserved.
Keywords: Trypanosoma cruzi; Cell invasion; Metacyclic trypomastigote; Oral infection; Gp90 surface molecule
1. Introduction
Studies aimed at clarifying the molecular mechanisms ofTrypanosoma cruzi infection have been performed in vitro,using various types of mammalian cells. To what extent thephenomena observed in cultured cells are relevant in T. cruziinfection in vivo remains to be determined. Using tissueculture-derived trypomastigotes, which are equivalent to the
parasite forms circulating in the blood, Toma et al. [1] foundno correlation between infectivity in vitro and the para-sitemia produced in mice. As multiple factors, not present inculture conditions, are expected to act in vivo, in manyinstances the results obtained in vitro may not be predictiveof infection in mammalian hosts. A number of surface and/orsecreted parasite molecules, implicated in the crucial step ofhost cell invasion, have been identified and characterized. Intissue culture trypomastigotes, the laminin-binding surfaceglycoprotein Tc-85 [2,3] and enzymes that are expressed atthe parasite surface and/or secreted, such as trans-sialidase
* Corresponding author. Tel.: +55 11 5576 4532; fax: +55 11 5571 1095.E-mail address: [email protected] (N. Yoshida).
Microbes and Infection 8 (2006) 36–44
www.elsevier.com/locate/micinf
1286-4579/$ - see front matter © 2005 Elsevier SAS. All rights reserved.doi:10.1016/j.micinf.2005.05.016

[4] and cruzipain [5], participate in target cell penetration invitro. Of these molecules, cruzipain was shown to play a rolein vivo by Engel et al. [6], who cured experimental T. cruziinfection by treating mice with peptide-fluoromethyl ketones,inhibitors that inactivate cruzipain.
Metacyclic trypomastigotes are the insect-derived T. cruzidevelopmental forms, responsible for natural infection of avariety of mammalian hosts. Studies with cultured counter-parts have shown that infectivity in mice correlates with theability to enter mammalian cells in vitro, and the varying ratesof infectivity of different T. cruzi isolates has been associatedwith differential expression of a set of surface glycoproteins[7,8]. Metacyclic forms of T. cruzi isolates that are poor invad-ers of host cells in vitro, invariably express high levels ofgp90 [8], a stage-specific molecule that negatively modulateshost cell invasion in vitro [9]. Highly invasive parasites expressreduced levels of gp90, concomitant with high expression ofgp82 [8]. Ggp82 is an adhesion molecule that promotes hostcell penetration by triggering Ca2+ mobilization. Ca2+ is mobi-lized in both cells [8] and is essential for T. cruzi internaliza-tion [10–12]. Gp82 has also been found to promote mucosalinfection upon oral inoculation [13]. The microepidemicsresponsible for the majority of acute cases of Chagas’ dis-ease recorded in the Brazilian Amazon are attributed to oralinfection. [14]. Consistent with this, mutant T. cruzi isolatesor clones deficient in gp82 are poorly infective in mice whenadministered orally [15,16].
Based on the preliminary observation that metacyclic formsof a T. cruzi isolate expressing high gp90 levels were poorlyinfective in vitro but highly infective in mice, we addressedthe question of discrepancy between in vivo and in vitro T.cruzi infectivity. Our data shed light on that question and alsovalidate the role of gp90 as a downmodulator of T. cruzi infec-tion in vivo.
2. Materials and methods
2.1. Parasites and mammalian cell invasion assays
Two T. cruzi isolates that have not been previously char-acterized were used in this study: 573 (MHOM/BR98/GOCH573), isolated in 1998 from the peripheral blood of a44 years old, acutely infected female who acquired the para-site by blood transfusion; and 587 (MHOM/BR99/GOCH587), isolated in 1999 from a congenitally infectednewborn. The CL isolate [17] was used as a control on thegrounds that metacyclic forms of this isolate are the mostextensively characterized. Parasites were maintained alter-nately in mice and liver infusion tryptose medium. To obtaincultures enriched in metacyclic trypomastigotes, parasiteswere also grown in Graces’s medium (Invitrogen). Metacy-clic forms, harvested from cultures in the stationary growthphase, were purified by passage through DEAE-cellulose col-umns, as described [18]. HeLa cells, a human carcinoma-derived epithelial cell line, were grown at 37 °C in Dulbec-
co’s minimal essential medium (DMEM), supplemented with10% fetal calf serum (FCS), streptomycin (100 µg/ml) andpenicillin (100 U/ml) in a humidified 5% CO2 atmosphere.Mammalian cell invasion assays were performed essentiallyas previously described [19]. Briefly, parasites were seededonto each well of 24-well plates containing round glass cov-erslips coated with 1.5 × 105 HeLa cells. After varying peri-ods of time, depending on the experiment, the duplicate cov-erslips were washed in PBS and stained with Giemsa. Thenumber of intracellular parasites was counted in at least500 Giemsa-stained cells.
2.2. Immunoblotting and detection of T. cruzi proteinaseactivity
To analyze the expression of metacyclic trypomastigotesurface molecules by immunoblotting, we followed the sameprocedure detailed elsewhere [19]. The final reaction wasrevealed by chemiluminescence, using the ECL Western blot-ting detection reagent and Hyperfilm-MP from Amersham.T. cruzi proteinase activity was determined as follows: 3 × 107
parasites were lyzed in 0.4% Triton X-100, solubilized insample buffer without 2-mercaptoethanol, then loaded onto10% SDS-polyacrylamide gel containing 0.1% co-polymerized gelatin as substrate. After electrophoresis, thegels were subjected to two 30 min washes with 2.5% TritonX-100 in 0.1 M acetate buffer, pH 6.0, to remove SDS, andincubated overnight in the same buffer without detergent. Thegel was stained with Coomassie blue R250 and destained forvisualization of white bands against a blue background.
2.3. Oral infection
T. cruzi metacyclic forms were inoculated by the intra-pharyngeal route into 4–5-week-old female Balb/c mice,through a plastic tube adapted to a 1 ml plastic syringe. Start-ing on day 13 post-inoculation of 4 × 105 parasites into eachmouse, parasitemia was monitored twice a week by examin-ing 5 µl peripheral blood samples collected from the tail usinga phase contrast microscope.
2.4. Preparation of histological sections
Groups of Balb/c mice were inoculated orally with 108
metacyclic trypomastigotes of different T. cruzi isolates. Fourdays post-infection, the stomach of each animal was col-lected and fixed in 10% neutral buffered formalin for 24 h.After processing by gradual dehydration in a graded ethanolsolution, followed by xylene immersion and embedding inparaffin, serial 5 µm tissue sections were cut and stained withhematoxylin and eosin for examination by light microscopy.
2.5. Recovery of metacyclic forms from mouse stomachafter oral inoculation
Balb/c mice were maintained overnight without food.Before oral inoculation of parasites, under anesthesia, the
37M. Cortez et al. / Microbes and Infection 8 (2006) 36–44

mouse stomach was exposed through a cut in the abdomen.With a knot, using a thread, the stomach was closed at thepylorus. The cut in the abdomen was sewed and the mice wereleft to fully recover from anesthesia. Then, they received 2 × 108
metacyclic trypomastigotes by intrapharingeal route. After 2 h,the cut was reopened, and an additional knot was made at thecardiac sphincter, in order to avoid the loss of gastric content.The stomach was removed from the abdominal cavity, placedin a well of six-well plate containing 1 ml of DMEM plus 20%FCS and was cut open. Avoiding as much as possible the solidmaterial, the supernatant was collected and the number of para-sites counted. About 50% of parasites were recovered.
2.6. Explant of cells from the mouse stomach
Balb/c mice, maintained overnight without food, werekilled to remove the stomach. The organ was cut open, washedseveral times with 5 ml PBS containing 100 µg/ml gentamy-cin (PBS/G) to discard its content. After transfer to a 50 mltube containing PBS/G, the organ was further washed byinversion, transferred to another tube and this washing pro-cedure was repeated through several cycles. The stomach wasplaced in a tube containing 5 ml DMEM plus 0.5 mg/ml col-lagenase, 20% FCS and 100 µg/ml gentamycin, and incu-bated at 37 °C for 4–5 h. Following the centrifugation of thereleased cells, the pellet was washed with DMEM containing20% FCS and the cells were resuspended in the same medium.Before invasion assays, the cells were maintained at 37 °Cfor 4 h to allow the recovery from the enzymatic treatment.As the explanted cells did not adhere well to surfaces, theinvasion assays with these cells were performed in suspen-sion, at 10:1 parasite/cell ratio, and the parasites that did noinvade were washed out by centrifugation. Samples were thenplaced onto glass slides, dried, fixed with methanol and stainedwith Giemsa.
2.7. Statistical analysis
The program INSTAT was used to determine significanceby Student’s t-test.
3. Results
3.1. In vitro infectivity of T. cruzi isolates and expressionof surface glycoproteins
To determine the in vitro infectivity of metacyclic formsof T. cruzi isolates 573 and 587, as compared to the controlCL isolate, invasion assays were performed by incubatingparasites with epithelial HeLa cells at 37 °C for 1 h, atparasite/cell ratio of 10:1. The number of internalized para-sites from isolates 573 and 587 was ~ threefold lower thanthat of the CL isolate (Fig. 1A). As the reduced ability ofmetacyclic forms to enter mammalian cells is associated withthe expression of surface glycoprotein, gp90 [8], which acts
as a down-regulator of cell invasion [9], the profile ofgp90 was examined. Metacyclic forms of isolates 573 and587 expressed higher levels of gp90 than the CL isolate(Fig. 1B). As regards the invasion-promoting molecule gp82,the three isolates expressed similar levels (Fig. 1B). In addi-tion to gp82, MAb 3F6 detected the glycoprotein gp30 inisolates 573 and 587, a molecule that plays a role in host cellinvasion of gp82-deficient isolates [15]. MAb 3F6 recog-nized a 25 kDa component in the CL isolate (Fig. 1B), whichmay be the unglycosylated counterpart of gp30 [15]. The threeisolates expressed mucin-like gp35/50 molecules at similarlevels (Fig. 1B).
3.2. Intracellular replication of T. cruzi isolates in vitroand cruzipain activity
We compared the course of HeLa cell infection by the dif-ferent isolates, up to the release of trypomastigotes. Metacy-clic forms were incubated with HeLa cells for 2 h and, afterwashings in PBS, the coverslips were reconstituted withmedium containing 2% FCS and maintained for up to 7 days,by changing the medium every other day. The number of intra-cellular parasites increased with time. By day 4, HeLa cellswere full of amastigotes and by day 5 the first trypomastig-
Fig. 1. In vitro infectivity of different T. cruzi isolates and profile of surfaceglycoproteins. A) Metacyclic forms of the indicated T. cruzi isolates wereincubated with HeLa cells for 1 h at 37 °C and the number of intracellularparasites was counted in at least 500 Giemsa-stained cells. Values aremean ± S.D. of seven independent experiments performed in duplicate. Thedifference observed between isolates 573/587 and CL was extremely signi-ficant (P < 0.0001), using the Student’s t-test. In panel B is shown the immu-noblot analysis of parasites using MAb 5E7 directed to gp90, MAb 3F6 thatrecognizes gp82 and gp30 molecules, and MAb 2B10 reacting with gp35/50,can be seen.Note the reduced expression of gp90 in the CL isolate.
38 M. Cortez et al. / Microbes and Infection 8 (2006) 36–44

otes were seen in the culture medium. The pattern of intrac-ellular parasite development was the same for isolates 573,587 and CL (Fig. 2A). In parallel, we examined the activityof cruzipain, the T. cruzi cysteine proteinase implicated inintracellular parasite replication [5]. The cruzipain activity,
detected using SDS-PAGE gels containing gelatin, was simi-lar in the three isolates (Fig. 2B).
3.3. Course of infection by T. cruzi isolates in miceinoculated orally with metacyclic forms
According to Hoft et al. [20], metacyclic forms given orallyto mice preferentially invade gastric mucosal epithelium, thisbeing the only port of entry for systemic T. cruzi infection.We have found that oral infection of Balb/c mice with meta-cyclic forms of the CL isolate leads to high parasitemia and~ 30% mortality [13]. Here we compared the course of oralinfection of isolates 573 and 587 to that of the CL isolate.From the results of in vitro infection assays (Figs. 1A and2A), we expected that parasitemia levels resulting from inocu-lation of isolates 573 and 587 would be comparable, and lowerthan those produced by the CL isolate. However, this was notthe case. In repeated experiments, metacyclic forms of iso-late 573 produced high parasitemia, killing ~ 40% of ani-mals, whereas reduced parasitemia and zero mortality wasobtained from infection with isolate 587 (Fig. 3A). Infectiv-ity of isolate 573 by the oral route was even higher than thatof the CL isolate. In A/Sn mice and out bred Swiss miceinfected orally, the results were similar, isolate 573 consis-tently producing much higher parasitemia than isolate 587(data not shown).
To examine the intracellular parasite development in vivo,Balb/c mice were inoculated orally with 108 metacyclic formsof isolate 573, 587 or CL. In this experiment we used a highnumber of parasites in the inoculum in order to visualize iso-late 587 parasites in the gastric mucosa. Four days post-inoculation, histological sections of the mouse stomach werestained with hematoxylin–eosin and examined for the pres-ence of parasites. In equivalent tissue sections of infected
Fig. 2. Intracellular replication of T. cruzi isolates in HeLa cells and cruzi-pain activity. A) Metacyclic trypomastigotes were incubated for 2 h withHeLa cells which, after washings in PBS and reconstitution with mediumcontaining 2% FCS, were maintained for up to 7 days. Shown are the cellscontaining replicating amastigotes of the indicated T. cruzi isolates, at 72 hpost-infection. B) Cruzipain activity was determined by analyzing parasiteextracts in SDS-PAGE gels containing gelatin. Comparable cruzipain acti-vity was detected in the three isolates.
Fig. 3. Course of infection in mice inoculated orally with different T. cruzi isolates. A) Balb/c mice were infected with metacyclic forms (4 × 105 parasites permouse) of the indicated T. cruzi isolates. Representative results of one out of three experiments are shown. Each data point corresponds to the meanparasitemia ± S.D. of six mice in each group. Isolate 573 produced the highest parasitemia throughout the course of acute infection. The difference in parasi-temia levels between isolate 573 and 587 was highly significant (P ≤ 0.0001 on days 17, 20, 31; P < 0.001 on days 24, 27) using the Student’s t-test. B) Gastricmucosal sections stained with hematoxylin and eosin were examined for replication of T. cruzi at fourth day post-inoculation of metacyclic forms. The numberof cells containing amastigote nests of isolae 573 or CL was significantly higher (P < 0.0001) than that of isolate 587. Values are the mean ± S.D. of infectedcells in 30 tissue sections from three animals (isolate 573), 30 sections from five animals (isolate CL) and 17 sections from four animals (isolate 587). In miceinfected with isolate 587 about 50% of the tissue sections were negative and these were not included in the analysis.
39M. Cortez et al. / Microbes and Infection 8 (2006) 36–44
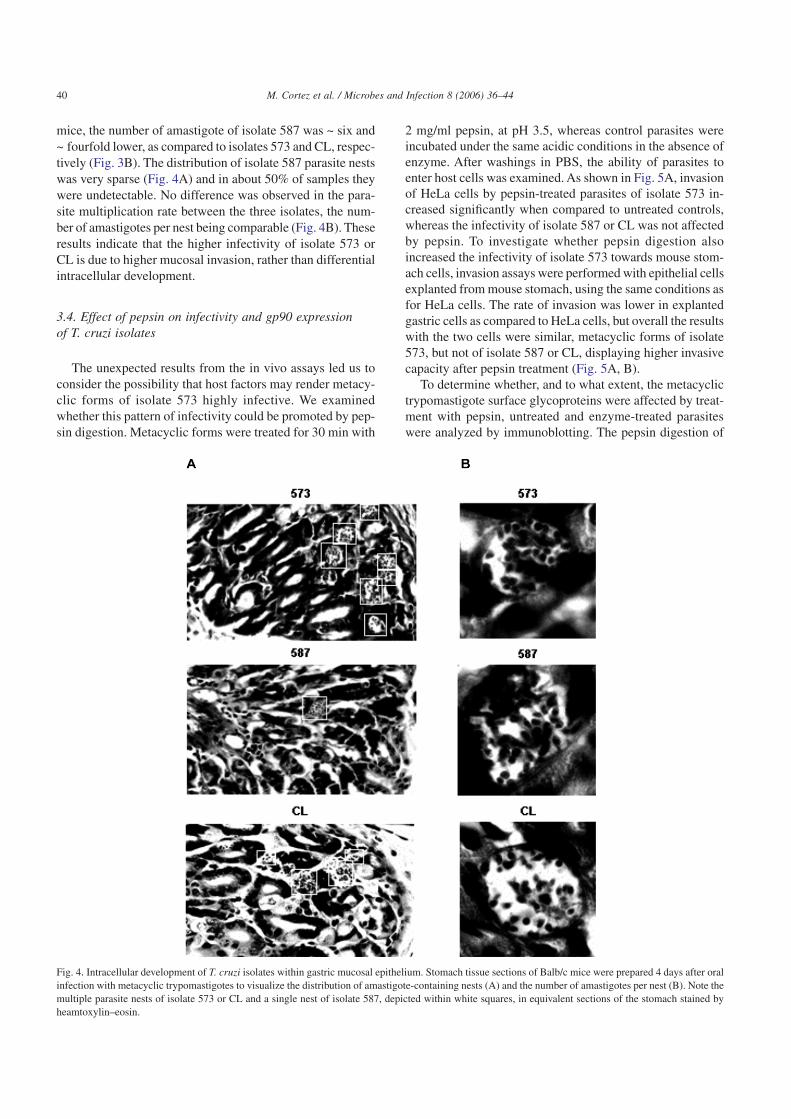
mice, the number of amastigote of isolate 587 was ~ six and~ fourfold lower, as compared to isolates 573 and CL, respec-tively (Fig. 3B). The distribution of isolate 587 parasite nestswas very sparse (Fig. 4A) and in about 50% of samples theywere undetectable. No difference was observed in the para-site multiplication rate between the three isolates, the num-ber of amastigotes per nest being comparable (Fig. 4B). Theseresults indicate that the higher infectivity of isolate 573 orCL is due to higher mucosal invasion, rather than differentialintracellular development.
3.4. Effect of pepsin on infectivity and gp90 expressionof T. cruzi isolates
The unexpected results from the in vivo assays led us toconsider the possibility that host factors may render metacy-clic forms of isolate 573 highly infective. We examinedwhether this pattern of infectivity could be promoted by pep-sin digestion. Metacyclic forms were treated for 30 min with
2 mg/ml pepsin, at pH 3.5, whereas control parasites wereincubated under the same acidic conditions in the absence ofenzyme. After washings in PBS, the ability of parasites toenter host cells was examined. As shown in Fig. 5A, invasionof HeLa cells by pepsin-treated parasites of isolate 573 in-creased significantly when compared to untreated controls,whereas the infectivity of isolate 587 or CL was not affectedby pepsin. To investigate whether pepsin digestion alsoincreased the infectivity of isolate 573 towards mouse stom-ach cells, invasion assays were performed with epithelial cellsexplanted from mouse stomach, using the same conditions asfor HeLa cells. The rate of invasion was lower in explantedgastric cells as compared to HeLa cells, but overall the resultswith the two cells were similar, metacyclic forms of isolate573, but not of isolate 587 or CL, displaying higher invasivecapacity after pepsin treatment (Fig. 5A, B).
To determine whether, and to what extent, the metacyclictrypomastigote surface glycoproteins were affected by treat-ment with pepsin, untreated and enzyme-treated parasiteswere analyzed by immunoblotting. The pepsin digestion of
Fig. 4. Intracellular development of T. cruzi isolates within gastric mucosal epithelium. Stomach tissue sections of Balb/c mice were prepared 4 days after oralinfection with metacyclic trypomastigotes to visualize the distribution of amastigote-containing nests (A) and the number of amastigotes per nest (B). Note themultiple parasite nests of isolate 573 or CL and a single nest of isolate 587, depicted within white squares, in equivalent sections of the stomach stained byheamtoxylin–eosin.
40 M. Cortez et al. / Microbes and Infection 8 (2006) 36–44

gp90 molecule of isolate 573 was so extensive that most of itwas degraded to small pieces, not detectable in SDS-PAGEgel, only a small fraction of it remaining as large 70–80 kDafragments (Fig. 5C). Gp90 molecules of isolates 587 and CLwere also susceptible to pepsin digestion, but only to someextent, so that they were mostly preserved as 70–80 kDa frag-ments (Fig. 5C, upper panel). To rule out the possibility ofunequal loading of parasite samples, the same blot was alsoreacted with MAb 3F6, which recognizes gp30 from isolates573 and 587 as well as the 25 kDa protein from CL isolate,all of which are resistant to pepsin digestion (Fig. 5C, lowerpanel). The gp82 molecule was similarly affected by pepsinin isolates 573 and 587, being partially digested to fragmentsof ~70 kDa (Fig. 5D). This partial digestion, with removal ofthe N-terminal domain preserves the host cell binding siteintact as well as the gastric mucin adhesion sites, which arelocated in the C-terminal half of the molecule [13]. CL iso-late gp82 was completely resistant to pepsin digestion(Fig. 5D).
3.5. Effect of gastric content on the infectivity of T. cruzimetacyclic forms
To determine how the contact with the gastric contentaffected the infectivity of different T. cruzi isolates, Balb/cmice were infected with metacyclic forms by oral route,according to the procedure described in Section 2, and theparasites recovered from the stomach were tested for infec-tivity towards HeLa cells. Compared to the control metacy-clic forms left in the incubator, the parasites of isolate 573,but not of isolate 587 or CL, recovered from mouse stomach,displayed higher infectivity (Fig. 6A). Western blot analysisof recovered parasites showed an extensive digestion ofgp90 in isolate 573, as opposed to more resistant gp90 in iso-lates 587 and CL (Fig. 6B), confirming the higher suscepti-bility of isolate 573 gp90 to gastric enzymes. Compatible withthe fact that in vivo several other factors, in addition to pep-sin, are acting on the parasite, some differences were foundin the profiles of gp90 digestion in vivo and in vitro (Figs. 5C
Fig. 5. Effect of pepsin on T. cruzi infectivity and the parasite surface profile. A) Metacyclic forms of the indicated T. cruzi isolates were either untreated ortreated with 2 mg/ml pepsin, at pH 3.5, washed in PBS and then incubated for 1 h at 37 °C with HeLa cells (A) or cells explanted from mouse stomach (B). Thenumber of intracellular parasites was counted in at least 500 Giemsa-stained cells. Values are mean ± S.D. of three experiments performed in duplicate. Theincrease in the number of internalized parasites of isolate 573 treated with pepsin, as compared to untreated controls, was significant (P = 0.01 in HeLa cells andP < 0.0005 in explanted cells). In (C) and (D), metacyclic forms, untreated or treated with pepsin, were processed for immunoblotting using MAb 5E7 (C, upperpanel), or MAb 3F6 (C lower panel and D). Note the extensive digestion of gp90 of isolate 573, as opposed to the relative resistance of gp90 of isolate 587 topepsin digestion.
41M. Cortez et al. / Microbes and Infection 8 (2006) 36–44

and 6B). As regards gp82, metacyclic forms recovered fromthe stomach preserved the molecule intact, irrespective of theT. cruzi isolate (data not shown).
4. Discussion
Our results provide evidence that host factors acting onthe parasite can modulate T. cruzi infection in vivo. Exten-sive digestion of gp90 by pepsin increases the ability of iso-late 573 metacyclic forms to enter HeLa cells as well as epi-thelial cells explanted from mouse stomach. Pepsin digestionmay occur in vivo, rendering the parasites highly efficient at
invading gastric mucosal epithelium, and this would explainthe discrepancy between in vitro and in vivo infectivity ofisolate 573. Expression of high levels of gp90, a moleculethat negatively modulates mammalian cell invasion [9], deter-mines the low infectivity of isolate 573 in vitro. However, inthe mouse stomach, most of gp90 may be removed from theparasite surface, facilitating invasion of epithelial cells. In sup-port of this notion is the finding that metacyclic forms of iso-late 573 recovered from the mouse stomach after oral infec-tion, and expressing lower gp90 levels, are more invasive thancontrol parasites. Metacyclic forms of T. cruzi isolates mayexpress different isoforms of gp90, which is encoded by amultigene family [21], the pepsin-resistant gp90 isoform beingpredominant in isolate 587.As the truncated recombinant pro-tein of gp90 without the N-terminal domain of the moleculebinds to HeLa cells in the same manner as the native gp90[7], the removal of the N-terminal portion of gp90 from iso-late 587 by pepsin treatment is expected to preserve the prop-erty of gp90 as a negative regulator of cell invasion, and thisis compatible with its poor infectivity in vitro and in vivo. Inthe case of CL isolate, the correlation between in vitro and invivo infectivity is consistent with the reduced expression ofgp90.
The ability of metacyclic trypomastigote gp90 to down-regulate host cell invasion is associated with the lack of Ca2+
signal-inducing activity. Intracellular Ca2+ mobilization, bothin the target cell and the parasite, is required for T. cruzi inter-nalization [8,12]. Binding of gp90 to mammalian cells doesnot trigger these Ca2+ responses [8]. We presume that the inter-action of gp90 with its receptor triggers an inhibitory path-way in both cells, in a similar way to cells of the immunesystem where, in addition to activation signals, signaling cas-cades acting as negative regulators can be induced [22,23].Like NK cell surface inhibitory receptors that antagonize acti-vation pathways using protein tyrosine phosphatases [22],metacyclic trypomastigote gp90 mediates the activation of aprotein tyrosine phosphatase that counteracts the action ofprotein tyrosine kinase in the parasite [24].
Activation of protein tyrosine kinase and Ca2+ mobiliza-tion is induced by the binding of gp82 to its receptor in para-sites that engage gp82 to enter target cells [25,26], as is thecase of the isolates analyzed in this study. In addition to itsrole in mammalian cell invasion in vitro [7], gp82 is impli-cated in gastric mucin adhesion and mucosal invasion in orallyinfected mice [13]. Gp82-mediated interaction with gastricmucin is probably the first step towards the penetration ofmetacyclic trypomastigotes into the underlying epithelialcells, in the same manner as the pathogenic potential of Shi-gella dysenteriae is correlated with its ability to preferen-tially adhere to colon mucin and to invade and multiply withinthe cells of the colon epithelium [27]. In accord with thishypothesis, recent studies have shown that T. cruzi isolatesdeficient in expression of gp82 are poorly infective when givenorally to mice. However, these gp82-deficient isolates effi-ciently infected HeLa cells [15]. In this case, the discrepancybetween in vitro and in vivo infectivity was ascribed to the
Fig. 6. Infectivity and surface profile of T. cruzi metacyclic forms recoveredfrom mouse stomach. A) Control parasites, and metacyclic forms recoveredfrom mice 2 h after oral infection, were incubated with HeLa cells for 1 h at37 °C. The number of intracellular parasites was counted in at least500 Giemsa-stained cells. Values are mean ± S.D. of three experiments per-formed in duplicate. The increase in the number of internalized parasites ofisolate 573 recovered from mouse stomach, as compared to controls, wassignificant (P < 0.005). B) Control and experimental parasites were analy-zed by immunobloting using MAb 5E7 directed to gp90. Note the extensivedigestion of gp90 from isolate 573, but not of isolates 587 and CL.
42 M. Cortez et al. / Microbes and Infection 8 (2006) 36–44

expression of gp30. This metacyclic stage surface moleculebinds to HeLa cells, inducing the Ca2+ response necessaryfor parasite internalization but, in contrast to gp82, binds inef-ficiently to gastric mucin, thus hampering mucosal infection[15]. The gp82 binding site to target cells, as well as to gas-tric mucin, is located in the carboxy-terminal half of the mol-ecule [13,28,29]. Therefore, the removal of its N-terminal por-tion by pepsin, in T. cruzi isolates 573 and 587, does notinterfere with the properties to bind to gastric mucin or toenter host cells.
Based on these findings, we visualize the following sce-nario: when T. cruzi metacyclic forms reach the mouse stom-ach, they resist destruction because they are protected bymucin-like gp35/50 molecules [30], which are abundant onthe surface and are resistant to protease digestion [31]. Meta-cyclic forms of CL isolate, which express gp90 at low levelsand a pepsin-resistant gp82, efficiently interact with gastricmucin and enter underlying epithelial cells. In the case ofisolate 573, upon extensive removal of gp90 by pepsin diges-tion, the parasites bind to gastric mucin in a gp82 dependentmanner, traverse the mucus layer and efficiently invade epi-thelial cells. Metacyclic forms of isolate 587 follow the sameroute, with the difference that the entrance into epithelial cellsis hampered by the presence of protease-resistant gp90. Thisdifferential invasion of gastric mucosal epithelium leads tothe difference in parasitemia levels in mice infected with iso-late 573/CL and 587.
Acknowledgements
We thank Dr. Ionizete Garcia da Silva for help in the iso-lation of T. cruzi isolate 587, Dr. José Daniel Lopes for read-ing the manuscript. This work was supported by Fundação deAmparo à Pesquisa do Estado de São Paulo (FAPESP) andConselho Nacional de Desenvolvimento Científico e Tec-nológico (CNPq), Brazil.
References
[1] K.H. Toma, I.P. Cerávolo, H.L. Guerra, M. Steindel, A.J. Romanha,Trypanosoma cruzi: parasitemia produced in mice does not seem to berelated to in vitro parasite–cell interaction, Int. J. Parasitol. 30 (2000)593–597.
[2] M.J.M.Alves, G.Abuin, V.Y. Kuwajima, W. Colli, Partial inhibition oftrypomastigote entry into cultured mammalian cells by monoclonalantibodies against a surface glycoprotein of Trypanosoma cruzi, Mol.Biochem. Parasitol. 21 (1986) 75–82.
[3] R. Giordano, R. Chammas, S.S. Veiga, W. Colli, M.J.M. Alves, Anacidic component of the heterogeneous Tc-85 protein family from thesurface of Trypanosoma cruzi is a laminin binding glycoprotein, Mol.Biochem. Parasitol. 65 (1994) 85–94.
[4] S. Schenkman, M. Jiang, G.W. Hart, V. Nussenzweig, A novel cellsurface trans-sialidase of Trypanosoma cruzi generates a stage-specific epitope required for invasion of mammalian cells, Cell 65(1991) 1117–1125.
[5] M.N. Meirelles, L. Juliano, E. Carmona, S.G. Silva, E.M. Costa,A.C. Murta, J. Scharfstein, Inhibitors of the major cysteinyl proteinase(gp57/51) impair host cell invasion and arrest the intracellular devel-opment of Trypanosoma cruzi in vitro, Mol. Biochem. Parasitol. 52(1992) 175–184.
[6] J.C. Engel, O.S. Doyle, I. Hsich, J.H. Mckerrow, Cysteine proteaseinhibitors cure an experimental Trypanosoma cruzi infection, J. Exp.Med. 188 (1998) 725–734.
[7] M.I. Ramirez, R.C. Ruiz, J.E. Araya, J. Franco da Silveira, N.Yoshida,Involvement of the stage-specific 82-kDa adhesion molecule of Try-panosoma cruzi metacyclic trypomastigotes in host cell invasion,Infect. Immun. 61 (1993) 3636–3641.
[8] R.C. Ruiz, S. Favoreto Jr., M.L. Dorta, M.E.M. Oshiro, A.T. Ferreira,P.M. Manque, N. Yoshida, Infectivity of Trypanosoma cruzi strains isassociated with differential expression of surface glycoproteins withdifferential Ca2+ signaling activity, Biochem. J. 330 (1998) 505–511.
[9] S. Málaga, N. Yoshida, Targeted reduction in expression of Trypano-soma cruzi surface glycoprotein gp90 increases parasite infectivity,Infect. Immun. 69 (2001) 353–359.
[10] S.N.J. Moreno, J. Silva, A.E. Vercesi, R. Docampo, Cytosolic-freecalcium elevation in Trypanosoma cruzi is required for cell invasion,J. Exp. Med. 180 (1994) 1535–1540.
[11] I. Tardieux, M.H. Nathanson, N.W. Andrews, Role in host cell inva-sion of Trypanosoma cruzi-induced cytosolic free Ca2+ transients, J.Exp. Med. 179 (1994) 1017–1022.
[12] M.L. Dorta, A.T. Ferreira, M.E.M. Oshiro, N. Yoshida, Ca2+ signalinduced by Trypanosoma cruzi metacyclic trypomastigote surfacemolecules implicated in mammalian cell invasion, Mol. Biochem.Parasitol. 73 (1995) 285–289.
[13] I. Neira, F.A. Silva, M. Cortez, N. Yoshida, Involvement of Trypano-soma cruzi metacyclic trypomastigote surface molecule gp82 in adhe-sion to gastric mucin and invasion of epithelial cells, Infect. Immun.71 (2003) 557–561.
[14] J.R. Coura, A.C.V. Junqueira, O. Fernandes, S.A.S. Valente,M.A. Miles, Emerging Chagas disease in Amazonian Brazil, TrendsParasitol. 18 (2002) 171–176.
[15] M. Cortez, I. Neira, D. Ferreira, A.O. Luquetti, A. Rassi, V.D. Atayde,N. Yoshida, Infection by Trypanosoma cruzi metacyclic forms defi-cient in gp82 but expressing a related surface molecule gp30, Infect.Immun. 71 (2003) 6184–6191.
[16] V.D. Atayde, I. Neira, M. Cortez, D. Ferreira, E. Freymuller,N. Yoshida, Molecular basis of non virulence of Trypanosoma cruziclone CL-14, Int. J. Parasitol. J. 34 (2004) 851–860.
[17] Z. Brener, E. Chiari, Variações morfológicas observadas em diferentesamostras de Trypanosoma cruzi, Rev. Inst. Med. Trop. Sao Paulo 5(1963) 220–224.
[18] M.M.G. Teixeira, N.Yoshida, Stage-specific surface antigens of meta-cyclic trypomastigotes of Trypanosoma cruzi identified by mono-clonal antibodies, Mol. Biochem. Parasitol. 18 (1986) 271–282.
[19] N. Yoshida, R.A. Mortara, M.F. Araguth, J.C. Gonzalez, M. Russo,Metacyclic neutralizing effect of monoclonal antibody 10D8 directedto the 35 and 50 kDa surface glycoconjugates of Trypanosoma cruzi,Infect. Immun. 57 (1989) 1663–1667.
[20] D.F. Hoft, P.L. Farrar, K. Kratz-Owens, D. Shaffer, Gastric invasionby Trypanosoma cruzi and induction of protective mucosal immuneresponses, Infect. Immun. 64 (1996) 3800–3810.
[21] F.R.S. Franco, G. Paranhos-Bacalla, L.M. Yamauchi, N. Yoshida,J. Franco da Silveira, Characterization of a cDNA clone encoding thecarboxy-terminal domain of a 90-kDa surface antigen of Trypano-soma cruzi metacyclic trypomastigotes, Infect. Immun. 61 (1993)4196–4201.
[22] E. Vivier, J.A. Nunès, F. Vély, Natural killer cell signaling pathways,Science 306 (2004) 1517–1519.
[23] A. Veillette, S. Latour, D. Davidson, Negative regulation of immu-noreceptor signaling, Annu. Rev. Immunol. 20 (2002) 669–707.
43M. Cortez et al. / Microbes and Infection 8 (2006) 36–44

[24] P.M. Manque, I. Neira, V.D. Atayde, E. Cordero, A.T. Ferreira,J. Franco da Silveira, M. Ramirez, N.Yoshida, Cell adhesion and Ca2+
signaling activity in stably transfected Trypanosoma cruzi epimastig-otes expressing the metacyclic stage-specific surface molecule gp82,Infect. Immun. 71 (2003) 1561–1565.
[25] S. Favoreto Jr., M.L. Dorta, N. Yoshida, Trypanosoma cruzi 175 kDaprotein tyrosine phosphorylation is associated with host cell invasion,Exp. Parasitol. 89 (1998) 188–194.
[26] N. Yoshida, S. Favoreto Jr., A.T. Ferreira, P.M. Manque, Signal trans-duction induced in Trypanosoma cruzi metacyclic trypomastigotesduring the invasion of mammalian cells, Braz. J. Med. Biol. Res. 33(2000) 269–278.
[27] P.S. Sudha, H. Devaraj, N. Devaraj, Adherence of Shigella dysente-riae 1 to human colonic mucin, Curr. Microbiol. 42 (2001) 381–387.
[28] F.R. Santori, M.L. Dorta, L. Juliano, M.A. Juliano, J. Franco daSilveira, R.C. Ruiz, N. Yoshida, Identification of a domain of Trypa-
nosoma cruzi metacyclic trypomastigote surface moleculegp82 required for attachment and invasion of mammalian cells, Mol.Biochem. Parasitol. 78 (1996) 209–216.
[29] P.M. Manque, D. Eichinger, M.A. Juliano, L. Juliano, J.E. Araya,N.Yoshida, Characterization of the cell adhesion site of Trypanosomacruzi metacyclic stage surface glycoprotein gp82, Infect. Immun. 68(2000) 478–484.
[30] S. Schenkman, M.A.J. Ferguson, N. Heise, M.L. Cardoso de Almeida,R.A. Mortara, N. Yoshida, Mucin-like glycoproteins linked to themembrane by glycosylphosphatidylinositol anchor are the majoracceptors of sialic acid in a reaction catalysed by trans-sialidase inmetacyclic forms of Trypanosoma cruzi, Mol. Biochem. Parasitol. 59(1993) 293–304.
[31] R.A. Mortara, S. Silva, M.F. Araguth, S.A. Blanco, N. Yoshida, Poly-morphism of the 35- and 50-kDa surface glycoconjugates of Trypano-soma cruzi metacyclic trypomastigotes, Infect. Immun. 60 (1992)4673–4678.
44 M. Cortez et al. / Microbes and Infection 8 (2006) 36–44




















