Expression of matrix metalloproteinases during murine chorioallantoic placenta maturation

Differential Expression of Matrix Metalloproteinasesand Their Tissue Inhibitors at the AdvancingPterygium Head
Nick Di Girolamo,1 Denis Wakefield,1 and Minas T. Coroneo2
PURPOSE. Pterygia are a proliferative and inflammatory growth of limbal epithelial stem cell origin,characterized by corneal tissue invasion and extensive matrix remodeling including the destructionof Bowman’s layer (BL). The purpose of this study was to determine the expression of matrixmetalloproteinases (MMPs) and tissue inhibitors of MMPs (TIMPs) at the advancing pterygium edge.
METHODS. Formalin-fixed, paraffin-embedded whole eyes (n 5 11) with pterygia attached, wereserially sectioned and analyzed immunohistochemically to determine the spatial distribution of fourMMPs and three TIMPs. Tear samples were collected from other patients with pterygia (n 5 11) anddisplayed by gelatin zymography.
RESULTS. Collagenase-1 was expressed by pterygium epithelial cells, corneal stromal fibroblasts andpterygium fibroblasts that had migrated between the epithelium and BL at the advancing pterygiumedge. Collagenase-3 and gelatinases A and B were detected in all pterygia, intensely stainingcolumnar epithelial cells directly adjacent to the denatured BL. In addition, gelatinase A immuno-reactivity was observed on BL. Immunoreactivity for TIMP-1 and -3 paralleled that of the gelatinases,with more intense staining in epithelial cells and fibroblasts where BL was absent. TIMP-2 wasfaintly detected in pterygium epithelial cells but intensely stained pterygium fibroblasts. GelatinaseB was the most abundant gelatinolytic enzyme present in tears, elevated approximately twofold ineyes with pterygia versus the contralateral control eyes.
CONCLUSIONS. This investigation is the first to identify the expression pattern of MMPs and TIMPs atthe advancing pterygium edge in specimens of human eyes and in tears derived from patients withpterygia. These enzymes may be responsible for the destruction of BL, and their pattern ofdifferential expression suggests that each may play a selective role in the pathogenesis of pterygia.(Invest Ophthalmol Vis Sci. 2000;41:4142–4149)
Pterygia represent an invasion of a wing of altered ocularsurface tissue into the normal cornea.1 Although thepathogenesis is still poorly understood, it has been pro-
posed that pterygia originate at the limbus and involve theactivation and proliferation of altered basal limbal epithelialstem cells that migrate and dissolve Bowman’s layer (BL).2
Despite the lack of knowledge regarding the pathogenesis ofpterygia, epidemiologic evidence suggests that exposure toUV-irradiation may be an initial trigger in the development ofthis lesion.3
Although there exists extensive literature on pterygia,controversy still surrounds the relative roles of the pterygiumfibroblast and epithelial cell in the pathogenesis of this disease.Recently, we have demonstrated the importance of the ptery-gium epithelial cell in the development of pterygia, because
this cell type was found to express abundant levels of matrixmetalloproteinases (MMPs) in resected tissue specimens and invitro, in a setting of inflammation.4 Lee et al.5 have also ob-served a marked upregulation of MMPs in pterygium fibroblastscompared with normal conjunctival fibroblasts. In addition,cytokines and growth factors, such as basic fibroblast growthfactor (bFGF), platelet-derived growth factor (PDGF), trans-forming growth factor (TGF)-b, and tumor necrosis factor(TNF)-a, have been localized to pterygium cells.6 Therefore, itis not unreasonable to propose that both cell types may act inconcert to degrade BL and other connective tissue structuralcomponents.
MMPs are a family of zinc-dependent endopeptidases ca-pable of denaturing most components of the extracellularmatrix.7 These enzymes share common structural and func-tional elements and are involved in physiological8,9 and patho-logic processes,10 including cancer,11 arthritis,12 and inflam-matory diseases of the eye.13,14 They are broadly divided intofour groups according to substrate specificity and include, thecollagenases, gelatinases, stromelysins, and the membrane-typeMMPs. MMPs are regulated at multiple levels including tran-scription, secretion, activation, and inhibition. The latter ac-complished by naturally occurring proteins called tissue inhib-itors of MMPs (TIMPs).15 The balance between the levels ofactivated enzymes and free TIMPs determines the overall MMPactivity. Maintenance of this equilibrium is essential, and any
From the 1Inflammation Research Unit, School of Pathology, Uni-versity of New South Wales; and the 2Department of Ophthalmology,Prince of Wales Hospital, Sydney, New South Wales, Australia.
Supported by the National Health and Medical Research Councilof Australia.
Submitted for publication April 19, 2000; revised August 2, 2000;accepted August 14, 2000.
Commercial relationships policy: N.Corresponding author: Nick Di Girolamo, Inflammation Research
Unit, School of Pathology, The University of New South Wales, Sydney2052, Australia. [email protected]
Investigative Ophthalmology & Visual Science, December 2000, Vol. 41, No. 134142 Copyright © Association for Research in Vision and Ophthalmology

disturbance in the balance is a critical determinant of proteol-ysis and tissue invasion.
The purposes of this study were to determine the expres-sion profile of MMPs and TIMPs in pterygia with intact under-lying corneal stroma, paying particular attention to the advanc-ing edge. In addition we analyzed the relative contribution ofthese proteins by both the pterygium fibroblast and epithelialcells, and finally we determined the gelatinolytic profile oftears derived from patients with pterygia.
MATERIALS AND METHODS
Donor Ocular Tissue
Whole eye specimens (n 5 11) were obtained from the De-partment of Anatomic Pathology, Prince of Wales Hospital,Sydney, Australia. All tissue specimens were formalin fixed andparaffin embedded. To determine the activation status of theMMPs, fresh pterygium specimens were obtained from surgeryand immediately placed in organ culture for 48 hours in serum-free Eagle’s minimum essential medium (EMEM, Trace Bio-sciences, Sydney, Australia). The supernatants were collectedand stored at 270°C for zymographic analysis. Only eyes withprimary pterygia and no signs of other ocular pathology orabnormality were obtained for this study. Patient details aresummarized in Table 1. All research protocols and patientswere treated in accordance with the tenets of the Declarationof Helsinki.
Collection of Tears from Patients with Pterygia
Unstimulated tear fluid (approximately 1–5 ml) was harvestedfrom patient eyes with primary pterygia (n 5 11) and controlsubjects (n 5 8) by placing a 10-ml capillary tube in the inferiorcul-de-sac of each eye. Because only small volumes could beextracted from an individual eye, protein standardization wasnot possible, because this assay would require most, if not all
the tear specimen. In addition, different volumes were oftencollected between the left and right eyes of the same patient,and diluted tear fluid often resulted in inaccurate protein mea-surements and weak or undetectable gelatinolytic bands in thezymograms. Tears were stored frozen at 270°C until used inzymography. Tears from the contralateral eyes (without ptery-gia) served as a control. All patients had unilateral lesions. Thepatients’ details are summarized in Table 2. Control subjectsincluded three women and five men, with a mean age of 33years.
Immunohistochemical Analysis
Whole human eyes with pterygia attached were seriallysectioned (2– 4 mm) and histologically evaluated by stainingwith H&E or sirius red, then processed immunohistochemi-cally as previously described.4 Briefly, sections were depar-affinized in xylene, rehydrated through decreasing gradedethanol, quenched for endogenous peroxidase, and thenincubated with preimmune serum from the secondary hostspecies. Antigen retrieval using proteinase K digestion13 wasrequired before the addition of the TIMP-2 antibody (Ab).Tissue sections were incubated with mouse primary mono-clonal Abs (see Table 3 for source and dilution) overnight at4°C, then extensively washed in 0.05 M Tris-buffered saline
TABLE 2. Tear Fluid from Patients with Pterygia
Patient Age/Sex Location/Eye Tears (ml)
DJ 40/M N/R 1.5JP 28/M N/L 1.3SD 40/F N/L 1.5NH 60/F N/R 2.7NM 32/M N/L 3.0SC 38/F N/L 3.5PP 29/M N/L 1.2CH 32/M N/L 1.3DW 66/M N/L 3.0RW 22/F N/R 2.5JL 44/M N/L 4.5
All pterygia were localized on the nasal side of the left (L) or theright (R) eye. The same volume of tears from both the pterygia affectedand the control contralateral eye from the one subject was analyzed byzymography. N, nasal.
TABLE 3. Antibodies Used for Immunohistochemical Analyses
Antibody Source Dilution Clone
Collagenase-1 ICN 1:500 41-1E5Collagenase-3 ORP 1:25 181-15A12Gelatinase A ICN 1:20 42-5D11Gelatinase B ICN 1:100 56-2A4TIMP-1 ICN 1:50 147-6D11TIMP-2 ICN 1:50 67-4H11TIMP-3 ORP 1:50 136-13H4Mouse IgG1 Dako 1:50 DAK-G01
All Abs were titrated using a range of concentrations, until optimaldilutions were established for the formalin-fixed ocular tissue. TheseAbs are directed against human antigens, show no cross-reactivity (asspecified by the manufacturer), and are all IgG1 subclass mouse mono-clonal Abs. Dako, ICN Biomedicals (Sydney, Australia) Oncogene Re-search Products (ORP; Cambridge, MA).
TABLE 1. Ocular Tissue Used for Histochemical and ZymographicAnalysis
Donor Age/Sex Location/Eye Cause of Death
1 81/F N/R Intracranial hemorrhage2 48/M N/R Multiple injuries3 62/M N/R Myocardial infarct4 71/M N/R Respiratory failure5 83/M N&T/R Aortic aneurysm6 67/M N/R Respiratory failure7 58/M N/L Carbon monoxide poisoning8 69/M N&T/L Heart disease9 44/F T/L Glioblastoma
10 60/M N/L Cardiac arrest11 78/F N/R Cardiac arrest12 53/M N/R13 49/F N/R14 28/M N/R15 60/M N/L
Histological examination by an ocular pathologist revealed noother ocular abnormalities. Other disease complications included dia-betes (donor 1), emphysema (donor 4), depression (donor 7), andosteoarthritis (donor 10). Tissue from patients 12 through 15 werefresh surgical specimens from living patients. N, nasal; T, temporal; R,right; L, left.
IOVS, December 2000, Vol. 41, No. 13 MMPs and TIMPs at the Invading Pterygium Edge 4143

(pH 7.6), before the addition of a biotinylated goat anti-mouse secondary Ab. Sections were again washed, incu-bated for 1 hour with horseradish peroxidase-conjugatedstreptavidin (Dako, Carpinteria, CA), and the immunoreac-tivity revealed by adding 3-amino-9-ethylcarbazole (Sigma,Sydney, Australia). Control reactions included, incubatingsections with an isotype Ab (see Table 3), omitting theprimary Ab or adding preimmune serum. Sections werecounterstained with hematoxylin.
Gelatin Substrate Zymography
Zymography was performed as previously described.4,13,14
Briefly, tear samples were thawed and an equal volume of tearsfrom eyes with and without pterygia were diluted with nonre-ducing sample buffer (10% sodium dodecyl sulfate [SDS], 4%sucrose, 0.25M Tris-HCl [pH 6.8], with 0.1% bromophenolblue) and loaded without boiling under nonreducing condi-tions. After electrophoresis, the gels were washed twice for 30minutes each in 2.5% Triton X-100 (Sigma). Gels were rinsedand incubated overnight at 37°C in substrate buffer (50 mMTris-HCl [pH 7.4], 10 mM CaCl2, and 0.02% NaN3), and stained(Coomassie Blue R-250; Bio-Rad, Sydney, Australia). Enzymaticactivity was identified as clear zones in a blue-stained back-ground. A low-range molecular weight standard (Bio-Rad) wasrun in adjacent lanes. MMP identity and activity were verifiedby running a sample of conditioned media derived from phor-bol myristate acetate (PMA, Sigma)–stimulated pterygium epi-thelial cells (a potent enhancer of MMP expression).13 Theaddition of EDTA (10 mM final) and 1,10-phenanthroline (1mM final; Sigma) completely abolished the gelatinolytic activ-ity. Photographs were taken (665 film Polaroid; Cambridge,MA) and the negative exposures scanned with a densitometer
(Hoefer Scientific, San Francisco, CA) to obtain semiquantita-tive data.
RESULTS
Histopathologic Features of theInvading Pterygium
Histologic features of pterygia included a multilayered pro-liferating squamous epithelium, (Figs. 1A, 1B) which de-creased in thickness at the advancing edge (Fig. 1C) wherethe pterygium encroached on the normal cornea. Behind theadvancing head there was an accumulation of fibrous and/orelastotic connective tissue matrix (Fig. 1A, dm). These re-gions usually contain inflammatory cells (micrographs notshown4) and blood vessels (Fig. 1A, v) that may provideboth stimulatory signals and nutrients for the proliferatingand invading tissue mass. A characteristic feature of pterygiais the loss of BL, a natural barrier that separates the cornealepithelium from the underlying stroma. This structure isnormally attached to the epithelial basement membrane(Fig. 1A, arrow) and is destroyed at the pterygium leadingedge (Figs. 1A, 1B, 1C, asterisks) by an as yet unknownmechanism. In regions void of BL or where BL has detachedfrom the underlying epithelium, clusters of pterygium fibro-blasts accumulate (Figs. 1B, through 1E, f). Dissolution of BLwas evident directly adjacent to pterygium epithelial cells(Figs. 1D, 1E, arrows), where BL was more irregular andfragmented (Figs. 1D, 1E, asterisks). The central corneaappeared normal, with intact BL and the absence pterygiumfibroblasts (Fig. 1C, inset).
FIGURE 1. Histologic features of human pterygia. Pterygium tissue was sectioned and stained with H&E (A, B, and C) or with sirius red (D, E),with (D) or without (E) polarized light to reveal typical representative histopathologic features of this lesion. (A, B, and C) Three contiguouscomposite photographs that show the pterygium body (A) extending to the advancing pterygium edge (C). No disease was observed at the centralcornea (inset, C). Arrows: Regions of intact BL; (p) regions where BL has become denatured. Typically, these are also areas where pterygiumfibroblasts accumulate (f). Other features include areas of denatured or elastotic matrix (dm) where blood vessels (v) are prominent. cs, cornealstroma; e, epithelium. Original magnification: (A, B, and C), 3500; (D, E), 3640.
4144 Di Girolamo et al. IOVS, December 2000, Vol. 41, No. 13

Expression of Collagenase-1 and -3 at the LeadingPterygium Edge
Having identified the invading pterygium edge in all tissuespecimens, we sectioned whole human eyes serially and ana-lyzed them immunohistochemically to determine the expres-sion profile of MMPs and TIMPs. Initially, tissue sections werestained for the collagenases. Immunoreactivity for collage-nase-1 (MMP-1) was abundant, intensely staining blood vesselsand detected along the entire length of the pterygium, predom-inantly within the pterygium epithelium as well as in ptery-gium fibroblasts (Figs. 2A, 2B, 2C). Reactivity for this proteasewas diminished toward the central cornea (Fig. 2C, inset). In
addition, collagenase-1 was expressed by some stromal fibro-blasts and often associated with the stromal connective tissue,suggesting that it may be sequestered on specific matrix com-ponents. In contrast to collagenase-1, collagenase-3 (MMP-13)was differentially expressed within the same pterygium speci-men. This enzyme stained more intensely the basal and colum-nar epithelium, particularly in the regions of fragmented BL,adjacent to pterygium fibroblasts, which themselves containedsome immunoreactivity (Figs. 2D, 2E, 2F). Collagenase-3 wasalso localized to vascular endothelial cells (Fig. 2D, v) and someinflammatory cells (micrographs not shown). Faint stainingwas detected in stromal fibroblasts, none was matrix associ-
FIGURE 2. Immunohistochemical localization of collagenases and gelatinases in the invading pterygia. Pterygium tissue was serially sectioned andstained for collagenase-1 (A, B, and C), collagenase-3 (D, E, and F), gelatinase A (G, H, and I), and gelatinase B (J, K, and L). Insets:Immunoreactivity at the central cornea with the respective Abs. Three consecutive photographs were taken along the length of the pterygium anda composite image compiled for each Ab. Red staining was regarded as specific immunoreactivity. Sections were counterstained with hematoxylin.Control samples included omitting the primary Ab (micrographs not shown) and incubating the tissue sections with an isotype control Ab (see Figs.3J, 3K, 3L). Similar results were obtained with all other pterygium tissue specimens analyzed. The labels and abbreviations used in this figure areidentical with those in Figure 1. Original magnification, 3500.
IOVS, December 2000, Vol. 41, No. 13 MMPs and TIMPs at the Invading Pterygium Edge 4145

ated, and minimal reactivity was detected at or near the centralcornea (Fig. 2F, inset).
Differential Expression of Gelatinases A and Bat the Pterygium Leading Edge
Gelatinase A (MMP-2) immunoreactivity was intense, particu-larly in the basal columnar epithelial cells, adjacent to regionsof denatured BL (Figs. 2G, 2H, 2I). However, only faint stainingfor this basement membrane–degrading enzyme was noted insuperficial epithelial layers and in pterygium fibroblasts, withsome reactivity in the central corneal epithelium (Fig. 2I, in-set). This enzyme was not expressed by the vascular endothe-lium or corneal stromal fibroblasts, nor was it associated withthe corneal stromal matrix (as was collagenase-1). Of note,gelatinase A staining was also found specifically associated withBL, in regions adjacent to the pterygium (Fig. 2G, arrow), butnot at the central cornea. In regions where BL was fragmented(Figs. 2G, 2H, 2I, asterisks), gelatinase A immunoreactivity wasdiffuse, and BL became indistinguishable from the stromalmatrix. Additional immunoreactivity was found within the fi-brous degenerative matrix of the pterygium body (Fig. 2G).Gelatinase B (MMP-9) staining was similar to that of gelatinaseA, most abundant in the basal epithelium. However, pterygiumfibroblasts, inflammatory cells, and the vascular endotheliumalso showed staining. In contrast to gelatinase A, gelatinase Bwas not found associated with BL or in the fibrous connectivetissue, nor was it detected in stromal fibroblasts (Figs. 2J, 2K,2L). Rarely was this proteinase detected at the central cornea.When it was, however, the intensity of staining was usually lessthan in the pterygium (Fig. 2L, inset).
Expressions of TIMPs 1 to 3 at the PterygiumLeading Edge
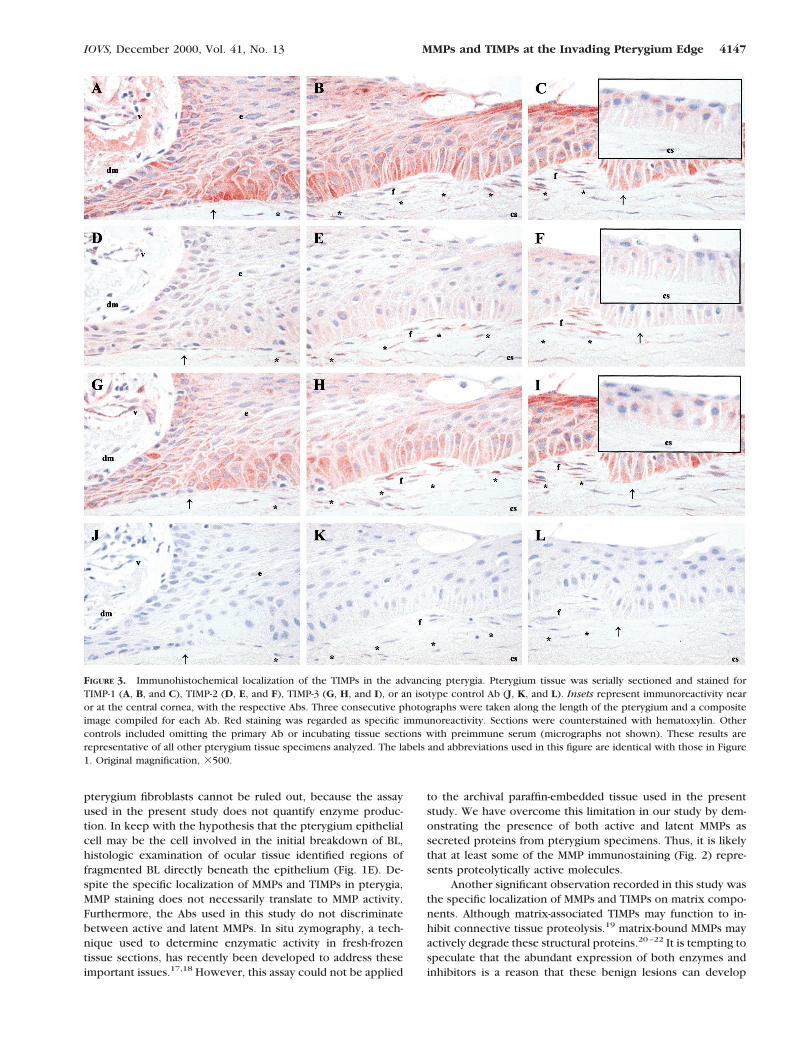
TIMP-1 expression closely resembled that of collagenase-1, inthat all pterygium epithelial cells contained some immunore-activity for this inhibitor. Similarly, the intensity of staining wasgreatest in the more basal epithelium, particularly in the vicin-ity of the denatured BL. Additional TIMP-1 signal was associ-ated with pterygium fibroblasts and fibrous bundles of thepterygium body and was faintly detected in some stromalfibroblasts (Figs. 3A, 3B, 3C). Weak epithelial cell reactivity wasdemonstrated at the central cornea (Fig. 3C, inset). TIMP-2reactivity was weak or undetectable in the pterygium epithe-lium, corneal stromal fibroblasts, and vascular endothelium butwas identified on pterygium fibroblasts (Fig. 3D, 3E, 3F) in 8 of11 specimens. TIMP-3 staining paralleled that of TIMP-1, de-tected in most pterygium epithelium, and diminished in inten-sity from basal to superficial and from pterygium to cornealepithelium. Pterygium fibroblasts were distinctly immunoreac-tive, as were vascular endothelial cells and inflammatory cells(Fig. 3G, 3F, 3I). Regions of specific connective tissue stainingby TIMP-3 were also observed in most specimens (micrographnot shown).4 This pattern of MMP and TIMP staining wasgenerally observed with all tissue specimens analyzed.
Secretion of Latent and Active MMPsfrom Pterygium Tissue
Of the four MMPs immunolocalized at the advancing pterygiumedge (Fig. 2), latent and active forms of three of these enzymeswere detected by gelatin zymography (Fig. 4). Progelatinase B(92 kDa) and progelatinase A (72 kDa) were detected in all four
pterygium tissue specimens. In addition, the active form ofeach gelatinase was displayed as a lower molecular weightspecies (gelatinase B, 83 kDa; gelatinase A, 66 kDa). Althoughother MMPs do not efficiently denature gelatin in this assaysystem, procollagenase-1 was detected as a faint doublet at 57and 54 kDa, along with its proteolytically active species whichmigrated to 45 kDa.
Gelatinase B in Tears from Eyes with Pterygia
Basal tear fluid was collected from patients with pterygia todetermine the gelatinolytic content. Two prominent gelatino-lytic activities were revealed: a 92-kDa and a 125-kDa bandcorresponding to gelatinase B and gelatinase B/TIMP complex,respectively. Densitometric analysis of these bands establishedthat on average, gelatinase B and the gelatinase B/TIMP com-plex were elevated at least 1.7- and 1.35-fold respectively ineyes with pterygia compared with the control eyes from thesame patients (Fig. 5A). Previously, we identified a similar125-kDa gelatinolytic band in aqueous humor samples frompatients with uveitis as an MMP-9/TIMP complex by Westernblot analysis.14 Collagenase-1 was not detected by this method,and gelatinase A was only faintly detected in 3 of 11 samples oftear fluid from eyes with pterygia. Although, gelatinase B andthe 125-kDa gelatinase/TIMP complex were detected in thetears of control subjects, densitometrically there was no signif-icant difference in the levels of this enzyme between the leftand right eyes of an individual (Fig. 5B).
DISCUSSION
Whereas previous studies used resected pterygium tissue4 or invitro assays of cultured pterygium epithelial cells4 and fibro-blasts,5 in the present study we used whole human eyes withattached pterygia to determine the expression of MMPs andTIMPs along the entire invading edge. Several novel observa-tions were recorded including the differential expression ofMMPs and TIMPs within the same tissue specimen along thelength of the lesion, the expression of collagenase-3, the local-ization of gelatinase A to BL, the relative contribution of MMPsand TIMPs by the pterygium fibroblast compared with thepterygium epithelial cell, the production of active and latentMMPs by pterygium tissue, and the detection of increasedlevels of gelatinase B in tears from patients with pterygia.
A characteristic feature of pterygia is the loss of BL, anatural collagenous barrier that separates the epithelium fromthe underlying stroma. The mechanism responsible for itsdestruction in pterygia is currently unknown. In the presentstudy, the overexpression of MMPs and TIMPs by pterygiumepithelial cells in regions without BL was a significant findingand suggests that these proteins may contribute to the matrixremodeling and BL destruction. The absence of BL also coin-cided with the presence of MMP- and TIMP-expressing ptery-gium fibroblasts. However, the staining intensity for any givenMMP or TIMP (with the exception of TIMP-2) was greatest inpterygium epithelial cells when compared with pterygium fi-broblasts. In a recent study, Dushku et al.16 identified theexpression of six different MMPs in altered limbal basal epithe-lial cells, whereas collagenase-1 was the only proteinase foundin pterygium fibroblasts. Taken together, these data suggestthat the epithelial cell is the more likely candidate involved inthe dissolution of BL. However, the potential involvement of
4146 Di Girolamo et al. IOVS, December 2000, Vol. 41, No. 13
pterygium fibroblasts cannot be ruled out, because the assayused in the present study does not quantify enzyme produc-tion. In keep with the hypothesis that the pterygium epithelialcell may be the cell involved in the initial breakdown of BL,histologic examination of ocular tissue identified regions offragmented BL directly beneath the epithelium (Fig. 1E). De-spite the specific localization of MMPs and TIMPs in pterygia,MMP staining does not necessarily translate to MMP activity.Furthermore, the Abs used in this study do not discriminatebetween active and latent MMPs. In situ zymography, a tech-nique used to determine enzymatic activity in fresh-frozentissue sections, has recently been developed to address theseimportant issues.17,18 However, this assay could not be applied
to the archival paraffin-embedded tissue used in the presentstudy. We have overcome this limitation in our study by dem-onstrating the presence of both active and latent MMPs assecreted proteins from pterygium specimens. Thus, it is likelythat at least some of the MMP immunostaining (Fig. 2) repre-sents proteolytically active molecules.
Another significant observation recorded in this study wasthe specific localization of MMPs and TIMPs on matrix compo-nents. Although matrix-associated TIMPs may function to in-hibit connective tissue proteolysis.19 matrix-bound MMPs mayactively degrade these structural proteins.20–22 It is tempting tospeculate that the abundant expression of both enzymes andinhibitors is a reason that these benign lesions can develop
FIGURE 3. Immunohistochemical localization of the TIMPs in the advancing pterygia. Pterygium tissue was serially sectioned and stained forTIMP-1 (A, B, and C), TIMP-2 (D, E, and F), TIMP-3 (G, H, and I), or an isotype control Ab (J, K, and L). Insets represent immunoreactivity nearor at the central cornea, with the respective Abs. Three consecutive photographs were taken along the length of the pterygium and a compositeimage compiled for each Ab. Red staining was regarded as specific immunoreactivity. Sections were counterstained with hematoxylin. Othercontrols included omitting the primary Ab or incubating tissue sections with preimmune serum (micrographs not shown). These results arerepresentative of all other pterygium tissue specimens analyzed. The labels and abbreviations used in this figure are identical with those in Figure1. Original magnification, 3500.
IOVS, December 2000, Vol. 41, No. 13 MMPs and TIMPs at the Invading Pterygium Edge 4147

over decades. In contrast, it has been reported that malignanttumors develop rapidly as a consequence of MMP overexpres-sion and TIMP downregulation.23
Of interest, staining for gelatinase A was found to beassociated with BL and in elastotic regions, whereas its struc-turally and functionally related molecule gelatinase B was not.Both gelatinases A and B contain fibronectin-like domains thatmediate their matrix-binding capacity.24 Although these pro-teinases were initially characterized based on their type IVcollagenolytic activity, data from other studies suggest thatgelatinase A acts as an interstitial collagenase, due to its abilityto cleave native type I collagen.25 In this regard, gelatinase A isa likely candidate involved in the fragmentation of BL. Perhapsit is not unreasonable to postulate that the activity of MMPs andTIMPs are essential in the development of pterygia; previousinvestigators have localized the same family of proteins indestructive corneal disorders18,26,27 and in migrating and re-generating epithelial cells in a rat model of corneal wound-ing.28
Although the principal function of TIMPs is to inhibitMMP activity,15 paradoxically, at least TIMP-2 is involved in theactivation of progelatinase A.29 TIMPs also possess growthfactor–like activity, and in this capacity they may be involved inpterygium proliferation; a recent study reported for cornealepithelial cells.30 Other functions attributed to TIMP-1 andTIMP-2 include, suppression31,32 or in the case of TIMP-3,induction of apoptosis,33 a process recently described in ptery-gia.34 TIMPs have also been reported to inhibit angiogene-sis,32,35 a function unlikely to be applicable to pterygia, be-cause these lesions characteristically contain an extensivevascular supply.4,36
The data presented in this investigation do not excludethe possible involvement of UV irradiation1,3,37,38 in the patho-genesis of pterygia. The increased expression of MMPs andTIMPs by basal pterygium epithelial cells (Figs. 2, 3) may beexplained by the peripheral light–focusing effect,1,37,38 whichpredicts that peripheral light entering the eye laterally has afocusing effect of approximately 20-fold at the limbus (theusual site of pterygia). Therefore, the basal epithelial cells maybe activated as they are struck from behind by the focusedlight.
Although other investigators have identified gelatinase Bin tears derived from patients with corneal graft failure39 andocular rosacea,40 this is the first study to document gelatinaseB levels in tears from patients with pterygia (Fig. 5). Further-more, levels of this protease were greater (1.7-fold) in eyeswith pterygia than in the contralateral control eyes, perhapsindicative of an abnormality on the ocular surface. Despite thedetection of gelatinase B and an MMP/TIMP complex, no col-lagenase-1 and no active MMPs were observed, whereas gela-tinase A was detected in only three tear samples.
The recent development of synthetic inhibitors of MMPactivity has provided a potentially new therapeutic strategy forthe treatment of cancer, which may be applicable to pterygia.The same agents have been applied to the eye to preventcorneal ulceration and reduce retinal neovascularization41,42 oradministered subcutaneously to prevent experimental autoim-mune uveoretinitis.43 Anti-MMP strategies, in conjunction withsurgical techniques, may be of future therapeutic use to reducethe rate of recurrence, severity of inflammation, tissue inva-sion, proliferation, and angiogenesis associated with pterygia.
Acknowledgments
The authors thank Tina Liakos for her assistance in the collection oftear samples.
References
1. Coroneo MT, Di Girolamo N, Wakefield D. The pathogenesis ofpterygia. Curr Opin Ophthalmol. 1999;10:282–288.
2. Dushku N, Reid TW. Immunohistochemical evidence that humanpterygia originate from an invasion of vimentin-expressing alteredlimbal epithelial basal cells. Curr Eye Res. 1994;13:473–481.
FIGURE 4. Matrix metalloproteinase activity in pterygia. Fresh ptery-gia was obtained from surgery and immediately placed in organ culturefor 48 hours. Supernatants from four different pterygium tissue spec-imens (patients 12–15, see Table 1) were displayed by gelatin-substratezymography (lanes 2 through 5). A molecular weight protein ladderwas run in parallel (lane 1). All samples demonstrated a similar profileof MMP activity, including the presence of pro- and active gelatinase B,gelatinase A, and collagenase-1.
FIGURE 5. Gelatinolytic activity in tears derived from patients withpterygia. (A) Tear samples from four patients with pterygia weredisplayed by zymography. Equal volume of tears from eyes with ptery-gia (lanes 2, 4, 6, and 8) and the corresponding control eyes (lanes 1,3, 5, and 7) were loaded. Lanes 1 and 2: patient DJ (1.5 ml of tears);lanes 3 and 4: patient SD (1.5 ml of tears); lanes 5 and 6: patient NM(3.0 ml of tears); lanes 7 and 8: patient SC (3.5 ml of tears). Conditionedmedia from PMA stimulated pterygium epithelial cells served as apositive control (lane 9). This gelatinolytic profile is representative ofthe tear fluid from all patients studied. (B) Tear samples from fourcontrol subjects were displayed by zymography. Equal volume of tearsfrom left eyes (lanes 1, 3, 5, and 7) and right eyes (lanes 2, 4, 6, and8) were loaded. Lanes 1 and 2: subject 1 (1.0 ml of tears); lanes 3 and4: subject 2 (1.0 ml of tears); lanes 5 and 6: subject 3 (2.5 ml of tears);lanes 7 and 8: subject 4 (2.7 ml of tears). The same gelatinolytic profilewas observed with the other four control subjects.
4148 Di Girolamo et al. IOVS, December 2000, Vol. 41, No. 13

3. Cameron ME. Geographic distribution of pterygia. Trans Ophthal-mol Soc Aust. 1962;22:67–81.
4. Di Girolamo N, McCluskey PJ, Lloyd A, Coroneo MT, Wakefield D.Expression of MMPs and TIMPs in human pterygia and culturedpterygium epithelial cells. Invest Ophthalmol Vis Sci. 2000;41:671–679.
5. Lee S-B, Li D-Q, Gunja–Smith Z, Liu YQ, Tan DTH, Tseng SCG.Increased expression and activity of MMP-1 and MMP-3 by culturedpterygium head fibroblasts [ARVO Abstract]. Invest OphthalmolVis Sci. 1999;40(4):S334. Abstract nr 1768.
6. Kria L, Ohira A, Amemiya T. Immunohistochemical localization ofbasic fibroblast growth factor, platelet derived growth factor,transforming growth factor-b and tumor necrosis factor-a in thepterygium. Acta Histochem. 1996;98:195–201.
7. Massova I, Kotra LP, Fridman R, Mobashery S. Matrixmetalloproteinases: structures, evolution, and diversification.FASEB J. 1998;12:1075–1095.
8. Stetler–Stevenson WG. Matrix metalloproteinases in angiogenesis:a moving target for therapeutic intervention. J Clin Invest. 1999;103:1237–1241.
9. Ray JM, Stetler–Stevenson WG. The role of matrix metalloprotein-ases and their inhibitors in tumour invasion, metastasis and angio-genesis. Eur Respir J. 1994;7:2062–2072.
10. Stetler–Stevenson WG. Dynamics of matrix turnover during patho-logic remodeling of the extracellular matrix. Am J Pathol. 1996;148:1345–1350.
11. Lampert K, Machein U, Regina M, Conca, W, Peter HH, Volk B.Expression of matrix metalloproteinases and their tissue inhibitorsin human brain tumors. Am J Pathol. 1998;153:429–437.
12. Firestein GS, Paine MM. Stromelysin and tissue inhibitor of metal-loproteinases gene expression in rheumatoid arthritis synovium.Am J Pathol. 1992;140:1309–1314.
13. Di Girolamo N, Lloyd A, McCluskey PJ, Filipic M, Wakefield D.Increased expression of matrix metalloproteinases in vivo in scle-ritis tissue and in vitro in cultured human scleral fibroblasts. Am JPathol. 1997;105:653–666.
14. Di Girolamo N, Verma MJ, McCluskey PJ, Lloyd A, Wakefield D.Increased matrix metalloproteinases in the aqueous humor ofpatients and experimental animals with uveitis. Curr Eye Res.1996;15:1060–1068.
15. Gomez DE, Alonso DF, Yoshiji H, Thorgeirsson UP. Tissue inhibi-tors of metalloproteinases: structure, regulation and biologicalfunctions. Eur J Cell Biol. 1997;74:111–122.
16. Dushku N, John MK, Schultz GS, Reid TW. Pterygia pathogenesis:corneal invasion by matrix metalloproteinase (MMP) expressingaltered limbal basal stem cells and activation of fibroblasts [ARVOAbstract]. Invest Ophthalmol Vis Sci. 2000;41(4):S451. Abstract nr2388.
17. Galis ZS, Sukhova GK, Libby P. Microscopic localization of activeproteases by in situ zymography: detection of matrix metallopro-teinase activity in vascular tissue. FASEB J. 1995;9:974–980.
18. Zhou L, Sawaguchi S, Twining SS, Sugar J, Feder RS, Yue BYJT.Expression of degradative enzymes and protease inhibitors incorneas with keratoconus. Invest Ophthalmol Vis Sci. 1998;39:1117–1124.
19. Fariss RN, Apte SS, Olsen BR, Iwata K, Milam AH. Tissue inhibitorof metalloproteinase-3 is a component of Bruch’s membrane of theeye. Am J Pathol. 1997;150:323–328.
20. Dahlen B, Shute J, Howarth P. Immunohistochemical localisationof matrix metalloproteinases MMP-3 and MMP-9 within the airwaysin asthma. Thorax. 1999;54:590–596.
21. Allen JA, Docherty AJP, Barker PJ, Huskisson NS, Reynolds JJ,Murphy G. Binding of gelatinases A and B to type-1 collagen andother matrix components. Biochem J. 1995;309:299–306.
22. Davis V, Persidskaia R, Regen–Baca L, et al. Matrix metalloprotein-ase-2 production and its binding to the matrix are increased inabdominal aortic aneurysms. Arterioscler Thromb Vasc Biol. 1998;18:1625–1633.
23. Ara T, Kusafuka T, Inoue M, Kuroda S, Fukuzawa M, Okada A.Determination of imbalance between MMP-2 and TIMP-2 in humanneuroblastoma by reverse-transcription polymerase chain reactionand its correlation with tumor progression. J Pediatr Surg. 2000;35:432–437.
24. Shipley JM, Doyle GA, Fliszar CJ, et al. The structural basis for theelastolytic activity of the 92-kDa and 72-kDa gelatinases: role of thefibronectin type II-like repeats. J Biol Chem. 1996;271:4335–4341.
25. Aimes RT, Quigley JP. Matrix metalloproteinase-2 is an interstitialcollagenase. J Biol Chem. 1995;270:5872–5876.
26. Kenney MC, Chwa M, Alba A, Saghizadeh M, Huang Z-S, Brown DJ.Localization of TIMP-1, TIMP-2, TIMP-3, gelatinase A and gelatinaseB in pathological human corneas. Curr Eye Res. 1998;17:238–246.
27. Brown D, Chwa M, Escobar M, Kenney MC. Characterization of themajor matrix degrading metalloproteinase of human cornealstroma: evidence for an enzyme/inhibitor complex. Exp Eye Res.1991;52:5–16.
28. Ye HQ, Azar DT. Expression of gelatinases A and B, and TIMPs 1and 2 during corneal wound healing. Invest Ophthalmol Vis Sci.1998;39:913–921.
29. Strongin AY, Collier I, Bannikov G, Marmer BL, Grant GA, Gold-berg GI. Mechanism of cell surface activation of 72-kDa type IVcollagenase. J Biol Chem. 1995;270:5331–5338.
30. Saika S, Kawashima Y, Okada Y, et al. Recombinant TIMP-1 and -2enhance the proliferation of rabbit corneal epithelial cells in vitroand the spreading of rabbit corneal epithelium in situ. Curr EyeRes. 1998;17:47–52.
31. Guedez L, Stetler–Stevenson WG, Wolff L, et al. In vitro suppres-sion of programmed cell death of B cells by tissue inhibitor ofmetalloproteinase-1. J Clin Invest. 1998;102:2002–2010.
32. Valente P, Fassina G, Melchiori A, et al. TIMP-2 over-expressionreduces invasion and angiogenesis and protects B16F10 melanomacells from apoptosis. Int J Cancer. 1998;75:246–253.
33. Baker AH, Zaltsman AB, George SJ, Newby AC. Divergent effects oftissue inhibitor of metalloproteinase-1, -2, or -3 overexpression onrat vascular smooth muscle cell invasion, proliferation, and deathin vitro. J Clin Invest. 1998;101:1478–1487.
34. Tan DTH, Tang WY, Liu YP, Goh H-S, Smith DR. Apoptosis andapoptosis related gene expression in normal conjunctiva andpterygium. Br J Ophthalmol. 2000;84:212–216.
35. Anand–Apte B, Pepper MS, Voest E, et al. Inhibition of angiogen-esis by tissue inhibitor of metalloproteinase-3. Invest OphthalmolVis Sci. 1997;38:817–823.
36. Seifert P, Sekundo W. Capillaries in the epithelium of pterygium.Br J Ophthalmol. 1998;82:77–81.
37. Coroneo MT, Muller–Stolzenburg NW, Ho A. Peripheral light fo-cusing by the anterior eye and the ophthalmohelioses. Ophthal-mic Surg. 1991;22:705–711.
38. Kwok LS, Coroneo MT. A model for pterygium formation. Cornea.1994;13:219–224.
39. Barro CD, Romanet J-P, Morel F. Gelatinase concentration in tearsof corneal-grafted patients. Curr Eye Res. 1998;17:174–182.
40. Afonso AA, Sobrin L, Monroy DC, Selzer M, Lokeshwar B, Pflug-felder SC. Tear fluid gelatinase B activity correlates with IL-1aconcentration and fluorescein clearance in ocular rosacea. InvestOphthalmol Vis Sci. 1999;40:2506–2512.
41. Schultz GS, Strelow S, Stern GA, et al. Treatment of alkali-injuredrabbit corneas with a synthetic inhibitor of matrix metalloprotein-ases. Invest Ophthalmol Vis Sci. 1992;33:3325–3331.
42. Das A, McLamore A, Song W, McGuire PG. Retinal neovasculariza-tion is suppressed with a matrix metalloproteinase inhibitor. ArchOphthalmol. 1999;117:498–503.
43. Wallace GR, Whiston RA, Stanford MR, Wells GMA, Gearing AJH,Clements JM. The matrix metalloproteinase inhibitor BB-1101 pre-vents experimental autoimmune uveoretinitis (EAU). Clin ExpImmunol. 1999;118:364–370.
IOVS, December 2000, Vol. 41, No. 13 MMPs and TIMPs at the Invading Pterygium Edge 4149
Copyright © 2022 FDOKUMEN