
Dynamic changes of matrix metalloproteinases and their tissue inhibitors in severe sepsis
Upload
independentCategory
view
1download
0
Expression of Matrix Metalloproteinases During MurineChorioallantoic Placenta MaturationTAMBET TEESALU,1,3 REGIS MASSON,4 PAUL BASSET,4 FRANCESCO BLASI,1,2 AND DANIELA TALARICO1,2*1 Dipartimento di Ricerca Biologica e Tecnologica (DIBIT), Istituto Scientifico S. Raffaele, Milano, Italia2Dipartimento di Genetica e Biologia dei Microorganismi, Universita di Milano, Milano, Italia3Department of Virology, Haartman Institute, Helsinki University, Helsinki, Finland4Institut de Genetique et de Biologie Moleculaire et Cellulaire, CNRS/INSERM/ULP, C.U. de Strasbourg, France
ABSTRACT A large body of experimentalevidence supports the participation of two groupsof extracellular proteases, matrix metalloprotein-ases (MMPs), and plasminogen activators/plas-min, in tissue remodeling in physiological andpathological invasion. In the late mouse placenta,several tissue remodeling and cell invasion pro-cesses take place. Spongiotrophoblast migrationinto maternal decidua, as well as decidual extra-cellular matrix remodeling require the coordi-nated action of extracellular proteolytic en-zymes. Via Northern and in situ hybridization, wehave analyzed the spatio-temporal expressionpatterns of members of the MMP family (strome-lysin-3, gelatinases A and B), as well as theirinhibitors TIMP-1, -2 and -3 in late murine pla-centa (days 10.5 to 18.5 of gestation). Gelatinaseactivity in placental extracts was assessed bysubstrate zymography. GelatinaseAand stromely-sin-3 were found to be prominently expressed indecidual tissue; shortly after midpregnancy, thedecidual expression patterns of gelatinase A andstromelysin-3 became overlapping with eachother, as well as with the expression domain ofTIMP-2. On the other hand, gelatinase B tran-scripts were expressed only by trophoblast giantcells at day 10.5, and were downregulated at laterstages. TIMP-1 and TIMP-3 transcripts were de-tected in decidual periphery at day 10.5, whilelater the expression was restricted to the endome-trial stroma and spongiotrophoblasts, respec-tively. The areas of stromelysin-3 expression werethe same (giant trophoblasts) or adjacent (de-cidua) to those where urokinase (uPA) tran-scripts were detected, suggesting a possible coop-eration between these proteinases in placentalremodeling.
We generated mice doubly deficient for strome-lysin-3 and uPA, and report here that these miceare viable and fertile. Furthermore, these ani-mals do not manifest obvious placental abnormali-ties, thereby suggesting the existence of compen-satory/redundant mechanisms involving otherproteolytic enzymes.
Our findings document the participation ofMMPs and their inhibitors in the process of late
murine placenta maturation, and warrant the char-acterization of other members of the MMP family,like membrane type-MMPs, in this process. DevDyn 1999; 214:248–258. r 1999 Wiley-Liss, Inc.
Key words: mouse; placenta; trophoblast; de-cidua; tissue remodeling; urokinase;stromelysin-3; gelatinase; TIMP
INTRODUCTION
Precisely controlled remodeling of extracellular ma-trix (ECM) is an essential aspect of normal animaldevelopment, pregnancy-induced changes in the femalereproductive system, and repair and regeneration ofdamaged tissues. ECM degradation during invasiveprocesses depends on the regulated action of extracellu-lar proteolytic enzymes (for reviews see Matrisian andHogan, 1990; Birkedal-Hansen, 1995; Werb, 1997). Twoclasses of proteinases, matrix metalloproteinases(MMPs) and plasminogen/plasminogen activators sys-tems, have been demonstrated to play major roles inECM remodeling in various physiological as well aspathological settings.
Matrix metalloproteinases are a growing group ofzinc-containing enzymes, which share a similar domainstructure; most MMPs are secreted as inactive precur-sors, which are converted into the active form by theproteolytic removal of a pro-domain; all MMPs areinhibited by naturally occurring tissue inhibitors ofmetalloproteinases (TIMPs) (Birkedal-Hansen, 1995;Jones and Walker, 1997). Gelatinases A and B (MMP-2and MMP-9) are members of the MMP family postu-lated to play a critical role in cell invasion. The associa-tion of gelatinases with cell invasion and malignancy islikely related to their ability to degrade basementmembrane collagen IV, as well as collagens V, VII andXI, fibronectin, laminin, elastin, proteoglycans, andentactin (Matrisian, 1990; Kleiner and Stetler-Steven-son, 1993; Aimes and Quigley, 1995). In contrast, littleis known about the physiological substrates of the most
*Correspondence to: Daniela Talarico, Dipartimento di RicercaBiologica e Tecnologica (DIBIT), Istituto Scientifico S. Raffaele, viaOlgettina 60, 20132 Milano, Italia. E-mail [email protected]
Received 5 July 1998; Accepted 21 December 1998
DEVELOPMENTAL DYNAMICS 214:248–258 (1999)
r 1999 WILEY-LISS, INC.

recently discovered member of the stromelysin subfam-ily, stromelysin-3. Stromelysin-3 has been found to beassociated with highly invasive tumors, as well as withprocesses of physiological tissue remodeling, and it isfrequently coexpressed with urokinase type plasmino-gen activator (uPA) (Lefebvre et al., 1992; Lefebvre etal., 1995; Basset et al., 1997).
In recent years, a number of transgenic/null mutantmice of ECM degrading proteinases have been gener-ated (reviewed by Carmeliet and Collen, 1996; Shapiro,1997; Edwards and Murphy, 1998). Transgenic miceoverexpressing proteolytic enzymes manifest alter-ations of target tissue architecture (D’Armiento et al.,1992; Sympson et al., 1995), corroborating the conceptof proteases as important players in several types ofpathology. Surprisingly, none of the genes encoding forextracellular proteases that have been inactivated, hasproven to be essential for murine embryonic develop-ment (Carmeliet and Collen, 1996; Shapiro, 1997; Itohet al., 1998; Masson et al., 1998; Vu et al., 1998). Thisfinding implies the existence of compensatory mecha-nisms and/or redundancy in the networks of extracellu-lar proteolysis. Double or multiple knockouts representa useful approach to shed further light on the interrela-tions and functional overlaps between different prote-ases during mouse development and physiology.
Mammalian uterus is a remarkably plastic organ,which during pregnancy undergoes a complex programof growth and remodeling in a short time window(Cross et al., 1994; Rinkenberger et al., 1997). Remodel-ing of the uterine ECM components, such as basallaminae and collagen fibers, is necessary to allowexpansion of the uterus to accommodate the growingembryo, as well as for the establishment of vascularand anchoring links between fetus and mother (Abraha-msohn and Zorn, 1993; Enders and Welsh, 1993).Extracellular proteolysis during mouse embryo implan-tation and early stages thereafter has been the subjectof a number of earlier studies (Sappino et al., 1989;Alexander et al., 1996; Teesalu et al., 1996; Afonso etal., 1997), while late stages of murine placental develop-ment are far less characterized. We have recentlyanalyzed the expression of urokinase type plasminogenactivator (uPA), as well as the functionally relatedmolecules uPAR, PAI-1, and a2MR/LRP, during placen-tal development (Teesalu et al., 1998).
In the current study, we have analyzed the system ofmatrix metalloproteinases in mouse placenta duringthe second half of pregnancy. We describe the spatio-temporal expression patterns of stromelysin-3, gelatin-ases A and B (MMP-2 and MMP-9) and their inhibitors(TIMP-1, TIMP-2, TIMP-3) in late (from day 10.5 ofgestation) murine placentas. In addition, we used gela-tin substrate zymography to characterize gelatinaseisoforms present in placental extracts. We found that inpostmidgestational placentas extensive decidual areas,undergoing tissue remodeling, were positive for thepresence of gelatinase A and stromelysin-3 transcripts;concomitant expression of TIMP-2 in an overlapping
pattern implies a role for this inhibitor in the regulationof activity of these MMPs. At the fetomaternal inter-face, stromelysin-3 was expressed by the uPA-positivegiant trophoblasts and by adjacent decidual cells, thussuggesting the existence of a functional cooperationbetween these two proteolytic enzymes. We report herethe generation of mice with the combined deficiency ofuPA and stromelysin-3 genes, which are however viableand fertile and show no obvious placental abnormali-ties.
RESULTSNorthern Blot Analysis of MMP/TIMPExpression
We first analyzed the expression of gelatinase A,gelatinase B, and stromelysin-3, as well as of theinhibitors TIMP-1, TIMP-2, and TIMP-3 in mouseplacentas by Northern blot analysis. To evaluate therelative changes in the level of expression of thedifferent transcripts, total RNA from whole placentas ofdays 10.5 to 18.5 of gestation was analyzed. Thisanalysis also allowed us to confirm the specificity of thecDNA probes, which were subsequently used for in situhybridization analysis.
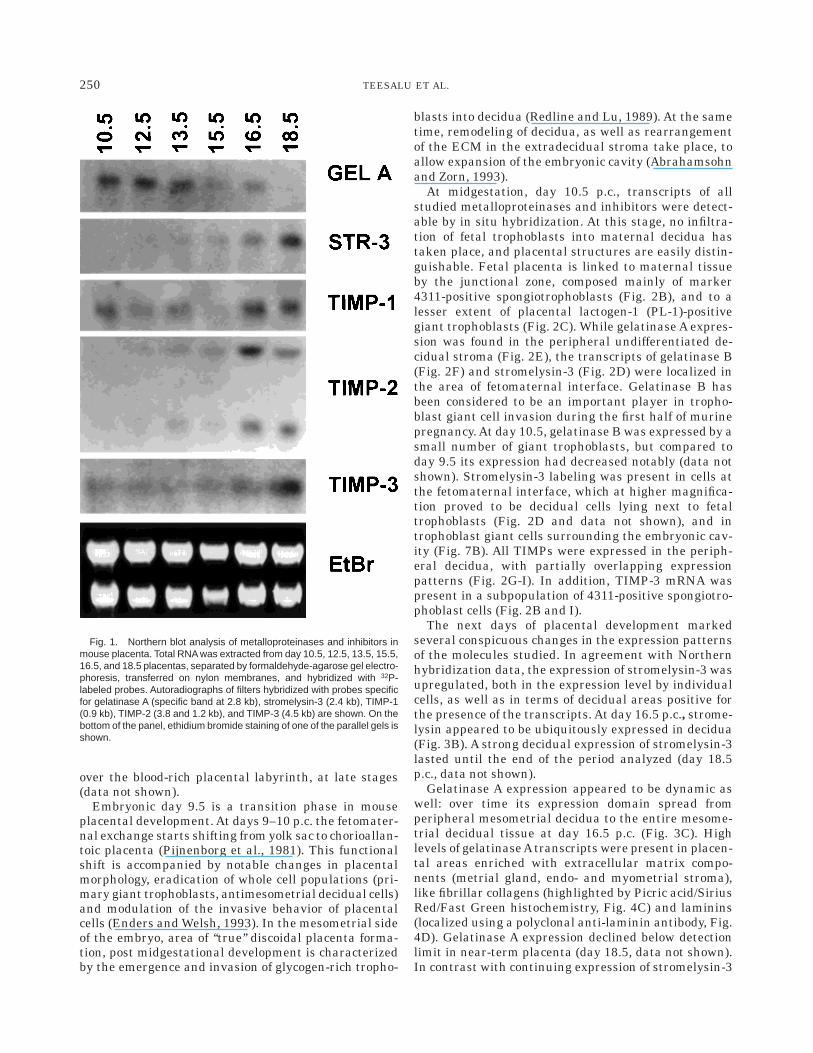
Gelatinase A transcripts were expressed at higherlevels at midgestation stages and declined in termplacenta (Fig. 1). On the other hand, no gelatinase Btranscripts could be detected at any of the stagesstudied (data not shown). Very low amounts of strome-lysin-3 transcripts were present in midgestation mu-rine placenta, while their level increased significantlyat later stages, and peaked at day 18.5 postcoitum(p.c.).
TIMP-1 transcripts were detected throughout thestages analyzed; however, there was no clear trend intheir levels during placental maturation. On the otherhand, the level of TIMP-2 transcripts, which wasrelatively low until day 15.5 p.c., was upregulated inday 16.5 p.c. placenta, and remained high at day 18.5.Also the expression of TIMP-3 transcripts increasedthroughout the period analyzed. Our data on TIMP-2and -3 expression are in agreement with the results ofthe study of Blavier and Declerck (1997). However,these authors were unable to detect TIMP-1 transcriptsin mouse placenta. We suggest that this is due toTIMP-1 expression being gradually excluded from thetrue placenta into extradecidual stroma (see next para-graph). In our preparation, this stroma was included,while in the experiments of Blavier and Declerck it wasdissected from the placental disk.
In Situ Analysis of MMP/TIMP Expression
We used RNA in situ hybridization analysis to deter-mine the expression patterns of gelatinases A and B,stromelysin-3, as well as the inhibitors TIMP-1, TIMP-2,and TIMP-3, on serial sections of day 9.5 to 18.5 mouseplacentas. As a control for the specificity of the signalwe used hybridization with sense riboprobes, whichshowed no signal, except for a weak unspecific labeling
249MMPs AND PLACENTA
over the blood-rich placental labyrinth, at late stages(data not shown).
Embryonic day 9.5 is a transition phase in mouseplacental development. At days 9–10 p.c. the fetomater-nal exchange starts shifting from yolk sac to chorioallan-toic placenta (Pijnenborg et al., 1981). This functionalshift is accompanied by notable changes in placentalmorphology, eradication of whole cell populations (pri-mary giant trophoblasts, antimesometrial decidual cells)and modulation of the invasive behavior of placentalcells (Enders and Welsh, 1993). In the mesometrial sideof the embryo, area of ‘‘true’’ discoidal placenta forma-tion, post midgestational development is characterizedby the emergence and invasion of glycogen-rich tropho-
blasts into decidua (Redline and Lu, 1989). At the sametime, remodeling of decidua, as well as rearrangementof the ECM in the extradecidual stroma take place, toallow expansion of the embryonic cavity (Abrahamsohnand Zorn, 1993).
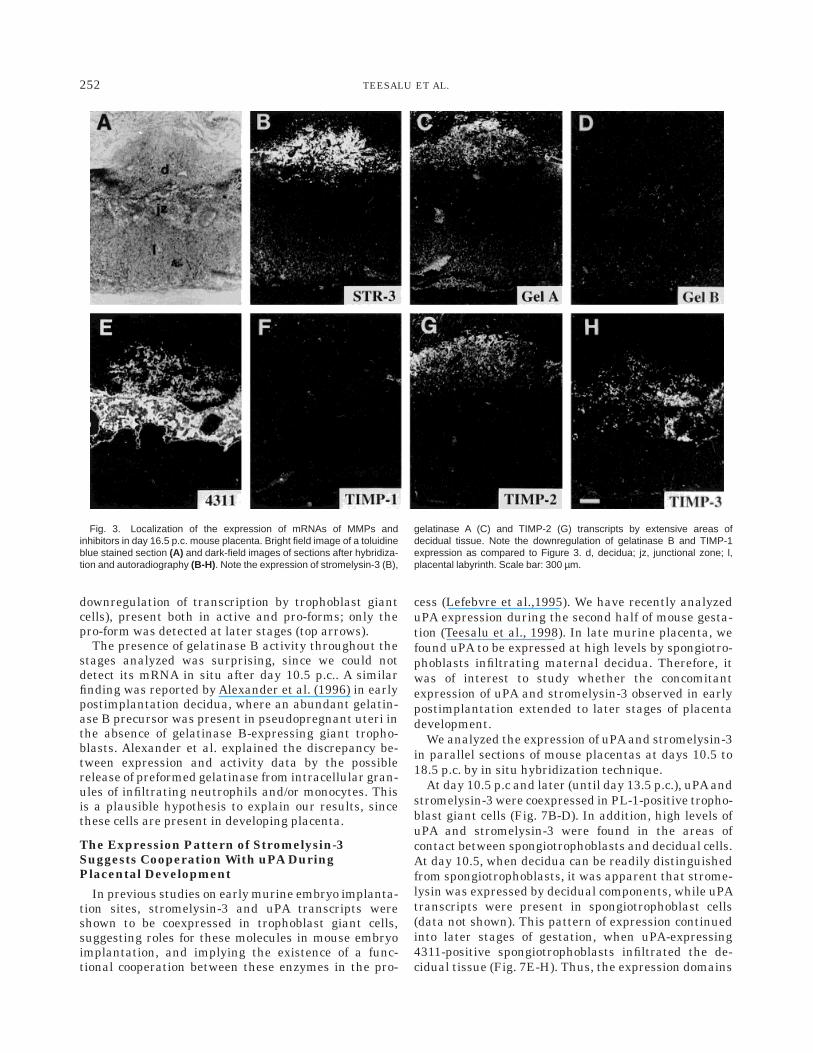
At midgestation, day 10.5 p.c., transcripts of allstudied metalloproteinases and inhibitors were detect-able by in situ hybridization. At this stage, no infiltra-tion of fetal trophoblasts into maternal decidua hastaken place, and placental structures are easily distin-guishable. Fetal placenta is linked to maternal tissueby the junctional zone, composed mainly of marker4311-positive spongiotrophoblasts (Fig. 2B), and to alesser extent of placental lactogen-1 (PL-1)-positivegiant trophoblasts (Fig. 2C). While gelatinase A expres-sion was found in the peripheral undifferentiated de-cidual stroma (Fig. 2E), the transcripts of gelatinase B(Fig. 2F) and stromelysin-3 (Fig. 2D) were localized inthe area of fetomaternal interface. Gelatinase B hasbeen considered to be an important player in tropho-blast giant cell invasion during the first half of murinepregnancy. At day 10.5, gelatinase B was expressed by asmall number of giant trophoblasts, but compared today 9.5 its expression had decreased notably (data notshown). Stromelysin-3 labeling was present in cells atthe fetomaternal interface, which at higher magnifica-tion proved to be decidual cells lying next to fetaltrophoblasts (Fig. 2D and data not shown), and introphoblast giant cells surrounding the embryonic cav-ity (Fig. 7B). All TIMPs were expressed in the periph-eral decidua, with partially overlapping expressionpatterns (Fig. 2G-I). In addition, TIMP-3 mRNA waspresent in a subpopulation of 4311-positive spongiotro-phoblast cells (Fig. 2B and I).
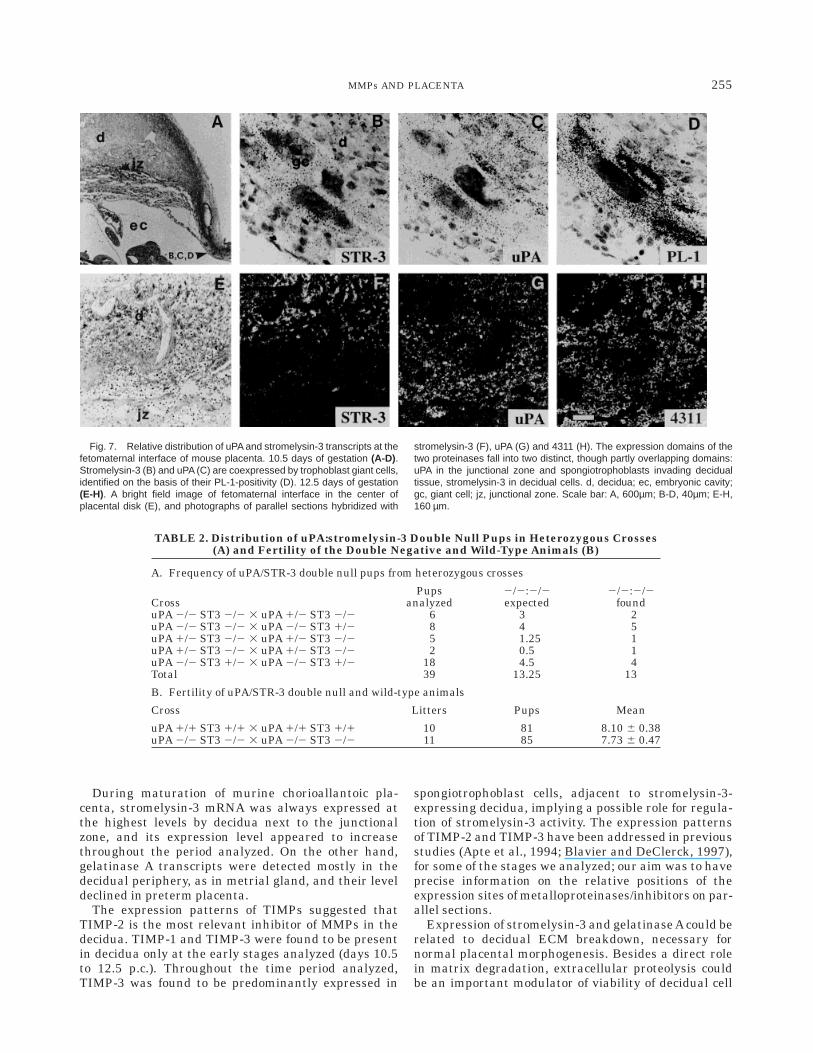
The next days of placental development markedseveral conspicuous changes in the expression patternsof the molecules studied. In agreement with Northernhybridization data, the expression of stromelysin-3 wasupregulated, both in the expression level by individualcells, as well as in terms of decidual areas positive forthe presence of the transcripts. At day 16.5 p.c., strome-lysin appeared to be ubiquitously expressed in decidua(Fig. 3B). A strong decidual expression of stromelysin-3lasted until the end of the period analyzed (day 18.5p.c., data not shown).
Gelatinase A expression appeared to be dynamic aswell: over time its expression domain spread fromperipheral mesometrial decidua to the entire mesome-trial decidual tissue at day 16.5 p.c. (Fig. 3C). Highlevels of gelatinaseAtranscripts were present in placen-tal areas enriched with extracellular matrix compo-nents (metrial gland, endo- and myometrial stroma),like fibrillar collagens (highlighted by Picric acid/SiriusRed/Fast Green histochemistry, Fig. 4C) and laminins(localized using a polyclonal anti-laminin antibody, Fig.4D). Gelatinase A expression declined below detectionlimit in near-term placenta (day 18.5, data not shown).In contrast with continuing expression of stromelysin-3
Fig. 1. Northern blot analysis of metalloproteinases and inhibitors inmouse placenta. Total RNA was extracted from day 10.5, 12.5, 13.5, 15.5,16.5, and 18.5 placentas, separated by formaldehyde-agarose gel electro-phoresis, transferred on nylon membranes, and hybridized with 32P-labeled probes. Autoradiographs of filters hybridized with probes specificfor gelatinase A (specific band at 2.8 kb), stromelysin-3 (2.4 kb), TIMP-1(0.9 kb), TIMP-2 (3.8 and 1.2 kb), and TIMP-3 (4.5 kb) are shown. On thebottom of the panel, ethidium bromide staining of one of the parallel gels isshown.
250 TEESALU ET AL.

and gelatinase A in postmidgestation stages, gelatinaseB was grossly down-regulated in placenta after day10.5, with no transcripts detectable from day 11.5 p.c.on (data not shown).
The spatial expression of the TIMPs during theprocess of placental maturation appeared to be alsoprecisely regulated. The expression of TIMP-1 mRNA,which was present in the peripheral decidua at day 10.5p.c., appeared to be excluded from maternal deciduaduring later stages (with no positive cells remaining atday 13.5 p.c.), and became restricted to subregions ofthe stroma surrounding some mesometrial blood ves-sels (data not shown). TIMP-2 was expressed by de-cidua throughout the stages analyzed, initially in pe-riphery, later spreading over more extensive areas (Fig.3G). At day 17.5 p.c. its expression also appeared in thejunctional zone spongiotrophoblasts (data not shown),as observed previously (Blavier and DeClerck, 1997).TIMP-3 was at the early stages (days 10.5–13.5 p.c.)expressed by both decidual tissue and trophoblast cells.With the progression of pregnancy, decidual expressionof TIMP-3 declined. At the same time, its expression bythe cells of spongiotrophoblast lineage was upregulated(Fig. 3H), as observed previously by Apte et al. (1994).
Summarizing, the in situ hybridization data show afine regulation of MMPs and inhibitors in postmidgesta-tion murine placenta (as summarized in Table 1 andFig. 5). During late murine placentation, gelatinase Aand stromelysin-3 are expressed in the regions offetomaternal interface undergoing remodeling. Theproducts of these genes appear to be operating mainlyin maternal decidua, where their degradative activitiescan be controlled by the inhibitor TIMP-2 and, for theearlier stages (days 10.5–12.5 p.c.), also by TIMP-1 andTIMP-3.
Zymographic Analysis of Gelatinaseson SDS-PAGE
The production of gelatinases at days 9.5–18.5 p.c.was evaluated by gelatin zymography of extracts ofwhole placentas (placental disk and adjacent myo- andendometrial components) (Fig. 6). As a positive controlfor gelatinases we used conditioned medium of HT1080human fibrosarcoma cells (Sato et al., 1994).
The pro-forms of both gelatinases were detectedthroughout the stages analyzed. Gelatinase A wasalways present only in pro-form (lower arrows), whilegelatinase B was initially (day 9.5 p.c., before gross
Fig. 2. Localization of the expression of mRNAs of MMPs andinhibitors in day 10.5 p.c. mouse placenta. Bright field image showingplacenta morphology (A), and dark-field images showing the expressionpatterns of markers of spongiotrophoblasts (4311, B) and giant tropho-
blasts (PL-1, C) as well as the expression patterns of stromelysin-3 (D),gelatinase A (E), gelatinase B (F), and inhibitors TIMP-1 (G), TIMP-2 (H),and TIMP-3 (I). db, decidua basalis; jz, junctional zone. Scale barcorresponds to 650 µm.
251MMPs AND PLACENTA
downregulation of transcription by trophoblast giantcells), present both in active and pro-forms; only thepro-form was detected at later stages (top arrows).
The presence of gelatinase B activity throughout thestages analyzed was surprising, since we could notdetect its mRNA in situ after day 10.5 p.c.. A similarfinding was reported by Alexander et al. (1996) in earlypostimplantation decidua, where an abundant gelatin-ase B precursor was present in pseudopregnant uteri inthe absence of gelatinase B-expressing giant tropho-blasts. Alexander et al. explained the discrepancy be-tween expression and activity data by the possiblerelease of preformed gelatinase from intracellular gran-ules of infiltrating neutrophils and/or monocytes. Thisis a plausible hypothesis to explain our results, sincethese cells are present in developing placenta.
The Expression Pattern of Stromelysin-3Suggests Cooperation With uPA DuringPlacental Development
In previous studies on early murine embryo implanta-tion sites, stromelysin-3 and uPA transcripts wereshown to be coexpressed in trophoblast giant cells,suggesting roles for these molecules in mouse embryoimplantation, and implying the existence of a func-tional cooperation between these enzymes in the pro-
cess (Lefebvre et al.,1995). We have recently analyzeduPA expression during the second half of mouse gesta-tion (Teesalu et al., 1998). In late murine placenta, wefound uPA to be expressed at high levels by spongiotro-phoblasts infiltrating maternal decidua. Therefore, itwas of interest to study whether the concomitantexpression of uPA and stromelysin-3 observed in earlypostimplantation extended to later stages of placentadevelopment.
We analyzed the expression of uPA and stromelysin-3in parallel sections of mouse placentas at days 10.5 to18.5 p.c. by in situ hybridization technique.
At day 10.5 p.c and later (until day 13.5 p.c.), uPA andstromelysin-3 were coexpressed in PL-1-positive tropho-blast giant cells (Fig. 7B-D). In addition, high levels ofuPA and stromelysin-3 were found in the areas ofcontact between spongiotrophoblasts and decidual cells.At day 10.5, when decidua can be readily distinguishedfrom spongiotrophoblasts, it was apparent that strome-lysin was expressed by decidual components, while uPAtranscripts were present in spongiotrophoblast cells(data not shown). This pattern of expression continuedinto later stages of gestation, when uPA-expressing4311-positive spongiotrophoblasts infiltrated the de-cidual tissue (Fig. 7E-H). Thus, the expression domains
Fig. 3. Localization of the expression of mRNAs of MMPs andinhibitors in day 16.5 p.c. mouse placenta. Bright field image of a toluidineblue stained section (A) and dark-field images of sections after hybridiza-tion and autoradiography (B-H). Note the expression of stromelysin-3 (B),
gelatinase A (C) and TIMP-2 (G) transcripts by extensive areas ofdecidual tissue. Note the downregulation of gelatinase B and TIMP-1expression as compared to Figure 3. d, decidua; jz, junctional zone; l,placental labyrinth. Scale bar: 300 µm.
252 TEESALU ET AL.

Fig. 4. Gelatinase A gene is actively transcribed by cells in the regionsof the placental disk which are relatively enriched in ECM. Brightfieldimage of a Toluidine Blue counterstained gelatinase A-hybridized section(A), and darkfield image of the same section (B). Brightfield microphoto-graph of adjacent section stained with picric acid/Sirius Red/Fast Green
(C); collagen fibers appear red. Adjacent section treated for lamininimmunohistochemistry (D); laminin appears as a brown DAB precipitate.Note that high levels of gelatinase A transcripts are found in the areaswhich contain elevated amounts of collagen/laminin (arrows). d, decidua;mg, metrial gland; ue, uterine epithelium. Scale bar: 60µm
TABLE 1. Summary of the Expression of MMPs and TIMPs Analyzedin Postmidgestation Mouse Placentaa
Gene 10.5 days p.c. 16.5 days p.c. 18.5 days p.c.Gelatinase A 111 11 n.d.
Peripheral decidua Decidua
Gelatinase B 1 n.d. n.d.Giant trophoblasts
Stromelysin-3 11 111 111Decidua at fetomaternal
interfaceDecidua Decidua
Giant trophoblasts
TIMP-1 11 12 12Peripheral decidua Extradecidual stroma Extradecidual stroma
TIMP-2 11 11 1Peripheral decidua Decidua Decidua
Spongiotrophoblasts
TIMP-3 11 11 11Peripheral deciduaSpongiotrophoblasts Spongiotrophoblasts Spongiotrophoblasts
aExpression levels are indicated by 1 to 111; n.d.: not detectable.
253MMPs AND PLACENTA

of the two proteases in this area appeared to becomplementary rather than overlapping.
Generation of uPA:stromelysin-3Double Null Mice
We bred uPA null (Carmeliet et al., 1994) and strome-lysin-3 null mice (Masson et al., 1998) in order to
generate animals with a combined deficiency of the twoproteases. The distribution of homozygous double nega-tive pups followed the pattern expected on the basis ofnormal Mendelian inheritance. Double null animalswere viable, and fertile (Table 2).
Analysis of embryo implantation sites/placentas atstages 7.5, 8.5, 9.5, 12.5, 13.5 and 18.5 p.c., did notreveal apparent histological abnormalities (data notshown). Moreover, no significant differences were ob-served in trophoblast invasion (as judged by immunohis-tochemistry with an anti-cytokeratin polyclonal anti-body) or distribution of laminins (as demonstrated byanti-laminin polyclonal antibody) (data not shown).
DISCUSSION
In the current study, we have analyzed the spatio-temporal expression patterns of the matrix metallopro-teinases stromelysin-3, gelatinases A and B, and theinhibitors TIMP-1, -2, and -3, in late murine placenta.Our results show a precise regulation of the protein-ases, as well as inhibitors, in the area of fetomaternalinterface. These data suggest that the function of theMMP system in mouse placenta formation could extendwell beyond the first half of the pregnancy.
We found that gelatinase A and stromelysin-3 tran-scripts are expressed throughout the stages of latepregnancy analyzed (days 10.5–18.5 p.c.). Gelatinase B,considered to be important for embryo implantationand early trophoblast invasion (Alexander et al., 1996),was expressed only during the first stage studied ingiant trophoblast cells, and appeared to be present inthe active form.
Fig. 6. Expression of gelatinolytic activities during mouse placentamaturation. Lysates of whole placentas of the indicated stages ofgestation were analysed by gelatin zymography as described in Materialsand Methods. As a positive control for gelatinases, conditioned medium ofHT-1080 cells was used. Because of the activation of gelatinases in theSDS sample buffer, both active and precursor gelatinases can bedetected by gelatin zymography. The doublets of arrows point at activitiesof gelatinase A (lower doublet) and B (upper doublet). In each pair, theupper arrow indicates the proform of gelatinase and the lower arrowindicates cleaved, active proteinase. Note the presence of stable amountof precursor forms of both gelatinase A and B throughout the stagesanalysed and the disappearance of active gelatinase B after day 9.5.
Fig. 5. Schematic representation of the central region of placental disk(excluding endo- and myometrium), and relative positions of the expres-sion domains of gelatinases, stromelysin-3, uPA, and TIMPs in mouseplacentas of days 10.5 and 16.5 of pregnancy. During this time window,rapid growth and differentiation of the junctional zone and gradualdisappearance of giant trophoblasts takes place. After midpregnancy,nests of morphologically distinct glycogen-rich spongiotrophoblasts emergein the junctional zone, and these cells subsequently continue invading the
maternal decidua. Trophoblast giant cells (day 10.5 p.c.) express tran-scripts of stromelysin-3, gelatinase B and uPA. uPA and TIMP-3 mRNAsare located on spongiotrophoblasts and invasive glycogen cells, whereasstromelysin-3 transcripts are detected close to the fetomaternal interfaceat 10.5 p.c., and over the all decidua at day 16.5 p.c. Gelatinase A andTIMP-2 transcripts at 10.5 days were localized in peripheral decidua; theexpression domain spread progressively, and covered extensive areas atday 16.5 p.c.
254 TEESALU ET AL.
During maturation of murine chorioallantoic pla-centa, stromelysin-3 mRNA was always expressed atthe highest levels by decidua next to the junctionalzone, and its expression level appeared to increasethroughout the period analyzed. On the other hand,gelatinase A transcripts were detected mostly in thedecidual periphery, as in metrial gland, and their leveldeclined in preterm placenta.
The expression patterns of TIMPs suggested thatTIMP-2 is the most relevant inhibitor of MMPs in thedecidua. TIMP-1 and TIMP-3 were found to be presentin decidua only at the early stages analyzed (days 10.5to 12.5 p.c.). Throughout the time period analyzed,TIMP-3 was found to be predominantly expressed in
spongiotrophoblast cells, adjacent to stromelysin-3-expressing decidua, implying a possible role for regula-tion of stromelysin-3 activity. The expression patternsof TIMP-2 and TIMP-3 have been addressed in previousstudies (Apte et al., 1994; Blavier and DeClerck, 1997),for some of the stages we analyzed; our aim was to haveprecise information on the relative positions of theexpression sites of metalloproteinases/inhibitors on par-allel sections.
Expression of stromelysin-3 and gelatinase A could berelated to decidual ECM breakdown, necessary fornormal placental morphogenesis. Besides a direct rolein matrix degradation, extracellular proteolysis couldbe an important modulator of viability of decidual cell
Fig. 7. Relative distribution of uPA and stromelysin-3 transcripts at thefetomaternal interface of mouse placenta. 10.5 days of gestation (A-D).Stromelysin-3 (B) and uPA (C) are coexpressed by trophoblast giant cells,identified on the basis of their PL-1-positivity (D). 12.5 days of gestation(E-H). A bright field image of fetomaternal interface in the center ofplacental disk (E), and photographs of parallel sections hybridized with
stromelysin-3 (F), uPA (G) and 4311 (H). The expression domains of thetwo proteinases fall into two distinct, though partly overlapping domains:uPA in the junctional zone and spongiotrophoblasts invading decidualtissue, stromelysin-3 in decidual cells. d, decidua; ec, embryonic cavity;gc, giant cell; jz, junctional zone. Scale bar: A, 600µm; B-D, 40µm; E-H,160 µm.
TABLE 2. Distribution of uPA:stromelysin-3 Double Null Pups in Heterozygous Crosses(A) and Fertility of the Double Negative and Wild-Type Animals (B)
A. Frequency of uPA/STR-3 double null pups from heterozygous crosses
CrossPups
analyzed2/2:2/2expected
2/2:2/2found
uPA 2/2 ST3 2/2 3 uPA 1/2 ST3 2/2 6 3 2uPA 2/2 ST3 2/2 3 uPA 2/2 ST3 1/2 8 4 5uPA 1/2 ST3 2/2 3 uPA 1/2 ST3 2/2 5 1.25 1uPA 1/2 ST3 2/2 3 uPA 1/2 ST3 2/2 2 0.5 1uPA 2/2 ST3 1/2 3 uPA 2/2 ST3 1/2 18 4.5 4Total 39 13.25 13
B. Fertility of uPA/STR-3 double null and wild-type animals
Cross Litters Pups Mean
uPA 1/1 ST3 1/1 3 uPA 1/1 ST3 1/1 10 81 8.10 6 0.38uPA 2/2 ST3 2/2 3 uPA 2/2 ST3 2/2 11 85 7.73 6 0.47
255MMPs AND PLACENTA

populations, by altering cell anchorage, as well as bygenerating bioactive fragments of ECM components orthrough the release of cytokines.
While decidua and extradecidual maternal struc-tures were relatively enriched in gelatinase A tran-scripts, gelatin substrate zymography showed that thebulk of the proteinase was present in its pro-form.Gelatinase A activation is a finely regulated proteolyticevent, which is believed to be mediated by cell surfacemembrane-type metalloproteinase-1 (MT1-MMP). MT1-MMP and TIMP-2 form a cell surface complex, whichacts as a receptor of pro-gelatinase A (Strongin et al.,1995; Butler et al., 1998). In this cascade, TIMP-2 playsa central regulatory role: in its absence no pro-gelatinase A can bind to the cell surface, and itsactivation is blocked; at large excess of TIMP-2 noproteolytically active MT1-MMP, competent to initiatethe activation of gelatinase A, is available. We foundthat in decidua, large amounts of gelatinase A andTIMP-2 are coexpressed, suggesting that the balance ofTIMP-2 is relatively shifted towards the inhibition ofpro-gelatinase A cleavage. It is possible that the pres-ence of pro-gelatinase A in the decidua forms a pool ofpotential proteolytic activity, which is rapidly recruitedunder physiological conditions (at delivery) and inpathology (preterm delivery). Since for zymography weused extracts of whole placentas, together with sur-rounding myometrial/endometrial components, the ex-istence of important local differences in gelatinase Aactivation patterns can not be excluded.
In human placenta, expression of high amounts ofMT1-MMP by extravillous trophoblasts has been re-ported (Nawrocki et al., 1996). In murine placenta at12.5 and 14.5 days, MT1-MMP and TIMP-2 transcriptswere found to colocalize to the junctional region be-tween uterine deciduum and spongiotrophoblasts (Apteet al., 1997).
Trophoblast/Stromal Cooperationin Decidual Remodeling
During many invasive processes, extracellular prote-ases are expressed by both invasive cells and adjacentstroma. The stroma of a solid tumor is distinct fromnon-cancerous tissue surrounding the tumor, andthough not malignant per se, it plays a key role intumor progression and pathogenesis (Wernert, 1997). Anumber of proteases have been shown to be expressedby malignant tumor cells, as well as by tumoral stroma(Heppner et al., 1996; Andreasen et al., 1997). In thecurrent study, we have shown that in placenta, proteo-lytic enzymes are expressed by both the invasive tropho-blasts, as well as by decidual stromal cells in thevicinity (stromelysin-3 and gelatinase A). Thus, theprocess of ECM remodeling taking place in mouseplacenta is characterized by cooperation between inva-sive cells and stromal components, similar to what hasbeen observed in tumors.
The concept that tissue remodeling is associated withmany independent proteolytic pathways, has become
well established (Werb, 1997). Coexpression of a num-ber of extracellular proteases in a variety of processeshas been observed, thereby suggesting a functionalcooperation between the enzymes. uPA and stromely-sin-3 have been shown to be coexpressed in numerousphysiological and pathological processes, such as tropho-blast giant cell invasion, mammary gland involution,bone ossification, tumor invasion, etc. (Lefebvre et al.,1992; Wolf et al., 1993; Lefebvre et al., 1995; Maquoi etal., 1997). Expression of stromelysin-3 has been foundin both invasive cells (trophoblast giant cells andpossibly osteoblasts) (Lefebvre et al., 1995), as well asin stroma surrounding invasive cells (as in the case ofcarcinomas) (Linder et al., 1998). In the current study,we show that in late murine placenta stromelysin-3 ispresent in the invasive trophoblast cells, as well as indecidual stroma undergoing invasion.
In order to address directly the functional meaning ofuPA and stromelysin coexpression in mouse placenta,we have generated mice with a combined deficiency ofuPA and stromelysin-3 genes. Mice with single deficien-cies for each of these enzymes are viable and fertile,albeit a number of abnormalities have been describedfor uPA null mice (reviewed by Carmeliet and Collen,1996). If a functional cooperation/mutual compensationbetween the two enzymes exists, stromelysin-3/uPAdouble null mice might manifest a more severe pheno-type than single mutants. Our finding that double nullmice are viable and fertile, shows that these twoproteinases are not absolutely required for mouse pla-cental development, and suggests that in single knock-outs no compensatory mechanisms of crucial impor-tance between these proteinases have evolved.
MATERIALS AND METHODSAnimals and Sections
Adult CD1 female mice were caged with males in theevening and monitored for the appearance of a vaginalplug the next morning. Pregnant animals were sacri-ficed and placentas were collected and subsequentlyeither snap-frozen in liquid nitrogen or fixed withfreshly prepared 4% paraformaldehyde in PBS (pH 7.4)overnight at 4°C. Fixed tissues were dehydrated, paraf-fin-embedded according to routine histological proce-dures and sectioned at 6 µM. Serial sections weremounted on gelatin-coated slides.
uPA -/- (Carmeliet et al., 1994) and wild type mice ofthe same background strain were kindly provided byDr. Peter Carmeliet. Generation of the stromelysin-3mice has been described by Masson et al. (1998). uPAand stromelysin-3 genes are located in different chromo-somes, thus we used breeding for generation of doublenull animals. Pups were genotyped by Southern blotanalysis of tail DNA.
In Situ Hybridization
35S-UTP-labeled RNA probes were prepared as de-scribed previously (Teesalu et al., 1996). The followinganti-sense probes were used: gelatinase A, gelatinase B,
256 TEESALU ET AL.

TIMP-1, TIMP-2, TIMP-3 (for a detailed description seeAlexander et al., 1996); stromelysin-3 (Lefebvre et al.,1992); and uPA, PL-1, and 4311 (for details see Teesaluet al., 1996). The labeled RNAs were reduced to approxi-mately 100 bases by alkaline hydrolysis, neutralized,extracted with phenol-chloroform, purified from unincor-porated nucleotides by filtration through SephadexG-50 column and ethanol precipitated. In situ hybridiza-tion of serial sections of placentas was performedessentially as described previously (Wilkinson et al.,1987). After hybridization, the slides were dipped intoKodak NTB2 emulsion, exposed at 4°C for 1–3 weeksand after standard development procedures stainedwith toluidine blue and mounted in DPX mountant.Slides were observed and photographs taken using aZeiss Axiophot microscope.
Northern Blot Analysis
Total RNA from mouse placentas was obtained usingguanidinium thiocyanate-phenol-chloroform extractionprocedure.
RNA was separated on 1% agarose gels and trans-ferred to nylon membranes by capillary transfer; hybrid-ization was performed using standard techniques.
Immunohistochemistry
Immunohistochemistry was performed with the Vec-tastain peroxidase staining kit (Vector Laboratories,Burlingame, CA) according to the manufacturers in-structions. The following commercial rabbit polyclonalantibodies were used: anti-cytokeratin antibody (DAKO,Glostrup, Denmark, catalog number Z0622) and anti-laminin antibody (Sigma, St. Louis, MO; L-9393).
Zymography on Polyacrylamide Gels
Placental extracts were analyzed by gelatin zymogra-phy of polyacrylamide gels (adopted with modificationsfrom Behrendtsen et al., 1992). Whole placentas werepowdered in liquid nitrogen and solubilized in 10 mMTris (pH 7.5), 0.25% Triton X-100. HT1080 conditionedmedium (positive control of gelatinolysis) was preparedby incubating the cells for 48 hr in serum free medium.Samples were electrophoresed on non-reducing 10%polyacrylamide-SDS gels, copolymerized with sub-strate (1 mg/ml type B collagen). After electrophoresis,gels were washed twice for 15 min in 2.5% Triton X-100,incubated in 10 mM CaCl2, 50 mM Tris (pH 7.6) for24–40 hr at 37°C and stained with Coomassie Blue. Theareas of proteolytic activity appeared as clear areas inthe background of blue staining.
ACKNOWLEDGMENTS
Dr. Zena Werb is gratefully acknowledged for provid-ing the cDNA probes for gelatinases and TIMPs. We aregrateful to Marie Christine Rio and Cornelius F.M. Sierfor critically reading the manuscript. Tambet Teesaluwas supported by fellowships from Telethon (Rome) andNordic Council of Ministers (Copenhagen). This workwas supported by grants of E.U. (Biomed II; contract
BMH4CT96–0017) to F.B. and P.B., AIRC (AssociazioneItaliana per la Ricerca sul Cancro) and MURST to F.B.P.B. thanks the following agencies for grant support:Institut National de la Sante et de la Recherche Medi-cale, the Centre Hospitalier Universitaire Regional, theBristol-Myers Squibb Pharmaceutical Research Insti-tute, the Association pour la Recherche sur le Cancer, laLigue Nationale Francaise contre le Cancer, the Comitedu Haut-Rhin, the Fondation de France, BIOTECH 2(contract ERBBIO4CT96–0464) program, and a grantto P. Chambon from the Fondation Jeantet.
REFERENCESAbrahamsohn PA, Zorn TM. 1993. Implantation and decidualization
in rodents. J Exp Zool 266:603–628.Afonso S, Romagnolo L, Babiarz B. 1997. The expression and function
of cystatin C and cathepsin L during mouse embryo implantationand placentation. Development 124:3415–3425.
Aimes RT, Quigley JP. 1995. Matrix metalloproteinase-2 is an intersti-tial collagenase. J Biol Chem 270:5872–5876.
Alexander CM, Hansell EJ, Behrendtsen O, Flannery ML, KishnaniNS, Hawkes SP, Werb Z. 1996. Expression and function of matrixmetalloproteinases and their inhibitors at the maternal-embryonicboundary during mouse embryo implantation. Development 122:1723–1736.
Andreasen PA, Kjoller L, Christensen L, Duffy MJ. 1997. The uroki-nase-type plasminogen activator in cancer metastasis: a review. IntJ Cancer 72:1–22.
Apte SS, Hayashi K, Seldin MF, Mattei M-G, Hayashi M, Olsen BR.1994. Gene encoding a novel murine tissue inhibitor of metallopro-teinases (TIMP), is expressed in developing mouse epithelia, carti-lage and muscle, and is located on chromosome 10. Dev Dyn200:177–197.
Apte SS, Fukai N, Beier DR, Olsen BR. 1997. The matrix metallopro-teinase-14 (MMP-14) gene is structurally distinct from other MMPgenes and is co-expressed with the TIMP-2 gene during mouseembryogenesis. J Biol Chem 272:25511–25517.
Basset P, Bellocq J-P, Lefebvre O, Noel A, Chenard M-P., Wolf C,Anglard P, Rio, M-C. 1997. Stromelysin-3: a paradigm for stroma-derived factors implicated in carcinoma progression. Crit RevOncology 26:43–53.
Behrendtsen O, Alexander CM, Werb Z. 1992. Metalloproteasesmediate extracellular matrix degradation by cells from mouseblastocyst outgrowths. Development 114:447–456.
Birkedal-Hansen H. 1995. Proteolytic remodeling of extracellularmatrix. Curr Opin Cell Biol 7:728–735.
Blavier L, DeClerck YA. 1997. Tissue inhibitor of metalloproteinases-2is expressed in the interstitial matrix in adult mouse organs andduring embryonic development. Mol Biol Cell 8:1513–1527.
Butler GS, Butler MJ, Atkinson SJ, Will H, Tamura T, van WestrumSS, Crabbe T, Clements J, d’Ortho MP, Murphy G. 1998. TheTIMP-2 membrane type 1 metalloproteinase ‘‘receptor’’ regulatesthe concentration and efficient activation of progelatinase A. J BiolChem 273:871–880.
Carmeliet P, Collen D. 1996. Targeted gene manipulation and transferof the plasminogen and coagulation systems in mice. Fibrinolysis10:195–213.
Carmeliet P, Schoonjans L, Kieckens L, Ream B, Degen J, Bronson R,De Vos R, van den Oord JJ, Collen D, Mulligan RC. 1994. Physiologi-cal consequences of loss of plasminogen activator gene function inmice. Nature 368:419–424.
Cross JC, Werb Z, Fisher S. 1994. Implantation and the placenta: keypieces of the development puzzle. Science 266:1508–1518.
D’Armiento J, Dalal SS, Okada I, Berg RA, Chada K. 1992. Collage-nase expression in the lungs of transgenic mice causes pulmonaryemphysema. Cell 71:955–961.
Edwards DR, Murphy G. 1998. Proteases - invasion and more. Nature394:527–528.
Enders AC, Welsh AO. 1993. Structural interactions of trophoblast
257MMPs AND PLACENTA

and uterus during hemochorial placenta formation. J Exp Zool266:578–587.
Heppner KJ, Matrisian LM, Jensen RA, Rodgers WH. 1996. Expres-sion of most matrix metalloproteinase family members in breastcancer represents a tumor-induced host response. Am J Pathol149:273–282.
Jones JL, Walker RA. 1997. Control of matrix metalloproteinaseactivity in cancer. J Pathol 183:377–379.
Itoh T, Tanioka M,Yoshida T, Nishimoto H, Itohara S. 1998. Reducedangiogenesis and tumor progression in gelatinase A-deficient mice.Cancer Res 58:1048–1051.
Kleiner DE, Stetler-Stevenson WG. 1993. Structural biochemistry andactivation of matrix metalloproteinases. Curr Opin Cell Biol 5:891–897.
Lefebvre O, Wolf C, Limacher JM, Hutin P, Wendling C, LeMeur M,Basset P, and Rio MC. 1992. The breast cancer-associated stromely-sin-3 gene is expressed during mouse mammary gland apoptosis. JCell Biol 199:997–1002.
Lefebvre O, Regnier C, Chenard M-P, Wendling C, Chambon P, BassetP, Rio MC. 1995. Developmental expression of mouse stromelysin-3mRNA. Development 121:947–955.
Linder C, Bystrom P, Engel G, Auer G, Aspenblad U, Strander H,Linder S. 1998. Correlation between basic fibroblast growth factorimmunostaining of stromal cells and stromelysin-3 mRNA expres-sion in human breast carcinoma. Br J Cancer 77:941–945.
Maquoi E, Polette M, Nawrocki B, Bischof P, Noel A, Pintiaux A,Santavicca M, Schaaps J-P, Pijnenborg R, Birembaut P, Foidart J-M.1997. Expression of Stromelysin-3 in the human placenta andplacental bed. Placenta 18:277–285.
Masson R, Lefebvre O, Noel A, El Fahime M, Chernard M-P, WendlingC, Kebers F, LeMeur M, Dierich A, Foidart J-M, Basset P, Rio MC.1998. In vivo evidence that the stromelysin-3 metalloproteinasecontributes in a paracrine manner to epithelial cells malignancy. JCell Biol 140:1535–1543.
Matrisian LM. 1990. Metalloproteinases and their inhibitors in matrixremodeling. Trends Genet 6:121–125.
Matrisian LM, Hogan BLM. 1990. Growth factor-regulated proteasesand extracellular matrix remodeling during mammalian develop-ment. Curr Top Dev Biol 24:219–257.
Nawrocki B, Polette M, Marchand V, Maquoi E, Beorchia A, TournierJM, Foidart JM, Birembaut P. 1996. Membrane-type matrix metal-loproteinase expression at the site of human placentation. Placenta17:565–572.
Pijnenborg R, Robertson WB, Brosens I, Dixon G. 1981. Review article:
trophoblast invasion and the establishment of haemochorical placen-tation in man and laboratory animals. Placenta 2:71–91.
Redline RW, Lu CY. 1989. Localization of fetal major histocompatibil-ity complex antigens and maternal leukocytes in murine placenta.Implications for maternal-fetal immunological relationship. LabInvest 61:27–36.
Rinkenberger JL, Cross JC, Werb Z. 1997. Molecular genetics ofimplantation in the mouse. Dev Gen 21:6–20.
Sappino AP, Huarte J, Belin D, Vassalli JD. 1989. Plasminogenactivators in tissue remodeling and invasion: mRNA localization inmouse ovaries and implanting embryos. J Cell Biol 109:2471–2479.
Sato H, Takino T, Okada Y, Cao J, Shinagawa A, Yamamoto E, Seiki M.1994. A matrix metalloproteinase expressed on the cell surface ofinvasive tumor cells. Nature 370:61–65.
Shapiro SD. 1997. Mighty mice: transgenic technology ‘‘knocks out’’questions of matrix metalloproteinase function. Matrix Biol:15:527–533.
Strongin AY, Collier I, Bannikov G, Marmer BL, Grant GA, GoldbergGI. 1995. Mechanism of cell surface activation of 72 kDa type IVcollagenase. Isolation of the activated form of the membrane metal-loprotease. J Biol Chem 27:5331–5338.
Sympson CJ, Bissell MJ, Werb Z. 1995. Mammary gland tumorformation in transgenic mice overexpressing stromelysin-1. SeminCancer Biol 3:159–163.
Teesalu T, Blasi F, Talarico D. 1996. Embryo implantation in themouse: fetomaternal coordination in the pattern of expression ofuPA, uPAR, PAI-1, and a2MR/LRP genes. Mech Dev 56:103–116.
Teesalu T, Blasi F, Talarico D. 1998.Expression and function of theurokinase type plasminogen activator during mouse hemochorialplacental development. Dev Dyn 213:27–38.
Vu TH, Shipley JM, Bergers G, Berger JE, Helms JA, Hanahan D,Shapiro SD, Senior SD, Werb Z. 1998. MMP-9/gelatinase B is a keyregulator of growth plate angiogenesis and apoptosis of hypertro-phic chondrocytes. Cell 93:411–422.
Werb Z. 1997. ECM and cell surface proteolysis: regulating cellularecology. Cell 91:439–442.
Wernert N. 1997. The multiple roles of tumour stroma. Virchows Arch430:433–443.
Wilkinson DG, Bailes JA, McMahon AP. 1987. Expression of theproto-oncogene int-1 is restricted to specific neural cells in thedeveloping mouse embryo. Cell 50:79–88.
Wolf C, Rouyer N, Lutz Y, Adida C, Loriot M, Bellocq J-P, Chambon P,Basset P. 1993. Stromelysin-3 belongs to a subgroup of proteinasesexpressed in breast carcinoma fibroblastic cells and possibly impli-cated in tumor progression. Proc Natl Acad Sci USA 90:1843–1847.
258 TEESALU ET AL.
Copyright © 2022 FDOKUMEN




















