![[Revised] Revisiting Verb Aspect in T'boli](https://static.fdokumen.com/doc/165x107/631ef9e50ff042c6110c9f71/revised-revisiting-verb-aspect-in-tboli.jpg)
Adsorption of matrix metalloproteinases onto biomedical polymers: a new aspect in biological...
11
J. Biomater. Sci. Polymer Edn, Vol. 19, No. 1, pp. 19–29 (2008) VSP 2008. Also available online - www.brill.nl/jbs Adsorption of matrix metalloproteinases onto biomedical polymers: a new aspect in biological acceptance FILIPPO RENÒ, VINCENZINA TRAINA and MARIO CANNAS ∗ Department of Medical Science, Human Anatomy Laboratory, University of Eastern Piedmont, via Solaroli 17, I-28100 Novara, Italy Abstract—Matrix metalloproteinases (MMPs) are zinc-dependent enzymes involved in the remod- elling of connective tissues during the development and wound healing. Moreover, two MMPs, Gelatinase A (MMP-2) and Gelatinase B (MMP-9), are also present in body fluids such as blood and urine and, therefore, they can be in contact with implanted biomaterials and can be adsorbed onto their surface. In order to test this hypothesis disks of different polymers (polystyrene (PS), polyvinyl chloride (PVC), poly(D, L-lactide) (PLA), polymethyl methacrylate (PMMA) and poly(2- hydroxyethyl methacrylate) (PHEMA)) have been exposed to human plasma and adsorbed proteins have been eluted and analyzed. Using Western blot and substrate zymography analysis, we observed that both MMP-2 and MMP-9 adsorbed onto the surfaces of all the polymers, especially hydrophilic ones (PMMA and PHEMA) and PLA, in both the active and inactive forms. Furthermore, we observed that adhesion of human granulocyte neutophils to PMMA, the polymer that adsorbed the higher quan- tity of MMP-2 and MMP-9 compared to the others, was reduced by more that 50% by the presence of a gelatinase inhibitor. This data suggest a surprising role of these absorbed enzymes in the adhesion of neutrophil onto some polymeric biomaterials surface and, therefore, in the setting of inflammation. Key words: Matrix metalloproteinases; polymers; adsorption; cell adhesion; neutrophil. INTRODUCTION Biomaterials surfaces are often exposed to concentrated solutions of proteins such as blood or other body fluids after prosthesis implantation, and protein adsorption from these fluids can elicit adverse host response such as blood coagulation and complement activation [1]; on the other hand, cell adhesion to surface depends on the availability of specific protein-binding sites [2]. Adsorption of plasma protein such as immunoglobulin G (IgG), albumin and fibrinogen has been extensively stud- ied due to their relatively large concentration in plasma and their ability to modulate ∗ To whom correspondence should be addressed. E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Adsorption of matrix metalloproteinases onto biomedical polymers: a new aspect in biological...

J. Biomater. Sci. Polymer Edn, Vol. 19, No. 1, pp. 19–29 (2008) VSP 2008.Also available online - www.brill.nl/jbs
Adsorption of matrix metalloproteinases onto biomedicalpolymers: a new aspect in biological acceptance
FILIPPO RENÒ, VINCENZINA TRAINA and MARIO CANNAS ∗
Department of Medical Science, Human Anatomy Laboratory, University of Eastern Piedmont,via Solaroli 17, I-28100 Novara, Italy
Abstract—Matrix metalloproteinases (MMPs) are zinc-dependent enzymes involved in the remod-elling of connective tissues during the development and wound healing. Moreover, two MMPs,Gelatinase A (MMP-2) and Gelatinase B (MMP-9), are also present in body fluids such as bloodand urine and, therefore, they can be in contact with implanted biomaterials and can be adsorbedonto their surface. In order to test this hypothesis disks of different polymers (polystyrene (PS),polyvinyl chloride (PVC), poly(D,L-lactide) (PLA), polymethyl methacrylate (PMMA) and poly(2-hydroxyethyl methacrylate) (PHEMA)) have been exposed to human plasma and adsorbed proteinshave been eluted and analyzed. Using Western blot and substrate zymography analysis, we observedthat both MMP-2 and MMP-9 adsorbed onto the surfaces of all the polymers, especially hydrophilicones (PMMA and PHEMA) and PLA, in both the active and inactive forms. Furthermore, we observedthat adhesion of human granulocyte neutophils to PMMA, the polymer that adsorbed the higher quan-tity of MMP-2 and MMP-9 compared to the others, was reduced by more that 50% by the presence ofa gelatinase inhibitor. This data suggest a surprising role of these absorbed enzymes in the adhesionof neutrophil onto some polymeric biomaterials surface and, therefore, in the setting of inflammation.
Key words: Matrix metalloproteinases; polymers; adsorption; cell adhesion; neutrophil.
INTRODUCTION
Biomaterials surfaces are often exposed to concentrated solutions of proteins suchas blood or other body fluids after prosthesis implantation, and protein adsorptionfrom these fluids can elicit adverse host response such as blood coagulation andcomplement activation [1]; on the other hand, cell adhesion to surface depends onthe availability of specific protein-binding sites [2]. Adsorption of plasma proteinsuch as immunoglobulin G (IgG), albumin and fibrinogen has been extensively stud-ied due to their relatively large concentration in plasma and their ability to modulate
∗To whom correspondence should be addressed. E-mail: [email protected]

20 F. Renò et al.
cell response in the inflammatory process [3]. Body fluids contain also many otherproteins that can modulate cell behavior such as matrix metalloproteinases (MMPs)but their ability to be adsorbed onto artificial surfaces has not been investigated.
MMPs are a family of structurally related zinc-dependent endopeptidases [4],which can degrade all ECM components and which activity is essential for a normalwound healing [5]. MMPs are secreted by stimulated cell as inactive proenzymesand activated successively through rather complex mechanisms [4]. Therefore,MMPs are present in both active and inactive forms in body fluids such as plasma[6, 7], saliva [8] and urine [9] both in physiological and pathological conditionsat different concentrations ranging from to 50 to 200 µg/mg protein. Among theMMPs the gelatinases, gelatinase A (MMP-2, 64 kDa) and gelatinase B (MMP-9,72 kDa), are present in human plasma and their activity seems to play a key role ininflammation [10].
In this paper we evaluated the adsorption of MMP-2 and MMP-9 present inthe human plasma onto the surface of 5 biopolymers with different wettabilityusing immunoblotting and gelatin substrate zymography to reveal their presenceand activity. Moreover, we investigated if the presence and activity of MMP-2 andMMP-9 could alter human neutrophiles adhesion onto biopolymers surface, a keyevent in the inflammatory response to biomaterials [11].
MATERIALS AND METHODS
Preparation of polymers
Polystyrene (PS), poly(methyl methacrylate) (PMMA) and poly(D,L-lactide)(Sigma-Aldrich) were dissolved in chloroform to 5% (w/v) solutions. Poly(2-hydroxyethyl methacrylate) (PHEMA) was dissolved in 1:1 tetrahydrofuran/65%(v/v) ethanol solution to produce a 5% (w/v) solution. Poly(vinyl chloride) was dis-solved in tetrahydrofuran to 5% (w/v) solution. Polymer films were cast onto glassdishes, dried overnight at 37◦C and cut in disks with a surface of 0.4 cm2.
Plasma and granulocyte preparation
Plasma and granulocyte neutrophil were separated from blood obtained from6 healthy donors (2 men and 4 women, average age ± SD = 30 ± 6 years).
Granulocytes were obtained from peripheral blood by a modification of themethod of Boyum [12]. EDTA collected human peripheral venous blood (20 ml)was divided in two aliquots of 10 ml. The first aliquot was diluted 1:1 withPBS (pH 7.4), layered onto a Ficoll-Hypaque density gradient and centrifugedfor 20 min at 2000 rpm to separate mononuclear cells from erythrocytes andgranulocytes. The mononuclear fraction was discharged and erythrocytes werethen lysed using an ammonium chloride lysing solution (150 mM NH4Cl, 10 mM

MMP adsorption onto biomedical polymers 21
NaHCO3, 1 mM EDTA, pH 7.4) for 20 min at 4◦C. Pellet containing granulocyteswas centrifuged twice in sterile phosphate buffer (PBS), cells were counted inoptical microscopy using the Trypan blue exclusion test (viability > 98%) andsuspended at a concentration of 1 × 106 cells/ml in serum free RPMI 1640 mediumin polypropylene tubes. For some experiments 10% (v/v) of human plasma wasadded to RPMI medium.
The second blood aliquot was used to obtain platelet-poor plasma (PPP). Briefly,undiluted blood was centrifuged at 200×g for 10 min to obtain platelet-rich plasma(PRP). PRP was then centrifuged at 1600 × g for 10 min to separate platelets andto obtain PPP.
Plasma protein adsorption onto polymeric surfaces
Protein adsorption assay was performed in triplicate using PS, PVC, PLA, PMMAand PHEMA disks that were covered with 200 µl of undiluted PPP and incubatedfor 1 h at 37◦C.
At the end of incubation, plasma was removed and disks were washed threetimes with PBS. Adsorbed proteins were collected by incubating samples withof 2% sodium dodecyl sulfate (SDS) solution in PBS (pH 7.4) for 4 hours atroom temperature and under vigorous shaking. Amount of adsorbed proteins wasmeasured in triplicate using a protein quantification kit (BCA; Pierce). The sampleoptical density was read at 562 nm against a calibration curve created using bovineserum albumin (BSA, 25–2000 µg/ml). The results were expressed as µg totalprotein absorbed/cm2 ± SD.
Patterns of adsorbed proteins have been analysed using SDS-polyacrylamide gelelectrophoresis (PAGE). Protein samples were denatured by boiling for 3 min,centrifuged at 12 000×g and then a 10 µl volume was loaded onto a 10% separatingpolyacrylamide gel with a 4% stacking gel. Electrophoresis was conducted at 100 Vby an electrophoresis system (Bio-Rad Mini-Protean II). Proteins were stained by aSilver Stain Plus kit (Bio-Rad). A densitometric analysis of the bands seen on gelswas performed using the Scion Image 4.02 software and size band was expressed asarbitrary units (a.u.).
Western blotting
For the blot analysis of MMP-2 and MMP-9, 50 µl of protein solution, detachedfrom polymers, were dissolved into the electrophoresis sample buffer (25 mM Trisbase, 191 mM glycine, 0.35 mM SDS). After boiling 20-µl aliquots of the sampleswere run on 7.5% SDS-polyacrylamide gels and transferred to nitrocellulosemembrane (Hybond, Amersham). Non-specific binding was eliminated using PBSsupplemented with 5% non-fat dry milk and 0.1% Tween-20 (Amersham PharmaciaBiotech) for 60 min. The membranes were incubated overnight at 4◦C underconstant shaking with a primary mouse anti-human monoclonal antibody against

22 F. Renò et al.
MMP-2 (1:400; Chemicon) or a primary mouse anti-human monoclonal againstMMP-9 (1:400; Chemicon). After extensive washing, blots were incubated witha peroxidase-labeled rabbit secondary antibody (1:2000; Amersham PharmaciaBiotech) and developed with the ECL system (Amersham Pharmacia Biotech).
Gelatin zymography
Protein samples were analyzed for measurement of MMP-2 and MMP-9 presenceand activity by zymography [5]. Briefly, Gelatinase A (MMP-2) and Gelatinase B(MMP-9) activity were detected by zimogram analysis using SDS-polyacrylamidegels co-polymerized with 0.2% gelatin. Deassorbed protein samples were mixedwith sample buffer and electrophoresed directly without boiling or reduction.Following electrophoresis, SDS was extracted from the polyacrylamide gel withTriton X-100, and the gel was incubated in 0.05 M Tris (pH 7.5), containing 5mM CaCl2 and 5 mM ZnCl2 at 37◦C overnight. Gels were then stained withCoomassie blue and destained. Both proenzyme and active gelatinases weredetected as clear bands against the blue background of the stained gelatin. Positivecontrols for gelatinase A and B (Chemicon) were used to identify the two enzymes.A densitometric analysis of the bands seen on gels was performed using the NIHImage 1.62 software and size band was expressed as arbitrary units (a.u.).
Neutrophil adhesion
A volume of 200 µl of granulocytes suspension (1 × 106 cells/ml) in plasma freeor supplemented with 10% human serum RPMI 1640 was added to PS and PMMApolymer disks uncoated or coated with plasma as previously described for 1 h at37◦C. At the end of the incubation disks were gently washed twice with PBS andadherent cells were fixed with a solution of 11% (v/v) glutharaldeide in PBS for15 min at room temperature. In some experiments the MMP-2/MMP-9 Inhibitor I(Calbiochem) was added to a final concentration of 0.5 µM.
Cell adhesion was quantified using crystal violet assay and adherent cell morphol-ogy was evaluated using acridine orange (AO) staining.
Samples for crystal violet assay were washed twice with deionized water anddried overnight at 37◦C, stained with crystal violet for 30 min at room temperature,washed and dried again. At the end 1 ml of a 11% solution of acetic acidwas added to each sample for 30 min under shaking in order to extract the dye.The optical density of the samples was read at 594 nm using a Beckman DU-68spectrophotometer.
In some experiments adherent cells were fixed with a solution of 3.7% formalineand 3% saccarose in PBS. Afterwards cells were stained for 5 min with AOsolution (0.025%, w/v) in PBS, washed three times with PBS and observed under afluorescence microscope at a magnification of 400×.

MMP adsorption onto biomedical polymers 23
Statistical analysis
Statistical analysis of variance (ANOVA) was used; the significance of differencesbetween means was assessed using the Newman–Keuls multiple range test, takingP < 0.05 as the minimum level of significance.
RESULTS
Protein adsorption onto polymers surface
The surface of biopolymers binds a different amount of plasma protein according totheir wettability. In fact, as shown in Fig. 1A hydrophilic polymers such as PMMAand PHEMA bind the highest quantity of adsorbed protein, 684 ± 136 µg/cm2 and
Figure 1. (A) BCA results of protein adsorption to the materials incubated 30 min at roomtemperature with undiluted PPP. Optical densities were read at 562 nm against a calibration curveusing BSA. Results are expressed as µg protein/cm2 ± SD. *P < 0.05; **P < 0.001 comparedto PS. (B) Silver-stained SDS-PAGE patterns of protein layers adhered to biomaterials, after in vitroincubation with PPP. White arrow = proteins around 66–70 kDa.

24 F. Renò et al.
717 ± 60 µg/cm2, respectively, while the most hydrophobic polymer used (PS)adsorbed only 76 ± 3 µg/cm2. The qualitative analysis of the adsorbed proteins wasconducted by SDS-PAGE using 7% polyacrylamide gels (Fig. 1B). The analysis ofthe silver staining pattern of the proteins eluted from the tested materials showed forall polymers a main band at about 66–70 kDa, tentatively identified as albumin band(with arrow). Other evident bands around 200 kDa were observed in the sampleseluted from PMMA and PHEMA (black arrow).
Identification of MMP-2 and MMP-9 adsorbed onto polymers
The presence of MMP-2 and MMP-9 among the protein adsorbed onto polymerssurface and eluted with 2% SDS was investigated using both Western immunoblot-ting and substrate zymography. In Fig. 2 the immunoblotting showed that bothMMP-2 and MMP-9 were present in the human plasma and they adsorbed onto allthe polymers tested both in the inactive (72 kDa band for MMP-2, 92 kDa bandfor MMP-9) and active (64 kDa band for MMP-2, 78 kDa band for MMP-9) form.The densitometric analysis showed that PMMA was the polymer more capable tobind both MMP-2 and MMP-9 compared to the others. In fact, the band size ex-pressed in a.u. ± SD observed for MMP-9 bands in three different experiments forPS samples was 555 ± 31, while it was 4136 ± 105, 1232 ± 54 and 1577 ± 51for PMMA, PHEMA and PLA, respectively. The MMP-9 bands present in PVCsamples showed an O.D. of 671 ± 70.
Similar results were observed for MMP-2 bands. In fact PMMA samples showeda band size of 4724±238, higher than observed for PS (855±96), PVC (215±64),PLA (3851 ± 309) and PHEMA (1759 ± 50).
Immunoblotting data were almost completely confirmed by gelatin substratezymography. In Fig. 3A presence and activity of MMP-2 and MMP-9 wasdemonstrated using 10 µl of eluted protein SDS solution/lane. In the zymogram
Figure 2. Western immunoblotting for MMP-2 and MMP-9 adsorbed onto the different polymers.Molecular mass scale in kDa.

MMP adsorption onto biomedical polymers 25
Figure 3. (A) Representative zymogram of protein eluted from polymers (10 µl of protein elutedsolution/lane). (B) Densitometric analysis of the bands seen on three different zymograms fromthere experiments was performed using the NIH Image 1.62 software and size band was expressedas arbitrary units (a.u.) ± SD. **P < 0.001 compared to PS.
the PS samples showed a very faint presence of both enzymes, while the highestpresence was observed for PMMA samples. Moreover, in the zymogram a typicalband around 180 kDa formed by MMP-9 dimers or by MMP-9 bound with TIMP-1[13] was observed. The same band was not observed in the immunoblottinganalysis. The highest presence of MMPs onto PMMA samples was confirmed bydensitometric analysis performed on three different zymograms from there differentexperiments that showed a band size ± SD for MMP-9 and MMP-2, respectively,of 319 181 ± 11 051 and 91 945 ± 9232 a.u. (Fig. 3B).
Effect of MMP-2 and MMP-9 inhibitor on neutrophil adhesion onto polymers
The proteins adsorbed onto a biomaterial surface are able to modulate the recruit-ment of macrophages and neutrophils to the implant surface [14]; therefore, we

26 F. Renò et al.
Figure 4. Quantification of neutrophil adhesion onto PS and PMMA in the absence of plasma, in thepresence of 10% human plasma and after surface preconditioning with human plasma, in the absence(white columns) and presence (black columns) of MMPs inhibitor I. **P < 0.001.
evaluated the adhesion of human neutrophils to the surfaces of the two polymersthat have shown, respectively, the lower (PS) and the higher (PMMA) presenceand activity of MMP-2 and MMP-9. The adhesion of neutrophils has been tested inserum-free medium, in 10% human plasma medium and after preconditioning of thepolymers with PPP as described in Materials and Methods. In these three conditionsan inhibitor of the activity of MMP-2 and MMP-9 (MMP-2/MMP-9 Inhibitor I, Cal-biochem) was also added at a final concentration of 0.5 µM. The quantification ofthe neutrophil adhesion was calculated measuring the optical density of the sam-ples treated as described in Materials and Methods. As shown in Fig. 4, the neu-trophil adhesion to PS surface in the absence of the MMPs inhibitor (open columns)was very low in every condition compared to the adhesion that occurred onto thePMMA surface. In particular onto PMMA the adhesion in the presence of 10%human plasma in the medium increased almost three times compared to the plasma-free condition and it increased almost 7 times after preconditioning of the polymersurface with PPP. The addition of MMP-2/MMP-9 Inhibitor I (0.5 µM) to the neu-trophile suspension (filled columns) had no significant effects on the cell adhesion
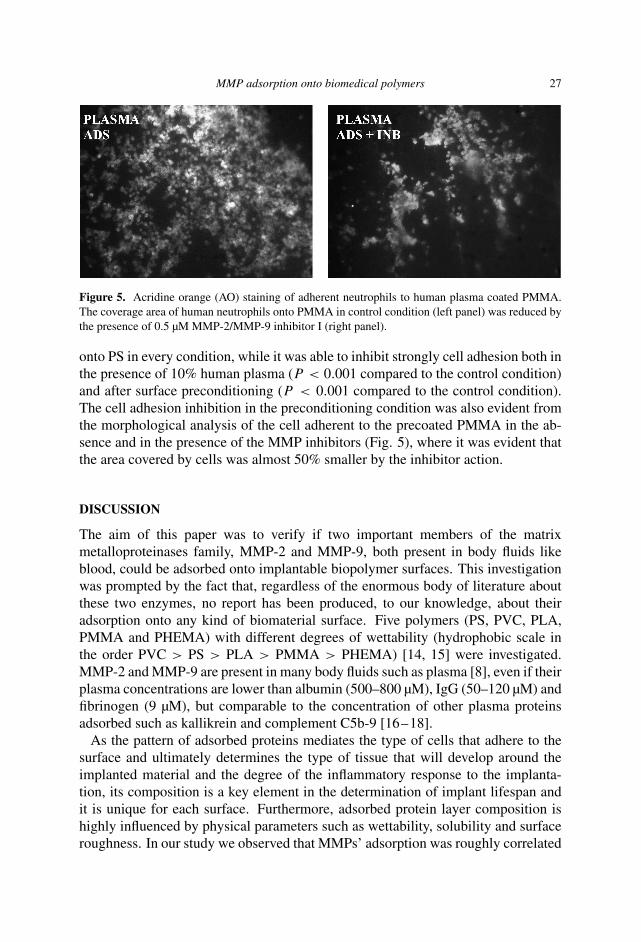
MMP adsorption onto biomedical polymers 27
Figure 5. Acridine orange (AO) staining of adherent neutrophils to human plasma coated PMMA.The coverage area of human neutrophils onto PMMA in control condition (left panel) was reduced bythe presence of 0.5 µM MMP-2/MMP-9 inhibitor I (right panel).
onto PS in every condition, while it was able to inhibit strongly cell adhesion both inthe presence of 10% human plasma (P < 0.001 compared to the control condition)and after surface preconditioning (P < 0.001 compared to the control condition).The cell adhesion inhibition in the preconditioning condition was also evident fromthe morphological analysis of the cell adherent to the precoated PMMA in the ab-sence and in the presence of the MMP inhibitors (Fig. 5), where it was evident thatthe area covered by cells was almost 50% smaller by the inhibitor action.
DISCUSSION
The aim of this paper was to verify if two important members of the matrixmetalloproteinases family, MMP-2 and MMP-9, both present in body fluids likeblood, could be adsorbed onto implantable biopolymer surfaces. This investigationwas prompted by the fact that, regardless of the enormous body of literature aboutthese two enzymes, no report has been produced, to our knowledge, about theiradsorption onto any kind of biomaterial surface. Five polymers (PS, PVC, PLA,PMMA and PHEMA) with different degrees of wettability (hydrophobic scale inthe order PVC > PS > PLA > PMMA > PHEMA) [14, 15] were investigated.MMP-2 and MMP-9 are present in many body fluids such as plasma [8], even if theirplasma concentrations are lower than albumin (500–800 µM), IgG (50–120 µM) andfibrinogen (9 µM), but comparable to the concentration of other plasma proteinsadsorbed such as kallikrein and complement C5b-9 [16–18].
As the pattern of adsorbed proteins mediates the type of cells that adhere to thesurface and ultimately determines the type of tissue that will develop around theimplanted material and the degree of the inflammatory response to the implanta-tion, its composition is a key element in the determination of implant lifespan andit is unique for each surface. Furthermore, adsorbed protein layer composition ishighly influenced by physical parameters such as wettability, solubility and surfaceroughness. In our study we observed that MMPs’ adsorption was roughly correlated

28 F. Renò et al.
to polymer wettability as MMP-2 and MMP-9 bound to both hydrophilic (PMMAand PHEMA) and hydrophobic (PLA) polymers, suggesting that also structure andsolubility of the proteins and their interplay with each other could be involved in thecontrol of this phenomenon. It is noteworthy to observe that both inactive and activeenzymes were adsorbed onto the polymeric surface, suggesting the presence of insitu enzymatic activity. MMP-2 and MMP-9 are two enzymes able to cleave almostevery component of the extracellular matrix with different kinetics [19] and bothenzymes are able to modify the structure of cytokines, growth factors and receptorspresent on the cell surface [20, 21], thus acting as “modulators” of the immunolog-ical response [13] and of the neutrophil trafficking into inflamed tissues [22].
MMPs can be activated mainly by other MMPs [4] or by the plasminogen–plasminsystem [4]. Plasminogen is considered to play a central role in the inflammatoryresponse to biomaterials, as it is involved in the recruitment of macrophages andneutrophils at the site of biomaterial implantation [23]. In order to test if theactivity of MMP-2 and MMP-9 could be involved in neutrophil adhesion we usedPS (low level MMPs adsorbed) and PMMA (high level MMPs adsorbed) as modelsurfaces and we observed that in the presence of a MMPs inhibitor, the MMP-2/MMP-9 Inhibitor I, neutrophil adhesion to PMMA was strongly inhibited only inthe presence of plasma protein in the medium or preadsorbed onto the surfacesinvestigated. These findings therefore suggest that the presence and activity ofboth MMPs could be important for the neutrophil adhesion that normally occursminutes after biomaterial implantation [24]. The mechanism(s) of neutrophiladhesion facilitation by MMP-2 and MMP-9 could be related to the activation ofreceptors or other adhesion factors exposed on the neutrophil plasma membraneby MMPs activity. A similar effect has been observed for endothelin-1 (ET-1)that is cleaved by MMP-2 and rapidly up-regulates the expression of the Mac-1adhesion receptor (macrophage antigen receptor, CD11b/CD18) [22] that increasesneutrophil adhesion to the endothelial cells. Of course it is possible that the MMPsactivity is not essential for neutrophil adhesion. In fact it has been demonstrated thatneutrophil strongly adhere to PMMA through the Mac-1 adhesion receptor [11] andthis receptor can bind different proteins such as fibrinogen [25], vitronectin andfibronectin [24]. MMP-2 and MMP-9 present an unique protein structure amongthe MMPs, with the presence of fibronectin-like domains [26, 27] that could berecognized and bound by Mac-1. In order to clarify the mechanism(s) of action ofthe absorbed MMP-2 and MMP-9 activity further investigations are necessary and itis possible that in part both the proposed mechanisms collaborate to the unexpectedrole of these enzymes in the neutrophil adhesion on polymeric surfaces.
CONCLUSIONS
Our paper showed for the first time that two important enzymes, MMP-2 andMMP-9, present in human plasma and other body fluids, are adsorbed ontoimplantable biopolymers such as PLA, PMMA and PHEMA, both in the inactive

MMP adsorption onto biomedical polymers 29
and active form carrying an enzymatic activity on the polymers surface. Theunexpected finding of our research was that the presence and activity of MMP-2and MMP-9 plays an important role in the modulation of human neutrophil adhesionthat was strongly reduced by the addition of a MMPs inhibitor. Therefore, even ifthe MMPs mechanism(s) of action have to be further investigated, a role for theseenzymes in the inflammatory response to the biomaterials implantation has to beconsidered.
REFERENCES
1. M. B. Gorbet and M. V. Sefton, Biomaterials 25, 5681 (2004).2. C. J. Wilson, R. E. Clegg, D. I. Leavesley and M. J. Pearcy, Tissue Eng. 11, 18 (2005).3. Y. Takami, S. Yamane, K. Makinouchi, G. Otsuka, J. Glueck, R. Benkowski and Y. Nose,
J. Biomed. Mater. Res. 40, 24 (1998).4. G. I. Murray, J. Pathol. 195, 135 (2001).5. S. Arumugam, Y. C. Jang, C. Chen-Jensen, N. S. Gibran and F. F. Isik, Surgery 125, 587(1999).6. D. P. Papadopoulos, T. K. Makris, P. G. Krespi, M. Poulakou, O. G. Papazachou, A. N.
Hatzizacharias, D. Perrea and V. Votteas, Eur. Cytokine Netw. 16, 211 (2005).7. M. G. Tutton, M. L. George, S. A. Eccles, S. Burton, R. I. Swift and A. M. Abulafi, Int. J. Cancer
107, 541 (2003).8. M. Makela, T. Salo, V. J. Uitto and H. Larjava, J. Dent. Res. 73, 1397 (1994).9. K. M. Thrailkill, S. Kumar, C. K. Rosenberg, K. J. Auten and J. L. Fowlkes, Pediatr. Nephrol.
13, 223 (1999).10. G. Opdenakker, P. E. Van den Steen and J. Van Damme, Trends Immunol. 22, 571 (2001).11. H. Peltroche-Llacsahuanga, S. Schmidt, N. Schnitzler, R. Lutticken and G. Haase, J. Immunol.
Methods 258, 13 (2001).12. A. Boyum, Scand. J. Clin. Lab. Invest. 97, 77 (1968).13. G. Opdenakker, P. E. Van den Steen, B. Dubois, I. Nelissen, E. Van Coillie, S. Masure, P. Proost
and J. Van Damme, J. Leukoc. Biol. 69, 851 (2001).14. M. Santin, M. Cannas, M. A. Wassall and S. P. Denyer, J. Mater. Sci. Mater. Med. 9, 135 (1998).15. F. Reno, V. Aina, S. Gatti and M. Cannas, Biomaterials 25, 5594 (2005).16. R. M. Cornelius and J. L. Brash, Biomaterials 20, 341 (1999).17. J. Hong, K. Nilsson Ekdahl, H. Reynolds, R. Larsson and B. Nilsson, Biomaterials 20, 603
(1999).18. N. Yahyapour, C. Eriksson, P. Malmberg and H. Nygren, Biomaterials 25, 3171 (2004).19. C. Streuli, Curr. Opin. Cell Biol. 11, 634 (1999).20. J. L. Fowlkes and M. K. Winkler, Cytokine Growth Factor Rev. 13, 277 (2002).21. L. J. McCawley and L. M. Matrisian, Curr. Opin. Cell Biol. 13, 534 (2001).22. C. Fernandez-Patron, C. Zouki, R. Whittal, J. S. Chan, S. T. Davidge and J. G. Filep, FASEB J.
15, 2230 (2001).23. S. J. Busuttil, V. A. Ploplis, F. J. Castellino, L. Tang, J. W. Eaton and E. F. Plow, J. Thromb.
Haemost. 2, 1798 (2004).24. J. Lundahl, C. M. Skold, G. Hallden, M. Hallgren and A. Eklund, Scand. J. Immunol. 44, 143
(1996).25. V. K. Lishko, N. P. Podolnikova, V. P. Yakubenko, S. Yakovlev, L. Medved, S. P. Yadav and T. P.
Ugarova, J. Biol. Chem. 279, 44897 (2004).26. R. T. Aimes and J. P. Quigley, J. Biol. Chem. 270, 5872 (1995).27. G. I. Goldberg, I. E. Collier, A. Z. Eisen, G. A. Grant, B. L. Marmer and S. M. Wilhelm, Matrix
1 (Suppl.), 25 (1992).




















