
Metalloproteinases 2 and -9 activity during promotion and progression stages of rat liver...
11
ORIGINAL PAPER Metalloproteinases 2 and -9 activity during promotion and progression stages of rat liver carcinogenesis Kelly Silva Furtado Æ Paulo Wagner Pires Æ Luis Antonio Justulin Jr. Æ Maria Aparecida Marchesan Rodrigues Æ Sergio Luis Felisbino Æ Luis Fernando Barbisan Received: 26 September 2008 / Accepted: 17 November 2008 Ó Springer Science+Business Media B.V. 2008 Abstract Activity of metalloproteinases 2 and 9 (MMP-2 and 9) during promotion and progression of rat liver car- cinogenesis was investigated in a modified resistant hepatocyte model. Development of preneoplastic liver lesions positive for glutathione S-transferase 7-7-(GST-P 7- 7-positive PNL) and tumors besides hepatocytes positive for proliferating cell nuclear antigen (PCNA) were quan- tified and compared to MMP-2 and-9 activity using gelatin zymography. Marked increases in GST-P 7-7-positive PNL development, PCNA labeling indices, MMP-2 (pro, inter- mediate and active forms) and pro-MMP-9 activity were observed after proliferative stimulus induced by 2-ace- tylaminofluorene (2-AAF) exposure cycles. After 2-AAF withdrawal, increase in MMP-2 activity was detected only in neoplastic mixed lesions, whereas active MMP-9 was increased in both PLN and neoplastic tissues. Our findings suggest that MMP-2 may be associated with proliferative events induced by 2-AAF rather than with selective growth of PNL and that MMP-9 could be associated with pro- gression of PNL and neoplastic mixed lesions. Keywords Rat liver carcinogenesis Resistant hepatocyte model Metalloproteinases Preneoplastic and neoplastic lesions Introduction Rat liver carcinogenesis is a multistep and complex process that includes initiation, promotion and progression stages (Pitot 2001). Foci of altered hepatocytes and hyperplastic nodules have been described as putative preneoplastic lesions (PNL) in various experimental models of rat liver carcinogenesis and it is argued that they are capable of progressing to hepatocellular adenomas and carcinomas (Bannasch and Zerban 1992; Hasegawa and Ito 1994). Recently, different types of PNL with similar morpholog- ical and biochemical changes in the hepatocellular phenotype were identified in human chronic liver diseases associated with hepatocellular carcinoma (Su and Ban- nasch 2003; Libbrecht et al. 2005). The resistant hepatocyte (RH) model is one of the most classical for the study of the different steps of rat liver carcinogenesis (Farber and Sarma 1987). In this model, diethylnitrosamine (DEN)-initiating agent induces single cells and small foci of putative initiated hepatocytes that are promoted with 2-acetylaminofluorene (2-AAF), asso- ciated with a 70% partial hepatectomy (PH) (Farber and Sarma 1987). 2-AAF is a mito-inhibitor to normal hepa- tocytes but does not inhibit a subpopulation of the glutathione S-transferase (GST-P) 7-7-positive hepato- cytes. Partial hepatectomy acts as a mitotic stimulus resulting in activation of hepatic stem cells (oval cells) and a rapid growth of the initiated cells subpopulation into larger GST-P 7-7-positive lesions. Some lesions eventually progress into grossly visible nodules and, over months, K. S. Furtado M. A. M. Rodrigues School of Medicine, Department of Pathology, UNESP Sa ˜o Paulo State University, Botucatu, SP 18618-000, Brazil P. W. Pires Department of Cell Biology, Institute of Cell Biology, UNICAMP, Campinas, SP 13083-950, Brazil K. S. Furtado L. A. Justulin Jr. S. L. Felisbino L. F. Barbisan (&) Department of Morphology, Institute of Biosciences, UNESP Sa ˜o Paulo State University, Botucatu, SP 18618-000, Brazil e-mail: [email protected] 123 J Mol Hist DOI 10.1007/s10735-008-9206-x
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Metalloproteinases 2 and -9 activity during promotion and progression stages of rat liver...

ORIGINAL PAPER
Metalloproteinases 2 and -9 activity during promotionand progression stages of rat liver carcinogenesis
Kelly Silva Furtado Æ Paulo Wagner Pires Æ Luis Antonio Justulin Jr. ÆMaria Aparecida Marchesan Rodrigues Æ Sergio Luis Felisbino ÆLuis Fernando Barbisan
Received: 26 September 2008 / Accepted: 17 November 2008
� Springer Science+Business Media B.V. 2008
Abstract Activity of metalloproteinases 2 and 9 (MMP-2
and 9) during promotion and progression of rat liver car-
cinogenesis was investigated in a modified resistant
hepatocyte model. Development of preneoplastic liver
lesions positive for glutathione S-transferase 7-7-(GST-P 7-
7-positive PNL) and tumors besides hepatocytes positive
for proliferating cell nuclear antigen (PCNA) were quan-
tified and compared to MMP-2 and-9 activity using gelatin
zymography. Marked increases in GST-P 7-7-positive PNL
development, PCNA labeling indices, MMP-2 (pro, inter-
mediate and active forms) and pro-MMP-9 activity were
observed after proliferative stimulus induced by 2-ace-
tylaminofluorene (2-AAF) exposure cycles. After 2-AAF
withdrawal, increase in MMP-2 activity was detected only
in neoplastic mixed lesions, whereas active MMP-9 was
increased in both PLN and neoplastic tissues. Our findings
suggest that MMP-2 may be associated with proliferative
events induced by 2-AAF rather than with selective growth
of PNL and that MMP-9 could be associated with pro-
gression of PNL and neoplastic mixed lesions.
Keywords Rat liver carcinogenesis � Resistant
hepatocyte model � Metalloproteinases � Preneoplastic
and neoplastic lesions
Introduction
Rat liver carcinogenesis is a multistep and complex process
that includes initiation, promotion and progression stages
(Pitot 2001). Foci of altered hepatocytes and hyperplastic
nodules have been described as putative preneoplastic
lesions (PNL) in various experimental models of rat liver
carcinogenesis and it is argued that they are capable of
progressing to hepatocellular adenomas and carcinomas
(Bannasch and Zerban 1992; Hasegawa and Ito 1994).
Recently, different types of PNL with similar morpholog-
ical and biochemical changes in the hepatocellular
phenotype were identified in human chronic liver diseases
associated with hepatocellular carcinoma (Su and Ban-
nasch 2003; Libbrecht et al. 2005).
The resistant hepatocyte (RH) model is one of the most
classical for the study of the different steps of rat liver
carcinogenesis (Farber and Sarma 1987). In this model,
diethylnitrosamine (DEN)-initiating agent induces single
cells and small foci of putative initiated hepatocytes that
are promoted with 2-acetylaminofluorene (2-AAF), asso-
ciated with a 70% partial hepatectomy (PH) (Farber and
Sarma 1987). 2-AAF is a mito-inhibitor to normal hepa-
tocytes but does not inhibit a subpopulation of the
glutathione S-transferase (GST-P) 7-7-positive hepato-
cytes. Partial hepatectomy acts as a mitotic stimulus
resulting in activation of hepatic stem cells (oval cells) and
a rapid growth of the initiated cells subpopulation into
larger GST-P 7-7-positive lesions. Some lesions eventually
progress into grossly visible nodules and, over months,
K. S. Furtado � M. A. M. Rodrigues
School of Medicine, Department of Pathology, UNESP Sao
Paulo State University, Botucatu, SP 18618-000, Brazil
P. W. Pires
Department of Cell Biology, Institute of Cell Biology,
UNICAMP, Campinas, SP 13083-950, Brazil
K. S. Furtado � L. A. Justulin Jr. � S. L. Felisbino �L. F. Barbisan (&)
Department of Morphology, Institute of Biosciences, UNESP
Sao Paulo State University, Botucatu, SP 18618-000, Brazil
e-mail: [email protected]
123
J Mol Hist
DOI 10.1007/s10735-008-9206-x

tumors arise (Sell and Dunsford 1989; Tiwawech et al.
1991).
The liver extracellular matrix (ECM) is not only a
passive structural support, since it plays key roles in pro-
viding a structural framework and maintaining the
differentiated phenotype and normal function of hepato-
cytes, sinusoidal, endothelial and stellate cells (Martinez-
Hernandez and Amenta 1993). ECM turnover is a vital step
in tissue remodeling that accompanies both physiological
and pathological processes, including aging, fibrosis and
tumor development (Brew et al. 2000; Visse and Nagase
2003; Chakraborti et al. 2003; Vihinen et al. 2005; Ra and
Parks 2007). Matrix metalloproteinases (MMPs) are a
family of zinc-dependent endopeptidases that degrade most
ECM components in both normal and pathophysiological
processes, and their activity is tightly regulated by the
tissue inhibitors of metalloproteinases (TIMPs) (Brew et al.
2000; Visse and Nagase 2003; Chakraborti et al. 2003;
Vihinen et al. 2005; Ra and Parks 2007). MMPs are syn-
thesized as inactive pro-forms which require subsequent
cleavage in order to become active (Visse and Nagase
2003; Ra and Parks 2007). Based on their substrate spec-
ificity and structural similarity, MMPs are often classified
into six subgroups: collagenases, gelatinases, stromelysins,
matrilysin, membrane-type MMPs and other MMPs (Vi-
hinen et al. 2005; Ra and Parks 2007). Perfect regulation of
ECM degradation is essential for tissue homeostasis
maintenance, and an imbalance between TIMPs and MMPs
rates may participate in tumor progression and metastasis
in several tissues and organs (Brew et al. 2000; Chakraborti
et al. 2003).
A small number of studies investigated the potential role
of MMPs in experimental liver carcinogenesis. Wood and
Archer (2001) examined the role of gelatinases during the
early PNL growth in both Fischer 344 (susceptible) and
Copenhagen—Cop (resistant) rats. These authors reported
a higher increase of hepatic MMP-2 and -9 activity in
Fischer 344 rats than in Cop rats, nevertheless no inhibition
on liver regeneration or early GST-P 7-7-positive PNL
growth after PH was observed when the Fischer 344 rats
were treated with batimastat, a MMP inhibitor. Thus, the
increase of hepatic gelatinases activity could be non
essential for the early PNL expansion in DEN-initiated and
2AAF-treated rats. Gao et al. (2002) described an increase
in MMP-2 mRNA and in pro and active MMP-2 and pro-
MMP-9 activity during HCC development in rats initiated
with DEN.
Therefore, the present study was designed to evaluate
the activity and in situ immunohistochemistry expression
of metalloproteinases 2 and 9 at the promotion/progression
stages of liver carcinogenesis in male Wistar rats submitted
to a modified RH model. In addition, MMP-2 and -9
findings were compared to hepatic cell proliferation, p21ras
and cyclooxygenase-2 (COX-2) protein immunoreactivity,
PNL development and tumor phenotypes.
Materials and methods
Animals and treatments
Four-week-old male Wistar rats were obtained from Multi-
disciplinary Center for Biological Investigation (CEMIB,
UNICAMP Campinas-SP, Brazil) and housed in polypro-
pylene cages (five animals/cage) covered with metallic grids
in a room maintained at 22 ± 2�C, 55 ± 10% humidity and
with a 12-h light-dark cycle. They were fed commercial
Purina chow (LABINA, Paulınia, SP, Brazil) and water ad
libitum during a 2-week acclimatization period. Then, the
animals were randomly allocated into four experimental
groups (Fig. 1): non-treated group (G1), a single dose of
200 mg/kg b.w. of diethylnitrosamine (DEN, Sigma-
Aldrich Co., St. Louis Mo, USA)-initiated group (G2),
DEN-initiated plus 0.02% 2-acetylaminofluorene (2-AAF,
Sigma-Aldrich Co., St. Louis Mo, USA, 1 cycle of 3 weeks
and 3 cycles of 2 weeks)-promoted group (G3) and 2-AAF-
treated group (G4). All animals were subjected to 70%
partial hepatectomy (PH) at week 3 and sacrificed at weeks
8, 20 or 35. Body weight and food/water intake were mea-
sured twice a week during the entire experimental period.
In this study, as Wistar non-isogenic strain is more
resistant than Fischer 344 isogenic strain to liver carcino-
genesis (Asamoto et al. 1989; Pascale et al. 2002), we
introduced three additional cycles of treatment with 2-AAF
to increase the development of hepatic preneoplastic lesions
and the tumor burden in male Wistar rats. Besides DEN plus
2-AAF-treated group (main group), we also introduced all
control groups including non-treated and DEN or 2-AAF-
treated groups. The protocols used were consistent with the
Ethical Principles for Animal Research adopted by the
Brazilian College of Animal Experimentation (COBEA).
puorG
1G
2G
3G
4G
86431 412101 028161 53skeeW
ss
s
s
s
s
s
s
s
s
s
s
staRforebmuN=n,ecifircaS=s
teidlasaB
FAA-2%20.0sulpteidlasaB
.p.i,.w.bgk/gm002NED
.p.i%9.0lCaN
ymotcetapehlaitrap%07
02=n
02=n
03=n
03=n
Fig. 1 Experimental design (for details see ‘‘Materials and
methods’’)
J Mol Hist
123

Tissue processing, histology and immunohistochemical
procedures
Immediately before necropsy, blood samples were collected
and the serum levels of alanine aminotransferase (ALT) and
albumin were measured spectrometrically to monitor
hepatocellular injury and function (Ramaiah 2007). At
necropsy, the liver was removed and weighted, and repre-
sentative samples were either fixed in 4% phosphate-
buffered formalin during 24 h for paraffin embedding or
frozen in liquid nitrogen for gelatin zymography. The par-
affin blocks were cut into 5-lm-thick sections and stained
with hematoxylin–eosin (HE) for histological analysis,
Gomori’s silver impregnation for reticulin fibers evaluation
and for immunohistochemical analysis of glutathione S-
transferase P form (GST-P 7-7), proliferating cell nuclear
antigen (PCNA) and metalloproteinases 2 (MMP-2) or 9
(MMP-9), p21ras and cyclooxygenase-2 (COX-2) markers.
In HE staining, PNL and neoplastic lesions (hepatocel-
lular adenoma or carcinoma) were classified according to
previous published criteria (Bannasch and Zerban 1992).
PCNA, GST-P 7-7, MMP-2, MMP-9, p21ras and COX-2
expression were immunohistochemically detected using the
avidin–biotin–peroxidase method. Briefly, deparaffinized
5-lm-thick serial liver sections on poly-L-lysine coated
slides were treated with 3% H2O2 in phosphate-buffered
saline for 15 min, nonfat milk for 60 min, anti-PCNA
(Dako Corporation, Carpinterie, CA, USA, clone PC10,
1:200 dilution), anti-GST-P 7-7 (Medical and Biological
Laboratories Co., Tokyo, Japan, clone 311, 1:1000 dilu-
tion), anti-MMP-2 and MMP-9 (Santa Cruz Biotechnology,
California, USA, clones C-19 and C-20, respectively, 1:200
dilution), p21ras (Santa Cruz, clone SC 32, 1:25 dilution)
or COX- 2 (Cayman Chemical Co, MI, EUA, 1:200 dilu-
tion) antibodies, biotinylated horse anti-mouse IgG, anti-
goat or anti-rabbit IgG antibodies (Dako Corporation,
Carpinterie, CA, USA, 1:200 dilution) for 60 min, and
avidin–biotin–peroxidase solution (Vector Laboratories
Inc, CA, USA, 1:1:50 dilution). Antigen retrieval for
MMP-2 and 9, p21ras and COX-2 was performed using
0.01 M citrate buffer (pH 6.0) boiling in microwave oven
(1,300 W) twice for 5 min each. Chromogen color was
accomplished with 3,30-diaminobenzidine tetrahydrochro-
ride (DAB, Sigma–Aldrich Co., St. Louis MO, USA) as the
substrate to demonstrate the sites of peroxidase binding.
The slides were counterstained with Harris’s hematoxylin.
Gelatin zymography for MMP-2 and MMP-9
All hepatic samples stored in liquid nitrogen for gelatin
zymography analysis were histologically checked for the
presence of preneoplastic and/or neoplastic lesions. Samples
from liver tumors did not contain surrounding parenchyma,
since they were easily identified macroscopically and iso-
lated. Thus, we compared liver samples containing liver
tumors or an extensive development of PNL (G3 and G4
groups), or few PNL lesions (G2 group) or unaltered liver
parenchyma (G1 group).
Frozen liver samples (*300 mg) were mechanically
homogenized in 50 mM Tris buffer pH 7.5 plus 1% triton-
X 100 and CaCl2 10 mM with a Polytron for 30 s at 4�C,
centrifuged and protein extracted on supernatant was
quantified by Bradford method. Aliquots (30 lg protein)
from liver extracts (non-altered, containing PNL or neo-
plastic lesions) were subjected to electrophoresis in gelatin
containing polyacrylamide (8% acrylamide) gels in the
presence of SDS under non-reducing conditions (Pires
et al. 2008). The gelatin substrate was present at 0.1% final
concentration in the gel. The gels (0.75 mm thick) were
electrophoresed for 2 h at 100 V, 4�C, in a Bio-Rad
MiniProtean II system (Bio-Rad Laboratories, Inc., Rich-
mond, CA, USA). Following electrophoresis, the gels were
washed by gentle shaking at room temperature with 2.5%
Triton X-100 (two changes) for 1 h. The gels were incu-
bated overnight (16 h) in 50 mM Tris–HCl (pH 8.4)
containing 5 mM CaCl2 and 1 lM ZnCl2 at 37�C. After
incubation, the gels were stained by Coomassie Brilliant
Blue R-250. Areas of proteolysis appeared as clear zones
against a blue background. Molecular mass determinations
were made with reference to prestained protein standards
(Bio-Rad Laboratories) co-electrophoresed into the gels.
The integrated optical density (IOD) spectrophotometry
was determinated in an Image Master VDS version 3.0
(Pharmacia Biotechnology, Piscataway, NJ, USA). The
values of IOD were analyzed statistically and plotted in
histograms.
GST-P positive PNL morphometry and PCNA labeling
analysis
PNL was detected by GST-P expression in liver sections
(3). GST-P 7-7-positive PNL were measured using a Nikon
photomicroscope (Microphot-FXA, Tokyo, Japan) con-
nected to a KS-300 apparatus (Kontron Elektronic,
Munich, Germany). Data from morphometric analysis of
putative preneoplastic liver lesions positive for GST-P 7-7
are frequently presented as number per liver area (cm2) or
per volume (cm3). Both two- and three-dimensionally
expressed quantitative results for GST-P 7-7 positive PNL
were found to be adequate to demonstrate the modifying
potential of test chemicals on chemically induced hepato-
carcinogenesis in rodents (Imaida et al. 1989). As the
immunohistochemical detection of GST-P 7-7-positive
PNL was performed in various fragments of all liver lobes
reminiscent after partial hepatectomy, our results were
presented as number per cm2, mean size (mm2) and
J Mol Hist
123

percentage of liver area occupied by these lesions (% liver
area). (Pinheiro et al. 2003; Pires et al. 2008).
PCNA labeling indices (PCNA LI %) for each animal
were estimated by counting the number of immunohisto-
chemically labeled hepatocytes nuclei (S-Phase) per total
number of counted hepatocytes (*2,000) in normal or
bearing PNL parenchyma (Pinheiro et al. 2003; Pires et al.
2008). PCNA LI% was also measured in neoplastic lesions
diagnosed at week 35.
Statistical analysis
Statistical analysis was performed using the Jandel Sigma
Stat Software (Jandel Corporation, San Rafael, CA, USA).
The body weight, body weight-gain, absolute and relative
liver, PCNA-labeling, GST-P 7-7-positive PNL and IOD
data and tumor multiplicity were analyzed by ANOVA test
or Kruskal–Wallis test. The incidence of different types of
PNL and neoplastic lesions was examined using v2 test or
the Fischer exact test. Data were considered statistically
significant when P \ 0.05.
Results
General findings
Body weight from animals of non-treated group (G1) and
DEN-treated group (G2) progressively increased during all
experimental periods. However, there was a marked
reduction in food intake (data not shown) and body-weight
gain in DEN plus 2-AAF-treated group (G3) and 2-AAF-
treated group (G4) during the cycles of 2-AAF treatment
(P \ 0.001, Fig. 2). A significant reduction in final body
weight was observed in the DEN plus 2-AAF-treated group
(G3) in all periods analyzed (weeks 8, 20 and 35)
(P \ 0.001, Table 1). The reduced food intake and body
weight gain observed in the 2-AAF-treated groups (G3 and
G4) is due to severe hepatoxicity induced by 2-AAF
treatment (Bitsch et al. 2000).
The relative liver weights from 2-AAF-treated groups
(G3 and G4) were significantly higher than the non-treated
group (G1) and DEN-treated group (G2) mainly at weeks
20 and 35 (P \ 0.001, Table 1). The alanine aminotrans-
ferase (ALT) and albumin serum levels did not differ
among the experimental groups at weeks 8, 20 and 35
(Table 1).
Preneoplastic and neoplastic lesions
The main hepatic lesions observed in carcinogen-treated
animals were PNL characterized by altered foci of hepato-
cytes and hyperplastic nodules with clear cell and/or
eosinophilic or basophilic cell phenotypes. Neoplastic
lesions included adenomas, hepatocarcinomas (HCC), cho-
langiomas, cholangiocarcinomas and mixed neoplastic
lesions like hepatocholangiocellular adenomas and carci-
nomas (Table 2). Benign and malignant lesions were
identified at weeks 20 and 35 predominantly in the DEN plus
2-AAF-treated group (G3) (Table 2). Some animals from
DEN-treated groups (G2 and G3) and/or 2AAF-treated
groups (G3 and G4) developed kidney adenomas and car-
cinomas or sebaceous and squamous cell carcinomas in the
Zymbal’s gland, respectively. These chemically induced
neoplasms are demonstrative of the organotropic activity of
DEN and 2-AAF on non-liver targets (Verna et al. 1996 a, b).
A rapid development of GST-P 7-7-positive PNL (i.e.,
mainly in mean size and percentage of occupied liver area)
and neoplastic lesions (i.e., mixed lesions, adenomas, HCC
and cholangioma and cholangiocarcinoma) was observed in
this modified RH model (Table 3). 2-AAF is a genotoxic
hepatocarcinogen with both initiating (weak) and promoting
(strong) activities on rat liver carcinogenesis (Tiwawech
et al. 1991, Verna et al. 1996b). Specially at week 20, the
promoting potential of this hepatocarcinogen was well
characterized in the 2-AAF-treated group with an increase of
6.75 times the liver area occupied by PNL when compared to
week 8 (Table 3). In 2-AAF-treated animals, the prolifera-
tive stimulus induced by partial hepatectomy resulted in oval
cell hyperplasia and an expansive growth of initiated cells
into larger GST-P 7-7-positive PNL in the absence of
hepatic inflammation, necrosis and fibrosis. However, there
was a reduction in the expansive growth of GST-P 7-7-
positive PNL after 2-AAF withdrawal (Table 3), probably
as a consequence of a decrease in cell proliferation and
remodeling (Tiwawech et al. 1991). In contrast, a modest
development of GST-P 7-7-positive lesions and rare GST-P
7-7-positive PNL were observed in the liver from DEN-
treated group (G2) and non-treated group (G1), respectively,
0
100
200
300
400
500
600
0 2 4 6 8 10 12 14 16 18 20 30 32 35
(G1) Control
(G2) DEN
(G3) DEN+2-AAF
(G4) 2-AAF
Weeks
)g(thgie
wydob
naeM
2-AAF 2-AAF 2-AAF2-AAF
Fig. 2 Effects of treatments on body weight. A significant reduction
in body weight was observed in groups treated with DEN plus 2-AAF.
(G3 \ G1 = G2 = G4, P \ 0.001) DEN, diethylnitrosamine
(200 mg/kg, b.w., i.p.); 2-AAF, 2-acethylaminofluorene (0.02% in
diet)
J Mol Hist
123
mainly at weeks 20 and 35 (Table 3). The finding of few
GST-P 7-7-positive PNL in the non-treated group (G1) has
been observed in senile rats (Hasegawa and Ito 1994).
The distribution of reticular fibers in the non-affected
liver parenchyma was restricted to Disse’s spaces, adjacent
to centrolobular veins and in the portal space (data not
shown). An increase in both thickness and number of
reticular fibers was observed at the compressive border that
embraces some neoplastic lesions like adenomas, HCC and
mixed neoplastic lesions, especially in the DEN plus
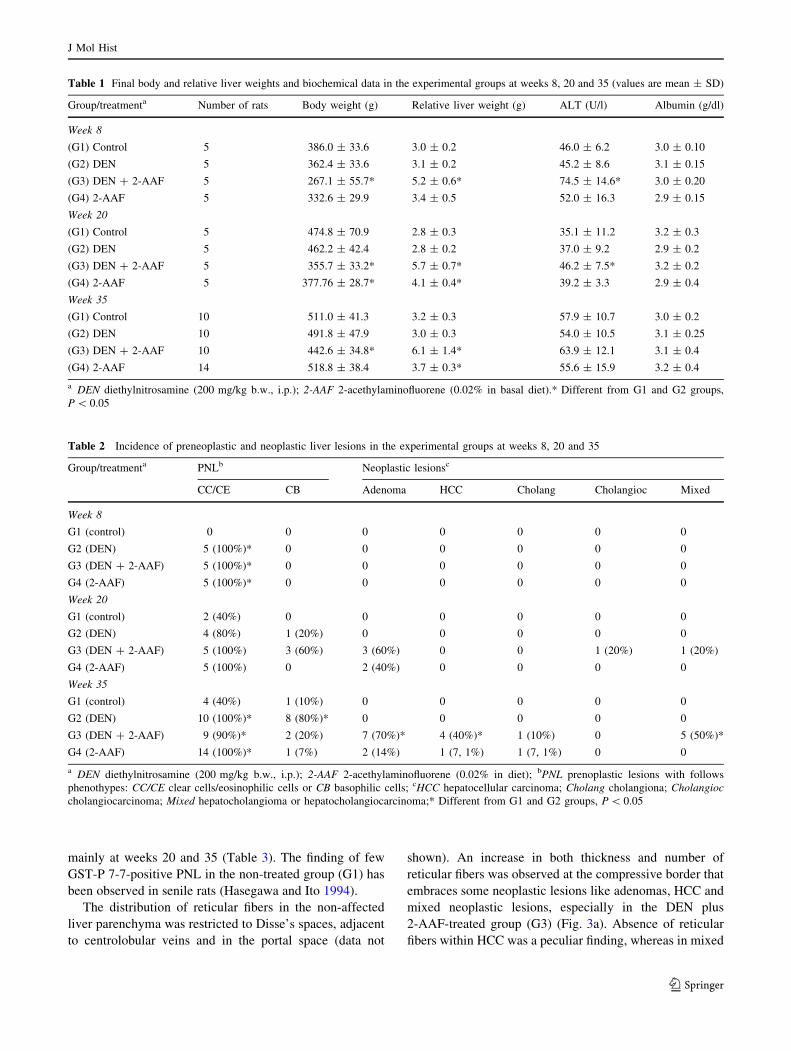
2-AAF-treated group (G3) (Fig. 3a). Absence of reticular
fibers within HCC was a peculiar finding, whereas in mixed
Table 1 Final body and relative liver weights and biochemical data in the experimental groups at weeks 8, 20 and 35 (values are mean ± SD)
Group/treatmenta Number of rats Body weight (g) Relative liver weight (g) ALT (U/l) Albumin (g/dl)
Week 8
(G1) Control 5 386.0 ± 33.6 3.0 ± 0.2 46.0 ± 6.2 3.0 ± 0.10
(G2) DEN 5 362.4 ± 33.6 3.1 ± 0.2 45.2 ± 8.6 3.1 ± 0.15
(G3) DEN ? 2-AAF 5 267.1 ± 55.7* 5.2 ± 0.6* 74.5 ± 14.6* 3.0 ± 0.20
(G4) 2-AAF 5 332.6 ± 29.9 3.4 ± 0.5 52.0 ± 16.3 2.9 ± 0.15
Week 20
(G1) Control 5 474.8 ± 70.9 2.8 ± 0.3 35.1 ± 11.2 3.2 ± 0.3
(G2) DEN 5 462.2 ± 42.4 2.8 ± 0.2 37.0 ± 9.2 2.9 ± 0.2
(G3) DEN ? 2-AAF 5 355.7 ± 33.2* 5.7 ± 0.7* 46.2 ± 7.5* 3.2 ± 0.2
(G4) 2-AAF 5 377.76 ± 28.7* 4.1 ± 0.4* 39.2 ± 3.3 2.9 ± 0.4
Week 35
(G1) Control 10 511.0 ± 41.3 3.2 ± 0.3 57.9 ± 10.7 3.0 ± 0.2
(G2) DEN 10 491.8 ± 47.9 3.0 ± 0.3 54.0 ± 10.5 3.1 ± 0.25
(G3) DEN ? 2-AAF 10 442.6 ± 34.8* 6.1 ± 1.4* 63.9 ± 12.1 3.1 ± 0.4
(G4) 2-AAF 14 518.8 ± 38.4 3.7 ± 0.3* 55.6 ± 15.9 3.2 ± 0.4
a DEN diethylnitrosamine (200 mg/kg b.w., i.p.); 2-AAF 2-acethylaminofluorene (0.02% in basal diet).* Different from G1 and G2 groups,
P \ 0.05
Table 2 Incidence of preneoplastic and neoplastic liver lesions in the experimental groups at weeks 8, 20 and 35
Group/treatmenta PNLb Neoplastic lesionsc
CC/CE CB Adenoma HCC Cholang Cholangioc Mixed
Week 8
G1 (control) 0 0 0 0 0 0 0
G2 (DEN) 5 (100%)* 0 0 0 0 0 0
G3 (DEN ? 2-AAF) 5 (100%)* 0 0 0 0 0 0
G4 (2-AAF) 5 (100%)* 0 0 0 0 0 0
Week 20
G1 (control) 2 (40%) 0 0 0 0 0 0
G2 (DEN) 4 (80%) 1 (20%) 0 0 0 0 0
G3 (DEN ? 2-AAF) 5 (100%) 3 (60%) 3 (60%) 0 0 1 (20%) 1 (20%)
G4 (2-AAF) 5 (100%) 0 2 (40%) 0 0 0 0
Week 35
G1 (control) 4 (40%) 1 (10%) 0 0 0 0 0
G2 (DEN) 10 (100%)* 8 (80%)* 0 0 0 0 0
G3 (DEN ? 2-AAF) 9 (90%)* 2 (20%) 7 (70%)* 4 (40%)* 1 (10%) 0 5 (50%)*
G4 (2-AAF) 14 (100%)* 1 (7%) 2 (14%) 1 (7, 1%) 1 (7, 1%) 0 0
a DEN diethylnitrosamine (200 mg/kg b.w., i.p.); 2-AAF 2-acethylaminofluorene (0.02% in diet); bPNL prenoplastic lesions with follows
phenothypes: CC/CE clear cells/eosinophilic cells or CB basophilic cells; cHCC hepatocellular carcinoma; Cholang cholangiona; Cholangioccholangiocarcinoma; Mixed hepatocholangioma or hepatocholangiocarcinoma;* Different from G1 and G2 groups, P \ 0.05
J Mol Hist
123

neoplastic lesions an increase in thickness and number of
reticular fibers was observed in areas associated with
cholangiocellular lesions (Fig. 3d).
MMP-2 and-9, p21ras and COX-2 immunoreactivity
and gelatinases
The immunolabeling for MMP-2 and -9, both pro and
active forms, performed at week 35 showed that these
enzymes were expressed in non-parenchymal cells, in
hepatocytes from PNL and adenomas (data not shown), and
hepatocholangiocellular adenomas and carcinomas
(Fig. 3b, c, e, f). All HCC analyzed at week 35 did not
expressed MMP-2 and -9. However, the immunoreactivity
for these MMPs was observed in the surrounding cells at
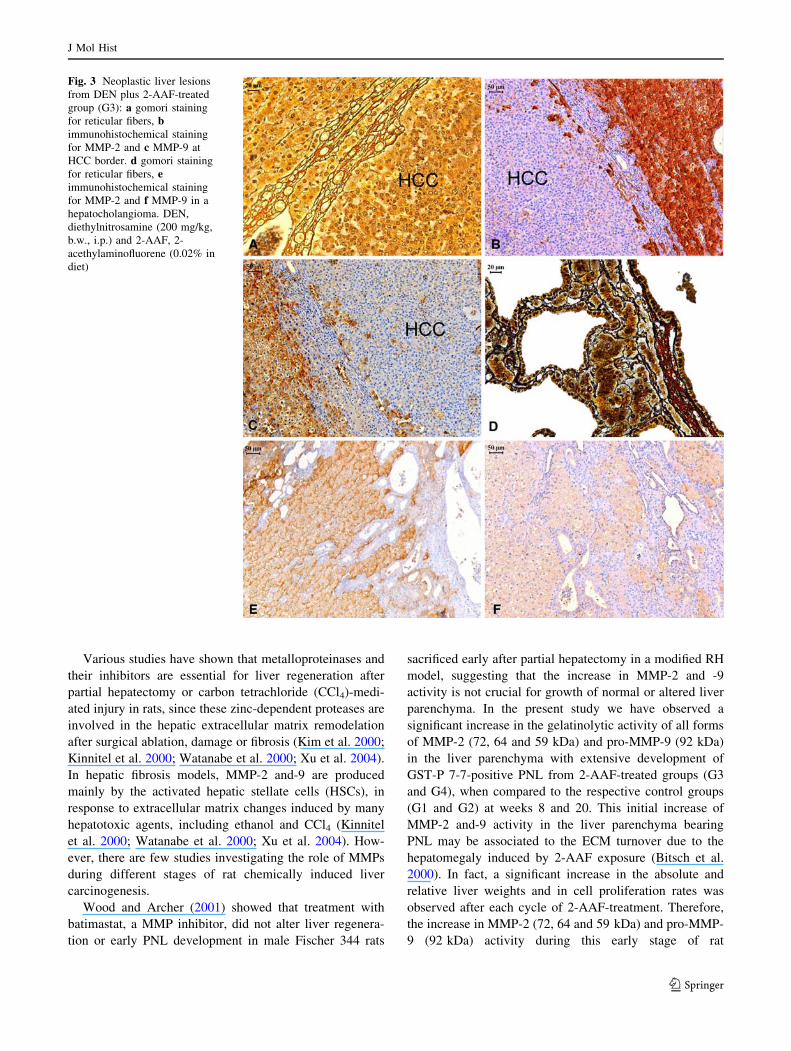
the tumor margin (Fig. 3b, C). At week 35, co-expression
of p21ras, COX-2 and MMP-9 was observed in most
neoplastic lesions from G3 and G4 groups. However, in
cholangiocellular areas of mixed lesions and in cholan-
giomas and cholangiocarcinomas, the MMP-9
immunoreactivity in bile ductal neoplastic cells was not
associated to p21ras and COX-2 co-expression (Fig. 4).
Data regarding metalloproteinases 2 and 9 (MMP-2 and
-9) activity in all experimental periods are shown in
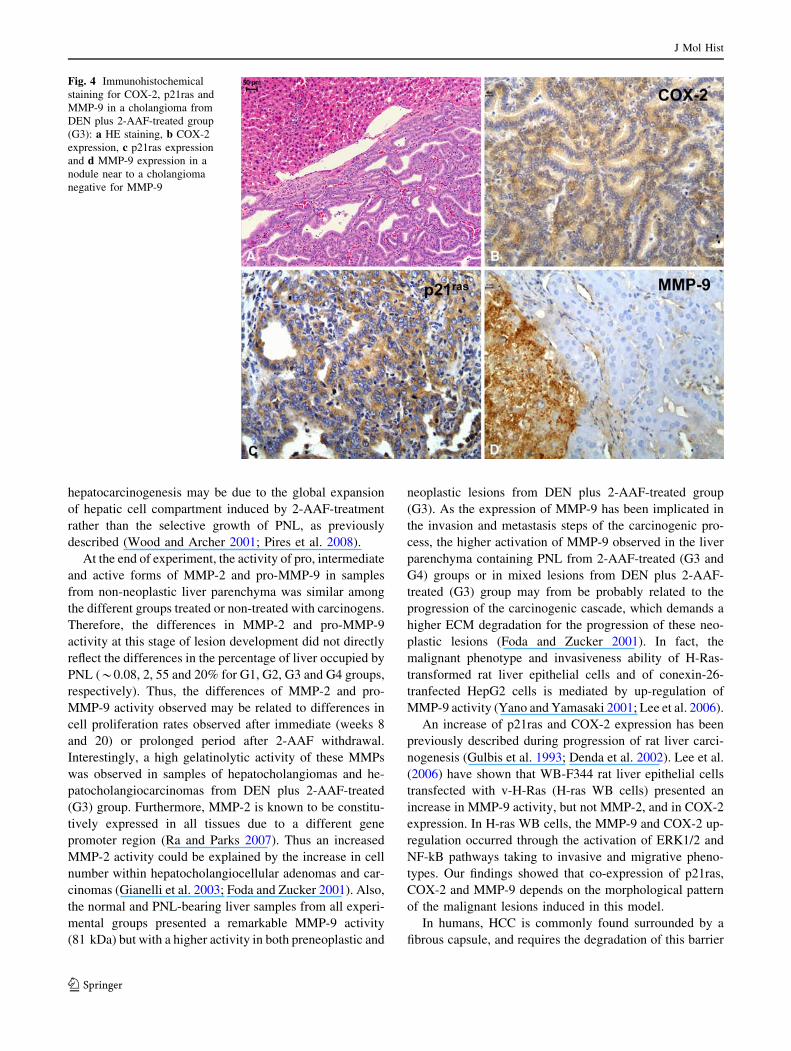
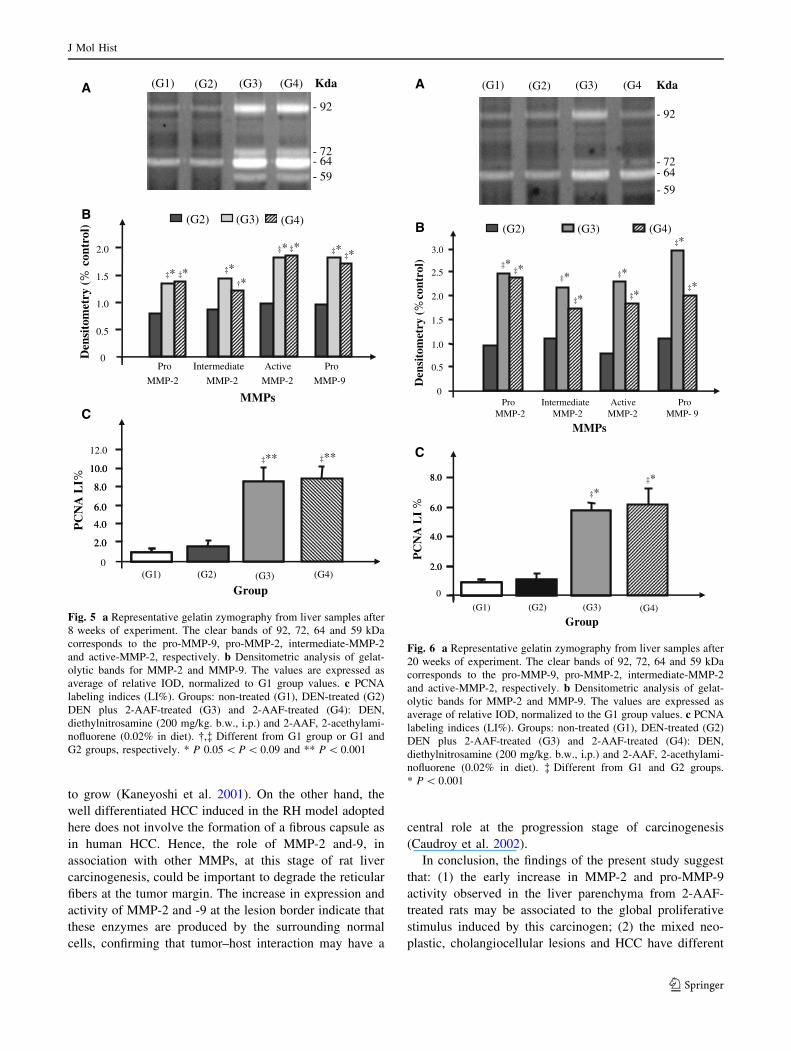
Figs. 5, 6 and 7. A significant increase in the activity of pro
(72 kDa), intermediate (64 kDa) and active forms (59 kDa)
of MMP-2 and pro-MMP-9 (92 kDa) was observed in both
2-AAF-treated groups (G3 and G4) at weeks 8 and 20
(0.002 \ P \ 0.05) but not at week 35, after a 15-week
period of 2-AAF withdrawal. However, in neoplastic
lesions (i.e., predominantly hepatocholangiocellular ade-
nomas and carcinomas) these enzymatic activities
remained high after the 2-AAF withdrawal. The active
form (81 kDa) of MMP-9 was detected only at week 35,
especially in the liver parenchyma from 2-AAF-treated
groups (G3 and G4) and in mixed neoplastic lesions from
DEN plus 2-AAF-treated group (G3).
PCNA analysis
The rates of hepatic cell proliferation were significantly
higher in liver parenchyma from 2-AAF-treated groups (G3
and G4) than in the respective control groups (G1 and G2)
in all experimental periods (0.05 \ P \ 0.001, Figs. 5c, 6c
and 7c). A significant reduction in PCNA LI% was
observed in the liver parenchyma from 2-AAF-treated
groups (G3 and G4) at week 35, after a 15-week period of
2-AAF withdrawal. In contrast, the neoplastic lesions from
DEN plus 2-AAF-treated group (G3) maintained high
PCNA labeling indices at week 35 (Fig. 7c).
Discussion
The present study showed the dynamic changes in
metalloproteinases -2 and -9 activity during the promotion
and progression stages of chemically induced liver carci-
nogenesis in male Wistar rats submitted to a modified
resistant hepatocyte (RH) model.
Table 3 Analysis of GST-P 7-7-positive lesions in liver of the different experimental groups at weeks 8, 20 and 35 (values are mean ± SD)
Group/treatmenta Number of rats GST-P 7-7-positive datab
Number/cm2 Mean size (mm2) % Liver area
Week 8
G1 (Control) 05 0 0 0
G2 (DEN) 05 7.87 ± 4.04* 0.05 ± 0.01* 0.53 ± 0.33*
G3 (DEN ? 2-AAF) 05 34.71 ± 15.99*,** 1.74 ± 0.76*,** 62.69 ± 4.19*,**
G4 (2-AAF) 05 5.38 ± 2.41*,**,*** 0.86 ± 0.43*,**,*** 3.96 ± 1.05*,**,***
Week 20
G1 (Control) 05 0.66 ± 1.23 0.01 ± 0.01 0.02 ± 0.04
G2 (DEN) 05 24.73 ± 5.41* 0.02 ± 0.003* 1.11 ± 0.74*
G3 (DEN ? 2-AAF) 05 35.01 ± 5.52*,** 0.44 ± 0.08*,** 69.27 ± 9.59*,**
G4 (2-AAF) 05 27.31 ± 7.11*,*** 0.28 ± 0.16*,** 26.99 ± 16.22*,**,***
Week 35
G1 (Control) 10 1.83 ± 1.61 0.01 ± 0.01 0.08 ± 0.07
G2 (DEN) 10 25.98 ± 4.81* 0.03 ± 0.01 2.07 ± 0.56*
G3 (DEN ? 2-AAF) 10 20.11 ± 9.79* 0.51 ± 0.10*,** 55.20 ± 11.62*,**
G4 (2-AAF) 14 32.36 ± 5.95*,*** 0.17 ± 0.08*,**,*** 20.64 ± 8.95*,**,***
a DEN diethylnitrosamine (200 mg/kg b.w., i.p.); 2-AAF 0.02% 2-acethylaminofluorene in diet; bGST-P-positive lesions PNL and neoplastic
lesions (adenoma, hepatocellular carcinoma and mixed lesions); *,**,*** Different from G1, G2 or G3, respectively, 0.005 \ P \ 0.001
J Mol Hist
123
Various studies have shown that metalloproteinases and
their inhibitors are essential for liver regeneration after
partial hepatectomy or carbon tetrachloride (CCl4)-medi-
ated injury in rats, since these zinc-dependent proteases are
involved in the hepatic extracellular matrix remodelation
after surgical ablation, damage or fibrosis (Kim et al. 2000;
Kinnitel et al. 2000; Watanabe et al. 2000; Xu et al. 2004).
In hepatic fibrosis models, MMP-2 and-9 are produced
mainly by the activated hepatic stellate cells (HSCs), in
response to extracellular matrix changes induced by many
hepatotoxic agents, including ethanol and CCl4 (Kinnitel
et al. 2000; Watanabe et al. 2000; Xu et al. 2004). How-
ever, there are few studies investigating the role of MMPs
during different stages of rat chemically induced liver
carcinogenesis.
Wood and Archer (2001) showed that treatment with
batimastat, a MMP inhibitor, did not alter liver regenera-
tion or early PNL development in male Fischer 344 rats
sacrificed early after partial hepatectomy in a modified RH
model, suggesting that the increase in MMP-2 and -9
activity is not crucial for growth of normal or altered liver
parenchyma. In the present study we have observed a
significant increase in the gelatinolytic activity of all forms
of MMP-2 (72, 64 and 59 kDa) and pro-MMP-9 (92 kDa)
in the liver parenchyma with extensive development of
GST-P 7-7-positive PNL from 2-AAF-treated groups (G3
and G4), when compared to the respective control groups
(G1 and G2) at weeks 8 and 20. This initial increase of
MMP-2 and-9 activity in the liver parenchyma bearing
PNL may be associated to the ECM turnover due to the
hepatomegaly induced by 2-AAF exposure (Bitsch et al.
2000). In fact, a significant increase in the absolute and
relative liver weights and in cell proliferation rates was
observed after each cycle of 2-AAF-treatment. Therefore,
the increase in MMP-2 (72, 64 and 59 kDa) and pro-MMP-
9 (92 kDa) activity during this early stage of rat
Fig. 3 Neoplastic liver lesions
from DEN plus 2-AAF-treated
group (G3): a gomori staining
for reticular fibers, bimmunohistochemical staining
for MMP-2 and c MMP-9 at
HCC border. d gomori staining
for reticular fibers, eimmunohistochemical staining
for MMP-2 and f MMP-9 in a
hepatocholangioma. DEN,
diethylnitrosamine (200 mg/kg,
b.w., i.p.) and 2-AAF, 2-
acethylaminofluorene (0.02% in
diet)
J Mol Hist
123
hepatocarcinogenesis may be due to the global expansion
of hepatic cell compartment induced by 2-AAF-treatment
rather than the selective growth of PNL, as previously
described (Wood and Archer 2001; Pires et al. 2008).
At the end of experiment, the activity of pro, intermediate
and active forms of MMP-2 and pro-MMP-9 in samples
from non-neoplastic liver parenchyma was similar among
the different groups treated or non-treated with carcinogens.
Therefore, the differences in MMP-2 and pro-MMP-9
activity at this stage of lesion development did not directly
reflect the differences in the percentage of liver occupied by
PNL (*0.08, 2, 55 and 20% for G1, G2, G3 and G4 groups,
respectively). Thus, the differences of MMP-2 and pro-
MMP-9 activity observed may be related to differences in
cell proliferation rates observed after immediate (weeks 8
and 20) or prolonged period after 2-AAF withdrawal.
Interestingly, a high gelatinolytic activity of these MMPs
was observed in samples of hepatocholangiomas and he-
patocholangiocarcinomas from DEN plus 2-AAF-treated
(G3) group. Furthermore, MMP-2 is known to be constitu-
tively expressed in all tissues due to a different gene
promoter region (Ra and Parks 2007). Thus an increased
MMP-2 activity could be explained by the increase in cell
number within hepatocholangiocellular adenomas and car-
cinomas (Gianelli et al. 2003; Foda and Zucker 2001). Also,
the normal and PNL-bearing liver samples from all experi-
mental groups presented a remarkable MMP-9 activity
(81 kDa) but with a higher activity in both preneoplastic and
neoplastic lesions from DEN plus 2-AAF-treated group
(G3). As the expression of MMP-9 has been implicated in
the invasion and metastasis steps of the carcinogenic pro-
cess, the higher activation of MMP-9 observed in the liver
parenchyma containing PNL from 2-AAF-treated (G3 and
G4) groups or in mixed lesions from DEN plus 2-AAF-
treated (G3) group may from be probably related to the
progression of the carcinogenic cascade, which demands a
higher ECM degradation for the progression of these neo-
plastic lesions (Foda and Zucker 2001). In fact, the
malignant phenotype and invasiveness ability of H-Ras-
transformed rat liver epithelial cells and of conexin-26-
tranfected HepG2 cells is mediated by up-regulation of
MMP-9 activity (Yano and Yamasaki 2001; Lee et al. 2006).
An increase of p21ras and COX-2 expression has been
previously described during progression of rat liver carci-
nogenesis (Gulbis et al. 1993; Denda et al. 2002). Lee et al.
(2006) have shown that WB-F344 rat liver epithelial cells
transfected with v-H-Ras (H-ras WB cells) presented an
increase in MMP-9 activity, but not MMP-2, and in COX-2
expression. In H-ras WB cells, the MMP-9 and COX-2 up-
regulation occurred through the activation of ERK1/2 and
NF-kB pathways taking to invasive and migrative pheno-
types. Our findings showed that co-expression of p21ras,
COX-2 and MMP-9 depends on the morphological pattern
of the malignant lesions induced in this model.
In humans, HCC is commonly found surrounded by a
fibrous capsule, and requires the degradation of this barrier
Fig. 4 Immunohistochemical
staining for COX-2, p21ras and
MMP-9 in a cholangioma from
DEN plus 2-AAF-treated group
(G3): a HE staining, b COX-2
expression, c p21ras expression
and d MMP-9 expression in a
nodule near to a cholangioma
negative for MMP-9
J Mol Hist
123
to grow (Kaneyoshi et al. 2001). On the other hand, the
well differentiated HCC induced in the RH model adopted
here does not involve the formation of a fibrous capsule as
in human HCC. Hence, the role of MMP-2 and-9, in
association with other MMPs, at this stage of rat liver
carcinogenesis, could be important to degrade the reticular
fibers at the tumor margin. The increase in expression and
activity of MMP-2 and -9 at the lesion border indicate that
these enzymes are produced by the surrounding normal
cells, confirming that tumor–host interaction may have a
central role at the progression stage of carcinogenesis
(Caudroy et al. 2002).
In conclusion, the findings of the present study suggest
that: (1) the early increase in MMP-2 and pro-MMP-9
activity observed in the liver parenchyma from 2-AAF-
treated rats may be associated to the global proliferative
stimulus induced by this carcinogen; (2) the mixed neo-
plastic, cholangiocellular lesions and HCC have different
(G1) (G2) (G3) (G4)
- 92
- 72- 64- 59
Kda)lortnoc
%(yrte
moti sneD
(G2) (G3) (G4)
0
0.5
1.0
1.5
2.0
Pro
MMP-2
Intermediate
MMP-2
Active
MMP-2
Pro
MMP-9
MMPs
‡*
2.0
4.0
6.0
8.0
10.0
12.0
%IL
AN
CP
Group
2.0
4.0
6.0
8.0
10.0
(G1) (G2) (G4)(G3)0
‡* ‡*†*
‡* ‡* ‡*‡*
‡** ‡**
A
B
C
Fig. 5 a Representative gelatin zymography from liver samples after
8 weeks of experiment. The clear bands of 92, 72, 64 and 59 kDa
corresponds to the pro-MMP-9, pro-MMP-2, intermediate-MMP-2
and active-MMP-2, respectively. b Densitometric analysis of gelat-
olytic bands for MMP-2 and MMP-9. The values are expressed as
average of relative IOD, normalized to G1 group values. c PCNA
labeling indices (LI%). Groups: non-treated (G1), DEN-treated (G2)
DEN plus 2-AAF-treated (G3) and 2-AAF-treated (G4): DEN,
diethylnitrosamine (200 mg/kg. b.w., i.p.) and 2-AAF, 2-acethylami-
nofluorene (0.02% in diet). �,� Different from G1 group or G1 and
G2 groups, respectively. * P 0.05 \ P \ 0.09 and ** P \ 0.001
(G1) (G2) (G3) (G4
- 92
- 72- 64
- 59
Kda
2.0
4.0
6.0
8.0
2.0
4.0
6.0
8.0
% IL
AN
CP
Group(G1) (G2) (G4)(G3)
0
ActiveMMP-2
0
0.5
1.0
1.5
2.0
2.5
3.0
(G2) (G3) (G4)
ProMMP-2
IntermediateMMP-2
ProMMP- 9
)l ortnoc%(
yr temotisne
DMMPs
‡*‡*
‡*
‡*‡* ‡*
‡*‡*
‡*‡*
A
B
C
Fig. 6 a Representative gelatin zymography from liver samples after
20 weeks of experiment. The clear bands of 92, 72, 64 and 59 kDa
corresponds to the pro-MMP-9, pro-MMP-2, intermediate-MMP-2
and active-MMP-2, respectively. b Densitometric analysis of gelat-
olytic bands for MMP-2 and MMP-9. The values are expressed as
average of relative IOD, normalized to the G1 group values. c PCNA
labeling indices (LI%). Groups: non-treated (G1), DEN-treated (G2)
DEN plus 2-AAF-treated (G3) and 2-AAF-treated (G4): DEN,
diethylnitrosamine (200 mg/kg. b.w., i.p.) and 2-AAF, 2-acethylami-
nofluorene (0.02% in diet). � Different from G1 and G2 groups.
* P \ 0.001
J Mol Hist
123
patterns of immunoreactivity for MMP-2 and 9, p21ras and
COX-2 and 3) the activation of pro-MMP-9 is the better
marker associated to the progression of GST-P- positive
PNL. As RAS and COX-2 are key genes associated to PNL
progression to HCC, further studies on the role of these
pathways should be carried out in this model.
Acknowledgments This work was supported by CAPES, FAPESP
and TOXICAM. Kelly S. Furtado was recipient of a fellowship from
CAPES.
References
Asamoto M, Tsuda H, Kagawa M, de Camargo JL, Ito N, Nagase S
(1989) Strain differences in susceptibility to 2-acetylaminoflu-
orene and phenobarbital promotion of rat hepatocarcinogenesis
in a medium-term assay system: quantitation of glutathione S-
transferase P-positive foci development. Jpn J Cancer Res
80:939–944
Bannasch P, Zerban H (1992) Predictive value of hepatic preneo-
plastic lesions as indicators of carcinogenic response. In: Vainio
H, Magee PN, McGregor DB, McMichal AJ (eds) Mechanism of
carcinogenesis in risk identification, vol 116. IARC Sci Publ,
Lyon, pp 389–427
Bitsch A, Hadjiolov N, Klohn P-C, Bergmann O, Zwirner-Baier I,
Neumann HG (2000) Dose response of early effects related to
tumor promotion of 2-acetylaminofluorene. Toxicol Sci 55:44–
51. doi:10.1093/toxsci/55.1.44
Brew K, Dinakarpandian D, Nagase H (2000) Tissue inhibitors of
metalloproteinases: evolution, structure and function. Biochim
Biophys Acta 1477:267–283
Caudroy S, Polette M, Nawrocki-Raby B, Cao J, Toole BP, Zucker S
et al (2002) EMMPRIN-mediated MMP regulation in tumor and
endothelial cells. Clin Exp Metastasis 19:697–702. doi:
10.1023/A:1021350718226
Chakraborti S, Mandal M, Das S, Mandal A, Chakraborti T (2003)
Regulation of matrix metalloproteinases: an overview. Mol Cell
Biochem 253:269–285. doi:10.1023/A:1026028303196
Denda A, Kitayama W, Murata A, Kishida H, Sasaki Y, Kusuoka O,
Tsujiuchi T et al (2002) Increased expression of cyclooxygenase-
2 protein during rat hepatocarcinogenesis caused by a choline-
deficient, L-amino acid-defined diet and chemopreventive effi-
cacy of a specific inhibitor, nimesulide. Carcinogenesis 23:245–
256. doi:10.1093/carcin/23.2.245
Farber E, Sarma DS (1987) Hepatocarcinogenesis: a dynamic cellular
perspective. Lab Investig 56:4–22
Foda HD, Zucker S (2001) Matrix metalloproteinases in cancer
invasion, metastasis and angiogenesis. Drug Discov Today
6:478–482. doi:10.1016/S1359-6446(01)01752-4
Gao Y, Zhang Z, Jiang Z-S, Fang SG, Sun EW, Wang Y et al (2002)
Dynamic changes of matrix metalloproteinases in rat liver during
the development of diethylnitrosamine-induced hepatocarci-
noma. Di Yi Jun Yi Da Xue Xue Bao 22:865–868
Gianelli G, Quaranta V, Antonaci S (2003) Tissue remodelling in
liver diseases. Histol Histopathol 18:1267–1274
Gulbis B, Alexandre K, Galand P (1993) Quantitative and/or
qualitative changes in the p21-H-ras post-translational products
in regenerating liver and during hepatocarcinogenesis. Int J
Cancer 55:837–840. doi:10.1002/ijc.2910550524
Hasegawa R, Ito N (1994) Hepatocarcinogenesis in the rat. In:
Waalkes MP, Ward JM (eds) Carcinogenesis. Raven Press Ltd,
New York, pp 39–65
Imaida K, Tatematsu M, Kato T, Tsuda H, Ito N (1989) Advantages
and limitations of stereological estimation of placental glutathi-
one S-transferase-positive rat liver cell foci by computerized
three-dimensional reconstruction. Jpn J Cancer Res 80:326–330
Kaneyoshi T, Nakatsukasa H, Higashi T, Fujiwara K, Naito I, Nouso
K et al (2001) Actual invasive potential of human hepatocellular
carcinoma revealed by in situ gelatin zymography. Clin Cancer
Res 7:4027–4032
-92
-64- 59
-72
-81
(G1) (G2) (G3) (G4) Kda (T1) (T2) (T3)
- 92
- 64- 59
- 72
- 81
--
Kda
ProMMP- 2
IntermediateMMP-2
ProMMP -9
ActiveMMP-9
0
2.0
4.0
6.0
8.0
10.0
(G2) (G3) (G4) Tumor (G3)
ActiveMMP-2
0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
% IL
AN
CP
Group
(G1) (G2) (G4)(G3) (G3)Tumor
)lortnoc %(
yrtemotisne
D
‡*
‡*
‡*
MMPs
‡* ‡*‡*
‡,§*
‡** ‡**
‡,§*
A
B
C
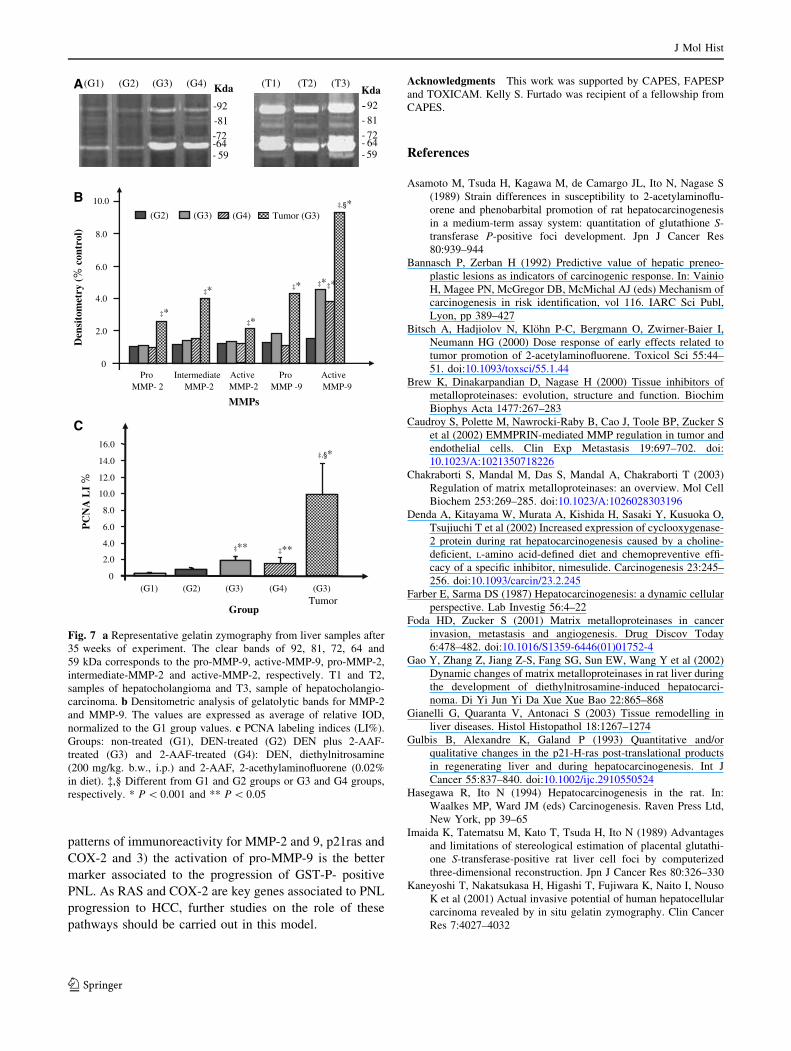
Fig. 7 a Representative gelatin zymography from liver samples after
35 weeks of experiment. The clear bands of 92, 81, 72, 64 and
59 kDa corresponds to the pro-MMP-9, active-MMP-9, pro-MMP-2,
intermediate-MMP-2 and active-MMP-2, respectively. T1 and T2,
samples of hepatocholangioma and T3, sample of hepatocholangio-
carcinoma. b Densitometric analysis of gelatolytic bands for MMP-2
and MMP-9. The values are expressed as average of relative IOD,
normalized to the G1 group values. c PCNA labeling indices (LI%).
Groups: non-treated (G1), DEN-treated (G2) DEN plus 2-AAF-
treated (G3) and 2-AAF-treated (G4): DEN, diethylnitrosamine
(200 mg/kg. b.w., i.p.) and 2-AAF, 2-acethylaminofluorene (0.02%
in diet). �,§ Different from G1 and G2 groups or G3 and G4 groups,
respectively. * P \ 0.001 and ** P \ 0.05
J Mol Hist
123

Kim T-H, Mars WM, Stolz DB, Michalopoulos GK (2000) Expres-
sion and activation of pro-MMP-2 and pro-MMP-9 during rat
liver regeneration. Hepatology 31:75–82. doi:10.1002/hep.
510310114
Kinnitel T, Mehde M, Grundmann A, Saile B, Scharf JG, Ramadori G
(2000) Expression of matrix metalloproteinases and their
inhibitors during hepatic tissue repair in the rat. Histochem Cell
Biol 113:443–453
Lee KW, Kim MS, Kang NJ, Kim DH, Surh YJ, Lee HJ et al (2006)
H-Ras selectively up-regulates MMP-9 and COX-2 through
activation of ERK1/2 and NF-kappaB: an implication for
invasive phenotype in rat liver epithelial cells. Int J Cancer
119:1767–1775. doi:10.1002/ijc.22056
Libbrecht L, Desmet V, Roskams T (2005) Preneoplastic lesions in
human hepatocarcinogenesis. Liver Int 25:16–27. doi:10.1111/
j.1478-3231.2005.01016.x
Martinez-Hernandez A, Amenta PS (1993) The hepatic extracellular
matrix I. Components and distribution in normal liver. Virchows
Arch A Pathol Anat Histopathol 423:1–11. doi:10.1007/BF01
606425
Pascale RM, Simile MM, De Miglio MR, Muroni MR, Calvisi DF,
Asara G, Casabona D et al (2002) Cell cycle deregulation in liver
lesions of rats with and without genetic predisposition to
hepatocarcinogenesis. Hepatology 35:1341–1350. doi:10.1053/
jhep.2002.33682
Pinheiro F, Faria RR, de Camargo JL, Spinardi-Barbisan ALT, da
Eira AF, Barbisan LF (2003) Chemoprevention of preneoplastic
liver foci development by dietary mushroom Agaricus blazei
Murrill in the rat. Food Chem Toxicol 41:1543–1550. doi:
10.1016/S0278-6915(03)00171-6
Pires PW, Furtado KS, Justulin LA Jr, Rodrigues MAM, Felisbino
SL, Barbisan LF (2008) Chronic ethanol intake promotes double
gluthatione S-transferase/transforming growth factor-a-positive
hepatocellular lesions in male Wistar rats. Cancer Sci 99:221–
228. doi:10.1111/j.1349-7006.2007.00677.x
Pitot HC (2001) Pathways of progression in hepatocarcinogenesis.
Lancet 358:859–860. doi:10.1016/S0140-6736(01)06038-X
Ra HJ, Parks WC (2007) Control of matrix metalloproteinase
catalytic activity. Matrix Biol 26:587–596. doi:10.1016/j.matbio.
2007.07.001
Ramaiah SK (2007) A toxicologist guide to the diagnostic interpre-
tation of hepatic biochemical parameters. Food Chem Toxicol
45:1551–1557. doi:10.1016/j.fct.2007.06.007
Sell S, Dunsford NA (1989) Evidence for the stem cell origin of
hepatocellular carcinoma and cholangiocarcinoma. Am J Pathol
134:1347–1363
Su Q, Bannasch P (2003) Relevance of hepatic preneoplasia for
human hepatocarcinogenesis. Toxicol Pathol 31:126–133. doi:
10.1080/01926230309732
Tiwawech D, Hasegawa R, Kurata Y, Tatematsu M, Shibata MA,
Thamavit W et al (1991) Dose-dependent effects of 2-acetyla-
minofluorene on hepatic foci development and cell proliferation
in rats. Carcinogenesis 12:985–990. doi:10.1093/carcin/12.6.985
Verna L, Whysner J, Williams GM (1996a) 2-acetylaminofluorene
mechanistic data and risk assessment: DNA reactivity, enhanced
cell proliferation and tumor initiation. Pharmacol Ther 71:83–
105. doi:10.1016/0163-7258(96)00063-0
Verna L, Whysner J, Williams GM (1996b) N-Nitrosodiethylamine
mechanistic data and risk assessment: bioactivation, DNA-
adduct formation, mutagenicity and tumor initiation. Pharmacol
Ther 71:57–81. doi:10.1016/0163-7258(96)00062-9
Vihinen P, Ala-Aho R, Kahari V-M (2005) Matrix metalloproteinases
as therapeutic targets in cancer. Curr Cancer Drug Targets
5:203–220. doi:10.2174/1568009053765799
Visse R, Nagase H (2003) Matrix metalloproteinases and tissue
inhibitors of metalloproteinases: structure, function and bio-
chemistry. Circ Res 92:827–839. doi:10.1161/01.RES.00000701
12.80711.3D
Watanabe T, Nijoka M, Hozawa S, Kameyama K, Hayashi T, Arai M
et al (2000) Gene expression of interstitial collagenase in both
progressive and recovery phase of rat liver fibrosis induced by
carbon tetrachloride. J Hepatol 33:224–235. doi:10.1016/S0168-
8278(00)80363-3
Wood GA, Archer MC (2001) Matrix metalloproteinases-2 and 9 do
not play a role in the grotwth of preneoplastic liver lesions in
F344 rats. Exp Biol Med (Maywood) 226:799–803
Xu GF, Pt Li, Wang XY, Jia X, Tian DL, Jiang LD et al (2004)
Dynamic changes in the expression of matrix metalloproteinases
and their inhibitors, TIMPs, during hepatic fibrosis induced by
alcohol in rats. World J Gastroenterol 10:3621–3627
Yano T, Yamasaki H (2001) Regulation of cellular invasion and
matrix metalloproteinase activity in HepG2 cell by connexin 26
transfection. Mol Carcinog 31:101–109. doi:10.1002/mc.1045
J Mol Hist
123




















