
Evaluation of Some Newer Matrix Metalloproteinases
15
25 Evaluation of Some Newer Matrix Metalloproteinases GILLIAN MURPHY, a,b VERA KNÄUPER, a SUSAN COWELL, c ROSALIND HEMBRY, a HEATHER STANTON, a GEORGINA BUTLER, a JOSÉ FREIJE, d ALBERTO M. PENDÁS, d AND CARLOS LÓPEZ-OTÍN d a School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, UK c Strangeways Research Laboratory, Cambridge CB1 4RN, UK d Departamento de Bioquimica y Biologia Molecular, Facultad de Medicina, Universidad de Oviedo, Oviedo, Spain ABSTRACT: Recombinant protein expression techniques have been utilized to facilitate the biochemical and cell biological characterization of human ma- trix metalloproteinases (MMPs). The importance of the membrane type 1 MMP (MMP 14) in the regulation of pericellular proteolysis, either directly or through the activation of MMP-2, MMP-9, and MMP-13 has been identified. Studies on an in vitro chondrocyte-like cell and an in vivo cartilage repair mod- el indicated that such MT1 MMP–regulated activation cascades are physiolog- ically feasible. MMP19 shows a limited sequence identity with other MMPs and may represent a novel subclass. However, analysis of the recombinant pro- tein identified a number of biochemical properties typical of the MMP family. Proteolysis of the extracellular matrix (ECM) is a key component of the inflamma- tory response, acting as a key effector in the modulation of the cell–cell and cell– ECM interactions underlying both disease and repair activities. The role of matrix metalloproteinases (MMPs) in matrix turnover has long been under scrutiny, and it has become evident that there is a bewildering array of these enzymes with appar- ently overlapping substrate specificities and expression patterns. A number of the MMPs that have been identified more recently have been biochemically and biolog- ically characterized in our laboratories in order to relate their function to that of the more established members of the MMP family. MEMBRANE-TYPE MATRIX METALLOPROTEINASES, MT-MMPS Four MT-MMPs have been cloned, MT1-MMP (MMP-14), MT2-MMP (MMP- 15), MT3-MMP (MMP-16), and MT4-MMP (MMP-17). 1–4 These membrane-asso- ciated members of the MMP family are expressed at low levels by many cell types. MT1-MMP is the most predominant and the most clearly regulated by cytokines and growth factors (M. Lafleur and D. Edwards, personal communication 5 ). However, b Address for correspondence: Gillian Murphy, School of Biological Sciences, University of East Anglia, Norwich, NR4 7TJ, UK. Phone, +44 1603 593811; fax, +44 1603 593874; e-mail: [email protected]
Transcript of Evaluation of Some Newer Matrix Metalloproteinases

25
Evaluation of Some Newer Matrix Metalloproteinases
GILLIAN MURPHY,
a,b
VERA KNÄUPER,
a
SUSAN COWELL,
c
ROSALIND HEMBRY,
a
HEATHER STANTON,
a
GEORGINA BUTLER,
a
JOSÉ FREIJE,
d
ALBERTO M. PENDÁS,
d
AND CARLOS LÓPEZ-OTÍN
d
a
School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, UK
c
Strangeways Research Laboratory, Cambridge CB1 4RN, UK
d
Departamento de Bioquimica y Biologia Molecular, Facultad de Medicina, Universidad de Oviedo, Oviedo, Spain
ABSTRACT: Recombinant protein expression techniques have been utilizedto facilitate the biochemical and cell biological characterization of human ma-trix metalloproteinases (MMPs). The importance of the membrane type 1MMP (MMP 14) in the regulation of pericellular proteolysis, either directly orthrough the activation of MMP-2, MMP-9, and MMP-13 has been identified.Studies on an
in vitro
chondrocyte-like cell and an
in vivo
cartilage repair mod-el indicated that such MT1 MMP–regulated activation cascades are physiolog-ically feasible. MMP19 shows a limited sequence identity with other MMPsand may represent a novel subclass. However, analysis of the recombinant pro-tein identified a number of biochemical properties typical of the MMP family.
Proteolysis of the extracellular matrix (ECM) is a key component of the inflamma-tory response, acting as a key effector in the modulation of the cell–cell and cell–ECM interactions underlying both disease and repair activities. The role of matrixmetalloproteinases (MMPs) in matrix turnover has long been under scrutiny, and ithas become evident that there is a bewildering array of these enzymes with appar-ently overlapping substrate specificities and expression patterns. A number of theMMPs that have been identified more recently have been biochemically and biolog-ically characterized in our laboratories in order to relate their function to that of themore established members of the MMP family.
MEMBRANE-TYPE MATRIX METALLOPROTEINASES, MT-MMP
S
Four MT-MMPs have been cloned, MT1-MMP (MMP-14), MT2-MMP (MMP-15), MT3-MMP (MMP-16), and MT4-MMP (MMP-17).
1–4
These membrane-asso-ciated members of the MMP family are expressed at low levels by many cell types.MT1-MMP is the most predominant and the most clearly regulated by cytokines andgrowth factors (M. Lafleur and D. Edwards, personal communication
5
). However,
b
Address for correspondence: Gillian Murphy, School of Biological Sciences, University ofEast Anglia, Norwich, NR4 7TJ, UK. Phone, +44 1603 593811; fax, +44 1603 593874; e-mail:[email protected]

26 ANNALS NEW YORK ACADEMY OF SCIENCES
both MT2-MMP and MT3-MMP share the ability to initiate the activation ofproMMP-2 with MT1-MMP. In contrast, MT4-MMP has negligible proMMP-2 pro-cessing activity (W. English, A. Merryweather, and G. Murphy, unpublished materi-al). Although there are a number of reports on the regulation of MT1-MMP mRNAby cytokines, this does not necessarily correlate with an increase in proMMP-2 acti-vation. Indeed, in our hands, the modulation of this MMP, in terms of the expressionof activity on proMMP-2, can occur at many levels other than the transcriptional one,including the extent of cell surface expression and aggregation, the extent of propep-tide processing and further proteolytic inactivation, and the levels of TIMP-2 present.The majority of these events are still not well understood and require clarification. Aspart of our studies on the potential for ECM regulation of MMP activity we lookedat the effect of fibronectin and laminin-1 on the proMMP-2 activation cascade inHT1080 cells.
6
Fibronectin, but not laminin, caused the activation of MMP-2 pro-duced by these cells. Culture of the cells on peptide fragments of fibronectin derivedfrom the cell-binding domain had a similar effect, as did culture on immobilized an-tibodies to the
α
5
and
β
1
integrin subunits. The data indicated that the activation ofMMP-2 is regulated by the nature of the ECM and that signals via the
α
5
β
1
receptormay be involved. Interestingly, another ligand for this integrin, vitronectin, did nothave the same effect as fibronectin. Levels of TIMP-2 expressed by the HT1080 cellsdid not vary between cells cultured on fibronectin or laminin, but the amount of MT1-MMP protein (but not mRNA) was increased almost two-fold. This increase waslargely attributed to an increase in levels of a truncated 45-kDa form of MT1-MMPrather than the 60-kDa active form (very little of the 63-kDa proform was detectablein any case). Parallel studies using gelatin zymography demonstrated that the upreg-ulation of the production of the 45-kDa form was concomitant with proMMP-2 acti-vation. Inhibition studies revealed that the truncation of MT1-MMP to a 45-kDa formis MMP-mediated, although not inhibited by TIMP-1. This implies that either MMP-2 or MT1-MMP could effect this processing, presenting the potential for a self-reg-ulatory termination to the activation cascade.
It is known that soluble forms of MT1-MMP and MT2-MMP are relatively effi-cient proteinases and degrade denatured interstitial collagens, cartilage aggrecan,perlecan, fibulin-1 and -2, fibronectin, vitronectin, nidogen, large tenascin-C, andlaminin-1, as well as a pro-tumor necrosis factor
α
-GST fusion protein.
7–9
In thecase of MT1-MMP, cleavage of native triple helical collagens I, II and III into thecharacteristic three-quarter, one-quarter fragments has been demonstrated, suggest-ing that this enzyme shares some characteristics with the collagenases.
8,9
C-terminaldeletion mutants of MT1-MMP are not able to cleave triple helical collagen—thatis, the mechanism of action involves the noncatalytic hemopexin domain, as de-scribed for the traditional human collagenases, MMPs 1, 8, and 13.
In order to assess whether MT1-MMP was functional at the surface of cells wetransiently transfected CHO cells, which make negligible amounts of this MMP,with an HCMV-driven expression vector expressing the cDNA.
10
Indirect immunof-luorescence showed that MT1-MMP was expressed at the surface of the cells. Trans-fected cells were cultured on thin films of Texas Red–labeled gelatin which had beenfixed to the culture vessel. Over 24 hours the cells degraded the gelatin, and lysiscould be detected by the appearance of dark patches in the labeled substrate film,specifically associated with cells expressing MT1-MMP. This focal degradation
27MURPHY
et al.
: NEWER MMP
S
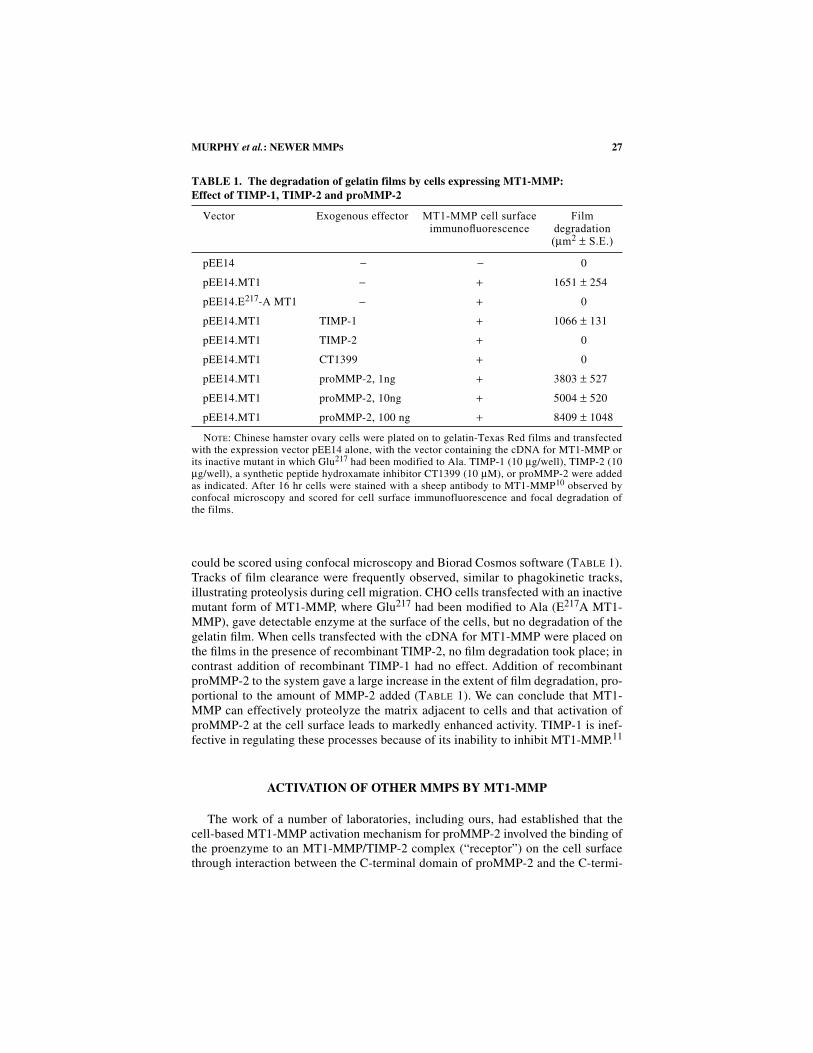
could be scored using confocal microscopy and Biorad Cosmos software (T
ABLE
1).Tracks of film clearance were frequently observed, similar to phagokinetic tracks,illustrating proteolysis during cell migration. CHO cells transfected with an inactivemutant form of MT1-MMP, where Glu
217
had been modified to Ala (E
217
A MT1-MMP), gave detectable enzyme at the surface of the cells, but no degradation of thegelatin film. When cells transfected with the cDNA for MT1-MMP were placed onthe films in the presence of recombinant TIMP-2, no film degradation took place; incontrast addition of recombinant TIMP-1 had no effect. Addition of recombinantproMMP-2 to the system gave a large increase in the extent of film degradation, pro-portional to the amount of MMP-2 added (T
ABLE
1). We can conclude that MT1-MMP can effectively proteolyze the matrix adjacent to cells and that activation ofproMMP-2 at the cell surface leads to markedly enhanced activity. TIMP-1 is inef-fective in regulating these processes because of its inability to inhibit MT1-MMP.
11
ACTIVATION OF OTHER MMPS BY MT1-MMP
The work of a number of laboratories, including ours, had established that thecell-based MT1-MMP activation mechanism for proMMP-2 involved the binding ofthe proenzyme to an MT1-MMP/TIMP-2 complex (“receptor”) on the cell surfacethrough interaction between the C-terminal domain of proMMP-2 and the C-termi-
TABLE 1. The degradation of gelatin films by cells expressing MT1-MMP:Effect of TIMP-1, TIMP-2 and proMMP-2
N
OTE
: Chinese hamster ovary cells were plated on to gelatin-Texas Red films and transfectedwith the expression vector pEE14 alone, with the vector containing the cDNA for MT1-MMP orits inactive mutant in which Glu
217
had been modified to Ala. TIMP-1 (10
µ
g/well), TIMP-2 (10
µ
g/well), a synthetic peptide hydroxamate inhibitor CT1399 (10
µ
M), or proMMP-2 were addedas indicated. After 16 hr cells were stained with a sheep antibody to MT1-MMP
10
observed byconfocal microscopy and scored for cell surface immunofluorescence and focal degradation ofthe films.
Vector Exogenous effector
MT1-MMP cell surface immunofluorescence
Film degradation (
µ
m
2
±
S.E.)
pEE14
− −
0
pEE14.MT1
− +
1651
±
254
pEE14.E
217
-A MT1
− +
0
pEE14.MT1 TIMP-1
+
1066
±
131
pEE14.MT1 TIMP-2
+
0
pEE14.MT1 CT1399
+
0
pEE14.MT1 proMMP-2, 1ng
+
3803
±
527
pEE14.MT1 proMMP-2, 10ng
+
5004
±
520
pEE14.MT1 proMMP-2, 100 ng
+
8409
±
1048

28 ANNALS NEW YORK ACADEMY OF SCIENCES
nal domain of TIMP-2.
5,7,11,12
Processing of proMMP-2 to a Leu
38
intermediateform may then be initiated by an adjacent free and active MT1-MMP molecule. Thisinitial cleavage event destabilizes the structure of the proMMP-2 propeptide domain,and autoproteolysis then proceeds in an MMP-2 concentration-dependent manner,which releases the rest of the propeptide domain and fully active MMP-2. In cell cul-ture studies, the enzyme concentration in solution is low, and deletion of either theproMMP-2 C-terminal domain or the transmembrane domain of MT1-MMP abol-ishes proMMP-2 activation, emphasizing that the binding mechanism involving theMT1-MMP/TIMP-2 complex on the cell surface acts as a concentration mechanism,which is crucial for the efficiency of activation. It appears that addition of small subequimolar amounts of TIMP-2 to cells expressing MT1-MMP can enhanceproMMP-2 activation, since this increases the concentration of the MT1-MMP/TIMP-2 receptor for proMMP-2 on the cell surface. However, at high TIMP-2 con-centrations all the MT1-MMP molecules on the cell surface are complexed withTIMP-2 and, although proMMP-2 binding occurs, no active MT1-MMP remains toinitiate processing. This concept has been substantiated by cell free kinetic studiesof the effects of the MT1-MMP/TIMP-2 “receptor” as a mode of concentratingproMMP-2 in order to promote autoproteolysis.
13
This suggests that proMMP-2 ac-tivation is regulated by the amount of TIMP-2 secreted by MT1-MMP expressingcells as well as by the extent of MT1-MMP activation. C-terminal domain mutantsof proMMP-2, TIMP-2, and MT1-MMP were used to further support the abovemechanism. Soluble constructs of full-length and catalytic domain MT1-MMP wereused to demonstrate that binding with TIMP-2 occurs primarily through N-terminaldomain interactions of both enzyme and inhibitor, leaving the C-terminal domain ofTIMP-2 free for interactions with proMMP-2. The rate of autolytic activation ofproMMP-2 initiated by MT1-MMP cleavage could be potentiated by concentrationof the proenzyme, for example, by binding to heparin.
13
Residues 568-631 of theproMMP-2 C-terminal domain are important in the formation of the heparin bindingsite, since replacement of this region with the corresponding stromelysin-1 sequenceabolished binding to heparin and the potentiation of activation. The same region ofMMP-2 was required for the binding of both its pro and active form to TIMP-2,whereas residues 418-474 were not required. A similar pattern was seen using cellmembrane–associated MT1-MMP: residues 568-631 were necessary for both thebinding and activation of MMP-2, whereas residues 418-474 were not. Neither re-gion was required for activation in solution. The addition of TIMP-2 to HT1080membrane preparations overexpressing MT1-MMP depleted of endogenous TIMP-2resulted in potentiation of proMMP-2 activation. This effect was dependent uponTIMP-2 binding to MT1-MMP, rather than to an independent membrane site. To-gether the data supported the hypothesis that TIMP-2 forms a receptor by complexformation with MT1-MMP and that this regulates the efficient generation of func-tionally active MMP-2.
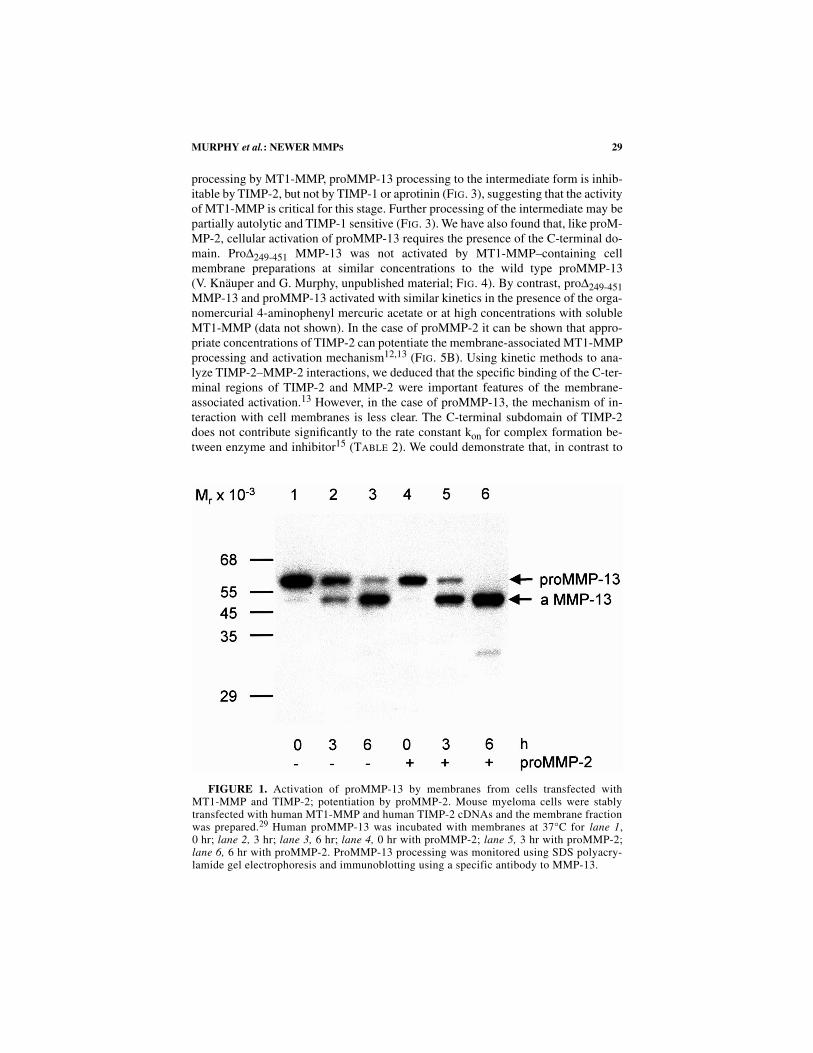
Besides the activation of proMMP-2 it has been shown that MT1-MMP may beresponsible for the activation of proMMP-13, either directly or via MMP-2activation
14
(F
IG
. 1). In both cases, processing of the prodomain of proMMP-13 oc-curs via a 56-kDa intermediate, yielding a final 48 kDa form. Similarly, the activationof proMMP-13 by MT1-MMP can potentiate the membrane processing of the cata-lytically inactive mutant of MMP-2, proE
375
-A MMP-2 (F
IG
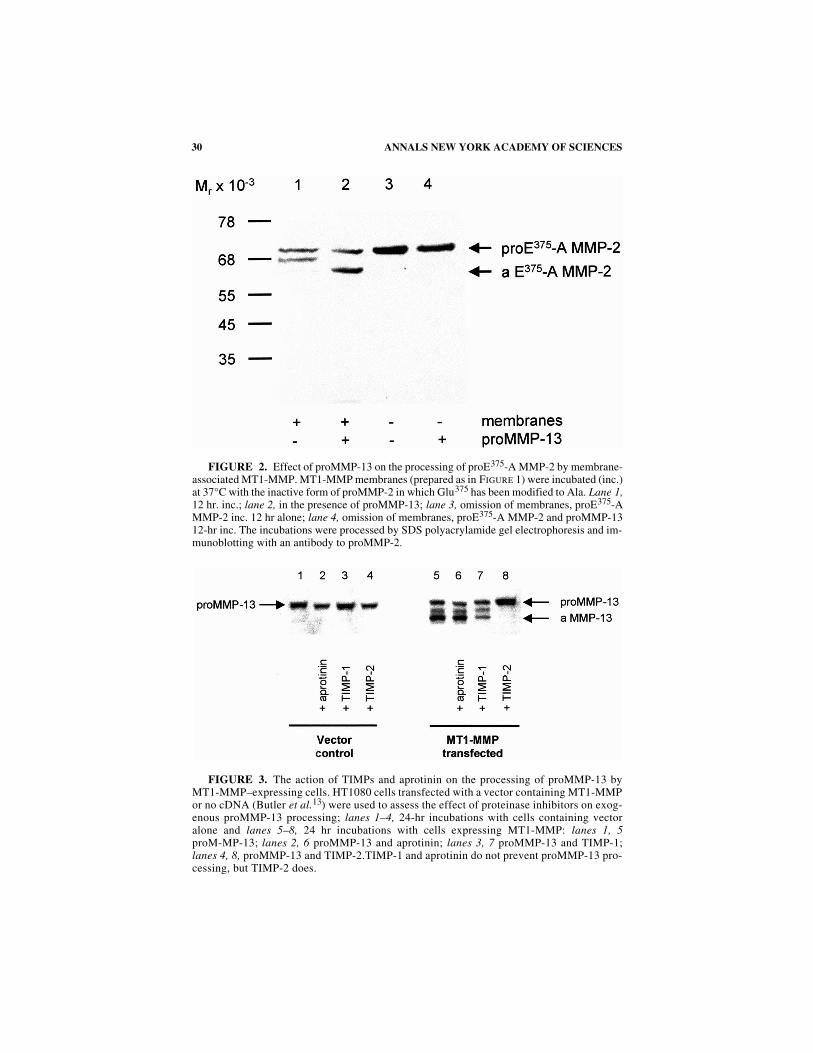
. 2). As for proMMP-2
29MURPHY
et al.
: NEWER MMP
S
processing by MT1-MMP, proMMP-13 processing to the intermediate form is inhib-itable by TIMP-2, but not by TIMP-1 or aprotinin (F
IG
. 3), suggesting that the activityof MT1-MMP is critical for this stage. Further processing of the intermediate may bepartially autolytic and TIMP-1 sensitive (F
IG
. 3). We have also found that, like proM-MP-2, cellular activation of proMMP-13 requires the presence of the C-terminal do-main. Pro
∆
249-451
MMP-13 was not activated by MT1-MMP–containing cellmembrane preparations at similar concentrations to the wild type proMMP-13(V. Knäuper and G. Murphy, unpublished material; F
IG
. 4). By contrast, pro
∆
249-451
MMP-13 and proMMP-13 activated with similar kinetics in the presence of the orga-nomercurial 4-aminophenyl mercuric acetate or at high concentrations with solubleMT1-MMP (data not shown). In the case of proMMP-2 it can be shown that appro-priate concentrations of TIMP-2 can potentiate the membrane-associated MT1-MMPprocessing and activation mechanism
12,13
(F
IG
. 5B). Using kinetic methods to ana-lyze TIMP-2–MMP-2 interactions, we deduced that the specific binding of the C-ter-minal regions of TIMP-2 and MMP-2 were important features of the membrane-associated activation.
13
However, in the case of proMMP-13, the mechanism of in-teraction with cell membranes is less clear. The C-terminal subdomain of TIMP-2does not contribute significantly to the rate constant k
on
for complex formation be-tween enzyme and inhibitor
15
(T
ABLE
2). We could demonstrate that, in contrast to
FIGURE 1. Activation of proMMP-13 by membranes from cells transfected withMT1-MMP and TIMP-2; potentiation by proMMP-2. Mouse myeloma cells were stablytransfected with human MT1-MMP and human TIMP-2 cDNAs and the membrane fractionwas prepared.29 Human proMMP-13 was incubated with membranes at 37°C for lane 1,0 hr; lane 2, 3 hr; lane 3, 6 hr; lane 4, 0 hr with proMMP-2; lane 5, 3 hr with proMMP-2;lane 6, 6 hr with proMMP-2. ProMMP-13 processing was monitored using SDS polyacry-lamide gel electrophoresis and immunoblotting using a specific antibody to MMP-13.
30 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 2. Effect of proMMP-13 on the processing of proE375-A MMP-2 by membrane-associated MT1-MMP. MT1-MMP membranes (prepared as in FIGURE 1) were incubated (inc.)at 37°C with the inactive form of proMMP-2 in which Glu375 has been modified to Ala. Lane 1,12 hr. inc.; lane 2, in the presence of proMMP-13; lane 3, omission of membranes, proE375-AMMP-2 inc. 12 hr alone; lane 4, omission of membranes, proE375-A MMP-2 and proMMP-1312-hr inc. The incubations were processed by SDS polyacrylamide gel electrophoresis and im-munoblotting with an antibody to proMMP-2.
FIGURE 3. The action of TIMPs and aprotinin on the processing of proMMP-13 byMT1-MMP–expressing cells. HT1080 cells transfected with a vector containing MT1-MMPor no cDNA (Butler et al.13) were used to assess the effect of proteinase inhibitors on exog-enous proMMP-13 processing; lanes 1–4, 24-hr incubations with cells containing vectoralone and lanes 5–8, 24 hr incubations with cells expressing MT1-MMP: lanes 1, 5proM-MP-13; lanes 2, 6 proMMP-13 and aprotinin; lanes 3, 7 proMMP-13 and TIMP-1;lanes 4, 8, proMMP-13 and TIMP-2.TIMP-1 and aprotinin do not prevent proMMP-13 pro-cessing, but TIMP-2 does.

31MURPHY
et al.
: NEWER MMP
S
proMMP-2, the MT1-MMP membrane activation of proMMP-13 is not potentiatedby carefully titrated concentrations of TIMP-2 (F
IG
. 5A). The question arose as towhether proMMP-13 and proMMP-2 compete for the same pool of membrane-boundMT1-MMP. The inactive proE
375
-A MMP-2 acts as an inhibitor of proMMP-2 pro-cessing by membranes unless low concentrations of TIMP-2 are present (F
IG
. 6A).This may be due to the formation of MT1-MMP/TIMP-2 complexes, which can bindthe excess proMMP-2. In earlier studies we showed that the isolated C-terminal do-main of MMP-2 inhibits cell membrane binding and activation of proMMP-2.
16
Fur-thermore, other work showed that administration of proE
375
-A MMP-2 with C127cells overexpressing proMMP-2 injected into the tail veins of nude mice preventedlung nodule formation of these cells.
17
In the same experiment described in F
IGURE
6proMMP-13 activation is potentiated by the addition of excess proE
375
-A MMP-2(F
IG
. 6C). In the presence of excess TIMP-2 and proE
375
-A MMP-2, the activationof proMMP-13 is potentiated, leading to partial processing of the proE
375
-A MMP-2(F
IG
. 6D). In contrast, during proMMP-2 activation under the same conditions,proE
375
-A MMP-2 is not processed (F
IG
. 6B). This suggests that specific interactionsbetween full-length proMMP-2 and MMP-13, promoted by the presence of TIMP-2,may be occurring. In the chondrosarcoma cell line SW1353, exposure to interleukin-1
β
(IL-1
β
) and oncostatin M cause an upregulation in the production of MMP-13 andof MMP-9. ProMMP-2 is made by the cells, but is not modulated by these cytokines.Treatment with concanavalin A (conA) upregulates MT1-MMP and proMMP-2, 9and 13 are activated.
18 Since both MMP-2 and MMP-13 can activate proMMP-9 invitro, it was necessary to identify their specific roles in this model system. Cells werestimulated with phorbol myristate acetate, PMA, which gives a large upregulation ofproMMP-9 production, but not MMP-13. In the presence of conA, activation ofMMP-2 occurred, but no active MMP-9 was detected. It could be concluded that
FIGURE 4. Comparison of the processing of proMMP-13 and pro∆249-451MMP-13 bymembrane-associated MT1-MMP. Recombinant proMMP-13 was incubated at 37°C for16 hr: lane 1, alone; lane 2, with an MT1-MMP–containing membrane preparation; lane 3,recombinant pro∆249-451 MMP-13 (deletion of the complete C-terminal domain15) incubat-ed alone; lane 4, with MT1-MMP membranes. The incubations were resolved by SDS poly-acrylamide gel electrophoresis and immunoblotting using an antibody to MMP-13.

32 ANNALS NEW YORK ACADEMY OF SCIENCES
MMP-13 is necessary for proMMP-9 activation in this system. To assess their effecton the activation cascade recombinant TIMPs were added to SW1353 cells treatedwith IL1β, oncostatin M, and ConA. Both TIMP-2 and TIMP-3 could inhibit the ac-tivation of proMMP-2, 9 and 13. TIMP-1 inhibited the activation of proMMP-9, butdid not prevent proMMP-2 or MMP-13 activation. The latter two were processed tointermediate products by cells treated with TIMP-1, implying that MT1-MMP wasactive but that further steps in the proteolytic cascade were not taking place (FIG. 7).
FIGURE 5. Activation of proMMP-13 and proMMP-2 by cell membranes containingMT1-MMP: effect of TIMP-2. (A) Lane 1, proMMP-13 incubated for 12 hr at 37°C alone;lane 2, proMMP-13 incubated in the presence of MT1-MMP-containing membranes; lane3, as lane 2 in the presence of 0.65 nM TIMP-2; lane 4, as lane 2 in the presence of 3.23 nMTIMP-2; lane 5, as lane 2 with 6.46 nM TIMP-2; lane 6, as lane 2 with 12.9 nM TIMP-2;lane 7, as lane 2 with 25.8 nM TIMP-2. Incubations were resolved by SDS-PAGE and im-munoblotting with an antibody to MMP-13. (B) As FIGURE 5A, but incubating proMMP-2with MT1-MMP membranes and increasing concentrations of TIMP-2. Incubations were re-solved by SDS-PAGE gelatin gel zymography. (C) As FIGURE 5A, but incubating proMMP-2 and proMMP-13 with MT1-MMP membranes and increasing concentrations of TIMP-2.Incubations were resolved to detect MMP-13 by immunoblotting as for FIGURE 5A.
TIMP-2 clearly potentiates the activity of membrane-bound MT1-MMP on proMMP-2at low doses and is inhibitory at higher doses, whereas it is inhibitory for proMMP-13 pro-cessing at all concentrations tested. However, in the presence of proMMP-2 higher concen-trations of TIMP-2 are needed to inhibit processing of MMP-13.
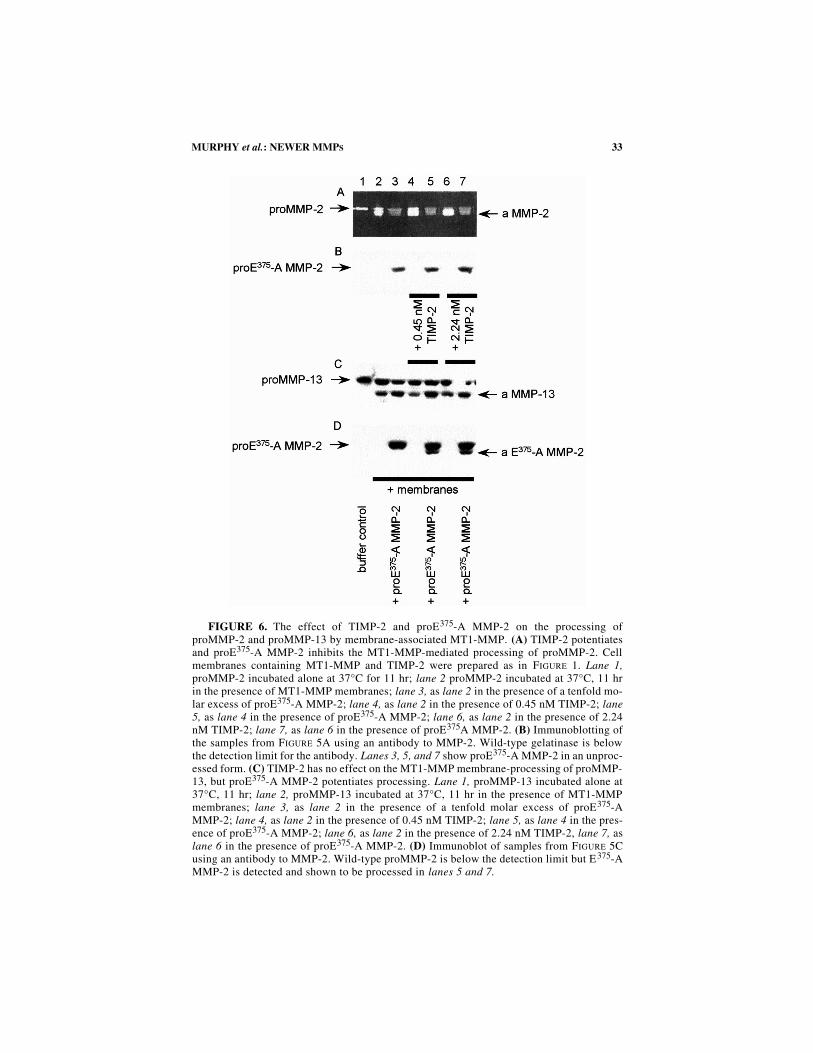
33MURPHY et al.: NEWER MMPS
FIGURE 6. The effect of TIMP-2 and proE375-A MMP-2 on the processing ofproMMP-2 and proMMP-13 by membrane-associated MT1-MMP. (A) TIMP-2 potentiatesand proE375-A MMP-2 inhibits the MT1-MMP-mediated processing of proMMP-2. Cellmembranes containing MT1-MMP and TIMP-2 were prepared as in FIGURE 1. Lane 1,proMMP-2 incubated alone at 37°C for 11 hr; lane 2 proMMP-2 incubated at 37°C, 11 hrin the presence of MT1-MMP membranes; lane 3, as lane 2 in the presence of a tenfold mo-lar excess of proE375-A MMP-2; lane 4, as lane 2 in the presence of 0.45 nM TIMP-2; lane5, as lane 4 in the presence of proE375-A MMP-2; lane 6, as lane 2 in the presence of 2.24nM TIMP-2; lane 7, as lane 6 in the presence of proE375A MMP-2. (B) Immunoblotting ofthe samples from FIGURE 5A using an antibody to MMP-2. Wild-type gelatinase is belowthe detection limit for the antibody. Lanes 3, 5, and 7 show proE375-A MMP-2 in an unproc-essed form. (C) TIMP-2 has no effect on the MT1-MMP membrane-processing of proMMP-13, but proE375-A MMP-2 potentiates processing. Lane 1, proMMP-13 incubated alone at37°C, 11 hr; lane 2, proMMP-13 incubated at 37°C, 11 hr in the presence of MT1-MMPmembranes; lane 3, as lane 2 in the presence of a tenfold molar excess of proE375-AMMP-2; lane 4, as lane 2 in the presence of 0.45 nM TIMP-2; lane 5, as lane 4 in the pres-ence of proE375-A MMP-2; lane 6, as lane 2 in the presence of 2.24 nM TIMP-2, lane 7, aslane 6 in the presence of proE375-A MMP-2. (D) Immunoblot of samples from FIGURE 5Cusing an antibody to MMP-2. Wild-type proMMP-2 is below the detection limit but E375-AMMP-2 is detected and shown to be processed in lanes 5 and 7.

34 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 7. Effect of TIMPs and thrombin on the activation of proMMP-13, MMP-2and MMP-9 by SW1353 cells. SW1353 cells were treated with IL1-β (10 ng/ml), oncostatinM (50 ng/ml), and conA (50 µg/ml in DMEM/Hams F-12, 0.2% lactalbumin hydrolysate for3 days to induce synthesis of proMMP-13 (lanes 1–3). TIMP-2 (3.3 µg/ml, lane 2), TIMP-1(0.42 µg/ml, lane 3), were added with the IL-1β and oncostatin M. ConA was replaced withthrombin (lane 4). (A) MMP-9 and MMP-2 were assessed by gelatin gel zymography (in-verted image). (B) MMP-13 was assessed by SDS polyacrylamide gel electrophoresis andimmunoblotting with an antibody to MMP-13.
In the presence of ConA activation of MMP-2, MMP-9 and MMP-13 was observed andTIMP-2 prevented all three processes. TIMP-1 prevented proMMP-9 activation, but notproMMP-2, nor MMP-13. Thrombin caused a modest processing of proMMP-2, but notMMP-9 or MMP-13.
TABLE 2. TIMP-2 shows weak interactions with the C-domain of human MMP-13
NOTE: Rate constants for the inhibition of active full-length MMP-13 and ∆249-451 MMP-13by full–length and truncated forms of TIMP-2 were determined using a quenched fluorescentpeptide substrate assay at 25°C.15
Inhibitor kon (× 10–6 M–1. s–1)
MMP-13 ∆249-451 MMP-13
TIMP-2 1.41 0.81
∆187-194 TIMP-2 1.80 0.69
∆128-194 TIMP-2 0.29 0.32

35MURPHY et al.: NEWER MMPS
This activation cascade may also operate in in vivo situations. For example, wehave been using a method for inducing cartilage repair in lesions that do not pene-trate the underlying bone19 in order to investigate the regulatory signals and under-lying mechanisms involved in cartilage repair. The method uses a staged delivery ofgrowth factors to attract cells into partial thickness defects, and the space to be re-paired is defined by filling the defect with a biodegradable matrix such as gelatin.Following formation of partial thickness defects in vivo in pigs, we monitored therepair process by immunofluorescence microscopy to determine which MMPs andTIMPs are expressed during the different stages of repair. At two days after wound-ing, macrophages identified using well-characterized cell marker antibodies wereobserved attached to the intact cartilage surface and to the cut cartilage matrix in thedefects. These macrophages were seen to synthesize and secrete MMP-9 focallyonto damaged but not onto intact cartilage surfaces. By day 8 defects were populatedwith increased numbers of predominantly macrophages containing MMP-9, andmacrophages synthesizing MMP-9 were seen to have penetrated the necrotic zonebordering the defect, and the cartilage matrix stained throughout the necrotic area.Also by day 8 chondrocytes adjacent to and below the necrotic areas had undergoneclonal expansion, and these clonal chondrocytes showed synthesis of MMP-13 in ad-dition to increased MMP-1 and MMP-3 (R. Hembry, J. Tyler and G. Murphy, unpub-lished material). MMP-13 is normally restricted to hypertrophic chondrocytes20,21
and has been shown to activate MMP-922: the presence of both these enzymes on theedges of the defects may well compromise mechanical function.
MATRIX METALLOPROTEINASE-19
One of the most recently cloned members of the MMP family is MMP-19.23,24
Its primary sequence suggested that it is a typical secreted protein with identifiablepropeptide, catalytic and hemopexin-like C-terminal domains, as found in other fam-ily members. Detailed sequence comparisons with other MMPs suggest that MMP-19 is not closely related to the collagenases or stromelysins and is the first memberof a new sub-class, a hypothesis that is upheld by its chromosomal location at q14on the long arm of chromosome 12.24 Three unusual sequence features are an oligo-glutamic acid containing “hinge” connecting the catalytic and C-domains, an appar-ently free cysteine in the catalytic domain, and an extended C-terminal “tail.” Thecloned protein was expressed as inclusion bodies in E. coli and solubilized, purified,and refolded. The proenzyme had a mol. wt. of 58 kDa and could be activated withtrypsin to a form that cleaved the general MMP peptide substrate Mca-Pro-Leu-Gly-Leu-Dpa-Ala-Arg-NH2 with a kcat/kM of 1.93 × 103 M–1⋅s–1. The proenzyme has apropensity to self-activate, which can be attributed to the fact that the propeptide“cysteine switch” motif is PRCGLED, as compared with PRCGXPD in most MMPs.Mutational studies by Park et al.25 have shown that modification of this Pro-residuein MMP-3 generates autoactivating forms. The N-terminal sequence of autoactivatedMMP-19 was found to be 99YLLLGRW. MMP-19 preferentially hydrolyzed aquenched fluorescent peptide “stromelysin substrate” Mca-Pro-Leu-Ala-Nva-Ala-Arg-NH2 over a collagenase-favored peptide sequence.24 MMP-19 shows little pro-pensity for the cleavage of extracellular matrix components, although activity

36 ANNALS NEW YORK ACADEMY OF SCIENCES
against large tenascin C and gelatin (preferential cleavage of the α2(I)-chain) hasbeen demonstrated. The activity versus gelatin is so limited that enzymatic activitycannot be demonstrated by gelatin zymography. Type I collagen and laminin are notcleaved (not shown). MMP-19 is able to degrade aggrecan, but cleaves it at a novelsite as well as that of other MMPs (J. Stracke, A. Merryweather, and G. Murphy, un-published material).26 The catalytic domain of the enzyme shows slow associationrate constants for binding TIMP-1 (kon = 1.22 × 104 M–1s–1) and TIMP-3 (kon = 3.73× 104 M–1s–1). TIMP-2 binding is considerably faster, but accurate values have notyet been determined (J. Stracke, V. Knäuper and G. Murphy, unpublished material).
MMP-19 is widely distributed at the mRNA level, transcripts being significantlydetectable by Northern analysis of adult human placenta, lung, pancreas, ovary,spleen, and intestine. This is unusual in that MMPs are not abundantly produced byadult cells in normal tissues. Preliminary studies of cell lines suggest that MMP-19
FIGURE 8. Cell surface–associated activation cascades for matrix metalloproteinases.The extracellular activation of most proMMPs is probably limited to the pericellular envi-ronment where cell-associated proteinase activities can function in an inhibitor-depleted en-vironment. Key initiators of the MMP activation cascades are thought to be: MT1-MMP(and perhaps MT2-MMP and MT3-MMP), which is activated intracellularly, and urokinase-like plasminogen activator, which is bound to its cell surface receptor. The production ofactive or partially active MMPs allows a cascade of autolytic and heterolytic cleavages tooccur, generating fully active enzymes. The efficiency of these interactions is dependent onmechanisms for the concentration of MMPs at the cell surface or by binding to matrix pro-teins.

37MURPHY et al.: NEWER MMPS
may be constitutively expressed, although upregulation by cytokines can occur (un-published data). It appears to be associated with the surface of cells, an observationfirst made by Sedlacek et al.,28 who described its localization on the surface of phy-tohaemagglutinin-stimulated peripheral blood leukocytes. (Interestingly, the North-ern analysis of tissues by Pendas et al.24 did not detect MMP-19 mRNA in leukocyteextracts.). However, MMP-19 does not have an identifiable transmembrane motif, al-though it has an extended C-terminal “tail” beyond the Cys472 residue, involved informing the typical MMP-β-propeller disulfide bond:
RPRTIDTTPSGNTTPSGTGITLDTTLSATETTFEY.
This sequence contains a putative N-glycosylation site and multiple Thr residues,which may be involved in cell surface interactions. Alternatively, the acidic hingeregion between the catalytic and C-domains may confer binding properties, for ex-ample, to cell surface proteoglycans. MMP-19 has also been identified as the autoan-tigen RASI-1, found in the synovial membranes of a patient with rheumatoidarthritis and a circulatory autologous antibody.27,28 By means of immunohistochem-ical techniques27 the protein appears to be localized to the blood vessels of the syn-ovium and of other normal tissues probably associated with smooth muscle cells.This enzyme is clearly an exciting addition to the MMP repertoire, meriting furtherwork on its biochemistry and cell biology as well as an in vivo approach to the anal-ysis of its function.
ACKNOWLEDGMENTS
We thank all the colleagues who have contributed to this work and Jill Gorton forhelp preparing the manuscript. Our work is funded by the Arthritis and RheumatismCampaign, the Wellcome Trust, the Nuffield Foundation and the Medical ResearchCouncil, U.K., and the European Union Biomed II Program.
REFERENCES
1. SATO, H., T. TAKINO, Y. OKADA, J. CAO, A. SHINAGAWA, E. YAMAMOTO & M. SEIKI.1994. A matrix metalloproteinase expressed on the surface of invasive tumour cells.Nature 370: 61–65.
2. TAKINO, T., H. SATO, A. SHINAGAWA & M. SEIKI. 1995. Identification of the secondmembrane-type matrix metalloproteinase (MT-MMP-2) gene from a human placentacDNA library—MT-MMPs form a unique membrane-type subclass in the MMP fam-ily. J. Biol. Chem. 270: 23013–23020.
3. WILL, H. & B. HINZMANN. 1995. cDNA sequence and mRNA tissue distribution of anovel human matrix metalloproteinase with a potential transmembrane segment. Eur.J. Biochem. 231: 602–608.
4. PUENTE, X.S., A.M. PENDAS, E. LLANO, G. VELASCO & C. LOPEZ-OTIN. 1996. Molec-ular cloning of a novel membrane-type matrix metalloproteinase from a humanbreast carcinoma. Cancer Res. 56: 944–949.
5. LOHI, J., K. LEHTI, J. WESTERMARCK, V.M. KÄHÄRI & J. KESKI-OJA. 1996. Regulationof membrane-type matrix metalloproteinase-1 expression by growth factors andphorbol 12-myristate 13-acetate. Eur. J. Biochem. 239: 239–247.

38 ANNALS NEW YORK ACADEMY OF SCIENCES
6. STANTON, H., J. GAVRILOVIC, J. ATKINSON, M.-P. D’ORTHO, K.M. YAMADA, L. ZARDI
& G. MURPHY. 1998. The activation of proMMP-2 (gelatinase A) by HT1080 fibro-sarcoma cells is promoted by culture on a fibronectin substrate and is concomitantwith an increase in processing of MT1-MMP (MMP-14) to a 45 kDa form. J. CellSci. 111: 2789–2798.
7. PEI, D.Q. & S.J. WEISS. 1996. Transmembrane-deletion mutants of the membrane-type matrix metalloproteinase-1 process progelatinase A and express intrinsicmatrix-degrading activity. J. Biol. Chem. 271: 9135–9140.
8. OHUCHI, E., K. IMAI, Y. FUJII, H. SATO, M. SEIKI & Y. OKADA. 1997. Membrane type1 matrix metalloproteinase digests interstitial collagens and other extracellularmatrix macromolecules. J. Biol. Chem. 272: 2446–2451.
9. D’ORTHO, M.P., H. WILL, S. ATKINSON, G. BUTLER, A. MESSENT, J. GAVRILOVIC,B. SMITH, R. TIMPL, L. ZARDI & G. MURPHY. 1997. Membrane-type matrix metallo-proteinases 1 and 2 exhibit broad-spectrum proteolytic capacities comparable tomany matrix metalloproteinases. Eur. J. Biochem. 250: 751–757.
10. D’ORTHO, M.P., H. STANTON, M. BUTLER, S.J. ATKINSON, G. MURPHY &R.M. HEMBRY. 1998. MT1-MMP on the cell surface causes focal degradation of gel-atin films. FEBS Lett. 421: 159–164.
11. WILL, H., S.J. ATKINSON, G.S. BUTLER, B. SMITH & G. MURPHY. 1996. The solublecatalytic domain of membrane type 1 matrix metalloproteinase cleaves the propep-tide of progelatinase A and initiates autoproteolytic activation—Regulation byTIMP-2 and TIMP-3. J. Biol. Chem. 271: 17119–17123.
12. STRONGIN, A.Y., I. COLLIER, G. BANNIKOV, B.L. MARMER, G.A. GRANT &G.I. GOLDBERG. 1995. Mechanism of cell surface activation of 72-kDa type IV colla-genase. Isolation of the activated form of the membrane metalloprotease. J. Biol.Chem. 270: 5331–5338.
13. BUTLER, G.S., M.J. BUTLER, S.J. ATKINSON, H. WILL, T. TAMURA, S.S. VAN
WESTRUM, T. CRABBE, J. CLEMENTS, M.P. D’ORTHO & G. MURPHY. 1998. TheTIMP2 membrane type 1 metalloproteinase “receptor” regulates the concentrationand efficient activation of progelatinase A—A kinetic study. J. Biol. Chem. 273:871–880.
14. KNÄUPER, V., H. WILL, C. LOPEZ-OTIN, B. SMITH, S.J. ATKINSON, H. STANTON,R.M. HEMBRY & G. MURPHY G. 1996. Cellular mechanisms for human procollage-nase-3 (MMP-13) activation: evidence that MT1-MMP (MMP-14) and gelatinase A(MMP-2) are able to generate active enzyme. J. Biol. Chem. 271: 17124–17131.
15. KNÄUPER, V., S. COWELL, B. SMITH, C. LOPEZ-OTIN, M. O’SHEA, H. MORRIS,L. ZARDI & G. MURPHY. 1997. The role of the C-terminal domain of human collage-nase-3 (MMP-13) in the activation of procollagenase-3, substrate specificity, and tis-sue inhibitor of metalloproteinase interaction. J. Biol. Chem. 272: 7608–7616.
16. WARD, R.V., S.J. ATKINSON, P.M. SLOCOMBE, A.J.P. DOCHERTY, J.J. REYNOLDS &G. MURPHY. 1991. Tissue inhibitor of metalloproteinases-2 inhibits the activation of72 kDa progelatinase by fibroblast membranes. Biochim. Biophys. Acta 1079: 242–246.

39MURPHY et al.: NEWER MMPS
17. COCKETT, M.I., G. MURPHY, M.I. BIRCH, J.P. O’CONNELL, T. CRABBE,A.T. MILLICAN, I.R. HART & A.J.P. DOCHERTY. 1997. Matrix metalloproteinasesand metastatic cancer. Biochem. Soc. Symp. 63: 295–313.
18. COWELL, S., V. KNÄUPER, M.L. STEWART, M.P. D’ORTHO, H. STANTON,R.M. HEMBRY, C. LOPEZ-OTIN, J.J. REYNOLDS & G. MURPHY. 1998. Induction ofmatrix metalloproteinase activation cascades based on membrane-type 1 matrix met-alloproteinase: associated activation of gelatinase A, gelatinase B and collagenase 3.Biochem. J. 331: 453–458.
19. HUNZIKER, E.B. & L.C. ROSENBERG. 1996. Repair of partial-thickness defects in artic-ular cartilage: Cell recruitment from the synovial membrane. J. Bone Joint Surg.78A: 721–733.
20. JOHANSSON, N., U. SAARIALHO-KERE, K. AIROLA, R. HERVA, L. NISSINEN,J. WESTERMARCK, E. VUORIO, J. HEINO & V.M. KÄHÄRI. 1997. Collagenase-3(MMP-13) is expressed by hypertrophic chondrocytes, periosteal cells, and osteo-blasts during human fetal bone development. Dev. Dyn. 208: 387–397.
21. STÅHLE-BÄCKDAHI, M., B. SANDSTEDT, K. BRUCE, A. LINDAHL, M.G. JIMENEZ,J.A. VEGA & C. LOPEZ-OTIN. 1997. Collagenase-3 (MMP-13) is expressed duringhuman fetal ossification and re-expressed in postnatal bone remodeling and in rheu-matoid arthritis. Lab. Invest. 76: 717–728.
22. KNÄUPER, V., B. SMITH, C. LOPEZ-OTIN & G. MURPHY. 1997. Activation of progelati-nase B (proMMP-9) by active collagenase-3 (MMP-13). Eur. J. Biochem. 248: 369–373.
23. COSSINS, J., T.J. DUDGEON, G. CATLIN, A.J.H. GEARING & J.M. CLEMENTS. 1996.Identification of MMP-18, a putative novel human matrix metalloproteinase. Bio-chem. Biophys. Res. Commun. 228: 494–498.
24. PENDAS, A.M., V. KNÄUPER, X.S. PUENTE, E. LLANO, M.G. MATTEI, S. APTE,G. MURPHY & C. LOPEZ-OTIN. 1997. Identification and characterization of a novelhuman matrix metalloproteinase with unique structural characteristics, chromosomallocation, and tissue distribution. J. Biol. Chem. 272: 4281–4286.
25. PARK, A.J., L.M. MATRISIAN, A.F. KELLS, R. PEARSON, Z. YUAN & M. NAVRE. 1991.Mutational analysis of the transin (rat stromelysin) autoinhibitor region demon-strates a role for residues surrounding the “cysteine switch”. J. Biol. Chem. 266:1584–1590.
26. HUGHES, C.E., B. CATERSON, A.J. FOSANG, P.J. ROUGHLEY & J.S. MORT. 1995. Mon-oclonal antibodies that specifically recognize neoepitope sequences generated by‘aggrecanase’ and matrix metalloproteinase cleavage of aggrecan: application tocatabolism in situ and in vitro. Biochem. J. 305: 799–804.
27. KOLB, C., S. MAUCH, H.H. PETER, U. KRAWINKEL & R. SEDLACEK. 1997. The matrixmetalloproteinase RASI-1 is expressed in synovial blood vessels of a rheumatoidarthritis patient. Immunol. Lett. 57: 83–88.
28. SEDLACEK, R., S. MAUCH, B. KOLB, C. SCHÄTZLEIN, H. EIBEL, H.H. PETER,J. SCHMITT & U. KRAWINKEL. 1998. Matrix metalloproteinase MMP-19 (RASI 1) isexpressed on the surface of activated peripheral blood mononuclear cells and isdetected as an autoantigen in rheumatoid arthritis. Immunobiology 198: 408–423.
29. ATKINSON, S.J., T. CRABBE, S. COWELL, R.V. WARD, M.J. BUTLER, H. SATO,M. SEIKI, J.J. REYNOLDS & G. MURPHY. 1995. Intermolecular autolytic cleavage cancontribute to the activation of progelatinase A by cell membranes. J. Biol. Chem.270: 30479–30485.




















