
2638.full.pdf - The Journal of Immunology
14
of May 28, 2022. This information is current as Migration Inflammatory Cytokines: Role in Cell (Matrix Metalloproteinase-9) Is Regulated by Human Mast Cell-Derived Gelatinase B Geczy, Jonathan P. Arm and Nicodemus Tedla Wakefield, H. Patrick McNeil, Weixing Yan, Carolyn Nick Di Girolamo, Ikuko Indoh, Nicole Jackson, Denis http://www.jimmunol.org/content/177/4/2638 doi: 10.4049/jimmunol.177.4.2638 2006; 177:2638-2650; ; J Immunol References http://www.jimmunol.org/content/177/4/2638.full#ref-list-1 , 28 of which you can access for free at: cites 62 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2006 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on May 28, 2022 http://www.jimmunol.org/ Downloaded from by guest on May 28, 2022 http://www.jimmunol.org/ Downloaded from
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of 2638.full.pdf - The Journal of Immunology

of May 28, 2022.This information is current as
MigrationInflammatory Cytokines: Role in Cell(Matrix Metalloproteinase-9) Is Regulated by Human Mast Cell-Derived Gelatinase B
Geczy, Jonathan P. Arm and Nicodemus TedlaWakefield, H. Patrick McNeil, Weixing Yan, Carolyn Nick Di Girolamo, Ikuko Indoh, Nicole Jackson, Denis
http://www.jimmunol.org/content/177/4/2638doi: 10.4049/jimmunol.177.4.2638
2006; 177:2638-2650; ;J Immunol
Referenceshttp://www.jimmunol.org/content/177/4/2638.full#ref-list-1
, 28 of which you can access for free at: cites 62 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

Human Mast Cell-Derived Gelatinase B (MatrixMetalloproteinase-9) Is Regulated by Inflammatory Cytokines:Role in Cell Migration1
Nick Di Girolamo,* Ikuko Indoh,* Nicole Jackson,* Denis Wakefield,* H. Patrick McNeil,*Weixing Yan,* Carolyn Geczy,* Jonathan P. Arm,† and Nicodemus Tedla2*
Mast cells are key effectors in the pathogenesis of inflammatory and tissue destructive diseases such as rheumatoid arthritis (RA).These cells contain specialized secretory granules loaded with bioactive molecules including cytokines, growth factors, and pro-teases that are released upon activation. This study investigated the regulation of matrix metalloproteinase MMP-9 (gelatinase B)in human mast cells by cytokines that are known to be involved in the pathogenesis of RA. Immunohistochemical staining ofsynovial tissue showed abundant expression of MMP-9 by synovial tissue mast cells in patients with RA but not in normal controls.The expression, activity, and production of MMP-9 in mast cells was confirmed by RT-PCR, zymography, and Western blottingusing cord blood-derived human mast cells (CB-HMC). Treatment of CB-HMC with TNF-� significantly increased the expressionof MMP-9 mRNA and up-regulated the activity of MMP-9 in a time- and dose-dependent manner. By contrast, IFN-� inhibitedMMP-9 mRNA and protein expression. The cytokine-mediated regulation of MMP-9 was also apparent in the human mast cellline (HMC-1) and in mouse bone marrow-derived mast cells. Furthermore, TNF-� significantly increased the invasiveness ofCB-HMC across Matrigel-coated membranes while the addition of IFN-�, rTIMP-1, or pharmacological MMP inhibitors signif-icantly reduced this process. These observations suggest that MMP-9 is not a stored product in mast cells but these cells arecapable of producing this enzyme under inflammatory conditions that may facilitate the migration of mast cell progenitors to sitesof inflammation and may also contribute to local tissue damage. The Journal of Immunology, 2006, 177: 2638–2650.
M ast cells are derived from hemopoietic progenitor cellsthat home to tissue as committed progenitors (1–3).Maturation and differentiation of mast cells occurs in
tissue in response to local production of stem cell factor (SCF)3
(4). Mast cells can also undergo significant change in number or inphenotype during allergic or nonallergic inflammation (1, 5–9).These cells play a crucial role in IgE-dependent immune responsesthat mediate immediate hypersensitivity reactions associated withallergic phenomena and host resistance to parasites (5, 10). Theyalso participate in innate immunity to bacterial infection (11) andplay a direct role in the pathogenesis of a mouse model of inflam-matory arthritis (12). There is considerable circumstantial evidenceimplicating mast cells in the pathogenesis of human rheumatoidarthritis (RA). These include the production of inflammatory (13,14) and tissue destructive (15, 16) mediators. Moreover, a signif-icant increase in mast cell numbers has been documented in human
synovial tissue derived from patients with RA (8, 9). Despite theextensive knowledge regarding the mediators that regulate mastcell phenotype and number (10), factors that regulate the homingof mast cell progenitors from the bone marrow to the tissue ofresidence are not well-defined.
Matrix metalloproteinases (MMPs) are a family of neutral pro-teolytic enzymes active against all major components of the ex-tracellular matrix. To date, 24 members have been cloned andcharacterized in humans and are divided into four groups depend-ing on structure and substrate preference (17). Broadly, they in-clude the collagenases, gelatinases, stromelysins, and the mem-brane-type MMPs. Gelatinase B (92-kDa, MMP-9) and gelatinaseA (72-kDa, MMP-2) are the largest members of the MMP familyand are to date the only gelatinases identified. MMP docking mol-ecules such as CD44 for MMP-9 (18) and membrane-type MMPsfor MMP-2 (17) have been identified on the surface of variouscells. Cells use these proteinases to promote local proteolysis andto enhance their invasive potential (18, 19). Proteolytically activeand inducible levels of MMPs have been observed in synovial cellsfrom patients with RA (20).
MMP-9 is regulated posttranscriptionally at multiple levels. Forexample, its activity is neutralized by binding to naturally occur-ring tissue inhibitors of metalloproteinase (TIMP-1 to TIMP-4)with preferential binding observed between MMP-9 and TIMP-1(21). MMPs (including MMP-9) are generally synthesized and se-creted as latent soluble enzymes that require activation in the ex-tracellular space. Several studies have identified mast cell-derivedchymase and tryptase as potent activators of MMPs (22, 23).MMPs are also regulated at the level of transcription and theirexpression is modulated by a variety of stimuli including cytokines(20, 24, 25), growth factors (26), and cell-to-cell and cell-to-matrixinteractions (27–29). Interestingly, the expression of MMP-2
*Inflammatory Diseases Research Unit, School of Medical Sciences, University ofNew South Wales, Sydney, New South Wales, Australia; and †Division of Rheuma-tology, Immunology and Allergy, Brigham and Women’s Hospital and Department ofMedicine, Harvard Medical School, Boston, MA 02115
Received for publication January 28, 2005. Accepted for publication May 23, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the National Health and Medical Research Council ofAustralia (project grant), the Arthritis Foundation of Australia, and a PostdoctoralFellowship from the American Arthritis Foundation.2 Address correspondence and reprint requests to Dr. Nicodemus Tedla, InflammatoryDiseases Research Unit, School of Medical Sciences, University of New South Wales,Sydney, New South Wales, 2052 Australia. E-mail address: [email protected] Abbreviations used in this paper: SCF, stem cell factor; RA, rheumatoid arthritis;MMP, matrix metalloproteinase; rh, recombinant human; TIMP, tissue inhibitor ofmetalloproteinase; CB-HMC, cord blood derived-human mast cell; mBMMC, mousebone marrow-derived mast cell.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

rarely varies, and it is regarded as a housekeeping-like gene inMMP biology.
MMP-9 was first identified in neutrophils (30) but these gran-ulocytes lack TIMP-1 (31). MMP-9 is also a product of other in-flammatory leukocytes including T cells, macrophages, and eosin-ophils (32). Relevant to acute and chronic inflammation, MMP-9can cleave IL-8 and potentiate its activity as a neutrophil chemoat-tractant by at least 10-fold (18). There is now compelling evidencethat along with the arsenal of stored serine proteases, mast cells arealso a major source of MMPs such as MMP-1 (33), MMP-3 (34),and MMP-9 (23, 35–37). Although there is limited evidence forthe expression of MMP-9 in mast cells in rheumatoid synovium(35), its regulation in RA is poorly understood. In the currentstudy, we confirm MMP-9 expression in rheumatoid synovial mastcells and for the first time, demonstrate its regulation by TNF-�and IFN-� in cord blood-derived human mast cells (CB-HMC),bone marrow-derived mouse mast cells (mBMMC), and the humanmast cell line-1 (HMC-1). Furthermore, we provide evidence im-plicating TNF-inducible MMP-9 in the migration of mast cellsthrough matrix proteins.
Materials and MethodsPatients
Formalin-fixed paraffin-embedded archival synovial tissue from patientswith RA (n � 3) and normal controls (n � 3) was obtained from a tissuecollection held at the Department of Pathology, University of New SouthWales (Sydney, Australia). Institutional human ethics committee approvalwas obtained for this study.
Immunohistochemical studies
Serial sections (2–4 �m) of formalin-fixed paraffin-embedded synovialtissue were used for immunohistochemical studies. Specific mouse IgG1mAbs against MMP-9 (Ab-8) and mast cell tryptase (AA1) were purchasedfrom Oncogene Research Products and DakoCytomation, respectively. Anirrelevant mouse IgG1 negative control Ab was purchased from DakoCy-tomation. These Abs were used in a modified three-step alkaline phospha-tase staining procedure (38). In brief, paraffin-embedded adjacent sectionswere dewaxed and equilibrated with TBS (pH 8.0) followed by a 5-min Agretrieval by microwave in 0.01 M citrate buffer (pH 6.2). The citrate bufferwas allowed to cool, sections were re-equilibrated in TBS, then blockedwith 20% goat serum for 20 min at room temperature. Sections were in-cubated with anti-MMP-9 (4 �g/ml), anti-tryptase (5 �g/ml), or an isotypecontrol (5 �g/ml) Ab in 2% BSA/TBS overnight at 4°C. Sections werewashed extensively with TBS, and then incubated with a biotinylated goatanti-mouse IgG (Vector Laboratories) for 1 h at room temperature. Sec-tions were rinsed in TBS, and incubated with streptavidin-alkaline phos-phatase conjugate (Vector Laboratories) for 45 min at room temperature.Immunoreactivity was detected using an alkaline phosphatase substrate(Vector Red; Vector Laboratories) and the sections were briefly counter-stained with hematoxylin.
Morphometry
Overlapping images (1345 � 1033 pixels), each spanning an area of �1.5mm2, were taken from sections using an Olympus BX60 microscope witha �10 objective linked to Spot Advanced software version 3.5.6 for Win-dows (Diagnostic Instruments). Multiple images taken from each sectionwere stitched into a single continuous image using Photoshop version 8(Adobe Systems) software and the total surface area was measured. Adja-cent sections that were stained for tryptase and MMP-9 were overlapped,rotated until they were aligned, and tryptase-positive and MMP-9/tryptasedouble-positive cells were counted. Finally, the proportion of MMP-9-pos-itive mast cells from each tissue specimen was calculated. An area of1.56–2.43 mm2 from two to three images was sufficient to count over 100mast cells in RA synovium. However, it was necessary to count the entirenormal synovium tissue section (5.84–7.35 mm2) to acquire �100 mastcells.
Human mast cell cultures
Cord blood from human placentas was obtained after routine Caesariansection from the Australian Cord Blood Bank at the Royal Women’s Hos-pital (Sydney, Australia). Cord blood was collected in accordance with
established institutional guidelines and ethics committee approval. CB-HMCs were established from the mononuclear cell fraction of the cordblood as described (39, 40). In brief, heparin-treated cord blood was sedi-mented with 4.5% dextran solution to remove erythrocytes. Buffy coatswere layered onto Ficoll-Hypaque (Pharmacia) and the mononuclear cellinterface was obtained after centrifugation. After repeated washes withPBS containing 5 mM EDTA, mononuclear cells were suspended at 2 �106/ml in high glucose RPMI 1640 (Invitrogen Life Technologies) con-taining 10% FBS, 2 mM L-glutamine, 0.1 mM nonessential amino acids,100 U/ml penicillin, 100 mg/ml streptomycin, 2 �g/ml gentamicin (allfrom Sigma-Aldrich), and 0.2 �M 2-ME ((Invitrogen Life Technologies).Cells were cultured in the presence of 100 ng/ml SCF (gift from Amgen),50 ng/ml IL-6 (R&D Systems), and 10 ng/ml IL-10 (R&D Systems). Thenonadherent cells were transferred every week for up to 10 wk into culturemedium containing fresh cytokines. The purity and maturity of cells wasassessed weekly by flow cytometry using a fluorochrome-conjugatedmouse Ab directed against human c-kit (CD117; BD Pharmingen), bymetachromatic staining of cytospin preparations using toluidine blue, andby immunostaining for both tryptase (DakoCytomation) and chymase(Chemicon International) using acetone-fixed cytospin preparations. Ma-turity was defined by �95% CD117high cells, �95% toluidine blue posi-tivity, and positive immunostaining for tryptase and chymase. Once cellsreached maturity, no other immunocytochemical or functional differenceswere noted between 7- and 12-wk-old cells. Therefore, cells were used forthis study when they reached �95% toluidine blue positivity rather than aspecific number of weeks in culture.
The human mast cell leukemia cell line (HMC-1) was provided Dr. J.Butterfield (Division of Allergic Diseases and Internal Medicine, MayoClinic, Rochester, MN) and cultured in the same medium as the CB-HMC(RPMI 1640 complete medium without growth factors). Because of thefunctional heterogeneity of the HMC-1 bulk culture, a limiting dilutionassay was performed to isolate individual HMC-1. Cells from passages 5to 8 of the newly propagated clones were used.
Mouse mast cell cultures
mBMMC were obtained by culturing primary bone marrow cells fromfemale 6- to 8-wk-old BALB/c mice (BRC) in 50% RPMI 1640 completemedium, 30% WEHI-3B supernatant (as a source of murine IL-3) and 20%3T3 supernatant (as a source of murine SCF) (41). The nonadherent cellswere transferred every week for up to 4 wk into fresh culture medium. Thepurity and maturity of cells was assessed weekly by flow cytometry usingfluorochrome-conjugated rat Ab against mouse c-kit (CD117; BD Pharm-ingen) and by metachromatic staining of cytospin preparations using tolu-idine blue. Maturity was defined by �95% CD117high cells and �95%toluidine blue positivity. Cells reached �95% maturity within 3–5 wk andno differences in morphology and function were noted between 3- and5-wk cells. Cells were used when they reached �95% toluidine blue pos-itivity rather than a specific number of weeks in culture.
MMP-9 production by human and mouse mast cells aftercytokine stimulation
CB-HMC were washed twice with medium alone and resuspended at 2 �106 cells/ml in assay medium (2.5% FBS/RPMI 1640 and SCF (100 ng/ml)). HMC-1 and mBMMC were resuspended in the same medium and atthe same density, but without SCF. Human mast cells were incubated with25 ng/ml TNF-� (R&D Systems) or 25 ng/ml IFN-� (Endogen) in 24-wellflat-bottom Falcon plates at 37°C and 5% CO2. mBMMC were incubatedwith 25 ng/ml recombinant mouse TNF-� (R&D Systems), 12.5 ng/mlrecombinant mouse IFN-� (R&D Systems), or the combination of both for24–36 h. For some experiments, RNA was extracted and supernatants har-vested for gelatin-substrate zymography and Western blotting, and eachpreparation was stored at �70°C.
To assess the effects of IFN-� on TNF-�-mediated induction of MMP-9,CB-HMC were incubated with the optimal concentration of TNF-� (25ng/ml) and increasing doses of IFN-� (0.005–50 ng/ml). Total RNA wasextracted after 4 h for RT-PCR analysis and supernatants were collected24 h after stimulation for zymography. To determine whether production ofMMP-9 from CB-HMC was due to de novo protein synthesis, cells werestimulated with 25 ng/ml TNF-� and coincubated with varying concentra-tions of cycloheximide (0–0.5 �g/ml; Sigma-Aldrich) for 24 h.
Quantitation of histamine released by CB-HMC
Degranulation of CB-HMC was induced after passive IgE sensitization andanti-IgE activation as described elsewhere (40). In brief, CB-HMC werewashed twice with RPMI 1640 and resuspended at 2 � 106 cells/ml in
2639The Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

culture medium containing 100 ng/ml SCF. Cells were primed with semi-purified human myeloma IgE (10 �g/ml; Chemicon International) and IL-4(10 ng/ml; Endogen) for 5 days. After washing with PBS containing 0.1%BSA (ICN Biomedicals), cells were activated with 1 �g/ml rabbit anti-human IgE Ab in 0.1% BSA/PBS for 30 min at 37°C in 5% CO2. For someexperiments, cells were treated for 4–8 h with varying concentrations ofA23187 (Sigma-Aldrich) or 50 ng/ml phorbol ester (PMA; Sigma-Aldrich). Supernatants were harvested and stored at �70°C before hista-mine determination, zymography, and Western blot analysis (see below).Some cells were collected by centrifugation, resuspended in RPMI 1640,and lysed by rapid freeze/thawing (three cycles). Histamine in the super-natant and cell pellet fractions was measured by ELISA (ICN Biomedi-cals). Percentage of histamine release was quantitated by the equation:histamine in supernatant/(histamine in supernatant � histamine inpellet) � 100.
Gelatin-substrate zymography
Zymography was performed as previously described (25, 33, 42, 43). Su-pernatants standardized for cell numbers were diluted 1/4 using serum-freeculture medium and 8 �l of nonreducing sample buffer (0.25M Tris-HCl(pH 6.8), 10% SDS, 4% sucrose, 0.1% bromphenol blue) was added to 25�l of diluted culture supernatant and loaded without boiling onto 10%SDS-PAGE gels containing 1 mg/ml gelatin (Sigma-Aldrich). After elec-trophoresis, gels were rinsed twice for 45 min in 2.5% Triton X-100(Sigma-Aldrich), incubated overnight at 37°C in substrate buffer (50 mMTris-HCl (pH 7.4), 10 mM CaCl2, and 0.02% NaN3), stained with Coo-massie Blue R-250 (Bio-Rad) for 1.5 h, then destained (3 � 50 min) toexpose gelatinolytic bands. A low range molecular mass protein ladder(Bio-Rad) was run in adjacent lanes. MMP identity was verified by addingproteinase inhibitors such as 10 mM EDTA, 1 mM 10-phenanthroline(Sigma-Aldrich), and 2 mM 4(�2-aminoethyl) benzene-sulfonyl fluoride(ABSF; Sigma-Aldrich) to the substrate buffer. Gelatinolytic bands weresemiquantified with the Gel Doc 2000 and the Quantity One program(Bio-Rad).
Western blotting
Western blotting was performed as previously described (25, 33, 43). Cul-ture supernatant from control or cytokine-stimulated CB-HMC (25 �l) waselectrophoretically separated on SDS-PAGE using 4% stacking and 10%resolving gels under nonreducing conditions. Proteins were transferred toPolyScreen polyvinylidene difluoride transfer membranes (PerkinElmerLife Sciences), blocked in 5% skim milk powder in TBST for 1 h, washedbriefly in TBST, then incubated with 5 �g/ml primary mAb directedagainst human MMP-9 (Ab-8; Calbiochem-Novabiochem) diluted in 5%BSA/TBST for 1 h at room temperature. Membranes were then extensivelywashed in TBST (3 � 5 min) and incubated with a 1/2000 dilution ofHRP-conjugated rabbit anti-mouse Ab (DakoCytomation) for 1 h at roomtemperature. Membranes were again washed (3 � 5 min) in TBST, thenplaced in chemiluminescent reagent for 1 min (Western Lightning;PerkinElmer Life Science) and exposed to Hyperfilm MP (Amersham Bio-sciences). A prestained low molecular mass protein ladder (Bio-Rad) wasrun in adjacent lanes.
RNA extraction and RT-PCR
Total RNA was extracted (RNAgents Total RNA Extraction kit; Promega)from control and cytokine-stimulated CB-HMC after 4 h. Reverse tran-scription was performed using the Preamplification System for First StrandcDNA Synthesis kit (Invitrogen Life Technologies) and as previously de-scribed (25, 33, 43) using Superscript III. Aliquots (4 �l) of cDNA wereamplified by PCR using 100 nM each of the forward 5�-TTC AAG GCTGGG AAG TAC TG-3� and reverse 5�-TCC GGA GGG CCC CGG TCACCT-3� primers for MMP-9, TIMP-1, and GAPDH (43). A 2-min hot startat 95oC was performed to denature the double stranded cDNA, followed by37 cycles of PCR (each cycle: 95°C, 30 s; 55°C, 30 s; 72°C, 30 s) and thereactions terminated with a 2-min extension at 72°C. Cycle number waspredetermined so that the products generated were within the linear portionof the amplification curve. PCR products were visualized on 2% agarosegels, precast with ethidium bromide and semiquantified with the Gel Doc2000 and the Quantity One program (Bio-Rad). A 100-bp ladder (Invitro-gen Life Technologies) was run in adjacent lanes.
Migration of CB-HMC through Matrigel
CB-HMC were washed twice in RPMI 1640 and resuspended at 2 �106cells/ml in RPMI 1640 complete medium containing 100 ng/ml SCF.Cells were preincubated for 6 h in the same medium with or withoutTNF-� (25 ng/ml), IFN-� (25 ng/ml), or a combination of TNF-� and
IFN-� in 15-ml round-bottom Falcon tubes at 37°C and 5% CO2. Cellswere washed once with 10 ml of binding buffer (1% BSA, 25 mM HEPESin RPMI 1640) then adjusted to 1.5 � 106 cells/ml. Polyvinylpyrrolidone-free membranes (10 �m pore, 25 � 80 mm; Osmonics) were coated witha 1/6 dilution of Matrigel (Sigma-Aldrich) in 75 mM HEPES/RPMI 1640for 2 h at room temperature then 10 �g/ml human fibronectin (InvitrogenLife Technologies) was applied for 1 h at room temperature and allowed toair dry.
Varying concentrations (10�10 to 10�8 M) of the mast cell chemoat-tractant C5a (R&D Systems) or binding buffer alone in a final volume of30 �l were dispensed in triplicate into the lower well of a 48-well chemo-taxis chamber (Neuro Probe). A suspension of cytokine-stimulated CB-HMC (50 �l) was dispensed into the upper wells of the chemotaxis cham-ber that was separated by a coated membrane. Chambers weresubsequently incubated at 37°C in a humid 5% CO2 incubator for 14 h, themembranes were removed, rinsed in PBS, and the upper surface gentlyscraped over with a wiper blade to remove noninvading cells. Membraneswere stained with Diff Quick according the manufacturer’s instruction (LabAids), mounted on glass slides and coverslipped. Cells that had invaded tothe lower surface were counted under high power (�400) using a lightmicroscope. Three randomly selected fields were counted from each welland each treatment consisted of triplicate wells giving nine counts pertreatment. The migration index from three independent experiments wasthen calculated by dividing the mean cell counts for each treatment dividedby counts from corresponding medium only control.
To determine whether the migration of CB-HMC through Matrigel wasindeed MMP dependent, migration assays were performed in the presenceof several MMP inhibitors. A broad-spectrum synthetic MMP inhibitor(BB3103) was provided by Dr. A. J. Gearing (British Biotech, Oxford,U.K.). Recombinant human (rh) TIMP-1 and TIMP-2 were purchased fromCalbiochem-Novabiochem and dexamethasone was purchased fromSigma-Aldrich. Briefly, CB-HMC were preincubated in RPMI 1640 com-plete medium that contained 100 ng/ml SCF and TNF-� (25 ng/ml) for 6 hat 37°C in 5% CO2. Cells were then washed once with 10 ml of bindingbuffer and adjusted to 1.5 � 106 cells/ml in binding buffer containingBB3103 (0–10��), rhTIMP-1/-2 (0–5 �g/ml), or dexamethasone (0–5��). Cells in binding medium alone were used as positive control. Todetermine toxicity, a 100-�l cell suspension containing each inhibitor at therespective concentration was dispensed into the wells of a 96-well flat-bottom plate. Chemotaxis chambers and 96-well plates were incubated for14 h at 37°C in 5% CO2. Membranes were rinsed, stained, and migrated cellscounted as described above. Supernatants from the 96-well plates were har-vested for zymographic analysis and cells were used for viability assays usinga fluorometric, 96-well plate LIVE/DEAD Viability assay kit (MolecularProbes).
Cell surface expression of C5a receptor and c-kit on CB-HMCby flow cytometry
CB-HMC (2 � 106/ml) incubated in RPMI 1640 complete medium con-taining 100 ng/ml SCF with or without TNF-� (25 ng/ml) for 6 h at 37°Cin 5% CO2 were washed with 10 ml of binding buffer and an aliquot (1 �106 cells) was sampled (6-h time point). The remaining cells (1 � 106
cells/ml) were transferred to a 96-well flat-bottom plate in binding buffercontaining 5% FBS and incubated at 37°C in 5% CO2 for 14 h. Beforestaining, cells were washed with cold PBS containing 0.05% NaN3 and 1%BSA (PAB buffer) and resuspended in the same buffer at 2 � 106/ml.Human serum (10% final) was added and cells (50 �l) were incubated for30 min at room temperature with 5 �g/ml unconjugated mouse IgG2a mAbto C5a R (CD88) (clone 7F3; a gift from Prof. C. Mackay, Garvan Institutefor Medical Research, Sydney, Australia), control mouse IgG2a (5 �g/ml;Serotec), saturating amounts of PE-conjugated mouse IgG1 mAb to c-kit(CD117; BD Pharmingen), or with PE-conjugated control mouse IgG1 (BDPharmingen). After washing in cold PAB buffer, 500 �l of 1% parafor-maldehyde in PBS was added to the cell suspensions and stained withconjugated Abs before storage at 4°C in the dark. Cells stained with uncon-jugated Abs were incubated on ice for 45 min with 10 �l (10 �g/ml) ofFITC-conjugated F(ab�)2 goat anti-mouse IgG (F(ab�)2-specific), secondaryAb (Jackson ImmunoResearch Laboratories). Cells were washed twice withPAB buffer, fixed with 1% paraformaldehyde in PBS, and analyzed by usinga FACScan flow cytometer (BD Biosciences). A unimodal shift in fluores-cence intensity of cells stained with specific Abs compared with cells stainedwith the isotype-matched negative control Abs was considered positive.
Statistical analysis
Significance of mast cell degranulation in response to A23187, PMA, andIgE cross-linking was determined using two-tailed t tests. Statistical sig-nificance in the activity of MMP-9 in mast cells treated with various doses
2640 INDUCTION OF MMP-9 BY TNF-� IN HUMAN MAST CELLS
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
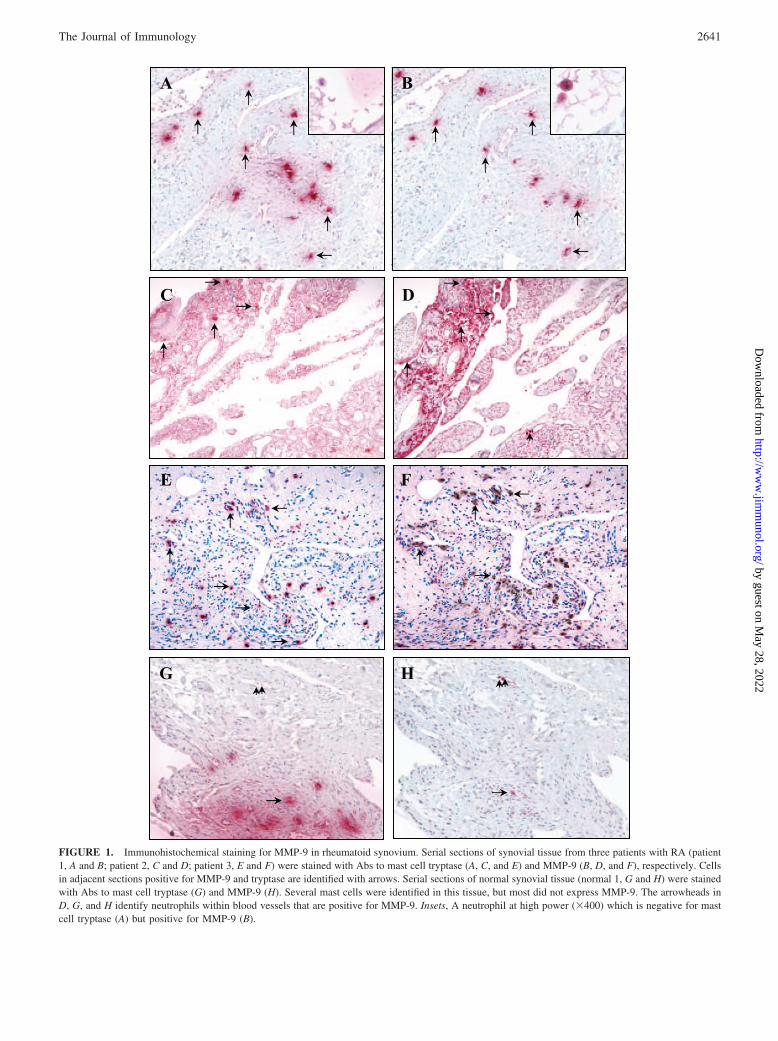
FIGURE 1. Immunohistochemical staining for MMP-9 in rheumatoid synovium. Serial sections of synovial tissue from three patients with RA (patient1, A and B; patient 2, C and D; patient 3, E and F) were stained with Abs to mast cell tryptase (A, C, and E) and MMP-9 (B, D, and F), respectively. Cellsin adjacent sections positive for MMP-9 and tryptase are identified with arrows. Serial sections of normal synovial tissue (normal 1, G and H) were stainedwith Abs to mast cell tryptase (G) and MMP-9 (H). Several mast cells were identified in this tissue, but most did not express MMP-9. The arrowheads inD, G, and H identify neutrophils within blood vessels that are positive for MMP-9. Insets, A neutrophil at high power (�400) which is negative for mastcell tryptase (A) but positive for MMP-9 (B).
2641The Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

of TNF-� or IFN-� at multiple time points was determined using theKruskal-Wallis test followed by Dunn’s multiple comparison posttest.Changes in migration between treatment groups were analyzed usingANOVA with Tukey’s posttest. Modulation of cell migration after treat-ment with MMP-9 inhibitors was determined using a two-tailed t test. Avalue of p 0.05 was considered significant.
ResultsProduction of MMP-9 by human mast cells in vivo
Upon stimulation, mast cells release a battery of enzymes that aregenerally stored in specific granules. In addition to the well-char-acterized serine proteases, previous studies have identified mem-bers of the MMPs in these cells (22, 33–37, 44). Initially, it wasour intention to confirm these primarily in vitro observations invivo, using synovial tissue obtained from patients with RA byimmunohistochemistry. Staining for MMP-9 (Fig. 1, B, D, and F)was observed in tryptase-positive (Fig. 1A, C, and E) mast cells inthe RA synovium whereas in normal human synovium, despite thepresence of several tryptase-positive mast cells (Fig. 1G), therewas limited immunoreactivity for MMP-9 in these cells (Fig.1H). MMP-9 was also identified in synovial lining cells, mac-rophages (data not shown) and intravascular neutrophils in RAtissue (Fig. 1, B, inset, D, arrowhead) and normal synovium(Fig. 1H, arrowhead). Quantitative analysis of tissue sections
demonstrated substantially more tryptase-positive mast cells inRA synovium with an average of 57.3 12 cells/mm2 com-pared with 13.0 1.8 cells/mm2 in normal synovium (Table I).There were also higher proportions of MMP-9-positive mastcells in RA tissue (65.4 9.5%) compared with normal syno-vium (1.1 1.1%) (Table I).
Characterization of CB-HMCs
To confirm expression of MMP-9 by human mast cells, primarymast cells were propagated from cord blood and characterized.These cells displayed typical morphological, phenotypic, andfunctional characteristics of mast cells including dense granularcytoplasm (Fig. 2A), intense metachromatic staining (�95%)with acidic toluidine blue (Fig. 2A), and abundant expression(�97%) of tryptase (Fig. 2B) and 95–99% chymase positively(Fig. 2C). Flow cytometric analysis demonstrated high intensitycell surface expression for c-kit/CD117 on �95% of these cells(Fig. 2D).
Production of MMP-9 by CB-HMC
CB-HMCs were used to determine whether MMP-9 is stored inmast cell granules or whether it is newly synthesized and se-creted upon mast cell activation. To test the former hypothesis,
FIGURE 2. Characterization of hu-man cord blood-derived mast cells. Cy-tospins from 8-wk-old CB-HMC werestained metachromatically with tolu-idine blue (A) or histochemically withAbs to mast cell tryptase (B) and chy-mase (C). Flow cytometric analysis oncell suspension (D) was also performedto show surface c-kit (CD117) expres-sion. Results are representative of over10 independent mast cell cultures fromdifferent donors.
Table I. Proportion of MMP-9-positive mast cells in synovial tissue of RA patients and control subjectsa
SubjectsTryptase-Positive
Mast Cells% MMP-9-Positive
Mast CellsTotal Area Counted
(mm2) Mast Cells/mm2
RA1 118 66.1 1.77 67RA2 110 55.5 2.43 45RA3 102 74.5 1.56 65N1 91 2.2 7.35 12.4N2 84 1.2 5.84 14.4N3 97 0 9.01 10.8
a RA, Synovial tissue obtained from rheumatoid arthritis; N, normal synovial tissue obtained from patients undergoingpostinjury knee surgery.
2642 INDUCTION OF MMP-9 BY TNF-� IN HUMAN MAST CELLS
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
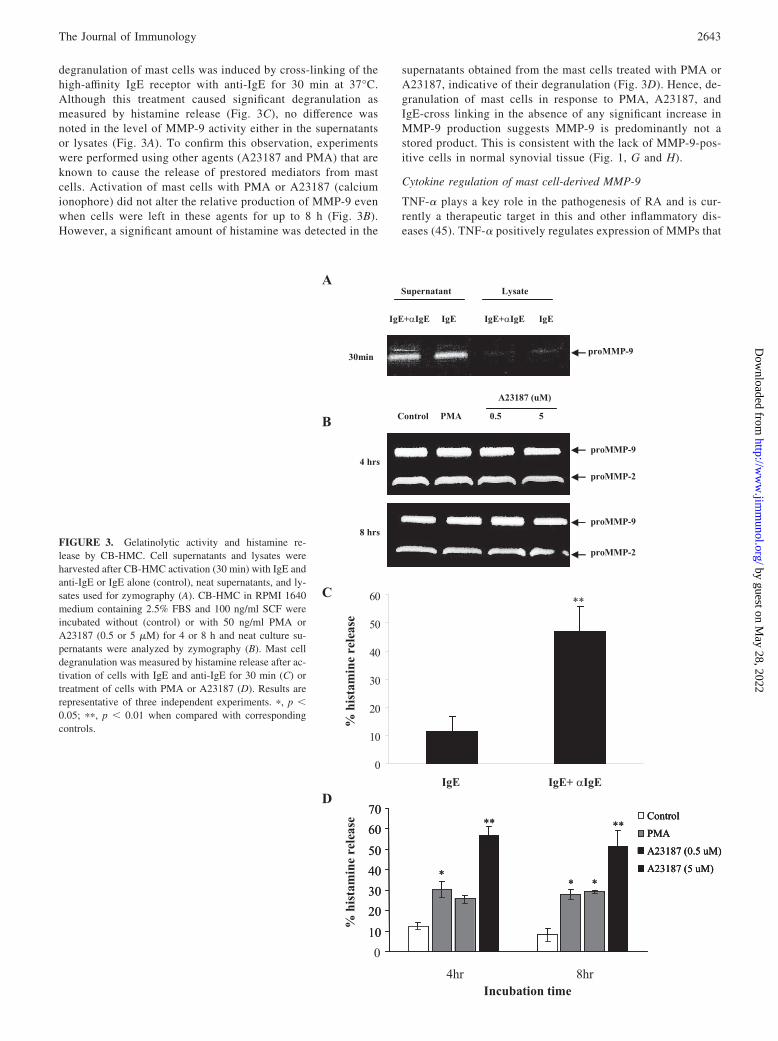
degranulation of mast cells was induced by cross-linking of thehigh-affinity IgE receptor with anti-IgE for 30 min at 37°C.Although this treatment caused significant degranulation asmeasured by histamine release (Fig. 3C), no difference wasnoted in the level of MMP-9 activity either in the supernatantsor lysates (Fig. 3A). To confirm this observation, experimentswere performed using other agents (A23187 and PMA) that areknown to cause the release of prestored mediators from mastcells. Activation of mast cells with PMA or A23187 (calciumionophore) did not alter the relative production of MMP-9 evenwhen cells were left in these agents for up to 8 h (Fig. 3B).However, a significant amount of histamine was detected in the
supernatants obtained from the mast cells treated with PMA orA23187, indicative of their degranulation (Fig. 3D). Hence, de-granulation of mast cells in response to PMA, A23187, andIgE-cross linking in the absence of any significant increase inMMP-9 production suggests MMP-9 is predominantly not astored product. This is consistent with the lack of MMP-9-pos-itive cells in normal synovial tissue (Fig. 1, G and H).
Cytokine regulation of mast cell-derived MMP-9
TNF-� plays a key role in the pathogenesis of RA and is cur-rently a therapeutic target in this and other inflammatory dis-eases (45). TNF-� positively regulates expression of MMPs that
FIGURE 3. Gelatinolytic activity and histamine re-lease by CB-HMC. Cell supernatants and lysates wereharvested after CB-HMC activation (30 min) with IgE andanti-IgE or IgE alone (control), neat supernatants, and ly-sates used for zymography (A). CB-HMC in RPMI 1640medium containing 2.5% FBS and 100 ng/ml SCF wereincubated without (control) or with 50 ng/ml PMA orA23187 (0.5 or 5 �M) for 4 or 8 h and neat culture su-pernatants were analyzed by zymography (B). Mast celldegranulation was measured by histamine release after ac-tivation of cells with IgE and anti-IgE for 30 min (C) ortreatment of cells with PMA or A23187 (D). Results arerepresentative of three independent experiments. �, p 0.05; ��, p 0.01 when compared with correspondingcontrols.
2643The Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

are responsible for tissue destruction that characterizes RA.Therefore, we determined whether this cytokine induced theexpression of MMP-9 in mast cells that are typically present inrheumatoid synovium and are believed to play a key role in thepathogenesis of the disease. Treatment of CB-HMCs withTNF-� caused significant up-regulation of MMP-9 in a time-dependent manner (peak 18 h) (Fig. 4). By contrast, treatmentof these cells with IFN-� significantly suppressed MMP-9 pro-duction (Fig. 4A) with the greatest decrease noted at the 24-htime point (Fig. 4B). This is consistent with previous studiesthat have demonstrated the down-regulation MMP-9 by IFN-�in macrophages (46, 47).
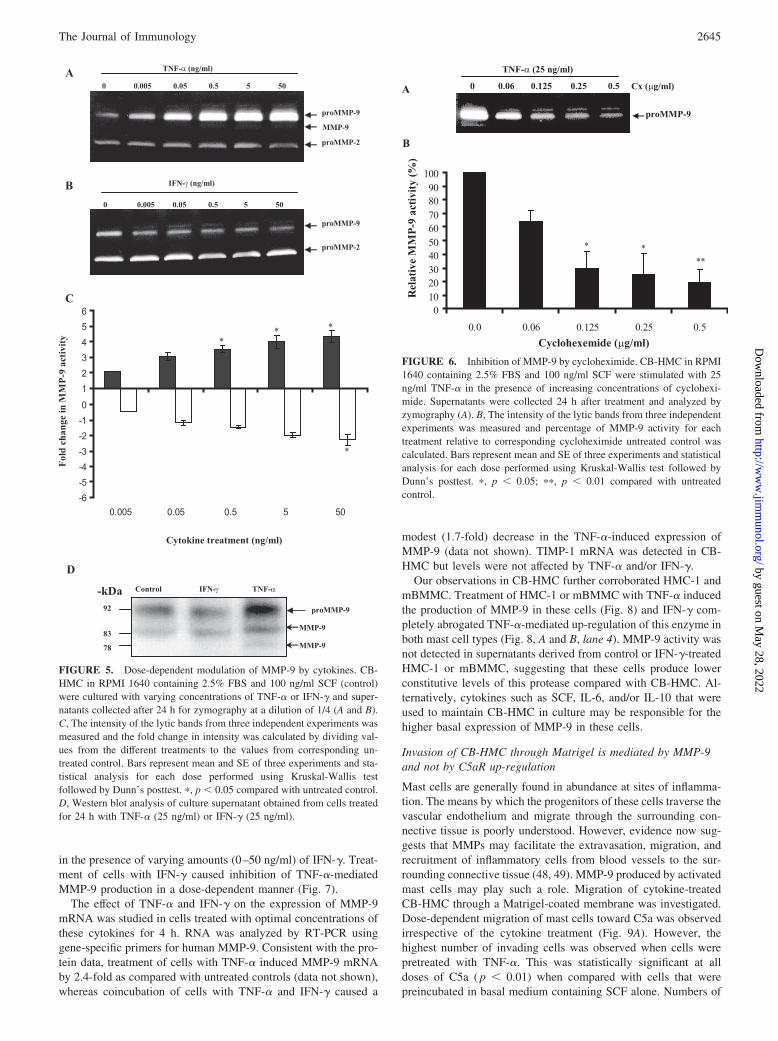
Following the kinetic studies, CB-HMCs were treated for24 h with varying concentrations of TNF-� (0 –50 ng/ml) andsupernatants analyzed by zymography. MMP-9 production wassignificantly induced by TNF-� in a dose-dependent manner,while MMP-2 remained unchanged (Fig. 5A). MaximumMMP-9 production was observed when cells were stimulatedwith 5–50 ng/ml TNF-� (Fig. 5C). Conversely, when CB-HMCs were treated with IFN-�, constitutive MMP-9 levels sig-nificantly decreased (Fig. 5B), while MMP-2 was not affected.Maximum inhibition was observed with 5–50 ng/ml IFN-�(Fig. 5C).
To confirm the identity of the 92-kDa gelatinolytic band as ametalloproteinase, zymograms were incubated in substrate
buffer containing EDTA (metal chelator) or 1,10-phenanthroline (anMMP inhibitor). Both agents abolished all activity associated withMMP-2 and -9 (data not shown, Refs. 42 and 43)), whereas the serineprotease inhibitor ABSF had no effect on the activity of MMP-2 and-9 (data not shown). Moreover, stimulation with TNF-� not only in-creased expression of the 92-kDa band, but also induced the appear-ance of a minor 83-kDa component (Fig. 4A, 24- to 72-h arrows; Fig.5A, arrow), a size typical of activated MMP-9. Western blotting con-firmed the identity of the 92-kDa band as MMP-9 that was increasedafter treatment with TNF-� (Fig. 5D). An immunoreactive band at 83kDa that corresponds to the active form of MMP-9 was observedfollowing TNF-� stimulation (Fig. 5D). Interestingly, TIMP-1, a nat-ural inhibitor of MMP-9 protein, was not detected in the same super-natants by Western blotting (data not shown) suggesting that activa-tion of mast cells by TNF-� might be biased toward the process ofincreased tissue permeability and degradation.
To determine whether MMP-9 production in CB-HMC was dueto de novo protein synthesis, cells were stimulated with 25 ng/mlTNF-� and coincubated with varying concentrations of cyclohex-imide (0–0.5 �g/ml). MMP-9 was dose-dependently inhibited bycycloheximide and this was visualized by a decrease in intensity ofthis proteinase (Fig. 6A). Inhibition of MMP-9 was statisticallysignificant with 0.125–0.5 �g/ml cycloheximide (Fig. 6B).
The effects of IFN-� on the TNF-�-mediated induction ofMMP-9 were examined by stimulating cells with 25 ng/ml TNF-�
FIGURE 4. Time course production of MMP-9 in cytokine-stimulated CB-HMC. CB-HMC in RPMI 1640 containing 2.5% FBS and 100 ng/ml SCFwere stimulated with either 25 ng/ml TNF-� or 25 ng/ml IFN-� and supernatants were collected at the indicated time points for zymographic analysis (A).Small arrows in A identify active MMP-9 (83 kDa) evident at 24–72 h after TNF-� stimulation. B, The intensity of the lytic bands from three independentexperiments was measured and the fold change in intensity was calculated by dividing values from the different treatments to the values from correspondinguntreated control. Bars represent mean and SE of three experiments and statistical analysis for each time point performed using Kruskal-Wallis test followedby Dunn’s posttest. �, p 0.05; ��, p 0.01 compared with untreated control.
2644 INDUCTION OF MMP-9 BY TNF-� IN HUMAN MAST CELLS
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
in the presence of varying amounts (0–50 ng/ml) of IFN-�. Treat-ment of cells with IFN-� caused inhibition of TNF-�-mediatedMMP-9 production in a dose-dependent manner (Fig. 7).
The effect of TNF-� and IFN-� on the expression of MMP-9mRNA was studied in cells treated with optimal concentrations ofthese cytokines for 4 h. RNA was analyzed by RT-PCR usinggene-specific primers for human MMP-9. Consistent with the pro-tein data, treatment of cells with TNF-� induced MMP-9 mRNAby 2.4-fold as compared with untreated controls (data not shown),whereas coincubation of cells with TNF-� and IFN-� caused a
modest (1.7-fold) decrease in the TNF-�-induced expression ofMMP-9 (data not shown). TIMP-1 mRNA was detected in CB-HMC but levels were not affected by TNF-� and/or IFN-�.
Our observations in CB-HMC further corroborated HMC-1 andmBMMC. Treatment of HMC-1 or mBMMC with TNF-� inducedthe production of MMP-9 in these cells (Fig. 8) and IFN-� com-pletely abrogated TNF-�-mediated up-regulation of this enzyme inboth mast cell types (Fig. 8, A and B, lane 4). MMP-9 activity wasnot detected in supernatants derived from control or IFN-�-treatedHMC-1 or mBMMC, suggesting that these cells produce lowerconstitutive levels of this protease compared with CB-HMC. Al-ternatively, cytokines such as SCF, IL-6, and/or IL-10 that wereused to maintain CB-HMC in culture may be responsible for thehigher basal expression of MMP-9 in these cells.
Invasion of CB-HMC through Matrigel is mediated by MMP-9and not by C5aR up-regulation
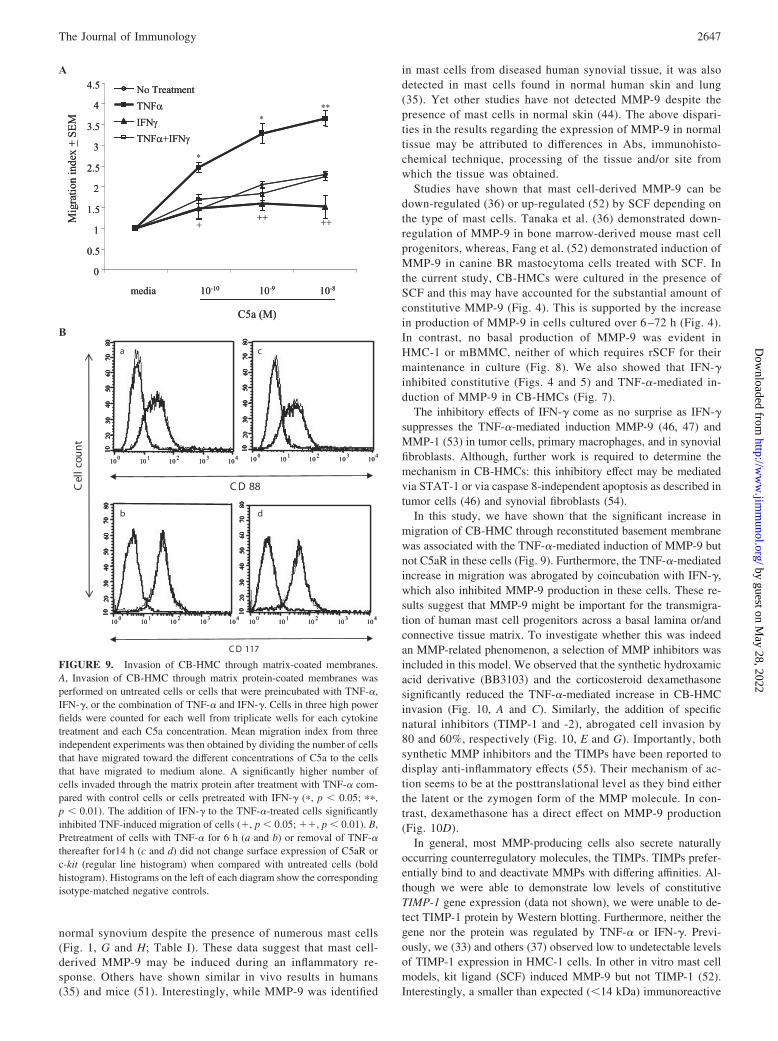
Mast cells are generally found in abundance at sites of inflamma-tion. The means by which the progenitors of these cells traverse thevascular endothelium and migrate through the surrounding con-nective tissue is poorly understood. However, evidence now sug-gests that MMPs may facilitate the extravasation, migration, andrecruitment of inflammatory cells from blood vessels to the sur-rounding connective tissue (48, 49). MMP-9 produced by activatedmast cells may play such a role. Migration of cytokine-treatedCB-HMC through a Matrigel-coated membrane was investigated.Dose-dependent migration of mast cells toward C5a was observedirrespective of the cytokine treatment (Fig. 9A). However, thehighest number of invading cells was observed when cells werepretreated with TNF-�. This was statistically significant at alldoses of C5a ( p 0.01) when compared with cells that werepreincubated in basal medium containing SCF alone. Numbers of
FIGURE 5. Dose-dependent modulation of MMP-9 by cytokines. CB-HMC in RPMI 1640 containing 2.5% FBS and 100 ng/ml SCF (control)were cultured with varying concentrations of TNF-� or IFN-� and super-natants collected after 24 h for zymography at a dilution of 1/4 (A and B).C, The intensity of the lytic bands from three independent experiments wasmeasured and the fold change in intensity was calculated by dividing val-ues from the different treatments to the values from corresponding un-treated control. Bars represent mean and SE of three experiments and sta-tistical analysis for each dose performed using Kruskal-Wallis testfollowed by Dunn’s posttest. �, p 0.05 compared with untreated control.D, Western blot analysis of culture supernatant obtained from cells treatedfor 24 h with TNF-� (25 ng/ml) or IFN-� (25 ng/ml).
FIGURE 6. Inhibition of MMP-9 by cycloheximide. CB-HMC in RPMI1640 containing 2.5% FBS and 100 ng/ml SCF were stimulated with 25ng/ml TNF-� in the presence of increasing concentrations of cyclohexi-mide. Supernatants were collected 24 h after treatment and analyzed byzymography (A). B, The intensity of the lytic bands from three independentexperiments was measured and percentage of MMP-9 activity for eachtreatment relative to corresponding cycloheximide untreated control wascalculated. Bars represent mean and SE of three experiments and statisticalanalysis for each dose performed using Kruskal-Wallis test followed byDunn’s posttest. �, p 0.05; ��, p 0.01 compared with untreatedcontrol.
2645The Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
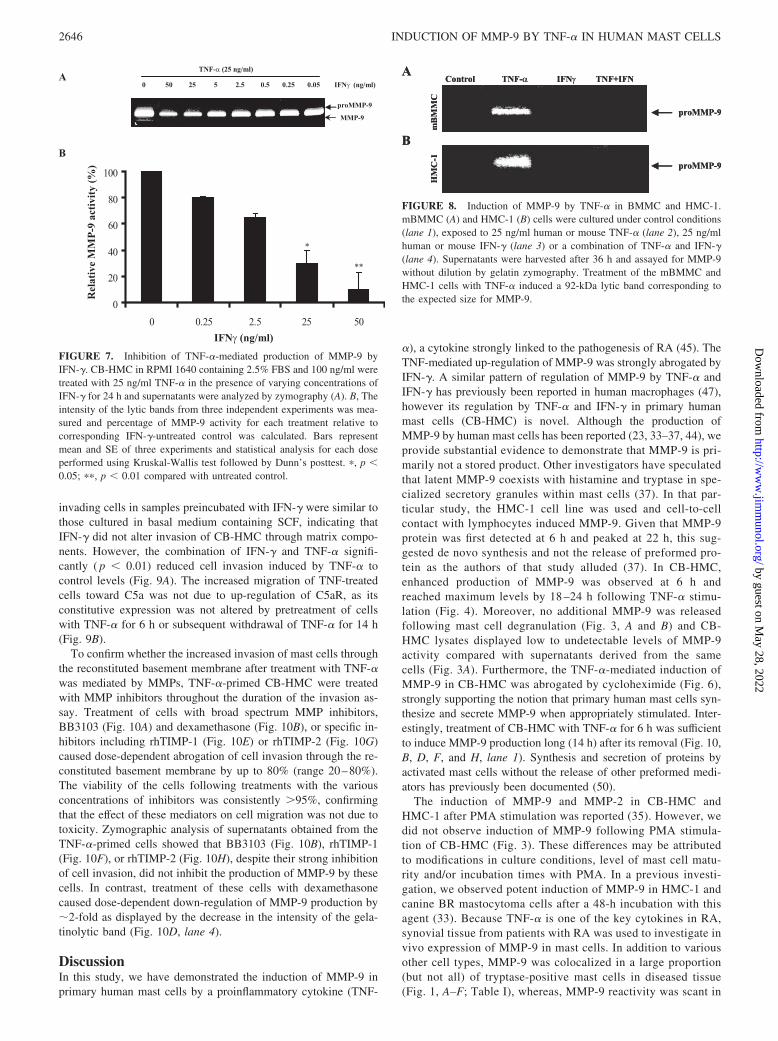
invading cells in samples preincubated with IFN-� were similar tothose cultured in basal medium containing SCF, indicating thatIFN-� did not alter invasion of CB-HMC through matrix compo-nents. However, the combination of IFN-� and TNF-� signifi-cantly ( p 0.01) reduced cell invasion induced by TNF-� tocontrol levels (Fig. 9A). The increased migration of TNF-treatedcells toward C5a was not due to up-regulation of C5aR, as itsconstitutive expression was not altered by pretreatment of cellswith TNF-� for 6 h or subsequent withdrawal of TNF-� for 14 h(Fig. 9B).
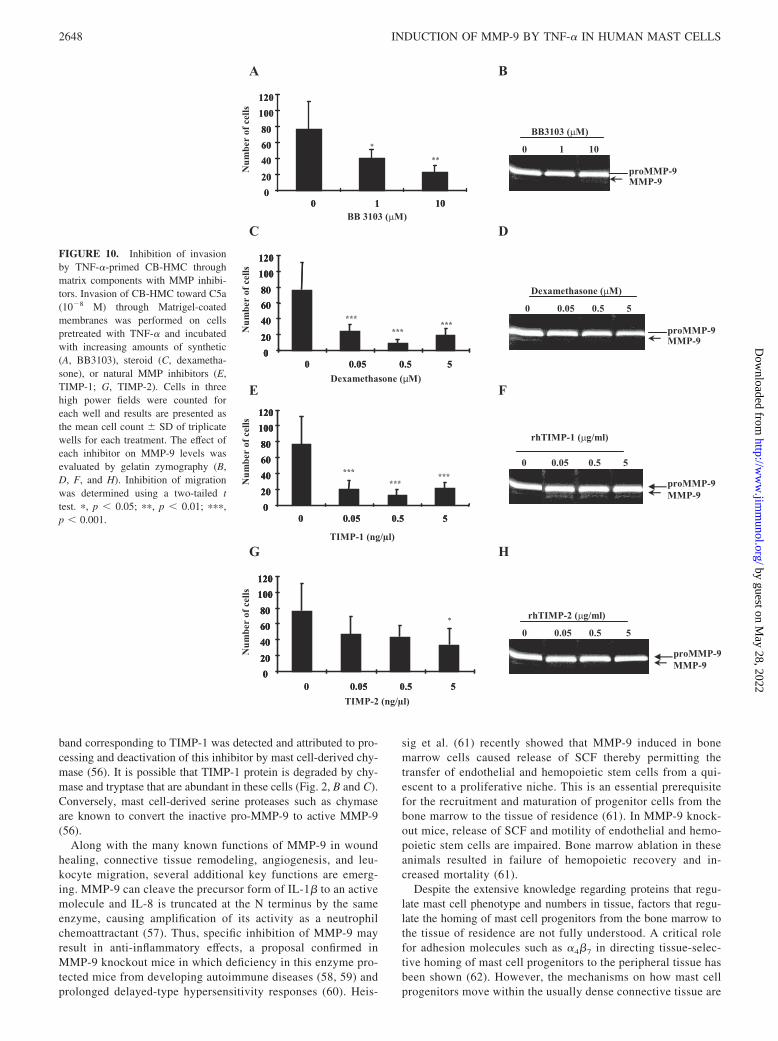
To confirm whether the increased invasion of mast cells throughthe reconstituted basement membrane after treatment with TNF-�was mediated by MMPs, TNF-�-primed CB-HMC were treatedwith MMP inhibitors throughout the duration of the invasion as-say. Treatment of cells with broad spectrum MMP inhibitors,BB3103 (Fig. 10A) and dexamethasone (Fig. 10B), or specific in-hibitors including rhTIMP-1 (Fig. 10E) or rhTIMP-2 (Fig. 10G)caused dose-dependent abrogation of cell invasion through the re-constituted basement membrane by up to 80% (range 20–80%).The viability of the cells following treatments with the variousconcentrations of inhibitors was consistently �95%, confirmingthat the effect of these mediators on cell migration was not due totoxicity. Zymographic analysis of supernatants obtained from theTNF-�-primed cells showed that BB3103 (Fig. 10B), rhTIMP-1(Fig. 10F), or rhTIMP-2 (Fig. 10H), despite their strong inhibitionof cell invasion, did not inhibit the production of MMP-9 by thesecells. In contrast, treatment of these cells with dexamethasonecaused dose-dependent down-regulation of MMP-9 production by�2-fold as displayed by the decrease in the intensity of the gela-tinolytic band (Fig. 10D, lane 4).
DiscussionIn this study, we have demonstrated the induction of MMP-9 inprimary human mast cells by a proinflammatory cytokine (TNF-
�), a cytokine strongly linked to the pathogenesis of RA (45). TheTNF-mediated up-regulation of MMP-9 was strongly abrogated byIFN-�. A similar pattern of regulation of MMP-9 by TNF-� andIFN-� has previously been reported in human macrophages (47),however its regulation by TNF-� and IFN-� in primary humanmast cells (CB-HMC) is novel. Although the production ofMMP-9 by human mast cells has been reported (23, 33–37, 44), weprovide substantial evidence to demonstrate that MMP-9 is pri-marily not a stored product. Other investigators have speculatedthat latent MMP-9 coexists with histamine and tryptase in spe-cialized secretory granules within mast cells (37). In that par-ticular study, the HMC-1 cell line was used and cell-to-cellcontact with lymphocytes induced MMP-9. Given that MMP-9protein was first detected at 6 h and peaked at 22 h, this sug-gested de novo synthesis and not the release of preformed pro-tein as the authors of that study alluded (37). In CB-HMC,enhanced production of MMP-9 was observed at 6 h andreached maximum levels by 18 –24 h following TNF-� stimu-lation (Fig. 4). Moreover, no additional MMP-9 was releasedfollowing mast cell degranulation (Fig. 3, A and B) and CB-HMC lysates displayed low to undetectable levels of MMP-9activity compared with supernatants derived from the samecells (Fig. 3A). Furthermore, the TNF-�-mediated induction ofMMP-9 in CB-HMC was abrogated by cycloheximide (Fig. 6),strongly supporting the notion that primary human mast cells syn-thesize and secrete MMP-9 when appropriately stimulated. Inter-estingly, treatment of CB-HMC with TNF-� for 6 h was sufficientto induce MMP-9 production long (14 h) after its removal (Fig. 10,B, D, F, and H, lane 1). Synthesis and secretion of proteins byactivated mast cells without the release of other preformed medi-ators has previously been documented (50).
The induction of MMP-9 and MMP-2 in CB-HMC andHMC-1 after PMA stimulation was reported (35). However, wedid not observe induction of MMP-9 following PMA stimula-tion of CB-HMC (Fig. 3). These differences may be attributedto modifications in culture conditions, level of mast cell matu-rity and/or incubation times with PMA. In a previous investi-gation, we observed potent induction of MMP-9 in HMC-1 andcanine BR mastocytoma cells after a 48-h incubation with thisagent (33). Because TNF-� is one of the key cytokines in RA,synovial tissue from patients with RA was used to investigate invivo expression of MMP-9 in mast cells. In addition to variousother cell types, MMP-9 was colocalized in a large proportion(but not all) of tryptase-positive mast cells in diseased tissue(Fig. 1, A–F; Table I), whereas, MMP-9 reactivity was scant in
FIGURE 7. Inhibition of TNF-�-mediated production of MMP-9 byIFN-�. CB-HMC in RPMI 1640 containing 2.5% FBS and 100 ng/ml weretreated with 25 ng/ml TNF-� in the presence of varying concentrations ofIFN-� for 24 h and supernatants were analyzed by zymography (A). B, Theintensity of the lytic bands from three independent experiments was mea-sured and percentage of MMP-9 activity for each treatment relative tocorresponding IFN-�-untreated control was calculated. Bars representmean and SE of three experiments and statistical analysis for each doseperformed using Kruskal-Wallis test followed by Dunn’s posttest. �, p 0.05; ��, p 0.01 compared with untreated control.
FIGURE 8. Induction of MMP-9 by TNF-� in BMMC and HMC-1.mBMMC (A) and HMC-1 (B) cells were cultured under control conditions(lane 1), exposed to 25 ng/ml human or mouse TNF-� (lane 2), 25 ng/mlhuman or mouse IFN-� (lane 3) or a combination of TNF-� and IFN-�(lane 4). Supernatants were harvested after 36 h and assayed for MMP-9without dilution by gelatin zymography. Treatment of the mBMMC andHMC-1 cells with TNF-� induced a 92-kDa lytic band corresponding tothe expected size for MMP-9.
2646 INDUCTION OF MMP-9 BY TNF-� IN HUMAN MAST CELLS
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
normal synovium despite the presence of numerous mast cells(Fig. 1, G and H; Table I). These data suggest that mast cell-derived MMP-9 may be induced during an inflammatory re-sponse. Others have shown similar in vivo results in humans(35) and mice (51). Interestingly, while MMP-9 was identified
in mast cells from diseased human synovial tissue, it was alsodetected in mast cells found in normal human skin and lung(35). Yet other studies have not detected MMP-9 despite thepresence of mast cells in normal skin (44). The above dispari-ties in the results regarding the expression of MMP-9 in normaltissue may be attributed to differences in Abs, immunohisto-chemical technique, processing of the tissue and/or site fromwhich the tissue was obtained.
Studies have shown that mast cell-derived MMP-9 can bedown-regulated (36) or up-regulated (52) by SCF depending onthe type of mast cells. Tanaka et al. (36) demonstrated down-regulation of MMP-9 in bone marrow-derived mouse mast cellprogenitors, whereas, Fang et al. (52) demonstrated induction ofMMP-9 in canine BR mastocytoma cells treated with SCF. Inthe current study, CB-HMCs were cultured in the presence ofSCF and this may have accounted for the substantial amount ofconstitutive MMP-9 (Fig. 4). This is supported by the increasein production of MMP-9 in cells cultured over 6 –72 h (Fig. 4).In contrast, no basal production of MMP-9 was evident inHMC-1 or mBMMC, neither of which requires rSCF for theirmaintenance in culture (Fig. 8). We also showed that IFN-�inhibited constitutive (Figs. 4 and 5) and TNF-�-mediated in-duction of MMP-9 in CB-HMCs (Fig. 7).
The inhibitory effects of IFN-� come as no surprise as IFN-�suppresses the TNF-�-mediated induction MMP-9 (46, 47) andMMP-1 (53) in tumor cells, primary macrophages, and in synovialfibroblasts. Although, further work is required to determine themechanism in CB-HMCs: this inhibitory effect may be mediatedvia STAT-1 or via caspase 8-independent apoptosis as described intumor cells (46) and synovial fibroblasts (54).
In this study, we have shown that the significant increase inmigration of CB-HMC through reconstituted basement membranewas associated with the TNF-�-mediated induction of MMP-9 butnot C5aR in these cells (Fig. 9). Furthermore, the TNF-�-mediatedincrease in migration was abrogated by coincubation with IFN-�,which also inhibited MMP-9 production in these cells. These re-sults suggest that MMP-9 might be important for the transmigra-tion of human mast cell progenitors across a basal lamina or/andconnective tissue matrix. To investigate whether this was indeedan MMP-related phenomenon, a selection of MMP inhibitors wasincluded in this model. We observed that the synthetic hydroxamicacid derivative (BB3103) and the corticosteroid dexamethasonesignificantly reduced the TNF-�-mediated increase in CB-HMCinvasion (Fig. 10, A and C). Similarly, the addition of specificnatural inhibitors (TIMP-1 and -2), abrogated cell invasion by80 and 60%, respectively (Fig. 10, E and G). Importantly, bothsynthetic MMP inhibitors and the TIMPs have been reported todisplay anti-inflammatory effects (55). Their mechanism of ac-tion seems to be at the posttranslational level as they bind eitherthe latent or the zymogen form of the MMP molecule. In con-trast, dexamethasone has a direct effect on MMP-9 production(Fig. 10D).
In general, most MMP-producing cells also secrete naturallyoccurring counterregulatory molecules, the TIMPs. TIMPs prefer-entially bind to and deactivate MMPs with differing affinities. Al-though we were able to demonstrate low levels of constitutiveTIMP-1 gene expression (data not shown), we were unable to de-tect TIMP-1 protein by Western blotting. Furthermore, neither thegene nor the protein was regulated by TNF-� or IFN-�. Previ-ously, we (33) and others (37) observed low to undetectable levelsof TIMP-1 expression in HMC-1 cells. In other in vitro mast cellmodels, kit ligand (SCF) induced MMP-9 but not TIMP-1 (52).Interestingly, a smaller than expected (14 kDa) immunoreactive
FIGURE 9. Invasion of CB-HMC through matrix-coated membranes.A, Invasion of CB-HMC through matrix protein-coated membranes wasperformed on untreated cells or cells that were preincubated with TNF-�,IFN-�, or the combination of TNF-� and IFN-�. Cells in three high powerfields were counted for each well from triplicate wells for each cytokinetreatment and each C5a concentration. Mean migration index from threeindependent experiments was then obtained by dividing the number of cellsthat have migrated toward the different concentrations of C5a to the cellsthat have migrated to medium alone. A significantly higher number ofcells invaded through the matrix protein after treatment with TNF-� com-pared with control cells or cells pretreated with IFN-� (�, p 0.05; ��,p 0.01). The addition of IFN-� to the TNF-�-treated cells significantlyinhibited TNF-induced migration of cells (�, p 0.05; ��, p 0.01). B,Pretreatment of cells with TNF-� for 6 h (a and b) or removal of TNF-�thereafter for14 h (c and d) did not change surface expression of C5aR orc-kit (regular line histogram) when compared with untreated cells (boldhistogram). Histograms on the left of each diagram show the correspondingisotype-matched negative controls.
2647The Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
band corresponding to TIMP-1 was detected and attributed to pro-cessing and deactivation of this inhibitor by mast cell-derived chy-mase (56). It is possible that TIMP-1 protein is degraded by chy-mase and tryptase that are abundant in these cells (Fig. 2, B and C).Conversely, mast cell-derived serine proteases such as chymaseare known to convert the inactive pro-MMP-9 to active MMP-9(56).
Along with the many known functions of MMP-9 in woundhealing, connective tissue remodeling, angiogenesis, and leu-kocyte migration, several additional key functions are emerg-ing. MMP-9 can cleave the precursor form of IL-1� to an activemolecule and IL-8 is truncated at the N terminus by the sameenzyme, causing amplification of its activity as a neutrophilchemoattractant (57). Thus, specific inhibition of MMP-9 mayresult in anti-inflammatory effects, a proposal confirmed inMMP-9 knockout mice in which deficiency in this enzyme pro-tected mice from developing autoimmune diseases (58, 59) andprolonged delayed-type hypersensitivity responses (60). Heis-
sig et al. (61) recently showed that MMP-9 induced in bonemarrow cells caused release of SCF thereby permitting thetransfer of endothelial and hemopoietic stem cells from a qui-escent to a proliferative niche. This is an essential prerequisitefor the recruitment and maturation of progenitor cells from thebone marrow to the tissue of residence (61). In MMP-9 knock-out mice, release of SCF and motility of endothelial and hemo-poietic stem cells are impaired. Bone marrow ablation in theseanimals resulted in failure of hemopoietic recovery and in-creased mortality (61).
Despite the extensive knowledge regarding proteins that regu-late mast cell phenotype and numbers in tissue, factors that regu-late the homing of mast cell progenitors from the bone marrow tothe tissue of residence are not fully understood. A critical rolefor adhesion molecules such as �4�7 in directing tissue-selec-tive homing of mast cell progenitors to the peripheral tissue hasbeen shown (62). However, the mechanisms on how mast cellprogenitors move within the usually dense connective tissue are
FIGURE 10. Inhibition of invasionby TNF-�-primed CB-HMC throughmatrix components with MMP inhibi-tors. Invasion of CB-HMC toward C5a(10�8 M) through Matrigel-coatedmembranes was performed on cellspretreated with TNF-� and incubatedwith increasing amounts of synthetic(A, BB3103), steroid (C, dexametha-sone), or natural MMP inhibitors (E,TIMP-1; G, TIMP-2). Cells in threehigh power fields were counted foreach well and results are presented asthe mean cell count SD of triplicatewells for each treatment. The effect ofeach inhibitor on MMP-9 levels wasevaluated by gelatin zymography (B,D, F, and H). Inhibition of migrationwas determined using a two-tailed ttest. �, p 0.05; ��, p 0.01; ���,p 0.001.
2648 INDUCTION OF MMP-9 BY TNF-� IN HUMAN MAST CELLS
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

unclear. In this study, we have demonstrated increased migra-tion of CB-HMC through reconstituted basement membranethat was associated with increased production of MMP-9 (Figs.9A and 10). Thus, MMP-9 produced by mast cell progenitorsmay enable these cells to migrate through connective tissue andrelease SCF by a membrane cleavage mechanism thereby gen-erating conditions that sustain mast cell maturation and differ-entiation. It is possible that the significant increase in mast cellsobserved during allergic or nonallergic inflammation (1, 5, 6, 8,9) might be due to TNF-�-induced MMP-9 production that fa-cilitates migration of mast cell progenitors and causes localrelease of SCF. This suggestion is supported by our demonstra-tion of increased numbers of MMP-9-positive mast cells in in-flamed synovial tissue. Modulation of MMP-9 production bymast cells might therefore be a useful therapeutic strategy tominimize the inflammatory reaction in RA.
AcknowledgmentsWe acknowledge Dr. John Hunt (Department of Pathology, University ofNew South Wales) for providing us with normal and diseased synovialtissue, Dr. Taline Hampartzoumian for her technical and editorial advice,and Paul Halasz for his assistance in histomorphometry.
DisclosuresThe authors have no financial conflict of interest.
References1. Nakano, T., T. Sonoda, C. Hayashi, A. Yamatodani, Y. Kanayama,
T. Yamamura, H. Asai, T. Yonezawa, Y. Kitamura, and S. J. Galli. 1985. Fate ofbone marrow-derived cultured mast cells after intracutaneous, intraperitoneal andintravenous transfer into genetically mast cell-deficient W/Wv mice: evidence thatcultured mast cells can give rise to both connective tissue type and mucosal mastcells. J. Exp. Med. 162: 1025–1043.
2. Metcalf, D. D., D. Baram, and Y. A. Mekori. 199. Mast cells. Physiol. Rev. 77:1033–1079.
3. Williams, C. M., and S. J. Galli. 2000. The diverse potential effector and immu-noregulatory roles of mast cells in allergic diseases. J. Allergy Clin. Immunol.105: 847–859.
4. Galli, S. J., K. M. Zsebo, and E. N. Geissler. 1994. The kit ligand, stem cellfactor. Adv. Immunol. 55: 1–16.
5. Madden, K. B., J. F. Urban Jr., H. J. Ziltener, J. W. Schrader, F. D. Finkelman,and I. M. Katona. 1991. Antibodies to IL-3 and IL-4 suppress helminth-inducedintestinal mastocytosis. J. Immunol. 147: 1387–1391.
6. Friend, D. S., N. Ghildyal, K. F. Austen, M. F. Gurish, R. Matsumoto, andR. L. Stevens. 1996. Mast cells that reside at different locations in the jejunum ofmice infected with Trichinella spiralis exhibit sequential changes in their granuleultrastructure and chymase phenotype. J. Cell Biol. 135: 279–290.
7. Galli, S. J. 2000. Mast cells and basophils. Curr. Opin. Hematol. 7: 32–39.8. Malone, D. G., R. L. Wilder, A. M. Saaverdra-Delgado, and D. D. Metcalfe.
1987. Mast cell numbers in rheumatoid synovial tissues: correlations with quan-titative measures of lymphocytic infiltration and modulation by anti-inflammatorytherapy. Arthritis Rheum. 30: 130–137.
9. Gotis-Graham, I., and H. P. McNeil. 1997. Mast cell responses in rheumatoidsynovium: association of the MCTC subset with matrix turnover and clinicalprogression. Arthritis Rheum. 40: 479–489.
10. Abraham, S. N., and R. Malaviya. 1997. Mast cells in infection and immunity.Infect. Immun. 65: 3501–3508.
11. Echtenacher, B., D. N. Mannuel, and L. Hultner. 1996. Critical protective role ofmast cells in a model of acute septic peritonitis. Nature 381: 75–77.
12. Lee, D. M., D. S. Friend, M. F. Gurish, C. Benolst, D. Mathis, and M. B. Brenner.2002. Mast cells: a cellular link between autoantibodies and inflammatory arthri-tis. Science 297: 1689–1692.
13. He, S., M. D. Gaca, and A. F. Walls. 2001. The activation of synovial mast cells:modulation of histamine release by tryptase and chymase and their inhibitors.Eur. J. Pharmacol. 412: 223–229.
14. Woolley, D. E., and L. C. Tetlow. 2000. Mast cell activation and its relation toproinflammatory cytokine production in the rheumatoid lesion. Arthritis Res. 2:65–74.
15. Tetlow, L. C., and D. E. Woolley. 1995. Mast cells, cytokines and metallopro-teinases at the rheumatoid lesion: dual immunolocalisation studies. Ann. Rheum.Dis. 54: 896–903.
16. Tetlow, L. C., N. Harper, T. Dunningham, M. A. Morris, H. Bertfield, andD. E. Woolley. 1998. Effects of induced mast cell activation on prostaglandin Eand metalloproteinase production by rheumatoid synovial tissue in vitro. Ann.Rheum. Dis. 57: 25–32.
17. Sternlicht, M. D., and Z. Werb. 2001. How matrix metalloproteinases regulatecell behaviour. Annu. Rev. Cell Dev. Biol. 17: 463–516.
18. Yu, Q., and I. Stamenkovic. 2000. Cell surface-localized matrix metalloprotein-ase-9 proteolytically activates TGF-� and promotes tumor invasion and angio-genesis. Genes Dev. 14: 163–176.
19. Baumann, P., P. Zigrino, C. Mauch, D. Breitkreutz, and R. Nischt. 2000. Mem-brane-type 1 matrix metalloproteinase-mediated progelatinase A activation innon-tumorigenic and tumorigenic human keratinocytes. Br. J. Cancer 83:1387–1393.
20. Mengshol, J. A., S. M. Kimberlee, and C. E. Brinckerhoff. 2002. Matrix metal-loproteinases as therapeutic targets in arthritic diseases. Arthritis Rheum. 46:13–20.
21. Brew, K., D. Dinakarpandian, and H. Nagase. 2000. Tissue inhibitors of metal-loproteinases: evolution, structure and function. Biochim. Biophys. Acta 1477:267–283.
22. Lohi, J., I. Harvima, and J. Keski-Oja. 1992. Pericellular substrates of humanmast cell tryptase: 72,000 dalton gelatinase and fibronectin. J. Cell. Biochem. 50:337–349.
23. Fang, K. C., W. W. Raymond, S. C. Lazarus, and G. H. Caughey. 1996. Dogmastocytoma cells secrete a 92-kD gelatinase activated extracellularly by mastcell chymase. J. Clin. Invest. 97: 1589–1596.
24. Ries, C., and P. E. Petrides. 1995. Cytokine regulation of matrix metalloprotein-ase activity and its regulatory dysfunction in disease. Biol. Chem. Hoppe Seyler376: 345–355.
25. Di Girolamo, N., P. McCluskey, A. Lloyd, M. T. Coroneo, and D. Wakefield.2000. Expression of MMPs and TIMPs in human pterygia and cultured pterygiumepithelial cells. Invest. Ophthalmol. Vis. Sci. 41: 671–679.
26. Vaday, G. G., H. Schor, M. A. Rahat, N. Lahat, and O. Lider. 2001. Transforminggrowth factor-� suppresses tumor necrosis factor �-induced matrix metallopro-teinase-9 expression in monocytes. J. Leukocyte Biol. 69: 613–621.
27. Lacraz, S., P. Isler, E. Vey, H. G. Welgus, and J. M. Dayer. 1994. Direct contactbetween T lymphocytes and monocytes is a major pathway for induction ofmetalloproteinase expression. J. Biol. Chem. 269: 22027–22033.
28. Aoudjit, F., E. F. Potworowski, and Y. St-Pierre. 1998. Bi-directional inductionof matrix metalloproteinase-9 and tissue inhibitor of matrix metalloproteinase-1during T lymphoma/endothelial cell contact: implication of ICAM-1. J. Immunol.160: 2967–2973.
29. Horwitz, A. R., and Z. Werb. 1998. Cell-to-cell contact and extracellular matrixcell adhesion and the extracellular matrix: recent progress and emerging themes.Curr. Opin. Cell Biol. 10: 562–565.
30. Sopata, I., and A. M. Dancewics. 1974. Presence of a gelatin-specific proteinaseand its latent form in human leucocytes. Biochim. Biophys. Acta 370: 510–523.
31. Hibbs, M. S., K. A. Hasty, J. M. Seyer, A. H. Kang, and C. L. Mainardi. 1985.Biochemical and immunological characterization of the secreted forms of humanneutrophil gelatinase. J. Biol. Chem. 260: 2493–2500.
32. Okada, S., H. Kita, T. J. George, G. J. Gleich, and K. M. Leiferman. 1997.Migration of eosinophils through basement membrane components in vitro:role of matrix metalloproteinase-9. Am. J. Respir Cell Mol. Biol. 17:519 –528.
33. Di Girolamo, N., and D. Wakefield. 2000. In vitro and in vivo expression ofinterstitial collagenase/MMP-1 by human mast cells. Dev. Immunol. 7:131–142.
34. Brownell, E., L. Fiorentino, G. Jolly, K. Wolfe, S. Kincaid, P. Seperack, andD. Visco. 1995. Immunolocalization of stromelysin-related protein in murinemast cell granules. Int. Arch. Allergy Immunol. 107: 333–335.
35. Kanbe, N., A. Tanaka, M. Kanbe, A. Itakura, M. Kurosawa, and H. Matsuda.1999.Human mast cells produce matrix metalloproteinase 9. Eur. J. Immunol. 29:2645–2649.
36. Tanaka, A., K. Aria, Y. Kitamura, and H. Matsuda. 1999. Matrix metallopro-teinase-9 production, a newly identified function of mast cell progenitors, isdownregulated by c-kit receptor activation. Blood 94: 2390–2395.
37. Baram, D., G. G. Vaday, P. Salamon, I. Drucker, R. Hershkoviz, andY. A. Mekori. 2001. Human mast cells release metalloproteinase-9 on contactwith activated T cells: juxtacrine regulation by TNF-�. J. Immunol. 167:4008–4016.
38. Tedla, N., K. Gibson, H. P. McNeil, D. Cosman, L. Borges, and J. P. Arm. 2002.The expression of activating and inhibitory leukocyte immunoglobulin-like re-ceptors in rheumatoid synovium. Am. J. Pathol. 160: 425–431.
39. Ochi, H., W. M. Hirani, Q. Yuan, D. S. Friend, K. F. Austen, and J. A. Boyce.1999. T helper cell type 2 cytokine-mediated comitogenic responses and CCR3expression during differentiation of human mast cells in vitro. J. Exp. Med. 190:267–280.
40. Hsieh, F. H., B. K. Lam, J. F. Penrose, K. F. Austen, and J. A. Boyce. 2001. Thelper cell type 2 cytokines coordinately regulate immunoglobulin E dependentcysteinyl leukotriene production by human cord blood-derived mast cells: pro-found induction of leukotriene C4 synthase expression by interleukin 4. J. Exp.Med. 193: 123–133.
41. Wang, H. W., N. Tedla, A. R. Lloyd, D. Wakefield, and H. P. McNeil. 1998. Mastcell activation and migration to lymph nodes during induction of an immuneresponse in Mice. J. Clin. Invest. 102: 1617–1626.
42. Di Girolamo, N., A. Lloyd, P. J. McCluskey, M. Filipic, and D. Wakefield.1997. Increased expression of matrix metalloproteinases in vivo in scleritistissue and in vitro in cultured human scleral fibroblasts. Am. J. Pathol. 105:653– 666.
43. Di Girolamo, N., M. T. Coroneo, and D. Wakefield. 2003. UVB-elicited induc-tion of MMP-1 expression in human ocular surface epithelial cells is mediatedthrough the ERK1/2 MAPK-dependent pathway. Invest. Ophthalmol. Vis. Sci. 44:4705–4714.
2649The Journal of Immunology
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

44. Coussens, L. M., W. W. Raymond, G. Bergers, M. Laig-Webster,O. Behrendtsen, Z. Werb, G. H. Caughey, and D. Hanahan. 1999. Inflammatorymast cells up-regulate angiogenesis during squamous epithelial carcinogenesis.Genes Dev. 13: 1382–1397.
45. DiMartino, M., C. Wolff, W. High, G. Stroup, S. Hoffman, J. Laydon, J. C. Lee,D. Bertolini, W. A. Gallowawy, M. J. Crimmin, et al. 1997. Anti-arthritic activityof hydroxamic acid-based pseudopeptide inhibitors of matrix metalloproteinasesand TNF� processing. Inflamm. Res. 46: 211–215.
46. Ma, Z., H. Qin, and E. N. Benveniste. 2001. Transcriptional suppression of ma-trix metalloproteinase-9 gene expression by IFN-� and IFN-�: critical role ofSTATS-1�. J. Immunol. 167: 5150–5159.
47. Saren, P., H. G. Welgus, and P. T. Kovanen. 2001. TNF-� and IL-1� selectivelyinduce expression of 92-kDa gelatinase by human macrophages. J. Immunol. 157:4159–4165.
48. Keck, T., J. H. Balcom, 4th, C. Fernandez del Castello, B. A. Antoniu, andA. L. Warshaw. 2002. MMP-9 promotes neutrophil migration and alveolar cap-illary leakage in pancreatitis associated lung injury in the rat. Gastroenterology122: 188–201.
49. Faveeuw, C., G. Preece, and A. Ager. 2001. Transendothelial migration of lym-phocytes across high endothelial venules into lymph nodes is affected by metal-loproteinases. Blood 98: 688–695.
50. Kandere-Grzybowska, K., R. Letourneau, D. Kempuraj, J. Donelan,S. Poplawski, W. Boucher, A. Athanassiou, and T. C. Theoharides. 2003. IL-1induces vesicular secretion of IL-6 without degranulation from human mast cells.J. Immunol. 171: 4830–4836.
51. Coussens, L. M., C. L. Tinkle, D. Hanahan, and Z. Werb. 2000. MMP-9 suppliedby bone marrow-derived cells contributes to skin carcinogenesis. Cell 103:481–490.
52. Fang, K. C., P. J. Wolters, M. Steinhoff, A. Bidgol, J. L. Blount, andG. H. Caughey. 1999. Mast cell expression of gelatinase A and B is regulated bykit ligand and TGF-�. J. Immunol. 162: 5528–5535.
53. Alvaro-Gracia, J. M., N. J. Zvaiffer, and G. S. Firestein. 1990. Cytokines inchronic inflammatory arthritis. V. Mutual antagonism between interferon-�and tumor necrosis factor-� on HLA-DR expression, proliferation, collag-
enases production, and granulocyte macrophage colony-stimulating factorproduction by rheumatoid arthritis synoviocytes. J. Clin. Invest. 86:1790 –1798.
54. Zhou, M., Y. Zhang, J. A. Ardans, and L. M. Wahl. 2003. Interferon-� differ-entially regulates monocyte matrix metalloproteinase-1 and -9 through tumornecrosis factor-� and caspase 8. J. Biol. Chem. 278: 45406–45413.
55. Kumagai, K., I. Ohno, S. Okada, Y. Ohkawara, K. Suzuki, T. Shinya, H. Nagase,K. Iwata, and K. Shirato. 1999. Inhibition of matrix metalloproteinases preventsallergen-induced airway inflammation in a murine model of asthma. J. Immunol.162: 4212–4219.
56. Frank, B. T., J. C. Rossall, G. H. Caughey, and K. C. Fang. 2001. Mast cell tissueinhibitor of metalloproteinase-1 is cleaved and inactivated extracellularly by�-chymase. J. Immunol. 166: 2783–2792.
57. Opdenakker G., P. E. Van den Steen, and J. Van Damme. 2001. Gelatinase B: atuner and amplifier of immune functions. Trends Immunol. 22: 571–579.
58. Dubois, B., S. Masure, U. Hurtenbach, L. Paemen, H. Heremans,J. Van den Oord, R. Sciot, T. Meinhardt, G. Hammerling, G. Opdenakker, andB. Arnold. 1999. Resistance of young gelatinase B-deficient mice to experimentalautoimmune encephalomyelitis and necrotizing tail lesions. J. Clin. Invest. 104:1507–1515.
59. Liu, Z., J. M. Shipley, T. H. Vu, X. Zhou, L. A. Diaz, Z. Werb, and R. M. Senior.1998. Gelatinase B-deficient mice are resistant to experimental bullous pemphi-goid. J. Exp. Med. 188: 475–482.
60. Wang, M., X. Qin, J. S. Mudgett, T. A. Ferguson, R. M. Senior, andH. G. Welgus. 1999. Matrix metalloproteinase deficiencies affect contact hyper-sensitivity: stromelysin-1 deficiency prevents the response and gelatinase B de-ficiency prolongs the response. Proc. Natl. Acad. Sci. USA 96: 6885–6889.
61. Heissig, B., K. Hattori, S. Dias, M. Friedrich, B. Ferris, N. R. Hackett,R. G. Crystal, P. Besmer, D. Lyden, M. A. Moore, et al. 2002. Recruitment ofstem and progenitor cells from the bone marrow niche requires MMP-9 mediatedrelease of kit-ligand. Cell 109: 625–637.
62. Gurish, M. F., H. Tao, J. P. Abonia, A. Arya, D. S. Friend, C. M. Parker, andK. F. Austen. 2001. Intestinal mast cell progenitors require CD49d�7 (�4�7) fortissue-specific homing. J. Exp. Med. 194: 1243–1252.
2650 INDUCTION OF MMP-9 BY TNF-� IN HUMAN MAST CELLS
by guest on May 28, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from




















