
Ultrastructural study shows morphologic features of apoptosis and para-apoptosis in megakaryocytes...
20
CHAPTER VI Ultrastructural study shows morphologic features of apoptosis and para-apoptosis in megakaryocytes from patients with idiopathic thrombocytopenic purpura Ewout J. Houwerzijl 1 , Nel R. Blom 1,2 , Johannes J. L. van der Want 2 , Mariet T. Esselink 1 , Jan J. Koornstra 4 , Jan W. Smit 1 , Henk Louwes 3 , Edo Vellenga 1 , Joost Th. M. de Wolf 1 1 Department of Hematology, University Medical Center Groningen 2 Laboratory for Cell Biology and Electron Microscopy, University of Groningen 3 Department of Nuclear Medicine, Martini Hospital, Groningen 4 Department of Medical Oncology, University Medical Center Groningen, The Netherlands Blood. 2004;103:500-506
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Ultrastructural study shows morphologic features of apoptosis and para-apoptosis in megakaryocytes...

CHAPTER VI
Ultrastructural study shows morphologic features of
apoptosis and para-apoptosis in megakaryocytes from
patients with idiopathic thrombocytopenic purpura
Ewout J. Houwerzijl1, Nel R. Blom
1,2, Johannes J. L. van der Want
2,
Mariet T. Esselink1, Jan J. Koornstra
4, Jan W. Smit
1, Henk Louwes
3,
Edo Vellenga1, Joost Th. M. de Wolf
1
1Department of Hematology, University Medical Center Groningen
2Laboratory for Cell Biology and Electron Microscopy, University of Groningen
3Department of Nuclear Medicine, Martini Hospital, Groningen
4Department of Medical Oncology, University Medical Center Groningen,
The Netherlands
Blood. 2004;103:500-506

CHAPTER VI
100
ABSTRACT
To investigate whether altered megakaryocyte morphology contributes to reduced platelet production in idiopathic thrombocytopenic purpura (ITP), ultrastructural analy-sis of megakaryocytes was performed in 11 ITP patients. Ultrastructural abnormalities
compatible with (para-)apoptosis were present in 78% ± 14% of ITP megakaryocytes, which could be reversed by in vivo treatment with prednisone and intravenous immu-noglobulin. Immunohistochemistry of bone marrow biopsies of ITP patients with ex-tensive apoptosis showed an increased number of megakaryocytes with activated cas-
pase-3 compared with normal (28% ± 4% versus 0%). No difference, however, was observed in the number of bone marrow megakaryocyte colony-forming units (ITP,
118 ± 93/105 bone marrow cells; versus controls, 128 ± 101/105 bone marrow cells; P = 0.7). To demonstrate that circulating antibodies might affect megakaryocytes, suspen-sion cultures of CD34+ cells were performed with ITP or normal plasma. Morphology compatible with (para-)apoptosis could be induced in cultured megakaryocytes with ITP plasma (2 of 10 samples positive for antiplatelet autoantibodies). Finally, the plasma glycocalicin index, a parameter of platelet and megakaryocyte destruction, was
increased in ITP (57 ± 70 versus 0.7 ± 0.2; P = 0.009) and correlated with the propor-tion of megakaryocytes showing (para-)apoptotic ultrastructure (P = 0.02; r = 0.7). In conclusion, most ITP megakaryocytes show ultrastructural features of (para-)apoptosis, probably due to action of factors present in ITP plasma.

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
101
INTRODUCTION
Idiopathic, or immune, thrombocytopenic purpura (ITP) is an autoimmune disease characterized by isolated thrombocytopenia in an otherwise healthy person. The thrombocytopenia in ITP is caused by accelerated platelet destruction due to the action of antiplatelet immunoglobulin G (IgG) autoantibodies that bind to antigens on the platelet cell membrane. The platelets are subsequently destroyed by tissue macro-phages, predominantly in the spleen.1 As a result of the accelerated destruction, platelet survival is usually greatly shortened and platelet production is thought to be compensa-torily increased.2,3 However, there is also evidence that platelet production can be im-paired in ITP. This was demonstrated in platelet kinetic studies using radiolabeled platelets.4-6 The reduced platelet production rate might be mediated by the action of antiplatelet antibodies, which can bind to megakaryocytes in the bone marrow.7-9 Re-cent in vitro studies support this concept showing that human megakaryocyte colony formation and proplatelet formation is inhibited10 and that a reduced expansion of megakaryocytic progenitors can be observed especially in the presence of certain anti-platelet glycoprotein antibodies.11 However, despite the evidence of a reduced platelet production in several ITP patients, numbers of megakaryocytes in the bone marrow are usually normal or increased.6 This is compatible with the finding that plasma throm-bopoietin (TPO) levels in ITP patients are not significantly different from healthy con-trols, indicating that the total megakaryocytic mass has not been changed in ITP. Inves-tigating the relationship between thrombokinetic parameters and the glycocalicin index (GCI), a parameter of platelet destruction,12 we recently demonstrated that there is an inverse correlation between the platelet production rate and the GCI.13 These results suggest that despite the normal number of megakaryocytes in the bone marrow an in-creased destruction of platelets and/or megakaryocytes might occur. These findings support the concept of ineffective thrombopoiesis in the bone marrow. To investigate whether apoptosis or other forms of programmed cell death are responsible for this ineffective thrombopoiesis, we examined the ultrastructure of bone marrow megaka-ryocytes from ITP patients with electron microscopy. The results demonstrate that, independent of the refractoriness of ITP to therapy, in all patients most bone marrow megakaryocytes are extensively damaged, showing ultrastructural abnormalities of apoptosis and para-apoptosis.

CHAPTER VI
102
PATIENTS, MATERIALS AND METHODS
Patients
Eleven adult patients with ITP were investigated. The diagnosis required that an other-wise healthy person, after history, physical examination, complete blood count, and examination of the peripheral blood smear, had an isolated thrombocytopenia (platelets
less than 100 × 109/L) of undetermined etiology. Patients with associated systemic disease were excluded. The University Hospital Groningen’s institutional review board approved the study protocol, and all patients gave informed consent.
Light microscopy of bone marrow megakaryocytes
In all patients bone marrow was aspirated from the sternum or posterior superior iliac spine and stained according to May-Grünwald Giemsa. Normal bone marrow was ob-tained from nonhematologic patients undergoing cardiac surgery after informed con-sent. The number of megakaryocytes was rated as normal (1 megakaryocyte per 1 to 3
low-power fields [16 × objective]), increased (more than 2 megakaryocytes per low-power field), or decreased (1 megakaryocyte per 5 to 10 low-power fields).6 Megaka-
ryocyte morphology was studied with a 100 × objective.
Electron microscopy of bone marrow megakaryocytes
Fresh bone marrow cells were washed in RPMI 1640 (BioWhittaker Europe, Verviers, Belgium), pelleted, and subsequently fixed in 2% glutaraldehyde in 0.1 M phosphate buffer for 24 hours at 4°C. Cells were dehydrated, osmicated, and embedded in Epon 812 according to routine procedures. Semi thin sections (0.5-1 µm) were inspected light microscopically to select megakaryocytes. To examine the ultrastructure in detail and to identify the developmental stage of the morphologically recognizable megakar-yocytes (stage I, II, or III), electron microscopy (Philips 201, Philips, Eindhoven, The Netherlands) was performed. The major criteria for classifying megakaryocytes into different stages are the quality and quantity of cytoplasm and the size, lobulation, and chromatin pattern of the nucleus. The immature, or stage I, megakaryocyte (mega-karyoblast) is characterized by a large round, indented, or bilobed nucleus, prominent nucleoli and cytoplasm containing scattered mitochondria, abundant free ribosomes, variable amounts of rough endoplasmic reticulum (RER), a small Golgi complex, a few α-granules, and rudiments of the demarcation membrane system (DMS). The maturing,

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
103
or stage II, megakaryocyte (promegakaryocyte) contains a lobulated nucleus with gradually condensed chromatin. Comparing with stage I the Golgi complex enlarges, the RER increases, the DMS penetrates the entire cytoplasm, and the number of gra-nules increases. Stage III, or mature, megakaryocytes are very large cells (40-56 µm) in which the nucleus is pushed to one pole of the cell, nucleoli are absent, and granular cytoplasm is abundant. The well-developed DMS divides the cytoplasm into platelet fields. The Golgi complex and RER are greatly reduced.14 A total of 30 megakaryo-cytes per sample were examined.
Ultrastructural characteristics of apoptosis and para-apoptosis
The ultrastructure of megakaryocytes was examined for the presence of apoptotic and para-apoptotic cell death. Ultrastructurally apoptosis is characterized by margination of condensed chromatin, nuclear fragmentation, and the formation of apoptotic bodies.15 Para-apoptosis, nonclassical apoptosis, is a specific morphologic type of non-necrotic cell death16-18 and is characterized by cytoplasmic vacuolization, condensed chromatin (but not early margination of the chromatin), and swollen mitochondria. Characteristics of apoptosis like surface blebbing and the formation of apoptotic bodies do not appear.
Immunohistochemistry
To identify apoptosis in megakaryocytes, immunohistochemical staining was per-formed with an antibody detecting activated caspase-3 as previously reported.19 For immunohistochemical staining, serial 3-µm– thick sections were cut from paraffin-embedded bone marrow biopsies. After deparaffinization in xylene, antigen retrieval was performed using microwave heating at 700 W for 10 minutes in EDTA buffer. Following blocking of endogenous peroxidase with 3% hydrogen peroxide for 30 mi-nutes, the primary antibody was applied for 1 hour at room temperature. To identify activated caspase-3, immunostaining was used with a rabbit polyclonal antibody (1:100; New England Biolabs, Beverly MA). Subsequently, slides were incubated for 30 mi-nutes with appropriate secondary and tertiary antibodies with streptavidin conjugated peroxidase (DAKO, Glostrup, Denmark). Peroxidase activity was visualized with dia-minobenzidine. Slides were counterstained with hematoxylin. As a positive control, a sample of colorectal carcinoma was included. As negative controls, slides were immu-nostained in the absence of the primary antibody. Evaluation of extension of staining was performed by light microscopy. Slides were evaluated by at least 2 independent investigators. If the evaluations did not agree, they were re-evaluated under a multi-headed microscope. Samples were scored as negative (ie, absence of detectable cyto-plasmic staining) or positive. A total of 30 megakaryocytes per sample were examined.

CHAPTER VI
104
Megakaryocyte progenitor cell assay
Megakaryocyte colony-forming units (CFU-Mks) were evaluated quantitatively using a commercially available kit (MegaCult-C; StemCell Technologies, Vancouver, BC,
Canada), according to manufacturer’s instructions. In short, 1 × 105 mononuclear bone marrow cells from ITP patients and healthy controls were seeded per double chamber culture slide in serumfree medium containing thrombopoietin (TPO; 50 ng/mL), inter-leukin-3 (IL-3; 10 ng/mL), IL-6 (10 ng/mL), and collagen (1.1 mg/mL). Cultures were incubated for 12 days, followed by dehydration, fixation, and immunocytochemical staining on slides. Megakaryocyte colonies were detected using antiglycoprotein IIb/IIIa (anti-GPIIb/IIIa) antibody (CD41) and the alkaline phosphatase (APAAP) de-tection system. Cultures were scored for the presence of pure megakaryocytic colonies consisting of at least 5 nucleated cells expressing GPIIb/IIIa without any negatively stained cells (mixed and non-Mk colonies). Colonies were divided into megakaryocyte progenitors with low proliferative capacity, when colonies contained 5 to 20 megaka-ryocytes, and progenitors with high proliferative capacity, when CFU-Mks contained more than 20 megakaryocytes per colony.20
Isolation of CD34+ cells
Peripheral blood cells were obtained from leukapheresis samples of a patient undergo-ing autologous peripheral blood stem cell mobilization after informed consent. Cells were collected by apheresis during the regeneration phase after high-dose cyclophos-phamide in the presence of granulocyte colony-stimulating factor (G-CSF) in patients with multiple myeloma. CD34+ cells were isolated by positive selection using Isolex-300 (Baxter, Deerfield, IL) magnetic cell-sorting system according to manufacturer’s instructions. At the end of the procedure, CD34+ cell purity was reanalyzed by flow cytometry using anti–HPCA-2 and was more than 90%.
Suspension cultures
Purified normal CD34+ cells were seeded at 2 × 105 cells per milliliter in liquid serum-free expansion medium (StemSpan SFEM, Stem Cell Technologies) plus growth fac-tors (TPO 20 ng/mL and stem cell factor [SCF] 10 ng/mL) in 24-well plates (Costar, Cambridge, MA). Cultures were performed for 7 and 12 days in the presence of 10% plasma from ITP patients, 10% plasma from healthy controls, and without plasma. All cultures were performed at 37°C under a 5% CO2 in air–humidified atmosphere. Viable cells were evaluated by trypan blue exclusion, and cell counts were determined using a hemocytometer.

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
105
Antiplatelet antibody detection
A commercially available enzyme-linked immunosorbent assay (ELISA; PakAuto, GTI, Brookfield, WI) was used to detect antibodies reactive with GPIIb/IIIa, GPIb/IX, and GPIa/IIa in plasma of ITP patients according to manufacturer’s instructions. Test re-sults showing absorbance values equal to or greater than twice the value obtained for the mean of the negative controls were regarded as positive.
Plasma glycocalicin
Plasma glycocalicin (GC) concentrations were measured by enzyme immunoassay (EIA; Takara Shuzo, Ohtsu, Japan). Citrate-anticoagulated blood was processed within 2 hours after blood collection. Because GC levels are dependent on the platelet count,
the GC index (GCI) was calculated by using the following formula: (GC [µg/mL] ×
[250 × 109/L]) / individual platelet count (109/L).12,21 Normal value of the GCI is
0.7±0.2.
Statistical analysis
Data are reported as mean ± SD. Differences between groups were calculated using the Mann-Whitney U test. Correlations were calculated using the Spearman rank correla-tion test. P values below 0.05 were considered significant.
RESULTS
Patients
Eight female and 3 male patients with ITP were studied. Mean age was 34 ± 19 years.
The mean platelet count at the time of study was 21 × 109/L ± 30 × 109/L. Eight pa-
tients (73%) had platelet counts below 15 × 109/L (Table 1). During the time of study no patients were on treatment. Four patients had undergone a splenectomy before they were studied.
Light microscopy of bone marrow megakaryocytes
Eight patients (73%) had an increased number and 3 patients a normal number of megakaryocytes in the bone marrow. In smears made from bone marrow aspirates for clinical use no abnormalities were found in megakaryocyte morphology.

CHAPTER VI
106
Table 1. Characteristics of ITP patients.
Antiplatelet GP antibodies
Patient Age, y Sex Plt, × 109/L GCI GPIIb/IIIa GPIa/IIa GPIb/IX
1 37 F 80 3 Negative Negative Negative
2 22 F 2 226 Positive Positive Positive
3 20 F 4 68 Negative Negative Negative
4 19 M 4 84 Negative Negative Negative
5 28 M 9 21 Negative Negative Negative
6 63 F 12 13 Negative Negative Negative
7 13 F 5 54 Negative Negative Negative
8 41 F 10 ND ND ND ND
9 17 F 5 47 Positive Negative Negative
10 47 F 80 3 Negative Negative Negative
11 69 M 20 11 Negative Negative Negative
Normal values: platelet count, 150 × 109/L to 350 × 10
9/L; GCI, 0.7 ± 0.2. Plt indicates platelets; GCI,
glycocalicin index; GP, glycoprotein; F, female; M, male; and ND, not determined.
Ultrastructure and immunohistochemistry of bone marrow megakaryocytes
ITP patients versus healthy controls. Electron microscopic study of bone marrow megakaryocytes of ITP patients and healthy controls demonstrated that the percentage of stage III (mature) megakaryocytes was significantly decreased in ITP patients (n=11)
(48% ± 21%) compared with healthy controls (n = 4) (72% ± 11%; P = 0.05) The per-centage of stage I (immature) megakaryocytes in ITP patients was not significantly
different from controls (12% ± 6% versus 11% ± 7%; P = 0.8), while stage II (maturing) megakaryocytes were significantly increased in ITP patients compared with controls
(40% ± 19% versus 17% ± 5%; P = 0.02). Ultrastructural examination of megakaryocytes in all ITP patients showed extensive
damage in 37% ± 37% of stage I, 65% ± 37% of stage II, and 84% ± 16% of stage III
megakaryocytes (Figure 1). Of all stages together, 78% ± 14% were morphologically abnormal. The morphologic alterations consisted primarily of cytoplasmic vacuoliza-tion due to mitochondrial swelling and distended DMS, and chromatin condensation without margination, all ultrastructural features of para-apoptosis. Megakaryocytes showing these abnormalities all had an intact, mostly thickened, peripheral zone, which did not seem to contain any functional cellular material (ie, organelles or DMS).

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
107
Figure 1. Percentage of damaged ITP megakaryocytes in different stages of differentiation. Signifi-
cantly more stage III (mature) megakaryocytes than stage I (immature) megakaryocytes showed the
characteristic ultrastructural features of (para-)apoptosis. Data presented as mean ± SD.
Continuity between the extracellular space and the DMS was only very rarely observed. The cytoplasm of these damaged cells lacked platelet territories. Practically all abnor-mal but none of the normal ITP megakaryocytes were surrounded by neutrophils or macrophages, some being in a state of phagocytosis (Figure 2B).
Most ultrastructural alterations in damaged ITP megakaryocytes consisted of cha-racteristics of para-apoptosis. Morphologic features of apoptosis were observed in 4 of 11 ITP patients and only in stage III megakaryocytes (Figure 3B). Of stage III megaka-
ryocytes, 62% ± 31% showed ultrastructural features of apoptosis, 31% ± 31% of para-
apoptosis, while 7% ± 8% were morphologically normal. In these 4 patients, all stage I and most stage II megakaryocytes were morphologically intact (15% to 20% of stage II megakaryocytes showing features of para-apoptosis).
In more than 90% of the abnormal megakaryocytes all ultrastructural characteristics of apoptosis and para-apoptosis were found. The amount and type of alterations in the megakaryocytes were not correlated with the platelet count.
To confirm the process of apoptosis, immunohistochemical staining for activated caspase-3 was performed on bone marrow biopsies from ITP patients (n = 2, nos. 6 and 10; Table 1) whose megakaryocytes showed ultrastructural features of apoptosis. A total of 25% and 30% of the megakaryocytes of these patients were positive for acti-vated caspase-3, while megakaryocytes of bone marrow biopsies from healthy controls
0
20
40
60
80
100
Immature Maturing Mature
p=0.0008
% (
pa
ra)
ap
oto
tic
pre
cu
rso
rs
(pa
ra)a
po
pto
tic
me
ga
ka
ryo
cyte
s (%
)
Immature Maturing Mature
P = 0.0008

CHAPTER VI
108
(n = 4) were negative (Figure 4).
ITP patients before and after treatment. In 2 patients the effects of prednisone or high-dose immunoglobulin could be analyzed. In the first patient the bone marrow aspiration was repeated 2 weeks after start of prednisone treatment. In this period the
platelet count had increased to 120 × 109/L. Ultrastructural examination of this bone marrow sample demonstrated a distinct reduction in the proportion of megakaryocytes showing programmed cell death (in stage I/II from 100% to 55%, in stage III megakar-yocytes from 100% to 61%). The percentage of stage III megakaryocytes increased from 20% to 26%. In the second patient bone marrow examination was repeated after high-dose intravenous immunoglobulin therapy, which increased the platelet count
from 20 to 160 × 109/L. The percentage of stage III megakaryocytes increased from 20% to 43%, which coincided with a diminished number of damaged megakaryocytes (stage I/II: from 80% to 35%; stage III from 50% to 23%).
Quantification of megakaryocyte progenitor cells
In view of the observed abnormalities in megakaryocytes, we were interested in whe-ther this was also reflected at progenitor level — that is, a reduced number of megakar-yocyte progenitors (CFU-Mks). The total number of CFU-Mks per 105 bone marrow
cells in ITP was not significantly different from healthy controls (n = 10) (118 ± 93
versus 128 ± 101; P = 0.7). In addition, there was no significant difference in the num-ber of progenitors with low and high proliferative capacity between ITP and normal control bone marrow (P = 0.7 and 0.9, respectively).
Megakaryocytes derived from CD34+ cells after incubation with normal and ITP
plasma
To investigate whether normal megakaryopoiesis can be influenced by factors in ITP plasma, normal CD34+ cells were incubated with 10% plasma from 3 ITP patients (nos. 1, 2, and 10; Table 1) and 3 healthy controls and cultured for 7 and 12 days in serum-free medium containing TPO and SCF. After 7 days of incubation with ITP or normal plasma, electron microscopy of the suspension cultures showed only immature mega-karyocytes without abnormalities. After 12 days of culture, similar cytoplasmic and
nuclear alterations as observed in the ITP megakaryocytes were seen in 60% ± 14% of
the megakaryocytes derived from incubation with ITP plasma (42% ± 23% with apop-
totic and 18% ± 9% with para-apoptotic ultrastructure). Ninety percent of megakaryo-cytes incubated with normal plasma were morphologically normal, and ultrastructural features of para-apoptosis were not observed.

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
109
Figure 2. Ultrastructure of normal and ITP
megakaryocytes.
A. Normal megakaryoblast (stage I mega-
karyocyte) showing a lobulated nucleus
(N). In the cytoplasm, characteristic de-
marcation membrane system (asterisks)
and normal mitochondria (arrowheads)
can be found. Original magnification, x
3000.
B. Megakaryoblast of an ITP patient in the
process of para-apoptosis; N indicates
nucleus. The cell has an intact enlarged
peripheral margin (large arrow). Original
magnification, x 4500.
C. This inset of panel B shows a higher
magnification with mitochondria, some of
which are slightly swollen (open arrow-
heads) and others which have completely
collapsed cristae, which appear as empty
vacuoles (asterisks). The enlarged peri-
pheral margin in the cytoplasm is free of
organelles (open arrow). Original magnifi-
cation, x 15 000.
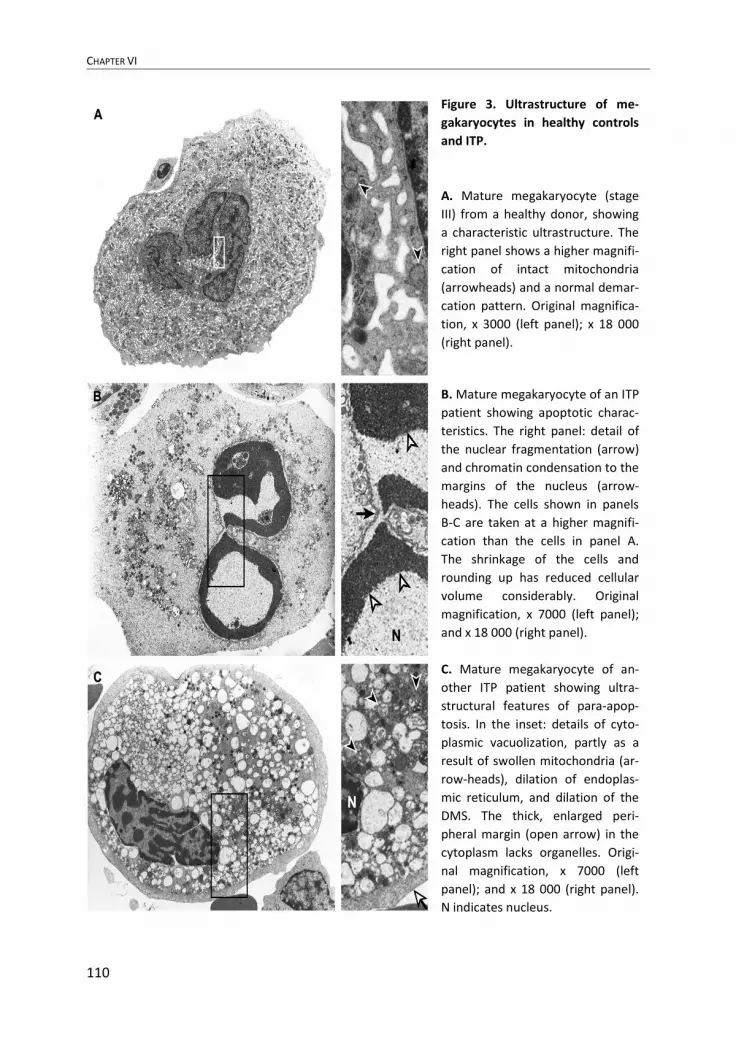
CHAPTER VI
110
Figure 3. Ultrastructure of me-
gakaryocytes in healthy controls
and ITP.
A. Mature megakaryocyte (stage
III) from a healthy donor, showing
a characteristic ultrastructure. The
right panel shows a higher magnifi-
cation of intact mitochondria
(arrowheads) and a normal demar-
cation pattern. Original magnifica-
tion, x 3000 (left panel); x 18 000
(right panel).
B. Mature megakaryocyte of an ITP
patient showing apoptotic charac-
teristics. The right panel: detail of
the nuclear fragmentation (arrow)
and chromatin condensation to the
margins of the nucleus (arrow-
heads). The cells shown in panels
B-C are taken at a higher magnifi-
cation than the cells in panel A.
The shrinkage of the cells and
rounding up has reduced cellular
volume considerably. Original
magnification, x 7000 (left panel);
and x 18 000 (right panel).
C. Mature megakaryocyte of an-
other ITP patient showing ultra-
structural features of para-apop-
tosis. In the inset: details of cyto-
plasmic vacuolization, partly as a
result of swollen mitochondria (ar-
row-heads), dilation of endoplas-
mic reticulum, and dilation of the
DMS. The thick, enlarged peri-
pheral margin (open arrow) in the
cytoplasm lacks organelles. Origi-
nal magnification, x 7000 (left
panel); and x 18 000 (right panel).
N indicates nucleus.

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
111
Antiplatelet antibodies
Plasma from 2 of 10 patients tested positive for the presence of antiplatelet antibodies (Table 1). Plasma used for incubation studies was derived from patient nos. 1, 2, and 10. The plasma from patient no. 2 tested positive for antibodies reactive with GPIIb/IIIa, GPIb/IX, and GPIa/IIa. In the plasma of the other 2 patients, test results for antibodies were negative.
Plasma GCI
GCI in ITP patients (n = 10) was significantly increased compared with healthy con-
trols (n = 16) (57 ± 70 versus 0.7 ± 0.2; P = 0.009) (Table 1). There was a significant correlation between the GCI and the proportion of ITP megakaryocytes with mor-phologic features of (para-)apoptosis (P = 0.02; r = 0.7) (Figure 5).
Figure 4. Immunohistochemical detection of activated caspase-3 in ITP megakaryocytes.
A. Normal bone marrow megakaryocytes are negative for activated caspase-3. Some neutrophil
granulocytes are positive (arrows). B and C. Megakaryocytes of 2 ITP patients with extensive ultra-
structural features of apoptosis are shown. These megakaryocytes stain positive for activated cas-
pase-3. Original magnification, x 400.

CHAPTER VI
112
Figure 5. Correlation between percentage of damaged megakaryocytes and glycocalicin index
(GCI). The GCI is significantly correlated with the proportion of megakaryocytes showing morphol-
ogic features of (para-)apoptosis (P = 0.02; r = 0.7).
DISCUSSION
In the present study extensive ultrastructural alterations in megakaryocytes from ITP patients are described. Previous studies on megakaryocyte morphology and ultrastruc-ture in ITP show conflicting results. Earlier light microscopic observations showed megakaryocytes with degenerative characteristics and greatly reduced platelet produc-tion, although the number of megakaryocytes was increased in ITP patients compared with healthy controls.22 Ultrastructural studies of megakaryocytes in ITP showed re-sults varying from moderate vacuolization, deficient formation of demarcation mem-brane system (DMS), and invaginations in the peripheral membrane23 to practically normal morphology of megakaryocytes.24,25 Stahl et al26 found that 50% to 75% of the ultrastructurally examined megakaryocytes from 4 ITP patients had extensive damage,
0 50 100 150 200 250
0
20
40
60
80
100
Glycocalicin index
% (
pa
ra)a
po
pto
tic
pre
cu
rso
rs
Glycocalicin index
(pa
ra)a
po
pto
tic
me
ga
ka
ryo
cyte
s (%
)

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
113
with marked dilation of the DMS and a disrupted peripheral zone. ITP megakaryocytes with an intact peripheral zone did not show any morphologic alterations. The present investigation, however, reveals ITP megakaryocytes with extensive abnormalities within an intact and often thickened peripheral zone. These cells are all surrounded by neutrophils and macrophages, suggesting an inflammatory response against these megakaryocytes.
The ultrastructural alterations observed in the present study are compatible with the morphologic criteria for apoptosis and para-apoptosis. Para-apoptosis, first described by Asher et al,16 is characterized by condensed chromatin and cytoplasmic vacuoliza-tion due to swollen mitochondria and endoplasmic reticulum. In contrast to necrosis, there is no cell membrane disruption. Recently, Sperandio et al defined a form of para-apoptosis (which they called paraptosis) with similar ultrastructure by cellular charac-teristics and response to apoptosis inhibitors.18 They found that this form of nonapop-totic cell death is TUNEL (terminal deoxynucleotidyl transferase-mediated nick-end labeling)–negative and resistant to caspase inhibitors and Bcl-xL and is driven by an alternative caspase-9 activity that is APAF-1 independent.
In our study about 80% of mature (stage III) megakaryocytes, in which most of the platelet production occurs, showed ultrastructural features of apoptosis and para-apoptosis. Significantly fewer stage I megakaryocytes showed morphologic alterations, indicating that immature megakaryocytes are less affected by (para-)apoptotic cell death. This is further supported by the finding that the total number of CFU-Mks in our ITP patients was not significantly different from normal. Previous studies on the num-ber of CFU-Mks in chronic ITP showed conflicting results, depending on the assay that was used, the stage (acute or chronic) of the disease, and the refractoriness of the ITP to therapy. Several authors27,28 found a significant increase in number of CFU-Mks in chronic ITP patients. Others29,30 reported a reduced number of CFU-Mks in chronic ITP. The present group of ITP patients was heterogeneous, consisting of patients re-sponding to prednisone and splenectomy and patients with recurrent disease after sple-nectomy.
Morphologic changes compatible with (para-)apoptosis similar to those found in megakaryocytes from ITP patients could be induced in normal CD34+ cells that were cultured to megakaryocytes in the presence of ITP plasma. The data from these in-cubation studies are consistent with earlier phase-contrast microscopic studies of ITP megakaryocytes in which cytoplasmic vacuolization, loss of granularity, and lack of platelet release were observed. In these studies the morphologic alterations in the megakaryocytes could be reproduced in vivo by infusing healthy controls with ITP plasma.31 The results suggest that the defect in ITP is not only at the level of platelets

CHAPTER VI
114
but also at the level of megakaryocytes. This is further supported by the finding that during successful treatment with prednisone the number of damaged megakaryocytes decreases. Surprisingly, this was also the case in an ITP patient treated with intra-venous immunoglobulin, indicating that this treatment not only protects peripheral blood platelets from destruction by macrophages but might also protect megaka-ryocytes against destruction by bone marrow macrophages. A possible explanation for this finding could be a reduced concentration of antiplatelet antibodies, because recent observations in a rat model of immune thrombocytopenia showed that intravenous immunoglobulin administration can lead to increased clearance of an antiplatelet anti-body.32 Because antiplatelet antibodies bind to antigens present on the surface of megakaryocytes,8 it is likely that this interaction plays an important role in initiating the cascade of programmed cell death. However, in only 20% of ITP patients’ plasma were antiplatelet antibodies detected. This may in part be due to the limited sensitivity of the ELISA test for detecting antiplatelet autoantibodies. Previous studies, however, have also demonstrated a low frequency of free circulating autoantibodies,33 suggesting that alternative mechanisms for initiating cell death are of importance, including the release of cytokines.34
The presence of extensive ultrastructural abnormalities in megakaryocytes indicates that ITP is essentially different from other autoimmune cytopenias such as autoimmune hemolytic anemia (AIHA). In contrast to AIHA, which presents in practically all cases with clear erythroid hyperplasia,35,36 bone marrow examination in ITP shows normal numbers of megakaryocytes in a large number of patients,37 suggesting that ITP mega-karyocytes might be affected contrary to erythroid precursors in AIHA. Only a small proportion of AIHA patients has reticulocytopenia, which is associated with an in-creased apoptosis of erythroid progenitors in the bone marrow.38
In a recent study6 megakaryocyte levels were normal in 65% of 141 ITP patients and increased in only 33%. Twenty-four percent had an increased platelet production rate (PPR; ie, the number of platelets entering the circulation), and only 8% had an increased PPR in conjunction with an increased number of megakaryocytes. Moreover, there are many reports about a reduced PPR in a subgroup (30% to 50%) of ITP pa-tients.4,5,6,39,40 Experiments in which thrombokinetic studies were repeated after sple-nectomy and prednisone treatment showed that both forms of treatment can induce a distinct increase in PPR,41 indicating that the release of platelets to the circulation (ie, PPR) in active ITP is relatively depressed, perhaps due to the action of autoantibodies directed against megakaryocytes.
Besides the results of reduced platelet production in platelet kinetic studies, the pre-sent study gives additional evidence for reduced platelet production by injured mega-

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
115
karyocytes. In a recent study13 we observed a significant inverse correlation between the PPR and the glycocalicin index (GCI) in ITP patients. The GCI is used as a pa-rameter of platelet destruction. A low PPR is associated with an elevated GCI. These findings reflect an increased release of GPIb complex in the circulation, as a result of shedding of the receptor complex from megakaryocytes and/or platelets destructed in the bone marrow. In the present data a significant correlation was observed between the percentage of megakaryocytes in programmed cell death and the GCI, underscoring the view that the low PPR in ITP might be due to an increase in (para-)apoptosis of plate-let-producing megakaryocytes.
In summary, the present study demonstrates that in ITP patients a large majority of morphologically recognizable megakaryocytes show ultrastructural alterations com-patible with (para-)apoptosis, which can be mimicked by incubating normal dif-ferentiated CD34+ cells with ITP plasma.
ACKNOWLEDGEMENTS
We are grateful to Nynke Zwart, Department of Pathology, University Hospital Gron-ingen, The Netherlands, for performing immunohistochemistry. We thank Drs Vrugt, pathologist (Martini Hospital), and Rosati, pathologist (University Hospital Groningen) for providing bone marrow samples and for critically reviewing the bone marrow slides.

CHAPTER VI
116
References
1. Cines DB, Blanchette VS. Immune thrombocytopenic purpura. N Engl J Med.
2002;346:995-1008. 2. Harker LA. Thrombokinetics in idiopathic thrombocytopenic purpura. Br J Haematol.
1970;19:95-104. 3. Branehög I, Kutti J, Ridell B, Swolin B, Weinfeld A. The relation of thrombokinetics
to bone marrow megakaryocytes in idiopathic thrombocytopenic purpura (ITP). Blood. 1975;45:551-562.
4. Grossi A, Vannucchi AM, Casprini P, et al. Different patterns of platelet turnover in chronic idiopathic thrombocytopenic purpura. Scand J Haematol. 1983;31:206-214.
5. Ballem PJ, Segal GM, Stratton JR, Gernsheimer T, Adamson JW, Slichter SJ. Mecha-nisms of thrombocytopenia in chronic autoimmune thrombocytopenic purpura. J Clin Invest. 1987;80:33-40.
6. Louwes H, Zeinali Lathori OA, Vellenga E, de Wolf JThM. Platelet kinetic studies in patients with idiopathic thrombocytopenic purpura. Am J Med. 1999;106:430-434.
7. McKenna JL, Pisciotta AV. Fluorescence of megakaryocytes in idiopathic thrombocy-topenic purpura (ITP) stained with fluorescent antiglobulin serum. Blood. 1962;19:664-675.
8. McMillan R, Luiken GA, Levy R, Yelenosky R, Longmire RL. Antibody against megakaryocytes in idiopathic thrombocytopenic purpura. JAMA. 1978;239:2460-2462.
9. Hoffman R, Zaknoen S, Tang HH, et al. An antibody cytotoxic to megakaryocyte pro-genitor cells in a patient with immune thrombocytopenic purpura. N Engl J Med. 1985;312:1170-1174.
10. Takahashi R, Sekine N, Nakatake T. Influence of monoclonal antiplatelet glycoprotein antibodies on in vitro human megakaryocyte colony formation and proplatelet forma-tion. Blood. 1999;93: 1951-1958.
11. Chang M, Nakagawa PA, Williams SA, et al. Immune thrombocytopenic purpura (ITP) plasma and purified ITP monoclonal autoantibodies inhibit megakaryocytopoiesis in vitro. Blood. 2003;102: 887-895.
12. Steinberg MH, Kelton JG, Coller BS. Plasma glycocalicin. N Engl J Med. 1987; 317:1037-1042.
13. Houwerzijl EJ, Louwes H, Smit JW, Esselink MT, Vellenga E, de Wolf JTM. Glyco-calicin-index is elevated in ITP patients with a decreased platelet production rate in the platelet kinetic studies [abstract]. Blood. 2001;98:525a.
14. Bentfeld-Barker ME, Bainton DF. Ultrastructure of rat megakaryocytes after prolonged thrombocytopenia. J Ultrastruct Res. 1977;61:201-214.
15. Kerr JFR, Winterford CM, Harmon BV. Apoptosis: its significance in cancer and can-cer therapy. Cancer. 1994;73:2013-2026.

APOPTOSIS AND PARA-APOPTOSIS OF MEGAKARYOCYTES FROM ITP PATIENTS
117
16. Asher E, Payne CM, Bernstein C. Evaluation of cell death in EBV-transformed lym-phocytes using agarose gel electrophoresis, light microscopy and electron microscopy. II. Induction of non-classic apoptosis (“para-apoptosis”) by tritiated thymidine. Leuk Lymphoma. 1995;19:107-119.
17. Tome ME, Baker AF, Powis G, Payne CM, Briehl MM. Catalase-overexpressing thy-mocytes are resistant to glucocorticoid-induced apoptosis and exhibit increased net tu-mor growth. Cancer Res. 2001;61:2766-2773.
18. Sperandio S, de Belle I, Bredesen DE. An alternative, nonapoptotic form of pro-grammed cell death. Proc Natl Acad Sci U S A. 2000;97:14376-14381.
19. Grossmann J, Walther K, Artinger M, Rümmele P, Woenckhaus M, Schölmerich J. Induction of apoptosis before shedding of human intestinal epithelial cells. Am J Gas-troenterol. 2002;97:1421-1428.
20. Drayer AL, Sibinga CT, Blom NR, De Wolf JT, Vellenga E. The in vitro effects of cytokines and migration of megakaryocyte progenitor. Br J Haematol. 2000;109:776-784.
21. Beer JH, Büchi L, Steiner B. Glycocalicin: a new assay—the normal plasma levels and its potential usefulness in selected diseases. Blood. 1994;83:691-702.
22. Dameshek W, Miller EB. The megakaryocytes in idiopathic thrombocytopenic purpura, a form of hypersplenism. Blood. 1946;1:27-50.
23. Falcão L, Caen J, Gautier A. Etude évolutive de l’ultrastructure des mégakaryocytes dans un cas de purpura thrombopénique avant, 12 jours et 5 mois après splénectomie. Blut. 1969;19:156-171.
24. Ridell B, Branehög I. The ultrastructure of the megakaryocytes in idiopathic thrombo-cytopenic purpura (ITP) in relation to thrombokinetics. Pathol Eur. 1976;11;179-187.
25. Lambertenghi-Deliliers G, Polli N, Pozzoli E, Nava M, Soligo D. Ultrastructural altera-tions of megakaryocytes in some hematologic disorders. Tumori. 1981;67:151-157.
26. Stahl CP, Zucker-Franklin D, McDonald TP. Incomplete antigen cross-reactivity be-tween platelets and megakaryocytes: relevance to ITP. Blood. 1986;67:421-428.
27. Sugiyama H, Yagita M, Takahashi T, et al. Megakaryocytopoiesis in idiopathic throm-bocytopenic purpura. Nippon Ketsueki Gakkai Zasshi. 1987;50:119-128.
28. Wang W, Matsuo T, Yoshida S, et al. Colony forming unit-megakaryocyte (CFU-meg) numbers and serum thrombopoietin concentrations in thrombocytopenic disorders: an inverse correlation in myelodysplastic syndromes. Leukemia. 2000;14:1751-1756.
29. Abgrall JF, El-Kassar N, Berthou C, et al. In vitro megakaryocyte colony formation in patients with idiopathic thrombocytopenic purpura: differences between acute and chronic ITP. Int J Cell Cloning. 1992;10:28-32.
30. Abgrall JF, Berthou C, Sensebé L, Le Niger C, Escoffre M. Decreased in vitro mega-karyocyte colony formation in chronic idiopathic thrombocytopenic purpura. Br J Haematol. 1993;85:803-804.

CHAPTER VI
118
31. Pisciotta AV, Stefanini M, Dameshek W. Studies on platelets. X. Morphologic charac-teristics of megakaryocytes by phase contrast microscopy in normals and in patients with idiopathic thrombocytopenic purpura. Blood. 1953;8:703-723.
32. Hansen RJ, Balthasar JP. Effects of intravenous immunoglobulin on platelet count and antiplatelet antibody disposition in a rat model of immune thrombocytopenia. Blood. 2002;100:2087-2093.
33. McMillan R. Autoantibodies and autoantigens in chronic immune thrombocytopenic purpura. Semin Hematol. 2000;37:239-248.
34. Semple JW, Lazarus AH, Freedman J. The cellular immunology associated with auto-immune thrombocytopenic purpura: an update. Transfus Sci. 1998;19:245-251.
35. Liesveld JL, Rowe JM, Lichtman MA. Variability of the erythropoietic response in autoimmune hemolytic anemia: analysis of 109 cases. Blood. 1987;69:820-826.
36. Stefanelli M, Barosi G, Cazzola M, Orlandi E. Quantitative assessment of erythropoi-esis in haemolytic disease. Br J Haematol. 1980;45:297-308.
37. Diggs LW, Hewlett JB. A study of the bone marrow from thirty-six patients with idio-pathic hemorrhagic (thrombopenic) purpura. Blood. 1948;3:1090-1104.
38. Van de Loosdrecht AA, Hendriks DW, Blom NR, Smit JW, De Wolf JT, Vellenga E. Excessive apoptosis of bone marrow erythroblasts in a patient with autoimmune haemolytic anaemia with reticulocytopenia. Br J Haematol. 2000;108:313-315.
39. Heyns Adu P, Badenhorst PN, Lotter MG, Pieters H, Wessels P, Kotze HF. Platelet turnover and kinetics in immune thrombocytopenic purpura: results with autologous 111In-labeled platelets and homologous 51Cr-labeled platelets differ. Blood. 1986;67:86-92.
40. Siegel RS, Rae JL, Barth S, et al. Platelet survival and turnover: important factors in predicting response to splenectomy in immune thrombocytopenic purpura. Am J He-matol. 1989;30:206-212.
41. Louwes H, Vellenga E, Houwerzijl EJ, de Wolf JT. Effects of prednisone and splenec-tomy in patients with idiopathic thrombocytopenic purpura: only splenectomy induces a complete remission. Ann Hematol. 2001;80:728-732




















