
Programmed Cell Death (Apoptosis) in Cord Blood Lymphocytes
11
Journal of Clinical Immunology, Vol. 17, No. 1, 1997 Programmed Cell Death (Apoptosis) in Cord Blood Lymphocytes SUDEEPTA AGGARWAL,1 ABHA GUPTA,2 SHIGEKAZU NAGATA,3 and SUDHIR GUPTA1' 4 Accepted: October 17, 1996 Cord blood lymphocytes are functionally immature and have deficient immune responses. In order to determine whether the process of programmed cell death is distinct between cord blood and peripheral blood lymphocytes, we analyzed the expression of fas and box (apoptosis promoting genes) and bcl-2 and bcl-xL (apoptosis inhibiting genes) at protein or mRNA- levels using flow cytometry and quantitative PCR methods, respectively. The susceptibility of T cell subsets from cord blood and adult peripheral blood to undergo apoptosis following restimulation with anti-CD3 or anti-Fas monoclonal antibodies was also studied. We observed that cord blood T cell subsets expressed lower levels of Fas and Bcl-2, a low bcl-2/bax ratio, and higher bcl-xL compared to peripheral blood. Additionally, upon primary stimulation with anti-CD3, cord blood T cell subsets were more resistant to apoptosis compared to peripheral blood. In contrast, rechallenge of previously stimulated lymphocytes with anti-CD3 monoclonal antibody triggered apoptosis in a larger proportion of T cells from cord blood as compared to peripheral blood, whereas the number of T cells undergoing anti-Fas-induced programmed cell death were lower in cord blood compared to peripheral blood. KEY WORDS: Cord blood; apoptosis; Fas; Bcl-2. INTRODUCTION The immune system of neonates in humans and animals is functionally immature (reviewed in Refs. 1-5). There is a decreased lymphocyte proliferation response to T cell-dependent antigens, an increased susceptibility to induction of tolerance, an increased susceptibility to the immunosuppressive effects of cyclosporin A and glu- cocorticoids, and a decreased capacity of T cells to induce a graft-vs-host reaction. The control of apoptosis in lymphocytes is mediated by various gene products which mediate cell death or extend cell survival (6-11). The Fas and Fas ligand (FasL) are members of tumor necrosis factor receptor and tumor necrosis factor families, respectively, and trigger apoptosis upon cross-linking of the receptor in Fas+ apoptosis-sensitive cells (12). Bcl-2 and its family of homologues have differential regulatory effects on apoptosis in lymphoid cells (reviewed in Ref. 13). Overexpression of Bcl-2 and Bcl-xL protects cells from apoptosis (14, 15), whereas Bax promotes it (16). Activation through the T cell receptor (TCR) results in the elimination of a proportion of responding cells by a process known as activation-induced cell death (AICD) and Fas and FasL appear to be regulators of this process (17). In this study, we compared the expression of genes critical in the apoptotic process between human cord blood lymphocytes and peripheral lymphocytes from young adults. In addition, the ability of these lympho- cytes to survive upon TCR rechallenge with anti-CD3 and anti-Fas monoclonal antibodies was compared. MATERIALS AND METHODS Subjects Peripheral blood was obtained from healthy young volunteers. Informed consent was obtained from each subject. Cord blood was obtained from normal deliver- ies. Antibodies Anti-Fas (anti-CD95) monoclonal antibody (CH-11) for the induction of apoptosis (12, 18,19) was purchased from UBI, Lake Placid, NY. FITC-labeled anti-CD95 1 Basic and Clinical Immunology, University of California, Irvine, Irvine, California. 2 Western Medical Center, Santa Ana, California. 3 Osaka Bioscience Institute, Osaka, Japan. 4 To whom correspondence should be addressed at C-240, Basic and Clinical Immunology, Medical Sciences 1, University of California— Irvine, Irvine, California 92697. 63 0271-9142/97/0100-0063$12.50/0 © 1997 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Programmed Cell Death (Apoptosis) in Cord Blood Lymphocytes

Journal of Clinical Immunology, Vol. 17, No. 1, 1997
Programmed Cell Death (Apoptosis) in Cord BloodLymphocytes
SUDEEPTA AGGARWAL,1 ABHA GUPTA,2 SHIGEKAZU NAGATA,3 and SUDHIR GUPTA1'4
Accepted: October 17, 1996
Cord blood lymphocytes are functionally immature and havedeficient immune responses. In order to determine whether theprocess of programmed cell death is distinct between cordblood and peripheral blood lymphocytes, we analyzed theexpression of fas and box (apoptosis promoting genes) andbcl-2 and bcl-xL (apoptosis inhibiting genes) at protein ormRNA- levels using flow cytometry and quantitative PCRmethods, respectively. The susceptibility of T cell subsets fromcord blood and adult peripheral blood to undergo apoptosisfollowing restimulation with anti-CD3 or anti-Fas monoclonalantibodies was also studied. We observed that cord blood T cellsubsets expressed lower levels of Fas and Bcl-2, a lowbcl-2/bax ratio, and higher bcl-xL compared to peripheralblood. Additionally, upon primary stimulation with anti-CD3,cord blood T cell subsets were more resistant to apoptosiscompared to peripheral blood. In contrast, rechallenge ofpreviously stimulated lymphocytes with anti-CD3 monoclonalantibody triggered apoptosis in a larger proportion of T cellsfrom cord blood as compared to peripheral blood, whereas thenumber of T cells undergoing anti-Fas-induced programmedcell death were lower in cord blood compared to peripheralblood.
KEY WORDS: Cord blood; apoptosis; Fas; Bcl-2.
INTRODUCTION
The immune system of neonates in humans and animalsis functionally immature (reviewed in Refs. 1-5). Thereis a decreased lymphocyte proliferation response to Tcell-dependent antigens, an increased susceptibility toinduction of tolerance, an increased susceptibility to theimmunosuppressive effects of cyclosporin A and glu-
cocorticoids, and a decreased capacity of T cells toinduce a graft-vs-host reaction.
The control of apoptosis in lymphocytes is mediatedby various gene products which mediate cell death orextend cell survival (6-11). The Fas and Fas ligand(FasL) are members of tumor necrosis factor receptorand tumor necrosis factor families, respectively, andtrigger apoptosis upon cross-linking of the receptor inFas+ apoptosis-sensitive cells (12). Bcl-2 and its familyof homologues have differential regulatory effects onapoptosis in lymphoid cells (reviewed in Ref. 13).Overexpression of Bcl-2 and Bcl-xL protects cells fromapoptosis (14, 15), whereas Bax promotes it (16).
Activation through the T cell receptor (TCR) results inthe elimination of a proportion of responding cells by aprocess known as activation-induced cell death (AICD)and Fas and FasL appear to be regulators of this process(17). In this study, we compared the expression of genescritical in the apoptotic process between human cordblood lymphocytes and peripheral lymphocytes fromyoung adults. In addition, the ability of these lympho-cytes to survive upon TCR rechallenge with anti-CD3and anti-Fas monoclonal antibodies was compared.
MATERIALS AND METHODS
Subjects
Peripheral blood was obtained from healthy youngvolunteers. Informed consent was obtained from eachsubject. Cord blood was obtained from normal deliver-ies.
Antibodies
Anti-Fas (anti-CD95) monoclonal antibody (CH-11)for the induction of apoptosis (12, 18,19) was purchasedfrom UBI, Lake Placid, NY. FITC-labeled anti-CD95
1 Basic and Clinical Immunology, University of California, Irvine,Irvine, California.
2 Western Medical Center, Santa Ana, California.3 Osaka Bioscience Institute, Osaka, Japan.4 To whom correspondence should be addressed at C-240, Basic and
Clinical Immunology, Medical Sciences 1, University of California—Irvine, Irvine, California 92697.
630271-9142/97/0100-0063$12.50/0 © 1997 Plenum Publishing Corporation

64 AGGARWAL, GUPTA, NAGATA, AND GUPTA
Fig. 1. Fas expression in T cell subsets in cord blood and peripheralblood. Mononuclear cells from 10 cord blood and peripheral bloodsamples each were stained with FITC-anti-Fas and PE-anti-CD4 orPE-anti-CD8 mAbs. Lower proportions of cord blood (O) CD4+ andCD8+ T cells expressed Fas compared to peripheral blood (O). Filledsymbols and vertical lines represent means ± SD from 10 experiments•done in pairs. Lower fas mRNA expression in cord blood (CB)compared to peripheral blood (PB) lymphocytes. Results as cpm ± SDratios fas:f3-actin from quantitative PCR using 5 ;ul cDNA are shownin the inset.
monoclonal antibody was purchased from Kamiya Bio-chemicals, Thousand Oaks, CA. FITC-labeled anti-Bcl-2monoclonal antibody was obtained from Dako Corpora-tion, Carpentaria, CA. Per-CP-labeled anti-CD4 andanti-CD8 and PE-labeled anti-CD45RA and anti-CD45RO monoclonal antibodies were purchased fromBecton Dickinson, San Jose, CA. Anti-CD3 monoclonalantibody (OKT3) was obtained from Ortho Diagnostics,Raritan, NJ.
Primers
Primers used for RT-PCR were synthesized fromGibco BRL, Gaithersberg, MD.
fas: Sense, 5'-ATG CTG GGC ATC TGG ACC CTCCTA-3'; antisense, 5'-TCT GCA CTT GGT ATT CTGGGT CCG-3' (20); bcl-2: sense, 5'-CGA CGA CTT
CTC CCG CCG CTA CCG C-3'; antisense, 5'-CCGCAT GCT GGG GCC GTA CAG TTC C-3' (21); bcl-xL:sense, 5'-TTG GAC AAT GGA CTG GTG GA-3';antisense, 5'-GTA GAG TGG ATG GTC AGT G-3'(22). bax: sense, 5'-ATG GAC GGG TCC GGG GAGCAG CCC-3'; antisense, 5'-GGT GAG CAC TCC CGCCAC AAA GAT-3' (23).
Reagents
Propidium iodide, PHA, PMA, ionomycin, and acri-dine orange were purchased from Sigma Chemicals, St.Louis, MO. [a-32P]dATP was obtained from DuPont,Wilmington, DE. Metalloprotease inhibitors TIMP1 andTIMP2 were purchased from Calbiochem, San Diego,CA. Recombinant IL-2 was obtained from R&D Sys-tems, Minneapolis, MN.
Expression of Fas Ag and Bcl-2 Protein in T CellSubsets
Mononuclear cells (MNC) were washed twice withphosphate-buffered saline (PBS) and stained with FITC-conjugated anti-Fas monoclonal antibody and PE-conjugated anti-CD4 or anti-CD8 monoclonal antibodiesfor 45 min on ice. For Bcl-2 staining, the cells werewashed and fixed with 1% paraformaldehyde for 15 minat room temperature. Cells were washed again andpermeabilized with 70% methanol for 45 min on ice.After washing, cells were incubated with FITC-labeledanti-Bcl-2 and PE-labeled anti-CD4 or anti-CD8 mono-clonal antibodies. For triple-color analysis, Per-CP-labeled anti-CD4 or anti-CD8 and PE-labeled anti-CD45RA or anti-CD45RO monoclonal antibodies wereused. Following incubation, the cells were washed twicewith PBS and 5000 cells were acquired and analyzed bydual- or triple-color analysis using Consort 30 andFACScan Research Software, respectively (Becton Dick-inson).
Table I. Expression of Fas in T Cell Subsets"
Cord (n = 5)Young (« = 5)
% Fas-positive cells
CD4+
CD45RA+
3 ±210 ±2
CD45RO+
3 ±222 ±4
CD8+
CD45RA+
3 ± 112 ±4
CD45RO+
5 ±225 ±4
" Mononuclear cells (1 X 106 cells) were stained with monoclonal antibodies as described under Materials and Methods. Data are represented as themean ± SD percentage of Fas+ cells which were CD4* (CD45RA+/CD45RO+) or CD8+ (CD45RA+/CD45RO+) T cell types.
Journal of Clinical Immunology, Vol. 17, No. 1,1997
APOPTOSIS IN CORD BLOOD LYMPHOCYTES 65
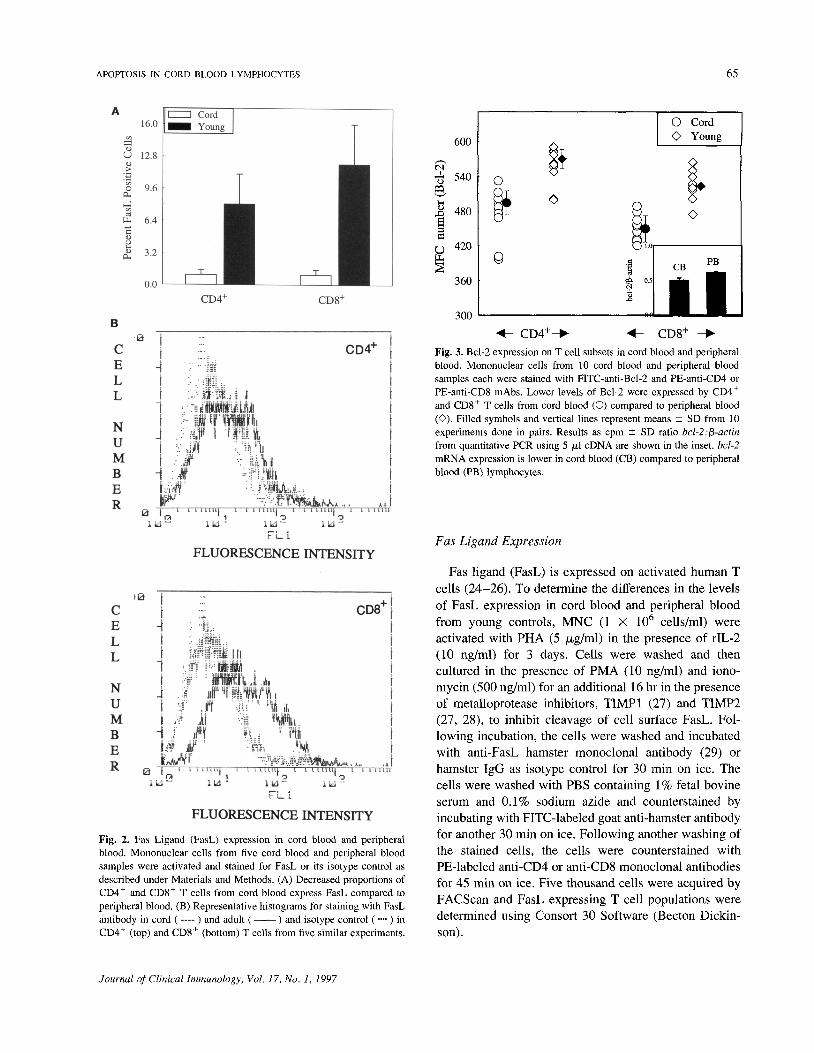
Fig. 2. Fas Ligand (FasL) expression in cord blood and peripheralblood. Mononuclear cells from five cord blood and peripheral bloodsamples were activated and stained for FasL or its isotype control asdescribed under Materials and Methods. (A) Decreased proportions ofCD4+ and CD8+ T cells from cord blood express FasL compared toperipheral blood. (B) Representative histograms for staining with FasLantibody in cord ( ---- ) and adult (—— ) and isotype control (••••) inCD4+ (top) and CD8+ (bottom) T cells from five similar experiments.
Fig. 3. Bcl-2 expression on T cell subsets in cord blood and peripheralblood. Mononuclear cells from 10 cord blood and peripheral bloodsamples each were stained with FITC-anti-Bcl-2 and PE-anti-CD4 orPE-anti-CD8 mAbs. Lower levels of Bcl-2 were expressed by CD4+
and CD8+ T cells from cord blood (O) compared to peripheral blood(O). Filled symbols and vertical lines represent means ± SD from 10experiments done in pairs. Results as cpm ± SD ratio bcl-2:fi-actinfrom quantitative PCR using 5 jul cDNA are shown in the inset. bcl-2mRNA expression is lower in cord blood (CB) compared to peripheralblood (PB) lymphocytes.
Fas Ligand Expression
Fas ligand (FasL) is expressed on activated human Tcells (24-26). To determine the differences in the levelsof FasL expression in cord blood and peripheral bloodfrom young controls, MNC (1 X 106 cells/ml) wereactivated with PHA (5 |U,g/ml) in the presence of rIL-2(10 ng/ml) for 3 days. Cells were washed and thencultured in the presence of PMA (10 ng/ml) and iono-mycin (500 ng/ml) for an additional 16 hr in the presenceof metalloprotease inhibitors, TIMP1 (27) and TIMP2(27, 28), to inhibit cleavage of cell surface FasL. Fol-lowing incubation, the cells were washed and incubatedwith anti-FasL hamster monoclonal antibody (29) orhamster IgG as isotype control for 30 min on ice. Thecells were washed with PBS containing 1% fetal bovineserum and 0.1% sodium azide and counterstained byincubating with FITC-labeled goat anti-hamster antibodyfor another 30 min on ice. Following another washing ofthe stained cells, the cells were counterstained withPE-labeled anti-CD4 or anti-CD8 monoclonal antibodiesfor 45 min on ice. Five thousand cells were acquired byFACScan and FasL expressing T cell populations weredetermined using Consort 30 Software (Becton Dickin-son).
Journal of Clinical Immunology, Vol. 17, No. 1, 1997

66 AGGARWAL, GUPTA, NAGATA, AND GUPTA
Table II. Expression of Bcl-2 in T Cell Subsets"
Cord (n = 5)Young (n = 5)
Mean fluorescence channel number
CD4+
CD45RA+
558 ± 13596 ± 10
CD45RO+
517 ± 25570 ± 8
CD8+
CD45RA+
554 ± 12585 ± 16
CD45RO+
542 ± 28561 ± 16
"Mononuclear cells (1 X 106 cells) were stained with monoclonal antibodies as described under Materials and Methods. Data are represented asmean ± SD fluorescence channel intensity of Bcl-2+ cells which were CD4+ (CD45RA+/CD45RO+) or CD8+ (CD45RA+/CD45RO+) T celltypes.
Quantitative Polymerase Chain Reaction (PCR)
Two hundred nanograms of total cellular RNA wasextracted from cord blood and peripheral blood. cDNA
Fig. 4. Expression of bcl-xL and bax mRNA in cord blood andperipheral blood lymphocytes. bcl-xL (A) and bax (B) mRNA expres-sion analyzed by quantitative PCR performed using bcl-xL- or bax-specific primers on two samples each from cord blood and peripheralblood lymphocytes. Vertical bars represent mean cpm ± SD ratios forgraded amounts (1, 5, and 10 pi, respectively, for bcl-xL or bax; 1 ;u,lfor j3-actin) of cDNA. All reactions were set up in triplicate.
was synthesized in a solution containing 100 ng ofrandom hexamers, 50 mM Tris-HCl, 75 mM KC1, 3 mMMgCl2, 10 mM DTT, a 500 /iM concentration of eachdNTP, and 10 U reverse transcriptase (RT) for 45 min atroom temperature. PCR was carried out in triplicateswith graded amounts of cDNA. fas-, bcl-2-, bcl-xL-, orbax-specific primers were each used at a 32 pM concen-tration. Each cycle of PCR included 1 min of denatur-ation at 94°C, 1 min of primer annealing at 60°C, and 2min of extension/synthesis at 72°C. Primers for fi-actinwere used as internal controls. For quantitation, 2 ptCi (1Ci = 37 GBq) of [a-32P]ATP was added to each reactionmixture. All experiments were performed in triplicate.PCR was carried out with a Thermal Cycler (Perkin-Elmer, CT) according to the kit instructions (PerkinElmer, Branchburg, NJ.). PCR products were separatedon 6% TBE gels and stained with ethidium bromide.Bands corresponding to each specific primer were ex-cised from gels and the amount of radioactivity incorpo-rated was determined by liquid scintillation counter.
DNA Staining
MNC (1 X 106) were washed with Dulbecco's PBS(DPBS) and incubated in the presence of FITC-labeledanti-CD4 or anti-CD8 monoclonal antibodies for 45 minon ice. Following incubation, the cells were washedtwice with DPBS containing 0.1% BSA (DPBS-BSA)and fixed in 0.25% paraformaldehyde for 15 min at roomtemperature in dark. After washing, cells were resus-pended in 70% ethanol and incubated overnight at-20°C. Following incubation, cells were washed twicewith DPBS-BSA and incubated in sodium citrate buffer(0.1%) containing 0.1% Triton X-100, 50 jug/ml RNaseA, and 50 jug/ml propidium iodide for 30 min at roomtemperature in dark. Ten thousand cells were acquiredand analyzed using FACScan.
TUNEL Assay
A TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end-labeling) assay was done
Journal of Clinical Immunology, Vol. 17, No. 1, 1997
APOPTOSIS IN CORD BLOOD LYMPHOCYTES 67
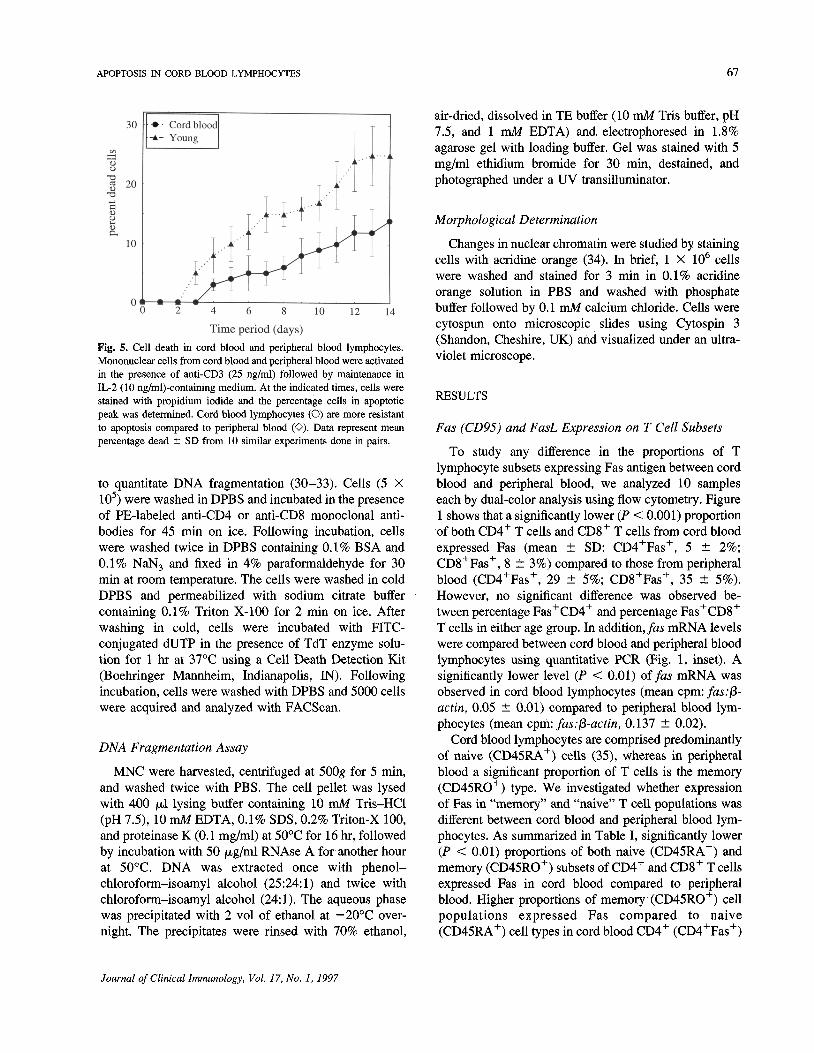
Fig. 5. Cell death in cord blood and peripheral blood lymphocytes.Mononuclear cells from cord blood and peripheral blood were activatedin the presence of anti-CD3 (25 ng/ml) followed by maintenance inIL-2 (10 ng/ml)-containing medium. At the indicated times, cells werestained with propidium iodide and the percentage cells in apoptoticpeak was determined. Cord blood lymphocytes (O) are more resistantto apoptosis compared to peripheral blood (O). Data represent meanpercentage dead ± SD from 10 similar experiments done in pairs.
to quantitate DNA fragmentation (30-33). Cells (5 X105) were washed in DPBS and incubated in the presenceof PE-labeled anti-CD4 or anti-CD8 monoclonal anti-bodies for 45 min on ice. Following incubation, cellswere washed twice in DPBS containing 0.1% BSA and0.1% NaN3 and fixed in 4% paraformaldehyde for 30min at room temperature. The cells were washed in coldDPBS and permeabilized with sodium citrate buffercontaining 0.1% Triton X-100 for 2 min on ice. Afterwashing in cold, cells were incubated with FITC-conjugated dUTP in the presence of TdT enzyme solu-tion for 1 hr at 37°C using a Cell Death Detection Kit(Boehringer Mannheim, Indianapolis, IN). Followingincubation, cells were washed with DPBS and 5000 cellswere acquired and analyzed with FACScan.
DNA Fragmentation Assay
MNC were harvested, centrifuged at 500g for 5 min,and washed twice with PBS. The cell pellet was lysedwith 400 /id lysing buffer containing 10 mM Tris-HCl(pH 7.5), 10 mM EDTA, 0.1% SDS, 0.2% Triton-X 100,and proteinase K (0.1 mg/ml) at 50°C for 16 hr, followedby incubation with 50 jug/ml RNAse A for another hourat 50°C. DNA was extracted once with phenol-chloroform-isoamyl alcohol (25:24:1) and twice withchloroform-isoamyl alcohol (24:1). The aqueous phasewas precipitated with 2 vol of ethanol at -20°C over-night. The precipitates were rinsed with 70% ethanol,
air-dried, dissolved in TE buffer (10 mM Tris buffer, pH7.5, and 1 mM EDTA) and electrophoresed in 1.8%agarose gel with loading buffer. Gel was stained with 5mg/ml ethidium bromide for 30 min, destained, andphotographed under a UV transilluminator.
Morphological Determination
Changes in nuclear chromatin were studied by stainingcells with acridine orange (34). In brief, 1 X 106 cellswere washed and stained for 3 min in 0.1% acridineorange solution in PBS and washed with phosphatebuffer followed by 0.1 mM calcium chloride. Cells werecytospun onto microscopic slides using Cytospin 3(Shandon, Cheshire, UK) and visualized under an ultra-violet microscope.
RESULTS
Fas (CD95) and FasL Expression on T Cell Subsets
To study any difference in the proportions of Tlymphocyte subsets expressing Fas antigen between cordblood and peripheral blood, we analyzed 10 sampleseach by dual-color analysis using flow cytometry. Figure1 shows that a significantly lower (P < 0.001) proportionof both CD4+ T cells and CD8+ T cells from cord bloodexpressed Fas (mean ± SD: CD4+Fas+, 5 ± 2%;CD8+Fas+, 8 ± 3%) compared to those from peripheralblood (CD4+Fas+, 29 ± 5%; CD8+Fas+, 35 ± 5%).However, no significant difference was observed be-tween percentage Fas+CD4+ and percentage Fas+CD8+
T cells in either age group. In addition, fas mRNA levelswere compared between cord blood and peripheral bloodlymphocytes using quantitative PCR (Fig. 1, inset). Asignificantly lower level (P < 0.01) of fas mRNA wasobserved in cord blood lymphocytes (mean cpm: fas:fi-actin, 0.05 ± 0.01) compared to peripheral blood lym-phocytes (mean cpm: fas:fB-actin, 0.137 ± 0.02).
Cord blood lymphocytes are comprised predominantlyof naive (CD45RA+) cells (35), whereas in peripheralblood a significant proportion of T cells is the memory(CD45RO+) type. We investigated whether expressionof Fas in "memory" and "naive" T cell populations wasdifferent between cord blood and peripheral blood lym-phocytes. As summarized in Table I, significantly lower(P < 0.01) proportions of both naive (CD45RA+) andmemory (CD45RO+) subsets of CD4+ and CD8+ T cellsexpressed Fas in cord blood compared to peripheralblood. Higher proportions of memory (CD45RO+) cellpopulations expressed Fas compared to naive(CD45RA+) cell types in cord blood CD4+ (CD4+Fas+)
Journal of Clinical Immunology, Vol. 17, No. 1, 1997
68 AGGARWAL, GUPTA, NAGATA, AND GUPTA
and CD8+ (CDS+Fas+) T cell subsets, though thedifferences were not significant (P > 0.01). In contrast,in peripheral blood, significantly (P < 0.01) higherproportions of memory (CD45RO+) cell types expressedFas compared to naive (CD45RA+) types in CD4+
(CD4+Fas+) and CD8+ (CD8+Fas+) T cells.Fas ligand (FasL) is expressed only on activated T
cells. We compared the proportions of T cell subsetsexpressing FasL between cord blood and peripheralblood. Mononuclear cells were stimulated with PHA andIL-2 for 72 hr, followed by activation with PMA andionomycin for an additional 16 hr. Data from fivesubjects in Fig. 2A show that a significantly lowered(P < 0.01) proportion of CD4+ and CD8+ T cell subsetsin cord blood expresses FasL (CD4+FasL+, 2 ± 1%;CD8+FasL+, 3 ± 1%) compared to peripheral blood(CD4+FasL+, 10 ± 1%; CD8+FasL+, 12 ± 2%).However, no significant difference was found betweenCD4+FasL+ and CD8+FasL+ T cell subsets in each agegroups. A representative histogram of FasL expression incord blood and peripheral lymphocytes is shown in Fig.2B.
Bcl-2 Expression on T Cell Subsets
Bcl-2 is involved in the survival of T cells and plays arole in maintaining cellular homeostasis. We examinedthe levels of Bcl-2 protein on cord blood and peripheralblood T cell subsets. Figure 3 shows data for Bcl-2staining from 10 subjects each from cord blood andperipheral blood. Cord blood T cell subsets expressedsignificantly (P < 0.01) lower levels of Bcl-2 (mean ±SD MFC number: CD4+ T cells, 494 ± 21; CD8+ Tcells, 449 ± 23) compared to peripheral blood (CD4+ Tcells, 570 ± 16; CD8+ T cells, 523 ± 25). However, nosignificant difference was found in Bcl-2 levels betweenCD4+ T cells and CD8+ T cells in both age groups.
A dissociation has been observed between Bcl-2protein and mRNA levels in several cell lines (36). Todetermine whether changes in Bcl-2 protein levels areassociated with changes at mRNA levels, we analyzedbcl-2 mRNA expression in cord blood lymphocytes andperipheral blood from young adults. As shown in Fig. 3(inset), cord blood lymphocytes expressed low (P <0.01) bcl-2 mRNA levels (bcl-2.-fiactin, 0.51 ± 0.03cpm) compared to peripheral blood (bcl-2.-fiactin,0.67 ± 0.02 cpm).
Bcl-2 expression at the protein levels in "memory"and "naive" cell populations was determined using tri-ple-color flow cytometric analysis on five samples eachfrom cord blood and peripheral blood. Results are shownin Table II. A decrease in Bcl-2 level was observed in
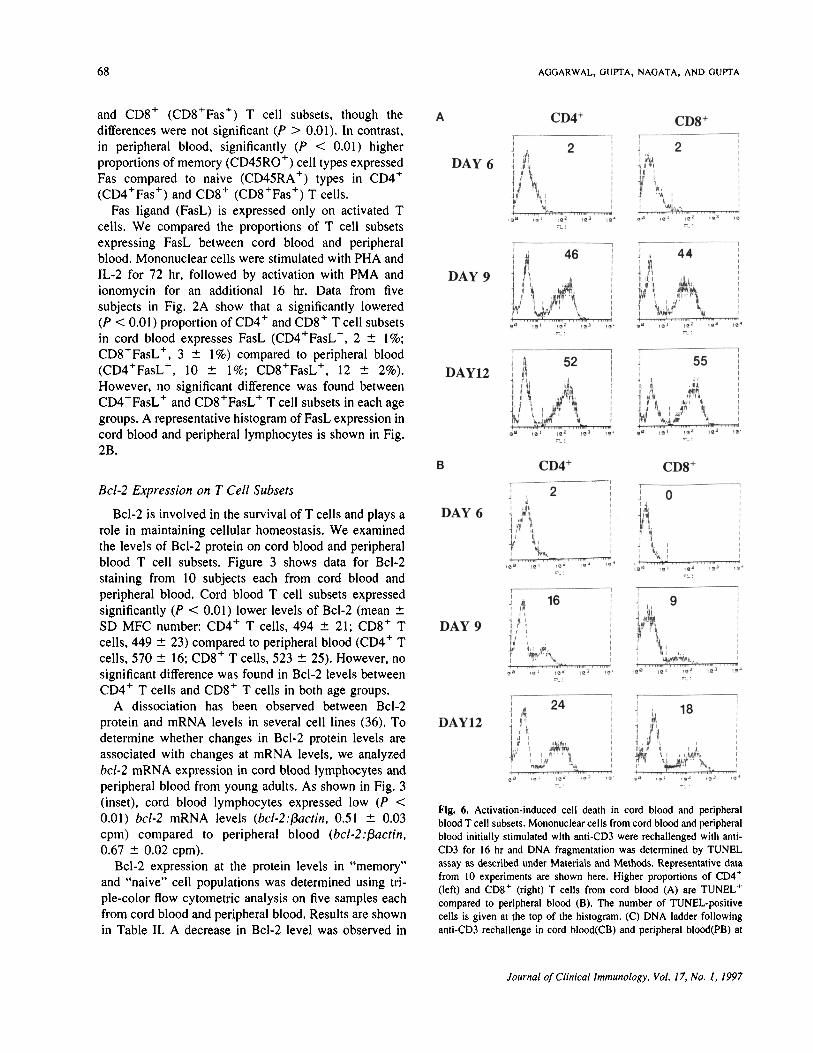
Fig. 6. Activation-induced cell death in cord blood and peripheralblood T cell subsets. Mononuclear cells from cord blood and peripheralblood initially stimulated with anti-CD3 were rechallenged with anti-CD3 for 16 hr and DNA fragmentation was determined by TUNELassay as described under Materials and Methods. Representative datafrom 10 experiments are shown here. Higher proportions of 004*(left) and CDS* (right) T cells from cord blood (A) are TUNEL*compared to peripheral blood (B). The number of TUNEL-positivecells is given at the top of the histogram. (C) DNA ladder followinganti-CD3 rechallenge in cord blood(CB) and peripheral blood(PB) at
Journal of Clinical Immunology, Vol. 17, No. 1, 1997

APOPTOSIS IN CORD BLOOD LYMPHOCYTES 69
Fig. 6. (Continued) days 6, 9, and 12 of culture. (D) Acridine orange staining of cells undergoing apoplosis in cord blood (CB; top) and peripheralblood (PB; bottom) following anti-CD3 rechallenge at days 6, 9, and 12 of culture. Original magnification, X40.
memory cell populations in CD4+ (P < 0.01) and CD8 +
(P < 0.02) T cell subsets (CD4+CD45RO+Bcl-2 andCD8+CD45RO+Bcl-2) from cord blood lymphocytescompared to peripheral blood. Furthermore, lower ex-pression of Bcl-2 was observed in naive cell populationsfrom cord blood (CD4 + CD45RA + Bcl-2 andCD8+CD45RA+Bcl-2) compared to peripheral blood(P < 0.01 and P < 0.02, respectively). Additionally,
CD4+ naive cell types (CD4+CD45RA+) expressedhigher Bcl-2 compared to memory cells(CD4+CD45RO+) in both age groups (P < 0.01 andP < 0.001, respectively), whereas in cord blood CD8+ Tcell types, no significant difference in Bcl-2 expressionwas found between naive (CD8+CD45RA+) and mem-ory (CD8+CD45RO+) cells (P > 0.5), whereas, differ-ences were significant in peripheral blood (P < 0.05).
Journal of Clinical Immunology, Vol. 17, No. 1, 1997
70 AGGARWAL, GUPTA, NAGATA, AND GUPTA
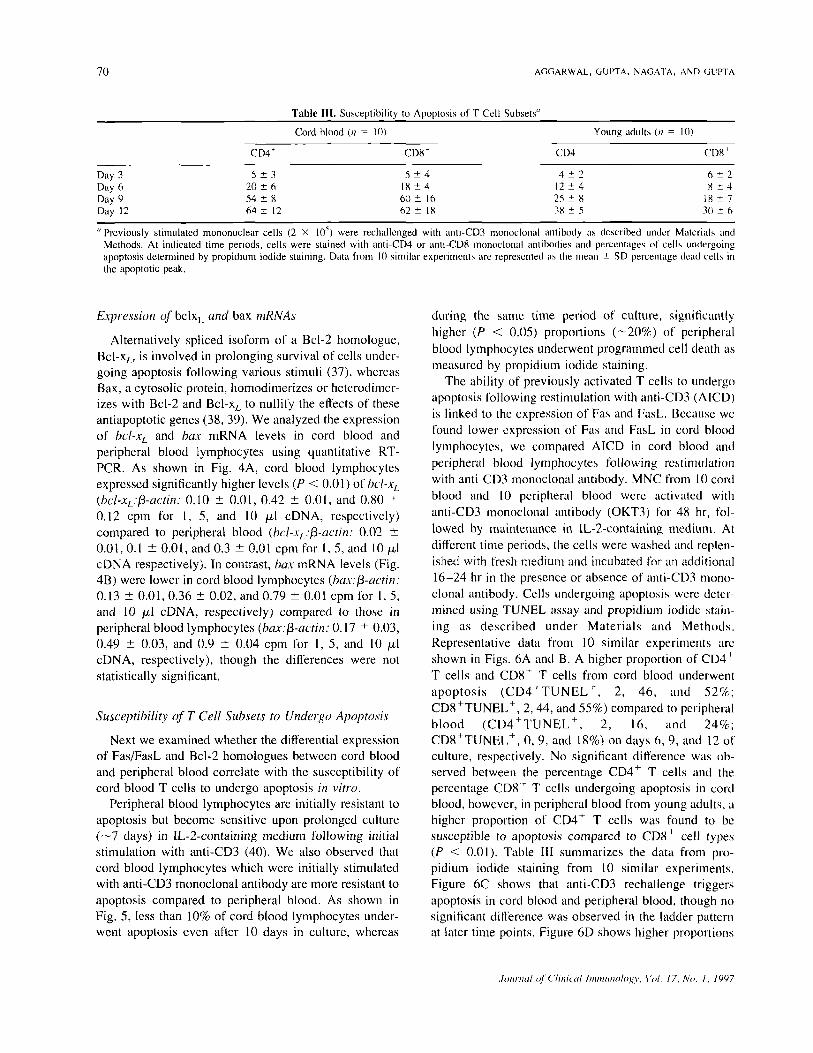
Table III. Susceptibility to Apoptosis of T Cell Subsets"
Day 3Day 6Day 9Day 12
Cord blood (n = 10)
CD4"
5 ± 320 ±654 ± 864 ± 12
CD8'
5 ±418 ± 460 ± 1662 ± 18
Young adults (n = 10)
CD41
4± 212 ± 425 ± 838 ± 5
CD8'
6 ± 28 ± 4
IX ± 73(1 ± 6
"Previously stimulated mononuclear cells (2 X 10~) were rechallenged with anti-CD3 monoclonal antibody as described under Materials andMethods. At indicated time periods, cells were stained with anti-CD4 or anti-CD8 monoclonal antibodies and percentages of cells undergoingapoptosis determined by propidium iodide staining. Data from 10 similar experiments are represented as the mean ± SD percentage dead cells inthe apoptotic peak.
Expression of bclxL and bax mRNAs
Alternatively spliced isoform of a Bcl-2 homologue,Bcl-xL, is involved in prolonging survival of cells under-going apoptosis following various stimuli (37), whereasBax, a cytosolic protein, homodimerizes or heterodimer-izes with Bcl-2 and Bcl-xL to nullify the effects of theseantiapoptotic genes (38, 39). We analyzed the expressionof bcl-xL and bax mRNA levels in cord blood andperipheral blood lymphocytes using quantitative RT-PCR. As shown in Fig. 4A, cord blood lymphocytesexpressed significantly higher levels (P < 0.01) of bcl-xL
(bd-xL:$-actin: 0.10 ± 0.01, 0.42 ±0.01, and 0.80 ±0.12 cpm for 1, 5, and 10 /il cDNA, respectively)compared to peripheral blood (bcl-xL:fi-actin: 0.02 ±0.01, O.I ± 0.01, and 0.3 ± 0.01 cpm for I, 5, and 10 /xlcDNA respectively). In contrast, bax mRNA levels (Fig.4B) were lower in cord blood lymphocytes (bax:fi-actin:0.13 ± 0.01, 0.36 ± 0.02, and 0.79 ± 0.01 cpm for 1, 5,and 10 /itl cDNA, respectively) compared to those inperipheral blood lymphocytes (bax:fi-actin: 0.17 ± 0.03,0.49 ± 0.03, and 0.9 ± 0.04 cpm for 1, 5, and 10 /nlcDNA, respectively), though the differences were notstatistically significant.
Susceptibility of T Cell Subsets to Undergo Apoptosis
Next we examined whether the differential expressionof Fas/FasL and Bcl-2 homologues between cord bloodand peripheral blood correlate with the susceptibility ofcord blood T cells to undergo apoptosis in vitro.
Peripheral blood lymphocytes are initially resistant toapoptosis but become sensitive upon prolonged culture(~7 days) in IL-2-containing medium following initialstimulation with anti-CD3 (40). We also observed thatcord blood lymphocytes which were initially stimulatedwith anti-CD3 monoclonal antibody are more resistant toapoptosis compared to peripheral blood. As shown inFig. 5, less than 10% of cord blood lymphocytes under-went apoptosis even after 10 days in culture, whereas
during the same time period of culture, significantlyhigher (P < 0.05) proportions (—20%) of peripheralblood lymphocytes underwent programmed cell death asmeasured by propidium iodide staining.
The ability of previously activated T cells to undergoapoptosis following restimulation with anti-CD3 (A1CD)is linked to the expression of Fas and FasL. Because wefound lower expression of Fas and FasL in cord bloodlymphocytes, we compared AICD in cord blood andperipheral blood lymphocytes following restimulationwith anti-CD3 monoclonal antibody. MNC from 10 cordblood and 10 peripheral blood were activated withanti-CD3 monoclonal antibody (OKT3) for 48 hr, fol-lowed by maintenance in IL-2-containing medium. Atdifferent time periods, the cells were washed and replen-ished with fresh medium and incubated for an additional16-24 hr in the presence or absence of anti-CD3 mono-clonal antibody. Cells undergoing apoptosis were deter-mined using TUNEL assay and propidium iodide stain-ing as described under Materials and Methods.Representative data from 10 similar experiments areshown in Figs. 6A and B. A higher proportion of CD4 'T cells and CD8 + T cells from cord blood underwentapoptosis (CD4+TUNELh, 2, 46, and 52%;CD8+TUNEL+, 2, 44, and 55%) compared to peripheralblood (CD4 + TUNEL + , 2, 16, and 24%;CD8+TUNEL+, 0, 9, and 18%) on days 6, 9, and 12 ofculture, respectively. No significant difference was ob-served between the percentage CD4+ T cells and thepercentage CD8 h T cells undergoing apoptosis in cordblood, however, in peripheral blood from young adults, ahigher proportion of CD4+ T cells was found to besusceptible to apoptosis compared to CD8 ' cell types(P < 0.01). Table III summarizes the data from pro-pidium iodide staining from 10 similar experiments.Figure 6C shows that anti-CD3 rechallenge triggersapoptosis in cord blood and peripheral blood, though nosignificant difference was observed in the ladder patternat later time points. Figure 6D shows higher proportions
Journal of Clinical Immunology: Vol. 17. No. 1, 1997

APOPTOSIS IN CORD BLOOD LYMPHOCYTES 71
Fig. 7. Anti-Fas-induced cell death in cord blood and peripheral bloodT cell subsets. Previously stimulated mononuclear cells from cordblood and peripheral blood were rechallenged with anti-Fas monoclo-nal antibody or its isotype control (IgM) as described under Materialsand Methods. Percentage cells undergoing apoptosis was determinedusing propidium iodide staining. (A) Significantly lowered proportionsof CD4+ and CD8+ T cells from cord blood undergo anti-Fas-inducedapoptosis compared to peripheral blood. Bars represent the mean ± SDpercentage increase in dead cells in the presence of anti-Fas antibodyover IgM controls.
of apoptotic nuclei in cord blood lymphocytes followinganti-CD3 rechallenge compared to peripheral blood.
To confirm that the killing we observed was Fasmediated, previously activated T cells were restimulatedwith anti-Fas monoclonal antibody. As shown in Fig. 7A,an increased proportion of CD4+ and CD8+ T cellsunderwent apoptosis in the presence of anti-Fas antibodyin both age groups. However, the percentage increase incord blood CD4+ T cells (8 ± 3%) and in CD8+ T cells(16 ± 4%) was significantly less (P < 0.1) compared toCD4+ T cells (71 ± 8%) and CD8+ T cells (68 ± 10%)from peripheral blood. Figures 7B and C show represen-tative histograms from propidium iodide-stained CD4+
and CD8+ T cells with or without anti-Fas restimulationin cord blood and peripheral blood, respectively.
DISCUSSION
Programmed cell death (apoptosis) plays a major rolein the maintenance of the immune system (41, 42). Themajority of cells leaving the thymus die by a process ofapoptosis (43). Dysregulation of programmed cell deathleads to many disease states in both experimental ani-mals and humans (44-46). In the present study, we haveshown that there is a differential expression of genesregulating apoptosis including fas, bcl-2, bcl-xL, and baxbetween cord blood and peripheral blood. Furthermore,we show that the cord blood T cells are more susceptible
Fig. 7. (Continued) (B, C) Left panels show the percentage apoptoticcells in the presence of isotype IgM control for cord blood (CB) andperipheral blood (PB) CD4+ T cells (B) and CD8+ T cell subsets (C).Right panels show the percentage cells undergoing apoptosis in thepresence of anti-Fas monoclonal antibody. The percentage of apoptoticcells is shown at the top of histograms. A representative of 10 similarexperiments is shown.
to activation-induced anti-CD3-mediated apoptosis com-pared to peripheral blood.
The process of activation-induced cell death in matureT cells is important in regulating peripheral immuneresponses and requires expression of functional Fas andFas ligand (FasL) by T cell populations (47). Defectivetranscription of Fas and FasL has been shown to beassociated with lymphoproliferation and autoimmunephenotypes in Ipr and gld mice, respectively (48, 49). Inthis study we have shown that in cord blood, significantlylower proportions of CD4+ and CD8+ T cells expressFas and FasL compared to peripheral blood T cells.Decreased expression of Fas was also observed in cord
Journal of Clinical Immunology, Vol. 17, No. 1, 1997

72 AGGARWAL, GUPTA, NAGATA, AND GUPTA
blood T cell memory and naive cell subpopulations.Shinohara et al. (50) have also reported decreasedproportions of Fas in cord blood CD4+ and CD8+ T cellsubsets.
Recent reports have shown that increased Fas/FasLexpression results in increased sensitivity to apoptosis(51, 52). We observed that cord blood T cell subsets aremore resistant to apoptosis upon primary stimulationcompared to peripheral blood. This is in agreement withour Fas/FasL expression data and suggests that cordblood T cell subsets undergo Fas-dependent apoptosis.Bcl-2 and Bcl-xt are involved in enhancing cell survivalin cells undergoing Fas-dependent apoptosis (22). Wefound that cord blood lymphocytes express low levels ofBcl-2, a low bcl-2/bax ratio, and high bcl-xL compared toperipheral blood. In addition, cord blood T cells showedincreased susceptibility to secondary stimulation withanti-CD3 monoclonal antibody compared to peripheralblood. Therefore, it is possible that in cord blood, highbcl-xL levels may be responsible for rescue from apopto-sis upon primary stimulation, whereas the low Bcl-2expression and low bcl-2/bax ratio are responsible forincreased cell death following secondary stimulation.Our results are in disagreement with the studies inneonatal mice by Adkins et al, (53), who showed that Tcells from neonatal mice are more susceptible to apopto-sis upon primary stimulation and resistant to secondarystimulation. Moreover, in these mice >70% of T cellsexpressed Fas antigen and the authors suggest that theincreased expression of Fas antigen on naive T cellsfound on neonatal mice is responsible for the increasedapoptosis following primary stimulation. However, lev-els of Bcl-2, Bcl-xL, or Bax were not measured, whichcould explain the differences between Adkins et al. andthe present study. Furthermore, species differences mightalso account for this discrepancy.
Differential sensitivity for anti-Fas-induced apoptosishas been found in human peripheral lymphocytes (40).We found that activated T cells acquire sensitivity foranti-Fas-mediated apoptosis during culture in vitro, anobservation made by others (40, 54). However, cordblood T cells were found to be significantly less sensitiveto anti-Fas-mediated apoptosis compared to peripheralblood. A possible explanation for this observation is thatdue to lower expression of Fas/FasL on cord blood Tcells even after activation, enough Fas receptor or FasLmolecules are not available for effective cross-linking ofreceptor and, hence, triggering of cell death. Further-more, Fas-independent AICD mechanisms for apoptosishave been reported (55). It is possible that both Fas-dependent and Fas-independent mechanisms regulateprogrammed cell death of T cells in cord blood lympho-
cytes; Fas-mediated apoptosis is decreased, whereasFas-independent apoptosis is increased in cord blood Tcells. The mechanisms underlying abnormalities of bothFas-dependent and Fas-independent apoptosis in cordblood are under investigation.
REFERENCES
I .
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
Stutman O: Ontogeny of T cells. Clin Immunol Allergy 5:191-196, 1985Takahashi N, Imanishi K, Nishida H, Uchiyama T: Evidence forimmunologic immaturity of cord blood T cells. Cord blood T cellsare susceptible to tolerance induction to in vitro stimulation with asuperantigen. J Immunol 155:5213-5219, 1995Aggarwal S, Gupta A, Gupta S: Differential expression of P-glycoprotein in cord blood and peripheral blood lymphocytesubpopulations. Int J Oncol 8:1063-1068, 1996McDougall RM, Suitters AJ, Smith H, Yacob MH. Rose ML:Increased cyclosporin sensitivity of T cells from cord bloodcompared with those from the adult. Clin Exp Immunol 95:519-524, 1994Kavelaars A, Zijlstra J, Bakkar JM, Van Rees EP, Visser GH,Zegers BJ, Heijnen CJ: Increased dexamethasone sensitivity ofneonatal leukocytes: Different mechanisms of glucocorticoid inhi-bition of T cell proliferation in adult and neonatal cells. EurJ Immunol 25:1346-1351, 1995Cohen JJ, Duke RC, Fadok VA, Sellins KS: Apoptosis andprogrammed cell death in immunity. Annu Rev Immunol 10:267-293, 1992Boise LH, Gottschalk AR, Quintans J, Thompson CB: Bcl-2 andBcl-2 related proteins in apoptosis regulation. Curr Topics Micro-biol Immunol 200:107-121, 1995Korsmeyer S3: Regulators of cell death. Trends Genet 11:101 -105,1995Nagata S, Golstein P: The Fas death factor. Science 267:1449-1456, 1995Osborne BA: Induction of genes during apoptosis: Examples fromimmune system. Semin Cancer Biol 6:27-33, 1995Reed JC: Bcl-2 family proteins: Regulators of chemoresistance incancer. Toxicol Lett 82-83:155-158, 1995Itoh N, Yonehara S, Ishil A, Yonehara M, Mizushima S-I,Sameshima M, Hase A, Seto Y, Nagata S: The polypeptideencoded by the cDNA for human cell surface antigen Fas canmediate apoptosis. Cell 66:233-243, 1991Reed JC: Bcl-2: Prevention of apoptosis as a mechanism of drugresistance. Hematol Oncol Clin North Am 9:451-474, 1995Hockenbery D, Nunez G, Milliman C, Schreiber RD, KorsmeyerSJ: Bcl-2 is an inner mitochondrial membrane protein that blocksprogrammed cell death. Nature 348:334-336, 1990Minn AJ, Rudin CM, Boise LH, Thompson CB: Expression ofBcl-x, can confer multidrug resistance phenotype. Blood 86:1903-1910, 1995Krajewski S, Blomvqvist C, Franssila K, Krajewska M. WaseniusV-M, Niskanen E, Reed JC: Reduced expression of pro-apoptoticgene Bax is associated with poor response rates to combinationchemotherapy and shorter survival in women with metastaticbreast adenocarcinoma. Cancer Res 55:4471-4478, 1995Alderson MR, Tough TW, Davis-Smith T, Braddy S, Falk B,Schooley KA, Goodwin RG, Smith CA, Ramsdell F. Lynch DH:Fas ligand mediates activation-induced cell death in human Tlymphocytes. J Exp Med 181:71-77, 1995
Journal of Clinical Immunology, Vol. 17, No. 1, 1997

APOPTOSIS IN CORD BLOOD LYMPHOCYTES 73
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
Yonehara S, Ishii A, Yonehara M: A cell killing monoclonal antibody(anti-Fas) to a cell surface antigen co-downregulated with the receptorof tumor necrosis factor. J Exp Med 169:1747-1756, 1989Kobayashi N, Hamamoto Y, Yamamoto N, Ishii A, Yonehara M,Yonehara S: Anti-Fas monoclonal antibody is cytocidal to humanimmunodeficiency virus infected cells without augmenting viralreplication. Proc Natl Acad Sci USA 87:9620-9624, 1990Nagafuji K, Shibuya T, Harada M, Mizuno S-I, Takenaka K,Miyamoto T, Okamura T, Gondo H, Niho Y: Functional expres-sion of Fas antigen (CD95) on hematopoietic progenitor cells.Blood 86:883-889, 1995Kitada S, Miyashita T, Tanaka S, Reed JC: Investigations ofantisense oligonucleotides targeted against bcl-2 RNAs. AntisenseRes Dev 3:157-169, 1993Boise LH, Gonzalez-Garcia M, Postema CE, Ding L, Lindsten T,Turka LA, Mao X, Nunez G, Thompson CB: Bcl-x, a Bcl-2-relatedgene that functions as a dominant regulator of apoptotic cell death.Cell 74:597-608, 1993Martinez-Valdez H, Guret C, de Bouteiller O, Fugier I, BanchereauJ, Liu Y-J: Human germinal center B cells express the apoptosis-inducing genes fas, c-myc, p53, and bax but not the survival genebcl-2. i Exp Med 183:971-977, 1995Suda T, Takahashi T, Golstein P, Nagata S: Molecular cloning andexpression of the Fas ligand, a novel member of the tumor necrosisfactor family. Cell 75:1169-1178, 1993Lynch DH, Ramsdell F, Alderson MR: Fas and FasL in homeo-static regulation of immune responses. Immunol Today 16:569-572, 1995Vignaux F, Vivier E, Malissen B, Depraetere V, Nagata S,Goldstein P: TCR/CD3 coupling to Fas based cytotoxicity. J ExpMed 181:781-786, 1995Liotta LA, Stetler-Stevenson WG: Metalloproteinases and cancerinvasion. Semin Cancer Biol 1:99-106, 1990Boone TC, Johnson MJ, De Clerck YA, Langley KE: cDNAcloning and expression of a metalloproteinase inhibitor related totissue inhibitor of metalloproteinases. Proc Natl Acad Sci USA87:2800-2804, 1990Tanaka M, Suda T, Haze K, Nakamura N, Sato K, Kimura F,Motoyashi K, Drummond AH, Nagata S: Fas ligand in humanserum. Nature Med 2:317-322, 1996Gavrieli Y, Sherman Y, Ben-Sasson SA: Identification of pro-grammed cell death in situ via specific labeling of nuclear DNAfragmentation. J Cell Biol 119:493-501, 1992Sgonc R, Boeck G, Dietrich H, Gruber J, Recheis H, Wick G:Simultaneous determination of cell surface antigens and apoptosis.Trends Genet 10:41-42, 1994Mochizuki H, Nakamura N, Nishi K, Mizuno Y: Apoptosis isinduced by l-methyl-4-phenylpyridinium ion (MPP) in ventralmesencephalic striatal co-culture in rat. Neurosci Lett 170:191-194, 1994Portera-Cailliau C, Sung CH, Nathans J, Adler R: Apoptoticphotoreceptor cell death in mouse models of retinitis pigmemosa.Proc Natl Acad Sci USA 91:974-978, 1994Guilhot S, Miller T, Cornman G, Isom HC: Apoptosis induced bytumor necrosis factor-a in rat hepatocyte cell lines expressinghepatitis B virus. Am J Pathol 148:801-814, 1996Chheda S, Palkowetz KH, Rassin DK, Goldman AS: Deficientquantitative expression of CD45 isoforms on CD4+ and CDS ' Tcell subpopulations and subsets of CD45RA<lim)CD45RO(l"wl Tcells in newborn blood. Biol Neonate 69:128-132, 1996Chleq-Deschamps CM, LeBrun DP, Huie P, Besnier DP, WarnkeRA, Sibley RK, Cleary ML: Topographical dissociation of bcl-2
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
messenger RNA and protein expression in human lymphoidtissues. Blood 81:293-298, 1993Nunez G, Merino R, Grillot D, Gonzalez-Garcia M: Bcl-2 andBcl-x: Regulatory switches for lymphoid death and survival.Immunol Today 15:582-588, 1994Oltvai ZN, Milliman CL, Korsmeyer SJ: Bcl-2 heterodimerizes invivo with a conserved homolog, Bax, that accelerates programmedcell death. Cell 47:609-619, 1993Yin XM, Oltvai ZN, Korsmeyer SJ: Heterodimerization with Baxis required for Bcl-2 to repress cell death. Curr Topics MicrobiolImmunol 194:331-338, 1995Owen-Schaub LB, Yonehara S, Crump WL III, Grimm E: DNAfragmentation and cell death is selectively triggered in activatedhuman lymphocytes by Fas antigen engagement. Cell Immunol140:197-205, 1995Golstein P, Ojcius DM, Young JD-E: Cell death mechanism andthe immune system. Immunol Rev 121:129-165, 1991Cohen JJ, Duke RC: Apoptosis and programmed cell death inimmunity. Annu Rev Immunol 10:267-293, 1992Smith SW, McLaughin KA, Osborne BA: Molecular events inthymocyte apoptosis. Curr Topics Microbiol Immunol 200:147-162, 1995Fisher GH, Rosenberg FJ, Straus SE, Dale JK, Middelton LA, LinAY, Strober W, Lenardo MJ, Puck JM: Dominant interfering fasgene mutations impair apoptosis in a human autoimmune lympho-proliferative syndrome. Cell 81:935-946, 1995Lynch DH, Ramsdell F, Alderson MR: Fas and FasL in thehomeostatic regulation of immune responses. Immunol Today16:569-574, 1995Cohen PL, Eisenberg RA: Lpr and gld: Single gene models ofsystemic autoimmunity and lymphoproliferative disease. AnnuRev Immunol 9:243-262, 1991Russell JH: Activation-induced death of mature T cells in theregulation of immune responses. Curr Opin Immunol 7:382-388,1995Wanatabe-Fukunaga R, Brannan CL, Copeland NG, Jenkins NA,Nagata S: Lymphoproliferation disorder in mice explained bydefects in Fas antigen that mediates apoptosis. Nature 356:314-317, 1992Takashi R, Tanaka M, Brannan CL, Jenkins NA, Copeland NG,Suda T, Nagata S: Generalized lymphoproliferative disease in micecaused by a point mutation in the Fas ligand. Cell 76:969-976, 1994Shinohara S, Sawada T, Nishioka Y, Tohma S, Kisaki T, Inoue T,Ando K, Ikeda M, Fuji H, Ito K: Differential expression of Fasantigen and Bcl-2 protein on CD4+ T cells, CD8+ T cells andmonocytes. Cell Immunol 163:303-308, 1995Ozaizu N, Pahwa S: Role of apoptosis in HIV disease pathogen-esis. J Clin Immunol 15:217-231, 1995Kotani T, Aratake Y, Kondo S, Tamura K, Ohtaki S: Expression offunctional Fas antigen on adult T-cell leukemia. Leuk Res 18:305-310, 1994Adkins B, Chun K, Hamilton K, Nassiri M: Naive murine neonatalT cells undergo apoptosis in response to primary stimulation.J Immunol 157:1343-1349, 1996Brunner T, Mogil RJ, LaFace D, Yoo NJ, Mahboubi A, EcheverriF, Martin SJ, Force WR, Lynch DH, Ware CF, Green DR: Cellautonomous Fas(CD95)/Fas-ligand interaction mediates activation-induced apoptosis in T cell hybridomas. Nature 373:441-444, 1995Tucek-Szabo CL, Andjelic S, Lacy E, Elkon KB, Nikolic-Zugic J:Surface T cell Fas receptor/CD95 regulation, in vivo activation,and apoptosis. Activation-induced death can occur without Fasreceptor. J Immunol 156:192-200, 1996
Journal of Clinical Immunology, Vol. 17, No. 1, 1997




















