
Endoleaks after endovascular aneurysm repair lead to nonuniform intra-aneurysm sac pressure
Upload
independentCategory
view
1download
0
NVESTIGATION into the causes of the genesis, growth, andrupture of intracranial aneurysms is relevant. The mostfrequently occurring aneurysms are saccular and con-
sidered to be the combined result of a multifactorial patho-genetic complex.36 Both congenital and acquired factorshave been investigated.5,17,34,35,40 Hemodynamic stress at ar-terial bifurcations, congenital medial defects, and degen-erative arterial wall changes are believed to contribute toaneurysm development.9 Other factors, such as hyperten-sion, smoking, atherosclerosis, and alcohol intake, are com-monly related to the pathogenesis and eventual rupture ofintracranial aneurysms.5 Several researchers have positedthat these aneurysms may be associated with a number ofgenetic factors whose role in lesion origin and pathogenesisappears to be increasingly relevant.34,35 A number of authorshave described families with two or more members affect-ed by a ruptured or an unruptured intracranial aneurysm.24,30,
34,40 In several hereditary disorders an association with in-tracranial aneurysms has been clearly described.1,6,10,39 De-spite the progress in research, however, basic genetic mech-anisms related to intracranial aneurysms remain unclear.
Other authors have suggested that apoptosis (genetical-ly programmed cell death) may be involved in aneurysmgrowth and rupture. In a number of congenital and/oracquired pathological conditions, apoptotic phenomenamay become pronounced, damaging arterial walls.14,20 Anattempt to delineate the mechanisms of apoptotic smooth-muscle cell death in intracranial aneurysms has been pro-posed by Stehbens,37 who studied the involvement of phos-phorylated c-Jun NH2-terminal kinase/c-Jun pathways.
Much literature covers the basic science of apoptosis.8,12,
21,22,25,29,32,33,41,43–45,47 Briefly, it functions as a homeostaticregulator of an appropriate cell population, maintaining asteady state by balancing cell replication and death. Ap-parently, to initiate the apoptotic mechanism a cell muststart novel gene expression and the synthesis of new pro-teins.42,45 Different mechanisms may induce a cell to startapoptosis.8,41,42,45 Often the trigger comes from the extracel-lular environment, which indicates that the locus of pro-grammed cell death is outside the cell. Perhaps apoptosiscan be modulated by manipulating the genes involved in itsregulation. It is a relevant phenomenon in vascular pathol-ogy, and there is evidence that the balance between cellproliferation and apoptosis regulates the number of cells invascular walls.2,3 High levels of apoptosis have been iden-tified in the perinatal phases of remodeling in arteries inlambs.7 Furthermore, apoptosis has been related to athero-
J Neurosurg 101:1018–1025, 2004
1018
Role of apoptosis in intracranial aneurysm rupture
LUIGI PENTIMALLI, M.D., PH.D., ANDREA MODESTI, M.D., ANDREA VIGNATI, M.D.,ENRICO MARCHESE, M.D., PH.D., ALESSIO ALBANESE, M.D., FEDERICO DI ROCCO, M.D.,ANNA COLETTI, PH.D., PAOLO DI NARDO, M.D., CRISTINA FANTINI, PH.D.,BARBARA TIRPAKOVA, M.D., AND GIULIO MAIRA, M.D.
Department of Neurosurgery, Catholic University, Institute of Experimental Medicine and BiochemicalSciences; and Department of Internal Medicine, Tor Vergata University, Rome, Italy
Object. Mechanisms involved in the rupture of intracranial aneurysms remain unclear, and the literature on apoptosis inthese lesions is extremely limited. The hypothesis that apoptosis may reduce aneurysm wall resistance, thus contributingto its rupture, warrants investigation. The authors in this study focused on the comparative evaluation of apoptosis in rup-tured and unruptured intracranial aneurysms. Peripheral arteries in patients harboring the aneurysms and in a group of con-trols were also analyzed.
Methods. Between September 1999 and February 2002, specimens from 27 intracranial aneurysms were studied. In 13of these patients apoptosis was also evaluated in specimens of the middle meningeal artery (MMA) and the superficial tem-poral artery (STA). The terminal deoxynucleotidyl transferase–mediated deoxyuridine triphosphate nick-end labeling tech-nique was used to study apoptosis via optical microscopy; electron microscopy evaluation was performed as well. Apop-totic cell levels were related to patient age and sex, aneurysm volume and shape, and surgical timing.
Significant differences in apoptosis were observed when comparing ruptured and unruptured aneurysms. High levels ofapoptosis were found in 88% of ruptured aneurysms and in only 10% of unruptured lesions (p , 0.001). Elevated apop-tosis levels were also detected in all MMA and STA specimens obtained in patients harboring ruptured aneurysms, where-as absent or very low apoptosis levels were observed in MMA and STA specimens from patients with unruptured aneu-rysms. A significant correlation between aneurysm shape and apoptosis was found.
Conclusions. In this series, aneurysm rupture appeared to be more related to elevated apoptosis levels than to thevolume of the aneurysm sac. Data in this study could open the field to investigations clarifying the causes of aneurysm en-largement and rupture.
KEY WORDS • apoptosis • intracranial aneurysm • subarachnoid hemorrhage
I
J. Neurosurg. / Volume 101 / December, 2004
Abbreviations used in this paper: MMA = middle meningealartery; SAH = subarachnoid hemorrhage; STA = superficial tempo-ral artery; TUNEL = terminal deoxynucleotidyl transferase–mediat-ed deoxyuridine triphosphate nick-end labeling technique.
12_04_JNS.2 11/22/04 11:03 AM Page 1018

sclerotic plaque formation12 and observed in cultured hu-man vascular smooth–muscle cells.4 In abdominal aortic an-eurysms in humans, a significant reduction in the density ofsmooth-muscle cells has been reported and even proposedas evidence of apoptosis.15,23 These data permit one to spec-ulate on the contribution of apoptosis to aneurysm wall de-generation.
Nonetheless, very few studies have been performed onthe role of apoptosis in the growth and rupture of intra-cranial aneurysms. In 1998 Kondo, et al.,20 using TUNEL,demonstrated the role of apoptosis in the disappearance ofsmooth-muscle cells during the development and growth ofexperimental cerebral aneurysms in rats. Furthermore, evi-dence of apoptosis in intracranial aneurysms has been dem-onstrated using TUNEL in four postmortem cases.14 Sakaki,et al.,31 found that ruptured aneurysm walls exhibited well-defined evidence of DNA fragmentation, which indicatesapoptosis. The object of the present study was to verify thehypothesis that apoptotic phenomena are relevant in rup-tured intracranial aneurysms and that they may representthe local expression of more extensive vascular enduranceinvolving the entire arterial system. For this reason wefocused on the evaluation of apoptotic phenomena ex vivoin the domes of ruptured and unruptured intracranial aneu-rysms, in arterial specimens located outside the subarach-noid spaces—namely the MMA or the STA obtained inpatients with ruptured and unruptured intracranial aneu-rysms—and in specimens of MMA and STA from patientssurgically treated for nonvascular pathologies. The TUNELwas used for the quantitative analysis of apoptotic cells. Todetermine the type of apoptotic mechanisms involved andthe primary reasons for aneurysm growth and rupture, westudied apoptotic phenomena by using electron microscopyas well.
Materials and MethodsBetween September 1999 and February 2002, specimens from 27
consecutive cases of intracranial aneurysms were collected duringsurgery at our neurosurgical center. Exclusion from the study de-pended on purely surgical reasons, namely when no specimen wascollectable (for example, when reaching the aneurysm required co-agulation of the aneurysm sac or the positioning of many aneurysmclips that actually covered the entire aneurysm sac) and when the siteof the aneurysm sac rendered its removal unacceptable for the pa-tient’s safety. In all collected specimens, the sampling site corre-sponded to the aneurysm dome.
Aneurysm specimens were obtained in nine men (33%) and 18women (67%), whose median age was 57.1 years (range 29–78years). Seventeen aneurysms (63%) were ruptured and 10 were un-ruptured (37%). All patients underwent routine preoperative four-vessel angiography.
In 13 (seven with ruptured and six with unruptured lesions) of the27 patients, a specimen of the MMA or the STA was also collected.In seven patients surgically treated via pterional craniotomy for braintumor (five patients) and subacute subdural hematoma (two pa-tients)—not aneurysm or vascular malformation pathologies—spec-imens of the same arteries were obtained as controls. These proce-dures were performed because they were judged completely safe.
Only two patients with ruptured aneurysms from among all thosewith ruptured or unruptured aneurysms and all control volunteersneeded intensive care unit admission before surgery.
All aneurysms and arterial specimens were studied using TUNELand electron microscopy. Specimens were immediately pretreated inthe operating room, eliminating from their surface blood clots and/orother biological materials that could invalidate a pure study of the an-eurysm dome.
For the TUNEL, obtained tissues were immediately fixed in 4%buffered formalin for temporary storage and were then processed.The molecular biological–histochemical method used was both sen-sitive and specific for the detection of DNA fragmentation in aneu-rysm walls. This technique, known as “TUNEL,”11,20 is useful for insitu identification of programmed cell death, permitting quantifica-tion of apoptosis in cell populations. The technique is based on thespecific binding of terminal deoxynucleotidyl transferase to 39-OHends of DNA and the consequent in situ synthesis of a polydeoxy-nucleotide polymer. The new 39-OH DNA ends generated by DNAfragmentation are the labeled targets as they are specifically locatedin morphologically identifiable nuclei and apoptotic bodies in cellsconstituting the aneurysm wall. Apoptotic cells are those containingapoptotic bodies.11 The manufacturer’s instructions for the apoptosisdetection kit (Vasotacs kit, Gaithersburg, MD) were followed. Fiveto seven slides per aneurysm were examined using light microscopy.Three separate observers conducted a double-blind overall evalua-tion of the slides. These observers were also unaware of whether thespecimen had been obtained in a patient harboring a ruptured or anunruptured aneurysm or in a control volunteer.
Apoptotic cells were defined by the presence of TUNEL-positivenuclei.14,28,31,38 The number of apoptotic cells in the high-power (3400) field was counted in different areas of the specimen and ex-pressed as a proportion of the total number of cells observed in thefield. The major drawback of TUNEL is that cell labeling is not spe-cific for apoptosis. In fact, a positive TUNEL reaction occurs in allapoptotic cells but also in cells subjected to DNA repair and necro-sis16,27; therefore, in the present study, we used electron microscopyto confirm apoptosis in ruptured and unruptured aneurysms and inspecimens obtained in control volunteers.
A scale to quantify apoptosis in aneurysm tissue was developedin cooperation with the Department of Epidemiology and MedicalStatistics at our university. We evaluated the specimens as follows:Class A4 (Fig. 1 left), numerous diffuse apoptotic cells in all ob-served microscopic fields (. 75%); Class A3 (Fig. 1 center), mas-sive focal areas of apoptotic cells, which tended to localize near thevessel’s lumen, mixed with apparently normal zones (from . 50 to75%); Class A2, presence of apoptotic cells (from 15 to 50%); ClassA1, modest number of apoptotic cells in all observed microscopicfields (, 15%); Class A0 (Fig. 1 right), absence of apoptosis or spo-radic presence apoptotic cells.
For electron microscopy, tissue samples of the aneurysm domesand the arteries were fixed overnight in cold (4˚C) Tyrode buffercontaining 2% glutaraldehyde. After rinsing with cold buffer for atleast 1 hour, tissues were postfixed for 1 to 2 hours in 1% osmiumtetroxide in cold Tyrode buffer and rinsed for 30 to 60 minutes.Specimens were dehydrated through graded (70, 95, and 100%) eth-anol for 30 minutes for each cycle at room temperature, incubatedtwice in propylene oxide (30 minutes for each cycle), and infiltratedovernight in a 1:1 mixture of propylene oxide/Polybed 812 resin.Tissues were embedded in Polybed 812 resin blocks for 1 hour atroom temperature, incubated overnight at 45˚C, and sectioned aftercooling at room temperature. Thin slices were cut using a diamondknife, placed on Formvar-coated copper grids, and examined on atransmission electron microscope (CM10; Philips, Eindhoven, TheNetherlands).
Differences in apoptosis related to variables, such as the patient’ssex and age and the aneurysm’s shape and dimensions, were ana-lyzed to establish predictive factors that might correlate with the de-velopment and rupture of an aneurysm incidentally diagnosed.
The chi-square test was used in statistical analyses of our results.Probability values less than 0.05 were considered statistically signif-icant.
ResultsThe TUNEL
Aneurysms. Results of the light microscopy evaluation(TUNEL) showed relevant apoptosis levels (Classes A3–A4) in 15 (88%) of 17 ruptured aneurysm walls (Table 1);the other two ruptured aneurysms appeared to be ClassesA0–A1. In all unruptured aneurysms, apoptosis was absent
J. Neurosurg. / Volume 101 / December, 2004
Apoptosis in intracranial aneurysms
1019
12_04_JNS.2 11/22/04 11:03 AM Page 1019

or scarce except in one case, which, interestingly, involveda patient harboring another aneurysm that had previouslybled and whose analysis showed elevated apoptosis (Class-es A3–A4). In this same case, a specimen from the MMAshowed Classes A3–A4 apoptosis (Fig. 2). Analyses of thedistribution of apoptosis in ruptured compared with unrup-tured aneurysms was statistically significant (p , 0.001;Table 2).
Arterial Walls. High apoptosis levels (Classes A3–A4)were detected in all seven arterial specimens (STA orMMA) obtained in patients harboring ruptured aneurysms(Table 3). In five unruptured aneurysms, very low or absentapoptosis levels were detected in the STA or MMA. In onlyone specimen (Case 22), the elevated apoptosis level in theperipheral artery did not match the condition in the unrup-tured aneurysm. No arterial specimen obtained in patientswithout an aneurysm showed the presence of apoptosis.These results are highly statistically significant (p , 0.001).
Surgical Timing and Apoptosis
In this study we evaluated the possibly variable and un-accounted effect of hemorrhage surrounding the aneurysm
dome as it related to the timing of surgery. The cases weresubdivided into two groups: Group 1 consisted of aneu-rysms surgically treated within 72 hours after SAH, andGroup 2 consisted of lesions surgically treated more than10 days after SAH. Analysis of our results indicated thatthere was no statistical difference between the two groups interms of apoptosis (Table 4).
Aneurysm Shape and Volume and Apoptosis
On the basis of angiographic evidence matched with theintraoperative appearance, aneurysms were classified as fol-lows: 1) regularly shaped (that is, round or oval), 16 cases(60%); and 2) irregularly shaped (that is, multilobate anduneven shapes), 11 cases (40%; Table 1). Of the 16 regular-ly shaped aneurysms, nine (56%) were unruptured and sev-en (44%) were ruptured. Levels of apoptosis were high insix aneurysms (38%) and low in 10 (62%). Of 11 irregular-ly shaped aneurysms, 10 (91%) were ruptured and only one(9%) was unruptured; apoptosis levels were elevated in allof the ruptured aneurysms and was absent in the irregular-ly shaped unruptured lesion. A statistically significant rela-tionship (p , 0.05) between aneurysm shape and apoptosislevel was found (Fig. 3). Among 19 aneurysms smaller than1 cm, 10 (53%) were associated with an SAH and demon-strated elevated levels of apoptosis in the walls. One aneu-rysm (5%) was unruptured and showed increased levels ofapoptosis. Eight lesions (42%) were unruptured and showedlow or absent levels of apoptosis. Of eight aneurysms larg-er than 1 cm, seven (87%) ruptured; only five had elevatedlevels of apoptosis. The statistical evaluation of these datademonstrated no significant differences in apoptosis levelsand aneurysm volumes (Table 5).
Patient Age and Sex and Apoptosis
Results of statistical analysis showed no significant cor-relation between patient age and sex and the level of apop-tosis.
Electron Microscopy
Results of ultrastructural analysis revealed complete dis-organization in all of the wall layers in ruptured aneurysms.
L. Pentimalli, et al.
1020 J. Neurosurg. / Volume 101 / December, 2004
FIG. 1. Photomicrographs showing apoptosis—indicated by the darkly marked cell nuclei—in the aneurysm wall fol-lowing TUNEL. Left: Ruptured aneurysm wall with massive and diffuse presence of apoptosis (Class A4). Center:Ruptured aneurysm wall with massive focal areas of apoptosis mixed with apparently normal zones. Apoptotic regionstended to localize near the vessel’s lumen (Class A3). Right: Unruptured aneurysm wall with no apoptosis (Class A0).H & E, original magnification 3 10 (B), 3 20 (A and C).
FIG. 2. Photomicrograph demonstrating Class A4 apoptosis in aspecimen from the MMA. H & E, original magnification 3 10.
12_04_JNS.2 11/22/04 11:03 AM Page 1020
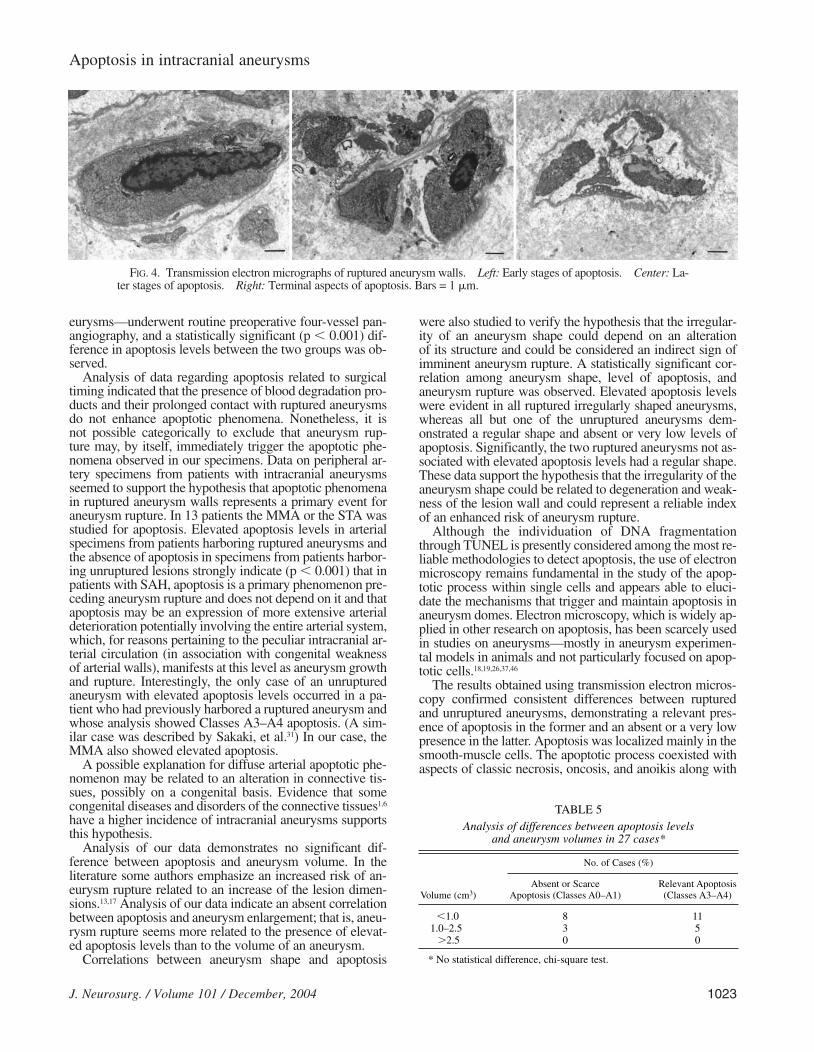
Smooth-muscle cells from ruptured aneurysms showed acomplete sequence of the apoptotic process, and electron-dense and shriveled cytoplasm was observed together withcondensation of mitochondria and disorganization of cyto-skeletal architecture. During the early stages of apoptosis(Fig. 4 left), progressive aggregation of chromatin was not-ed in the nuclei. Later stages of apoptosis (Fig. 4 center)were characterized by a more evident aggregation of nucle-ar chromatin along with a progressive reduction in the cy-toplasm mass, presence of cytoplasm vacuoles, and disor-ganization of contractile filaments. At this apoptotic stage,we also observed partial detachment of the cells from theirbasal membranes.
A progressive reduction in cellular volume caused by thedecrease in cytoplasm was evident throughout the entireapoptotic process. Swollen mitochondria and the presenceof apoptotic bodies were also peculiar to the late stages ofapoptosis. Finally, nuclei with condensed chromatin havinga doughnut or horseshoe shape and progressive disorgani-zation of the entire cell body were detectable in smooth-muscle cell layers (Fig. 4 right). The apoptotic process was
also apparent in fibroblasts residing in the stroma of rup-tured aneurysms (Fig. 5 left). The half-moon shape formedby the accumulation of the euchromatin on the nuclearmembrane is characteristic of the apoptotic process (Fig. 5right).
Fibroblasts in ruptured aneurysms showed progressiveaggregation of nuclear chromatin and different aspects ofnuclear pyknosis. Moreover, they displayed intense cyto-
J. Neurosurg. / Volume 101 / December, 2004
Apoptosis in intracranial aneurysms
1021
TABLE 1Apoptosis levels in 27 aneurysms and 20 arterial specimens*
Aneurysms Arteries
Shape Apoptosis Level Apoptosis Level
Case No. Status Vol (cm3) RS IS Classes A0–A1 Classes A3–A4 Classes A0–A1 Classes A3–A4
1 R 1.0–2.5 1 1 12 R 1.0–2.5 1 1 13 R ,1.0 1 1 14 R ,1.0 1 15 R ,1.0 1 16 R 1.0–2.5 1 17 R ,1.0 1 1 18 R ,1.0 1 1 19 R 1.0–2.5 1 1 1
10 R 1.0–2.5 1 111 R 1.0–2.5 1 112 R 1.0–2.5 1 113 R ,1.0 1 114 R ,1.0 1 115 R ,1.0 1 116 R ,1.0 1 117 R ,1.0 1 117† R ,1.0 1 118 U ,1.0 1 1 119 U ,1.0 1 1 120 U ,1.0 1 1 121 U ,1.0 1 122 U ,1.0 1 1 123 U ,1.0 1 1 124 U ,1.0 1 125 U 1.0–2.5 1 126 U ,1.0 1 127 U ,1.0 1 127† U ,1.0 1 128 C 129 C 130 C 131 C 132 C 133 C 134 C 1
* C = control volunteer, so not applicable; IS = irregular shape; R = ruptured; RS = regular shape; U = unruptured; Vol = volume. † Arterial specimen only.
TABLE 2Analysis of the distribution of apoptosis in ruptured
compared with unruptured aneurysms*
No. of Cases (%)
Aneurysm No. of Absent or Scarce Relevant ApoptosisType Cases Apoptosis (Classes A0–A1) (Classes A3–A4)
ruptured 17 2 (12) 15 (88)unruptured 10 9 (90) 1 (10)
* p , 0.001, chi-square test.
12_04_JNS.2 11/22/04 11:03 AM Page 1021

plasmic disorganization characterized by the presence ofvacuolization, reduced volume, swollen mitochondria, andrupture of the plasma membrane. Cytoplasm vacuoles ofdifferent dimensions, lipid droplets, dilated nuclear mem-branes, and nuclear pores were observed, as opposed to thenormal morphological features of fibroblasts in the stro-ma of unruptured aneurysms. Note that apoptosis often wasoverlapped by or coexisted with morphological features ofnecrosis, which was characterized by cells with swelling cy-toplasm, ruptured plasma membrane, and dispersed cellularorganelles.
In ruptured aneurysms, analysis of extracellular matrixorganization revealed partial degradation of collagen fibers.Signs of this degradation are electrolucent areas locatedin the stroma very close to remodeling extracellular matrixzones, which are characterized by thinner collagen fiberswith a different orientation (Fig. 6). Few inflammatorycells—mostly granulocytes and macrophages, with differ-ent degrees of activation—were visible in the stroma.
The final aspect of apoptosis was characterized by chro-matin agglomerates, basal membrane rupture, and cyto-plasm degeneration.
Results of the ultrastructural study in the majority of un-ruptured aneurysms, in accordance with the data obtainedusing TUNEL, showed no evidence of apoptosis.
Discussion
Vascular remodeling through apoptotic cell death playsan important role in vascular disease.15 Sakaki, et al.,31 ob-served DNA fragmentation in intracranial aneurysms inhumans. Henderson, et al.,15 reported apoptotic cell deathin the walls of abdominal aortic aneurysms. These find-ings indicate that apoptotic processes seem to contribute toaneurysms. Note that the aneurysms were studied usingTUNEL. Although many apoptotic cells were detected in
the domes of ruptured intracranial aneurysms, a quantitativeevaluation of apoptotic phenomena was not described andmultiple variables were not related to apoptosis. Moreover,the literature consists of no studies aimed at clarifying therole of apoptosis as a primary cause rather than a conse-quence of aneurysm rupture. Our study targeted such re-search and we endeavored to analyze the ultimate role ofapoptotic phenomena in the pathogenesis of intracranial an-eurysm rupture. For this reason, differences in apoptoticphenomena in the walls of ruptured compared with unrup-tured intracranial lesions were investigated. Apoptosis inthe peripheral arteries was also studied in these same pa-tients and in those affected by nonvascular pathological en-tities.
In the present study we demonstrated apoptotic phe-nomena in aneurysm walls through the use of TUNEL andelectron microscopy. It was not difficult to differentiateapoptotic from necrotic cells, based on data already report-ed in the literature.8,32,41,44
The thin walls of ruptured aneurysm domes appear to becomposed of spindle-shaped fibrous cells and numerousapoptotic cells. The number of these cells is greater nearthe rupture site. In the aneurysm dome, the cells contain-ing apoptotic bodies appear clearly differentiated from sur-rounding areas and submerged by abundant fibrous ma-terial.
The typical degeneration or absence of the internal elas-tic membrane along with apoptosis in the smooth-musclecells in the aneurysm dome could indicate a deterioration ofthe aneurysm wall that may secondarily cause its rupture.
Statistical analyses of our data revealed a clear, high cor-relation (p , 0.001) for the association between high levelsof apoptosis (Classes A3–A4) and aneurysm rupture, indi-cating that elevated apoptosis may cause weakness in andrupture of aneurysm walls. In two (12%) of 17 cases, how-ever, rupture occurred independent of elevated levels ofapoptosis in the aneurysm walls, probably due to factors re-lated to hemodynamic variations. Elevated apoptosis levels(Classes A3–A4) were also detected in one of 10 unrup-tured aneurysms, which could indicate that only one of theten unruptured aneurysms that had been surgically treatedwas really at risk for rupture.
We excluded the possibility that angiographic contrastmedia could have induced apoptosis because all of our pa-tients—those with ruptured and those with unruptured an-
L. Pentimalli, et al.
1022 J. Neurosurg. / Volume 101 / December, 2004
TABLE 3Apoptosis levels in arterial specimens in 20 cases*
No. of Cases (%)
Absent or Scarce RelevantNo. of Apoptosis Apoptosis
Artery Origin Cases (Classes A0–A1) (Classes A3–A4)
patient w/ ruptured aneurysm 7 0 (0) 7 (100)patient w/ unruptured artery 6 5 (83) 1 (17)control volunteers 7 7 (100) 0 (0)
* p , 0.001, chi-square test.
TABLE 4Analysis of surgical timing and apoptosis in 17 cases*
No. of Cases (%)
Absent or Scarce Relevant ApoptosisSurgery Apoptosis (Classes A0–A1) (Classes A3–A4)
w/in 72 hrs after SAH 0 6.10 days after SAH 2 9
* No statistical difference, chi-square test.
FIG. 3. Bar graph depicting the relationship between aneurysmshape and apoptosis. A statistically significant correlation was found(p , 0.05, chi-square test). Black bars represent the presence ofapoptosis and hatched bars its absence.
12_04_JNS.2 11/22/04 11:03 AM Page 1022
eurysms—underwent routine preoperative four-vessel pan-angiography, and a statistically significant (p , 0.001) dif-ference in apoptosis levels between the two groups was ob-served.
Analysis of data regarding apoptosis related to surgicaltiming indicated that the presence of blood degradation pro-ducts and their prolonged contact with ruptured aneurysmsdo not enhance apoptotic phenomena. Nonetheless, it isnot possible categorically to exclude that aneurysm rup-ture may, by itself, immediately trigger the apoptotic phe-nomena observed in our specimens. Data on peripheral ar-tery specimens from patients with intracranial aneurysmsseemed to support the hypothesis that apoptotic phenomenain ruptured aneurysm walls represents a primary event foraneurysm rupture. In 13 patients the MMA or the STA wasstudied for apoptosis. Elevated apoptosis levels in arterialspecimens from patients harboring ruptured aneurysms andthe absence of apoptosis in specimens from patients harbor-ing unruptured lesions strongly indicate (p , 0.001) that inpatients with SAH, apoptosis is a primary phenomenon pre-ceding aneurysm rupture and does not depend on it and thatapoptosis may be an expression of more extensive arterialdeterioration potentially involving the entire arterial system,which, for reasons pertaining to the peculiar intracranial ar-terial circulation (in association with congenital weaknessof arterial walls), manifests at this level as aneurysm growthand rupture. Interestingly, the only case of an unrupturedaneurysm with elevated apoptosis levels occurred in a pa-tient who had previously harbored a ruptured aneurysm andwhose analysis showed Classes A3–A4 apoptosis. (A sim-ilar case was described by Sakaki, et al.31) In our case, theMMA also showed elevated apoptosis.
A possible explanation for diffuse arterial apoptotic phe-nomenon may be related to an alteration in connective tis-sues, possibly on a congenital basis. Evidence that somecongenital diseases and disorders of the connective tissues1,6
have a higher incidence of intracranial aneurysms supportsthis hypothesis.
Analysis of our data demonstrates no significant dif-ference between apoptosis and aneurysm volume. In theliterature some authors emphasize an increased risk of an-eurysm rupture related to an increase of the lesion dimen-sions.13,17 Analysis of our data indicate an absent correlationbetween apoptosis and aneurysm enlargement; that is, aneu-rysm rupture seems more related to the presence of elevat-ed apoptosis levels than to the volume of an aneurysm.
Correlations between aneurysm shape and apoptosis
were also studied to verify the hypothesis that the irregular-ity of an aneurysm shape could depend on an alterationof its structure and could be considered an indirect sign ofimminent aneurysm rupture. A statistically significant cor-relation among aneurysm shape, level of apoptosis, andaneurysm rupture was observed. Elevated apoptosis levelswere evident in all ruptured irregularly shaped aneurysms,whereas all but one of the unruptured aneurysms dem-onstrated a regular shape and absent or very low levels ofapoptosis. Significantly, the two ruptured aneurysms not as-sociated with elevated apoptosis levels had a regular shape.These data support the hypothesis that the irregularity of theaneurysm shape could be related to degeneration and weak-ness of the lesion wall and could represent a reliable indexof an enhanced risk of aneurysm rupture.
Although the individuation of DNA fragmentationthrough TUNEL is presently considered among the most re-liable methodologies to detect apoptosis, the use of electronmicroscopy remains fundamental in the study of the apop-totic process within single cells and appears able to eluci-date the mechanisms that trigger and maintain apoptosis inaneurysm domes. Electron microscopy, which is widely ap-plied in other research on apoptosis, has been scarcely usedin studies on aneurysms—mostly in aneurysm experimen-tal models in animals and not particularly focused on apop-totic cells.18,19,26,37,46
The results obtained using transmission electron micros-copy confirmed consistent differences between rupturedand unruptured aneurysms, demonstrating a relevant pres-ence of apoptosis in the former and an absent or a very lowpresence in the latter. Apoptosis was localized mainly in thesmooth-muscle cells. The apoptotic process coexisted withaspects of classic necrosis, oncosis, and anoikis along with
J. Neurosurg. / Volume 101 / December, 2004
Apoptosis in intracranial aneurysms
1023
TABLE 5Analysis of differences between apoptosis levels
and aneurysm volumes in 27 cases*
No. of Cases (%)
Absent or Scarce Relevant ApoptosisVolume (cm3) Apoptosis (Classes A0–A1) (Classes A3–A4)
,1.0 8 111.0–2.5 3 5
.2.5 0 0
* No statistical difference, chi-square test.
FIG. 4. Transmission electron micrographs of ruptured aneurysm walls. Left: Early stages of apoptosis. Center: La-ter stages of apoptosis. Right: Terminal aspects of apoptosis. Bars = 1 mm.
12_04_JNS.2 11/22/04 11:03 AM Page 1023

the presence of extracellular matrix degradation and basalmembrane detachment. The ultrastructural features of apop-totic cells revealed early cytoplasm modification, whichmay be related both to the effects of the apoptotic stimulusand to a lack of energy necessary to complete the apoptoticprocess.
In the wall of ruptured aneurysms it was possible to iden-tify myoepithelial cells demonstrating relevant features ofapoptosis in the smooth-muscle tissue. The different phasesof the apoptotic process were observed. Such features al-so appeared in unruptured aneurysms, although more rarelyand more sparsely. This common pathway of myoepithelialcellular apoptotic degeneration, detected in both rupturedand unruptured aneurysm walls, strongly indicates that theapoptotic features observed must be regarded as precedinganeurysm rupture and do not constitute a simple epiphe-nomenon subsequent to aneurysm rupture. These observa-tions support a possible authentic role of apoptotic phenom-ena in the progressive weakness and rupture of aneurysmwalls.
Cellular alterations together with extracellular matrix re-modeling and detachment of smooth-muscle cells from thebasal membrane, which were detected on electron micros-copy, could play a fundamental role in the growth and rup-ture of aneurysms.
The association between relevant apoptotic features andaneurysm rupture indicates that these events may be relatedto the rupture of aneurysms.
Conclusions
Mechanisms underlying intracranial aneurysm growthand rupture remain unclear. Presently, we are unable defin-itively to define the real role played by apoptosis in an-eurysm walls, although our observations revealed that theapoptotic phenomenon constitutes an active element andpossibly plays a fundamental part in reducing the resistanceof aneurysm walls, thus contributing to the genesis of theirdilation and rupture.
Data collected in relation to the peripheral arteries indi-cated that apoptosis is a primary phenomenon involving theentire arterial system more diffusely and precedes aneurysmrupture and does not depend on it. Although these observa-tions are preliminary, if they are confirmed by additionaldata, they may support a role for apoptosis as a parameterfor predicting the risk of rupture in incidentally diagnosedaneurysms and in multiple aneurysms. Moreover, the mod-ulation of apoptosis—already being studied in oncology—could offer innovative therapeutic opportunities for intra-cranial aneurysms.
References
1. Baker CJ, Fiore A, Connolly ES Jr, et al: Serum elastase and al-pha-1-antitrypsin in ruptured and unruptured cerebral aneurysms.Neurosurgery 37:56–62, 1995
2. Bennett MR, Evan GI, Newby AC: Deregulated expression of thec-myc oncogene abolishes inhibition of proliferation of rat vascu-lar smooth muscle cells by serum reduction, interferon-g, heparin,and cyclic nucleotide analogues and induces apoptosis. Circ Res74:525–536, 1994
3. Bennett MR, Evan GI, Schwartz SM: Apoptosis of rat vascularsmooth muscle cells is regulated by p53-dependent and -indepen-dent pathways. Circ Res 77:266–273, 1995
4. Bjorkerud S, Bjorkerud B, Joelsson M: Structural organization ofreconstituted human arterial smooth muscle tissue. ArteriosclerThromb 14:644–651, 1994
5. Bonita R: Cigarette smoking, hypertension and the risk of sub-arachnoid hemorrhage: a population-based case-control study.Stroke 17:831–835, 1986
L. Pentimalli, et al.
1024 J. Neurosurg. / Volume 101 / December, 2004
FIG. 5. Transmission electron micrographs of ruptured aneurysm walls. Left: Apoptosis in fibroblasts in the stroma,indicated by progressive aggregation of nuclear chromatin. Right: Half-moon shape determined by the accumulation ofthe euchromatin on the nuclear membrane and characteristic of the apoptotic process. Bar = 1 mm (left), 0.5 mm (right).
FIG. 6. Transmission electron micrograph showing extracellularmatrix organization in a ruptured aneurysm. Bar = 0.5 mm.
12_04_JNS.2 11/22/04 11:03 AM Page 1024

6. Child AH: Marfan syndrome—current medical and geneticknowledge: how to treat and when. J Card Surg 12 (Suppl 2):131–136, 1997
7. Cho A, Courtman DW, Langille BL: Apoptosis (programmed celldeath) in arteries of the neonatal lamb. Circ Res 76:168–175,1995
8. Cohen JJ: Apoptosis. Immunol Today 14:126–130, 19939. Ferguson GG: Physical factors in the initiation, growth and rup-
ture of Human intracranial saccular aneurysms. J Neurosurg 37:666–677, 1972
10. Gabow PA: Autosomal dominant polycystic kidney disease. NEngl J Med 329:332–342, 1993
11. Gavrieli Y, Sherman Y, Ben-Sasson SA: Identification of pro-grammed cell death in situ via specific labeling of nuclear DNAfragmentation. J Cell Biol 119:493–501, 1992
12. Geng YJ, Libby P: Evidence for apoptosis in advanced humanatheroma. Colocalization with interleukin-1 beta-converting en-zyme. Am J Pathol 147:251–266, 1995
13. Hademenos GJ, Massoud TF, Turjman F, et al: Anatomical andmorphological factors correlating with rupture of intracranial an-eurysms in patients referred for endovascular treatment. Neurora-diology 40:755–760, 1998
14. Hara A, Yoshimi N, Mori H: Evidence for apoptosis in humanintracranial aneurysms. Neurol Res 20:127–130, 1998
15. Henderson EL, Geng YJ, Sukhova GK, et al: Death of smoothmuscle cells and expression of mediators of apoptosis by T lym-phocytes in human abdominal aortic aneurysms. Circulation 99:96–104, 1999
16. Hughes SE: Detection of apoptosis using in situ markers for DNAstrand breaks in the failing human heart. Fact or epiphenomenon?J Pathol 201:181–186, 2003
17. Juvela S, Porras M, Poussa K: Natural history of unruptured intra-cranial aneurysms: probability of and risk factors for aneurysmrupture. J Neurosurg 93:379–387, 2000
18. Kim C, Cervos-Navarro J, Kikuchi H, et al: Alterations in cerebralvessels in experimental animals and their possible relationship tothe development of aneurysms. Surg Neurol 38:331–337, 1992
19. Kojima M, Handa H, Hashimoto N, et al: Early changes of exper-imentally induced aneurysms in rats: scanning electron micro-scopic study. Stroke 17:835–841, 1986
20. Kondo S, Hashimoto N, Kikichi H, et al: Apoptosis of medialsmooth muscle cell in the development of saccular cerebral aneu-rysms in rats. Stroke 29:181–189, 1998
21. Kondo S, Ishizaka Y, Okada T, et al: FADD gene therapy formalignant gliomas in vitro and in vivo. Hum Gene Ther 9:1599–1608, 1998
22. Kontogeorgos G, Sambaziotis D, Piaditis G, et al: Apoptosis inhuman pitutary adenomas: a morphologic and in situ end-labelingstudy. Mod Pathol 10:921–926, 1997
23. Lopez-Candales A, Holmes DR, Liao S, et al: Decreased vascularsmooth muscle cell density in medial degeneration of human ab-dominal aortic aneurysms. Am J Pathol 150:993–1007, 1997
24. Lozano AM, Leblanc R: Familial intracranial aneurysms. J Neu-rosurg 66:522–528, 1987
25. Lyons SK, Clarke AR: Apoptosis and carcinogenesis. Br MedBull 53:554–569, 1997
26. Nagata I, Handa H, Hasimoto N, et al: Experimentally inducedcerebral aneurysms in rats: VII. Scanning electron microscopestudy. Surg Neurol 16:291–296, 1981
27. Otsuki Y, Li Z, Shibata MA: Apoptotic detection methods—frommorphology to gene. Prog Histochem Cytochem 38:275–339,2003
28. Qureshi AI, Suri MFK, Ostrow PT, et al: Apoptosis as a formof cell death in intracerebral hemorrhage. Neurosurgery 52:1041–1048, 2003
29. Riccioni T, Cirielli C, Wang X, et al: Adenovirus-mediated wild-type p53 overexpression inhibits endothelial cell differentiation invitro and angiogenesis in vivo. Gene Ther 5:747–754, 1998
30. Ronkainen A, Hernesniemi JA, Tromp G: Special features of fa-milial intracranial aneurysms: report of 215 familial aneurisms.Neurosurgery 37:43–47, 1995
31. Sakaki T, Kohmura E, Kishiguchi T, et al: Loss and apoptosis ofsmooth muscle cells in intracranial aneurysms. Studies with in situDNA end labeling and antibody against single-stranded DNA.Acta Neurochir 139:469–475, 1997
32. Sanderson CJ: Morphological aspects of lymphocyte mediated cy-totoxicity, in Clark WR, Golstein P (eds): Mechanisms of Cell-Mediated Cytotoxicity. New York: Plenum Press, 1982, pp 3–21
33. Savill J: Apoptosis in disease. Eur J Clin Invest 24:715–723,1994
34. Schievink WI: Genetics and aneurysm formation. NeurosurgClin N Am 9:485–495, 1998
35. Schievink WI, Schaid DJ, Rogers HM, et al: On the inheritance ofintracranial aneurisms. Stroke 25:2028–2037, 1994
36. Stehbens WE: Etiology of intracranial berry aneurysms. J Neuro-surg 70:823–831, 1989
37. Stehbens WE: The ultrastructure of experimental aneurysms inrabbits. Pathology 17:87–95, 1985
38. Takagi Y, Ishikawa M, Nozaki K, et al: Increased expression ofphosphorylated c-Jun amino-terminal kinase and phosphorylatedc-Jun in human cerebral aneurysms: role of the c-Jun amino-ter-minal kinase/c-Jun pathway in apoptosis of vascular walls. Neu-rosurgery 51:997–1004, 2002
39. Takenaka K, Yamakawa H, Sakai H, et al: Angiotensin I-convert-ing enzyme gene polymorphism in intracranial saccular aneurysmindividuals. Neurol Res 20:607–611, 1998
40. Ter Berg HWM, Dippel DWJ, Limburg M, et al: Familial intracra-nial aneurysms. A review. Stroke 23:1024–1030, 1992
41. Vaux DL: Toward an understanding of the molecular mecha-nisms of physiological cell death. Proc Natl Acad Sci USA 90:786–789, 1993
42. Williams GT, Smith CA: Molecular regulation of apoptosis: ge-netic controls on cell death. Cell 74:777–779, 1993
43. Wiman KG: New p53-based anti-cancer therapeutic strategies.Med Oncol 15:222–228, 1998
44. Wyllie AH: Cell death: a new classification separating apoptosisfrom necrosis, in Bowenn ID, Lockshin RA (eds): Cell Death inBiology and Pathology. London: Chapman and Hall, 1981, pp9–34
45. Wyllie AH, Morris RG, Smith AL, et al: Chromatin cleavage inapoptosis: association with condensed chromatin morphology anddependence on macromolecular synthesis. J Pathol 142:67–77,1984
46. Yamazoe N, Hashimoto N, Kikuchi H, et al: Elastic skeleton ofintracranial cerebral aneurysms in rats. Stroke 21:1722–1726,1990
47. Yuan JY, Horvitz HR: The Caenorhabditis elegans genes ced-3and ced-4 act cell autonomously to cause programmed cell death.Dev Biol 138:33–41, 1990
Manuscript received April 28, 2004.Accepted in final form July 30, 2004.This research was supported by the NANDO PERETTI Foun-
dation, Zurich, Switzerland, Associazione Terapie NeurochirurgicheAvanzate onlus, and the Institute of Neurosurgery, Catholic Univer-sity, Rome, Italy.
Address reprint requests to: Luigi Pentimalli, M.D., Ph.D., Istitutodi Neurochirurgia, Policlinico “A. Gemelli,” Largo Agostino Gem-elli 8, 00168 Roma, Italy. email: [email protected].
J. Neurosurg. / Volume 101 / December, 2004
Apoptosis in intracranial aneurysms
1025
12_04_JNS.2 11/22/04 11:03 AM Page 1025
Copyright © 2022 FDOKUMEN




















