
PACAP and VIP prevent apoptosis in schwannoma cells
7
Research Report PACAP and VIP prevent apoptosis in schwannoma cells Alessandro Castorina a,b , Adriana Tiralongo a , Salvatore Giunta a , Maria Luisa Carnazza a , Guido Rasi c , Velia D'Agata a,b, ⁎ a Department of Anatomy, Diagnostic Pathology, Legal Medicine, Hygiene and Public Health, University of Catania, Italy b Neuropharmacology PhD program, University of Catania, Catania, Italy c Institute of Neurobiology and Molecular Medicine ARTOV, CNR, Rome, Italy ARTICLE INFO ABSTRACT Article history: Accepted 8 September 2008 Available online 20 September 2008 Pituitary adenylate cyclase activating polypeptide (PACAP) and vasoactive intestinal peptide (VIP) are structurally endogenous peptides showing rich profile of biological activities. These peptides bind specific membrane receptors belonging to the superfamily of G protein- coupled receptors, the PAC1 and VPAC type receptors. Although these receptors have been identified in oligodendrocytes progenitors cells, to date the effects of PACAP and VIP in Schwann cells are still unknown. In the present study we investigated the expression of these neuropeptides as well as their receptors in a schwannoma cell line. RT-PCR and western blot analysis demonstrated that both PAC1 and VPAC2 receptors, but also PACAP peptide were expressed. To study the physiological effects mediated by PAC1/VPAC receptors, we evaluated their role in preventing apoptotic cell death induced by serum deprivation. Treatment with 100 nM PACAP38 and 100 nM VIP increased survival of serum- deprived schwannoma cells. Anti-apoptotic effects of these peptides were correlated to changes in BCL2 and BAX gene expression. Our results suggested that both PACAP38 and VIP could act as trophic factors in Schwann cells. © 2008 Elsevier B.V. All rights reserved. Keywords: PACAP VIP Apoptosis Schwannoma cells 1. Introduction Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) and Vasoactive Intestinal Polypeptide (VIP) are structurally related neuropeptides widely distributed in the central and peripheral nervous system (Vaudry et al., 2000). PACAP and VIP are potent activators of adenylyl cyclase (Arimura, 1998) and belong to a family of neuropeptides which include secretine and glucagone. They bind to specific membrane receptors which are classified into two types: PAC1 type (including at least eight different splice variants) and VPAC type (including VPAC1 and VPAC2 subtypes). PAC1 receptors preferable bind PACAP, recognizing poorly VIP. VPAC type receptors recognize similarly and with high affinity PACAP and VIP. PAC1/VPAC receptors comprise seven transmembrane domains and are coupled to G proteins (Zhou et al., 2002) which are associated with several transduction systems like adenylyl cyclase, phospholipase C (Arimura, 1998), Erk and MAPK pathways (Jamen et al., 2002). PAC1 and VPAC type receptors are involved in proliferation, differentiation and survival of normal and cancer cell lines. Previous reports suggested that PACAP exerts developmental actions. In fact, PACAP gene expression and PACAP immuno- reactivity are widely distributed in neurons within the embryonic and neonatal rat brain (Nielsen et al., 1998a, 1998b; Waschek et al., 1998; Skoglösa et al., 1999). Activation of PAC1 and VPAC type receptors has shown to regulate proli- BRAIN RESEARCH 1241 (2008) 29 – 35 ⁎ Corresponding author. Department of Anatomy, Diagnostic Pathology, Legal Medicine, Hygiene and Public Health, Via S. Sofia, 87, 95123 Catania, Italy. Fax: +39 095 3782046. E-mail address: [email protected] (V. D'Agata). 0006-8993/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2008.09.035 available at www.sciencedirect.com www.elsevier.com/locate/brainres
Transcript of PACAP and VIP prevent apoptosis in schwannoma cells

B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
PACAP and VIP prevent apoptosis in schwannoma cells
Alessandro Castorinaa,b, Adriana Tiralongoa, Salvatore Giuntaa, Maria Luisa Carnazzaa,Guido Rasic, Velia D'Agataa,b,⁎aDepartment of Anatomy, Diagnostic Pathology, Legal Medicine, Hygiene and Public Health, University of Catania, ItalybNeuropharmacology PhD program, University of Catania, Catania, ItalycInstitute of Neurobiology and Molecular Medicine ARTOV, CNR, Rome, Italy
A R T I C L E I N F O
⁎ Corresponding author. Department of AnatoCatania, Italy. Fax: +39 095 3782046.
E-mail address: [email protected] (V. D'Ag
0006-8993/$ – see front matter © 2008 Elsevidoi:10.1016/j.brainres.2008.09.035
A B S T R A C T
Article history:Accepted 8 September 2008Available online 20 September 2008
Pituitary adenylate cyclase activating polypeptide (PACAP) and vasoactive intestinal peptide(VIP) are structurally endogenous peptides showing rich profile of biological activities. Thesepeptides bind specific membrane receptors belonging to the superfamily of G protein-coupled receptors, the PAC1 and VPAC type receptors. Although these receptors have beenidentified in oligodendrocytes progenitors cells, to date the effects of PACAP and VIP inSchwann cells are still unknown. In the present study we investigated the expression ofthese neuropeptides as well as their receptors in a schwannoma cell line. RT-PCR andwestern blot analysis demonstrated that both PAC1 and VPAC2 receptors, but also PACAPpeptide were expressed. To study the physiological effects mediated by PAC1/VPACreceptors, we evaluated their role in preventing apoptotic cell death induced by serumdeprivation. Treatment with 100 nM PACAP38 and 100 nM VIP increased survival of serum-deprived schwannoma cells. Anti-apoptotic effects of these peptides were correlated tochanges in BCL2 and BAX gene expression. Our results suggested that both PACAP38 and VIPcould act as trophic factors in Schwann cells.
© 2008 Elsevier B.V. All rights reserved.
Keywords:PACAPVIPApoptosisSchwannoma cells
1. Introduction
PituitaryAdenylateCyclase-Activating Polypeptide (PACAP) andVasoactive Intestinal Polypeptide (VIP) are structurally relatedneuropeptides widely distributed in the central and peripheralnervous system (Vaudry et al., 2000). PACAP and VIP are potentactivators of adenylyl cyclase (Arimura, 1998) and belong to afamily of neuropeptideswhich include secretine andglucagone.
They bind to specific membrane receptors which areclassified into two types: PAC1 type (including at least eightdifferent splice variants) and VPAC type (including VPAC1 andVPAC2 subtypes). PAC1 receptors preferable bind PACAP,recognizing poorly VIP. VPAC type receptors recognize similarly
my, Diagnostic Pathology
ata).
er B.V. All rights reserved
and with high affinity PACAP and VIP. PAC1/VPAC receptorscomprise seven transmembrane domains and are coupled to Gproteins (Zhou et al., 2002) which are associated with severaltransduction systems like adenylyl cyclase, phospholipase C(Arimura, 1998), Erk and MAPK pathways (Jamen et al., 2002).
PAC1 andVPAC type receptors are involved in proliferation,differentiation and survival of normal and cancer cell lines.Previous reports suggested that PACAP exerts developmentalactions. In fact, PACAP gene expression and PACAP immuno-reactivity are widely distributed in neurons within theembryonic and neonatal rat brain (Nielsen et al., 1998a,1998b; Waschek et al., 1998; Skoglösa et al., 1999). Activationof PAC1 and VPAC type receptors has shown to regulate proli-
, Legal Medicine, Hygiene and Public Health, Via S. Sofia, 87, 95123
.
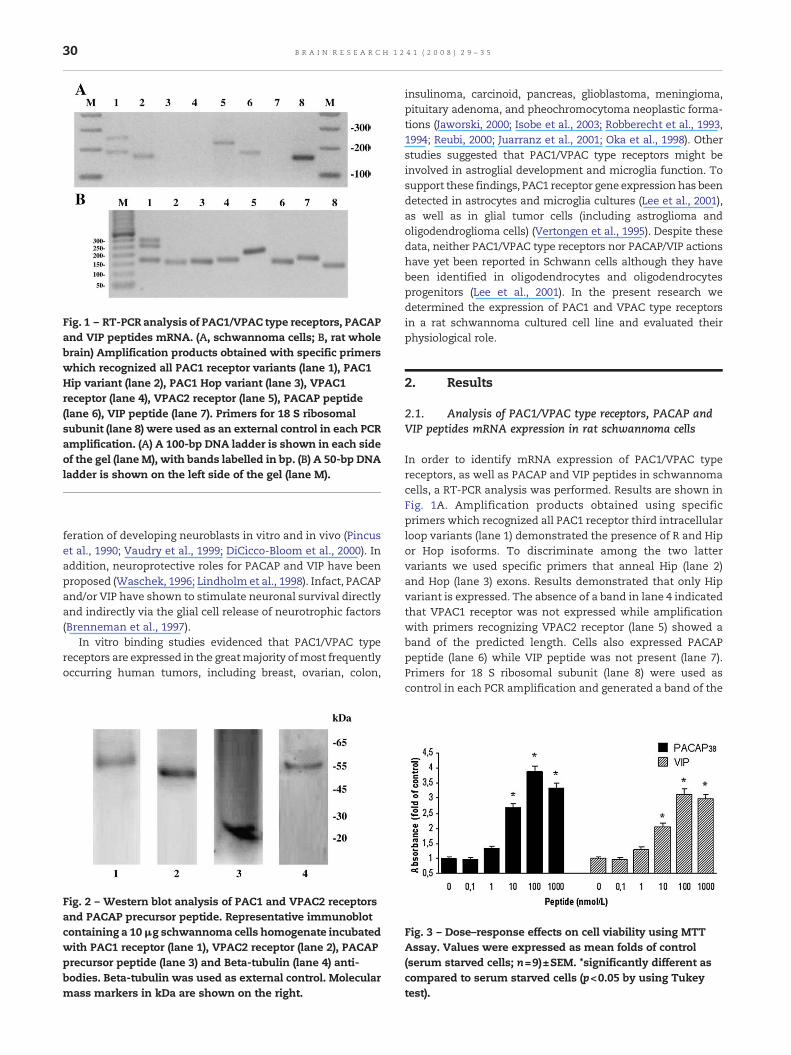
Fig. 1 – RT-PCR analysis of PAC1/VPAC type receptors, PACAPand VIP peptides mRNA. (A, schwannoma cells; B, rat wholebrain) Amplification products obtained with specific primerswhich recognized all PAC1 receptor variants (lane 1), PAC1Hip variant (lane 2), PAC1 Hop variant (lane 3), VPAC1receptor (lane 4), VPAC2 receptor (lane 5), PACAP peptide(lane 6), VIP peptide (lane 7). Primers for 18 S ribosomalsubunit (lane 8) were used as an external control in each PCRamplification. (A) A 100-bp DNA ladder is shown in each sideof the gel (lane M), with bands labelled in bp. (B) A 50-bp DNAladder is shown on the left side of the gel (lane M).
30 B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
feration of developing neuroblasts in vitro and in vivo (Pincuset al., 1990; Vaudry et al., 1999; DiCicco-Bloom et al., 2000). Inaddition, neuroprotective roles for PACAP and VIP have beenproposed (Waschek, 1996; Lindholm et al., 1998). Infact, PACAPand/or VIP have shown to stimulate neuronal survival directlyand indirectly via the glial cell release of neurotrophic factors(Brenneman et al., 1997).
In vitro binding studies evidenced that PAC1/VPAC typereceptors are expressed in the greatmajority ofmost frequentlyoccurring human tumors, including breast, ovarian, colon,
Fig. 2 – Western blot analysis of PAC1 and VPAC2 receptorsand PACAP precursor peptide. Representative immunoblotcontaining a 10μg schwannoma cells homogenate incubatedwith PAC1 receptor (lane 1), VPAC2 receptor (lane 2), PACAPprecursor peptide (lane 3) and Beta-tubulin (lane 4) anti-bodies. Beta-tubulin was used as external control. Molecularmass markers in kDa are shown on the right.
insulinoma, carcinoid, pancreas, glioblastoma, meningioma,pituitary adenoma, and pheochromocytoma neoplastic forma-tions (Jaworski, 2000; Isobe et al., 2003; Robberecht et al., 1993,1994; Reubi, 2000; Juarranz et al., 2001; Oka et al., 1998). Otherstudies suggested that PAC1/VPAC type receptors might beinvolved in astroglial development and microglia function. Tosupport these findings, PAC1 receptor gene expression has beendetected in astrocytes and microglia cultures (Lee et al., 2001),as well as in glial tumor cells (including astroglioma andoligodendroglioma cells) (Vertongen et al., 1995). Despite thesedata, neither PAC1/VPAC type receptors nor PACAP/VIP actionshave yet been reported in Schwann cells although they havebeen identified in oligodendrocytes and oligodendrocytesprogenitors (Lee et al., 2001). In the present research wedetermined the expression of PAC1 and VPAC type receptorsin a rat schwannoma cultured cell line and evaluated theirphysiological role.
2. Results
2.1. Analysis of PAC1/VPAC type receptors, PACAP andVIP peptides mRNA expression in rat schwannoma cells
In order to identify mRNA expression of PAC1/VPAC typereceptors, as well as PACAP and VIP peptides in schwannomacells, a RT-PCR analysis was performed. Results are shown inFig. 1A. Amplification products obtained using specificprimers which recognized all PAC1 receptor third intracellularloop variants (lane 1) demonstrated the presence of R and Hipor Hop isoforms. To discriminate among the two lattervariants we used specific primers that anneal Hip (lane 2)and Hop (lane 3) exons. Results demonstrated that only Hipvariant is expressed. The absence of a band in lane 4 indicatedthat VPAC1 receptor was not expressed while amplificationwith primers recognizing VPAC2 receptor (lane 5) showed aband of the predicted length. Cells also expressed PACAPpeptide (lane 6) while VIP peptide was not present (lane 7).Primers for 18 S ribosomal subunit (lane 8) were used ascontrol in each PCR amplification and generated a band of the
Fig. 3 – Dose–response effects on cell viability using MTTAssay. Values were expressed as mean folds of control(serum starved cells; n=9)±SEM. *significantly different ascompared to serum starved cells (p<0.05 by using Tukeytest).
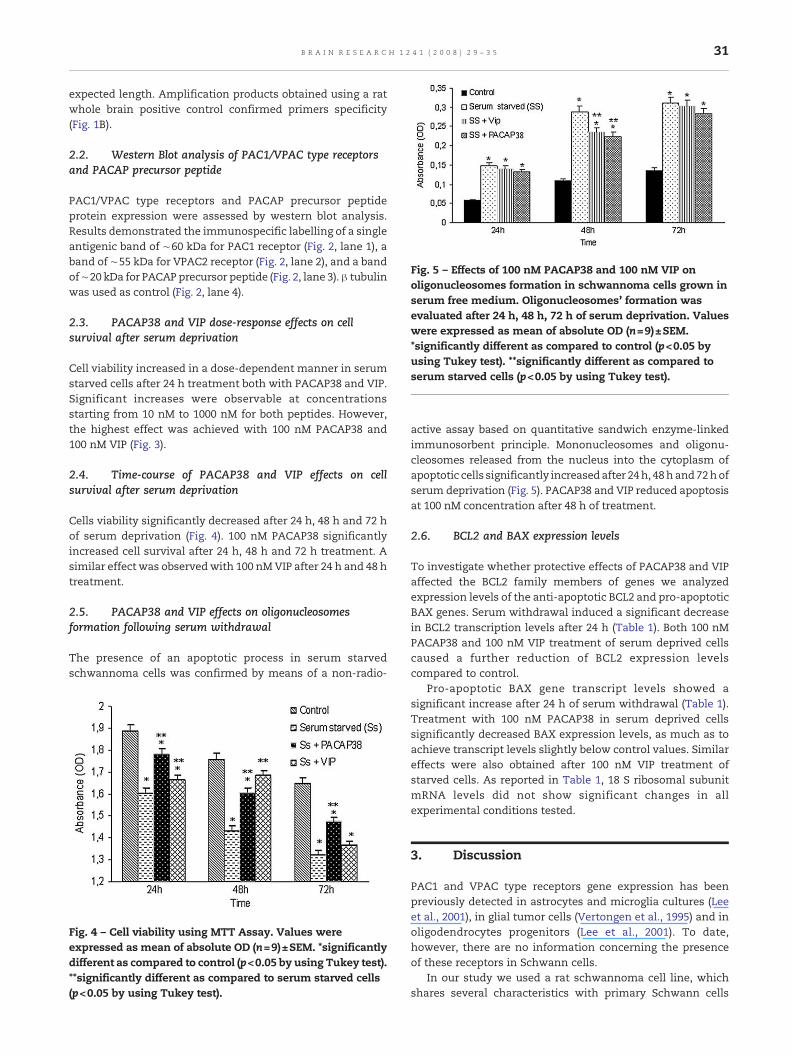
Fig. 5 – Effects of 100 nM PACAP38 and 100 nM VIP onoligonucleosomes formation in schwannoma cells grown inserum free medium. Oligonucleosomes' formation wasevaluated after 24 h, 48 h, 72 h of serum deprivation. Valueswere expressed as mean of absolute OD (n=9)±SEM.*significantly different as compared to control (p<0.05 byusing Tukey test). **significantly different as compared toserum starved cells (p<0.05 by using Tukey test).
31B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
expected length. Amplification products obtained using a ratwhole brain positive control confirmed primers specificity(Fig. 1B).
2.2. Western Blot analysis of PAC1/VPAC type receptorsand PACAP precursor peptide
PAC1/VPAC type receptors and PACAP precursor peptideprotein expression were assessed by western blot analysis.Results demonstrated the immunospecific labelling of a singleantigenic band of ∼60 kDa for PAC1 receptor (Fig. 2, lane 1), aband of ∼55 kDa for VPAC2 receptor (Fig. 2, lane 2), and a bandof∼20 kDa for PACAP precursor peptide (Fig. 2, lane 3). β tubulinwas used as control (Fig. 2, lane 4).
2.3. PACAP38 and VIP dose-response effects on cellsurvival after serum deprivation
Cell viability increased in a dose-dependent manner in serumstarved cells after 24 h treatment both with PACAP38 and VIP.Significant increases were observable at concentrationsstarting from 10 nM to 1000 nM for both peptides. However,the highest effect was achieved with 100 nM PACAP38 and100 nM VIP (Fig. 3).
2.4. Time-course of PACAP38 and VIP effects on cellsurvival after serum deprivation
Cells viability significantly decreased after 24 h, 48 h and 72 hof serum deprivation (Fig. 4). 100 nM PACAP38 significantlyincreased cell survival after 24 h, 48 h and 72 h treatment. Asimilar effect was observedwith 100 nMVIP after 24 h and 48 htreatment.
2.5. PACAP38 and VIP effects on oligonucleosomesformation following serum withdrawal
The presence of an apoptotic process in serum starvedschwannoma cells was confirmed by means of a non-radio-
Fig. 4 – Cell viability using MTT Assay. Values wereexpressed as mean of absolute OD (n=9)±SEM. *significantlydifferent as compared to control (p<0.05 by using Tukey test).**significantly different as compared to serum starved cells(p<0.05 by using Tukey test).
active assay based on quantitative sandwich enzyme-linkedimmunosorbent principle. Mononucleosomes and oligonu-cleosomes released from the nucleus into the cytoplasm ofapoptotic cells significantly increasedafter 24h, 48hand72hofserum deprivation (Fig. 5). PACAP38 and VIP reduced apoptosisat 100 nM concentration after 48 h of treatment.
2.6. BCL2 and BAX expression levels
To investigate whether protective effects of PACAP38 and VIPaffected the BCL2 family members of genes we analyzedexpression levels of the anti-apoptotic BCL2 and pro-apoptoticBAX genes. Serum withdrawal induced a significant decreasein BCL2 transcription levels after 24 h (Table 1). Both 100 nMPACAP38 and 100 nM VIP treatment of serum deprived cellscaused a further reduction of BCL2 expression levelscompared to control.
Pro-apoptotic BAX gene transcript levels showed asignificant increase after 24 h of serum withdrawal (Table 1).Treatment with 100 nM PACAP38 in serum deprived cellssignificantly decreased BAX expression levels, as much as toachieve transcript levels slightly below control values. Similareffects were also obtained after 100 nM VIP treatment ofstarved cells. As reported in Table 1, 18 S ribosomal subunitmRNA levels did not show significant changes in allexperimental conditions tested.
3. Discussion
PAC1 and VPAC type receptors gene expression has beenpreviously detected in astrocytes and microglia cultures (Leeet al., 2001), in glial tumor cells (Vertongen et al., 1995) and inoligodendrocytes progenitors (Lee et al., 2001). To date,however, there are no information concerning the presenceof these receptors in Schwann cells.
In our study we used a rat schwannoma cell line, whichshares several characteristics with primary Schwann cells

Table 1 – BCL2 and BAX expression levels after 100 nM PACAP38 and 100 nM VIP treatment in serum deprived cells
Gene Treatment Copy number/ng RNA+SEM
BCl2 Control 166,68±7,87Serum starved (SS) 130,80±6,82 ⁎
SS+PACAP 38 79,07±6,25⁎, ⁎⁎
SS+VIP 59,94±7,45⁎, ⁎⁎
BAX Control 2146,25±49,90Serum starved (SS) 2746,66±84,73 ⁎
SS+PACAP 38 1992,08±84,62 ⁎⁎
SS+VIP 1701,08±62,02⁎, ⁎⁎
18 S Control 628,33±49,81Serum starved (SS) 750,33±90,52SS+PACAP 38 613,33±37,26SS+VIP 629,66±40,11
Transcript levels are reported as the mean number of copies (n=5)/ng mRNA±SEM at 24 h.⁎ Significantly different as compared to control (p<0.05 by using Tukey test).⁎⁎ Significantly different as compared to serum starved cells (p<0.05 by using Tukey test).
32 B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
(Bansal and Pfeiffer, 1987; Hai et al., 2002). RT-PCR andwestern blot analysis demonstrated that this cell lineexpressed PAC1 receptor, in particular its transcript variantsPAC1-Hip and PAC1-R, VPAC2 receptor and PACAP precursorpeptide. We also evaluated the functional role mediated bythese receptors by testing their trophic action in a conditionof serum deprivation. Viability was reduced by serum with-drawal and cells underwent apoptosis. Previous reportsdemonstrated that PACAP38 and VIP exert a protective actionin different cell types (Lindholm et al., 1998; Onoue et al.,2002; Pincus et al., 1990; Vaudry et al., 2002). Such effect wasmaximal at 100 nM concentration for both peptides. Based onthese results we used 100 nM PACAP38 and 100 nM VIP toevaluate their role in schwannoma cell's survival. BothPACAP38 or VIP were able to increase cell viability and toreduce apoptotic cell death.
Programmed cell death is a physiological event that involvesactivation of several genes including BCL2 family members.They are highlighted both for their apoptosis-regulatingcapacity and also for their effect on cell cycle progression.Cumulative data indicate that cell cycle control is linked to celldeath regulation (Gil-Gomez et al., 1998; Huang et al., 1997;Vairo et al., 1996; Vaux et al., 1988). The relationship is complexand context dependent. BCL2 family members include a groupof genes encoding proteins that can either inhibit or promoteapoptosis. Among these genes BCL2 and BAX play a key role inregulating cell cycle and cell death.
BCL2andBAXtranscript productsaremitochondrialproteinwhich have the ability to form homo- and hetero-dimers. Thebalance betweenpro- and anti-apoptotic proteins is believed tobe the critical factor regulating apoptosis (Ge et al., 2004).
The BCL2 anti-apoptotic gene has parallel effects on celldeath and cell cycle inhibition by maintaining cell survival atthe expense of proliferation (Zinkel et al., 2006). The pro-apoptotic molecule BAX is the converse of BCL2, in fact itpromotes both cell death and cell cycle (Yin et al., 1997). Ourresults demonstrated that the anti-apoptotic effects ofPACAP38 and VIP peptides were correlated to changes inthese BCL2 family members. However, the analysis waslimited to the characterization of BCL2 and BAX expression
levels and it cannot be ruled out that the trophic action ofPACAP38 and VIP affect the expression of other genes involvedin apoptotic process.
The present results confirmed previously reported anti-apoptotic actions mediated by PAC1 and VPAC receptors(Brenneman, 2007). This is the first report on the directsurvival promoting effects of PACAP38 and VIP in schwan-noma cells. Further experiments should be performed toconfirm if the two peptides could exert similar actions onhealthy Schwann cells.
4. Experimental procedures
4.1. Cell culture
The study was carried out in rat schwannoma cells (CRL-2768)obtained from American Type Culture Collection (Rockville,MD, USA). Cells were cultured in Dulbecco's modified Eagle'smedium supplemented with 10% heat-inactivated fetalbovine serum and 2-mM L-glutamine, 100 U/ml penicillin,and 200-μg/ml streptomycin (Sigma-Aldrich,USA). Cells wereincubated at 37 °C in a humidified atmosphere with 5% CO2.
4.2. Analysis of mRNA expression by RT-PCR
Total RNA extracts were isolated by 1 ml TRIzol reagent(Invitrogen) and 0.2 ml chloroform and precipitated with0.5 ml isopropanol. Pellet was washed with 75% ethanol andair dried. Single stranded cDNAs were synthesized incubatingtotal RNA (5 μg) with SuperScript III Rnase H-reverse trans-criptase (200 U/μl) (Invitrogen); Oligo-(dT)20 primer (100 nM)(Invitrogen); 1 mM dNTP mix (Invitrogen), dithiothreitol (DTT,0.1M), Recombinant RNase-inhibitor (40 U/μl) at 42 °C for 1 h ina final volume of 20 μl. Reaction was terminated by incubationof samples at 70 °C for 10 min.
Aliquots of cDNA were amplified using specific primers forPAC1/VPAC type receptors, PACAP and VIP peptides. Primersused to identify PAC1 receptors' isoforms, VPAC1 receptor,VPAC2 receptor, PACAP peptide, VIP peptide, BCL2, BAX and

Table 2 – Primer sequences
Gene Forward Reverse bp length
PAC1 receptor GeneBank accession no Z23272 ATCGGCATCATCATCATCCT TTCCCTCTTGCTGACGTTCT 339 (Hip–Hop)253 (Hop)251(Hip)168 (R)
PAC1 Hip variant GeneBank accession no Z23272 ATCGGCATCATCATCATCCT AGGAAAGGGGGTGAATGCT 158PAC1 Hop variant GeneBank accession no Z23272 AAATGCTACTGCAAGCCACA TTCCCTCTTGCTGACGTTCT 165VPAC1 receptor GeneBank accession no NM_012685 GTGAAGACCGGCTACACCAT TGAAGAGGGCCATATCCTTG 178VPAC2 receptor GeneBank accession no NM_017238 AGAGCCATCTCTGTGCTGGT AGGTAGGCCAGGAAACACCT 221PACAP peptide GeneBank accession no NM_016989 ACAGCGTCTCCTGTTCACCT CCTGTCGGCTGGGTAGTAAA 179VIP peptide GeneBank accession no XM_217838 CAGAAGCAAGCCTCAGTTCC CATTCTCCGCTAAGGCATTC 197Bcl2 GeneBank accession no NM_016993 AAGCTGTCACAGAGGGGCTA CTCTCAGGCTGGAAGGAGAA 101Bax GeneBank accession no NM_017059 TGTTTGCTGATGGCAACTTC GATCAGCTCGGGCACTTTAG 10418 S Ribosomial subunit GeneBank accession no X57529 CAGAAGGACGTGAAGGATGG CAGTGGTCTTGGTGTGCTGA 159
Forward and reverse primers were selected from the 5′ and 3′ region of each genemRNA. The expected length of PCR products is indicated in theright column.
33B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
18 S ribosomial subunit were based on the reported sequences(NCBI). Oligonucleotide sequences are listed in Table 2. EachPCR reaction contained 0.4 μM specific primers, 200 μMdNTPs,1.25 U AmpliTaq Gold DNA polymerase and GeneAmp buffercontaining 2.5 mM MgCl2+ (Applied Biosystem). PCR wasperformed using the following three cycle programs: (I)denaturation of cDNA (1 cycle: 95 °C for 12 min); (II)amplification (40 cycles: 95 °C for 30 s, 60 °C for 30 s, 72 °Cfor 45 s); (III) final extension (1 cycle: 72 °C for 7 min).Amplification products were separated by electrophoresis in a1.8% agarose gel in 0.045 M Tris–borate/1 mM EDTA buffer.Bands were excised from the gel, subcloned in a vector andsequenced to confirm their identity. A positive control wasperformed using rat whole brain cDNAs to validate primersspecificity.
4.3. Western blot analysis
Crude extract was prepared by homogenizing cells in a buffercontaining 20 mM Tris (pH 7.4), 2 mM EDTA, 0.5 mM EGTA;50 mM mercaptoethanol, 0.32 mM sucrose and a proteaseinhibitor cocktail (Roche Diagnostics) using a Teflon-glasshomogenizer and then sonicated. Protein concentrations weredetermined by Bradford's method (Bradford, 1976) using BSAas a standard. Sample proteins (10 μg) were diluted in 4Xsodium dodecyl sulphate (SDS) protein gel loading solution(Invitrogen), boiled for 5 min, separated on 12% tris glycine gel(Invitrogen) by electrophoresis and processed as previouslydescribed by Pascale et al., 1996. PAC1 receptor rabbitpolyclonal antibody (raised against amino acids 61–115 ofPAC1 receptor of human origin, Santa Cruz Biotechnology Inc),VPAC2 receptor rabbit polyclonal antibody (raised againstamino acids 101–150 of VPAC2 receptor of human origin, SantaCruz Biotechnology Inc), PACAP peptide rabbit polyclonalantibody (raised against amino acids 101–176 of PACAP ofhuman origin, Santa Cruz Biotechnology Inc) and β-tubulinrabbit polyclonal antibody (raised against amino acids 210–444of β-tubulin of human origin, Santa Cruz Biotechnology Inc)were diluted 1:200, while the secondary antibody (goatantirabbit IgG2b HRP, Amersham Biosciences) was used at1:5000. Blots were developed using enhanced chemilumines-cence technique (Amersham Biosciences). No signal was
detected when the primary antibody was omitted (data notshown).
4.4. MTT test
To assess cell viability, we used the cell proliferation kit I(MTT) following manufacturer's instructions (Roche). Serumstarved cells were treated with a range of concentrations(0, 0.1, 1, 10, 100 and 1000 nM) of both PACAP38 (SigmaAldrich)and VIP (Sigma Aldrich) for 24 h andwith 100 nM PACAP38 and100 nM VIP for 24 h, 48 h and 72 h. Cells were seeded into 96-well plates at a concentration of 1×105 cells/well. DMEMcontaining 0.5 mg/ml 3-[4,5-dimethylthiazol-2-yl]-2,5-diphe-nyltetrazolium bromide (MTT) (Sigma Aldrich) was added ineach well. Following incubation for 2 h at 37 °C, medium wasremoved, and 100 μl of DMSOwas added. Formazan formed bythe cleavage of the yellow tetrazolium salt MTTwasmeasuredspectrophotometrically by absorbance change at 550–600 nmusing a microplate reader.
4.5. Immunodetection of oligonucleosomes
Mononucleosomes and oligonucleosomes released from thenucleus into the cytoplasm of apoptotic cells were detected byusing a sandwich enzyme-linked immunosorbent assay (TheCell Death Detection ELISAPLUS 10× ,Roche Applied Sciences).The assay is based on quantitative sandwich enzyme-linkedimmunosorbent assay principle, with rat monoclonal anti-body directed against DNA histones. For sample preparation,untreated, serum starved, 100 nM PACAP38 or 100 nM VIPtreated rat schwannoma cells were harvested by trypsiniza-tion. Cells were lysed in incubation buffer for 30 min. Thelysate was centrifuged at 20000 g×10 min. Supernatant wasdiluted to yield 1×104 cell equivalents/ml and used forimmunodetection. The assay was performed as follows: (1)an antibody that react with the histone H1, H2A, H2B, H3 andH4 was fixed on the wall of the microplate module providedwith the kit; (2) samples prepared as described above wereadded to the plate containing the immobilized anti-histoneantibody; (3) anti-DNA monoclonal antibodies conjugated toperoxidase were added, to allow their binding to the DNA partof nucleosomes; and (4) after removal of unbound peroxidase

34 B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
conjugate, the amount of peroxidase retained in theimmunocomplex was determined photometrically with 2,2′-azino-di(3-ethylbenzthiazoline sulfonate) as a substrate.
4.6. Measurement of BCL2 and BAX levels by quantitativeReal Time PCR
Aliquots of cDNA (400 ng) from untreated, serum starved,100 nM PACAP38 or 100 nM VIP treated cells and externalstandards at known amounts (purified PCR products, rangingfrom 102 to 108 copies) were amplified in parallel reactions,using primers indicated in Table 2. mRNA levels of thereference gene, 18 S ribosomial subunit, were measured ineach amplification. Each PCR reaction contained 0.5 μMprimers, 1.6 mMMgCl2+, 1X Light Cycler-FastStart DNA MasterSYBR Green I (Roche Diagnostic). Amplifications were per-formed using a Light Cycler 1.5 instrument (Roche Diagnostic)with the following program setting : (I) cDNA denaturation(1 cycle: 95 °C for 10 min); (II) quantification (45 cycles:95 °C for 10 s, 57 °C for 7 s, 72 °C for 5 s); (III) melting curveanalysis (1 cycle: 95 °C for 0 s, 65 °C for 15 s, 95 °C for 0 s);(IV) cooling (1 cycle: 40 °C for 30 s). Quantification wasobtained by comparing the fluorescence emitted by PCRproducts at unknown concentration with the fluorescenceemitted by external standards at known concentration. Forthis analysis, fluorescence values, measured in the log-linearphase of amplification, were estimated with the secondderivativemaximummethod using Light Cycler Data Analysissoftware. PCR products specificity was evaluated by meltingcurve analysis followed by gel electrophoresis.
Expression levels were assessed by analyzing the meancopy number/ng mRNA for each gene.
4.7. Statistical analysis
Data are reported as mean±SEM. One-way analysis ofvariance (ANOVA) was used to compare differences amonggroups followed by Tukey test to evaluate which groups werestatistically different. The level of significance for all statisticaltests was p≤0.05.
Acknowledgments
These experiments were supported by the PhD program inNeuropharmacology, University of Catania, Medical School.
We thank Mr P. Asero for his technical support and Dr. F.Murabito for his administrative support.
R E F E R E N C E S
Arimura, A., 1998. Perspectives on pituitary adenylate cyclaseactivating polypeptide (PACAP) in the neuroendocrine,endocrine, and nervous systems. Jpn. J. Physiol. 48, 301–331.
Bansal, R., Pfeiffer, S.E., 1987. Regulated galactolipid synthesis andcell surface expression in Schwann cell line D6P2 T.J. Neurochem. 49, 1902–1911.
Bradford, M.M., 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72, 248–254.
Brenneman, D.E., Phillips, T.M., Festoff, B.W., Gozes, I., 1997.Identity of neurotrophic molecules released from astroglia byvasoactive intestinal peptide. Ann. N.Y. Acad. Sci. 814, 167–173.
Brenneman, D.E., 2007. Neuroprotection: a comparative view ofvasoactive intestinal peptide and pituitary Adenylatecyclase-activating polypeptide. Peptides 28, 1720–1726.
DiCicco-Bloom, E., Deutsch, P.J., Maltzman, J., Zhang, J., Pintar, J.E.,Zheng, J., et al., 2000. Autocrine expression and ontogeneticfunctions of the PACAP ligand/receptor system duringsympathetic development. Dev. Biol. 219, 197–213.
Ge, Y., Belcher, S.M., Pierce, D.R., Light, K.M., 2004. Alteredexpression of Bcl2, Bad and Bax mRNA occurs in the ratcerebellum within hours after ethanol exposure onpostnatal day 4 but not on postnatal day 9. Mol. Brain Res. 129,124–134.
Gil-Gomez, G., Berns, A., Brady, H.J., 1998. A link between cell cycleand cell death: Bax and Bcl-2 modulate Cdk2 activation duringthymocyte apoptosis. EMBO J. 17, 7209–7218.
Hai, M., Muja, N., DeVries, G.H., Quarles, R.H., Patel, P.I., 2002.Comparative analysis of Schwann cell lines asmodel systems formyelin gene transcription studies. J. Neurosci. Res. 69, 497–508.
Huang, D.C., O'Reilly, L.A., Strasser, A., Cory, S., 1997. Theanti-apoptosis function of Bcl-2 can be genetically separatedfrom its inhibitory effect on cell cycle entry. EMBO J. 16,4628–4638.
Isobe, K., Tatsuno, I., Yashiro, T., Nanmoku, T., Takekoshi, K.,Kawakami, Y., et al., 2003. Expression of mRNA for PACAP andits receptors in intra- and extra- adrenal humanpheochromocytomas and their relationship to catecholaminesynthesis. Regul. Pept. 110, 213–217.
Jamen, F., Bouschet, T., Laden, J.C., Bockaert, J., Brabet, P., 2002.Up-regulation of the PACAP type-1 receptor (PAC1) promoter byneurotrophins in rat PC12 cells and mouse cerebellar granulecells via the Ras/mitogen-activated protein kinase cascade.J. Neurochem. 82, 1199–1207.
Jaworski, D.M., 2000. Expression of pituitary adenylatecyclase-activating polypeptide (PACAP) and thePACAP-selective receptor in cultured rat astrocytes, humanbrain tumors, and in response to acute intracranial injury. CellTissue Res. 300, 219–230.
Juarranz, M.G., Bolaños, O., Gutiérrez-Cañas, I., Lerner, E.A.,Robberecht, P., Carmena, M.J., et al., 2001. Neuroendocrinedifferentiation of the LNCaP prostate cancer cell line maintainsthe expression and function of VIP and PACAP receptors. CellSignal 13, 887–894.
Lee,M., Lelievre, V., Zhao, P., Torres,M., Rodriguez,W., Byun, J., et al.,2001. Pituitary adenylyl cyclase-activating polypeptidestimulates DNA synthesis but delays maturation ofoligodendrocyte progenitors. J. Neurosci. 21, 3849–3859.
Lindholm, D., Skoglösa, Y., Takei, N., 1998. Developmentalregulation of pituitary adenylate cyclase activating polypeptide(PACAP) and its receptor 1 in rat brain: function of PACAP as aneurotrophic factor. Ann. N.Y. Acad. Sci. 865, 189–196.
Nielsen, H.S., Hannibal, J., Fahrenkrug, J., 1998a. Embryonicexpression of pituitary adenylate cyclase-activatingpolypeptide in sensory and autonomic ganglia and in spinalcord of the rat. J. Comp. Neurol. 394, 403–415.
Nielsen, H.S., Hannibal, J., Fahrenkrug, J., 1998b. Prenatalexpression of pituitary adenylate cyclase activatingpolypeptide (PACAP) in autonomic and sensory ganglia andspinal cord of rat embryos. Ann. N.Y. Acad. Sci. 865, 533–536.
Oka, H., Jin, L., Reubi, J.C., Qian, X., Scheithauer, B.W., Fujii, K., et al.,1998. Pituitary adenylate-cyclase-activating polypeptide(PACAP) binding sites and PACAP/vasoactive intestinalpolypeptide receptor expression in human pituitary adenomas.Am. J. Pathol. 153, 1787–1796.
Onoue, S., Endo, K., Ohshima, K., Yajima, T., Kashimoto, K., 2002.The neuropeptide PACAP attenuates beta-amyloid(1-42)-induced toxicity in PC12 cells. Peptides 23, 1471–1478.

35B R A I N R E S E A R C H 1 2 4 1 ( 2 0 0 8 ) 2 9 – 3 5
Pascale, A., Fortino, I., Covoni, S., Trabucchi, M., Wetsel, W.C.,Battaini, F., 1996. Functional impairment in protein kinase C byRACK1 (receptor for activated C kinase 1) deficiency in aged ratbrain cortex. J. Neurochem. 67, 2471–2477.
Pincus, D.W., DiCicco-Bloom, E.M., Black, I.B., 1990. Vasoactiveintestinal peptide regulation of neuroblast mitosis andsurvival: role of cAMP. Brain Res. 514, 355–357.
Reubi, J.C., 2000. In vitro evaluation of VIP/PACAP receptors inhealthy and diseased human tissues. Clinical implicationsAnn. N.Y. Acad. Sci. 921, 1–25.
Robberecht, P., Vertongen, P., Velkeniers, B., De Neef, P.,Vergani, P., Raftopoulos, C., et al., 1993. Receptors for pituitaryadenylate cyclase activating peptides in human pituitaryadenomas. J. Clin. Endocrinol. Metab. 77, 1235–1239.
Robberecht, P., Woussen-Colle, M.C., Vertongen, P., De Neef, P.,Hou, X., Salmon, I., et al., 1994. Expression of pituitaryadenylate cyclase activating polypeptide (PACAP) receptors inhuman glial cell tumors. Peptides 15, 661–665.
Skoglösa, Y., Takei, N., Lindholm, D., 1999. Distribution of pituitaryadenylate cyclase activating polypeptide mRNA in thedeveloping rat brain. Brain Res. Mol. Brain Res. 65, 1–13.
Vairo, G., Innes, K.M., Adams, J.M., 1996. Bcl-2 has a cell cycleinhibitory function separable from its enhancement of cellsurvival. Oncogene 13, 1511–1519.
Vaudry, D., Gonzalez, B.J., Basille, M., Fournier, A., Vaudry, H., 1999.Neurotrophic activity of pituitary adenylate cyclase-activatingpolypeptide on rat cerebellar cortex during development. Proc.Natl. Acad. Sci. U. S. A. 96, 9415–9420.
Vaudry, D., Gonzalez, B.J., Basille, M., Yon, L., Fournier, A.,Vaudry, H., 2000. Pituitary adenylate cyclase-activating
polypeptide and its receptors: from structure to functions.Pharmacol. Rev. 52, 269–324.
Vaudry, D., Pamantung, T.F., Basille, M., Rousselle, C., Fournier, A.,Vaudry, H., et al., 2002. PACAP protects cerebellar granuleneurons against oxidative stress-induced apoptosis. Eur. J.Neurosci. 15, 1451–1460.
Vaux, D.L., Cory, S., Adams, J.M., 1988. Bcl-2 gene promoteshaemopoietic cell survival and cooperates with c-myc toimmortalize pre-B cells. Nature 335, 440–442.
Vertongen, P., D'Haens, J., Michotte, A., Velkeniers, B.,Van Rampelbergh, J., Svoboda, M., et al., 1995. Expression ofpituitary adenylate cyclase activating polypeptide andreceptors in human brain tumors. Peptides 16, 713–719.
Waschek, J.A., 1996. VIP and PACAP receptor-mediated actions oncell proliferation and survival. Ann. N.Y. Acad. Sci. 805,290–300.
Waschek, J.A., Casillas, R.A., Nguyen, T.B., DiCicco-Bloom, E.M.,Carpenter, E.M., Rodriguez, W.I., 1998. Neural tube expressionof pituitary adenylate cyclase-activating peptide (PACAP) andreceptor: potential role in patterning and neurogenesis. Proc.Natl. Acad. Sci. U. S. A. 95, 9602–9607.
Yin, C., Knudson, C.M., Korsmeyer, S.J., Van Dyke, T., 1997. Baxsuppresses tumorigenesis and stimulates apoptosis in vivo.Nature 385, 637–640.
Zhou, C.J., Shioda, S., Yada, T., Inagaki, N., Pleasure, S.J.,Kikuyama, S., 2002. PACAP and its receptors exert pleiotropiceffects in the nervous system by activating multiple signalingpathways. Curr. Protein Pept. Sci. 3, 423–439.
Zinkel, S., Gross, A., Yang, E., 2006. BCL2 family in DNA damageand cell cycle control. Cell Death Differ. 13, 1351–1359.




















