
Differential Gene Expression in the Activation and Maturation of Human Monocytes

Experimental Gerontology 46 (2011) 897–904
Contents lists available at ScienceDirect
Experimental Gerontology
j ourna l homepage: www.e lsev ie r.com/ locate /expgero
Two variants located in the upstream enhancer region of human UCP1 gene affectgene expression and are correlated with human longevity
Giuseppina Rose ⁎, Paolina Crocco, Patrizia D'Aquila, Alberto Montesanto, Dina Bellizzi, Giuseppe PassarinoDepartment of Cell Biology, University of Calabria, 87036, Rende, Italy
⁎ Corresponding author. Tel.: +39 0984492931; fax:E-mail address: [email protected] (G. Rose).
0531-5565/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.exger.2011.07.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 February 2011Received in revised form 31 May 2011Accepted 24 July 2011Available online 30 July 2011
Section Editor: R. Westendorp
Keywords:Uncoupling proteinsThermogenesisEnergy storageLongevityGenetic polymorphisms
The brown fat specific UnCoupling Protein 1 (UCP1) is involved in thermogenesis, a process by which energyis dissipated as heat in response to cold stress and excess of caloric intake. Thermogenesis has potentialimplications for body mass control and cellular fat metabolism. In fact, in humans, the variability of the UCP1gene is associated with obesity, fat gain and metabolism. Since regulation of metabolism is one of the key-pathways in lifespan extension, we tested the possible effects of UCP1 variability on survival.Two polymorphisms (A-3826G and C-3740A), falling in the upstream promoter region of UCP1, were analyzedin a sample of 910 subjects from southern Italy (475 women and 435 men; age range 40–109). By analyzinghaplotype specific survival functions we found that the A-C haplotype favors survival in the elderly.Consistently, transfection experiments showed that the luciferase activity of the construct containing the A-Chaplotype was significantly higher than that containing the G-A haplotype. Interestingly, the different UCP1haplotypes responded differently to hormonal stimuli. The results we present suggest a correlation betweenthe activity of UCP1 and human survival, indicating once again the intricacy of mechanisms involved in energyproduction, storage and consumption as the key to understanding human aging and longevity.
+39 0984492911.
l rights reserved.
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
The UnCoupling Protein 1 (UCP1), also known as thermogenin,belongs to a family of anion transporters located in the innermembrane of mitochondria. By allowing the leak of protons throughthe inner mitochondrial membrane of Brown Adipose Tissue (BAT),UCP1 uncouples fuel oxidation from ATP synthesis. UCP1 activationleads to thermogenesis, which allows energy to be dissipated as heatin response to cold stress and excess caloric intake (Lowell andSpiegelman, 2000; Cannon and Nedergaard, 2004).
There is a huge body of evidence indicating that UCP1-mediatedthermogenesis in brown fat in rodents significantly contributes tocontrol of body temperature, body weight, energy expenditure,glucose and lipid metabolism (Sell et al., 2004; Cannon andNedergaard, 2010; Kozak et al., 2010). Moreover, since a consequenceof the mitochondrial uncoupling is the lowering of the mitochondrialmembrane potential, a potential role for thermogenin in preventingexcessive production of mitochondrial reactive oxygen species (ROS)has also been proposed (Cannon et al., 2006; Oelkrug et al., 2010;Dlasková et al., 2010).
In humans, UCP1 has been thought to be exclusively expressed inBAT which is well developed only in newborns and young children.However, some recent studies have shown thatUCP1 is also expressed,
albeit at low levels, in islet cells (Sale et al., 2007) and in thymocytes(Adams et al., 2008). Moreover, in response to certain stimuli such ascold exposure and sympathetic stimulation, human white adipocytescan acquire the features of brown cells (Tiraby et al., 2003).Furthermore, starting from the first study by Nedergaard et al.(2007), a series of reports provided evidence for the presence ofactive adipose tissue in adult humans (Zingaretti et al., 2009; Virtanenet al., 2009; Cypess et al., 2009; van Marken Lichtenbelt et al., 2009;Saito et al., 2009; Nedergaard et al., 2011). In addition, Yoneshiro et al.(2010) have reported that BAT is involved in cold-induced thermo-genesis in healthy adult men, and thus contributes to whole-bodyenergy expenditure. It may therefore be inferred from theseobservations that BAT thermogenesis may have a role in humanadult whole-body metabolism, and therefore also in aging andlongevity.
Aging is a very complex phenomenon, largely influenced by anincreased susceptibility to energy imbalance (Roberts and Rosenberg,2006). The dysregulation of energy intake and the progressive declineof all components of energy expenditure (that is, obligatory, activityinduced and facultative thermogenesis) with advancing age has beenwell established (Roberts and Rosenberg, 2006; Manini, 2010). Thedecline of resting metabolic rate (RMR), which represents the largestcomponent of energy expenditure, is typically associatedwith changesin body composition (i.e. increase in fatmass and the decline in fat-freemass). However, the analysis of energy expenditure shows that afteradjustment for differences in fat mass and fat-free mass, the bodycomposition accounts for a small proportion of the age associated RMR

Table 1Frequency distribution of two-marker haplotypes in the UCP1 region in the analyzedsample categorized according to the defined age classes. Standard errors (SE) ofhaplotype frequencies were calculated through 1000 bootstrap resampling cycles.
Age groups
Females
Haplotypes Age class 12n=400(40–72 years)
Age class 22n=341(73–91 years)
Age class 32n=236(N91 years)
Rel. freq.±SE Rel. freq.±SE Rel. freq.±SE
A-C 0.741±0.022 0.780±0.022 0.764±0.028G-A 0.088±0.014 0.092±0.016 0.081±0.017G-C 0.171±0.019 0.127±0.019 0.153±0.023A-A – – –
Males
Haplotypes Age class 12n=232(40–65 years)
Age class 22n=430(66–88 years)
Age class 32n=208(N88 years)
Rel. Freq. ± SE Rel. Freq. ± SE Rel. Freq. ± SE
A-C 0.728±0.029 0.760±0.020 0.773±0.030G-A 0.134±0.022 0.110±0.016 0.076±0.019G-C 0.139±0.024 0.131±0.016 0.152±0.026A-A – – –
898 G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
decline (Krems et al., 2005; Frisard et al., 2007; Ruggiero et al., 2008;Luhrmann et al., 2010; St-Onge and Gallagher, 2010). This indicatesthat in aging, resting metabolism may decrease through othermechanisms such as reduction in the mass and metabolic rate ofindividual organs and tissues. An increase in resting metabolic rate, asis observed in pathological conditions, accelerates aging and leads toearly mortality (Ruggiero et al., 2008; Jumpertz et al., 2011).Consistently, a reduction in metabolic rate, such as in caloricrestriction, improves several biological parameters and has a favorableimpact on longevity in both animal models and humans (Mattson andWan, 2005). Notwithstanding the above considerations, the linkbetween metabolic rate and aging and longevity is far fromstraightforward. Rizzo et al. (2005) showed that compared to agedsubjects, healthy long-lived subjects display energy expenditureparameters that are closer to the values of healthymiddle-aged adults.
It is also worth noting that aging is characterized by the progressiveincrease in oxidative stress due to the production of ROS during cellularmetabolism. The accumulation over time of oxidative damage to cellularcomponents affects normal cell functioning, and leads to impaired abilityto maintain baseline tissue energy homeostasis, and to the inability torespond adequately to physiological needs (Frisard and Ravussin, 2006).Therefore, metabolic control and stress response are interconnectedprocesses that likely contribute to the aging phenomenon.
According to the “Uncoupling to survive theory” (Brand, 2000)uncoupling modulates the aging process by affecting the body'smetabolic rate (mitochondrial proton leak accounts for 20–30% ofwhole-body resting energy expenditure, see Harper et al., 2002), andby protecting from oxidative stress. In this scenario, the UCP1uncoupling activity in BAT mitochondria, as well as the uncouplingactivity of other UCPs in different tissues, may be considered aphysiological process that by linking metabolic rate and ROSproduction, can modulate the rate of human aging and lifespan. It isworth noting that in a previous report we showed that a functionalpolymorphism located in the promoter region of the muscle-specificUCP3 gene (rs1800849), significantly affected hand grip strength, apredictor of disability and mortality, in an elderly population fromsouthern Italy (Crocco et al., 2011). Our report suggested a correlationbetween the uncoupling process and the regulation of muscleperformance in the elderly. Thus, the goal of this study, was to testthe hypothesis that aging and longevity are influenced by UCP1variants. Two polymorphisms, A-3826G and C-3740A, falling in theupstream enhancer region of UCP1, were studied in a cohort of middleaged and oldest old subjects from southern Italy. The functionalrelevance of the two polymorphisms in the context of haplotypes wasalso explored.
2. Materials and methods
2.1. Subjects and genotyping
A total of 910 (475 women and 435 men) unrelated individuals(age range: 40–109 years) participated in the present study. Thesubjects, born and currently living in Calabria (southern Italy), andwhose parents and grandparents were native of the same area, werecollected during several campaigns of recruitment (De Rango et al.,2010). All the subjects gave their informed consent to use theiranonymous genetic and phenotypic data for genetic studies on agingand longevity.
The UCP1 A-3826G polymorphism was genotyped by genomic PCRand BclI-RFLP analysis as previously described (Oppert et al., 1994). Togenotype the UCP1 C-3737A polymorphism 3–5 μl of the amplifiedDNA as above was digested by the restriction enzyme DdeI (3 U) for3 h at 37 °C as recommended by the manufacturer. The fragmentswere separated by electrophoresis on a 2% agarose gel and stainedwith ethidium bromide.
2.2. Genetic and statistical analyses
The Hardy–Weinberg equilibrium (HWE) was tested by using aMonte-Carlo approach based on 10,000 random allele permutationsfor each sample (Weir, 1996).
An expectation maximization (EM) algorithm, implemented in theArlequin software package (Excoffier et al., 2005) was used to obtainmaximum likelihood estimation of multilocus haplotype frequenciesfor UCP1 genotypes. Standard deviations of the haplotype frequencieswere estimated by a parametric bootstrap procedure (1000 replica-tions). The D′ linkage disequilibrium coefficient was assessed usingArlequin software package (Excoffier et al., 2005). Diplotypes wereconstructed based on the population genotypes in the UCP1 geneusing a pseudo-Bayesian approach (ELB algorithm) implemented inthe Arlequin software package (Excoffier et al., 2005).
In order to model haplotype/diplotype frequency as a function ofage, we carried out a binomial logistic regression analysis, aspreviously reported by Tan and co-workers (Tan et al., 2001). Forthis purpose the following model was used:
g πi xð Þ½ � = lnπi xð Þ
1−πi xð Þ� �
= βi0 + βi1x: 1
In Eq. (1) g[πi(x)] represents a logit transformation of thefrequency of the carriers of the haplotype hi at age x, πi(x). Thus, theeffect of the haplotype hi on survival is represented by the slopeparameter βi1: if it is significantly different from 0 the frequency of thecarrier of the haplotype hi goes up if βi1N0, or down if βi1b0.
For the diplotype frequencies, the previous model can bereformulated as:
g πij xð Þh i
= lnπij xð Þ
1−πij xð Þ
!= βij0 + βi j1x;
where the effect of the diplotype hi/hj on survival is represented by theslope parameter βij1: if it is significantly different from 0 the frequencyof the carriers of the diplotype hi/hj goes up if βij1N0, or down if βij1b0.
The binomial logistic regression model has been appliedto estimate the effect of each of the three reconstructed haplotypes(A-C, G-A and G-C) and of the relevant diplotypes on survival.
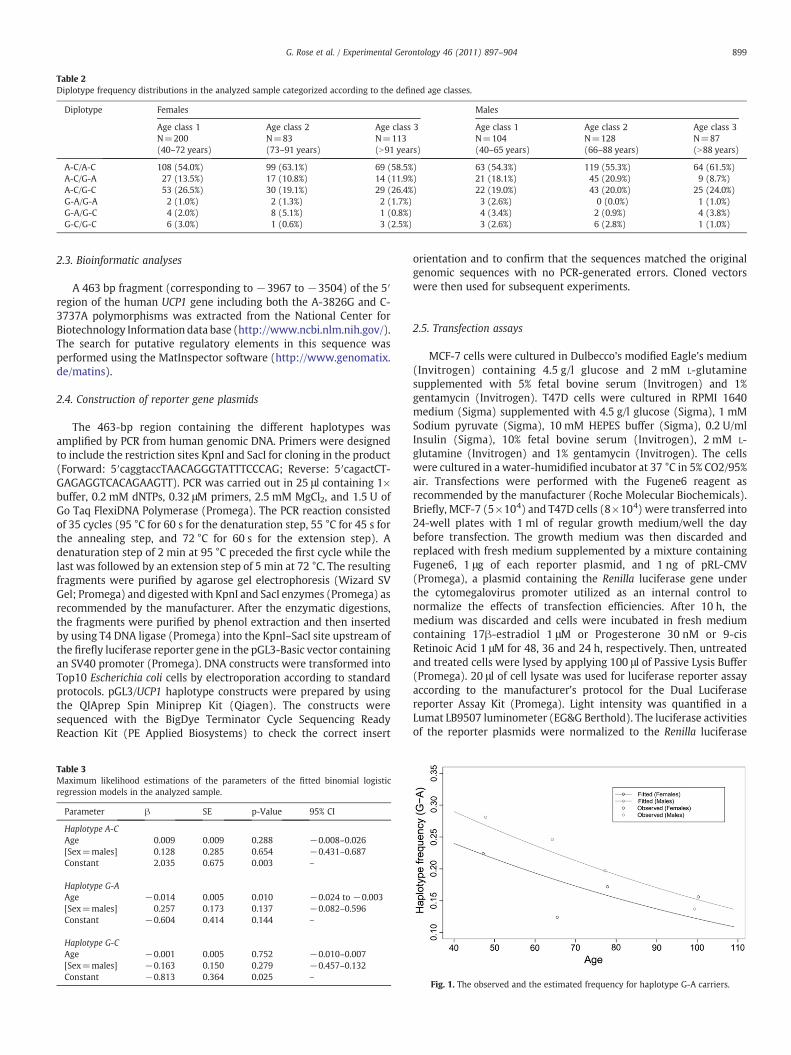
Table 2Diplotype frequency distributions in the analyzed sample categorized according to the defined age classes.
Diplotype Females Males
Age class 1N=200(40–72 years)
Age class 2N=83(73–91 years)
Age class 3N=113(N91 years)
Age class 1N=104(40–65 years)
Age class 2N=128(66–88 years)
Age class 3N=87(N88 years)
A-C/A-C 108 (54.0%) 99 (63.1%) 69 (58.5%) 63 (54.3%) 119 (55.3%) 64 (61.5%)A-C/G-A 27 (13.5%) 17 (10.8%) 14 (11.9%) 21 (18.1%) 45 (20.9%) 9 (8.7%)A-C/G-C 53 (26.5%) 30 (19.1%) 29 (26.4%) 22 (19.0%) 43 (20.0%) 25 (24.0%)G-A/G-A 2 (1.0%) 2 (1.3%) 2 (1.7%) 3 (2.6%) 0 (0.0%) 1 (1.0%)G-A/G-C 4 (2.0%) 8 (5.1%) 1 (0.8%) 4 (3.4%) 2 (0.9%) 4 (3.8%)G-C/G-C 6 (3.0%) 1 (0.6%) 3 (2.5%) 3 (2.6%) 6 (2.8%) 1 (1.0%)
899G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
2.3. Bioinformatic analyses
A 463 bp fragment (corresponding to −3967 to −3504) of the 5′region of the human UCP1 gene including both the A-3826G and C-3737A polymorphisms was extracted from the National Center forBiotechnology Information data base (http://www.ncbi.nlm.nih.gov/).The search for putative regulatory elements in this sequence wasperformed using the MatInspector software (http://www.genomatix.de/matins).
2.4. Construction of reporter gene plasmids
The 463-bp region containing the different haplotypes wasamplified by PCR from human genomic DNA. Primers were designedto include the restriction sites KpnI and SacI for cloning in the product(Forward: 5′caggtaccTAACAGGGTATTTCCCAG; Reverse: 5′cagactCT-GAGAGGTCACAGAAGTT). PCR was carried out in 25 μl containing 1×buffer, 0.2 mM dNTPs, 0.32 μM primers, 2.5 mM MgCl2, and 1.5 U ofGo Taq FlexiDNA Polymerase (Promega). The PCR reaction consistedof 35 cycles (95 °C for 60 s for the denaturation step, 55 °C for 45 s forthe annealing step, and 72 °C for 60 s for the extension step). Adenaturation step of 2 min at 95 °C preceded the first cycle while thelast was followed by an extension step of 5 min at 72 °C. The resultingfragments were purified by agarose gel electrophoresis (Wizard SVGel; Promega) and digestedwith KpnI and SacI enzymes (Promega) asrecommended by the manufacturer. After the enzymatic digestions,the fragments were purified by phenol extraction and then insertedby using T4 DNA ligase (Promega) into the KpnI–SacI site upstream ofthe firefly luciferase reporter gene in the pGL3-Basic vector containingan SV40 promoter (Promega). DNA constructs were transformed intoTop10 Escherichia coli cells by electroporation according to standardprotocols. pGL3/UCP1 haplotype constructs were prepared by usingthe QIAprep Spin Miniprep Kit (Qiagen). The constructs weresequenced with the BigDye Terminator Cycle Sequencing ReadyReaction Kit (PE Applied Biosystems) to check the correct insert
Table 3Maximum likelihood estimations of the parameters of the fitted binomial logisticregression models in the analyzed sample.
Parameter β SE p-Value 95% CI
Haplotype A-CAge 0.009 0.009 0.288 −0.008–0.026[Sex=males] 0.128 0.285 0.654 −0.431–0.687Constant 2.035 0.675 0.003 –
Haplotype G-AAge −0.014 0.005 0.010 −0.024 to −0.003[Sex=males] 0.257 0.173 0.137 −0.082–0.596Constant −0.604 0.414 0.144 –
Haplotype G-CAge −0.001 0.005 0.752 −0.010–0.007[Sex=males] −0.163 0.150 0.279 −0.457–0.132Constant −0.813 0.364 0.025 –
orientation and to confirm that the sequences matched the originalgenomic sequences with no PCR-generated errors. Cloned vectorswere then used for subsequent experiments.
2.5. Transfection assays
MCF-7 cells were cultured in Dulbecco's modified Eagle's medium(Invitrogen) containing 4.5 g/l glucose and 2 mM L-glutaminesupplemented with 5% fetal bovine serum (Invitrogen) and 1%gentamycin (Invitrogen). T47D cells were cultured in RPMI 1640medium (Sigma) supplemented with 4.5 g/l glucose (Sigma), 1 mMSodium pyruvate (Sigma), 10 mM HEPES buffer (Sigma), 0.2 U/mlInsulin (Sigma), 10% fetal bovine serum (Invitrogen), 2 mM L-glutamine (Invitrogen) and 1% gentamycin (Invitrogen). The cellswere cultured in a water-humidified incubator at 37 °C in 5% CO2/95%air. Transfections were performed with the Fugene6 reagent asrecommended by the manufacturer (Roche Molecular Biochemicals).Briefly, MCF-7 (5×104) and T47D cells (8×104) were transferred into24-well plates with 1 ml of regular growth medium/well the daybefore transfection. The growth medium was then discarded andreplaced with fresh medium supplemented by a mixture containingFugene6, 1 μg of each reporter plasmid, and 1 ng of pRL-CMV(Promega), a plasmid containing the Renilla luciferase gene underthe cytomegalovirus promoter utilized as an internal control tonormalize the effects of transfection efficiencies. After 10 h, themedium was discarded and cells were incubated in fresh mediumcontaining 17β-estradiol 1 μM or Progesterone 30 nM or 9-cisRetinoic Acid 1 μM for 48, 36 and 24 h, respectively. Then, untreatedand treated cells were lysed by applying 100 μl of Passive Lysis Buffer(Promega). 20 μl of cell lysate was used for luciferase reporter assayaccording to the manufacturer's protocol for the Dual Luciferasereporter Assay Kit (Promega). Light intensity was quantified in aLumat LB9507 luminometer (EG&G Berthold). The luciferase activitiesof the reporter plasmids were normalized to the Renilla luciferase
Fig. 1. The observed and the estimated frequency for haplotype G-A carriers.
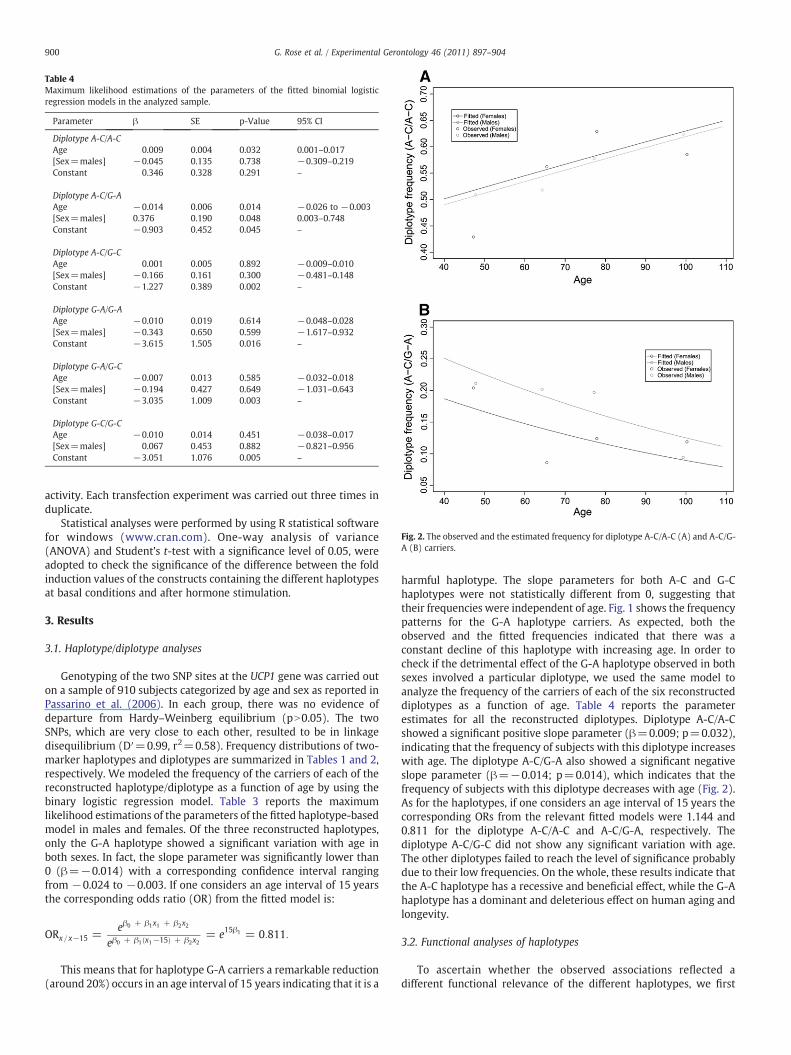
Table 4Maximum likelihood estimations of the parameters of the fitted binomial logisticregression models in the analyzed sample.
Parameter β SE p-Value 95% CI
Diplotype A-C/A-CAge 0.009 0.004 0.032 0.001–0.017[Sex=males] −0.045 0.135 0.738 −0.309–0.219Constant 0.346 0.328 0.291 –
Diplotype A-C/G-AAge −0.014 0.006 0.014 −0.026 to −0.003[Sex=males] 0.376 0.190 0.048 0.003–0.748Constant −0.903 0.452 0.045 –
Diplotype A-C/G-CAge 0.001 0.005 0.892 −0.009–0.010[Sex=males] −0.166 0.161 0.300 −0.481–0.148Constant −1.227 0.389 0.002 –
Diplotype G-A/G-AAge −0.010 0.019 0.614 −0.048–0.028[Sex=males] −0.343 0.650 0.599 −1.617–0.932Constant −3.615 1.505 0.016 –
Diplotype G-A/G-CAge −0.007 0.013 0.585 −0.032–0.018[Sex=males] −0.194 0.427 0.649 −1.031–0.643Constant −3.035 1.009 0.003 –
Diplotype G-C/G-CAge −0.010 0.014 0.451 −0.038–0.017[Sex=males] 0.067 0.453 0.882 −0.821–0.956Constant −3.051 1.076 0.005 –
Fig. 2. The observed and the estimated frequency for diplotype A-C/A-C (A) and A-C/G-A (B) carriers.
900 G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
activity. Each transfection experiment was carried out three times induplicate.
Statistical analyses were performed by using R statistical softwarefor windows (www.cran.com). One-way analysis of variance(ANOVA) and Student's t-test with a significance level of 0.05, wereadopted to check the significance of the difference between the foldinduction values of the constructs containing the different haplotypesat basal conditions and after hormone stimulation.
3. Results
3.1. Haplotype/diplotype analyses
Genotyping of the two SNP sites at the UCP1 gene was carried outon a sample of 910 subjects categorized by age and sex as reported inPassarino et al. (2006). In each group, there was no evidence ofdeparture from Hardy–Weinberg equilibrium (pN0.05). The twoSNPs, which are very close to each other, resulted to be in linkagedisequilibrium (D′=0.99, r2=0.58). Frequency distributions of two-marker haplotypes and diplotypes are summarized in Tables 1 and 2,respectively. We modeled the frequency of the carriers of each of thereconstructed haplotype/diplotype as a function of age by using thebinary logistic regression model. Table 3 reports the maximumlikelihood estimations of the parameters of the fitted haplotype-basedmodel in males and females. Of the three reconstructed haplotypes,only the G-A haplotype showed a significant variation with age inboth sexes. In fact, the slope parameter was significantly lower than0 (β=−0.014) with a corresponding confidence interval rangingfrom −0.024 to −0.003. If one considers an age interval of 15 yearsthe corresponding odds ratio (OR) from the fitted model is:
ORx= x−15 =eβ0 + β1x1 + β2x2
eβ0 + β1 x1−15ð Þ + β2x2= e15β1 = 0:811:
This means that for haplotype G-A carriers a remarkable reduction(around 20%) occurs in an age interval of 15 years indicating that it is a
harmful haplotype. The slope parameters for both A-C and G-Chaplotypes were not statistically different from 0, suggesting thattheir frequencies were independent of age. Fig. 1 shows the frequencypatterns for the G-A haplotype carriers. As expected, both theobserved and the fitted frequencies indicated that there was aconstant decline of this haplotype with increasing age. In order tocheck if the detrimental effect of the G-A haplotype observed in bothsexes involved a particular diplotype, we used the same model toanalyze the frequency of the carriers of each of the six reconstructeddiplotypes as a function of age. Table 4 reports the parameterestimates for all the reconstructed diplotypes. Diplotype A-C/A-Cshowed a significant positive slope parameter (β=0.009; p=0.032),indicating that the frequency of subjects with this diplotype increaseswith age. The diplotype A-C/G-A also showed a significant negativeslope parameter (β=−0.014; p=0.014), which indicates that thefrequency of subjects with this diplotype decreases with age (Fig. 2).As for the haplotypes, if one considers an age interval of 15 years thecorresponding ORs from the relevant fitted models were 1.144 and0.811 for the diplotype A-C/A-C and A-C/G-A, respectively. Thediplotype A-C/G-C did not show any significant variation with age.The other diplotypes failed to reach the level of significance probablydue to their low frequencies. On the whole, these results indicate thatthe A-C haplotype has a recessive and beneficial effect, while the G-Ahaplotype has a dominant and deleterious effect on human aging andlongevity.
3.2. Functional analyses of haplotypes
To ascertain whether the observed associations reflected adifferent functional relevance of the different haplotypes, we first

Fig. 3. Cloned sequence of the UCP1 gene containing the A-3828G (R) and the C-3737A (M) polymorphisms. Retinoic Acid-Response Element (RARE), Progesterone-ResponseElement (PRE), Activating Transcription Factor/cAMP Response Element binding protein (ATF/CREB), and Estrogen-Response Element (ERE) are underlined.
901G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
submitted a sequence of 463 bp (from −3967 to −3504) includingthe two polymorphic sites to the MatInspector online software. Asshown in Fig. 3, a putative retinoic acid response element (from−3842 to−3826), including the A-3826G variation, and a previouslycharacterized ATF/CREB binding element (from −3738 to −3733),carrying the C-3737A variation (Rousset et al., 2002), were predicted.Furthermore, a progesterone responsive element (PRE)-like sequence(from−3817 to−3804), and an estrogen responsive element (ERE)-like sequence (from −3713 to −3701), were also predicted in thevicinity of the two SNPs. Subsequently, using a luciferase reportergene assay we generated reporter constructs carrying the threehaplotypes and then tested their transcriptional activity in transientlytransfected MCF-7 and T47D cells, which specifically express theestradiol and progesterone receptors, respectively. Experiments werecarried out both under basal conditions and after hormonalstimulation. As a first step we estimated the maximum effectiveconcentration by quantitative dose–response experiments (data notshown). Fig. 4 shows that, under basal conditions, in both cell lines,the luciferase activity of the cells carrying either the A-C or the G-Aconstruct was significantly higher than cells carrying the haplotype G-C, which did not show any transcriptional activity. Moreover, theactivity of the A-C construct was higher than that of G-A.
With respect to basal conditions, a higher luciferase activity wasobserved for both the A-C (p=0.027) and G-A (p=0.046) constructsin MCF-7 cells treated with 9-cis retinoic acid (1 μM) for 24 h. Incontrast, A-C demonstrated a significantly decreased luciferaseactivity (p=0.028) when the same cells were treated with 17β-estradiol (1 μM) for 48 h (Fig. 4A and B). Moreover, in presence of17β-estradiol the G-A construct also showed a trend for reducedactivity which was not statistically inferior to basal transcription. Incontrast, the activity of the sole A-C construct increased when T47Dcells were treated with progesterone (30 nM) for 36 h (p=0.014;Fig. 4C).
4. Discussion
Human aging is accompanied by changes in energy balance andbody composition which are consequent to hormonal changes(Hughes et al., 2004). In this context, the role of BAT is particularlyimportant, since it regulates energy expenditure in the elderly. In fact,although the amount of BAT in human adults is low, it is responsiblefor 1–2% of the energy expenditure, which is enough to preventweight gain of about 2 kg per year (Lean, 1989). Consistently, aninverse relation between BAT and BMI, body fat, and visceral fat isobserved in aging (Mattson, 2010; Vijgen et al., 2011). UCP1, which isexpressed in BAT, has been observed to influence metabolism, fat
accumulation, and thermogenesis in mice models but also to beassociated with increased median survival and diminished age-related diseases (McDonald and Horwitz, 1999; Kontani et al., 2005;Gates et al., 2007).
For these reasons, a number of nucleotide variants in the UCP1gene have been described and analyzed in adult onset metabolicdisorders (Mori et al., 2001; Herrmann et al., 2003; Kim et al., 2005,2006). The A-3826G transition, which is in strong linkage disequilib-riumwith the C-3737A polymorphism (Rousset et al., 2002), has beenassociated with the percentage of body fat gain over time, highweight gain during adult life, and increased BMI in obese subjects (seeJia et al., 2010 and references therein). Here we report a relationshipbetweenUCP1 variability and survival at very old age. In particular, wefound that the frequency of the A-C/A-C diplotype increases, while theA-C/G-A frequency decreases as the population ages. By contrast, novariation in the A-C/G-C diplotype frequency was found. The A-3826Gand the C-3737A polymorphisms are located at the 5′ regionof the gene in proximity of a complex enhancer region (fromposition −3820 to −3470 upstream of the transcription start site)containing multiple and distinct cis-acting elements that mediatestrong drug-dependent transcriptional activation of the UCP1 gene(del Mar Gonzales-Barroso et al., 2000). The results of our in vitrotransfection experiments using luciferase reporter assays clearly pointto an effect of these haplotypes on UCP1 gene expression. The A-Chaplotype showed higher transcriptional activity compared to the G-Ahaplotype. No activity was observed for the GC haplotype. Theseresults seem to indicate a major role for the A-3826G polymorphismin modulating the transcriptional activity of the UCP1 gene. In linewith this, it has been shown that the -3826G allele is associated withreduced mRNA levels (Sramkova et al., 2007), while the C to Asubstitution at −3737, does not seem to substantially affecttranscriptional activity (Rousset et al., 2002).
It is worth noting that, although A-C and G-A haplotypes positivelyaffect UCP1 transcriptional activity compared to the GC haplotype,they have opposite effects on longevity. This apparently contrastingresult may be explained considering the haplotype-specific effectafter hormonal stimuli. In fact, progesterone up-regulates whereasestradiol down-regulates the transcriptional activity of the AChaplotype, while neither changes the transcriptional activity of GA.Therefore, it is likely that the real in vivo effect of these haplotypes oncomplex phenotypes such as aging and longevity is mediated by theintricacy of the different cellular and physiological stimuli. Similarly,this may explain why the G-C haplotype, lacking any transcriptionalactivity, also did not show significant changes in its frequency duringaging. It is evident that this observation calls for a better understand-ing of the different stimuli affecting UCP1 transcription. In this frame,
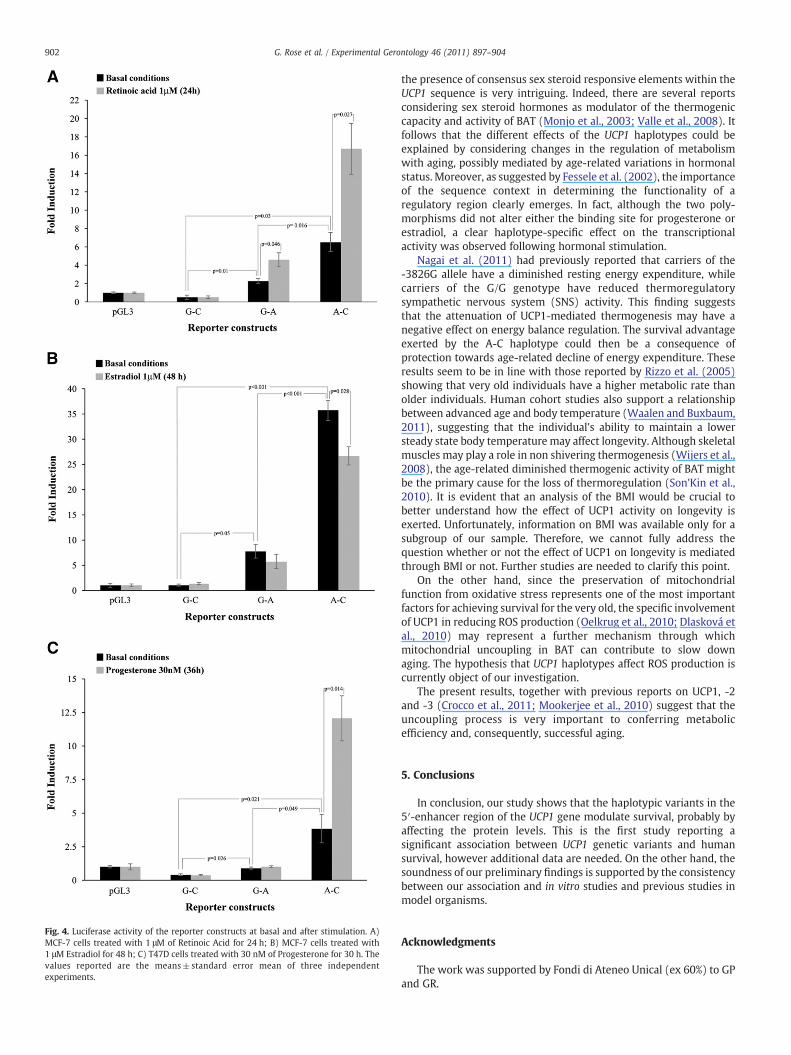
Fig. 4. Luciferase activity of the reporter constructs at basal and after stimulation. A)MCF-7 cells treated with 1 μM of Retinoic Acid for 24 h; B) MCF-7 cells treated with1 μM Estradiol for 48 h; C) T47D cells treated with 30 nM of Progesterone for 30 h. Thevalues reported are the means±standard error mean of three independentexperiments.
902 G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
the presence of consensus sex steroid responsive elements within theUCP1 sequence is very intriguing. Indeed, there are several reportsconsidering sex steroid hormones as modulator of the thermogeniccapacity and activity of BAT (Monjo et al., 2003; Valle et al., 2008). Itfollows that the different effects of the UCP1 haplotypes could beexplained by considering changes in the regulation of metabolismwith aging, possibly mediated by age-related variations in hormonalstatus. Moreover, as suggested by Fessele et al. (2002), the importanceof the sequence context in determining the functionality of aregulatory region clearly emerges. In fact, although the two poly-morphisms did not alter either the binding site for progesterone orestradiol, a clear haplotype-specific effect on the transcriptionalactivity was observed following hormonal stimulation.
Nagai et al. (2011) had previously reported that carriers of the-3826G allele have a diminished resting energy expenditure, whilecarriers of the G/G genotype have reduced thermoregulatorysympathetic nervous system (SNS) activity. This finding suggeststhat the attenuation of UCP1-mediated thermogenesis may have anegative effect on energy balance regulation. The survival advantageexerted by the A-C haplotype could then be a consequence ofprotection towards age-related decline of energy expenditure. Theseresults seem to be in line with those reported by Rizzo et al. (2005)showing that very old individuals have a higher metabolic rate thanolder individuals. Human cohort studies also support a relationshipbetween advanced age and body temperature (Waalen and Buxbaum,2011), suggesting that the individual's ability to maintain a lowersteady state body temperature may affect longevity. Although skeletalmuscles may play a role in non shivering thermogenesis (Wijers et al.,2008), the age-related diminished thermogenic activity of BAT mightbe the primary cause for the loss of thermoregulation (Son'Kin et al.,2010). It is evident that an analysis of the BMI would be crucial tobetter understand how the effect of UCP1 activity on longevity isexerted. Unfortunately, information on BMI was available only for asubgroup of our sample. Therefore, we cannot fully address thequestion whether or not the effect of UCP1 on longevity is mediatedthrough BMI or not. Further studies are needed to clarify this point.
On the other hand, since the preservation of mitochondrialfunction from oxidative stress represents one of the most importantfactors for achieving survival for the very old, the specific involvementof UCP1 in reducing ROS production (Oelkrug et al., 2010; Dlasková etal., 2010) may represent a further mechanism through whichmitochondrial uncoupling in BAT can contribute to slow downaging. The hypothesis that UCP1 haplotypes affect ROS production iscurrently object of our investigation.
The present results, together with previous reports on UCP1, -2and -3 (Crocco et al., 2011; Mookerjee et al., 2010) suggest that theuncoupling process is very important to conferring metabolicefficiency and, consequently, successful aging.
5. Conclusions
In conclusion, our study shows that the haplotypic variants in the5′-enhancer region of the UCP1 gene modulate survival, probably byaffecting the protein levels. This is the first study reporting asignificant association between UCP1 genetic variants and humansurvival, however additional data are needed. On the other hand, thesoundness of our preliminary findings is supported by the consistencybetween our association and in vitro studies and previous studies inmodel organisms.
Acknowledgments
The work was supported by Fondi di Ateneo Unical (ex 60%) to GPand GR.

903G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
References
Adams, A.E., Hanrahan, O., Nolan, D.N., Voorheis, H.P., Fallon, P., Porter, R.K., 2008.Images of mitochondrial UCP 1 in mouse thymocytes using confocal microscopy.Biochim. Biophys. Acta 1777, 115–117.
Brand, M.D., 2000. Uncoupling to survive? The role of mitochondrial inefficiency inageing. Exp. Gerontol. 35, 811–820.
Cannon, B., Nedergaard, J., 2004. Brown adipose tissue: function and physiologicalsignificance. Physiol. Rev. 84, 277–359.
Cannon, B., Nedergaard, J., 2010. Metabolic consequences of the presence or absence ofthe thermogenic capacity of brown adipose tissue in mice (and probably inhumans). Int. J. Obes. (Lond) 34 (Suppl. 1), S7–S16.
Cannon, B., Shabalina, I.G., Kramarova, T.V., Petrovic, N., Nedergaard, J., 2006.Uncoupling proteins: a role in protection against reactive oxygen species—ornot? Biochim. Biophys. Acta 1757, 449–458.
Crocco, P., Montesanto, A., Passarino, G., Rose, G., 2011. A common polymorphism in theUCP3 promoter influences hand grip strength in elderly people. Biogerontology 12(3), 265–271.
Cypess, A.M., Lehman, S., Williams, G., Tal, I., Rodman, D., Goldfine, A.B., Kuo, F.C., Palmer,E.L., Tseng, Y.H., Doria, A., Kolodny, G.M., Kahn, C.R., 2009. Identification andimportance of brown adipose tissue in adult humans. N. Eng. J. Med. 360, 1509–1517.
De Rango, F., Montesanto, A., Berardelli, M., Mazzei, B., Mari, V., Lattanzio, F., Corsonello,A., Passarino, G., 2010. To grow old in southern Italy: a comprehensive descriptionof the old and oldest old subjects in Calabria. Gerontology 57, 327–334.
del Mar Gonzales-Barroso, M., Pecqueur, C., Gelly, C., Sanchis, D., Alves-Guerra, M.C.,Bouillaud, F., Ricquier, D., Cassard-Doulcier, A.M., 2000. Transcriptional activationof the human UCP1 gene in a rodent cell line. Synergism of retinoids, isoproterenoloand thiazolidinedione is mediated by a multipartite response element. J. Biol.Chem. 275, 31722–31732.
Dlasková, A., Clarke, K.J., Porter, R.K., 2010. The role of UCP 1 in production of reactiveoxygen species by mitochondria isolated from brown adipose tissue. Biochim.Biophys. Acta 1797, 1470–1476.
Excoffier, L., Laval, G., Schneider, S., 2005. Arlequin (version 3.0): an integrated softwarepackage for population genetics data analysis. Evol. BioinformOnline 1, 47–50.
Fessele, S., Maler, H., Zischek, C., Nelson, P.J., Werner, T., 2002. Regulatory context is acrucial part of gene function. Trends Genet. 18, 60–63.
Frisard, M., Ravussin, E., 2006. Energy metabolism and oxidative stress: impact on themetabolic syndrome and the aging process. Endocrine 29, 27–32.
Frisard, M.I., Broussard, A., Davies, S.S., Roberts II, L.J., Rood, J., de Jonge, L., Fang, X.,Jazwinski, S.M., Deutsch, W.A., Ravussin, E., for the Louisiana Healthy Aging Study,2007. Aging, resting metabolic rate, and oxidative damage: results from theLouisiana Healthy Aging Study. J. Gerontol. A Biol. Sci. Med. Sci. 62, 752–759.
Gates, A.C., Bernal-Mizrachi, C., Chinault, S.L., Feng, C., Schneider, J.G., Coleman, T.,Malone, J.P., Townsend, R.R., Chakravarthy, M.V., Semenkovich, C.F., 2007.Respiratory uncoupling in skeletal muscle delays death and diminishes age-relateddisease. Cell. Metab. 6, 497–505.
Harper, M.E., Antoniou, A., Bevilacqua, L., Bezaire, V., Monemdjou, S., 2002. Cellularenergy expenditure and the importance of uncoupling. J. Anim. Sci. 80, E90–E97.
Herrmann, S.M., Wang, J.G., Staessen, J.A., Kertmen, E., Schmidt-Petersen, K., Zidek, W.,Paul, M., Brand, E., 2003. Uncoupling protein 1 and 3 polymorphisms are associatedwith waist-to-hip ratio. J. Mol. Med. 81, 327–332.
Hughes, V.A., Roubenoff, R., Wood, M., Frontera, W.R., Evans, W.J., Fiatarone Singh, M.A.,2004. Anthropometric assessment of 10-y changes in body composition in theelderly. Am. J. Clin. Nutr. 80, 475–482.
Jia, J.J., Tian, Y.B., Cao, Z.H., Tao, L.L., Zhang, X., Gao, S.Z., Ge, C.R., Lin, Q.Y., Jois, M., 2010.The polymorphisms of UCP1 genes associated with fat metabolism, obesity anddiabetes. Mol. Biol. Rep. 37, 1513–1522.
Jumpertz, R., Hanson, R.L., Sievers, M.L., Bennett, P.H., Nelson, R.G., Krakoff, J., 2011.Higher energy expenditure in humans predicts natural mortality. J. Clin.Endocrinol. Metab. 96, E972–E976.
Kim, K.S., Cho, D., Kim, Y.J., Choi, S.M., Kim, J.Y., Shin, S.U., Yoon, Y.S., 2005. The finding ofnew genetic polymorphism of UCP-1 A-1766G and its effects on body fataccumulation. Biochim. Biophys. Acta 1741, 149–155.
Kim, S.M., Han, J.H., Park, H.S., 2006. Prevalence of low HDL cholesterol levels andassociated factors among Koreans. Circ. J. 70, 820–826.
Kontani, Y., Wang, Y., Kimura, K., Inokuma, K.I., Saito, M., Suzuki-Miura, T., Wang, Z.,Sato, Y., Mori, N., Yamashita, H., 2005. UCP1 deficiency increases susceptibility todiet-induced obesity with age. Aging Cell 4, 147–155.
Kozak, L.P., Koza, R.A., Anunciado-Koza, R., 2010. Brown fat thermogenesis and bodyweight regulation in mice: relevance to humans. Int. J. Obes. (Lond) 34 (Suppl. 1),S23–S27.
Krems, C., Luhrmann, P.M., Strassburg, A., Hartmann, B., Neuhauser-Berthold, M., 2005.Lower resting metabolic rate in the elderly may not be entirely due to changes inbody composition. Eur. J. Clin. Nutr. 59, 255–262.
Lean, M.E., 1989. Brown adipose tissue in humans. Proc. Nutr. Soc. 48, 243–256.Lowell, B.B., Spiegelman, B.M., 2000. Towards a molecular understanding of adaptive
thermogenesis. Nature 404, 652–660.Luhrmann, P.M., Edelmann-Schafer, B., Neuhauser-Berthold, M., 2010. Changes in
resting metabolic rate in an elderly German population: cross-sectional andlongitudinal data. J. Nutr. Health Aging 14, 232–236.
Manini, T.M., 2010. Energy expenditure and aging. Ageing Res. Rev. 9, 1–11.Mattson, M.P., 2010. Perspective: does brown fat protect against diseases of aging?
Aging Res. Rev. 9, 69–76.Mattson, M.P., Wan, R., 2005. Beneficial effects of intermittent fasting and caloric
restriction on the cardiovascular and cerebrovascular systems. J. Nutr. Biochem. 16,129–137.
McDonald, R.B., Horwitz, B.A., 1999. Brown adipose tissue thermogenesis during agingand senescence. J. Bioenerg. Biomembr. 31, 507–516.
Monjo, M., Rodriguez, A.M., Palou, A., Roca, P., 2003. Direct effects of testosterone, 17betaestradiol, and progesterone on adrenergic regulation in cultured brownadipocytes: potential mechanism for gender-dependent thermogenesis. Endocri-nology 144, 4923–4930.
Mookerjee, S.A., Divakaruni, A.S., Jastroch, M., Brand, M.D., 2010. Mitochondrialuncoupling and lifespan. Mech. Ageing Dev. 131, 463–472.
Mori, H., Okazawa, H., Iwamoto, K., Maeda, E., Hashiramoto, M., Kasuga, M., 2001. Apolymorphism in the 5′ untranslated region and a Met229→Leu variant in exon 5of the human UCP1 gene are associated with susceptibility to type II diabetesmellitus. Diabetologia 44, 373–376.
Nagai, N., Sakane, N., Kotani, K., Hamada, T., Tsuzaki, K., Moritani, T., 2011. Uncouplingprotein 1 gene -3826 A/G polymorphism is associated with weight loss on a short-term, controlled-energy diet in young women. Nutr. Res. 31, 255–261.
Nedergaard, J., Bengtsson, T., Cannon, B., 2007. Unexpected evidence for activebrown adipose tissue in adult humans. Am. J. Physiol. Endocrinol. Metab. 293,E444–E452.
Nedergaard, J., Bengtsson, T., Cannon, B., 2011. Three years with adult human brownadipose tissue. Ann. N.Y. Acad. Sci. 1212, E20–E36.
Oelkrug, R., Kutschke, M., Meyer, C.W., Heldmaier, G., Jastroch, M., 2010. Uncouplingprotein 1 decreases superoxide production in brown adipose tissuemitochondria. J.Biol. Chem. 285, 21961–21968.
Oppert, J.M., Vohl, M.C., Chagnon, M., Dionne, F.T., Cassard-Doulcier, A.M., Ricquier, D.,Pérusse, L., Bouchard, C., 1994. DNA polymorphism in the uncoupling protein (UCP)gene and human body fat. Int. J. Obes. Relat. Metab. Disord. 18, 526–531.
Passarino, G., Montesanto, A., Dato, S., Giordano, S., Domma, F., Mari, V., Feraco, E., DeBenedictis, G., 2006. Sex and age specificity of susceptibility genes modulatingsurvival at old age. Hum. Hered. 62, 213–220.
Rizzo, M.R., Mari, D., Barbieri, M., Ragno, E., Grella, R., Provenzano, R., Villa, I., Esposito,K., Giugliano, D., Paolisso, G., 2005. Resting metabolic rate and respiratory quotientin human longevity. J. Clin. Endocrinol. Metab. 90, 409–413.
Roberts, S.B., Rosenberg, I., 2006. Nutrition and aging: changes in the regulation ofenergy metabolism with aging. Physiol. Rev. 86, 651–667.
Rousset, S., del Mar Gonzalez-Barroso, M., Gelly, C., Pecqueur, C., Bouillaud, F., Ricquier,D., Cassard-Doulcier, A.M., 2002. A new polymorphic site located in the humanUCP1 gene controls the in vitro binding of CREB-like factor. Int. J. Obes. Relat.Metab. Disord. 26, 735–738.
Ruggiero, C., Metter, E.J., Melenovsky, V., Cherubini, A., Najjar, S.S., Ble, A., Senin, U.,Longo, D.L., Ferrucci, L., 2008. High basal metabolic rate is a risk factor for mortality:the Baltimore Longitudinal Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 63,698–706.
Saito, M., Okamatsu-Ogura, Y., Matsushita, M., Watanabe, K., Yoneshiro, T., Nio-Kobayashi, J., Iwanaga, T., Miyagawa, M., Kameya, T., Nakada, K., Kawai, Y., Tsujisaki,M., 2009. High incidence of metabolically active brown adipose tissue in healthyadult humans: effects of cold exposure and adiposity. Diabetes 58, 1526–1531.
Sale, M.M., Hsu, F.C., Palmer, N.D., Gordon, C.J., Keene, K.L., Borgerink, H.M., Sharma, A.J.,Bergman, R.N., Taylor, K.D., Saad, M.F., Norris, J.M., 2007. The uncoupling protein 1gene, UCP1, is expressed in mammalian islet cells and associated with acute insulinresponse to glucose in African American families from the IRAS Family Study. BMCEndocr. Disord. 7, 1–10.
Sell, H., Deshaies, Y., Richard, D., 2004. The brown adipocyte: update on its metabolicrole. Int. J. Biochem. Cell. Biol. 36, 2098–2104.
Son'kin, V.D., Kirdin, A.A., Andreev, R.S., Akimov, E.B., 2010. Homeostatic non-shiveringthermogenesis in humans facts and hypotheses. Hum. Physiol. 36, 599–614.
Sramkova, D., Krejbichova, S., Vcelak, J., Vankova, M., Samalikova, P., Hill, M.,Kvasnickova, H., Dvorakova, K., Vondra, K., Hainer, V., Bendlova, B., 2007. TheUCP1 gene polymorphism A-3826G in relation to DM2 and body composition inCzech population. Exp. Clin. Endocrinol. Diab. 115, 303–307.
St-Onge, M.P., Gallagher, D., 2010. Body composition changes with aging: the cause orthe result of alterations in metabolic rate and macronutrient oxidation? Nutrition26, 152–155.
Tan, Q., Yashin, A.I., De Benedictis, G., Cintolesi, F., Rose, G., Bonafe, M., Franceschi, C.,Vach, W., Vaupel, J.W., 2001. A logistic regression model for measuring gene–longevity associations. Clin. Genet. 60, 463–469.
Tiraby, C., Tavernier, G., Lefort, C., Larrouy, D., Bouillaud, F., Ricquier, D., Langin, D., 2003.Acquirement of brown fat cell features by human white adipocytes. J. Biol. Chem.278, 33370–33376.
Valle, A., Santandreu, F.M., Garcìa-Palmer, F.J., Roca, P., Oliver, J., 2008. The serumlevels of 17β-estradiol, progesterone and triiodothyronine correlate with brownadipose tissue thermogenic parameters during aging. Cell. Physiol. Biochem. 22,337–346.
van Marken Lichtenbelt, W.D., Vanhommerig, J.W., Smulders, N.M., Drossaerts, J.M.,Kemerink, G.J., Bouvy, N.D., Schrauwen, P., Teule, G.J., 2009. Cold activated brownadipose tissue in healthy men. N. Engl. J. Med. 360, 1500–1508.
Vijgen, G.H., Bouvy, N.D., Teule, G.J., Brans, B., Schrauwen, P., van Marken Lichtenbelt,W.D., 2011. Brown adipose tissue in morbidly obese subjects. PLoS ONE 6 (2),e17247.
Virtanen, K.A., Lidell, M.E., Orava, J., Heglind, M., Westergren, R., Niemi, T., Taittonen, M.,Laine, J., Savisto, N.J., Enerbäck, S., Nuutila, P., 2009. Functional brown adiposetissue in healthy adults. N. Engl. J. Med. 360, 1518–1525.
Waalen, J., Buxbaum, J.N., 2011. Is older colder or colder older? The association of agewith body temperature in 18,630 individuals. J. Gerontol. A Biol. Sci. Med. Sci. 66(5), 487–492.
Weir, B.S., 1996. Genetic Data Analysis II: Methods for Discrete Population Genetic Data.Sinauer Associates, Sunderland, Mass.
904 G. Rose et al. / Experimental Gerontology 46 (2011) 897–904
Wijers, S.L.J., Schrauwen, P., Saris, W.H.M., van Marken Lichtenbelt, W.D., 2008. Humanskeletal muscle mitochondrial uncoupling is associated with cold induced adaptivethermogenesis. PLoS ONE 3, e1777.
Yoneshiro, T., Aita, S., Matsushita, M., Kameya, T., Nakada, K., Kawai, Y., Saito, M., 2010.Brown adipose tissue, whole-body energy expenditure, and thermogenesis inhealthy adult men. Obesity 19, 13–16.
Zingaretti, M.C., Crosta, F., Vitali, A., Guerrieri, M., Frontini, A., Cannon, B., Nedergaard, J.,Cinti, S., 2009. The presence of UCP1 demonstrates that metabolically activeadipose tissue in the neck of adult humans truly represents brown adipose tissue.FASEB J. 23, 3113–3120.
Copyright © 2022 FDOKUMEN




















