
Regulation ofGaxHomeobox Gene Transcription by a Combination of Positive Factors Including...
11
1995, 15(8):4272. Mol. Cell. Biol. V Andrés, S Fisher, P Wearsch and K Walsh factor 2. factors including myocyte-specific enhancer transcription by a combination of positive Regulation of Gax homeobox gene http://mcb.asm.org/content/15/8/4272 Updated information and services can be found at: These include: CONTENT ALERTS more» cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new articles http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on February 12, 2014 by guest http://mcb.asm.org/ Downloaded from on February 12, 2014 by guest http://mcb.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Regulation ofGaxHomeobox Gene Transcription by a Combination of Positive Factors Including...

1995, 15(8):4272. Mol. Cell. Biol.
V Andrés, S Fisher, P Wearsch and K Walsh factor 2.factors including myocyte-specific enhancertranscription by a combination of positive Regulation of Gax homeobox gene
http://mcb.asm.org/content/15/8/4272Updated information and services can be found at:
These include:
CONTENT ALERTS more»cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new articles
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
on F
ebruary 12, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY, Aug. 1995, p. 4272–4281 Vol. 15, No. 80270-7306/95/$04.0010Copyright 1995, American Society for Microbiology
Regulation of Gax Homeobox Gene Transcription by a Combinationof Positive Factors Including Myocyte-Specific Enhancer Factor 2
VICENTE ANDRES,1 STEVE FISHER,2 PAM WEARSCH,† AND KENNETH WALSH*
Division of Cardiovascular Research, St. Elizabeth’s Medical Center, Tufts University School of Medicine,Boston, Massachusetts 02135,1 Department of Physiology and Biophysics,
Case Western Reserve University, Cleveland, Ohio 441062
Received 8 December 1994/Returned for modification 9 February 1995/Accepted 19 May 1995
Homeobox-containing genes play an essential role in basic processes during embryogenesis and develop-ment, but little is known about the regulation of their expression. To elucidate regulatory networks that governhomeobox gene expression, we defined the core promoter of the mouse Gax homeobox gene and characterizedits interactions with cellular proteins. Transient transfection experiments revealed Gax promoter activity inseveral cell types. Deletion analysis defined a 138-bp minimal promoter fragment between positions 2125 and113 relative to the transcription initiation site. Mutagenesis and protein-DNA binding assays suggested thatat least three positive factors interact with this fragment and are required for transcriptional activity. One ofthese factors, HRF-1, recognizes a cis element consisting of an inverted palindromic motif. A second factor isSp1, that binds to a G/C-rich element. The third is the MADS box factor referred to as MEF2 or RSRF.Mutations in the MEF2/RSRF site had the greatest effect on transcription in cell types that expressed thehighest levels of endogenous MEF2 activity. Conversely, overexpression of MEF2A transactivated the Gaxpromoter more efficiently in cells lacking endogenous MEF2. These data provide evidence for a direct tran-scriptional link between members of the MADS and homeobox families of transcription factors.
Homeobox genes control cell fate determination and forma-tion of the general body plan in vertebrates and invertebrates(25, 30, 41, 51). Homeobox genes encode transcription factorsthat contain a characteristic homeodomain, a 60-amino-acidDNA-binding domain that is conserved through evolution. Au-to- and cross-regulatory interactions between homeobox geneproducts have been shown to be essential for proper regulationof Drosophila homeobox genes (6, 30, 32). Similar regulatorynetworks between mammalian homeobox genes may also exist(3, 59), but relatively little is known about homeobox generegulation by non-homeobox transcription factors. It has beenshown that the zinc finger protein Krox-20 directly activatesthe expression of Hoxb-2 (Hox-2.8) in rhombomeres r3 and r5during hindbrain development (49, 54). Homeobox gene tran-scription is also regulated by retinoic acid, which functionsthrough retinoic acid-responsive cis elements (31, 37, 46, 52).Another class of developmentally important genes is the
MADS superfamily of transcriptional regulators, named forthe yeast MCM1 factor, two plant homeotic genes, agamousand deficiens, and the serum response factor (50). All of thesetranscription factors share the MADS box, a highly conservedDNA-binding and dimerization domain. Recently, a new fam-ily of MADS transcription factors, named RSRF (related toserum response factor) (45) or MEF2 (myocyte-specific en-hancer factor 2) (58), has been identified. MEF2 proteins areencoded in mammals by four genes, MEF2a, MEF2b, MEF2c,andMEF2d (7), which display a complex pattern of alternativesplicing (7, 34, 38–40, 45, 58). While MEF2 transcripts areubiquitous, MEF2 proteins appear to be restricted to skeletal,cardiac, and smooth muscle cells and the brain, suggesting thatposttranscriptional mechanisms regulate MEF2 expression (7,
34, 39, 40, 45, 53, 58). Remarkably, MEF2 can activate themyogenic differentiation program when ectopically expressedin nonmuscle cells (26). The myogenic commitment may result,at least in part, from the ability of MEF2 to transcriptionallyactivate the myogenic bHLH factors by binding to regulatoryelements in their promoters (9, 14, 33, 57). All MEF2 proteinsbind to and activate transcription through a consensus CTA(A/T)4TA(G/A) binding site that occurs in many promoters andenhancers of muscle-specific and growth factor-regulatedgenes (13, 45).Previously, we identified a homeobox gene, Gax, which is
widely expressed in developing mouse embryos, including me-sodermal and ectodermal tissues, but becomes largely confinedto some cardiovascular tissues of adults (20). To identify up-stream regulators of homeobox genes, we have isolated andcharacterized the promoter of the mouse Gax gene. We showhere that at least three positive factors bind to specific se-quences within a minimal promoter fragment. One of thesefactors, MEF2, transactivates the Gax promoter in a site-de-pendent manner. These observations provide insights into ho-meotic gene regulatory networks by demonstrating a directtranscriptional link between a homeobox gene and a memberof the MADS family of transcription factors.
MATERIALS AND METHODS
Molecular cloning of mouse Gax genomic clones and sequence analysis. Amouse genomic library (M129) in the lFIX vector (Stratagene) was screenedwith two oligonucleotide probes complementary to the rat Gax cDNA (20),positions 71 to 110 and 284 to 340, respectively. A total of 1.53 106 phages wereplated, and plaques were lifted in quadruplicate onto nitrocellulose membranes.Pairs of membranes were hybridized at 608C for 16 h in 1% bovine serumalbumin–1 mM EDTA–7% sodium dodecyl sulfate (SDS)–0.5 M NaPO4 (pH7.5) with probes end labeled with [g-32P]ATP. Washes were initially performedat room temperature with 33 SSC (13 SSC is 0.15 M NaCl plus 0.015 M sodiumcitrate)–1% SDS and then at 608C in 13 SSC–1% SDS and then in 0.53SSC–1% SDS. The screening identified two plaques positive for both probes, onall four lifts, that were subsequently purified to homogeneity by three additionalrounds of screening in the manner described above. Restriction mapping indi-cated that both clones had an identical ;15.8-kb insert. Southern blot analysis
* Corresponding author. Mailing address: Division of Cardiovascu-lar Research, St. Elizabeth’s Medical Center, Tufts University Schoolof Medicine, 736 Cambridge St., Boston, MA 02135.† Present address: Department of Cell Biology, Duke University
Medical Center, Durham, NC 27710.
4272
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

revealed that a;5.3 kb XbaI restriction fragment contained the sequences of theoligonucleotide probes used for the screening. pB-Hx-1 was constructed bysubcloning this XbaI genomic fragment into pBluescriptIIKS1 (Stratagene).pB-Hx-1EcoD and pB-Hx-1HindD were generated by religating pB-Hx-1 afterdigestion with EcoRI and HindIII, respectively. Gax genomic sequences wereanalyzed with Sequenase (U.S. Biochemicals).Cell culture and construction and transfection of reporter genes. Rat aorta
embryonic A10 cells (American Type Culture Collection) and C3H mouse me-sodermal embryonic 10T1⁄2 cells (clone 8) (47) were passed in Dulbecco’s mod-ified Eagle’s medium supplemented with 10% (vol/vol) fetal calf serum (FCS).C2C12 myogenic cells (American Type Culture Collection) were passed in Dul-becco’s modified Eagle’s medium containing 20% FCS (growth medium). Toinduce differentiation, C2C12 cells were transferred to Dulbecco’s modifiedEagle’s medium supplemented with 2% heat-inactivated horse serum (differen-tiation medium). Primary rat aorta cells were isolated and grown essentially asdescribed by Pickering et al. (44). Transient transfection assays were performedwith plasmid DNA prepared with Qiagen columns (Qiagen Inc.). Unless other-wise stated, cells were transfected with N-[1-(2,3-dioleoyloxy)propyl]-N,N,N-tri-methylammoniummethylsulfate (DOTAP; Boehringer Mannheim). Cells weregrown to 60 to 80% confluence in 100-mm-diameter dishes, and the DNA-DOTAP mixture (typically, 10 mg of DNA–60 mg of DOTAP) was added to cellsfed 5 ml of Opti-MEM I (Gibco, BRL) containing 10% FCS. After 4 to 8 h, cellswere washed with Opti-MEM I and fed 10 ml of 10% FCS in Opti-MEM I. Cellswere harvested 48 to 72 h after addition of DNA-DOTAP. Because of problemsof cytoxicity with DOTAP, C2C12 cells were transfected by the calcium phos-phate method (19). C2C12 cells were maintained in growth medium throughoutthe transfection or transferred to differentiation medium for the last 2 days afterglycerol shock. Differences in transfection efficiency were corrected relative tothe level of alkaline phosphatase activity produced from a cotransfected internalcontrol plasmid (pSVAPAP; 0.2 to 1 mg per transfection), which contains thereporter gene under control of the simian virus 40 promoter-enhancer (24).Luciferase (Promega) and alkaline phosphatase (CSPD chemiluminescent sub-strate; Tropix) activities were measured with a Berthold Lumat LB9501 lumi-nometer. Each transfection was independently repeated as indicated in the figurelegends, and the mean 6 the standard error of the mean was plotted.To construct Gax-luciferase reporter genes, different Gax genomic fragments
were subcloned into the multiple cloning region of pGL2-Basic (Promega), apromoterless vector containing the luciferase gene. The chimeric plasmids weredesignated pGax(x/x)-L, where x indicates the coordinates of the Gax sequencerelative to the transcription initiation site. To construct pGax(23829/113)-L andpGax(21276/113)-L, the KpnI-PvuII inserts from pB-Hx-1 and pB-Hx-1HindD,respectively, were subcloned into pGL2-Basic vector which had been digestedwith XhoI, filled in with the Klenow enzyme, and digested with KpnI.pGax(2652/113)-L was generated by digestion of pGax(21276/113)-L withNheI-Asp718, followed by filling in and religation. Finally, pGax(23829/2396)-Lwas constructed by subcloning the KpnI-BglII insert from pB-Hx-1 into equallydigested pGL2-Basic. Other 59 deletion mutants were generated by the PCR withGax DNA templates and Gax-specific 59 primers containing a KpnI site and R1,a 39 primer containing a BglII site and nucleotides complementary to positions113 to 29 from the Gax gene (59-gccagatctCTGCTCAGATGTCAAGTGTGG-39). For construction of pGax(2125/113)-L site-directed mutants, the corre-sponding Gax genomic fragments were generated by PCR with R1 and thefollowing 59 primers that contained the desired mutations (mutated bases areunderlined): 59-GCCGGTACCAGCTTTAAATCCTAAGCCAGCCCCCAGC-39(L mt) 59-GCCGGTACCAGCGCTGGCTCCTAATTTAAACCCCAGC-39 (R/Sp1 mt), and 59-GCCGGTACCAGCGCTGGCTCCTAAGCCAGCAAACAGCTCAC-39 (Sp1 mt). The PCR products were phenol extracted, digested withKpnI and BglII, and subcloned into equally digested pGL2-Basic.To construct pGax(2652/113)-L T1M2 (MEF2 mutant), PCR products A
and B were generated with primers L1-R3 and L6-R1, respectively: L1, 59-GCCGGTACCGCTTAGCCTTGGTGCACAGC-39 (KpnI site); L6, 59-GCGAATTCTATAACCTCTCAGAGCC-39 (EcoRI site); R3, 59-GCGAATTCGTGTAATGTGAGCTGG-39 (EcoRI site). The PCR products were phenol extracted,digested with KpnI-EcoRI (PCR A) and EcoRI-BglII (PCR B), and ligated intoKpnI-BglII-digested pGL2-Basic. pGax(2652/113)-L T2M2 (TATA and MEF2mutant) was constructed similarly but with primers L1/R2 (PCR A) and L5/R1(PCR B): L5, 59-GCGAATTCTAACCTCTCAGAGCCATTTCC-39 (EcoRIsite); R2, 59-GCGAATTCGAGTGTAATGTGAGCTGG-39 (EcoRI site). Allreporter plasmids generated by PCR were verified by DNA sequencing. ThepMT2-MEF2A expression vector has been described by Yu et al. (58).Other reporter genes were derived from D56-luciferase, which contains the
c-fos minimal promoter, positions 256 to 1109, linked to the promoterlessluciferase gene (48). MEF2D56 has one copy of the muscle creatine kinase(MCK) enhancer MEF2 site upstream from the c-fos minimal promoter (53).TATA2D56, in which the c-fos TATA box has been replaced with a NotI site(GCGGCCGC), was constructed by cloning the following oligonucleotide inSalI-PvuII-digested D56-luciferase (mutated bases are underlined):
59-TCGACAATCCATCCATTCACAGCGCTTGCGGCCGCGCGCCAG-3939-GTTAGGTAGGTAAGTGTCGCGAACGCCGGCGCGCGGTC-59
To construct other D56-derived reporters, oligonucleotides with SalI-NotIoverhangs flanking the wild-type MEF2/TATA box from the Gax promoter(T1M1D56), or mutated derivatives that destroyed the MEF2 site (T1M2D56)or both the MEF2 and TATA-binding protein (TBP) sites (T2M2D56), werecloned in SalI-NotI-digested TATA2D56 (see the diagram in Fig. 9). All D56-derived reporter plasmids were verified by DNA sequencing.Transfection of embryonic chicken hearts in vivo. Stage 13 to 17 embryonic
chicken hearts were transfected with plasmid DNA as previously described, withminor modifications (15). In brief, fertilized White Leghorn chicken eggs wereobtained from Case Western Reserve University’s Squire Valleevue farm andincubated at 388C for 64 h. The entire egg contents were drained into a shallowwell of Saran Wrap within a plastic cup. One microgram of DNA in a volume of4 ml of water was mixed with 6 ml of Lipofectamine reagent (GIBCO, BRL).Under stereoscopic guidance, a small hole was cut in the vitelline membranesurrounding the heart, and ;1 ml of the DNA solution (100 ng of plasmid) wasdelivered via a 30-mm-bore micropipette onto the surface of the beating heart.The embryo was incubated for an additional 24 h, and then the heart wasseparated from the body and placed into 30 ml of a 1% Triton X-100-containinglysis buffer (Promega). The tissue was homogenized by trituration and brieflycentrifuged to remove debris. Ten microliters of the homogenate was used forluciferase assay (see above). Samples were typically assayed twice, and thedifference between the two determinations was less than 10%. A small percent-age of embryos had activity of less than twice the background in groups thatusually had activity. These embryos presumably did not transfect efficiently andwere excluded from analysis. It should be noted that under the conditions oftransfection, the embryos developed normally and the hearts increased in massby fivefold over the 24-h incubation period following DNA delivery.Primer extension analysis. Cellular RNA was isolated from A10 cells tran-
siently transfected with pGax(2174/113)-L by using RNAzol B (Biotecx Labo-ratories, Inc.). Cells were also cotransfected with 10 mg of pGax(23829/113)-Land 1 mg of pSV-neo, and RNA was prepared from the pool of neomycin-resistant cells selected with G418 (0.125 mg/ml; GIBCO, BRL). Analysis wasperformed on 35 mg of RNA with the Primer Extension M-MLV ReverseTranscriptase System (Promega) and a primer specific for the luciferase cDNA(59-GTCTAGAGGATAGAATGGC-39). The reaction products were electro-phoresed on a 7 M urea–6% (wt/vol) polyacrylamide gel, and the dried gel wasanalyzed with a Phosphorimager (Molecular Dynamics).Whole-cell extracts and electrophoretic mobility shift assays (EMSA). Syn-
thetic oligonucleotides used for EMSA were end labeled with [g-32P]ATP and T4polynucleotide kinase. For EMSA involving TBP,;400,000 cpm of probe (250 to650 pmol) and 20 ng of human recombinant TBP (Promega) were incubated andelectrophoresed as suggested by the manufacturer. Where indicated, an unla-beled competitor was added prior to TBP.The preparation of whole-cell extracts and the procedure for EMSA involving
cellular proteins were essentially as previously described (2), except that thebinding buffer was 40 mM N-2-hydroxyethylpiperazine-N9-2-ethanesulfonic acid(HEPES)–KOH (pH 7.8)–40 mM KCl–20% (wt/vol) glycerol–2 mM dithiothrei-tol–0.4 mM EDTA–1 mg of bovine serum albumin per ml. For EMSA involvingaddition of anti-Sp1 antibodies (Santa Cruz Biotech), 2 mg of antibody was addedafter incubation of the probe and extract, and the reaction was incubated on icefor an additional 20 min. Binding reactions were loaded onto a 6% (wt/vol)polyacrylamide gel (80:1 acrylamide/bisacrylamide ratio) and electrophoresed at200 V in 0.53 TBE (0.045 M Tris-borate, 0.001 M EDTA [pH 7.5]) with acooling system. The gel was then dried and exposed to X-ray film.
RESULTS
Isolation of mouse Gax 5 -flanking sequences. As a first steptoward the identification ofGax regulatory sequences, a mousegenomic library was screened with two oligonucleotide probesspanning positions 71 to 110 and 284 to 340 of the rat GaxcDNA (20). Two identical phage clones were isolated whichcontained an ;15.8-kb insert, and a 5,349-bp XbaI fragmentspanning the promoter and the cDNA sequences was identi-fied, subcloned, and sequenced (see Material and Methods).Figure 1A shows the sequence of the proximal promoter re-gion, the 59 untranslated region, and the beginning of thecoding sequence. We then performed primer extension analy-sis of cellular RNA to map the transcription initiation site.Initial attempts with poly(A)1 RNA templates from mouseembryos and adult tissues using Gax-specific primers were notsuccessful, perhaps because of the low abundance of Gax tran-scripts. Therefore, we isolated RNA from A10 cells transientlyand permanently transfected with reporter genes containingthe Gax promoter regions from positions 2174 to 113 and23829 to 113, respectively, fused to a promoterless luciferasereporter gene. Primer extension reactions were carried out
VOL. 15, 1995 HOMEOBOX GENE REGULATION BY MEF2 4273
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
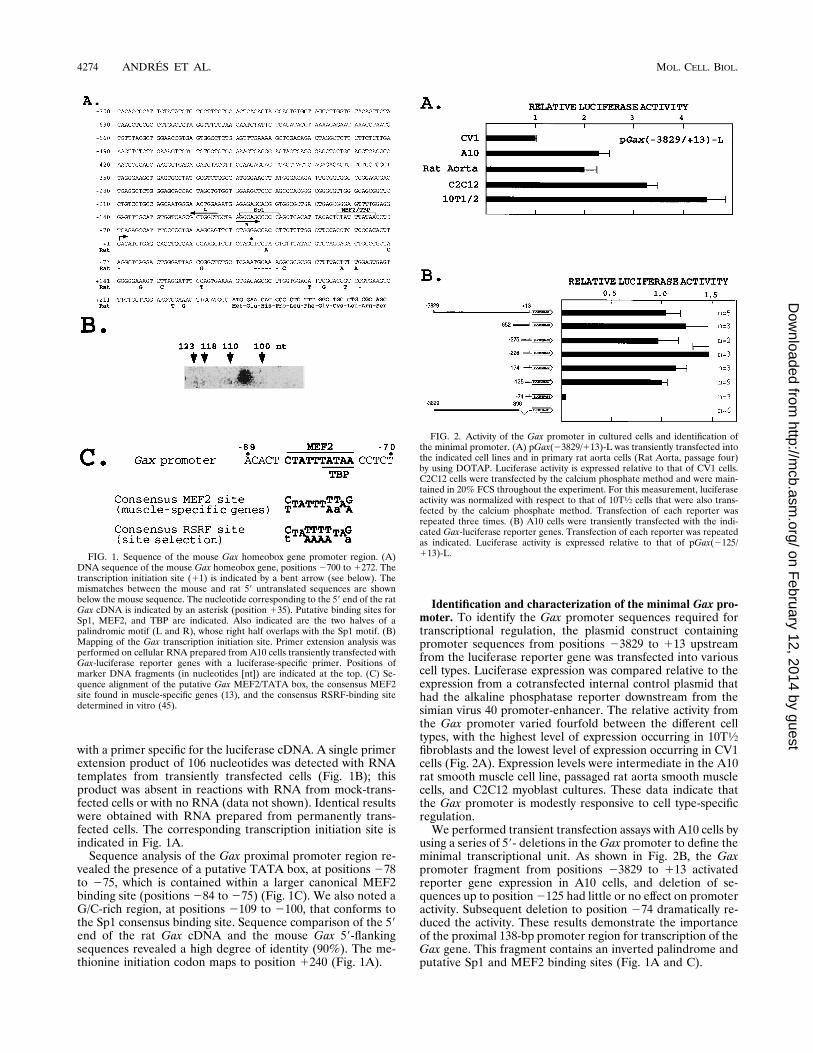
with a primer specific for the luciferase cDNA. A single primerextension product of 106 nucleotides was detected with RNAtemplates from transiently transfected cells (Fig. 1B); thisproduct was absent in reactions with RNA from mock-trans-fected cells or with no RNA (data not shown). Identical resultswere obtained with RNA prepared from permanently trans-fected cells. The corresponding transcription initiation site isindicated in Fig. 1A.Sequence analysis of the Gax proximal promoter region re-
vealed the presence of a putative TATA box, at positions 278to 275, which is contained within a larger canonical MEF2binding site (positions 284 to 275) (Fig. 1C). We also noted aG/C-rich region, at positions 2109 to 2100, that conforms tothe Sp1 consensus binding site. Sequence comparison of the 59end of the rat Gax cDNA and the mouse Gax 59-flankingsequences revealed a high degree of identity (90%). The me-thionine initiation codon maps to position 1240 (Fig. 1A).
Identification and characterization of the minimal Gax pro-moter. To identify the Gax promoter sequences required fortranscriptional regulation, the plasmid construct containingpromoter sequences from positions 23829 to 113 upstreamfrom the luciferase reporter gene was transfected into variouscell types. Luciferase expression was compared relative to theexpression from a cotransfected internal control plasmid thathad the alkaline phosphatase reporter downstream from thesimian virus 40 promoter-enhancer. The relative activity fromthe Gax promoter varied fourfold between the different celltypes, with the highest level of expression occurring in 10T1⁄2fibroblasts and the lowest level of expression occurring in CV1cells (Fig. 2A). Expression levels were intermediate in the A10rat smooth muscle cell line, passaged rat aorta smooth musclecells, and C2C12 myoblast cultures. These data indicate thatthe Gax promoter is modestly responsive to cell type-specificregulation.We performed transient transfection assays with A10 cells by
using a series of 59- deletions in theGax promoter to define theminimal transcriptional unit. As shown in Fig. 2B, the Gaxpromoter fragment from positions 23829 to 113 activatedreporter gene expression in A10 cells, and deletion of se-quences up to position2125 had little or no effect on promoteractivity. Subsequent deletion to position 274 dramatically re-duced the activity. These results demonstrate the importanceof the proximal 138-bp promoter region for transcription of theGax gene. This fragment contains an inverted palindrome andputative Sp1 and MEF2 binding sites (Fig. 1A and C).
FIG. 1. Sequence of the mouse Gax homeobox gene promoter region. (A)DNA sequence of the mouse Gax homeobox gene, positions 2700 to 1272. Thetranscription initiation site (11) is indicated by a bent arrow (see below). Themismatches between the mouse and rat 59 untranslated sequences are shownbelow the mouse sequence. The nucleotide corresponding to the 59 end of the ratGax cDNA is indicated by an asterisk (position 135). Putative binding sites forSp1, MEF2, and TBP are indicated. Also indicated are the two halves of apalindromic motif (L and R), whose right half overlaps with the Sp1 motif. (B)Mapping of the Gax transcription initiation site. Primer extension analysis wasperformed on cellular RNA prepared from A10 cells transiently transfected withGax-luciferase reporter genes with a luciferase-specific primer. Positions ofmarker DNA fragments (in nucleotides [nt]) are indicated at the top. (C) Se-quence alignment of the putative Gax MEF2/TATA box, the consensus MEF2site found in muscle-specific genes (13), and the consensus RSRF-binding sitedetermined in vitro (45).
FIG. 2. Activity of the Gax promoter in cultured cells and identification ofthe minimal promoter. (A) pGax(23829/113)-L was transiently transfected intothe indicated cell lines and in primary rat aorta cells (Rat Aorta, passage four)by using DOTAP. Luciferase activity is expressed relative to that of CV1 cells.C2C12 cells were transfected by the calcium phosphate method and were main-tained in 20% FCS throughout the experiment. For this measurement, luciferaseactivity was normalized with respect to that of 10T1⁄2 cells that were also trans-fected by the calcium phosphate method. Transfection of each reporter wasrepeated three times. (B) A10 cells were transiently transfected with the indi-cated Gax-luciferase reporter genes. Transfection of each reporter was repeatedas indicated. Luciferase activity is expressed relative to that of pGax(2125/113)-L.
4274 ANDRES ET AL. MOL. CELL. BIOL.
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

To further characterize the minimal Gax promoter, we ex-amined the promoter activities of a series of unidirectional 59deletion mutations with endpoints clustered within the2125 to113 fragment. The results of these experiments are shown inFig. 3. The activity of the 2110 to 113 and 2109 to 113fragments in A10 cells was ;25 to 30% of the activity of the2125 to113 fragment. It is of note that these deletion mutantslack the left side of an inverted palindrome (designated L).The functional relevance of the Sp1 binding site at positions2109 to 2100 was suggested by the further ;50% decrease inthe promoter activity of pGax(2104/113)-L relative to theactivity of pGax(2109/113)-L. Results similar to those de-scribed above were obtained in transfection assays performedwith primary cultures derived from rat aorta tissue and withdeveloping chick hearts, in which deletion of the sequences
containing the inverted palindrome and the putative Sp1 bind-ing site significantly reduced reporter gene expression. How-ever, one notable difference was the finding that pGax(284/113)-L, which contains the putative MEF2 site, was sevenfoldmore active than pGax(274/113)-L in developing chick hearttissue, but in transfected smooth muscle cells, both of theseconstructs produced similar low levels of reporter expression.To directly assess the functional role of the putative cis
elements within the promoter region from positions 2125 to275, we performed site-directed mutational analyses. A seriesof five mutations were tested in transfected A10 cells (Fig. 4).All mutations resulted in some degree of decreased activityrelative to the wild-type constructs. The largest effect on tran-scription was observed when the overlapping R/Sp1 elementwas mutated (Fig. 4, R/Sp1 mt and Sp1 mt). We also examinedthe effect of two mutations within the putative MEF2/TATAmotif. These mutations were designated T1M2 and T2M2,depending on whether the mutation altered the MEF2 motif orboth sequence motifs, respectively. Each mutation decreasedexpression by a factor of 2. Taken together, the above resultsdemonstrate that the activity of the minimal Gax promoterrequires the interaction of several positive factors with differ-ent cis-acting elements within the sequence from positions2125 to 275. These elements include an inverted palindromicmotif which overlaps a canonical Sp1 box and an A/T-richelement that conforms to the consensus binding site for bothMEF2 and TBP.Identification of factors that interact with the minimal Gax
promoter.We next performed EMSA with radiolabeled probesspanning theGax promoter region from positions2125 to275to identify the proteins that specifically bind to this region.First, we examined the cellular proteins that interact with theinverted palindromic motif located at positions 2121 to 2104by using the Gax-1 probe (Fig. 5A). As shown in Fig. 5B, amajor nucleoprotein complex was detected with extracts fromA10 cells (lane 4). We dubbed this DNA-binding activityHRF-1 (for homeobox-regulating factor 1). The apparentDNA-binding affinity of HRF-1 decreased significantly wheneither half of the palindrome was mutated in the Gax-1 probe(L mt and R mt; compare lanes 4 to 6), suggesting that HRF-1interacts with both elements. To rule out the involvement ofsequences flanking the palindromic motif, we performed
FIG. 3. Deletion analysis of the 2125 to 113 Gax minimal promoter. Thesequence of the 59 end of the Gax-luciferase reporter genes is shown at the top.Putative cis elements for Sp1, MEF2, and TBP are boxed. L and R refer to theleft and right halves of the inverted palindromic motif, respectively (see text).Transient transfections were performed with A10 cells, primary rat aorta cells(passage 4), and embryonic chicken hearts (see Materials and Methods fordetails). Luciferase activity is expressed relative to that of pGax(2125/113)-L.Transfection of each reporter was repeated as indicated. SMCs, smooth musclecells.
FIG. 4. Site-directed mutagenesis of the minimal Gax promoter fragment. A10 cells were transfected with the indicated Gax-luciferase reporter gene mutants.Mutations within the 2125 to 113 Gax genomic sequence affected the left half of the palindromic motif (L mt), the right half and the Sp1 motif (R/Sp1mt), and theSp1 motif (Sp1 mt). Mutations affecting the MEF2 consensus binding site (T1M2mt) or both the MEF2 and TATA elements (T2M2mt) were introduced into the2652to 113 construct. Luciferase activity is expressed relative to that of the corresponding wild-type plasmid, which was taken as 1. Transfection of each reporter wasrepeated as indicated.
VOL. 15, 1995 HOMEOBOX GENE REGULATION BY MEF2 4275
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
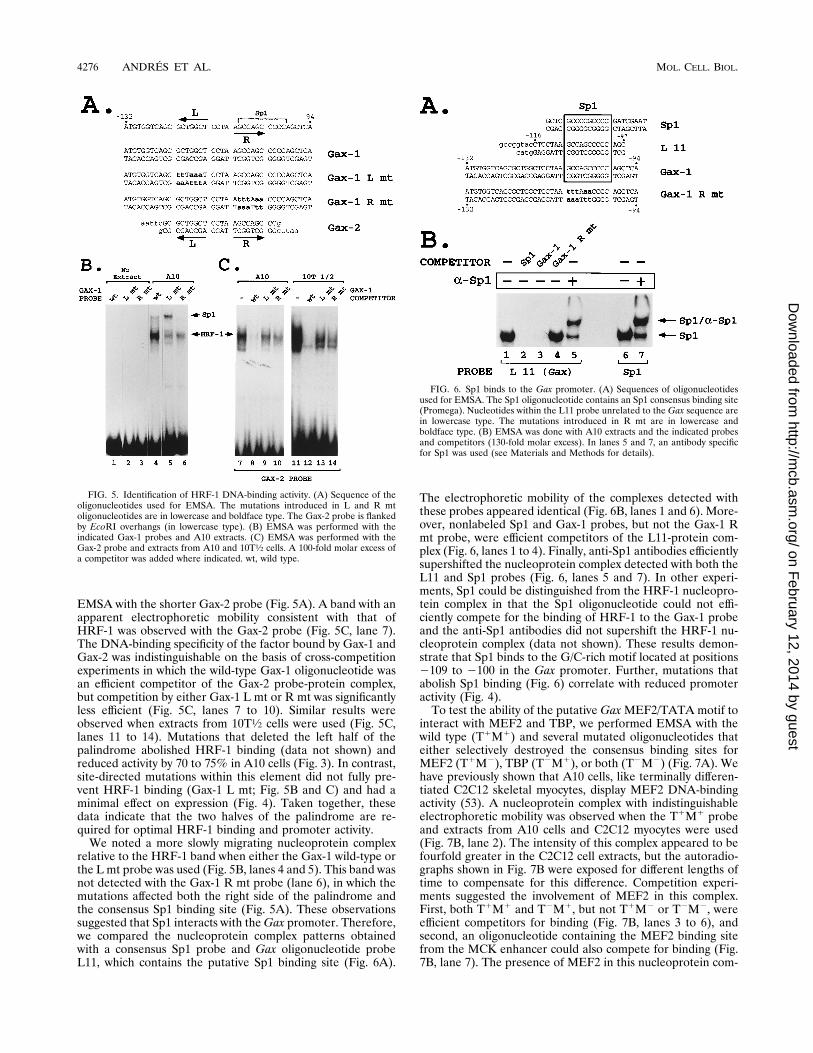
EMSA with the shorter Gax-2 probe (Fig. 5A). A band with anapparent electrophoretic mobility consistent with that ofHRF-1 was observed with the Gax-2 probe (Fig. 5C, lane 7).The DNA-binding specificity of the factor bound by Gax-1 andGax-2 was indistinguishable on the basis of cross-competitionexperiments in which the wild-type Gax-1 oligonucleotide wasan efficient competitor of the Gax-2 probe-protein complex,but competition by either Gax-1 L mt or R mt was significantlyless efficient (Fig. 5C, lanes 7 to 10). Similar results wereobserved when extracts from 10T1⁄2 cells were used (Fig. 5C,lanes 11 to 14). Mutations that deleted the left half of thepalindrome abolished HRF-1 binding (data not shown) andreduced activity by 70 to 75% in A10 cells (Fig. 3). In contrast,site-directed mutations within this element did not fully pre-vent HRF-1 binding (Gax-1 L mt; Fig. 5B and C) and had aminimal effect on expression (Fig. 4). Taken together, thesedata indicate that the two halves of the palindrome are re-quired for optimal HRF-1 binding and promoter activity.We noted a more slowly migrating nucleoprotein complex
relative to the HRF-1 band when either the Gax-1 wild-type orthe L mt probe was used (Fig. 5B, lanes 4 and 5). This band wasnot detected with the Gax-1 R mt probe (lane 6), in which themutations affected both the right side of the palindrome andthe consensus Sp1 binding site (Fig. 5A). These observationssuggested that Sp1 interacts with theGax promoter. Therefore,we compared the nucleoprotein complex patterns obtainedwith a consensus Sp1 probe and Gax oligonucleotide probeL11, which contains the putative Sp1 binding site (Fig. 6A).
The electrophoretic mobility of the complexes detected withthese probes appeared identical (Fig. 6B, lanes 1 and 6). More-over, nonlabeled Sp1 and Gax-1 probes, but not the Gax-1 Rmt probe, were efficient competitors of the L11-protein com-plex (Fig. 6, lanes 1 to 4). Finally, anti-Sp1 antibodies efficientlysupershifted the nucleoprotein complex detected with both theL11 and Sp1 probes (Fig. 6, lanes 5 and 7). In other experi-ments, Sp1 could be distinguished from the HRF-1 nucleopro-tein complex in that the Sp1 oligonucleotide could not effi-ciently compete for the binding of HRF-1 to the Gax-1 probeand the anti-Sp1 antibodies did not supershift the HRF-1 nu-cleoprotein complex (data not shown). These results demon-strate that Sp1 binds to the G/C-rich motif located at positions2109 to 2100 in the Gax promoter. Further, mutations thatabolish Sp1 binding (Fig. 6) correlate with reduced promoteractivity (Fig. 4).To test the ability of the putative GaxMEF2/TATA motif to
interact with MEF2 and TBP, we performed EMSA with thewild type (T1M1) and several mutated oligonucleotides thateither selectively destroyed the consensus binding sites forMEF2 (T1M2), TBP (T2M1), or both (T2M2) (Fig. 7A). Wehave previously shown that A10 cells, like terminally differen-tiated C2C12 skeletal myocytes, display MEF2 DNA-bindingactivity (53). A nucleoprotein complex with indistinguishableelectrophoretic mobility was observed when the T1M1 probeand extracts from A10 cells and C2C12 myocytes were used(Fig. 7B, lane 2). The intensity of this complex appeared to befourfold greater in the C2C12 cell extracts, but the autoradio-graphs shown in Fig. 7B were exposed for different lengths oftime to compensate for this difference. Competition experi-ments suggested the involvement of MEF2 in this complex.First, both T1M1 and T2M1, but not T1M2 or T2M2, wereefficient competitors for binding (Fig. 7B, lanes 3 to 6), andsecond, an oligonucleotide containing the MEF2 binding sitefrom the MCK enhancer could also compete for binding (Fig.7B, lane 7). The presence of MEF2 in this nucleoprotein com-
FIG. 5. Identification of HRF-1 DNA-binding activity. (A) Sequence of theoligonucleotides used for EMSA. The mutations introduced in L and R mtoligonucleotides are in lowercase and boldface type. The Gax-2 probe is flankedby EcoRI overhangs (in lowercase type). (B) EMSA was performed with theindicated Gax-1 probes and A10 extracts. (C) EMSA was performed with theGax-2 probe and extracts from A10 and 10T1⁄2 cells. A 100-fold molar excess ofa competitor was added where indicated. wt, wild type.
FIG. 6. Sp1 binds to the Gax promoter. (A) Sequences of oligonucleotidesused for EMSA. The Sp1 oligonucleotide contains an Sp1 consensus binding site(Promega). Nucleotides within the L11 probe unrelated to the Gax sequence arein lowercase type. The mutations introduced in R mt are in lowercase andboldface type. (B) EMSA was done with A10 extracts and the indicated probesand competitors (130-fold molar excess). In lanes 5 and 7, an antibody specificfor Sp1 was used (see Materials and Methods for details).
4276 ANDRES ET AL. MOL. CELL. BIOL.
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
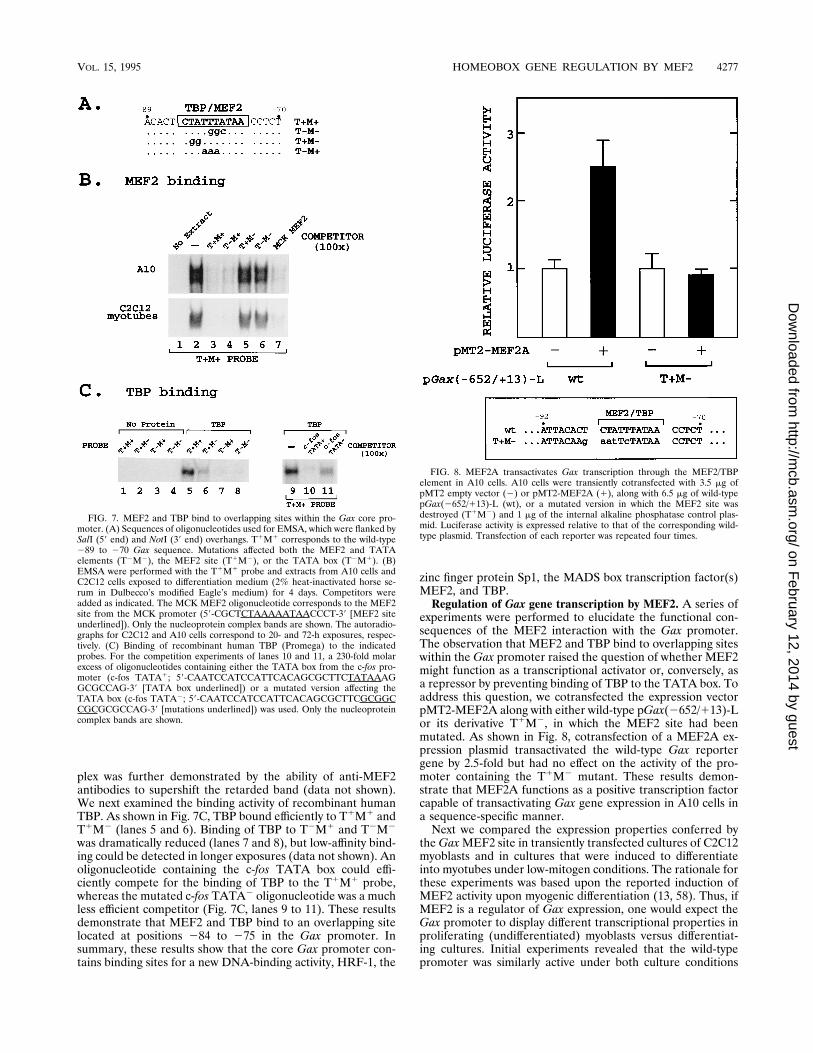
plex was further demonstrated by the ability of anti-MEF2antibodies to supershift the retarded band (data not shown).We next examined the binding activity of recombinant humanTBP. As shown in Fig. 7C, TBP bound efficiently to T1M1 andT1M2 (lanes 5 and 6). Binding of TBP to T2M1 and T2M2
was dramatically reduced (lanes 7 and 8), but low-affinity bind-ing could be detected in longer exposures (data not shown). Anoligonucleotide containing the c-fos TATA box could effi-ciently compete for the binding of TBP to the T1M1 probe,whereas the mutated c-fos TATA2 oligonucleotide was a muchless efficient competitor (Fig. 7C, lanes 9 to 11). These resultsdemonstrate that MEF2 and TBP bind to an overlapping sitelocated at positions 284 to 275 in the Gax promoter. Insummary, these results show that the core Gax promoter con-tains binding sites for a new DNA-binding activity, HRF-1, the
zinc finger protein Sp1, the MADS box transcription factor(s)MEF2, and TBP.Regulation of Gax gene transcription by MEF2. A series of
experiments were performed to elucidate the functional con-sequences of the MEF2 interaction with the Gax promoter.The observation that MEF2 and TBP bind to overlapping siteswithin the Gax promoter raised the question of whether MEF2might function as a transcriptional activator or, conversely, asa repressor by preventing binding of TBP to the TATA box. Toaddress this question, we cotransfected the expression vectorpMT2-MEF2A along with either wild-type pGax(2652/113)-Lor its derivative T1M2, in which the MEF2 site had beenmutated. As shown in Fig. 8, cotransfection of a MEF2A ex-pression plasmid transactivated the wild-type Gax reportergene by 2.5-fold but had no effect on the activity of the pro-moter containing the T1M2 mutant. These results demon-strate that MEF2A functions as a positive transcription factorcapable of transactivating Gax gene expression in A10 cells ina sequence-specific manner.Next we compared the expression properties conferred by
theGaxMEF2 site in transiently transfected cultures of C2C12myoblasts and in cultures that were induced to differentiateinto myotubes under low-mitogen conditions. The rationale forthese experiments was based upon the reported induction ofMEF2 activity upon myogenic differentiation (13, 58). Thus, ifMEF2 is a regulator of Gax expression, one would expect theGax promoter to display different transcriptional properties inproliferating (undifferentiated) myoblasts versus differentiat-ing cultures. Initial experiments revealed that the wild-typepromoter was similarly active under both culture conditions
FIG. 7. MEF2 and TBP bind to overlapping sites within the Gax core pro-moter. (A) Sequences of oligonucleotides used for EMSA, which were flanked bySalI (59 end) and NotI (39 end) overhangs. T1M1 corresponds to the wild-type289 to 270 Gax sequence. Mutations affected both the MEF2 and TATAelements (T2M2), the MEF2 site (T1M2), or the TATA box (T2M1). (B)EMSA were performed with the T1M1 probe and extracts from A10 cells andC2C12 cells exposed to differentiation medium (2% heat-inactivated horse se-rum in Dulbecco’s modified Eagle’s medium) for 4 days. Competitors wereadded as indicated. The MCK MEF2 oligonucleotide corresponds to the MEF2site from the MCK promoter (59-CGCTCTAAAAATAACCCT-39 [MEF2 siteunderlined]). Only the nucleoprotein complex bands are shown. The autoradio-graphs for C2C12 and A10 cells correspond to 20- and 72-h exposures, respec-tively. (C) Binding of recombinant human TBP (Promega) to the indicatedprobes. For the competition experiments of lanes 10 and 11, a 230-fold molarexcess of oligonucleotides containing either the TATA box from the c-fos pro-moter (c-fos TATA1; 59-CAATCCATCCATTCACAGCGCTTCTATAAAGGCGCCAG-39 [TATA box underlined]) or a mutated version affecting theTATA box (c-fos TATA2; 59-CAATCCATCCATTCACAGCGCTTCGCGGCCGCGCGCCAG-39 [mutations underlined]) was used. Only the nucleoproteincomplex bands are shown.
FIG. 8. MEF2A transactivates Gax transcription through the MEF2/TBPelement in A10 cells. A10 cells were transiently cotransfected with 3.5 mg ofpMT2 empty vector (2) or pMT2-MEF2A (1), along with 6.5 mg of wild-typepGax(2652/113)-L (wt), or a mutated version in which the MEF2 site wasdestroyed (T1M2) and 1 mg of the internal alkaline phosphatase control plas-mid. Luciferase activity is expressed relative to that of the corresponding wild-type plasmid. Transfection of each reporter was repeated four times.
VOL. 15, 1995 HOMEOBOX GENE REGULATION BY MEF2 4277
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
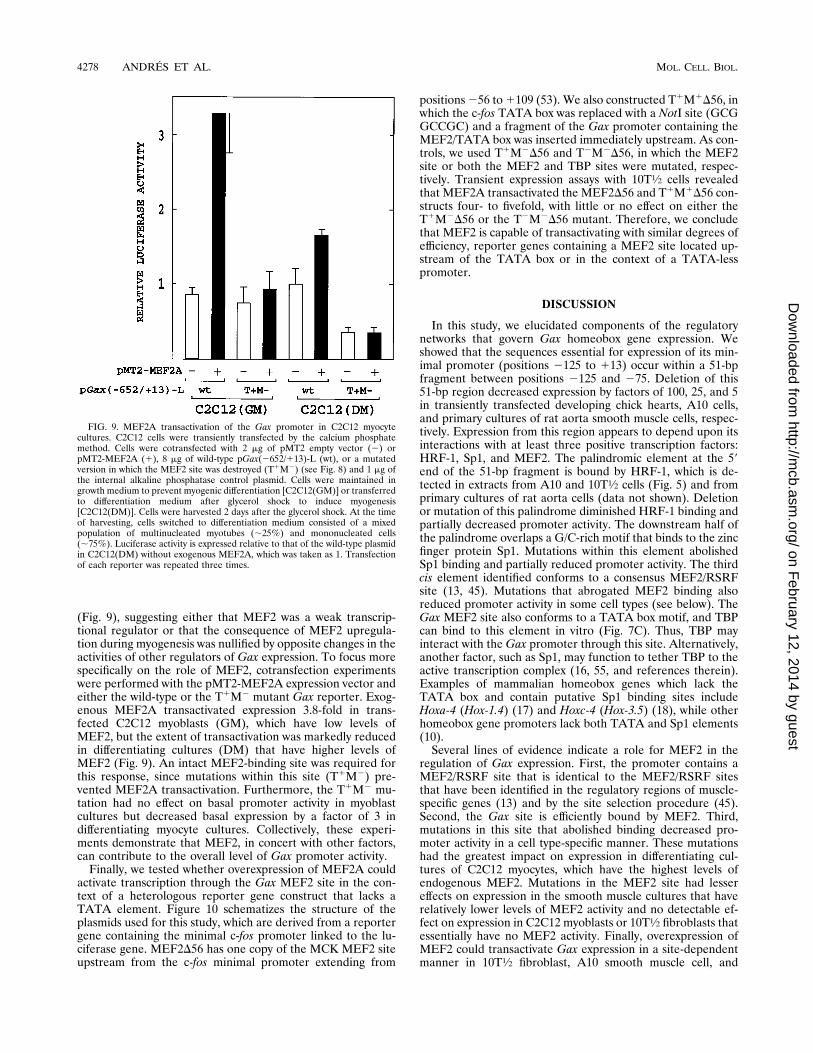
(Fig. 9), suggesting either that MEF2 was a weak transcrip-tional regulator or that the consequence of MEF2 upregula-tion during myogenesis was nullified by opposite changes in theactivities of other regulators of Gax expression. To focus morespecifically on the role of MEF2, cotransfection experimentswere performed with the pMT2-MEF2A expression vector andeither the wild-type or the T1M2 mutant Gax reporter. Exog-enous MEF2A transactivated expression 3.8-fold in trans-fected C2C12 myoblasts (GM), which have low levels ofMEF2, but the extent of transactivation was markedly reducedin differentiating cultures (DM) that have higher levels ofMEF2 (Fig. 9). An intact MEF2-binding site was required forthis response, since mutations within this site (T1M2) pre-vented MEF2A transactivation. Furthermore, the T1M2 mu-tation had no effect on basal promoter activity in myoblastcultures but decreased basal expression by a factor of 3 indifferentiating myocyte cultures. Collectively, these experi-ments demonstrate that MEF2, in concert with other factors,can contribute to the overall level of Gax promoter activity.Finally, we tested whether overexpression of MEF2A could
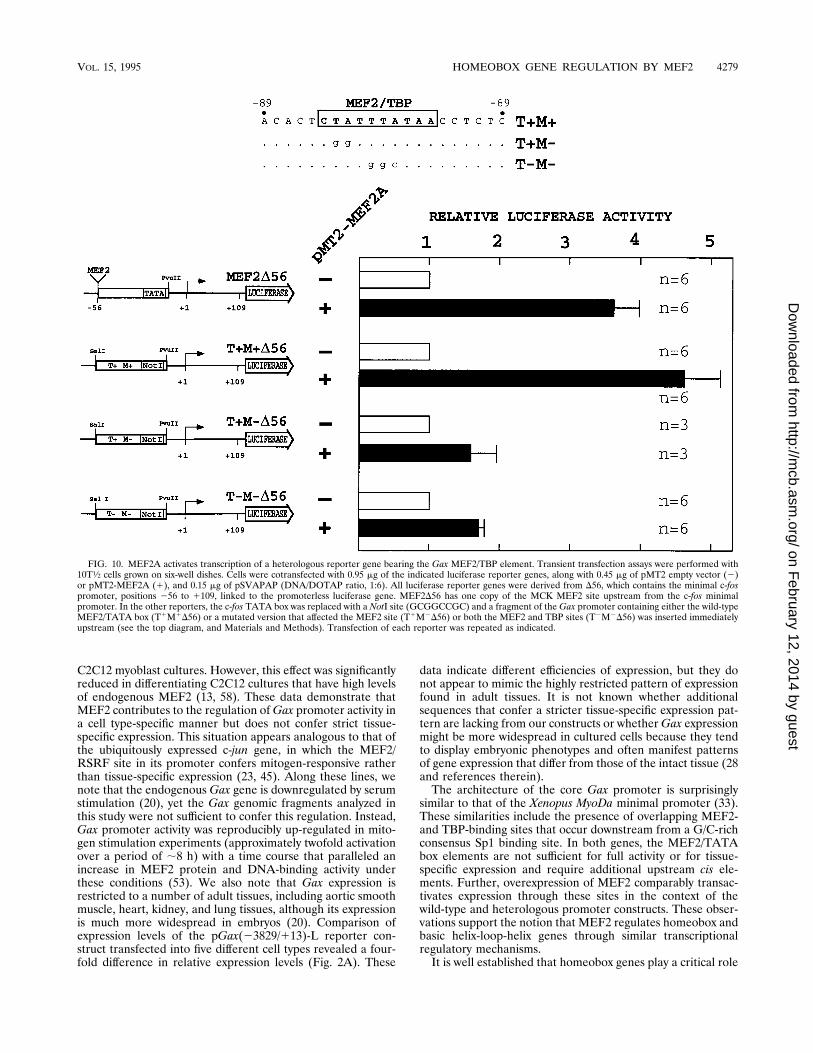
activate transcription through the Gax MEF2 site in the con-text of a heterologous reporter gene construct that lacks aTATA element. Figure 10 schematizes the structure of theplasmids used for this study, which are derived from a reportergene containing the minimal c-fos promoter linked to the lu-ciferase gene. MEF2D56 has one copy of the MCK MEF2 siteupstream from the c-fos minimal promoter extending from
positions256 to1109 (53). We also constructed T1M1D56, inwhich the c-fos TATA box was replaced with a NotI site (GCGGCCGC) and a fragment of the Gax promoter containing theMEF2/TATA box was inserted immediately upstream. As con-trols, we used T1M2D56 and T2M2D56, in which the MEF2site or both the MEF2 and TBP sites were mutated, respec-tively. Transient expression assays with 10T1⁄2 cells revealedthat MEF2A transactivated the MEF2D56 and T1M1D56 con-structs four- to fivefold, with little or no effect on either theT1M2D56 or the T2M2D56 mutant. Therefore, we concludethat MEF2 is capable of transactivating with similar degrees ofefficiency, reporter genes containing a MEF2 site located up-stream of the TATA box or in the context of a TATA-lesspromoter.
DISCUSSION
In this study, we elucidated components of the regulatorynetworks that govern Gax homeobox gene expression. Weshowed that the sequences essential for expression of its min-imal promoter (positions 2125 to 113) occur within a 51-bpfragment between positions 2125 and 275. Deletion of this51-bp region decreased expression by factors of 100, 25, and 5in transiently transfected developing chick hearts, A10 cells,and primary cultures of rat aorta smooth muscle cells, respec-tively. Expression from this region appears to depend upon itsinteractions with at least three positive transcription factors:HRF-1, Sp1, and MEF2. The palindromic element at the 59end of the 51-bp fragment is bound by HRF-1, which is de-tected in extracts from A10 and 10T1⁄2 cells (Fig. 5) and fromprimary cultures of rat aorta cells (data not shown). Deletionor mutation of this palindrome diminished HRF-1 binding andpartially decreased promoter activity. The downstream half ofthe palindrome overlaps a G/C-rich motif that binds to the zincfinger protein Sp1. Mutations within this element abolishedSp1 binding and partially reduced promoter activity. The thirdcis element identified conforms to a consensus MEF2/RSRFsite (13, 45). Mutations that abrogated MEF2 binding alsoreduced promoter activity in some cell types (see below). TheGax MEF2 site also conforms to a TATA box motif, and TBPcan bind to this element in vitro (Fig. 7C). Thus, TBP mayinteract with the Gax promoter through this site. Alternatively,another factor, such as Sp1, may function to tether TBP to theactive transcription complex (16, 55, and references therein).Examples of mammalian homeobox genes which lack theTATA box and contain putative Sp1 binding sites includeHoxa-4 (Hox-1.4) (17) and Hoxc-4 (Hox-3.5) (18), while otherhomeobox gene promoters lack both TATA and Sp1 elements(10).Several lines of evidence indicate a role for MEF2 in the
regulation of Gax expression. First, the promoter contains aMEF2/RSRF site that is identical to the MEF2/RSRF sitesthat have been identified in the regulatory regions of muscle-specific genes (13) and by the site selection procedure (45).Second, the Gax site is efficiently bound by MEF2. Third,mutations in this site that abolished binding decreased pro-moter activity in a cell type-specific manner. These mutationshad the greatest impact on expression in differentiating cul-tures of C2C12 myocytes, which have the highest levels ofendogenous MEF2. Mutations in the MEF2 site had lessereffects on expression in the smooth muscle cultures that haverelatively lower levels of MEF2 activity and no detectable ef-fect on expression in C2C12 myoblasts or 10T1⁄2 fibroblasts thatessentially have no MEF2 activity. Finally, overexpression ofMEF2 could transactivate Gax expression in a site-dependentmanner in 10T1⁄2 fibroblast, A10 smooth muscle cell, and
FIG. 9. MEF2A transactivation of the Gax promoter in C2C12 myocytecultures. C2C12 cells were transiently transfected by the calcium phosphatemethod. Cells were cotransfected with 2 mg of pMT2 empty vector (2) orpMT2-MEF2A (1), 8 mg of wild-type pGax(2652/113)-L (wt), or a mutatedversion in which the MEF2 site was destroyed (T1M2) (see Fig. 8) and 1 mg ofthe internal alkaline phosphatase control plasmid. Cells were maintained ingrowth medium to prevent myogenic differentiation [C2C12(GM)] or transferredto differentiation medium after glycerol shock to induce myogenesis[C2C12(DM)]. Cells were harvested 2 days after the glycerol shock. At the timeof harvesting, cells switched to differentiation medium consisted of a mixedpopulation of multinucleated myotubes (;25%) and mononucleated cells(;75%). Luciferase activity is expressed relative to that of the wild-type plasmidin C2C12(DM) without exogenous MEF2A, which was taken as 1. Transfectionof each reporter was repeated three times.
4278 ANDRES ET AL. MOL. CELL. BIOL.
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
C2C12 myoblast cultures. However, this effect was significantlyreduced in differentiating C2C12 cultures that have high levelsof endogenous MEF2 (13, 58). These data demonstrate thatMEF2 contributes to the regulation ofGax promoter activity ina cell type-specific manner but does not confer strict tissue-specific expression. This situation appears analogous to that ofthe ubiquitously expressed c-jun gene, in which the MEF2/RSRF site in its promoter confers mitogen-responsive ratherthan tissue-specific expression (23, 45). Along these lines, wenote that the endogenous Gax gene is downregulated by serumstimulation (20), yet the Gax genomic fragments analyzed inthis study were not sufficient to confer this regulation. Instead,Gax promoter activity was reproducibly up-regulated in mito-gen stimulation experiments (approximately twofold activationover a period of ;8 h) with a time course that paralleled anincrease in MEF2 protein and DNA-binding activity underthese conditions (53). We also note that Gax expression isrestricted to a number of adult tissues, including aortic smoothmuscle, heart, kidney, and lung tissues, although its expressionis much more widespread in embryos (20). Comparison ofexpression levels of the pGax(23829/113)-L reporter con-struct transfected into five different cell types revealed a four-fold difference in relative expression levels (Fig. 2A). These
data indicate different efficiencies of expression, but they donot appear to mimic the highly restricted pattern of expressionfound in adult tissues. It is not known whether additionalsequences that confer a stricter tissue-specific expression pat-tern are lacking from our constructs or whetherGax expressionmight be more widespread in cultured cells because they tendto display embryonic phenotypes and often manifest patternsof gene expression that differ from those of the intact tissue (28and references therein).The architecture of the core Gax promoter is surprisingly
similar to that of the Xenopus MyoDa minimal promoter (33).These similarities include the presence of overlapping MEF2-and TBP-binding sites that occur downstream from a G/C-richconsensus Sp1 binding site. In both genes, the MEF2/TATAbox elements are not sufficient for full activity or for tissue-specific expression and require additional upstream cis ele-ments. Further, overexpression of MEF2 comparably transac-tivates expression through these sites in the context of thewild-type and heterologous promoter constructs. These obser-vations support the notion that MEF2 regulates homeobox andbasic helix-loop-helix genes through similar transcriptionalregulatory mechanisms.It is well established that homeobox genes play a critical role
FIG. 10. MEF2A activates transcription of a heterologous reporter gene bearing the Gax MEF2/TBP element. Transient transfection assays were performed with10T1⁄2 cells grown on six-well dishes. Cells were cotransfected with 0.95 mg of the indicated luciferase reporter genes, along with 0.45 mg of pMT2 empty vector (2)or pMT2-MEF2A (1), and 0.15 mg of pSVAPAP (DNA/DOTAP ratio, 1:6). All luciferase reporter genes were derived from D56, which contains the minimal c-fospromoter, positions 256 to 1109, linked to the promoterless luciferase gene. MEF2D56 has one copy of the MCK MEF2 site upstream from the c-fos minimalpromoter. In the other reporters, the c-fos TATA box was replaced with a NotI site (GCGGCCGC) and a fragment of the Gax promoter containing either the wild-typeMEF2/TATA box (T1M1D56) or a mutated version that affected the MEF2 site (T1M2D56) or both the MEF2 and TBP sites (T2M2D56) was inserted immediatelyupstream (see the top diagram, and Materials and Methods). Transfection of each reporter was repeated as indicated.
VOL. 15, 1995 HOMEOBOX GENE REGULATION BY MEF2 4279
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

in the formation of skeletal, cardiac, and smooth muscles (8,21, 42). Null mutations in the homeobox gene tinman give riseto flies that lack heart and visceral (smooth) muscles (4, 5), andbagpipe, another homeobox gene, is also essential for the for-mation of visceral muscle (4). Further, mouse embryos withnull mutations in the mammalian homolog of tinman, referredto as NKX2.5 or Csx, show abnormal heart development (29,36). The involvement ofMEF2 in the specification of Drosoph-ila muscle is demonstrated by the complete absence of musclefibers in embryos carrying null mutations inDMEF2 (35). Sinceboth MEF2 and homeobox genes are involved in muscle for-mation, it is not surprising that members of these familiesconverge in regulatory networks. Indeed, protein-protein in-teractions between homeodomain proteins and MADS factorshave been demonstrated previously. Examples include the in-teraction of yeast homeodomain protein Mata-2 with SRF-likeprotein MCM1, which permits repression of the mating typea-specific genes through cooperative binding of the two pro-teins to operator sequences (1, 27, 43, 56). In addition, thehuman homeoprotein Phox can activate c-fos expression byfacilitating binding of the SRF to the CArG box (22). Finally,MEF2 and MHox, the mouse homolog of Phox, have overlap-ping DNA-binding specificities on the MCK enhancer, thusproviding the potential for antagonistic effects on MCK genetranscription (11, 12). Taken together, these observations es-tablish precedence for a functional convergence between ho-meoproteins and MADS factors, whereby they can influenceone another’s activities through cooperative protein-proteininteractions or mutually exclusive binding to overlapping targetsites. Our results provide further evidence for cross talk betweenhomeodomain and MADS proteins by demonstrating the directregulation of transcription through the interaction of MEF2 withcis elements within the Gax homeobox gene promoter.
ACKNOWLEDGMENTS
We thank Jason Lowry for DNA sequencing, Laura Wei for assis-tance with transfections, and Liz Kim for preparing the cultures ofprimary rat aorta cells. We also thank Jose Ayte, Marıa D. Chiara, andmembers of our laboratory for comments on the manuscript. V.A. isthe recipient of a postdoctoral fellowship from the American HeartAssociation (Massachusetts affiliate). K.W. is an established investiga-tor from the American Heart Association. This work was supported inpart by grants from the National Institutes of Health (AR40197 andHL50692) to K.W.
REFERENCES
1. Ammerer, G. 1990. Identification, purification, and cloning of a polypeptide(PRTF/GRM) that binds to mating-specific promoter elements in yeast.Genes Dev. 4:299–312.
2. Andres, V., B. Nadal-Ginard, and V. Mahdavi. 1992. Clox, a mammalian ho-meobox gene related to Drosophila cut, encodes DNA-binding regulatory pro-teins differentially expressed during development. Development 116:321–334.
3. Arcioni, L., A. Simeone, S. Guazzi, V. Zappavigna, E. Boncinelli, and F.Mavilio. 1992. The upstream region of the human homeobox gene HOX3Dis a target for regulation by retinoic acid and HOX homeoproteins. EMBOJ. 11:265–277.
4. Azpiazu, N., and M. Frasch. 1993. tinman and bagpipe: two homeo box genesthat determine cell fates in the dorsal mesoderm of Drosophila. Genes Dev.7:1325–1340.
5. Bodmer, R. 1993. The gene tinman is required for specification of the heartand visceral muscles in Drosophila. Development 118:719–729.
6. Botas, J. 1993. Control of morphogenesis and differentiation by HOM/Hoxgenes. Curr. Opin. Cell Biol. 5:1015–1022.
7. Breitbart, R. E., C. S. Liang, L. B. Smoot, D. A. Laheru, V. Mahdavi, and B.Nadal-Ginard. 1993. A fourth human MEF2 transcription factor, hMEF2D,is an early marker of the myogenic lineage. Development 118:1095–1106.
8. Buckingham, M. 1994. Molecular biology of muscle development. Cell 78:15–21.
9. Cheng, T. C., M. C. Wallace, J. P. Merlie, and E. N. Olson. 1993. Separableregulatory elements governing myogenin transcription in mouse embryogen-esis. Science 261:215–218.
10. Coletta, P. L., S. M. Shimeld, C. Chaudhuri, U. Muller, J. P. Clarke, andP. T. Sharpe. 1991. Characterization of the murine Hox-3.3 gene and itspromoter. Mech. Dev. 35:129–142.
11. Cserjesi, P., B. Lilly, L. Bryson, Y. Wang, D. A. Sassoon, and E. N. Olson.1992.MHox: a mesodermally restricted homeodomain protein that binds an essentialsite in the muscle creatine kinase enhancer. Development 115:1087–1101.
12. Cserjesi, P., B. Lilly, C. Hinkley, M. Perry, and E. N. Olson. 1994. Home-odomain protein MHox and MADS protein myocyte enhancer-binding fac-tor-2 converge on a common element in the muscle creatine kinase en-hancer. J. Biol. Chem. 269:16740–16745.
13. Cserjesi, P., and E. N. Olson. 1991. Myogenin induces the myocyte-specificenhancer binding factor MEF-2 independently of other muscle-specific geneproducts. Mol. Cell. Biol. 11:4854–4862.
14. Edmondson, D. G., T. C. Cheng, P. Cserjesi, T. Chakraborty, and E. N.Olson. 1992. Analysis of the myogenin promoter reveals an indirect pathwayfor positive autoregulation mediated by the muscle-specific enhancer factorMEF-2. Mol. Cell. Biol. 12:3665–3677.
15. Fisher, S. A., K. Walsh, and C. J. Forehand. Characterization of cardiac genecis-regulatory elements in the early stages of chicken heart morphogenesis.Submitted for publication.
16. Franklin, B., and R. Tjian. 1990. Mechanism of transcriptional activation bySp1: evidence for coactivators. Cell 61:1187–1197.
17. Galliot, B., M. Dolle, M. Vigneron, M. Featherstone, A. Baron, and D.Duboule. 1989. The mouse Hox-1.4 gene: primary structure, evidence forpromoter activity and expression during development. Development 107:343–359.
18. Geada, A. M. C., S. J. Gaunt, M. Azzawi, S. M. Shimeld, J. Pearce, and P. T.Sharpe. 1992. Sequence and embryonic expression of the murine Hox-3.5gene. Development 116:497–506.
19. Gorman, C., R. Padmanabhan, and B. H. Howard. 1983. High efficiencyDNA-mediated transformation of primate cells. Science 221:551–553.
20. Gorski, D. H., D. F. LePage, C. V. Patel, N. G. Copeland, N. A. Jenkins, andK. Walsh. 1993. Molecular cloning of a diverged homeobox gene that israpidly down-regulated during the G0/G1 transition in vascular smooth mus-cle cells. Mol. Cell. Biol. 13:3722–3733.
21. Gorski, D. H., C. V. Patel, and K. Walsh. 1993. Homeobox transcriptionfactor regulation in the cardiovascular system. Trends Cardiovasc. Med.3:184–190.
22. Grueneberg, D. A., S. Natesan, C. Alexandre, and M. Z. Gilman. 1992.Human and Drosophila homeodomain proteins that enhance the DNA-binding activity of the serum response factor. Science 257:1089–1095.
23. Han, T.-H., W. W. Lamph, and R. Prywes. 1992. Mapping of epidermalgrowth factor-, serum-, and phorbol ester-responsive sequence elements inthe c-jun promoter. Mol. Cell. Biol. 12:4472–4477.
24. Henthorn, P., P. Zervos, M. Raducha, H. Harris, and T. Kadesch. 1988.Expression of a human placental alkaline phosphatase gene in transfectedcells: use as a reporter for studies of gene expression. Proc. Natl. Acad. Sci.USA 85:6342–6346.
25. Holland, P. W., and B. L. M. Hogan. 1988. Expression of homeo box genesduring mouse development: a review. Genes Dev. 2:773–782.
26. Kaushal, S., J. W. Schneider, B. Nadal-Ginard, and V. Mahdavi. 1994.Activation of the myogenic lineage by MEF2A, a factor that induces andcooperates with MyoD. Science 266:1236–1240.
27. Keleher, C. A., C. Goutte, and A. D. Johnson. 1988. The yeast cell-type-specific repressor a2 acts cooperatively with a non-cell-type-specific protein.Cell 53:927–936.
28. Kitsis, R. N., and L. A. Leinwand. 1992. Discordance between gene regula-tion in vitro and in vivo. Gene Expression 2:313–318.
29. Komuro, I., and S. Izumo. 1993. Csx: a murine homeobox-containing genespecifically expressed in the developing heart. Proc. Natl. Acad. Sci. USA90:8145–8149.
30. Krumlauf, R. 1994. Hox genes in vertebrate development. Cell 78:191–201.31. Langston, A. W., and L. J. Gudas. 1992. Identification of a retinoic acid
responsive enhancer 39 of the murine homeobox gene Hox-1.6. Mech. Dev.38:217–228.
32. Lawrence, P. A., and G. Morata. 1994. Homeobox genes: their function inDrosophila segmentation and pattern formation. Cell 78:181–189.
33. Leibham, D., M.-W. Wong, T.-C. Cheng, S. Schroeder, P. A. Weil, E. N.Olson, and M. Perry. 1994. Binding of TFIID and MEF2 to the TATAelement activates transcription of the Xenopus MyoDa promoter. Mol. Cell.Biol. 14:686–699.
34. Leifer, D., D. Krainc, Y. T. Yu, J. McDermott, R. E. Breitbart, J. Heng, R. L.Neve, B. Kosofsky, B. Nadal-Ginard, and S. A. Lipton. 1993. MEF2C, aMADS/MEF2-family transcription factor expressed in a laminar distributionin cerebral cortex. Proc. Natl. Acad. Sci. USA 90:1546–1550.
35. Lilly, B., B. Zhao, G. Ranganayakulu, B. M. Paterson, R. A. Schulz, andE. N. Olson. 1995. Requirement of MADS domain transcription factorD-MEF2 for muscle formation in Drosophila. Science 267:688–693.
36. Lints, T. J., L. M. Parsons, L. Hartley, I. Lyons, and P. Harvey. 1993.NKX-2.5: a novel murine homeobox gene expressed in early heart progenitorcells and their myogenic descendants. Development 119:419–431.
37. Marshall, H., M. Studer, H. Popperl, S. Aparicio, A. Kuroiwa, S. Brenner,
4280 ANDRES ET AL. MOL. CELL. BIOL.
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

and R. Krumlauf. 1994. A conserved retinoic acid response element requiredfor early expression of the homeobox gene Hoxb-1. Nature (London) 370:567–571.
38. Martin, J. F., J. M. Miano, C. M. Hustad, N. G. Copeland, N. A. Jenkins, andE. N. Olson. 1994. A Mef2 gene that generates a muscle-specific isoform viaalternative mRNA splicing. Mol. Cell. Biol. 14:1647–1656.
39. Martin, J. F., J. J. Schwarz, and E. N. Olson. 1993. Myocyte enhancer factor(MEF) 2C: a tissue-restricted member of the MEF-2 family of transcriptionfactors. Proc. Natl. Acad. Sci. USA 90:5282–5286.
40. McDermott, J. C., M. C. Cardoso, Y.-T. Yu, V. Andres, D. Leifer, D. Krainc,S. A. Lipton, and B. Nadal-Ginard. 1993. hMEF2C gene encodes skeletalmuscle- and brain-specific transcription factors. Mol. Cell. Biol. 13:2564–2577.
41. McGinnis, W., and R. Krumlauf. 1992. Homeobox genes and axial pattern-ing. Cell 68:283–302.
42. Olson, E. N., and N. Rosenthal. 1994. Homeobox genes and muscle pattern-ing. Cell 79:9–12.
43. Passmore, S., R. Elble, and B.-R. Tye. 1989. A protein involved in minichro-mosome maintenance in yeast binds a transcriptional enhancer conserved ineukaryotes. Genes Dev. 3:921–935.
44. Pickering, J. G., P. A. Bacha, L. Weir, J. Jekanowski, J. C. Nichols, and J. M.Isner. 1993. Prevention of smooth muscle cell outgrowth from human ath-erosclerotic plaque by a recombinant cytotoxin specific for the epidermalgrowth factor receptor. J. Clin. Invest. 91:724–729.
45. Pollock, R., and R. Treisman. 1991. Human SRF-related proteins: DNA-binding properties and potential regulatory targets. Genes Dev. 5:2327–2341.
46. Popperl, H., and M. S. Featherstone. 1993. Identification of a retinoic acidresponse element upstream of the murine Hox-4.2 gene. Mol. Cell. Biol.13:257–265.
47. Reznikoff, C. A., D. W. Brankow, and C. Heidelberg. 1973. Establishment andcharacterization of a cloned line of C3H mouse embryo cells sensitive topostconfluence inhibition of division. Cancer Res. 33:3231–3238.
48. Santoro, I. M., and K. Walsh. 1991. Natural and synthetic DNA elementswithin the CArG motif differ in expression and protein-binding properties.Mol. Cell. Biol. 11:6296–6305.
49. Schneider-Maunoury, S., P. Topilko, T. Seitanidou, G. Levi, M. Cohen-Tannoudji, S. Pournin, C. Babinet, and P. Charnay. 1993. Disruption ofKrox-20 results in elimination of rhombomeres 3 and 5 in the developinghindbrain. Cell 75:1199–1214.
50. Schwarz-Sommer, Z., P. Huijser, W. Nacken, H. Saedler, and H. Sommer.1990. Genetic control of flower development: homeotic genes in Atirrhinummajus. Science 250:931–936.
51. Shashikant, C. S., M. F. Utset, S. M. Violette, T. L. Wise, P. Einat, J. W.Pendleton, K. Schugart, and F. H. Ruddle. 1991. Homeobox genes in mousedevelopment. Crit. Rev. Eukaryotic Gene Expression 1:207–245.
52. Studer, M., H. Popperl, H. Marshall, A. Kuroiwa, and R. Krumlauf. 1994.Role of a conserved retinoic acid response element in rhombomere restric-tion of Hoxb-1. Science 265:1728–1732.
53. Suzuki, E., K. Guo, M. Kolman, Y.-T. Yu, and K. Walsh. 1995. Seruminduction of MEF2/RSRF expression in vascular myocytes is mediated at thelevel of translation. Mol. Cell. Biol. 15:3415–3423.
54. Swiatek, P. J., and T. Gridley. 1993. Perinatal lethality and defects in hind-brain development in mice homozygous for a targeted mutation of the zincfinger gene Krox-20. Genes Dev. 7:2071–2084.
55. Swick, A. G., M. C. Blake, J. W. Kahn, and J. C. Azizkhan. 1989. Functionalanalysis of GC element binding and transcription in hamster dihydrofolatereductase gene promoter. Nucleic Acids Res. 17:9291–9304.
56. Vershon, A., and A. D. Johnson. 1993. A short, disordered protein regionmediates interactions between the homeodomain of the yeast a2 protein andthe MCM1 protein. Cell 72:105–112.
57. Yee, S. P., and P. W. Rigby. 1993. The regulation of myogenin gene expres-sion during the embryonic development of the mouse. Genes Dev. 7:1277–1289.
58. Yu, Y. T., R. E. Breitbart, L. B. Smoot, Y. Lee, V. Mahdavi, and B. Nadal-Ginard. 1992. Human myocyte-specific enhancer factor 2 comprises a groupof tissue-restricted MADS box transcription factors. Genes Dev. 6:1783–1798.
59. Zappavigna, V., A. Renucci, J.-C. Izpisua-Belmonte, G. Urier, C. Peschle,and D. Duboule. 1991. HOX4 genes encode transcription factors with po-tential auto- and cross-regulatory capacities. EMBO J. 10:4177–4187.
VOL. 15, 1995 HOMEOBOX GENE REGULATION BY MEF2 4281
on February 12, 2014 by guest
http://mcb.asm
.org/D
ownloaded from




















