
trongylocentrotus franciscanus growth using six growth functions
Upload
independentCategory
view
1download
0
CELLULAR IMMUNOLOGY 114,41-54 (1988)
Transforming Growth Factor@ Are Equipotent Growth Inhibitors of Interleukin-1 -Induced Thymocyte Proliferation
LARRY R. ELLINGSWORTH,DEBRANAKAYAMA,PATRICIA SEGARINI, JAMES DASCH, PEDRO CARRILLO, AND WENDY WAEGELL
Immunology Laboratory, Connective Tissue Research Laboratories, Collagen Corporation, 2500 Faber Place, Palo Alto, California 94303
Received October 16.1987; accepted January 5, 1988
The effects of two forms of transforming growth factor-& TGF-j31 and TGF@, upon the proliferative response of murine thymocytes were investigated in this study. TGF-fll and TGF- /32 were found to be equipotent growth inhibitor of interleukin- 1 (IL- I)- and phytohemaggluti- nin (PHA)-stimulated thymocytes when added at the initiation of the cultures. These factors suppressed the proliferative response in a dose-dependent fashion between 0.4 and 100 pM The proliferative response was maximally inhibited (90% inhibition) at 100 PM. The half-maximal inhibitory dose (ID& was 6 and 4 pM for TGF-Bl and TGF-82, respectively. These factors were less effective or ineffective at suppressing the proliferation of thymocytes which had been prestimulated for 24 to 48 hr by IL1 and PHA. Neither factor inhibited interleukin-2 (K-2)- dependent thymocyte proliferation or the proliferation of an IL-2dependent cytotoxic T cell line (CTL-L), suggesting that the anti-proliferative actions of these factors was by inhibition of celhtlar events triggered by ILl. Furthermore, anti-TGF-@l antibodies did neutralize the biological actions of TGF-/31 and these antibodies did block the binding of ‘251-labeled TGF-j31 to cell surface receptors showing that the inhibitory action is mediated through specific receptors for TGF-fil on thymocytes. These antibodies, however, did not neutralize the anti-proliferative action of TGF-82. Although TGF+l and TGF-j32 exhibit very similar biological activities, these molecules are antigenically different and, therefore, have different tertiary structures. o 19s~
Academic Press, Inc.
INTRODUCTION
Transforming growth factor-a (TGF-(Y) and TGF-/3 were originally described as protein factors which are produced by virally transformed cells which act synergisti- cally to impart a transformed phenotype (i.e., anchorage-independent growth and the loss of contact inhibition properties) to normal rat kidney (NRK) fibroblasts and other nontransformed cells ( l-6). TGF-a has been shown to be structurally and func- tionally related to epidermal growth factor (EGF) (7). In contrast to TGF-a, TGF-8 is a 26-kDa homodimer which is biochemically and functionally distinct from TGF- (Y or EGF (8- 11). TGF-/3 is produced by numerous transformed cells (1,3-6, 12, 13) and it has been purified from human placenta ( 14), human and porcine platelets ( 15, 16), bovine kidney ( 17) and bovine bone (18).
TGF-P was originally believed to be a single molecular entity. Recently, multiple forms of TGF-p have been purified from porcine platelets ( 16) and bovine bone ( 18- 2 1). These different forms have been referred to as TGF-/31 and TGF-P2 (16). A
41
0008-8749188 $3.00 Copyright 0 1988 by Academic Pms, Inc. All rigbfs of reprodunion in any form reserved.

42 ELLINGSWORTH ET AL.
partial amino acid sequence shows that TGF-P 1 and TGF-02 share 63% amino-ter- minal sequence identity (16,22).
Recently, TGF-/3 1 has been shown to be a potent inhibitor of the mitogenic respon- siveness of human tonsillar T cells (38) and B cells (39) and murine thymocytes (40). Kehrl et al. (38, 39) have shown that human T cells and B cells express TGF+l mRNA upon mitogenic stimulation and that the conditioned culture medium from these cells contains TGF-P-like biological activity. These investigators have proposed that TGF-01 may function in an autocrine manner to regulate the clonal growth of lymphocytes.
Little is known about functional or structural differences between TGF-61 and TGF-02. In this study, we have used an IL l-induced thymocyte proliferation assay method and neutralizing antibodies to compare the anti-proliferative action of TGF- /31 to TGF-@2. These studies show that both factors are equally potent inhibitors of IL- 1 -induced thymocyte proliferation and that the anti-proliferative action is medi- ated through specific receptors for TGF-Pl on thymocytes. Although TGF-PI and TGF-/32 were found to be functionally equivalent, antibodies which neutralized the biological activity of TGF+3 1 did not neutralize the biological activity of TGF+32. It is concluded that these factors likely have very different tertiary structures.
MATERIALS AND METHODS
Animals. Young (approximately 4 to 8 weeks old) C3H/HeJ mice were purchased from Jackson Laboratories (Bar Harbor, ME).
TGF-j.31 and TGF-/32. Bone-derived TGF-P 1 and TGF-@2 were purified from the noncollagenous, guanidine-HCl-soluble proteins of demineralized bovine metatarsal bone. These factors were purified by a combination of gel filtration, CM-cellulose cation-exchange chromatography, and Cl* reverse-phase HPLC using a previously described method ( 18). Protein concentrations were determined by comparing HPLC peaks of known protein standards.
NRK colony-forming assay. The biological activity of TGF-fil and TGF+?2 was determined by the NRK colony-forming assay method as described by Roberts et al. (25) using NRK clone 49F (CRL 1570) as indicator cells. The co-factor, EGF, was purchased from Bethesda Research Laboratories (Gaithersburg, MD) and it was used at a culture concentration of 5 rig/ml.
IL-l assay. Single cell suspensions of thymocytes were prepared and the cells were suspended in Earle’s minimal essential medium (Irvine Scientific, Santa Ana, CA) supplemented with 5% fetal calf serum, 100 U/ml penicillin, 100 pg/ml streptomycin, 2 rnA4 L-glutamine, and 5 X 10e5 It4 2-mercaptoethanol. The thymocytes ( lo6 cells/ well) were plated into 96-well microtiter plates (Costar 3596). The cells were activated with 1 .O pg/ml phytohemagglutinin (PHA, Sigma Chemical Co., St. Louis, MO) and the indicated amounts of IL- 1. Highly purified, endotoxin-free human blood mono- cyte-derived IL-1 or recombinant human IL1 (r&l) was purchased from Cistron Technology (Pine Brook, NJ). One unit of specific activity was defined as the amount of IL- 1 which resulted in the half-maximal proliferative response of the thymocytes. TGF-PI or TGF-/32 was added to the thymocyte cultures at the time and in the con- centrations indicated in each experiment. Unless otherwise indicated, the cells were cultured for 72 hr in a humidified incubator in 5% CO* at 37°C. Prior to harvesting, the cells were pulsed for 24 hr with 0.5 &i of [3H]thymidine per culture (Amersham,

TGF-(31 AND TGF-p2 AS IMMUNE SUPPRESSORS 43
Chicago, IL). The cultures were harvested with a semiautomated cell harvester (Otto Hiller Co., Madison, WI) and the amount of [3H]thymidine incorporation was deter- mined by standard liquid scintillation methods.
IL-2-dependent T cells. Thymocytes (4 X 1 06/ml) were preactivated in bulk culture for 24 hr with 8 U/ml IL- 1 and PHA. The cells were washed extensively with medium to remove the mitogens. The cells were recultured in 0.2 ml of medium containing 2 to 80 U/ml of human interleukin-2 (Boehringer-Mannheim, Indianapolis, IN) with the indicated amounts of either TGF-PI or TGF-/32. The cells were cultured for an additional 72 hr and pulsed for 24 hr with 0.5 &i of [3H]thymidine per culture prior to harvesting.
Antibodies CO TGF-PI. A New Zealand white rabbit was immunized by multiple intramuscular injections of highly purified TGF-/31. The injections were given monthly over a period of 6-9 months using 100 pg of TGF-/31 per injection. The primary immunization was in Freund’s complete adjuvant and the subsequent boosts were in Freund’s incomplete adjuvant (GIBCO Laboratories, Grand Island, NY). Ten days following the boosts, blood samples were collected by cardiac puncture and the antiserum was stored at -70°C.
Antibody purzjication. Antiserum IgG or normal rabbit serum IgG was purified using protein A-Sepharose (Pharmacia, Inc., Piscataway, NJ). The column-bound IgG was eluted with 0.1 A4 glycine-HCl (pH 2.0) and the antibody-containing frac- tions were neutralized with 4.0 M Tris base and dialyzed against 0.1 M phosphate- buffered saline (pH 7.0). The anti-TGF+l antibody titer was determined by the ELISA method described by Ellingsworth et af. (24). The anti-TGF-/31 antibody titer was l/8 1,000 against TGF-/3 1 and l/ 1200 against TGF-P2.
Antibody neutralization. The anti-TGF-@ 1 IgG was evaluated for neutralizing ac- tivity using the NRK colony-forming assay and the thymocyte proliferation assay. In these experiments, normal rabbit IgG or anti-TGF-Pl IgG was either preincubated for 2-3 hr with TGF-/31 or TGF-/32 or added simultaneously with TGF-@I or TGF- @2 to the cultures. The antibody concentrations are indicated in each experiment.
Radioiodination. TGF-P 1 was radiolabeled with 1 mCi of Na’251 ( 13- 16 mCi/pg, Amersham) using a method described elsewhere ( 11). The specific activity of the ra- diolabeled peptide ranged between 50 and 100 pCi/pg.
Receptor crosslinking. The TGF-P 1 receptor complex was evaluated by covalently crosslinking ‘251-labeled TGF-@l to the cell surface receptors on NRK fibroblasts, fresh thymocytes (unactivated), and thymocytes which had been activated with IL- 1 and PHA for 24 hr. ‘2SI-labeled TGF-/31 was crosslinked to the receptors using disuccinimidyl suberate (DSS, Pierce No. 2155). Brielly, 100 pM “‘I-labeled TGF- pl was incubated for 2-3 hr at 4°C with 0.5 to 1 X 10’ cells in Dulbecco’s minimal essential medium containing 25 mMHepes and 0.1% bovine serum albumin (Sigma). DSS was added to the cell suspension at a final concentration of 0.3 mM and incu- bated for 15 min at 4°C. The cells were washed three times with 10 rn&! Tris (pH 7.4) containing 250 mMsucrose and 1 mMethylenediaminetetraacetic acid (sucrose buffer) to remove the nonbound radiolabel and excess crosslinking agent. Cell mem- branes were then lysed in 100 ~1 of sucrose buffer containing 1% Triton, 1 rmV phe- nylmethylsulfonyl fluoride, and 5 mM benzamidine. The nuclei were pelleted ( 10,000 rpm, Beckman microfuge) for 1 min. The solubilized membranes were re- duced with sample buffer containing 100 mM DL-dithiothreitol (Sigma). The solubi- lized membranes were electrophoretically separated in a 4-10% linear SDS-poly-
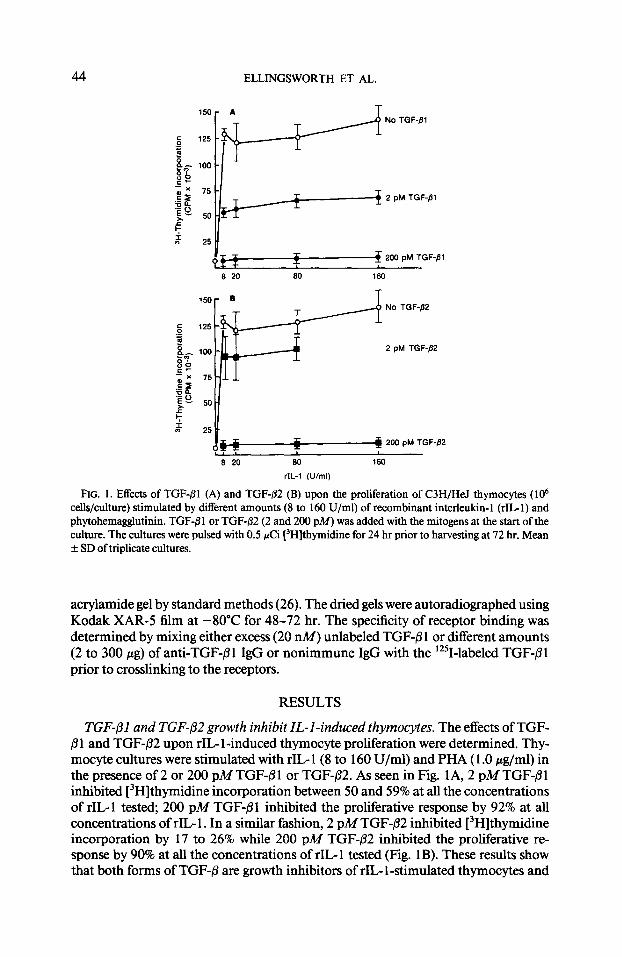
44 ELLINGSWORTH ET AL.
No TGF-fll
2 pt.4 TGF-Bl
u 0 $ 200 pM TGF-ISI
8 20 SO 160
100
75
2 pM TGF-02
25 $r# 200 pk.4 TGF-/32
s 20 so 160
rlL-I (U/ml)
FIG. 1. Effects of TGF-01 (A) and TGF-,!32 (B) upon the proliferation of C3H/HeJ thymocytes ( lo6 cells/culture) stimulated by different amounts (8 to 160 U/ml) of recombinant interleukin- 1 (rIL- 1) and phytohemagglutinin. TGF-61 or TGF-02 (2 and 200 p&f) was added with the mitogens at the start of the culture. The cultures were pulsed with 0.5 PCi (3H]thymidine for 24 hr prior to harvesting at 72 hr. Mean f SD of triplicate cultures.
acrylamide gel by standard methods (26). The dried gels were autoradiographed using Kodak XAR-5 film at -80°C for 48-72 hr. The specificity of receptor binding was determined by mixing either excess (20 nM) unlabeled TGFj3 1 or different amounts (2 to 300 pg) of anti-TGF-/31 IgG or nonimmune IgG with the ‘251-labeled TGF-/31 prior to crosslinking to the receptors.
RESULTS
TGF-01 and TGF+32 growth inhibit IL-l-induced thymocytes. The effects of TGF- B 1 and TGF-/32 upon rIG 1 -induced thymocyte proliferation were determined. Thy- mocyte cultures were stimulated with rIL- 1 (8 to 160 U/ml) and PHA ( 1 .O pg/ml) in the presence of 2 or 200 pM TGF-Pl or TGF-/32. As seen in Fig. 1 A, 2 pM TGF-@l inhibited [3H]thymidine incorporation between 50 and 59% at all the concentrations of rIL-1 tested, 200 pM TGF-01 inhibited the proliferative response by 92% at all concentrations of rIL- 1. In a similar fashion, 2 pM TGF-@2 inhibited [3H]thymidine incorporation by 17 to 26% while 200 pM TGF-P2 inhibited the proliferative re- sponse by 90% at all the concentrations of rIL 1 tested (Fig. 1 B). These results show that both forms of TGF-0 are growth inhibitors of rIL- 1 -stimulated thymocytes and
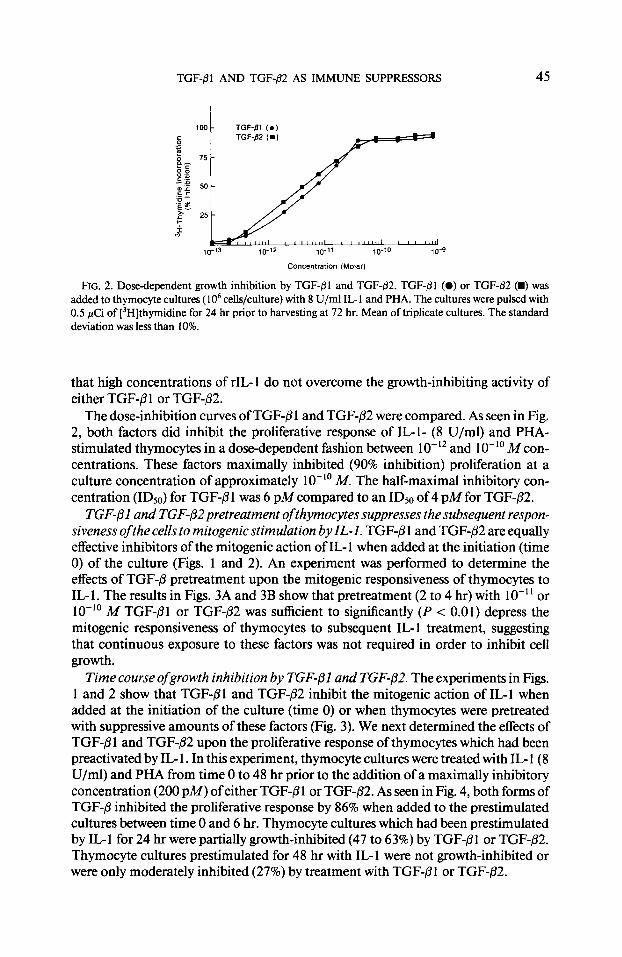
TGF-Pl AND TGF-/32 AS IMMUNE SUPPRESSORS 45
100 - TGF-,Tl (0)
6 TGF-82 (m)
PF
FIG. 2. Dose-dependent growth inhibition by TGF-fll and TGF-fl2. TGF-/31 (0) or TGF-82 (I) was added to thymocyte cultures ( 1 O6 cells/culture) with 8 U/ml IL- 1 and PHA. The cultures were pulsed with 0.5 pCi of [‘Hlthymidine for 24 hr prior to harvesting at 72 hr. Mean of triplicate cultures. The standard deviation was less than 10%.
that high concentrations of rIL- 1 do not overcome the growth-inhibiting activity of either TGF-P 1 or TGF-02.
The dose-inhibition curves of TGF-@I and TGF-P2 were compared. As seen in Fig. 2, both factors did inhibit the proliferative response of IL-l- (8 U/ml) and PHA- stimulated thymocytes in a dose-dependent fashion between lo-‘* and 10-l’ M con- centrations. These factors maximally inhibited (90% inhibition) proliferation at a culture concentration of approximately lo-” M. The half-maximal inhibitory con- centration (IDSo) for TGF-@ 1 was 6 pM compared to an ID50 of 4 p&f for TGF-P2.
TGF-/31 and TGF-02 pretreatment ofthymocytes suppresses the subsequent respon- siveness ofthecells to mitogenicstimulation by ZL-1. TGF-/31 and TGF-/32 are equally effective inhibitors of the mitogenic action of IL- 1 when added at the initiation (time 0) of the culture (Figs. 1 and 2). An experiment was performed to determine the effects of TGF-P pretreatment upon the mitogenic responsiveness of thymocytes to IL- 1. The results in Figs. 3A and 3B show that pretreatment (2 to 4 hr) with 10-l’ or lo-*’ M TGF-pl or TGF-P2 was sufficient to significantly (P < 0.01) depress the mitogenic responsiveness of thymocytes to subsequent IL- 1 treatment, suggesting that continuous exposure to these factors was not required in order to inhibit cell growth.
Time course ofgrowth inhibition by TGF$l and TGF-p2. The experiments in Figs. 1 and 2 show that TGF-PI and TGF$2 inhibit the mitogenic action of IL-l when added at the initiation of the culture (time 0) or when thymocytes were pretreated with suppressive amounts of these factors (Fig. 3). We next determined the effects of TGF-/31 and TGF-@2 upon the proliferative response of thymocytes which had been preactivated by IL- 1. In this experiment, thymocyte cultures were treated with IL- 1 (8 U/ml) and PHA from time 0 to 48 hr prior to the addition of a maximally inhibitory concentration (200 PM) ofeither TGF-/31 or TGF-fi2. As seen in Fig. 4, both forms of TGF-/3 inhibited the proliferative response by 86% when added to the prestimulated cultures between time 0 and 6 hr. Thymocyte cultures which had been prestimulated by IL1 for 24 hr were partially growth-inhibited (47 to 63%) by TGF-@l or TGF-/32. Thymocyte cultures prestimulated for 48 hr with IL- 1 were not growth-inhibited or were only moderately inhibited (27%) by treatment with TGF-P1 or TGF-82.
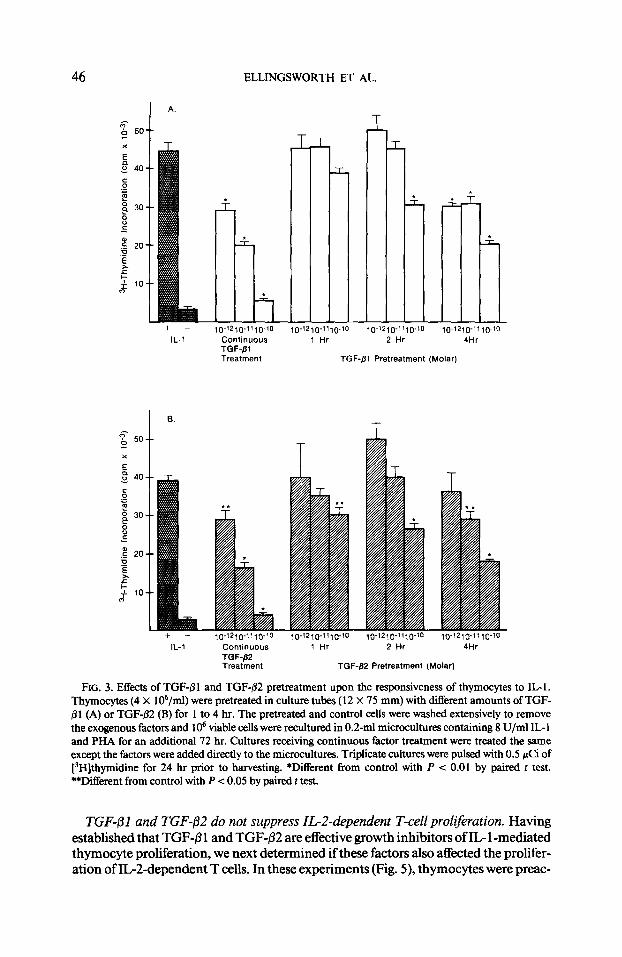
ELLINGSWORTH ET AL.
D-‘2l@“t,-~-‘0
Continuous TGF-fil Treatment
IL-l Continuous TGF-P2 Treatment
T- r
TGF-Pl Pretreatment (Molar)
,o-x2,0-“,0-‘0 ~0-12,0-11,0-‘0 10-‘2~fJ-“,0-‘0
1 Hr 2 Hr 4Hr
TGF-fl2 Pretreatment (Molarj
FIG. 3. Effects of TGF-8 1 and TGF+2 pretreatment upon the responsiveness of thymocytes to IL1 . Thymocytes (4 X 106/ml) were pretreated in culture tubes (12 X 75 mm) with different amounts of TGF- j31 (A) or TGF-@2 (B) for 1 to 4 hr. The pretreated and control cells were washed extensively to remove the exogenous factors and lo6 viable cells were reculturcd in 0.2-ml microcultures containing 8 U/ml IL- 1 and PHA for an additional 72 hr. Cultures receiving continuous factor treatment were treated the same except the factors were added directly to the microcultures. Triplicate cultures were pulsed with 0.5 pCi of [‘H]thymidine for 24 hr prior to harvesting. *Different from control with P < 0.01 by paired t test. **Different from control with P < 0.05 by paired t test.
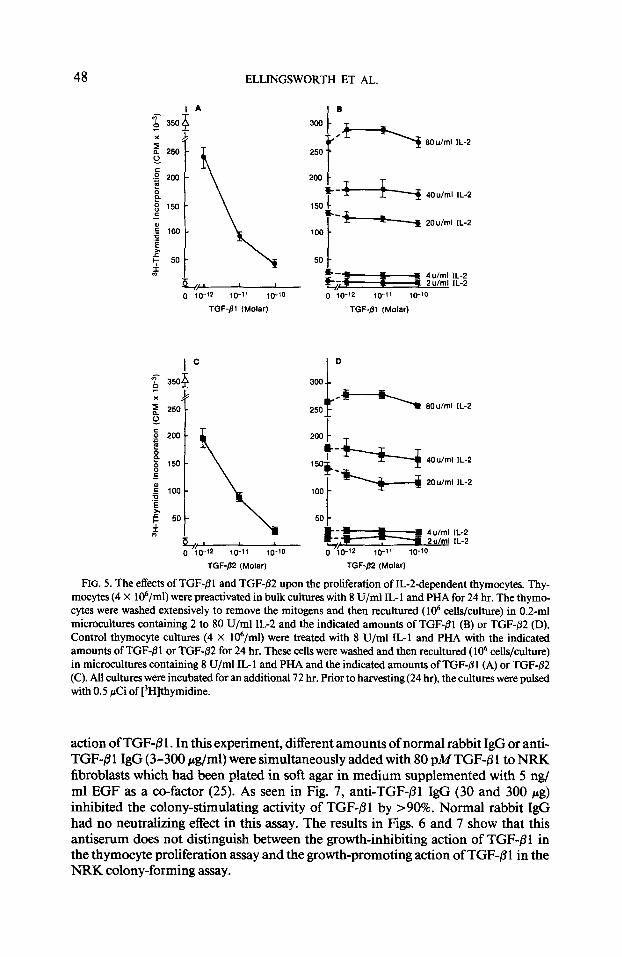
TGF-/31 and TGF-@2 do not suppress IL-2-dependent T-cell proliferation. Having established that TGF-@ 1 and TGF-j32 are effective growth inhibitors of IL- 1 -mediated thymocyte proliferation, we next determined if these factors also affected the prolifer- ation of IG2dependent T cells. In these experiments (Fig. 5), thymocytes were preac-
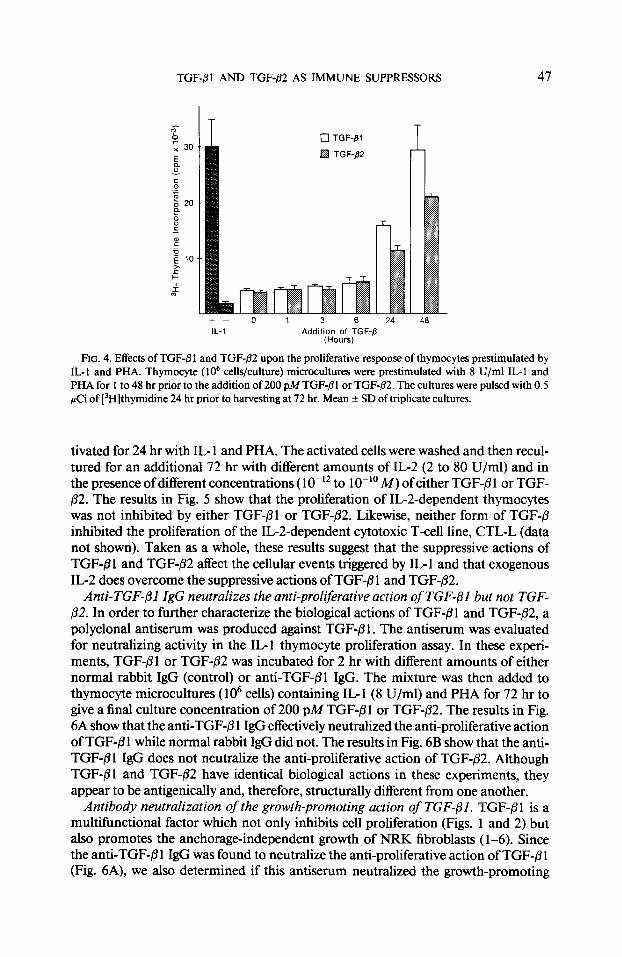
TGF-/31 AND TGF-82 AS IMMUNE SUPPRESSORS 47
0 TGF-/31
q TGF-/32
24 48 IL-1 Addition of TGF-fi
(Hours)
FIG. 4. Effects of TGF-fi 1 and TGF-02 upon the proliferative response of thymocytes prestimulated by IL-I and PHA. Thymocyte (lo6 cells/culture) microcultures were prestimulated with 8 U/ml IL-1 and PHA for 1 to 48 hr prior to the addition of 200 pMTGF-(31 or TGF-02. The cultures were pulsed with 0.5 PCi of [3H]thymidine 24 hr prior to harvesting at 72 hr. Mean f SD of triplicate cultures.
tivated for 24 hr with IL- 1 and PHA. The activated cells were washed and then recul- tured for an additional 72 hr with different amounts of IL-2 (2 to 80 U/ml) and in the presence of different concentrations ( lo-” to lo-” M) of either TGF-/3 1 or TGF- p2. The results in Fig. 5 show that the proliferation of IL-Zdependent thymocytes was not inhibited by either TGF-PI or TGF-82. Likewise, neither form of TGF-/3 inhibited the proliferation of the IL-Zdependent cytotoxic T-cell line, CTGL (data not shown). Taken as a whole, these results suggest that the suppressive actions of TGF-/3 1 and TGF-82 affect the cellular events triggered by IL- 1 and that exogenous IL-2 does overcome the suppressive actions of TGF+ 1 and TGF-02.
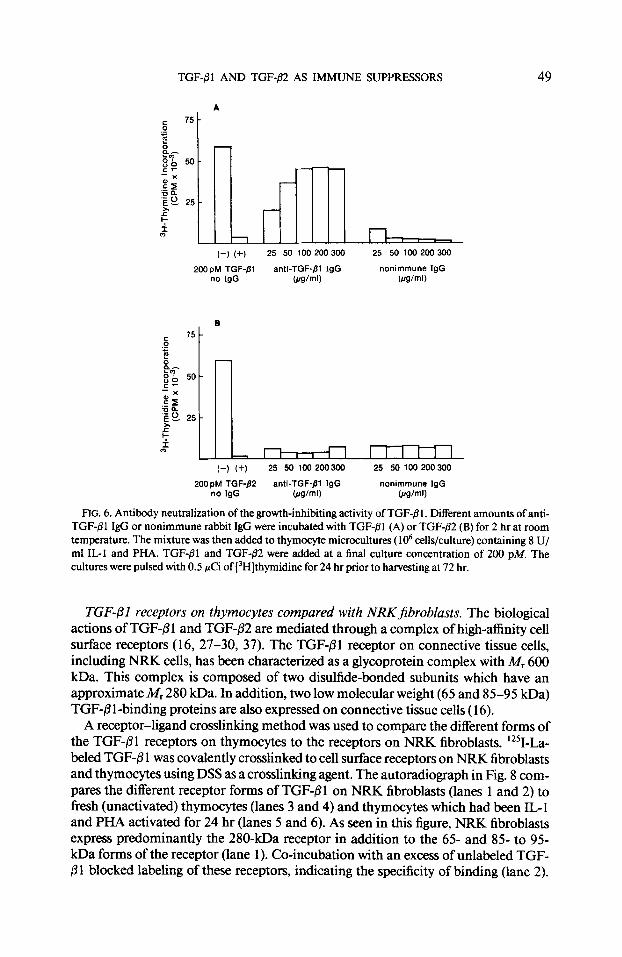
Anti-TGF-01 IgG neutralizes the anti-proliferative action of TGF-(31 but not TGF- p2. In order to further characterize the biological actions of TGF-B 1 and TGF-82, a polyclonal antiserum was produced against TGF+?l . The antiserum was evaluated for neutralizing activity in the IL-1 thymocyte proliferation assay. In these experi- ments, TGF-fll or TGF-P2 was incubated for 2 hr with different amounts of either normal rabbit IgG (control) or anti-TGF-j31 IgG. The mixture was then added to thymocyte microcultures ( lo6 cells) containing IL- I (8 U/ml) and PHA for 72 hr to give a final culture concentration of 200 pM TGF-P 1 or TGF-/32. The results in Fig. 6A show that the anti-TGF-81 IgG effectively neutralized the anti-proliferative action of TGF-B 1 while normal rabbit IgG did not. The results in Fig. 6B show that the anti- TGF-01 IgG does not neutralize the anti-proliferative action of TGF-p2. Although TGF-@I and TGF-02 have identical biological actions in these experiments, they appear to be antigenically and, therefore, structurally different from one another.
Antibody neutralization of the growth-promoting action of TGF-PI. TGF-/31 is a multifunctional factor which not only inhibits cell proliferation (Figs. 1 and 2) but also promotes the anchorage-independent growth of NRK fibroblasts (l-6). Since the anti-TGF-8 1 IgG was found to neutralize the anti-proliferative action of TGF-fi 1 (Fig. 6A), we also determined if this antiserum neutralized the growth-promoting
48 ELLINGSWORTH ET AL.
x +
g 250 -
.g 200 -
b P 8 150 - E
; loo-
B ly 50-
,I
$$iykyp 1 TGF-/31 (Molar)
lC lC $ 35op
x J
z 250 - z 250
1
&t f I 4u/ml IL-2 1 2u/ml IL-2
0 10-12 10-l’ lo-10
TGF-01 (Molar)
*Y SOu/ml IL-2
TGF-82 (Molar) TGF-/32 (Molar)
F~c. 5. The effects of TGF-01 and TGF-02 upon the proliferation of IL-Z-dependent thymocytes. Thy- mocytes (4 X 106/ml) were preactivated in bulk cultures with 8 U/ml IL-l and PHA for 24 hr. The thymo- cytes were washed extensively to remove the mitogens and then recultured ( lo6 cells/culture) in 0.2-ml microcultures containing 2 to 80 U/ml IL2 and the indicated amounts of TGF-01 (B) or TGF-02 (D). Control thymocyte cultures (4 X 106/ml) were treated with 8 U/ml IL1 and PHA with the indicated amounts of TGF-01 or TGF-@2 for 24 hr. These cells were washed and then recultured ( lo6 cells/culture) in microcultures containing 8 U/ml IL- 1 and PHA and the indicated amounts of TGF-j3 1 (A) or TGF-,92 (C). All cultures were incubated for an additional 72 hr. Prior to harvesting(24 hr), the cultures were pulsed with 0.5 &i of [3H]thymidine.
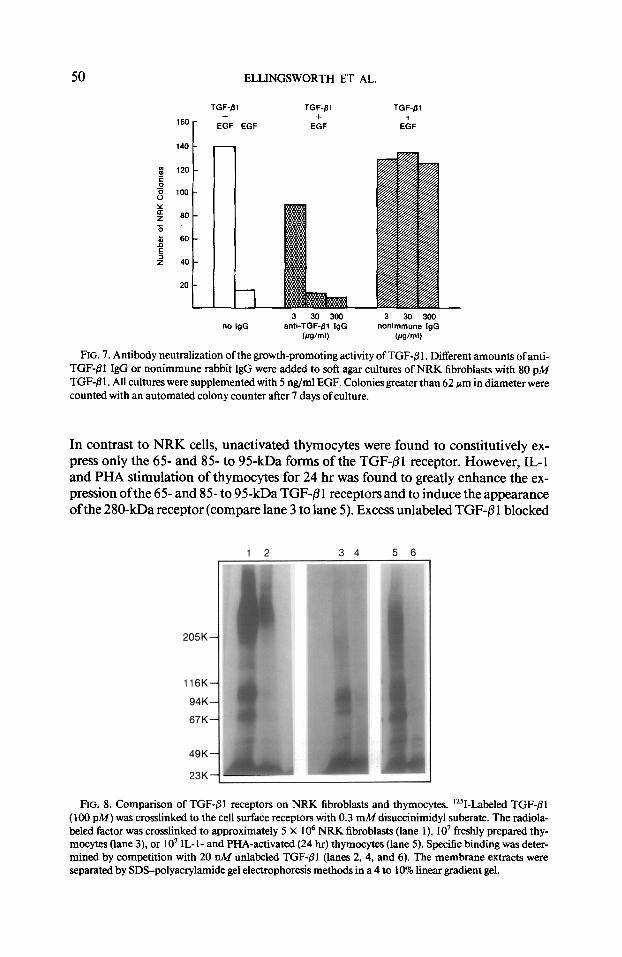
action of TGF-8 1. In this experiment, different amounts of normal rabbit IgG or anti- TGF-/I 1 IgG (3-300 &ml) were simultaneously added with 80 pA4 TGF-/3 1 to NRK fibroblasts which had been plated in soft agar in medium supplemented with 5 ngf ml EGF as a co-factor (25). As seen in Fig. 7, anti-TGF-/31 IgG (30 and 300 pg) inhibited the colony-stimulating activity of TGF-81 by >90%. Normal rabbit IgG had no neutralizing effect in this assay. The results in Figs. 6 and 7 show that this antiserum does not distinguish between the growth-inhibiting action of TGF-61 in the thymocyte proliferation assay and the growth-promoting action of TGF-8 1 in the NRK colony-forming assay.
TGF-,91 AND TGF-fl2 AS IMMUNE SUPPRESSORS 49
(-) (+I 25 50 100 200 300 25 50 100 200 300
200 pM TGF-/31 anti-TGF-fll IgG nonimmune IgG no IgG Wml) (Ilg/ml)
(3 (+) 25 50 100 200300 25 50 100 200 300
200 pM TGF-PP antl-TGF-81 IgG nonimmune IgG no IgG bdghl) 0.4VmU
FIG. 6. Antibody neutralization of the growth-inhibiting activity of TGF-j31. Different amounts of anti- TGF-j31 IgG or nonimmune rabbit IgG were incubate-d with TGF-Bl (A) or TGF-82 (B) for 2 hr at room temperature. The mixture was then added to thymocyte microcultures ( lo6 cells/culture) containing 8 U/ ml IL-I and PHA. TGF-PI and TGF-82 were added at a final culture concentration of 200 PM. The cultures were pulsed with 0.5 pCi of [‘Hlthymidine for 24 hr prior to harvesting at 72 hr.
TG’F-/31 receptors on thymocytes compared with NRKjibroblasts. The biological actions of TGF-8 1 and TGF-@2 are mediated through a complex of high-affinity cell surface receptors (16, 27-30, 37). The TGF-@l receptor on connective tissue cells, including NRK cells, has been characterized as a glycoprotein complex with A& 600 kDa. This complex is composed of two disulfide-bonded subunits which have an approximate M, 280 kDa. In addition, two low molecular weight (65 and 85-95 kDa) TGF-/3 1 -binding proteins are also expressed on connective tissue cells ( 16).
A receptor-ligand crosslinking method was used to compare the different forms of the TGF-01 receptors on thymocytes to the receptors on NRK fibroblasts. 12%La- beled TGF-@ 1 was covalently crosslinked to cell surface receptors on NRK fibroblasts and thymocytes using DSS as a crosslinking agent. The autoradiograph in Fig. 8 com- pares the different receptor forms of TGF-pl on NRK fibroblasts (lanes 1 and 2) to fresh (unactivated) thymocytes (lanes 3 and 4) and thymocytes which had been IL- 1 and PHA activated for 24 hr (lanes 5 and 6). As seen in this figure, NRK fibroblasts express predominantly the 280-kDa receptor in addition to the 65- and 85- to 95- kDa forms of the receptor (lane 1). Co-incubation with an excess of unlabeled TGF- /3 1 blocked labeling of these receptors, indicating the specificity of binding (lane 2).
50 ELLINGSWORTH ET AL.
TGF-,¶l TGF-~31 TGF-jl
160 + + +
EGF EGF EGF EGF
140
8 120 E _o s 100
g 60
z 5 60
5 z 40
20
3 30 300 3 30 300 no IgG anti-TGF-pl IgG nonimmune IgG
@g/ml) Celml)
FIG. 7. Antibody neutralization of the growth-promoting activity of TGF-@I. Different amounts of anti- TGF-01 IgG or nonimmune rabbit IgG were added to soft agar cultures of NRK fibroblasts with 80 pM TGF-@ 1. All cultures were supplemented with 5 rig/ml EGF. Colonies greater than 62 pm in diameter were counted with an automated colony counter alter 7 days of culture.
In contrast to NRK cells, unactivated thymocytes were found to constitutively ex- press only the 65 and 8% to 95kDa forms of the TGF-@1 receptor. However, IL-l and PHA stimulation of thymocytes for 24 hr was found to greatly enhance the ex- pression of the 65- and 85- to 95-kDa TGF-B 1 receptors and to induce the appearance of the 280-kDa receptor (compare lane 3 to lane 5). Excess unlabeled TGF-6 1 blocked
205K
116K
94K
67K
49K
23K
5 6
FIG. 8. Comparison of TGF-j31 receptors on NRK fibroblasts and thymocytes. ‘251-Labeled TGF-Bl (100 PM) was crosslinked to the cell surface receptors with 0.3 mM disuccinimidyl suberate. The radiola- beled factor was crosslinked to approximately 5 X IO6 NRK fibroblasts (lane 1), IO’ freshly prepared thy- mocytes (lane 3) or 10’ IL-l- and PHA-activated (24 hr) thymocytes (lane 5). Specific binding was deter- mined by competition with 20 nM unlabeled TGF-01 (lanes 2, 4, and 6). The membrane extracts were separated by SDS-polyacrylamide gel electrophoresis methods in a 4 to 10% linear gradient gel.
TGF-pl AND TGF-/32 AS IMMUNE SUPPRESSORS 51
116K
94K
67K
49K
94K
67K
49K
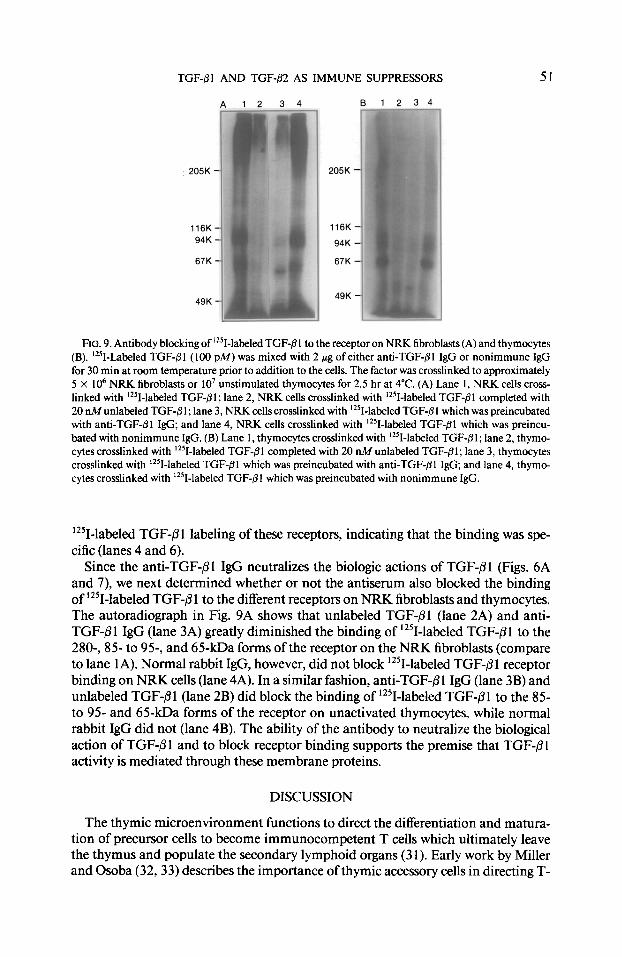
FIG. 9. Antibody blocking of ‘251-labeled TGF-fll to the receptor on NRK fibroblasts (A) and thymocytes (B). ‘251-Labeled TGF-/31 (100 PM) was mixed with 2 pg of either anti-TGF-@l IgG or nonimmune IgG for 30 min at room temperature prior to addition to the cells. The factor was crosslinked to approximately 5 x lo6 NRK fibroblasts or lo7 unstimulated thymocytes for 2.5 hr at 4°C. (A) Lane 1, NRK cells cross- linked with ‘*‘I-labeled TGF-@I; lane 2, NRK cells crosslinked with ‘251-labeled TGF$l completed with 20 nMunlabeled TGF-8 1; lane 3, NRK cells crosslinked with ‘251-labeled TGF-@I which was preincubated with anti-TGF-01 IgG; and lane 4, NRK cells crosslinked with ‘251-labeled TGF-/31 which was preincu- bated with nonimmune IgG. (B) Lane 1, thymocytes crosslinked with ‘251-labeled TGF-@I; lane 2, thymo- cytes crosslinked with ‘*‘I-labeled TGF-8 1 completed with 20 nM unlabeled TGF-,TI ; lane 3, thymocytes crosslinked with ‘251-labeled TGF-@I which was preincubated with anti-TGF-@l IgG; and lane 4, thymo- cytes crosslinked with ‘*‘I-labeled TGF-pl which was preincubated with nonimmune IgG.
1251-labeled TGF-PI labeling of these receptors, indicating that the binding was spe- cific (lanes 4 and 6).
Since the anti-TGF-01 IgG neutralizes the biologic actions of TGF-PI (Figs. 6A and 7), we next determined whether or not the antiserum also blocked the binding of 1251-labeled TGF-@ 1 to the different receptors on NRK fibroblasts and thymocytes. The autoradiograph in Fig. 9A shows that unlabeled TGF+l (lane 2A) and anti- TGF-01 IgG (lane 3A) greatly diminished the binding of 12?-labeled TGF-/31 to the 280-, 85 to 95-, and 65-kDa forms of the receptor on the NRK fibroblasts (compare to lane IA). Normal rabbit IgG, however, did not block ‘251-lab-eled TGF-/31 receptor binding on NRK cells (lane 4A). In a similar fashion, anti-TGF-01 IgG (lane 3B) and unlabeled TGF-@l (lane 2B) did block the binding of 12SI-labeled TGF-Pl to the 85- to 95- and 65-kDa forms of the receptor on unactivated thymocytes, while normal rabbit IgG did not (lane 4B). The ability of the antibody to neutralize the biological action of TGF-/?l and to block receptor binding supports the premise that TGF-@l activity is mediated through these membrane proteins.
DISCUSSION
The thymic microenvironment functions to direct the differentiation and matura- tion of precursor cells to become immunocompetent T cells which ultimately leave the thymus and populate the secondary lymphoid organs (3 1). Early work by Miller and Osoba (32,33) describes the importance of thymic accessory cells in directing T-

52 ELLINGSWORTH ET AL.
cell development. Indeed, numerous hormone-like protein factors have now been purified from thymic extracts. The biological actions and the developmental role of many of these factors have been characterized (see Refs. (34) and (35) for review). Although several T-cell growth-promoting factors have been identified, very little is known about the factors which limit the clonal growth of developing lymphocytes. Recently, we reported that TGF-@I is immunohistochemically associated with thymic accessory cells as well as lymphopoietic and hematopoietic cell populations in bone marrow and fetal liver (24), suggesting that this factor may have a role in the development of lymphopoietic and hematopoietic cells.
The association of TGF-81 with developing immune and hematopoietic cells has prompted us to investigate the role of TGF-01 and TGF-82 in the development, regulation, and function of the immune system. The evidence presented here shows that TGF-01 and TGF-@2 are equipotent antagonists of the mitogenic action of IL- 1 in a thymocyte proliferation assay. These factors inhibited thymocyte proliferation in a dose-dependent fashion over the dose range between 1 O-l2 and 10-r’ M. Kinetic studies show that these factors inhibit thymocyte proliferation when added at the start of the culture period (i.e., time 0 to 6 hr). TGF-/31 and TGF-/32 did not inhibit the proliferation of thymocytes which had been prestimulated by IL1 and PHA for 24 to 48 hr. Furthermore, neither factor inhibited the proliferation of IG2-dependent primary T cells (Fig. 4) or the proliferation of IL-Zdependent CTL-L cells (data not shown). These data demonstrate that the anti-proliferative action of TGF-Pl and TGF-82 occurs through the inhibition of cellular events triggered by IL-l and that exogenous IL-2 can overcome the suppressive actions of these factors.
The growth-inhibiting actions of TGF-/3 1 and TGF-/32 are not restricted to devel- oping T cells. We have recently reported that these factors may also have a regulatory role in B lymphopoiesis (36). These studies show that TGF-6 1 and TGF-/32 are potent inhibitors of the growth and differentiation of developing B cells. These factors were found to suppress &ght-chain gene expression, but did not affect heavy-chain gene expression or the expression of Ia antigen on developing B cells. Similar studies to evaluate the effects of TGF-/3 1 and TGF-P2 in myeloid and monocyte development, and the interaction of these factors with the different colony-stimulating factors, are currently in progress.
Although the mechanism whereby TGF-@l and TGF+32 suppress cell growth is not known, it is evident that the biological actions are mediated through a complex of high and low molecular weight cell surface receptors (16, 27-30). Cheifetz et al. ( 16) have used an affinity crosslinking method to show that the TGF-/3 1 receptor is a complex of three glycoproteins which have A& 280,85-95, and 65 kDa on connective tissue cells. Using a similar crosslinking method, we show that thymocytes also ex- press specific TGF-P 1 receptors. These results show that fresh, unstimulated thymo- cytes appear to constitutively express only the low molecular weight (65 and 85-95 kDa) forms of the TGF-61 receptor. Upon mitogen stimulation by IL- 1 and PHA for 24 hr, there was an apparent enhancement of the low molecular weight receptors (65 and 85-95 kDa) and the appearance of the 280-kDa receptor. These results suggest, therefore, that a regulatory relationship may exist between the growth-stimulating action of IL-l and the growth-suppressing action of TGF-Pl in developing T cells. Because cells of the immune system appear to have very few (250 to 500 receptors/ cell) TGF-B 1 receptors (38, 39), we were unable to accurately quantitate the number of receptors on resting and IL- 1 -activated thymocytes.

TGF-fll AND TGF-82 AS IMMUNE SUPPRESSORS 53
Multiple forms of TGF-/3 have been shown to be present in porcine ( 16) and bovine (2 1,22) tissues. The biological significance of these different forms of TGF-/3 remains unclear at this time. In this study, we were unable to show any significant difference in the potency or the biological action of either factor. It was evident, however, that TGF-Bl and TGF-P2 likely have very different tertiary structures since antibodies raised against TGF-P 1 only neutralized the biological action of TGF-fi 1, but did affect the biological action of TGF-P2. Additional comparative studies will be needed in order to determine if there are biological differences between these different forms of TGF-0.
In summary, the results presented here show that TGF-P 1 and TGF-02 are equipo- tent inhibitors of IL- 1 -mediated thymocyte proliferation. The anti-proliferative ac- tion of both factors was most effective at inhibiting the proliferation of thymocytes when added with IL-l/PHA at the start (O-6 hr) of the culture. Thymocytes which had been prestimulated for 24-48 hr by IL- 1 /PHA were less responsive or were refi-ac- tory to inhibition by these factors. IL-2-dependent T-cell proliferation was not inhib- ited by either TGF-01 or TGF-P2. It was shown that the anti-proliferative action of TGF-/3 1 is mediated through specific receptors for TGF-@ 1 on thymocytes. Mitogenic stimulation did appear to enhance the expression of the low molecular weight (65 and 85-95 kDa) TGF-fi 1 receptors and did stimulate the expression of the high molecular weight (280 kDa) form of the TGF-/31 receptor on thymocytes. These results suggest that a regulatory relationship may exist between the growth-stimulating action of IL- 1 and the growth-suppressing action of TGF-B 1 and that these factors may function to regulate the clonal growth of developing T cells.
REFERENCES
1. Anzano, M., Roberts, A., Meyers, C., Komoriya, A., Lamb, L., Smith, J., and Spom, M., Cancer Rex 42,4776, 1982.
2. As&an, R., Frolik, C., Roberts, A., Miller, D., and Spom, M., Cell 136,35, 1984. 3. Pircher, R., Lawrence, D., and Jullien, P., CancuRes. 44,5538, 1984. 4. Roberts, A., Anzano, M., Lamb, L., Smith, J., Frolik, C., Marquardt, H., Todaro, G., and Spom, M.,
Nature (London) 295,4 17, 1982. 5. Anzano, M., Roberts, A., Smith, J., Spom, M., and DeLarco, J., Proc. Natl. Acad. Sci. USA 80,6264,
1983. 6. DeLarco, J., and Todaro, G., Proc. Natl. Acad. Sci. USA 75,4001, 1978. 7. Reynolds, F., Todaro, G., Fryling, C., and Stephenson, J., Nature (London) 292,259, 198 1. 8. Derynck, R., Roberts, A., Winkler, A., Chen, E., and Goeddel, D., Ceil38,287, 1984. 9. Derynck, R., Jarrett, J., Chen, E., Eaton, D., Bell, J., Assoian, R., Roberts, A., Spom, M., and Goeddel,
D.,Nature(London)316,701, 1985. 10. Derynck, R., Jarrett, J., Chen, E., and Goeddel, D., J. Biol. Chem. 261,4377, 1986. Il. Frolik, C., Wakefield, L., Smith, D., and Spom, M., J. Biol. Chem. 259, 1095, 1984. 12. Roberts, A., Frolik, C., Anzano, M., and Spom, M., Fed. Proc. 42,262 1, 1983. 13. Roberts, A., Lamb, L., Newton, D., Spom, M., DeLarco, J., and Todaro, G., Proc. Natl. Acad. Sci.
USA 77,3494, 1980. 14. Frolik, C., Dart, L., Meyers, C., Smith, D., and Spom, M., Proc. Natl. Acad. Sci. USA 80,3676, 1983. 15. Assoian, R., Komoriya, A., Meyers, C., Miller, D., and Spom, M., J. Biol. Chem. 258,7 155, 1983. 16. Cheifetz, S., Weatherbee, J., Tsang, M., Anderson, J., Mole, J., Lucas, R.‘, and Massagud, J., Cell 48,
409,1987. 17. Roberts, A., Anzano, M., Meyers, C., Wideman, J., Blather, R., Pan, Y., Stein, S., Lehrman, S., Smith,
J., Lamb, L., and Spom, M., Biochemistry 22,5692, 1983. 18. Seyedin, S., Thomas, T., Thompson, A., Rosen, D., and Piez, K., Proc. Natl. Acad. Sci. USA 82,2267,
1985. 19. Seyedin, S., Thomas, A., Rosen, D., and Piez, K., J. Cell Biol. 97, 1950, 1983.

54 ELLINGSWORTH ET AL.
20. Thompson, A., Piez, K., and Seyedin, S., Exp. Cell Res. 157,483, 1985. 21. Seyedin, S., Thompson, A., Bentz, H., Rosen, D., McPherson, J., Conti, A., Siegel, N., Galluppi, G.,
and Piez, K., J. Biol. Chem. 261,5693, 1986. 22. Seyedin, S., Segarini, P., Rosen, D., Thompson, A., Bentz, H., and Graycar, J., J. Biol. Chem. 262,
1946,1987. 23. Spom, M., and Roberts, A., J. Cell. Biochem. (Suppl.) HA, 11 (abstract), 1987.
24. Ellingsworth, L., Brennan, J., Fok, R., Rosen, D., Bentz, H., Piez, K., and Seyedin, S., J. Biol. Chem. 261,12362,1986.
25. Roberts, A., Frolik, C., Anzano, M., Assoian, R., and Spom, M., In “Cell Culture Methods for Molecu- lar and Cell Biology” (D. Barnes, D. Sirbasku, and G. Sato, Eds.), pp. 18 1- 194. Alan R. Liss, New York, 1984.
26. Laemmli, U., Nature (London) 227,680,1970. 27. Tucker, R., Branum, E., Shipley, G., Ryan, R., and Moses, H., Proc. Natl. Acad. Sci. USA 81,6751,
1984. 28. Massague, J., 1. Biol. Chem. 260,7059, 1985. 29. Massague, J., Cancer Cells 3,73, 1985. 30. Massagui, J., and Like, B., J. Biol. Chem. 260,2636, 1985. 3 1. Owens, J., In “Ontogeny of Acquired Immunity,” p. 35. A Ciba Foundation Symposium, Elsevier/
North-Holland, New York/Amsterdam, 1972. 32. Miller, J., Lancet 2,748, 1961. 33. Osoba, D., and Miller, J., Nature (London) 199,653, 1963. 34. Dardenne, M., and Bach, J., In “The Thymus gland” (M. Kendall, Ed.), p. I 13. Academic Press, New
York, 198 1. 35. Low, T., and Goldstein, A., In “Methods in Enzymology” (G. Di Sabato, J. J. Langone, and H. Van
Vunakis, Eds.), Vol. 116, pp. 213-219. AcademicPress, Orlando, FL, 1985. 36. Lee, G., Ellingsworth, L., Gillis, S., Wall, R., and Kincade, P., J. Exp. Med., 166, 1290, 1987. 37. Segarini, P., Roberts, A., Rosen, D., and Seyedin, S., J. Biol. Chem., 262, 14655, 1987. 38. Kehrl, J., Wakefield, L., Roberts, A., Jakowlew, S., Alvarez-Mon, M., Derynck, R., Spom, M., and
Fauci, A., J. Exp. Med. 63, 1037, 1986. 39. Kehrl, J., Roberts, A., Wakefield, L., Jakowlew, S., Spom, M., and Fauci, A., J. Zmmunol. 137,3855,
1986. 40. Ristow, H., Proc. Natl. Acad. Sci. USA83,5531,1986.
Copyright © 2022 FDOKUMEN




















