
The beneficial effects of the Lab4 probiotic consortium in ...

ARTHRITIS & RHEUMATISMVol. 46, No. 3, March 2002, pp 806–817DOI 10.1002/art.10170© 2002, American College of Rheumatology
The Beneficial Effect of Interleukin-12 on Arthritis Induced byGroup B Streptococci Is Mediated by Interferon-� and
Interleukin-10 Production
Manuela Puliti,1 Christina von Hunolstein,2 Francesco Bistoni,1 Paolo Mosci,1
Graziella Orefici,2 and Luciana Tissi1
Objective. To assess the effect of interleukin-12(IL-12) administration on the evolution of systemicinfection and septic arthritis induced by group B strep-tococci (GBS) in mice.
Methods. CD1 mice were inoculated intrave-nously with arthritogenic strain 1/82 of type IV GBS.Exogenous murine IL-12 was administered intraperito-neally 18 hours or 5 days after infection with 1 � 107
GBS, at doses ranging from 0.5 to 2.5 �g per mouse.Mice were monitored daily for survival and for signs ofarthritis. In a subsequent set of experiments, mice werekilled at selected times for examination of bacterialclearance, histopathologic changes in the joints, andcytokine production.
Results. IL-12 administration before the onset ofclinical signs had a beneficial effect on GBS-inducedarthritis and was clearly dose-dependent. The 2.5-�gdose per mouse totally prevented death from GBS-induced arthritis. The decrease in pathology was asso-ciated with a reduction of the bacterial burden and achange in the cytokine profile. In particular, systemicand joint levels of interferon-� (IFN�) and IL-10 signif-icantly increased in mice treated with IL-12, whereas adecrease in IL-6 and IL-1� production was observed.The beneficial effects of IL-12, in terms of the incidenceand severity of articular lesions, were reversed by
coadministration of anti-IFN� or anti–IL-10–neutralizing antibodies.
Conclusion. The findings of this study demon-strate that IL-12 is important in controlling the cytokineproduction that leads to the evolution of GBS-inducedexperimental arthritis. The amelioration of articularlesions is mostly attributable to IL-12–induced IFN�,but with a relevant participation of IL-12–induced IL-10.
Group B streptococci (GBS) are a leading causeof life-threatening infections in neonates and younginfants (1). Invasive neonatal GBS infection has eitheran early (usually in the first 24 hours after birth) or a late(7 days after birth) onset. Common manifestations ofGBS disease in neonates include pneumonia, septice-mia, meningitis, bacteremia, and bone or joint infections(1). Invasive disease caused by GBS has also beenrecognized in adults (2,3).
Septic arthritis is one of the clinical manifesta-tions of late-onset GBS infection in neonates (1) andrequires prolonged antibiotic treatment to ensure anuncomplicated outcome. In adults, septic arthritiscaused by GBS infection is often associated with age andrisk factors such as diabetes mellitus, cancer, cardiovas-cular disease, chronic renal insufficiency, alcoholism,intravenous drug abuse, human immunodeficiency virusinfections, neurologic disease, and cirrhosis (4–7). GBSarthritis is usually hematogenously acquired, and themost frequently affected joints are the hip, ankle, andwrist (1).
In our mouse model of hematogenously inducedgroup B streptococcal arthritis, mice inoculated with thereference serotype IV GBS strain manifested clinicalsigns of arthritis, characterized by early onset and evo-lution from an acute exudative synovitis to permanentlesions, with irreversible joint damage and/or ankylosis
Supported by the Ministero dell’Universita e della RicercaScientifica 1999–2000, “Biotechnology-derived strategies of vaccina-tion and therapy,” Italy.
1Manuela Puliti, PhD, Francesco Bistoni, MD, Paolo Mosci,MD, Luciana Tissi, PhD: University of Perugia, Perugia, Italy; 2Chris-tina von Hunolstein, PhD, Graziella Orefici, MD: Istituto Superiore diSanita, Rome, Italy.
Address correspondence and reprint requests to LucianaTissi, PhD, Microbiology Section, Department of Experimental Med-icine and Biochemical Sciences, University of Perugia, Via del Gio-chetto, Perugia 06122, Italy. E-mail: [email protected].
Submitted for publication May 30, 2001; accepted in revisedform October 15, 2001.
806

(8). This laboratory mouse model offers outstandingpotential for GBS arthritis, in that bacteremia persistsfor more than 10 days after GBS infection, and localiza-tion of articular lesions is similar to those observed inhumans.
We initially demonstrated that induction of GBSarthritis depends on the viability and number of micro-organisms injected (8), the presence and amount ofbacterial capsule, as well as the presence of sialic acid inthe capsular polysaccharide (9). Moreover, in thismodel, the production of interleukin-6 (IL-6), IL-1�,and tumor necrosis factor � (TNF�) increased in re-sponse to GBS infection in sera and joints. In particular,a direct correlation between IL-6 and IL-1� concentra-tions in the joints and the severity of arthritis wasobserved (10). Subsequently, we found that interferon-�(IFN�) exerts detrimental effects on GBS-induced ar-thritis when administered before infection, while bene-ficial effects were observed when the cytokine was given18 hours after infection (11). Less severe joint pathologycorrelated with low levels of IL-6 and IL-1� in the joints,whereas rapid onset and worsening of articular lesionscorresponded to early and sustained levels of IL-6.
IL-12, a heterodimeric cytokine produced mainlyby mononuclear phagocytes and B cells (12,13), is apotent inducer of IFN� and directs the generation ofTh1 responses (14–16). It is therefore a major modula-tor of inflammation and immune responses and is likelyto play a significant role in the pathogenesis of infectionsand autoimmune diseases (16).
The participation of IL-12 in the arthritic processhas been extensively investigated. In type II collagen–induced arthritis (CIA), IL-12 possibly has a biphasicrole, such that early administration of low-dose IL-12 isproinflammatory, whereas late administration of high-dose IL-12 may be antiinflammatory due to an IL-10–dependent mechanism (17,18). Anti–IL-12 treatmentduring induction of CIA was shown to dramaticallyattenuate the severity of disease, although it did notlower the incidence of arthritis (19). Neutralizing anti–IL-12 antibodies significantly reduced the severity ofLyme arthritis, accompanied by decreased IFN� serumlevels, in C3H mice infected with Borrelia burgdorferi(20). Suppression of acute streptococcal cell wall–induced arthritis was observed upon endogenous IL-12blockade, and there was a concomitant reduction oflocal levels of IL-1�, IL-12, and IFN� and up-regulationof IL-10, IL-1 receptor antagonist protein, and messen-ger RNA levels in synovial tissues (21).
In the present study, we focused on the potentialrole of IL-12 in GBS-induced arthritis. Our data on in
vivo treatment with recombinant murine IL-12 demon-strate that IL-12 can beneficially influence the severityof GBS-induced arthritis, depending on the dose ofcytokine administered. Beneficial effects are associatedwith a decrease in the bacterial burden and a change inthe cytokine profile. These observations provide evi-dence that IL-12 is an important immunomodulator inthe pathogenesis of GBS-induced arthritis.
MATERIALS AND METHODS
Animals. Female outbred CD1 mice were obtainedfrom Charles River Breeding Laboratories (Calco, Italy).Eight-week-old to 10-week-old mice were used in the presentexperiments.
Microorganisms and induction of arthritis. Type IVGBS (reference strain GBS 1/82) was used throughout thestudy. For experimental infection, the microorganisms weregrown overnight at 37°C in Todd-Hewitt broth (Oxoid, Bas-ingstoke, UK) and then washed and diluted in RPMI 1640medium (Gibco Life Technologies, Milan, Italy). The inocu-lum size was estimated turbidimetrically. Viability counts wereperformed by plating on tryptic soy agar/5% sheep blood agar(blood agar) and overnight incubation under anaerobic condi-tions at 37°C.
A bacterial suspension was prepared in RPMI 1640medium. Mice were inoculated intravenously via the tail veinwith 1 � 107 GBS in a volume of 0.5 ml of RPMI 1640 permouse. Control mice were injected in the same way with 0.5 mlof RPMI 1640 medium.
Cytokines and antibodies. Recombinant murine IL-12(purchased from Sigma, Milan, Italy) was diluted according tothe manufacturer’s recommendations in phosphate bufferedsaline (PBS) containing 0.1% bovine serum albumin (BSA)and injected intraperitoneally in a 0.25 ml volume at differentdoses, ranging from 0.5 to 2.5 �g per mouse. IL-12 wasadministered 18 hours after infection, just before the onset ofclinical signs of arthritis. In another set of experiments, IL-12(2.5 �g per mouse) was administered to animals with estab-lished GBS arthritis 5 days (120 hours) after infection. Thetime and doses were chosen on the basis of our preliminaryresults and treatment schedules used in other experimentalmodels of arthritis or GBS infection (18,22). Control micereceived PBS plus BSA according to the same protocol.
Culture supernatants from the hybridoma lines R4-6A2 and JES5-2A5 (American Type Culture Collection, Rock-ville, MD) were used as a source of rat IgG1 monoclonalantibodies (mAb) to murine IFN� and IL-10, respectively.Antibody purification was performed by affinity chromatogra-phy with protein A–Sepharose CL-4B (Amersham PharmaciaBiotech, Uppsala, Sweden). The protein concentration wasdetermined by measuring the optical density at 280 nm. Theneutralizing activity was assessed in bioassays using L929 cellsfor anti-IFN� mAb (median neutralizing dose 0.045 �g/ml)and MC/9 cells for anti–IL-10 mAb (median neutralizing dose3 �g/ml). The mAb (1 mg per mouse) were given intraperito-neally at the time of IL-12 administration. These doses werechosen on the basis of preliminary titration experiments,showing complete neutralization of both in vivo cytokine
BENEFICIAL EFFECT OF IL-12 ON GBS-INDUCED ARTHRITIS 807

levels, as assessed by enzyme-linked immunosorbent assays(ELISAs). Control mice received normal rat IgG1 (Sigma).
Clinical evaluation of arthritis and mortality. Miceinjected with GBS and treated or not treated with IL-12 and/ormAb as described above were evaluated for signs of arthritisand for mortality. Mortality was recorded at 24-hour intervalsfor 30 days. After challenge, mice were examined daily by 2independent observers (LT and MP) for 2 months to evaluatethe presence of joint inflammation. Arthritis was defined asvisible erythema or swelling of at least 1 joint. The intensity ofarthritis in each limb was scored clinically (arthritis index),using a 0–3 scale, where 0 � normal, 1 � mild swelling anderythema, 2 � moderate swelling and erythema, and 3 �marked swelling, erythema, and/or ankylosis (maximum score12 per mouse). The arthritis index (mean � SD) was con-structed by dividing the total score by the number of animals ineach experimental group.
Leukocyte counts. Eighteen hours after GBS infection(or not), mice were treated with 2.5 �g of IL-12 or PBS permouse; 18 and 120 hours after this final treatment, mice werebled from the retroorbital sinus. Blood cell counts wereperformed in a hemocytometer after diluting anticoagulatedblood 1:20 in Turk’s solution. Differential cell counts (counting200 leukocytes per slide) were performed on blood smearsstained with May-Grunwald–Giemsa stain.
GBS growth in blood and joints. Blood and jointinfections in GBS-infected mice treated or not treated withIL-12 or with PBS plus BSA were determined by measuring thenumber of colony-forming units (CFU) at different times afterinoculation. Blood samples were obtained by retroorbital sinusbleeding before the mice were killed. Ten-fold dilutions inRPMI 1640 medium were prepared, and 0.1 ml of eachdilution was plated in triplicate on blood agar and incubatedunder anaerobic conditions for 24 hours. The number of CFUwas determined, and the results were expressed as the numberof CFU per ml of blood.
All wrist and ankle joints from each mouse wereremoved, weighed, and homogenized in toto in sterile RPMI1640 medium (1 ml/100 mg of joint weight). After homogeni-zation, all tissue samples were diluted and plated in triplicateon blood agar. The results were expressed as the number ofCFU per milliliter of joint homogenate.
Preparation of samples for cytokine assessment.Blood samples from the different experimental groups wereobtained by retroorbital sinus bleeding at different times afterinfection, before the mice were killed. Sera were stored at�80°C until analyzed. Joint tissues were prepared as previouslydescribed (10). Briefly, all wrist and ankle joints from eachmouse were removed and homogenized in toto in 1 ml/100 mgjoint weight of lysis medium (RPMI 1640 containing 2 mMphenylmethylsulfonyl fluoride and 1 �g/ml final concentrationof aprotinin, leupeptin, and pepstatin A). The homogenizedtissues were then centrifuged at 2,000g for 10 minutes. Super-natants were sterilized using a Millipore filter (0.45 �m) andstored at �80°C until analyzed.
To measure ongoing IL-12 production in the spleenduring GBS infection, the spleens from 3 animals were pooledand a single-cell suspension was prepared as previously de-scribed (11). Briefly, cells were washed and resuspended inRPMI 1640 containing 10% fetal calf serum (Gibco) to adensity of 5 � 106 cells/ml. One-milliliter aliquots were cul-
tured in 24-well plates for 24 hours at 37°C in a humidifiedatmosphere containing 5% CO2. Spleen cells from uninfectedanimals were used as controls. After culturing, supernatantswere removed and stored at �80°C until assayed.
Cytokine assays. Concentrations of IL-6, IL-1�, IFN�,IL-10, IL-12 p70, and TNF� in the biologic samples weremeasured with commercial ELISA kits (Amersham PharmaciaBiotech, Amersham, UK) according to the manufacturer’srecommendations. Results were expressed as picograms permilliliter of serum or supernatant from joint homogenates orfrom spleen cultures. The detection limits of the assays were 7pg/ml for IL-6, 3 pg/ml for IL-1�, 10 pg for IFN�, 12 pg/ml forIL-10, 5 pg/ml for IL-12, and 10 pg/ml for TNF�.
Histologic analysis. To confirm the clinical features ofarthritis, groups of mice inoculated with GBS and treated ornot treated with IL-12 were examined on selected days afterinfection for histopathologic features of arthritis. Ankle andwrist joints were removed aseptically, fixed in formalin (10%volume/volume) for 24 hours, decalcified in trichloroaceticacid (5% volume/volume) for 7 days, dehydrated, embedded inparaffin, sectioned at 3–4 �m, and stained with hematoxylinand eosin. Samples were examined under blinded conditions,and a histologic score was assigned to each joint based on theextent of inflammation, pannus formation, cartilage damage,and bone erosion. A scale of 0 (normal) to 4 (severe) was used.
Statistical analysis. Differences in the number ofCFU, white blood cell counts, and cytokine concentrationsbetween the groups of mice were analyzed by Student’s t-test.Between-group differences in survival data and incidence ofarthritis were analyzed by chi-square test, and differences inthe arthritis index by Student’s t-test. Each experiment wasrepeated 2–3 times. P values less than 0.05 were consideredsignificant.
RESULTS
Clinical course of GBS arthritis and assessmentof endogenous IL-12 production. As previously de-scribed (8) and as summarized in Figure 1A, the clinicalonset of arthritis occurred in 30% of the mice as early as24–30 hours after GBS infection. One week later, theincidence of arthritis increased to �80%, and the max-imum prevalence was observed on day 10 after inocula-tion, when 90% of the mice displayed clinical signs ofarthritis. A gradual increase in the severity of arthritiswas observed from day 1 to day 20 (Figure 1B); at latertime points, ankylosis was also observed. A 30% mortal-ity rate was recorded during the course of infection(Figure 2A).
Endogenous IL-12 was measured in sera, super-natants from the joints, and supernatants from culturedspleen cells explanted at various times after infectionwith GBS. No production of IL-12 was observed in anyof the biologic samples (data not shown).
Effect of IL-12 treatment on mortality and ar-thritis. To investigate the effect of IL-12 treatment onthe course of GBS infection, 3 different doses of the
808 PULITI ET AL

cytokine (0.5, 1.25, and 2.5 �g per mouse) were admin-istered intraperitoneally 18 hours after infection. In-fected control mice received PBS plus BSA (referred tohereafter as the PBS group) instead of IL-12. Mortalityrates and appearance of arthritis were evaluated. Asshown in Figure 2A, no significant differences in mor-tality rates between the control mice and the IL-12–treated mice were found, except when the cytokine wasadministered at the highest dose (2.5 �g per mouse),which allowed the survival of 100% of the animals.
A clear dose-dependent effect of IL-12 adminis-tration on the incidence and severity of arthritis wasevident in the control mice (Figures 2B and C). In fact,treatment of mice with the highest dose (2.5 �g permouse) dramatically reduced the incidence of arthritis,with only 25% of the mice showing a mild articular
swelling and erythema, and the arthritis index neverexceeding a mean value of 0.6 throughout the observa-tion period. Treatment with 1.25 �g of IL-12 per mousedid not alter the incidence of arthritis. However, asignificant decrease in the severity of arthritis was
Figure 1. Time course of arthritis in CD1 mice after intravenousinoculation of group B streptococci (GBS; 1 � 107 colony-formingunits per mouse). Mice were inoculated on day 0 and then examineddaily for the appearance of articular lesions. A, Incidence of GBSarthritis (percentage of mice with visible arthritis). B, Arthritis index(clinical severity of arthritis, evaluated as described in Materials andMethods). Values are the mean � SD of 3 separate experiments (n �20 mice per group).
Figure 2. Effect of exogenous interleukin-12 (IL-12) administrationon survival and the incidence and severity of group B streptococci(GBS)–induced arthritis in CD1 mice infected intravenously with GBS(1 � 107 colony-forming units per mouse). Murine recombinant IL-12or phosphate buffered saline plus 0.1% bovine serum albumin (PBS)was given intraperitoneally 18 hours after GBS infection. A, Survivalcurves. Deaths were recorded at 24-hour intervals for 30 days. Datarepresent the cumulative results of 3 separate experiments. B, Inci-dence of arthritis (percentage of mice with visible arthritis). C,Arthritis index (clinical severity of arthritis, evaluated as described inMaterials and Methods). Values are the mean � SD of 3 separateexperiments (n � 10 mice per group). ✳ � P � 0.001 versusPBS-treated mice; † � P � 0.05 versus PBS-treated mice.
BENEFICIAL EFFECT OF IL-12 ON GBS-INDUCED ARTHRITIS 809

Figure 3. Histologic features on day 7 after GBS infection in the joints of mice infected with GBS (1 � 107 colony-forming units per mouse) and,at 18 hours after infection, treated intraperitoneally with IL-12 or PBS. Joint sections were stained with hematoxylin and eosin and examined underblinded conditions. A and B, In mice treated with PBS, there is severe arthritis (histologic score 4), with massive subcutaneous, periarticular, andintraarticular infiltrates (A), and cartilage destruction and bone erosion (B). C and D, In mice treated with 1.25 �g of IL-12, there is moderatearthritis (histologic score 2), with periarticular and subcutaneous infiltrates (C), and a moderate cell influx in the articular cavity (D), but no jointdestruction. E and F, In mice treated with 2.5 �g of IL-12, arthritis is absent (E), and there are only a few inflammatory cells in the subcutaneoustissue (F). B, D, and F are higher-magnification views of the boxed areas in A, C, and E, respectively. (Original magnification � 2.5 in A, C, and E;� 10 in B, D, and F.) See Figure 2 for definitions.
810 PULITI ET AL
observed; on day 20, the arthritis index was 2.4 � 0.4(mean � SD), compared with 4.3 � 0.5 in the controlinfected mice. No changes in the incidence or severity ofarticular lesions were observed with the 0.5-�g dose permouse.
The beneficial effect of IL-12 administration wasconfirmed by histologic examination of the joints. Infact, on day 7 after challenge, a severe arthritis, with apurulent exudate filling the articular cavities (Figure 3A)and bone and cartilage erosion (Figure 3B), was ob-served in GBS-infected mice. In the joints of micetreated with 1.25 �g of IL-12, the inflammatory infiltratewas limited to the subcutaneous and periarticular tissues(Figure 3C), with a moderate cell influx in the articularcavity (Figure 3D), and no signs of cartilage or boneerosion. Administration of the highest dose of IL-12 (2.5�g per mouse) totally prevented the signs of inflamma-tion (Figure 3E), with only a few inflammatory cellspresent in the subcutaneous tissue (Figure 3F).
To further define the effect of IL-12 treatment onGBS-induced arthritis, the highest dose of the cytokine(2.5 �g per mouse) was administered on day 5 afterinfection to mice with established arthritis (mean � SDarthritis index 3.5 � 0.4). Control mice with comparablearthritis severity received PBS plus BSA instead ofIL-12. Although a slight improvement was observed inthe first 2 days after IL-12 treatment (mean arthritisindex of 3.0 � 0.3 versus 3.9 � 0.5 in control infectedmice), this effect was transient, since 5 days after treat-ment, no differences in the severity of arthritis wereevident between the 2 experimental groups (data notshown).
Effect of IL-12 administration on peripheralblood leukocyte counts. Since in vivo IL-12 administra-tion was reported to induce neutropenia, lymphopenia,
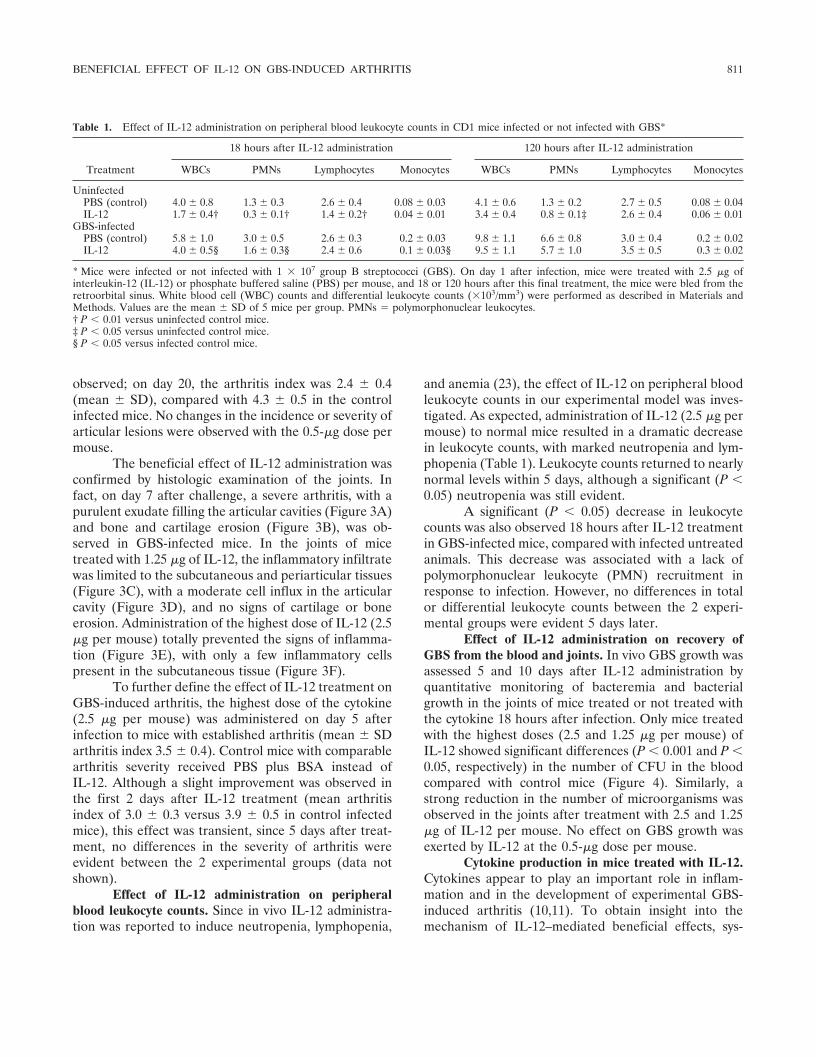
and anemia (23), the effect of IL-12 on peripheral bloodleukocyte counts in our experimental model was inves-tigated. As expected, administration of IL-12 (2.5 �g permouse) to normal mice resulted in a dramatic decreasein leukocyte counts, with marked neutropenia and lym-phopenia (Table 1). Leukocyte counts returned to nearlynormal levels within 5 days, although a significant (P �0.05) neutropenia was still evident.
A significant (P � 0.05) decrease in leukocytecounts was also observed 18 hours after IL-12 treatmentin GBS-infected mice, compared with infected untreatedanimals. This decrease was associated with a lack ofpolymorphonuclear leukocyte (PMN) recruitment inresponse to infection. However, no differences in totalor differential leukocyte counts between the 2 experi-mental groups were evident 5 days later.
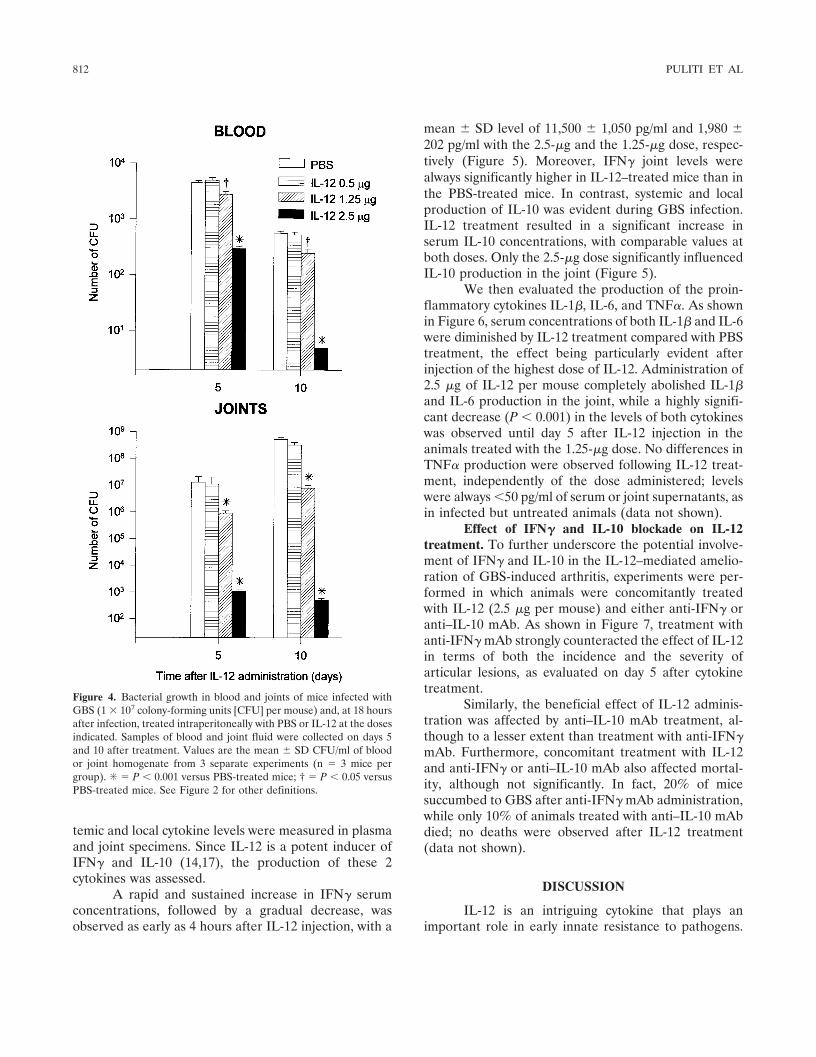
Effect of IL-12 administration on recovery ofGBS from the blood and joints. In vivo GBS growth wasassessed 5 and 10 days after IL-12 administration byquantitative monitoring of bacteremia and bacterialgrowth in the joints of mice treated or not treated withthe cytokine 18 hours after infection. Only mice treatedwith the highest doses (2.5 and 1.25 �g per mouse) ofIL-12 showed significant differences (P � 0.001 and P �0.05, respectively) in the number of CFU in the bloodcompared with control mice (Figure 4). Similarly, astrong reduction in the number of microorganisms wasobserved in the joints after treatment with 2.5 and 1.25�g of IL-12 per mouse. No effect on GBS growth wasexerted by IL-12 at the 0.5-�g dose per mouse.
Cytokine production in mice treated with IL-12.Cytokines appear to play an important role in inflam-mation and in the development of experimental GBS-induced arthritis (10,11). To obtain insight into themechanism of IL-12–mediated beneficial effects, sys-
Table 1. Effect of IL-12 administration on peripheral blood leukocyte counts in CD1 mice infected or not infected with GBS*
Treatment
18 hours after IL-12 administration 120 hours after IL-12 administration
WBCs PMNs Lymphocytes Monocytes WBCs PMNs Lymphocytes Monocytes
UninfectedPBS (control) 4.0 � 0.8 1.3 � 0.3 2.6 � 0.4 0.08 � 0.03 4.1 � 0.6 1.3 � 0.2 2.7 � 0.5 0.08 � 0.04IL-12 1.7 � 0.4† 0.3 � 0.1† 1.4 � 0.2† 0.04 � 0.01 3.4 � 0.4 0.8 � 0.1‡ 2.6 � 0.4 0.06 � 0.01
GBS-infectedPBS (control) 5.8 � 1.0 3.0 � 0.5 2.6 � 0.3 0.2 � 0.03 9.8 � 1.1 6.6 � 0.8 3.0 � 0.4 0.2 � 0.02IL-12 4.0 � 0.5§ 1.6 � 0.3§ 2.4 � 0.6 0.1 � 0.03§ 9.5 � 1.1 5.7 � 1.0 3.5 � 0.5 0.3 � 0.02
* Mice were infected or not infected with 1 � 107 group B streptococci (GBS). On day 1 after infection, mice were treated with 2.5 �g ofinterleukin-12 (IL-12) or phosphate buffered saline (PBS) per mouse, and 18 or 120 hours after this final treatment, the mice were bled from theretroorbital sinus. White blood cell (WBC) counts and differential leukocyte counts (�103/mm3) were performed as described in Materials andMethods. Values are the mean � SD of 5 mice per group. PMNs � polymorphonuclear leukocytes.† P � 0.01 versus uninfected control mice.‡ P � 0.05 versus uninfected control mice.§ P � 0.05 versus infected control mice.
BENEFICIAL EFFECT OF IL-12 ON GBS-INDUCED ARTHRITIS 811
temic and local cytokine levels were measured in plasmaand joint specimens. Since IL-12 is a potent inducer ofIFN� and IL-10 (14,17), the production of these 2cytokines was assessed.
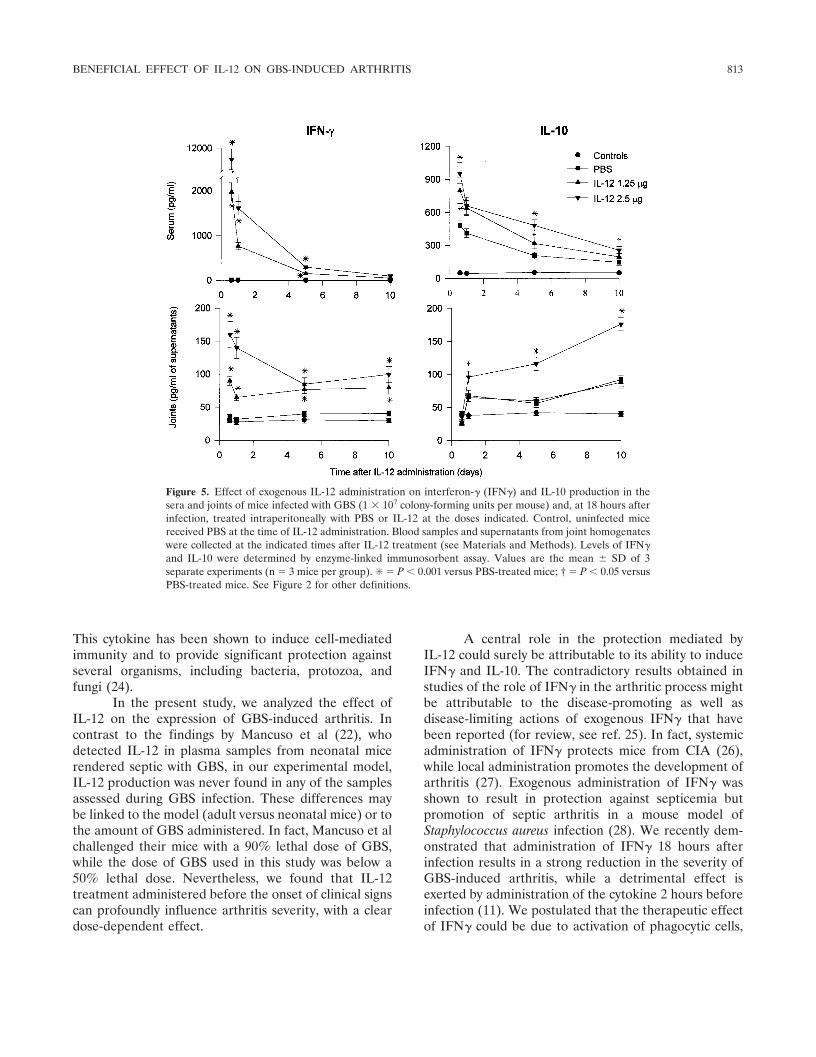
A rapid and sustained increase in IFN� serumconcentrations, followed by a gradual decrease, wasobserved as early as 4 hours after IL-12 injection, with a
mean � SD level of 11,500 � 1,050 pg/ml and 1,980 �202 pg/ml with the 2.5-�g and the 1.25-�g dose, respec-tively (Figure 5). Moreover, IFN� joint levels werealways significantly higher in IL-12–treated mice than inthe PBS-treated mice. In contrast, systemic and localproduction of IL-10 was evident during GBS infection.IL-12 treatment resulted in a significant increase inserum IL-10 concentrations, with comparable values atboth doses. Only the 2.5-�g dose significantly influencedIL-10 production in the joint (Figure 5).
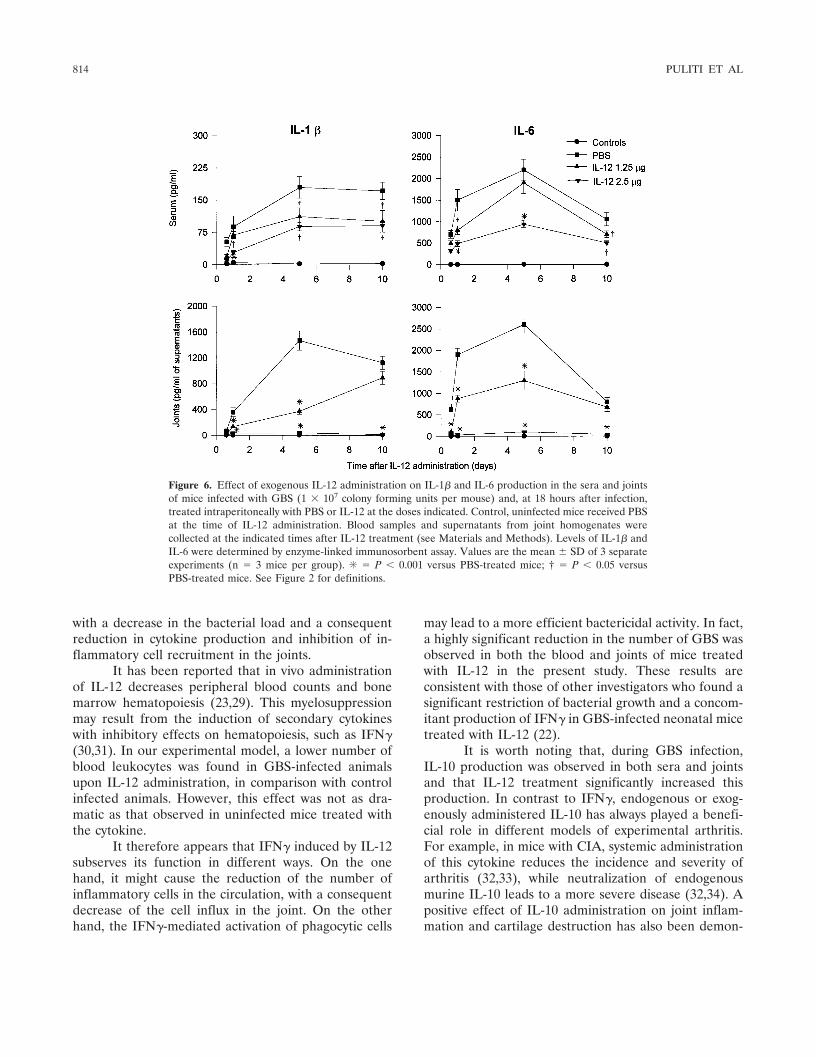
We then evaluated the production of the proin-flammatory cytokines IL-1�, IL-6, and TNF�. As shownin Figure 6, serum concentrations of both IL-1� and IL-6were diminished by IL-12 treatment compared with PBStreatment, the effect being particularly evident afterinjection of the highest dose of IL-12. Administration of2.5 �g of IL-12 per mouse completely abolished IL-1�and IL-6 production in the joint, while a highly signifi-cant decrease (P � 0.001) in the levels of both cytokineswas observed until day 5 after IL-12 injection in theanimals treated with the 1.25-�g dose. No differences inTNF� production were observed following IL-12 treat-ment, independently of the dose administered; levelswere always �50 pg/ml of serum or joint supernatants, asin infected but untreated animals (data not shown).
Effect of IFN� and IL-10 blockade on IL-12treatment. To further underscore the potential involve-ment of IFN� and IL-10 in the IL-12–mediated amelio-ration of GBS-induced arthritis, experiments were per-formed in which animals were concomitantly treatedwith IL-12 (2.5 �g per mouse) and either anti-IFN� oranti–IL-10 mAb. As shown in Figure 7, treatment withanti-IFN� mAb strongly counteracted the effect of IL-12in terms of both the incidence and the severity ofarticular lesions, as evaluated on day 5 after cytokinetreatment.
Similarly, the beneficial effect of IL-12 adminis-tration was affected by anti–IL-10 mAb treatment, al-though to a lesser extent than treatment with anti-IFN�mAb. Furthermore, concomitant treatment with IL-12and anti-IFN� or anti–IL-10 mAb also affected mortal-ity, although not significantly. In fact, 20% of micesuccumbed to GBS after anti-IFN� mAb administration,while only 10% of animals treated with anti–IL-10 mAbdied; no deaths were observed after IL-12 treatment(data not shown).
DISCUSSION
IL-12 is an intriguing cytokine that plays animportant role in early innate resistance to pathogens.
Figure 4. Bacterial growth in blood and joints of mice infected withGBS (1 � 107 colony-forming units [CFU] per mouse) and, at 18 hoursafter infection, treated intraperitoneally with PBS or IL-12 at the dosesindicated. Samples of blood and joint fluid were collected on days 5and 10 after treatment. Values are the mean � SD CFU/ml of bloodor joint homogenate from 3 separate experiments (n � 3 mice pergroup). ✳ � P � 0.001 versus PBS-treated mice; † � P � 0.05 versusPBS-treated mice. See Figure 2 for other definitions.
812 PULITI ET AL
This cytokine has been shown to induce cell-mediatedimmunity and to provide significant protection againstseveral organisms, including bacteria, protozoa, andfungi (24).
In the present study, we analyzed the effect ofIL-12 on the expression of GBS-induced arthritis. Incontrast to the findings by Mancuso et al (22), whodetected IL-12 in plasma samples from neonatal micerendered septic with GBS, in our experimental model,IL-12 production was never found in any of the samplesassessed during GBS infection. These differences maybe linked to the model (adult versus neonatal mice) or tothe amount of GBS administered. In fact, Mancuso et alchallenged their mice with a 90% lethal dose of GBS,while the dose of GBS used in this study was below a50% lethal dose. Nevertheless, we found that IL-12treatment administered before the onset of clinical signscan profoundly influence arthritis severity, with a cleardose-dependent effect.
A central role in the protection mediated byIL-12 could surely be attributable to its ability to induceIFN� and IL-10. The contradictory results obtained instudies of the role of IFN� in the arthritic process mightbe attributable to the disease-promoting as well asdisease-limiting actions of exogenous IFN� that havebeen reported (for review, see ref. 25). In fact, systemicadministration of IFN� protects mice from CIA (26),while local administration promotes the development ofarthritis (27). Exogenous administration of IFN� wasshown to result in protection against septicemia butpromotion of septic arthritis in a mouse model ofStaphylococcus aureus infection (28). We recently dem-onstrated that administration of IFN� 18 hours afterinfection results in a strong reduction in the severity ofGBS-induced arthritis, while a detrimental effect isexerted by administration of the cytokine 2 hours beforeinfection (11). We postulated that the therapeutic effectof IFN� could be due to activation of phagocytic cells,
Figure 5. Effect of exogenous IL-12 administration on interferon-� (IFN�) and IL-10 production in thesera and joints of mice infected with GBS (1 � 107 colony-forming units per mouse) and, at 18 hours afterinfection, treated intraperitoneally with PBS or IL-12 at the doses indicated. Control, uninfected micereceived PBS at the time of IL-12 administration. Blood samples and supernatants from joint homogenateswere collected at the indicated times after IL-12 treatment (see Materials and Methods). Levels of IFN�and IL-10 were determined by enzyme-linked immunosorbent assay. Values are the mean � SD of 3separate experiments (n � 3 mice per group). ✳ � P � 0.001 versus PBS-treated mice; † � P � 0.05 versusPBS-treated mice. See Figure 2 for other definitions.
BENEFICIAL EFFECT OF IL-12 ON GBS-INDUCED ARTHRITIS 813
with a decrease in the bacterial load and a consequentreduction in cytokine production and inhibition of in-flammatory cell recruitment in the joints.
It has been reported that in vivo administrationof IL-12 decreases peripheral blood counts and bonemarrow hematopoiesis (23,29). This myelosuppressionmay result from the induction of secondary cytokineswith inhibitory effects on hematopoiesis, such as IFN�(30,31). In our experimental model, a lower number ofblood leukocytes was found in GBS-infected animalsupon IL-12 administration, in comparison with controlinfected animals. However, this effect was not as dra-matic as that observed in uninfected mice treated withthe cytokine.
It therefore appears that IFN� induced by IL-12subserves its function in different ways. On the onehand, it might cause the reduction of the number ofinflammatory cells in the circulation, with a consequentdecrease of the cell influx in the joint. On the otherhand, the IFN�-mediated activation of phagocytic cells
may lead to a more efficient bactericidal activity. In fact,a highly significant reduction in the number of GBS wasobserved in both the blood and joints of mice treatedwith IL-12 in the present study. These results areconsistent with those of other investigators who found asignificant restriction of bacterial growth and a concom-itant production of IFN� in GBS-infected neonatal micetreated with IL-12 (22).
It is worth noting that, during GBS infection,IL-10 production was observed in both sera and jointsand that IL-12 treatment significantly increased thisproduction. In contrast to IFN�, endogenous or exog-enously administered IL-10 has always played a benefi-cial role in different models of experimental arthritis.For example, in mice with CIA, systemic administrationof this cytokine reduces the incidence and severity ofarthritis (32,33), while neutralization of endogenousmurine IL-10 leads to a more severe disease (32,34). Apositive effect of IL-10 administration on joint inflam-mation and cartilage destruction has also been demon-
Figure 6. Effect of exogenous IL-12 administration on IL-1� and IL-6 production in the sera and jointsof mice infected with GBS (1 � 107 colony forming units per mouse) and, at 18 hours after infection,treated intraperitoneally with PBS or IL-12 at the doses indicated. Control, uninfected mice received PBSat the time of IL-12 administration. Blood samples and supernatants from joint homogenates werecollected at the indicated times after IL-12 treatment (see Materials and Methods). Levels of IL-1� andIL-6 were determined by enzyme-linked immunosorbent assay. Values are the mean � SD of 3 separateexperiments (n � 3 mice per group). ✳ � P � 0.001 versus PBS-treated mice; † � P � 0.05 versusPBS-treated mice. See Figure 2 for definitions.
814 PULITI ET AL

strated in murine streptococcal cell wall–induced arthri-tis (35). Finally, IL-10 was found to modulate Borreliaburgdorferi–induced inflammation in vivo, sinceC57BL/6J mice deficient in IL-10 develop more severearthritis than wild-type C57BL/6J mice (36).
One of the actions of IL-10 is to down-regulate
many inflammatory mediators (37–39), including cyto-kines, whose participation in the pathogenesis of arthri-tis has been well documented (10,40–44). Here, theantiinflammatory properties of IL-10 seemed to play asignificant role in the amelioration of the arthritis due toIL-12 treatment, at least when this cytokine was admin-istered at the 2.5-�g dose per mouse. In fact, in this case,a complete abolition of IL-6 and IL-1� in the joint wasobserved.
Experiments performed with neutralizing anti-bodies further emphasize the involvement of IFN� andIL-10 in the IL-12–mediated beneficial effects on arthri-tis. Although both anti-IFN� and anti–IL-10 mAb re-versed the amelioration of GBS-induced arthritis, itappears that a major role in the protective effect ofIL-12 was played by IFN�, as assessed by the differencesin the arthritis index.
Cellular sources of IFN� and IL-10 producedafter IL-12 treatment are presently under investigation.IL-12 is known to induce IFN� secretion in different cellpopulations, including PMN (45), macrophages (46),natural killer cells (16), and dendritic cells (47). It hasbeen reported that treatment with IL-12 increases theproduction of IL-10 by PMN (48). Moreover, synovialcells may contribute to the production of IL-10, sincesynovial fibroblast-like cells are able to secrete IL-10(49), and up-regulation of messenger RNA levels ofIL-10 was observed in synovial tissues upon IL-12 ad-ministration in a mouse model of CIA (17). Thus, it isconceivable that all these cell populations, which areknown to be involved in joint inflammation (50–52), areresponsible for the IFN� and IL-10 levels observed soonafter IL-12 injection in GBS-infected animals.
One interesting observation of the present studyis the different kinetics of the appearance of IFN� andIL-10 in the joints. Indeed, while IFN� was producedearly and then subsequently decreased, the productionof IL-10 continuously increased during the observationperiod. It is likely that the temporal production of IFN�and IL-10 may also contribute to their effects in micewith GBS-induced arthritis.
In conclusion, we have demonstrated the benefi-cial role of IL-12 in GBS-induced arthritis at the desig-nated doses. The appearance and severity of GBS-induced arthritis are the byproducts of a multifactorialprocess involving the number of microorganisms andinflammatory cells that reach the joints and the produc-tion of proinflammatory cytokines; both factors areaffected by IL-12 administration through induction ofIFN� and IL-10. The finding that a single injection of ahigh dose of IL-12 in mice with established arthritis
Figure 7. Effect of neutralizing anticytokine antibodies on A, theincidence and B, the severity of arthritis in CD1 mice infectedintravenously with GBS (1 � 107 colony-forming units per mouse), andat 18 hours after infection, treated intraperitoneally with PBS or IL-12(2.5 �g per mouse). Anti–interferon-� (anti-IFN�) or anti–IL-10monoclonal antibodies (mAb; 1 mg per mouse) were administeredintraperitoneally at the same time as the PBS or IL-12. A, Incidence ofGBS-induced arthritis (percentage of mice with visible arthritis). B,Arthritis index (clinical severity of arthritis, evaluated as described inMaterials and Methods). The incidence and severity of arthritis wereevaluated on day 5 after IL-12 plus mAb administration. Values arethe mean � SD of 2 separate experiments (n � 10 mice per group).✳ � P � 0.001 versus PBS-treated mice; † � P � 0.05 versusPBS-treated mice; ‡ � P � 0.05 versus anti–IL-10 mAb–treated mice.See Figure 2 for other definitions.
BENEFICIAL EFFECT OF IL-12 ON GBS-INDUCED ARTHRITIS 815

results in a slight, although transient amelioration ofarticular lesions establishes the need for further investi-gation with prolonged treatment to evaluate the efficacyof IL-12 in the therapy of GBS-induced arthritis.
ACKNOWLEDGMENTS
The authors wish to thank Prof. Luigina Romani forcritical reading and revision of the manuscript and Mrs. EileenMahoney Zannetti for dedicated editorial assistance.
REFERENCES
1. Baker CJ, Edwards MS. Group B streptococcal infections. In:Remington JS, Klein JO, editors. Infectious diseases of the fetusand newborn infant. 4th ed. Philadelphia: WB Saunders; 1995. p.980–1054.
2. Cooper BW, Morganelli E. Group B streptococcal bacteremia inadults at Hartford Hospital 1991-1996. Conn Med 1998;62:515–7.
3. Gardam MA, Low DE, Saginur R, Miller MA. Group B strepto-coccal necrotizing fasciitis and streptococcal toxic shock-like syn-drome in adults. Arch Intern Med 1998;158:1704–8.
4. Straus C, Caplanne D, Bergemer AM, le Parc JM. Destructivepolyarthritis due to a group B streptococcus. Rev Rhum Engl Ed1997;64:339–41.
5. Schattner A, Vosti K. Recurrent group B streptococcal arthritis.Clin Rheumatol 1998;17:387–9.
6. Jackson LA, Hilsdon R, Farley MM, Harrison LH, Reingold AL,Plikaytis BD, et al. Risk factors for group B streptococcal diseasein adults. Ann Intern Med 1995;123:415–20.
7. Schattner A, Vosti KL. Bacterial arthritis due to �-hemolyticstreptococci of serogroups A, B, C, F, and G: analysis of 23 casesand a review of the literature. Medicine (Baltimore) 1998;77:122–39.
8. Tissi L, Marconi P, Mosci P, Merletti L, Cornacchione P, Rosati E,et al. Experimental model of type IV Streptococcus agalactiae(group B Streptococcus) infection in mice with early developmentof septic arthritis. Infect Immun 1990;58:3093–100.
9. Tissi L, von Hunolstein C, Bistoni F, Marangi M, Parisi L, OreficiG. Role of group B streptococcal capsular polysaccharides in theinduction of septic arthritis. J Med Microbiol 1998;47:717–23.
10. Tissi L, Puliti M, Barluzzi R, Orefici G, von Hunolstein C, BistoniF. Role of tumor necrosis factor �, interleukin-1� and interleu-kin-6 in a mouse model of group B streptococcal arthritis. InfectImmun 1999;67:4545–50.
11. Puliti M, von Hunolstein C, Bistoni F, Mosci P, Orefici G, Tissi L.Influence of interferon-� administration on the severity of exper-imental group B streptococcal arthritis. Arthritis Rheum 2000;43:2678–86.
12. D’Andrea A, Rengaraju M, Valiante NM, Chehimi J, Kubin M,Aste M, et al. Production of natural killer cell stimulatory factor(interleukin-12) by peripheral blood mononuclear cells. J Exp Med1992;176:1387–98.
13. Kobayashi M, Fitz L, Ryan M, Hewick RM, Clark SC, Chan SH,et al. Identification and purification of natural killer cell stimula-tory factor (NKSF), a cytokine with multiple biologic effects onhuman lymphocytes. J Exp Med 1989;170:827–45.
14. Wysocka M, Kubin M, Vieira LQ, Ozmen L, Carrotta G, Scott P,et al. Interleukin-12 is required for interferon-� production andlethality in lipopolysaccharide-induced shock in mice. Eur J Im-munol 1995;25:672–6.
15. Trinchieri G. Interleukin-12 and its role in the generation of Th1cells. Immunol Today 1993;14:335–8.
16. Trinchieri G. IL-12: a proinflammatory cytokine with immuno-
regulatory functions that bridges innate resistance and antigenspecific adaptive immunity. Annu Rev Immunol 1995;13:251–76.
17. Joosten LAB, Lubberts E, Helsen MMA, van den Berg WB. Dualrole of IL-12 in early and late stages of murine collagen type IIarthritis. J Immunol 1997;159:4094–102.
18. Kasama T, Yamazachi J, Hanaoka R, Miwa Y, Hatano Y,Kobayashi K, et al. Biphasic regulation of the development ofmurine type II collagen–induced arthritis by interleukin-12: possi-ble involvement of endogenous interleukin-10 and tumor necrosisfactor �. Arthritis Rheum 1999;42:100–9.
19. Malfait AM, Butler DM, Presky DH, Maini RN, Brennan FM,Feldmann M. Blockade of IL-12 during the induction of collagen-induced arthritis (CIA) markedly attenuates the severity of thearthritis. Clin Exp Immunol 1998;111:377–83.
20. Anguita J, Persing DH, Rincon H, Barthold SW, Fikrig E. Effectof anti-interleukin 12 treatment on murine Lyme borreliosis. J ClinInvest 1996;97:1028–34.
21. Joosten LAB, Helsen MMA, van den Berg WB. Blockade ofendogenous interleukin 12 results in suppression of murine strep-tococcal cell wall arthritis by enhancement of interleukin 10 andinterleukin 1 Ra. Ann Rheum Dis 2000;59:196–205.
22. Mancuso G, Cusumano V, Genovese F, Gambuzza M, Beninati C,Teti G. Role of interleukin 12 in experimental neonatal sepsiscaused by group B streptococci. Infect Immun 1997;65:3731–5.
23. Brunda MJ. Interleukin 12. J Leukoc Biol 1994;55:280–8.24. Romani L, Puccetti P, Bistoni F. Interleukin-12 in infectious
diseases. Clin Microbiol Rev 1997;10:611–36.25. Billiau A. Interferon-�: biology and role in pathogenesis. Adv
Immunol 1996; 62:61–130.26. Mauritz NJ, Holmdahl R, Jonsson R, van der Meide PH, Schey-
nius A, Klareskog L. Treatment with �-interferon triggers theonset of collagen arthritis in mice. Arthritis Rheum 1988;31:1297–304.
27. Nakajiama H, Takamori H, Hiyama Y, Tsukuda W. The effect oftreatment with interferon-� on type II collagen-induced arthritisfollowing treatment with anti-interferon-�. Clin Exp Immunol1990;81:441–5.
28. Zhao Y-X, Nilsson I-M, Tarkowski A. The dual role of interfer-on-� in experimental Staphylococcus aureus septicaemia versusarthritis. Immunology 1998;93:80–5.
29. Tare NS, Bowen S, Warrier RR, Carvajal DM, Benjamin WR,Riley JH, et al. Administration of recombinant interleukin-12 tomice suppresses hematopoiesis in the bone marrow but enhanceshematopoiesis in the spleen. J Interferon Cytokine Res 1995;15:377–83.
30. Bellone G, Trinchieri G. Dual stimulatory and inhibitory effect ofNK cell stimulatory factor/IL-12 on human hematopoiesis. J Im-munol 1994;153:930–7.
31. Eng VM, Car BD, Schnyder B, Lorenz M, Lugli S, Aguet M, et al.The stimulatory effects of interleukin (IL)-12 on hematopoiesisare antagonized by IL-12-induced interferon � in vivo. J Exp Med1995;181:1893–8.
32. Joosten LAB, Lubberts E, Durez P, Helsen MMA, Jacobs MJM,Goldman M, et al. Role of interleukin-4 and interleukin-10 inmurine collagen-induced arthritis: protective effect of interleu-kin-1� and interleukin-10 treatment on cartilage destruction.Arthritis Rheum 1997;40:249–60.
33. Walmsley M, Katsikis PD, Abney E, Parry S, Williams RO, MainiRN, et al. Interleukin-10 inhibition of the progression of estab-lished collagen-induced arthritis. Arthritis Rheum 1996;39:495–503.
34. Kasama T, Strieter RM, Lukacs NW, Lincoln PM, Burdick MD,Kunkel SL. Interleukin-10 expression and chemokine regulationduring the evolution of the murine type II collagen-inducedarthritis. J Clin Invest 1995;95:2868–76.
35. Lubberts E, Joosten LAB, Helsen MMA, van den Berg WB.Regulatory role of interleukin 10 in joint inflammation and
816 PULITI ET AL

cartilage destruction in murine streptococcal cell wall (SCW)arthritis: more therapeutic benefit with IL-4/IL-10 combinationtherapy than with IL-10 treatment alone. Cytokine 1998;10:361–9.
36. Brown JP, Zachary JF, Teuscher C, Weis JJ, Wooten RM. Dualrole of interleukin-10 in murine Lyme disease: regulation ofarthritis severity and host defense. Infect Immun 1999;67:5142–50.
37. Moore KW, O’Garra A, de Waal Malefyt R, Vieira P, MosmannTR. Interleukin-10. Annu Rev Immunol 1993;11:165–90.
38. Bogdan C, Vodovotz Y, Nathan C. Macrophage deactivation byinterleukin 10. J Exp Med 1991;174:1549–55.
39. Bessis N, Chiocchia G, Kollies G, Mirty A, Fournier C, FradeliziD, et al. Modulation of proinflammatory cytokine production intumor necrosis factor-� (TNF-�)-transgenic mice by treatmentwith cells engineered to secrete IL-4, IL-10 or IL-13. Clin ExpImmunol 1998;111:391–6.
40. Arend WP, Dayer J-M. Cytokine and cytokine inhibitors orantagonists in rheumatoid arthritis. Arthritis Rheum 1990;33:305–15.
41. Ridderstad A, Abedi-Valugerdi M, Moller E. Cytokines in rheu-matoid arthritis. Ann Med 1991;23:219–23.
42. Bremmel T, Abdelnour A, Tarkowski A. Histopathological andserological progression of experimental Staphylococcus aureusarthritis. Infect Immun 1992;60:2976–85.
43. Nietfeld JJ, Wilbrink B, Helle M, van Roy JLAM, Den Otter W,Swaak AJG, et al. Interleukin-1–induced interleukin-6 is requiredfor the inhibition of proteoglycan synthesis by interleukin-1 inhuman articular cartilage. Arthritis Rheum 1990;33:1695–701.
44. Green J, Schotland S, Sella Z, Kleeman CR. Interleukin-6 atten-
uates agonist-mediated calcium mobilization in murine osteoblas-tic cells. J Clin Invest 1994;93:2340–50.
45. Yeaman RG, Collins JE, Currie JK, Guyre PM, Wira CR, FangerMW. IFN-� is produced by polymorphonuclear neutrophils inhuman uterine endometrium and by cultured peripheral bloodpolymorphonuclear neutrophils. J Immunol 1998;160:5145–53.
46. Puddu P, Fantuzzi L, Borghi P, Varano B, Rainaldi G, Guiller-mard E, et al. IL-12 induces IFN-� expression and secretion inmouse peritoneal macrophages. J Immunol 1997;159:3490–7.
47. Stober D, Schirmbeck R, Reimann J. IL-12/IL-18 dependentIFN-� release by murine dendritic cells. J Immunol 2001;167:957–65.
48. Romani L, Mencacci A, Cenci E, Spaccapelo R, Del Sero G,Nicoletti I, et al. Neutrophil production of IL-12 and IL-10 incandidiasis and efficacy of IL-12 therapy in neutropenic mice.J Immunol 1997;158:5349–56.
49. Ritchlin C, Haas-Smith SA. Expression of interleukin 10 mRNAand protein by synovial fibroblastoid cells. J Rheumatol 2001;28:698–705.
50. Bresnihan B. Pathogenesis of joint damage in rheumatoid arthritis.J Rheumatol 1999;26:717–9.
51. Holmdahl R, Tarkowski A, Jonsson R. Involvement of macro-phages and dendritic cells in synovial inflammation of collageninduced arthritis in DBA/1 mice and spontaneous arthritis inMRL/lpr mice. Autoimmunity 1991;8:271–80.
52. Verdrengh M, Tarkowski A. Role of neutrophils in experimentalsepticemia and septic arthritis induced by Staphylococcus aureus.Infect Immun 1997;65;2517–21.
BENEFICIAL EFFECT OF IL-12 ON GBS-INDUCED ARTHRITIS 817
Copyright © 2022 FDOKUMEN




















