
Morphometrical and acoustical comparison between diploid and tetraploid green toads
Upload
independentCategory
view
2download
0
Plant Cell RepDOI 10.1007/s00299-006-0273-8
GENETIC TRANSFORMATION AND HYBRIDIZATION
The usefulness of the gfp reporter gene for monitoringAgrobacterium-mediated transformation of potato dihaploid andtetraploid genotypesElena Rakosy-Tican · Cristian M. Aurori ·Camelia Dijkstra · Ramona Thieme · Adriana Aurori ·Michael R. Davey
Received: 7 August 2006 / Revised: 6 November 2006 / Accepted: 16 November 2006C© Springer-Verlag 2006
Abstract Potato is one of the main targets for genetic im-provement by gene transfer. The aim of the present study wasto establish a robust protocol for the genetic transformationof three dihaploid and four economically important cultivarsof potato using Agrobacterium tumefaciens carrying the invivo screenable reporter gene for green fluorescent protein(gfp) and the marker gene for neomycin phosphotransferase(nptII). Stem and leaf explants were used for transforma-tion by Agrobacterium tumefaciens strain LBA4404 carry-ing the binary vector pHB2892. Kanamycin selection, visualscreening of GFP by epifluorescent microscopy, PCR am-plification of nptII and gfp genes, as well as RT-PCR andSouthern blotting of gfp and Northern blotting of nptII, wereused for transgenic plant selection, identification and analy-sis. Genetic transformation was optimized for the best per-forming genotypes with a mean number of shoots expressinggfp per explant of 13 and 2 (dihaploid line 178/10 and cv.‘Baltica’, respectively). The nptII marker and gfp reportergenes permitted selection and excellent visual screening oftransgenic tissues and plants. They also revealed the effectsof antibiotic selection on organogenesis and transformation
Communicated by L. Pena
E. Rakosy-Tican () · C. M. Aurori · A. AuroriBabes-Bolyai University, Plant Genetic Engineering Group,400006 Cluj-Napoca, Romaniae-mail: [email protected], [email protected]
C. Dijkstra · M. R. DaveySchool of Biosciences, University of Nottingham,Sutton Bonington Campus,Loughborough, LE12 5RD UK
R. ThiemeFederal Centre for Breeding Research on Cultivated Plants,Institute of Agricultural Crops,Groß Lusewitz, Germany
frequency, and the identification of escapes and chimerasin all potato genotypes. Silencing of the gfp transgene thatmay represent site-specific inactivation during cell differ-entiation, occurred in some transgenic shoots of tetraploidcultivars and in specific chimeric clones of the dihaploid line178/10. The regeneration of escapes could be attributed to ei-ther the protection of non-transformed cells by neighbouringtransgenic cells, or the persistence of Agrobacterium cells inplant tissues after co-cultivation.
Keywords Agrobacterium tumefaciens-mediatedtransformation . Chimeras . Green fluorescent proteinreporter gene . Neomycin phophotransferase marker gene .
Transgene silencing
Abbreviations GA: Gibberellic acid . gfp: Greenfluorescent protein gene . GFP: Green fluorescent protein .
gus: β-Glucuronidase gene . GUS: β-Glucuronidase .
luc: Luciferase gene . MS: Murashige and Skoog (1962) .
NAA: Naphthaleneacetic acid . nptII: Neomycinphosphotransferase gene . PCR: Polymerase chain reaction .
RT-PCR reverse: Transcriptase-polymerase chain reaction
Introduction
Potato, as the fourth most important crop in the world and afood security crop, is an important target for genetic improve-ment through classical and biotechnological approaches. Invitro culture and recombinant DNA technologies are avail-able to complement conventional breeding, seed multipli-cation and disease control. Although potato was one of thefirst crops to be transformed by Agrobacterium tumefaciens(Stiekema et al. 1988; Sheerman and Bevan 1988) and trans-genic plants have been generated in several potato cultivars
Springer

Plant Cell Rep
(Heeres et al. 2002), the efficiency of plant regeneration isdependent on genotype. Novel gene sequencing, cloning andinsertion, as well as functional genomics, are tools that willbenefit the improvement of economically important potatocvs. and dihaploid genotypes through reliable transforma-tion protocols. Moreover, refinement of transformation bya visible reporter gene, such as gfp, should prove useful intransferring genes of interest without the need for selectablemarkers, such as genes for antibiotic resistance, since thelatter raise public concerns, particularly in Europe (Halford2004).
Reporter genes that allow selection and visual detectionof transgene expression, such as gus and luc, have been usedextensively to monitor plant transformation. The most com-monly exploited gus gene with its product GUS (Jeffersonet al. 1987), has a crucial drawback because the histochem-ical assay is destructive for plant tissue and, consequently,cannot be exploited for analysis in vivo of transgene expres-sion. The luc gene that can be monitored in vivo requires anexogenous substrate, luciferin, and sophisticated equipment(Ow et al. 1986). In contrast, the gfp gene from the jelly-fish Aequorea victoria (Chalfie et al. 1994) does not needan exogenous substrate, can be monitored in vivo withoutan additional cofactor, and its expression is visible underfluorescence microscopy or even laboratory fluorescent illu-mination (Molinier et al. 2000). GFP expression is visiblein whole plants, permitting screening of primary transfor-mants and the discrimination of homozygous and hemizy-gous plants (Molinier et al. 2000; Ghorbel et al. 1999; Zhanget al. 2001). GFP has become useful for studying promoterexpression, monitoring of protein trafficking, cell compart-mentalisation, virus infection and virus-induced gene silenc-ing (Rizzuto et al. 1995; Kohler 1998; Ruiz et al. 1998),as well as monitoring transformation (Haselof et al. 1997;Rouwendal et al. 1997; Elliott et al. 1999; Monilier et al.2000; Kaeppler et al. 2001). It has been used as a marker forsomatic hybridization in Citrus (Olivares-Fuster 2002) andto produce green fluorescent flowers (Mercuri et al. 2001).However, in potato, gfp has been used only in stable chloro-plast transformation (Sidorov et al. 1999).
In this report, gfp and the selectable marker gene nptII,were used to monitor Agrobacterium tumefaciens-mediatedtransformation of dihaploid genotypes and economically im-portant tetraploid cvs. of potato. Gfp expression was re-liable as it allowed visual screening of expression fromcallus to whole plants, with in vivo discrimination of es-capes and chimeras, as well as genotype-dependent effectsof kanamycin on organogenesis. All genotypes were trans-formed, but leaf explants of the dihaploid line 178/10 andstem internode explants of the cv. ‘Baltica’ exhibited thegreatest transformation frequencies with mean values of13 and 2 transgenic shoots per initial explant, respectively.The utility of gfp, the limitations of kanamycin selection
and the possible causes of escapes and chimeras, are alsodiscussed.
Material and methods
Plant material and transformation
Leaf and stem explants of the dihaploid genotypes 178/10,224/1 and 227/5 were excised from in vitro propagated plants4 weeks after transfer of the stock plants to RMB5 medium(Menzel et al. 1981). Dihaploids were produced by partheno-genesis after crossing of the tetraploid cvs. ‘Amsel’, ‘Nicola’and ‘Sante’ with Solanum phureja at the Institute for PotatoResearch and Production, Brasov, Romania. The tetraploidcvs. ‘Desiree’ (used as reference), ‘Agave’ and ‘Delikat’(Norika, Gross Lusewitz, Germany) and ‘Baltica’ (Saka-Ragis-Pflanzenzucht, Saka-Ragis, Germany), were propa-gated in vitro on semi-solid MS-based medium enriched with1.15 g L−1 NH4NO3 (designed MS.5). Generally, internodalexplants from 4-week-old in vitro grown plants of tetraploidcvs. were inoculated with A. tumefaciens LBA4404 harbor-ing pHB2892 (provided by G. Hahne and J. Molinier) fortransformation (Fig. 1). Plasmid HB2892 contained the s-gfp gene, under the control of a double CaMV 35S promoter,adjacent to the left T-DNA border and the nos-nptII-nos se-lectable marker gene next to the right T-DNA border (Molin-ier et al. 2000). The maintenance of bacteria, co-cultivationof explants with bacteria and culture of stem and leaf ex-plants, were as described by Kumar (1995). Since LSR1 fol-lowed by LSR2 culture media (Kumar, 1995) induced limitedshoot regeneration from tetraploid cvs., MS-based mediumwith 16 g L−1 glucose, 0.5 mg L−1 folic acid, 0.05 mg L−1
biotin, 40 mg L−1 adenine, 0.02 mg L−1 GA3, 0.02 mg L−1
NAA, 2.0 mg L−1 zeatin riboside, and 0.75% (w/v) agarat pH 5.8 (designed MS-t medium) was assessed. Agrobac-terium growth was controlled by addition of 250 mg L−1
cefotaxime (Claforan; Hoechst-Roussel Pharmaceuticals,Frankfurt, Germany) to the culture medium with or with-out 50 mg L−1 kanamycin (Sigma–Aldrich, Poole, UK).Comparative experiments were undertaken with permanentkanamycin selection or with the elimination of kanamycinfrom the medium after 5 weeks of culture. Uninoculated ex-plants were cultured in the same way as explants inoculatedwith Agrobacterium. Cultures were maintained at 22 ± 2Cwith a 16 h photoperiod (90 µmol m−2 s−1, Daylight fluo-rescent illumination).
Transgenic tissue, plant selection and analysis
Putative transgenic callus and shoots were selected onkanamycin-containing medium followed by visualising gfpexpression by fluorescence microscopy (BX-60 microscope
Springer

Plant Cell Rep
BamH
BamHI
Nos nptII LB Lac Z
MCS CaMV Enhancer....CaMV Enhancer Ω
Nos RB
SGFP
MCS CaMV term
NcoI NcoI BamHIFig. 1 Structure of the T-DNAof pHB2892 with gfp and nptIIgenes (Molinier et al. 2000)
with wide band cube unit U-MWB; Olympus GmbH, Ham-burg, Germany) to discriminate the green fluorescence ofgfp from the native red fluorescence of chlorophyll. Eachregenerated shoot exhibiting green fluorescence was consid-ered putatively transgenic; shoots showing red chlorophyllautofluorescence were designated escapes. Shoots with dualgreen and red fluorescence in different tissues were chimeras.Three replicates of each experiment were set up, each with atleast 10–20 individual explants. Control and putative trans-genic shoots were analysed for integration of the nptII geneby PCR amplification and Northern blotting, and of gfp byPCR, RT-PCR and Southern blotting.
PCR amplification
Primers were manufactured and sequenced by MWGBiotech, Ebersberg, Germany.
The sequences used for nptII were 5′-AGA CAA TCGGCT GCT CTG AT-3′ and 5′-ATA CTT TCT CGG CAGGAG CA-3′ and for gfp 5′-AGG GCG ATG CCA CCT A-3′
and 5′-GAC TGG GTG CTC AGG TA-3′. Template ge-nomic DNA was extracted using a GenElute Plant Miniprepkit (Sigma–Aldrich). PCR was performed using RED TaqReady Mix (Sigma) according to the manufacturer’s instruc-tions. Amplification was performed in a DNA Thermal Cy-cler 480 (Perkin Elmer Applied Biosystems Division, War-rington, UK), with initial denaturing (1 cycle, 94C, 3 min),denaturating (35 cycles, 94C, 1 min), primer annealing [35cycles, 55C (nptII) or 53C (gfp), 1 min], primer exten-sion (35 cycles, 72C, 90 s), final extension (1 cycle, 72C,10 min) and holding at 4C (5 min to ∞ ).
RT-PCR analysis
RNA was extracted from approx. 100 mg leaf samples ofputatively transformed and non-transformed plants using anRNeasy r© Plant Mini Kit (Qiagen, Crawley, UK). RT-PCR
was peformed using a One Step RT-PCR Kit (Qiagen) ac-cording to the manufacturer’s instructions. The amplificationprogramme involved reverse transcription (1 cycle, 50C,30 min), polymerase activation (1 cycle, 94C, 15 min), de-naturation (35 cycles, 94C, 1 min) and primer annealing(35 cycles, 55C, 1 min). Subsequent conditions were as forPCR analysis.
NptII gene expression studies using Northern blot analysis
Total RNA (15 µg) was isolated from leaves using anRNeasy r© Plant Mini Kit (Qiagen). RNA was capillary blot-ted onto a nylon membrane (Roche Diagnostics Ltd., Lewes,UK). The membrane was prehybridised in DIG-Easy-Hybbuffer (Roche) at 68C for 1 h. For hybridisation, 10 mLof DIG-Easy-Hyb, to which had been added DIG-labelledRNA probes (100 ng mL−1; Roche), were used per 100 cm2
of membrane. Hybridisation was at 68C for 16 h. The mem-brane was washed for 2 × 5 min in 2 × SSC/0.1% SDS andfor 2 × 15 min in 0.1 × SSC/0.1% SDS at 68C. Themembrane was preincubated for 45 min in blocking solution(Roche) and incubated for 30 min with anti-DIG-AP con-jugate (diluted 1:10,000, v:v). Two 15 min washing stepspreceeded equilibration in detection buffer for 5 min. Detec-tion involved CDPStar (Roche) with a 20 min exposure toX-ray film (Kodak).
Southern blotting analysis for the gfp gene
Genomic DNA (10 µg) was isolated from leaves using aGenElute Plant Genomic DNA kit (Sigma–Aldrich). TheDNA was digested with EcoRI (Promega) according tothe manufacturer’s instructions. Digested DNA was capil-lary blotted onto nylon membrane (Roche Diagnostics Ltd.,Lewes, UK) and prehybridised in DIG-Easy-Hyb buffer(Roche) at 37C for 2 h. For hybridisation, 30 mL of DIG-Easy-Hyb, to which had been added DIG-labelled DNA
Springer

Plant Cell Rep
probes (100 ng mL−1; Roche), were used at 68C for 16 h.The membrane was washed twice (5 min each) in 2 ×SSC/0.1% SDS and twice (15 min each) in 0.1 × SSC/0.1%SDS at 65C. The membrane was preincubated for 45 minin blocking solution (Roche) and incubated for 30 min withanti-DIG-AP conjugate (diluted 1:10,000, v:v). Two 15 minwashing steps preceded equilibration in detection buffer for5 min. Detection involved CDPStar (Roche) with a 5 h ex-posure to X-ray film (Kodak).
Results
Although transformation efficiency was dependent on plantgenotype, all genotypes were transformed and regeneratedshoots expressed constitutively gfp fluorescence. The dihap-loids regenerated well from both leaf and stem explants onLSR1/LSR2 media, as recommended by Kumar (1995) forthe cv. ‘Desiree’, a model system in potato transformation.Leaf explants of tetraploid genotypes tended to be overgrownby Agrobacterium and were unable to form callus or shoots.Optimum regeneration was obtained for the four tetraploidcvs. when stem explants were cultured on MS-t medium,as these explants were less affected by Agrobacterium over-growth.
The dihaploid genotypes 178/10 and 224/1 regeneratedthe most shoots from leaf explants when kanamycin waseliminated from the medium after 5 weeks of culture (Fig.2A). Interestingly, the dihaploid genotype 227/5, which didnot regenerate shoots from uninoculated explants, did regen-erate a limited number of gfp-expressing shoots followingAgrobacterium inoculation and kanamycin selection (Fig.2A). Moreover, the percentage of gfp-expressing shoots wasdependent on genotype, but was greater with kanamycin se-lection for the majority of dihaploids (Fig. 2B). A relativelyhigh percentage of shoots not expressing gfp, ranging from30 to 100%, were also regenerated even under permanentkanamycin selection. The dihaploid 178/10, known to be themost responsive dihaploid genotype in vitro, also regener-ated the most shoots after Agrobacterium inoculation. Leafexplants were preferable for the dihaploid genotype 178/10
Table 1 Comparison of shoot regeneration from stem internodes oftetraploid potato cultivars, cultured on LSR1 followed by LSR2 media,or on MS-t medium
Potato tetraploid cv. Shoots per explantLSR1 followed by LSR2a MS-t medium
Agave 0 4 ± 0.5Baltica 16 ± 2.0 38 ± 4.0Delikat 0 5 ± 0.8Desiree 2 ± 0.5 5 ± 0.7
aKumar (1995). Data are means ± SEM; n = 30.
0
5
10
15
20
25
30
35
40
45
C A1 A2 C A1 A2
stem leaf
178/10
224/1
227/5
Sh
oo
ts p
er e
xpla
nt
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
S-224A1
S-227A1
S-227A2
L-178A1
L-224A1
L-224A2
L-227A1
L-227A2
gfp-
gfp+/-
gfp+
A
B
Fig. 2 Mean number of potato shoots regenerated on leaf or stemexplants A and the percentage of shoots regenerated on leaf (L) orstem (S) internodal explants B, after co-cultivation of explants withA. tumefaciens LBA4404 pHB2892. Dihaploid potato lines expressinggreen fluorescence (gfp + ), not expressing fluorescence (gfp − ) orchimeric for this trait (gfp + / − ), were assessed after 3 months ofculture on LSR medium with 250 mg L−1 cefotaxime. C = control;A1, selection on 50 mg L−1 kanamycin for the first 5 weeks of culture;A2, permanent selection on 50 mg L−1 kanamycin. (Bars = SEM)
with a mean number of regenerated shoots of 26 from uninoc-ulated explants and 34.5 per explant after Agrobacterium co-cultivation. Of the latter, 38% expressed gfp and 15% werechimeras when kanamycin was removed after 5 weeks ofculture (Fig. 2A and B).
Tetraploid cultivars also exhibited genotype-dependentregeneration, as well as optimum transformation efficiencyon MS-t medium (Fig. 3). Previous experiments showed thatmore shoots were regenerated for all tetraploid cvs. on MS-tmedium than on LSR1 medium followed by LSR2 medium(Table 1), although the latter sequence was recommendedby Kumar (1995) for the cv. ‘Desiree’. In all experiments,uninoculated (control) explants were unable to regenerateshoots on selection medium containing kanamycin. Prelim-inary experiments with A. tumefaciens EHA105 pGPTV re-vealed a tendency of Agrobacterium to over-grow leaf ex-plants (data not shown). Consequently, only stem internodal
Springer

Plant Cell Rep
0
5
10
15
20
25
30
35
40
45
Delikat Desiree Baltica Agave
Control
A1
A2
Sh
oo
ts p
er e
xpla
nt
0
10
20
30
40
50
60
70
80
90
100
Delikat Desiree Baltica Agave
% T
ran
sfo
rmat
ion
A 1
A 2
A
B
Fig. 3 Mean number of potato shoots regenerated on stem internodalexplants (A) and the percentage of regenerated shoots expressing gfp (B)in potato tetraploid cvs. following co-cultivation with A. tumefaciensLBA4404 pHB2892 and culture for 4 months on MS-t medium with250 mg L−1 cefotaxime. A1, selection on 50 mg L−1 kanamycin forthe first 5 weeks of culture; A2, permanent selection on 50 mg L−1
kanamycin. (Bars = SEM)
explants were co-cultivated with Agrobacterium LBA4404pHB2892 on MS-t medium. The most regenerated shootswere obtained for the cv. ‘Baltica’ (Fig. 3A), which showedno significant difference in the percentage of gfp express-ing shoots under permanent kanamycin selection, or whenkanamycin was eliminated from the medium after 5 weeks(Fig. 3B). The cvs. ‘Desiree’ and ‘Agave’ regenerated mostshoots per explant when kanamycin selection was not appliedafter 5 weeks of culture (Fig. 3A). The percentage of gfp ex-pressing shoots was for all cvs. and, particularly for the onesshowing poor shoot regeneration (‘Delikat’ and ‘Agave’), al-ways better under permanent kanamycin selection (Fig. 3B).The tetraploid cvs. showed polarity of regeneration fromstem internodal explants, with shoots developing at the api-cal pole and callus at the basal end, as illustrated for the cv.‘Baltica’ (Fig. 4). The most responsive genotypes, namelythe dihaploid 178/10 and the cv. ‘Baltica’, also regeneratedcallus and shoots more rapidly, the first shoots developing
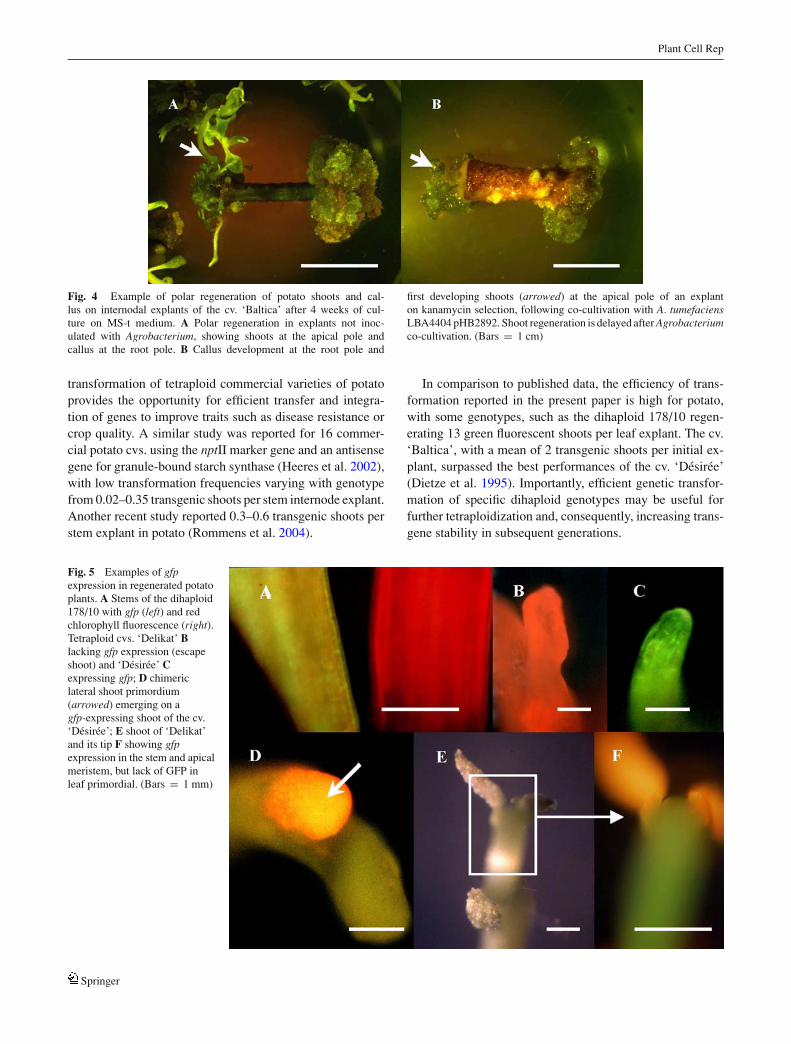
within 3–4 weeks of culture in control, uninoculated leafor stem explants. After Agrobacterium co-cultivation andkanamycin selection, the polarity of regeneration was main-tained, but shoot regeneration was delayed by 2–4 weeks,depending on the genotype (Fig. 4). A quite high percentage(30–100%) of regenerated shoots did not express gfp, as in di-haploids, and some chimeras were observed in tetraploid cul-tivars (Fig. 5). Chimeras were identified in the cv. ‘Desiree’,with low gfp expression in a secondary shoot (Fig. 5D); in thecv. ‘Delikat’, a plant was generated with a green fluorescentstem and meristem, but no GFP in the young leaves (Fig. 5Eand F). Interesting chimeric clones were identified in regen-erated shoots of the dihaploid genotype 178/10, includingsilencing of the gfp gene during apical bud development inone clone (Fig. 6), and silencing in collenchymatous cellsin another clone (Fig. 7A–D). A chimera exhibiting silenc-ing of gfp in the meristem of the axillary bud comparedto a transgenic clone of the same genotype expressing gfpconstitutively is shown in Fig. 7E and F.
NptII and gfp primers amplified 261 and 497 bp regionsof the respective genes. Molecular analysis of four chimericclones of the cv. ‘Delikat’, showed that all plants had inte-grated the nptII gene, as shown by PCR amplification (Fig.8). Northern analysis revealed that nptII expression was lessin the chimeric clone T3 as compared to the transgenic cloneT4 of the cv. ‘Delikat’ (Fig. 9). PCR amplification, RT-PCRconfirmed the integration and expression of the gfp gene inall the clones classified as transgenic by visual screening ofgreen fluorescence, and the absence of the gfp gene in theclones exhibiting the red autofluorescence of chlorophyll,considered as escapes (Figs. 10 and 11). The integration ofthe gfp gene in putative transgenic clones exhibiting greenfluorescence was confirmed by PCR and Southern blot anal-yses (Fig. 12), the latter showing the copy number to varybetween 2 in a chimeric plant and 5 in clones constitutivelyexpressing gfp. Neither gfp nor nptII were detected by PCRanalysis in controls or escapes (Figs. 8 and 10). RT-PCRanalysis of the gfp gene revealed no differences in its expres-sion in different transgenic clones of dihaploid or tetraploidgenotypes of potato (Fig. 11).
Discussion
GFP is a useful tool for monitoring the efficiency of trans-formation, the effects of antibiotics on organogenesis andtransgenic plant selection, and the occurrence of escapes andchimeras in potato dihaploid genotypes and tetraploid culti-vars. Previously, the cv. ‘Desiree’ was used extensively as amodel for potato transformation, but the German commercialpotato cultivars or dihaploid genotypes used in this study,were not subjected previously to Agrobacterium-mediatedgene transfer. Developing an efficient procedure for genetic
Springer
Plant Cell Rep
Fig. 4 Example of polar regeneration of potato shoots and cal-lus on internodal explants of the cv. ‘Baltica’ after 4 weeks of cul-ture on MS-t medium. A Polar regeneration in explants not inoc-ulated with Agrobacterium, showing shoots at the apical pole andcallus at the root pole. B Callus development at the root pole and
first developing shoots (arrowed) at the apical pole of an explanton kanamycin selection, following co-cultivation with A. tumefaciensLBA4404 pHB2892. Shoot regeneration is delayed after Agrobacteriumco-cultivation. (Bars = 1 cm)
transformation of tetraploid commercial varieties of potatoprovides the opportunity for efficient transfer and integra-tion of genes to improve traits such as disease resistance orcrop quality. A similar study was reported for 16 commer-cial potato cvs. using the nptII marker gene and an antisensegene for granule-bound starch synthase (Heeres et al. 2002),with low transformation frequencies varying with genotypefrom 0.02–0.35 transgenic shoots per stem internode explant.Another recent study reported 0.3–0.6 transgenic shoots perstem explant in potato (Rommens et al. 2004).
In comparison to published data, the efficiency of trans-formation reported in the present paper is high for potato,with some genotypes, such as the dihaploid 178/10 regen-erating 13 green fluorescent shoots per leaf explant. The cv.‘Baltica’, with a mean of 2 transgenic shoots per initial ex-plant, surpassed the best performances of the cv. ‘Desiree’(Dietze et al. 1995). Importantly, efficient genetic transfor-mation of specific dihaploid genotypes may be useful forfurther tetraploidization and, consequently, increasing trans-gene stability in subsequent generations.
Fig. 5 Examples of gfpexpression in regenerated potatoplants. A Stems of the dihaploid178/10 with gfp (left) and redchlorophyll fluorescence (right).Tetraploid cvs. ‘Delikat’ Blacking gfp expression (escapeshoot) and ‘Desiree’ Cexpressing gfp; D chimericlateral shoot primordium(arrowed) emerging on agfp-expressing shoot of the cv.‘Desiree’; E shoot of ‘Delikat’and its tip F showing gfpexpression in the stem and apicalmeristem, but lack of GFP inleaf primordial. (Bars = 1 mm)
Springer

Plant Cell Rep
Fig. 6 Expression of gfpduring stages of potato buddevelopment (A–F) in a putativetransgenic dihaploid clone178/10. (Bar = 2.5 mm)
A relatively high percentage of shoots not expressing gfpwere regenerated under permanent kanamycin selection inall potato genotypes tested. Molecular analysis revealed thatall shoots regenerated on kanamycin that did not show greenfluorescence, had not integrated the nptII gene. Such a highpercentage of these shoots may be possible because auxinpretreatment (10 mg L−1 NAA and 10 mg L−1 zeatin for2 days), as an initial step of the protocol (Kumar 1995),could act as a signal stimulating the regeneration of escapes.It also accounts for the polarity of regeneration as demon-strated for Citrus transformation (Domınguez et al. 2004).Moreover, as shown in Citrus (Pena et al. 2004), dedifferen-tiation is required to shift cells to a competent state for stabletransformation, and dedifferentiation is triggered by auxinsduring co-cultivation, as in Citrus, or as a pretreatment inthe current experiments. Another possible factor responsiblefor the escapes is the concentration of kanamycin, whichmay have been too low, although employed at the concentra-tion recommended previously for routine transformation ofthe cv. ‘Desiree’ (Kumar 1995). The persistence of agrobac-teria during shoot regeneration and the propagation of pu-
tative transgenic clones in vitro over prolonged time afterco-cultivation of explants with bacteria, may also accountfor the high percentage of escapes. Similar results have beenreported for Agrobacterium-mediated transformation of Cit-rus (Domınguez et al. 2004). The present results reinforcethe previous assumption (Domınguez et al. 2004) that selec-tion with kanamycin after Agrobacterium co-cultivation didnot prevent the regeneration of shoots that failed to integratethe nptII gene. However, in contrast to Citrus (Domınguezet al. 2004), PCR and RT-PCR for potato in the present studydid not show any case where escapes had integrated, but notexpressed, the nptII gene.
A kanamycin concentration of 50 mg L−1 and co-cultivation with Agrobacterium showed genotype-dependenteffects on organogenesis, reducing shoot yield and delayingshoot regeneration by 2–4 weeks, depending on the geno-type. Cefotaxime, used mainly at 250 mg L−1, also influ-enced organogenesis with stimulation of shoot regenerationin previous studies (data not presented). Other authors haverecommended, for systems that generate many escapes andchimeras, the use of a screenable strategy instead of lethal
Springer

Plant Cell Rep
Fig. 7 Different patterns ofexpression of gfp in stemsections of transgenic potatodihaploid 178/10. A Clone withconstitutive gfp expression. BStem with red chlorophyllfluorescence of a control(non-transformed) clone. CTransgenic clone where GFP isabsent from the collenchyma. DDetail of the same clone; arrowindicates collenchyma cellslacking gfp expression. EGeneral view of transgenicclone with constitutive gfpexpression in comparison with aclone F where GFP is absent inthe collenchyma and themeristem of an axillary bud(arrowed). (Bars = 1 mm,except D, where bar = 100 µm)
selection (Christou and McCabe 1992), with gfp expressionbeing preferred in previous experiments with other plantspecies (Ghorbel et al. 1999; Zhang et al. 2001). The presentresults demonstrate that the use of both gfp and nptII genesmay reveal the real picture of genotype-dependent transfor-mation efficiency. Observations of the effects of culture me-dia, antibiotics and Agrobacterium strains on organogenesis
may also be facilitated and permit rapid optimisation of trans-formation for a particular genotype. Such a transformationprotocol could be further used for efficient transfer of othergenes (Rakosy-Tican et al. 2004), eventually eliminating thenecessity to use genes for antibiotic resistance (De Vettenet al. 2003). The present data, based on PCR screening andSouthern blot analyses of transgenic shoots, also reinforce
Springer

Plant Cell Rep
261 bp
M C0 C+ T1 T2 T3 T4 T5 T6 T7 T8 C1 C2 C3 C4 C5
Fig. 8 PCR profiles generated using nptII primers for transgenic potatoclones and their respective control. M = 100 bp ladder; C0 = negativecontrol (water); C + = positive control (pHB2892) for nptII gene; T1–T4, T5–T6, T7 and T8 = clones of cvs. ‘Delikat’, ‘Desiree’, ‘Baltica’
and ‘Agave’ expressing gfp; C1–C5 = DNA from non-transgenicplants of cvs. ‘Agave’, ‘Delikat’, ‘Baltica’ (2) and ‘Desiree’, respec-tively
T 1 T 2 T 3 T 4C 1
Fig. 9 Northen blot analysis of transgenic potato clones. T1, T2 = cv.‘Agave’ expressing gfp; C1 = non-transformed clone of the cv.‘Agave’; T3 = a chimeric clone partially expressing gfp of the cv.‘Delikat’; T4 = transgenic clone expressing gfp of the cv. ‘Delikat’
previous reports in potato with 1–5% transformation effi-ciency (De Vetten et al. 2003), cabbage (Berthomieu et al.1994) and Citrus (Domınguez et al. 2004), in which PCRanalysis was proposed to identify transgenic plants withoutthe use of selectable markers, since the latter raise concerns inrelation to the environment and food safety. Furthermore, thepresent results reveal the key role of the culture medium andauxin pretreatment to increase transgenic shoot regenerationin responsive cvs. In contrast with the results of Domınguezet al. (2004) in Citrus, there was no molecular proof thattransformation of potato may occur at a greater frequencythan indicated by screening for the gfp reporter gene.
The regeneration of chimeras with green fluorescence insome tissues or cells but not in others, was also reportedin tobacco and analysed quantitatively, suggesting involve-ment of a special type of gene silencing (Bastar et al. 2004).Moreover, by analysing the intensity of gfp expression, ho-mozygous and hemizigous states could be visualised in to-bacco, with Mendelian segregation being reported in T1 andT2 generations (Molinier et al. 2000). Differences in theintensity of green fluorescence were not observed in the cur-rent experiments, and chimeras occurred only in some of thegenotypes at low frequency ( < 0.5%), except in the dihap-loid 178/10, where 15% of the shoots regenerated from leafexplants were chimeric. Site-specific integration effects, aswell as reduction in GFP protein synthesis or gene silenc-ing, may be the causes of the chimeras observed to date.In contrast to tobacco (Molinier et al. 2000), natural fluo-rescence that interfered with gfp expression was not evidentin potato. All transgenic shoots expressing gfp were stableduring repeated cloning in vitro, without any adverse ef-fects on growth or development. Variation was observed ingfp expression with silencing during bud development, in
M C+ C0 C1 C2 C+ C3 C4 C5 R1 T1 T2 T3 T4 T5 T6 T7 T8 T9 T10 T11 T12 T13
497 pb
Fig. 10 PCR profiles generated using gfp primers for transgenic potatoclones and their respective controls. M = 100 bp. ladder; C + = pos-itive control (pHB 2892) for gfp; C0 = negative control (water); C1–C5 = non-transgenic clones of the cvs. ‘Agave’, ‘Delikat’, ‘Baltica’,‘Desiree’ and the dihaploid 178/10, respectively; R1 = clone of the
cv. ‘Baltica’ not expressing gfp; transgenic clones expressing gfp of thecvs. ‘Agave’ (T1, T2), ‘Delikat’ (T3, T4), ‘Baltica’ (T5–T7), ‘Desiree’(T8) and the dihaploid 178/10 (T9); T10–T13 = chimeras of the cv.‘Delikat’
M C0 C+ T1 T2 T3 T4 T5 T6 T7 T8 T9 T10 T11 C1
Fig. 11 RT-PCR profiles generated using gfp primers for transgenicpotato clones and their respective controls. M = 100 bp. ladder;C0 = negative control (water); C + = positive control (pHB 2892) forgfp; C1 = non-transgenic clone of the cv. ‘Agave’; transgenic clones
expressing gfp of the cvs. ‘Agave’ (T1, T2), ‘Delikat’ (T3, T4), ‘Baltica’(T5, T6), ‘Desiree’ (T7) and the dihaploid 178/10 (T10); T8, T9 andT11 = chimeras of the cv. ‘Delikat’
Springer

Plant Cell Rep
M T1 T2 T 3 C1
2176 bp
1766 bp
1230 bp
653 bp
234 bp
Fig. 12 Southern analysis of DNA from selected transgenic potatoclones using a gfp probe. M = DIG-labelled DNA Molecular WeightMarker VI (Roche); T1 = a chimeric clone of the cv. ‘Delikat’ with acopy number of 2; T2, T3 = transgenic clones of the cvs. ‘Agave’ (T2)and ‘Desiree’ (T3) constitutively expressing gfp, both with five copiesof the gene; C1 = non-transformed clone of the cv. ‘Agave’
collenchymatous cells in some chimeric clones of the di-haploid 178/10. Silencing of gfp was also seen in lateralbranches and leaf primordia of specific tetraploid potato cvs.Such variation in gfp expression during development may beexplained by integration site-specific gene inactivation; suchplants may represent valuable material for further molecularinvestigations.
Preliminary data from the experiments with potato in-dicated that Agrobacterium strain EHA105 compared toLBA4404, may also affect explant response to co-cultivation,but further research is required to improve understanding ofAgrobacterium–plant tissue interaction, as indicated by Penaet al. (2004).
Visual screening of gfp expression may be used to opti-mise genetic transformation of specific cultivars of potato toeliminate the difficulties associated with kanamycin selec-tion. Selection on kanamycin-containing medium could notdiscriminate transgenic shoots from escapes and chimeras.Kanamycin has genotype-dependent effects on organogene-sis in potato and increasing its concentration for tighter se-lection may affect the regeneration of transgenic plants. Sucheffects can be monitored easily using the gfp reporter–nptIImarker system that may be suitable for other plant species.
In conclusion, a gfp reporter–nptII marker system wasused to optimise genetic transformation of dihaploid andtetraploid potato genotypes. Genotype-dependent responseto culture media, the effects of kanamycin selection andthe transformation protocol, as well as the frequency of es-capes, are important factors for establishing strategies to en-hance transformation efficiency in this tuberous crop. How-
ever, it was also found, as indicated previously (Domınguezet al. 2004), that there are several assumptions relating toAgrobacterium-mediated transformation that need to be crit-ically assessed. Transformation efficiency, based on resis-tance to a selective agent, may not reveal the actual fre-quency of transgenic plant regeneration in potato. However,screening for the expression of the gfp gene gave the realfigure for transformation in the present experiments. Impor-tantly, gfp allows identification of escapes and chimeras invivo, permitting further investigation of the causes for theregeneration of escapes in some genotypes, together withstudies on silencing and site-integration effects. Further-more, for vegetatively propagated crops that are known toregenerate chimeras and escapes (Domınguez et al. 2004),establishing a robust protocol for transformation, avoidingmarker based-selection of transgenic shoots, modifying theantibiotic treatment to reduce Agrobacterium persistence af-ter co-cultivation and screening transgenic events by PCR,are advisable to maximise transformation efficiencies andthe generation of marker-free plants.
Acknowledgements A German Academic Exchange Service(DAAD) Scholarship and a German–Romanian Bilateral Project areacknowledged for supporting part of this research (ER-T). The EUErasmus–Socrates programme provided partial support for CD. Dr. G.Hahne and Dr. J. Molinier kindly provided pHB2892.
References
Bastar MT, Luthar Z, Skof S, Bohanec B (2004) Quantitative determi-nation of mosaic GFP gene expression in tobacco. Plant Cell Rep22:939–944. DOI 10.1007/s00299-004-0782-2
Berthomieu P, Beclin C, Charlot F, Dore C, Jouanin L (1994) Routinetransformation of rapid cycling cabbage (Brassica oleracea)—molecular evidence for the regeneration of chimeras. Plant Sci96:223–235
Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC (1994) Greenfluorescent protein as a marker for gene expression. Science263:802–805. DOI 10.1126/science.8303295
Christou P, McCabe DE (1992) Prediction of germ-line transfor-mation events in chimeric R0 transgenic soybean plantlets us-ing tissue-specific expression patterns. Plant J 2:283–290. DOI10.1111/j.1365-313X.1992.00283.x
De Vetten N, Wolters AM, Raemakers K, Van Der Meer I, TerStege R, Heeres E, Heeres P, Visser R (2003) A transformationmethod for obtaining marker-free plants of a cross-pollinated andvegetatively propagated crop. Nat Biotechnol 21:439–442. DOI10.1038/nbt801
Dietze J, Blau A, Willmitzer L (1995) Agrobacterium-mediated trans-formation of potato (Solanum tuberosum). In: Potrykus I, Spangen-berg G (eds) Gene transfer to plants. Springer, Berlin HeidelbergNew York, pp 24–29
Domınguez A, Cervera M, Perez RM, Romero J, Fagoaga C,Cubero J, Lopez MM, Juz JA, Navaro L, Pena L (2004)Characterisation of regenerants obtained under selective con-ditions after Agrobacterium-mediated transformation of citrusexplants reveals production of silenced and chimeric plantsat unexpected high frequencies. Mol Breed 14:171–183. DOI10.1023/B:MOLB.0000038005.73265.61
Springer

Plant Cell Rep
Elliott AR, Campbell JA, Dugdale B, Brettel RIS, Grof CPI (1999)Green fluorescent protein facilitates rapid in vivo detection ofgenetically transformed plant cells. Plant Cell Rep 18:707–714.DOI 10.1007/s002990050647
Ghorbel R, Juarez J, Navarro L, Pena L (1999) Green fluorescent proteinas a screenable marker to increase the efficiency of generatingtransgenic woody fruit plants. Theor Appl Genet 99:350–358.DOI 10.1007/s001220051244
Halford NG (2004) Prospects for genetically modified crops. Ann ApplBiol 145:17–24. DOI 10.1111/j.1744-7348.2004.tb00355.x
Haselhof J, Siermering KR, Prasher DC, Hodge S (1997) Removal ofa cryptic intron and subcellular localization of green fluorescentprotein are required to mark transgenic Arabidopsis plants brightly.Proc Natl Acad Sci USA 94:2122–2127
Heeres P, Schippers-Rozenboom M, Jacobsen E, Visser RGF (2002)Transformation of a large number of potato varieties: genotype-dependent variation in efficiency and somaclonal variability. Eu-phytica 124:13–22. DOI 10.1023/A:1015689112703
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusion: β-glucuronidase as a sensitive and versatile gene fusion marker inhigher plants. EMBO J 6:3901–3907
Kaeppler HF, Carlson AR, Menon GK (2001) Routine utilization ofgreen fluorescent protein as a visual marker for cereal transforma-tion. In vitro Cell Dev Biol–Plant 37:120–126
Kohler R (1998) GFP for in vivo imaging of subcellular structures inplant cells. Trends Plant Sci 3:317–320
Kumar A (1995) Agrobacterium-mediated transformation of potatogenotypes. In: Gartland KMA, Davey MR (eds) Methods in molec-ular biology, vol 44. Humana Press Inc, New York Totowa, pp121–128
Menczel L, Nagy F, Kiss ZsR, Maliga P (1981) Streptomycin resistantand sensitive somatic hybrids of Nicotiana tabacum + Nicotianaknightiana correlation of resistance to N. tabacum plastids. TheorAppl Genet 59:191–195. DOI 10.1007/BF00264975
Mercuri A, Sacchetti A, De Benedetti A, Schiva T, Alberti S (2001)Green fluorescent flowers. Plant Sci 161:961–968
Molinier J, Himber C, Hahne G (2000) Use of green fluorescent proteinfor detection of transformed shoots and homozygous offspring.Plant Cell Rep 19:219–223. DOI 10.1007/s002990050002
Murashige T, Skoog F (1962) A revised medium for rapid growth andbio-assays with tobacco tissue cultures. Physiol Plant 15:473–497
Olivares-Fuster O, Pena L, Duran-Vila N, Navarro L (2002) Greenfluorescent protein as a visual marker in somatic hybridization.Ann Bot 89:491–497. DOI 10.1093/aob/mcf54
Ow DW, Wood KV, DeLuca M, Wet JR de, Helinski DR, Howell SH(1986) Transient and stable expression of the firefly luciferasegene in plant cell and transgenic plants. Science 234:856–859
Pena L, Perez RM, Cervera M, Juarez JA, Navarro L (2004) Earlyevents in Agrobacterium-mediated genetic transformation of Cit-rus explants. Ann Bot 94:67–74. DOI 10.1093/aob/mch117
Rakosy-Tican L, Aurori A, Aurori CM, Ispas G, Famelaer I (2004)Transformation of wild Solanum species resistant to late blightby using reporter gene gfp and msh2 genes. Plant Breed Seed Sci50:119–128
Rommens CM, Humara JM, Ye J, Yan H, Richael C, Zhang L, Perry R,Swords K (2004) Crop improvement through modification of theplant’s own genome. Plant Physiol 135:421–431. DOI 10.1104
Rizzuto R, Brini M, Pizzo P, Murgia M, Pozzan T (1995) Chimeric greenfluorescent protein as a tool for vusualizing subcellular organellesin living cells. Curr Biol 5:635–642. DOI 10.1016/SO960-9822(95)00128-x
Rouwendal GJA, Mendes O, Wolbert EJH, Douwe de Boer A (1997)Enhanced expression in tobacco of the gene encoding green fluo-rescent protein by modification of its codon usage. Plant Mol Biol33:989–999. DOI 10.1023/A:1005740823703
Ruiz MT, Voinnet O, Baulcombe DC (1998) Initiation and maintenanceof virus-induced gene silencing. Plant Cell 10:937–946
Sidorov VA, Kasten D, Pang S-Z, Hajdukiewicz PTJ, Staub JM, NehraNS (1999) Stable chloroplast trasnsformation in potato: use ofgreen fluorescent protein as a plastid marker. Plant J 19:209–216.DOI 10.1046/j.1365-313X.1999.00508.x
Sheerman S, Bevan MW (1988). A rapid transformation method forSolanum tuberosum using binary Agrobacterium tumefaciens vec-tors. Plant Cell Rep 7:13–16. DOI 10.1007/BF00272967
Stiekema WJ, Heidekamp F, Louwerse JD, Verhoeven HA, Dijkhuis P(1988) Introduction of foreign genes into potato cultivars Bintjeand Desiree using an Agrobacterium tumefaciens binary vector.Plant Cell Rep 7:47–50. DOI 10.1007/BF00272976
Zhang CL, Chen DF, McCormac AC, Scott NW, Elliot MC, SlaterA (2001) Use of the GFP reporter as a vital marker forAgrobacterium-mediated transformation of suger beet (Beta vul-garis L.). Mol Biotechnol 17:109–117 (ISSN 1073-6085)
Springer
Copyright © 2022 FDOKUMEN




















