
New transposons to generate GFP protein fusions in Candida albicans
6
New transposons to generate GFP protein fusions in Candida albicans Marcos Vinicios Salles Dias, Luis Roberto Basso Jr. 1 , Paulo Sergio Rodrigues Coelho ⁎ Departamento de Biologia Celular e Molecular e Bioagentes Patogênicos, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Av. dos Bandeirantes 3.900, Ribeirão Preto, SP, Brazil ABSTRACT ARTICLE INFO Article history: Received 6 November 2007 Received in revised form 11 March 2008 Accepted 11 March 2008 Available online 19 March 2008 Received by J.V. Moran Keywords: Candida albicans Transposon GFP Septin Protein localization Transposon elements are important tools for gene function analysis, for example they can be used to easily create genome-wide collections of insertion mutants. Transposons may also carry sequences coding for an epitope or fluorescent marker useful for protein expression and localization analysis. We have developed three new Tn5-based transposons that incorporate a GFP (green fluorescent protein) coding sequence to generate fusion proteins in the important fungal pathogen Candida albicans. Each transposon also contains the URA3 and Kan R genes for yeast and bacterial selection, respectively. After in vitro transposition, the insertional allele is transferred to the chromosomal locus by homologous recombination. Transposons Tn5- CaGFP and Tn5-CaGFP-URA3::FLIP can generate C-terminal truncated GFP fusions. A URA3 flipper recycling cassette was incorporated into the transposon Tn5-CaGFP-URA3::FLIP . After the induction of Flip recombinase to excise the marker, the heterozygous strain is transformed again in order to obtain a GFP-tagged homozygous strains. In the Tn5-CaGFP-FL transposon the markers are flanked by a rare-cutting enzyme. After in vitro transposition into a plasmid-borne target gene, the markers are eliminated by restriction digestion and religation, resulting in a construct coding for full-length GFP-fusion proteins. This transposon can generate plasmid libraries of GFP insertions in proteins where N- or C-terminal tagging may alter localization. We tested our transposon system by mutagenizing the essential septin CDC3 gene. The results indicate that the Cdc3 C-terminal extension is important for correct septin filament assembly. The transposons described here provide a new system to obtain global gene expression and protein localization data in C. albicans. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Candida albicans is an opportunistic pathogen and the fourth leading cause of nosocomial infections (Pfaller and Diekema, 2007). The sequencing of its diploid genome (Jones et al., 2004) and the development of molecular genetic tools such as prototrophic and dominant markers for homozygous gene deletions and reporters of gene expression (Berman and Sudbery, 2002; Noble and Johnson, 2007) have made it possible to determine gene function in a large- scale fashion (Bruno and Mitchell, 2004). In fungi as in other organisms, transposons are important tools for making libraries of insertion mutants carrying reporters of gene expression and tags for protein localization (Ross-Macdonald et al., 1997; Merkulov and Boeke,1998; Ross-Macdonald et al., 1999; Judson and Mekalanos, 2000; Goyard et al., 2001; Firon et al., 2003; Bestor, 2005). In C. albicans, two transposon systems have been developed that incorporates different genetic elements. The first one, a Tn7 transposon containing a URA3 auxotrophic marker and a lacZ reporter gene was used to in vitro mutagenize a library of genomic fragments (Uhl et al., 2003). After transposition, the mutant alleles were pooled and transferred to the genome by homologous recombination to obtain a heterozygous mutant collection. Eigtheen thousand strains were screened for defects of filamention in agar media due to haploinsufficiency, resulting in the identification of 146 genes probably involved in the yeast to hypha dimorphic transition. Another study using transposon mutagenesis was performed by Davis et al. (2002). In this case, the authors used a Tn7 transposon containing a UAU1 selectable marker that allows for homozygous gene disruption in a single step. The UAU1 cassette consists of two prototrophic markers: a URA3 gene that is interrupted by the ARG4 gene. After disruption of the first allele of a target gene, mitotic recombination will result in homozygosis of the mutation. Subse- quently, intramolecular recombination at one of the UAU1 markers will result in deletion of ARG4 and reconstitution of URA3(Enloe et al., 2000). The homozygous disrupted strains are selected in agar media without uridine and arginine. Many screens have been done Gene 417 (2008) 13–18 Abbreviations: GFP, green fluorescent protein; bGal, b-galactosidase; bp, base pair(s); BSA, bovine serum albumin; DMSO, dimethylsulfoxide; DTT, dithiothreitol; EDTA, ethylene diamine tetra-acetate; FLP, Flip recombinase; kb, 1000 bp; kDa, kilodalton(s); lacZ, Beta galactosidase gene; LB, Luria-Bertani (medium); ORF, open reading frame; SDS_PAGE, sodium dodecyl sulfate polyacrylamide-gel electrophoresis; PCR, polymerase chain reaction; PMSF, phenylmethylsulfonyl fluoride; SDS, sodium dodecyl sulfate; Tn, transposon; UTR, untranslated region(s); URA3, orotidine 5-phosphate decarboxylase; wt, wild type. ⁎ Corresponding author. Tel.: +5516 3602 3257; fax: +55 16 3633 1786. E-mail address: [email protected] (P.S.R. Coelho). 1 Present address: Department of Medicine, Division of Infectious Diseases, Oregon Health & Science University, 3181 S.W. Sam Jackson Park Rd. Portland, Oregon 97239-3098. 0378-1119/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.gene.2008.03.005 Contents lists available at ScienceDirect Gene journal homepage: www.elsevier.com/locate/gene
Transcript of New transposons to generate GFP protein fusions in Candida albicans

Gene 417 (2008) 13–18
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r.com/ locate /gene
New transposons to generate GFP protein fusions in Candida albicans
Marcos Vinicios Salles Dias, Luis Roberto Basso Jr. 1, Paulo Sergio Rodrigues Coelho ⁎Departamento de Biologia Celular e Molecular e Bioagentes Patogênicos, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Av. dos Bandeirantes 3.900, Ribeirão Preto,SP, Brazil
Abbreviations:GFP, greenfluorescent protein; bGal, bBSA, bovine serum albumin; DMSO, dimethylsulfoxidethylene diamine tetra-acetate; FLP, Flip recombinase; klacZ, Beta galactosidase gene; LB, Luria-Bertani (mediuSDS_PAGE, sodiumdodecyl sulfate polyacrylamide-gel elechain reaction; PMSF, phenylmethylsulfonyl fluoride; SDtransposon; UTR, untranslated region(s); URA3, orotidinwt, wild type.⁎ Corresponding author. Tel.: +55 16 3602 3257; fax:
E-mail address: [email protected] (P.S.R. Coelho1 Present address: Department of Medicine, Division
Health & Science University, 3181 S.W. Sam Jackso97239-3098.
0378-1119/$ – see front matter © 2008 Elsevier B.V. Alldoi:10.1016/j.gene.2008.03.005
A B S T R A C T
A R T I C L E I N F OArticle history:
Transposon elements are im Received 6 November 2007Received in revised form 11 March 2008Accepted 11 March 2008Available online 19 March 2008Received by J.V. Moran
Keywords:Candida albicansTransposonGFPSeptinProtein localization
portant tools for gene function analysis, for example they can be used to easilycreate genome-wide collections of insertion mutants. Transposons may also carry sequences coding for anepitope or fluorescent marker useful for protein expression and localization analysis. We have developedthree new Tn5-based transposons that incorporate a GFP (green fluorescent protein) coding sequence togenerate fusion proteins in the important fungal pathogen Candida albicans. Each transposon also containsthe URA3 and KanR genes for yeast and bacterial selection, respectively. After in vitro transposition, theinsertional allele is transferred to the chromosomal locus by homologous recombination. Transposons Tn5-CaGFP and Tn5-CaGFP-URA3::FLIP can generate C-terminal truncated GFP fusions. A URA3 flipper recyclingcassette was incorporated into the transposon Tn5-CaGFP-URA3::FLIP. After the induction of Flip recombinaseto excise the marker, the heterozygous strain is transformed again in order to obtain a GFP-taggedhomozygous strains. In the Tn5-CaGFP-FL transposon the markers are flanked by a rare-cutting enzyme. Afterin vitro transposition into a plasmid-borne target gene, the markers are eliminated by restriction digestionand religation, resulting in a construct coding for full-length GFP-fusion proteins. This transposon cangenerate plasmid libraries of GFP insertions in proteins where N- or C-terminal tagging may alter localization.We tested our transposon system by mutagenizing the essential septin CDC3 gene. The results indicate thatthe Cdc3 C-terminal extension is important for correct septin filament assembly. The transposons describedhere provide a new system to obtain global gene expression and protein localization data in C. albicans.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Candida albicans is an opportunistic pathogen and the fourthleading cause of nosocomial infections (Pfaller and Diekema, 2007).The sequencing of its diploid genome (Jones et al., 2004) and thedevelopment of molecular genetic tools such as prototrophic anddominant markers for homozygous gene deletions and reporters ofgene expression (Berman and Sudbery, 2002; Noble and Johnson,2007) have made it possible to determine gene function in a large-scale fashion (Bruno and Mitchell, 2004).
In fungi as in other organisms, transposons are important tools formaking libraries of insertion mutants carrying reporters of gene
-galactosidase; bp, base pair(s);e; DTT, dithiothreitol; EDTA,b, 1000 bp; kDa, kilodalton(s);m); ORF, open reading frame;ctrophoresis; PCR, polymeraseS, sodium dodecyl sulfate; Tn,e 5-phosphate decarboxylase;
+55 16 3633 1786.).of Infectious Diseases, Oregonn Park Rd. Portland, Oregon
rights reserved.
expression and tags for protein localization (Ross-Macdonald et al.,1997; Merkulov and Boeke, 1998; Ross-Macdonald et al., 1999; Judsonand Mekalanos, 2000; Goyard et al., 2001; Firon et al., 2003; Bestor,2005). In C. albicans, two transposon systems have been developedthat incorporates different genetic elements. The first one, a Tn7transposon containing a URA3 auxotrophic marker and a lacZ reportergene was used to in vitro mutagenize a library of genomic fragments(Uhl et al., 2003). After transposition, the mutant alleles were pooledand transferred to the genome by homologous recombination toobtain a heterozygous mutant collection. Eigtheen thousand strainswere screened for defects of filamention in agar media due tohaploinsufficiency, resulting in the identification of 146 genesprobably involved in the yeast to hypha dimorphic transition.Another study using transposon mutagenesis was performed byDavis et al. (2002). In this case, the authors used a Tn7 transposoncontaining a UAU1 selectable marker that allows for homozygousgene disruption in a single step. The UAU1 cassette consists of twoprototrophic markers: a URA3 gene that is interrupted by the ARG4gene. After disruption of the first allele of a target gene, mitoticrecombination will result in homozygosis of the mutation. Subse-quently, intramolecular recombination at one of the UAU1 markerswill result in deletion of ARG4 and reconstitution of URA3 (Enloeet al., 2000). The homozygous disrupted strains are selected in agarmedia without uridine and arginine. Many screens have been done

14 M.V.S. Dias et al. / Gene 417 (2008) 13–18
using a library of UAU1 transposed mutant strains to look for genesinvolved in different cellular process such as filamentous growth atalkaline pH, and chlamydospore and biofilm formation which areimportant virulence attributes (Davis et al., 2002; Nobile et al., 2003;Nobile and Mitchell, 2005).
A codon-optimized GFP (green-fluorescent protein) coding genehas been constructed for expression in C. albicans (Cormack et al.,1997). Several studies have demonstrated the usefulness of thismarker for evaluation of gene expression and for protein localizationeither in cultures or during infection (Morschhauser et al., 1998;Gerami-Nejad et al., 2001; Mao et al., 2003; Barelle et al., 2004; Greenet al., 2005; Barelle et al., 2006; Schaub et al., 2006). In this study, ourgoal was to construct new Tn5 based transposons bearing a C. albicansGFP reporter gene that can be used to make libraries of mutant strainsexpressing random GFP protein fusions.
2. Materials and methods
2.1. Strains and media
The C. albicans strains used in this study are indicated in Table 1.YPD (10 g yeast extract, 20 g peptone, 20 g glucose per liter) was usedfor routine growth of C. albicans. URA3+ transformants were selectedon media synthetic complete lacking uracil.
2.2. Construction of transposons
To generate Tn5CaGFP, we used the Tn5 transposon carried onplasmid pMOD-3bR6Kγori/MCSN (Epicentre) as a backbone. Thistransposon contains the conditional origin of replication R6Kγ and amultiple cloning site (MCS) flanked by the hyperactive 19 bp MosaicEnds (ME) that are specifically recognized by EZ::TN Transposase(Reznikoff, 2006). A ORF encoding a C. albicans-optimized GFP wasamplified by PCR using the plasmid pGFP-URA3 (Gerami-Nejad et al.,2001) as a template and oligonucleotide primers GFP-F containing aEcoRI restriction site at its 5′ end and GFP-R containing a SacIrestriction site at its 3′ end (Table 1). This PCR fragment was ligatedto pCR2.1-TOPO (Invitrogen). A 0.7 Kb EcoRI-SacI fragment encodingGFP was ligated to pMOD-3 digested with the same enzymes, whichyielded pMD1. The KanR gene was released from the vector pGPS3(New England Biolabs) as a EcoRV fragmentand ligated to pMD1previously digested with HindIII and made blunt with T4 DNA
Table 1Oligonucleotides and strains
Name Description
OligonucleotidesEcoRI GFP forward GAATTCGTCTAAAGGTGAAGAATTATTSacI GFP reverse GAGCTCCCAGACAATGTCTTTGCCGTTn5-GFP forward GGTACCCTGTCTCTTATACACATCTCTAAAGGTTn5-GFP reverse GAGCTCTTAGGCGCGCCCTTTGTACTA CCATGTn5-Kan forward TCTAGATGCTAAAGGAAGCGGAACTn5-Kan reverse CTGTCTCTTATACACATCTCCGGCGCGCTAAGGCdc3 F AAGCGGCCGCCTCACCTCACAAATAATTCAGGCdc3 R AAGCGGCCGCGTAGTTATAACCCTATTGGCCAFP-1 ATTCAGGCTGCGCAACTGTRP-1 GTCAGTGAGCGAGGAAGCGGAAGpMOD seq reverse GAGCCAATATGCGAGAAACACCCGAGAA
Bacterial strainsDH10B F− araD 139 Δ(ara, leu) 7679 deoR endA 1 g
ZΔM15 mcrA nupG Δ(mmr-hsdRMS-mcrBC)
Yeast strainsSC5314 Wild typeCAI4 SC5314 Δura3::imm434/Δura3::imm434Y126 CAI4 CaCdc3::GFP (PCR-mediated GFP tagg
to Gerami-Nejad et al. (2001)
polymerase (New England Biolabs), which yielded pMD2. The URA3gene was released from the vector CIp10-Mal2p (Backen et al., 2000)as 1.2 kb SacI–XbaI fragment and ligated to pMD2 previouslydigested with the same enzymes, yielding pMD3 which carriesTn5CaGFP.
To generate Tn5CaGFP-URA3:FLIP and CaTn5-GFP-FL transposonswe used Litmus28i (New England Biolabs) as the cloning vector. WePCR amplified a GFP ORF (without the start and stop codons) usingprimers Tn5-GFP-F (with a KpnI restriction site and a Tn5 19 bp MEat its 5′ end) and Tn5-GFP-R (with AscI and SacI restriction sites atits 5′ end) and the plasmid pGFP-URA3 (Gerami-Nejad et al., 2001) asa template. This PCR fragment was ligated to pCR2.1-TOPO. A KpnI–SacI fragment encoding GFP was ligated into Litmus28i whichyielded pMD4. The KanR gene was PCR amplified with primers Kan-Fand Kan-R using pCR2.1-TOPO as a template. The resulting PCRfragment containing an AscI site and the Tn5 19 bp ME at its 3′ endwas ligated to pCR2.1-TOPO. A EcoRI fragment containing KanR genewas ligated into pMD4 previously digested with EcoRI, which yieldedpMD5. A 1.2 Kb SacI–XbaI fragment carrying the URA3 gene fromvector CIp10-Mal2p was then cloned into pMD5 between the GFPand the KanR gene resulting in pMD6 which carries the transposonTn5CaGFP-URA3-FL. We generated Tn5CaGFP-URA3::FLIP by ligating aXhoI (blunt)–SacI fragment containing the URA3:FLP cassette fromplasmid pSFU1 (Morschhauser et al., 1999) into pMD4 linearizedwith SacI and XbaI (blunt).
2.3. CaCDC3 cloning
The primers Cdc3F and Cdc3R (Table 1) were used to amplify a1556 bp product fromwild type strain SC5314 genomic DNA. The finalproduct contains the CaCDC3 ORF flanked by 128 bp upstream and141 bp downstream genomic sequence. This product was digestedwith NotI and cloned into the vector pUC19 (New England Biolabs)previously digested with NotI.
2.4. In vitro transposition
The transposon Tn5CaGFP-URA3 was amplified from their hostplasmid by PCR using the primers FP-1 and RP-1 (Table 1). Thetransposition reaction was performed according to manufacturer'srecommendations (Epicentre Technologies, Madison, WI) with200 ng of target plasmid pUC19 plus CaCDC3 and a molar equivalent
Reference
This studyThis study
GAAGAATTATT This studyGGTAATA This study
This studyGCGACACGGAAAT This study
This studyTC This study
EpicentreEpicentreEpicentre
alU galK ΔlacX74 Λ-Φ80dlacrecA 1 rpsL
(Gillum et al., 1984)(Fonzi and Irwin, 1993)
ed at the C-terminus according (P. Coelho and M. Snyder,unpublished results)

15M.V.S. Dias et al. / Gene 417 (2008) 13–18
of purified transposon Tn5CaGFP. Electrocompetent E. coli DH10βwere transformed with the transposition reaction sample and platedon LB agar with either ampicillin (100 μg/mL) and kanamycin (30 μg/mL) to recover transposed clones, or ampicillin (100 μg/mL) alone toestablish the transposition efficiency. Clones with transpositionevents in CaCDC3 were verified by restriction analysis. Sequencingof the positive clones was done using primer pMOD reverse sequenceprimer (Table 1).
2.5. C. albicans transformation
C. albicans strain CAI4 was transformed with NotI-digestedplasmids carrying the CDC3 transposed alleles using a lithium acetateprotocol as previously described (Braun and Johnson, 1997). Trans-formants were selected in synthetic complete media lacking uracil.
2.6. Western blotting
Stationary phase C. albicans yeast cells were diluted into fresh YPDmedium and incubated at 30 °C for 4 h. Cells were pelleted, washedonce in 1ml of PBS and then in 1ml of lysis buffer (50mMTris–HCl pH7.5, 5 mM EDTA, 100 mM NaCl, 40 mg/ml pepstatin A, 40 mg/mlaprotinin, 20 mg/ml leupeptin, 0.5 mM PMSF, 0.1% β-mercaptoetha-nol). The cells were then pelleted and the supernatant discarded, twovolumes of acid washed glass beads were added to the pellet andfurther lysis buffer was added to cover the glass beads. Tubes werethen vortexed (5X 20 s) and then placed immediately on ice. Thesupernatant was collected after low speed centrifugation at 2000 rpmfor 2 min and then transferred to a new tube and centrifuged at14,000 rpm for a further 5 min at 4 °C. Protein concentrations weredetermined by Bradford assay. The supernatant was loaded onto a 12%SDS-PAGE and separated proteins were electroblotted to a Hybond Cnitrocellulose membrane (Amersham Biosciences). The blot wasincubated with rabbit anti-GFP primary antibody diluted 1:10,000 (agift from Frederico Gueiros-Filho) and then with goat anti-rabbitsecondary antibody conjugated to peroxidase (Jackson Laboratory)
Fig. 1. Schematic representation of the transposons constructed. Each transposon contains thexpression in C. albicans. In each case the transposon are flanked by Tn5 terminal repeats (Transposon Tn5CaGFP-URA3::FLIP contains a URA3-Flip cassette. (C) Transposon Tn5CaGFP-FLURA3 and KanR markers. SAP2p: SAP2 promoter. Frt: Flipase recognition element. Relevantindicate the size of each transposon in base pairs.
diluted 1:20,000. The antigen-antibody complexes were visualizedwith ECL (Amersham Biosciences).
2.7. Microscopy
Yeast cells were grown for 4 h at 30 °C. 1 mL samples were fixed in5% formaldehyde for 30 min, washed twice with PBS and resuspendedin mounting medium. Cells were visualized by confocal microscopy(Leica SP5) using GFP excitation and emission filters. Images wereprocessed using Adobe Photoshop 6.0.
3. Results and discussion
3.1. Transposons for GFP tagging
In this study, all transposons are based on the Tn5 bacterialtransposon (Reznikoff, 2006) that is flanked by minimal sequenceelements (19 bp mosaic ends) necessary for efficient in vitrotransposition. The transposon constructions are described in themethods section. The transposon Tn5CaGFP (Fig. 1A) carries a GFP genewith codons optimized for expression in C. albicans (Cormack et al.,1997) and URA3 auxotrophic and kanamycin resistance genes forselection of yeast and bacterial transformants, respectively. The GFPgene contains a stop codon and lacks amethionine initiation codon, soTn5CaGFP is useful to obtain random GFP fusions in a genomic DNAlibrary or in a cloned gene of interest when GFP is inserted in thecorrect orientation and frame of the target sequence. The transposedalleles can be transferred to the chromosome by homologousrecombination to generate heterozygous strains. A R6Kγ origin ofreplication lies between de kanamycin resistance gene and the rightterminal repeat. The transposon Tn5CaGFP-URA3::FLIP (Fig. 1B)incorporates a URA3::Flipper cassette that carries a Flip recombinase(Morschhauser et al., 1999). After in vitro transposition and transfor-mation to replace the first allele, yeast cells are induced with BSA toexpress Flip, which recombines the Frt flanking sites and consequentlyeliminates the URA3 marker. So the URA3 marker excision enables
e coding regions for the kanamycin resistance, the Ura3 protein and a GFP optimized forTn5 repeat is not drawn to scale). (A) Transposon Tn5CaGFP contains a R6kγ origin. (B)contains a GFP without the start and stop codons and AscI flanking sites for excision ofrestriction sites used in cloning are indicated by arrows. The numbers in parentheses

16 M.V.S. Dias et al. / Gene 417 (2008) 13–18
yeast cells to be transformed againwith the original construct in orderto produce a homozygous insertional mutant.
The two transposons described so far generate C-terminaltruncated fusions. However in cases where a sorting motif is locatedat the C-terminus of the protein, such as resident proteins in theendoplasmic reticulum and GPI-anchored proteins, the truncatedversions will not be correctly localized. We developed a transposonsystem (Tn5CaGFP-FL) that circumvents this problem. After in vitrotransposition and bacterial transformation, the transposed plasmidsare digested with the AscI enzyme to remove the markers, and thenreligated to reconstitute a reading frame that extends across the entiretarget gene and the GFP inserted sequence (without the stop codon).Thus the final religated plasmid will carry a gene coding for a full-length protein with a GFP insertion. The GFP domain is bordered byamino acid linkers encoded by the Tn5 MEs as well as the 9 bpduplication of the target sequence that is generated during thetransposition process. Plasmid libraries carrying random GFP inser-tions in a gene of interest can be generated by this approach. If theplasmids are integrative, it might be possible to transfer thetransposed alleles to the chromosome.
3.2. Testing the Tn5CaGFP transposon
As a target of the transposon Tn5CaGFP (Fig. 1A), we used aplasmid carrying the CDC3 coding sequence flanked by 128 bpupstream of the start codon and 141 bp downstream of stop codon.The product of the essential CDC3 gene is a member of the septinfamily of cell cortex proteins (Kim et al., 1991; Douglas et al., 2005;Versele and Thorner, 2005). The septin proteins in yeast assembleinto a filament ring at the mother bud neck and functions as ascaffold for recruiting several proteins involved in fundamentalcellular processes such as cytokinesis, cell polarity, and bud site
Fig. 2. (A) Domain structure of septin Cdc3. The transposon insertion points are indicatedprepared, treatedwith SDS sample buffer, loaded into SDS-PAGE, transferred to nitrocelluloseThe amino acid insertion point is indicated on the top of each lane.
selection (Douglas et al., 2005; Versele and Thorner, 2005). Aftertransposition, E. coli cells were transformed and plated in mediacontaining kanamycin and ampicillin to select for clones withtransposon insertions. The efficiency of transposition was 1%,which is relatively high if compared with results obtained by othergroups (Reznikoff et al., 2004). This indicates that the size of thetransposon and its genetic elements do not present any significantconstraint for the transposition. To verify which clones hadtransposon insertions in CDC3, we performed restriction andsequencing analysis. Of 96 clones obtained 40 presented insertionswithin CDC3, and among these 6 had in-frame GFP insertions(Fig. 2A). The number of in-frame insertions obtained is inaccordance with the prediction (1/6). We released the inserts byrestriction digestions of the flanking NotI sites and then transformedC. albicans cells. By homologous recombination, the CDC3–GFP fusionallele substitutes the wild type allele. One copy of the wild type allelestill remains after allele replacement since C. albicans is an obligatorydiploid. Western analysis revealed that all GFP-fusions wereexpressed although in lower levels compared to the full-lengthversion of CDC3–GFP fusion (Fig. 2B).
3.3. Subcellular localization of CDC3–GFP fusions
In C. albicans yeast cells, like S. cerevisiae yeast cells, septinslocalize in the cell periphery forming initially a cap at the incipientbud site in G1 phase (Sudbery, 2001; Warenda and Konopka, 2002;Sudbery et al., 2004). The assembly of a septin ring filament occursupon the bud emergence at the start of S phase. This ring expands toform a collar that extends in the neck from the mother to thedaughter sides. At the end of mitosis the septin collar splits into tworings that delineate the site of cytokinesis and septum formation.After the completion of mitosis and upon entry in G1 phase the
by triangles. (B) Western blot analysis of septin-GFP fusions. Yeast total extracts weremembrane and incubatedwith anti-GFP antibody. Marker sizes are indicated on the left.
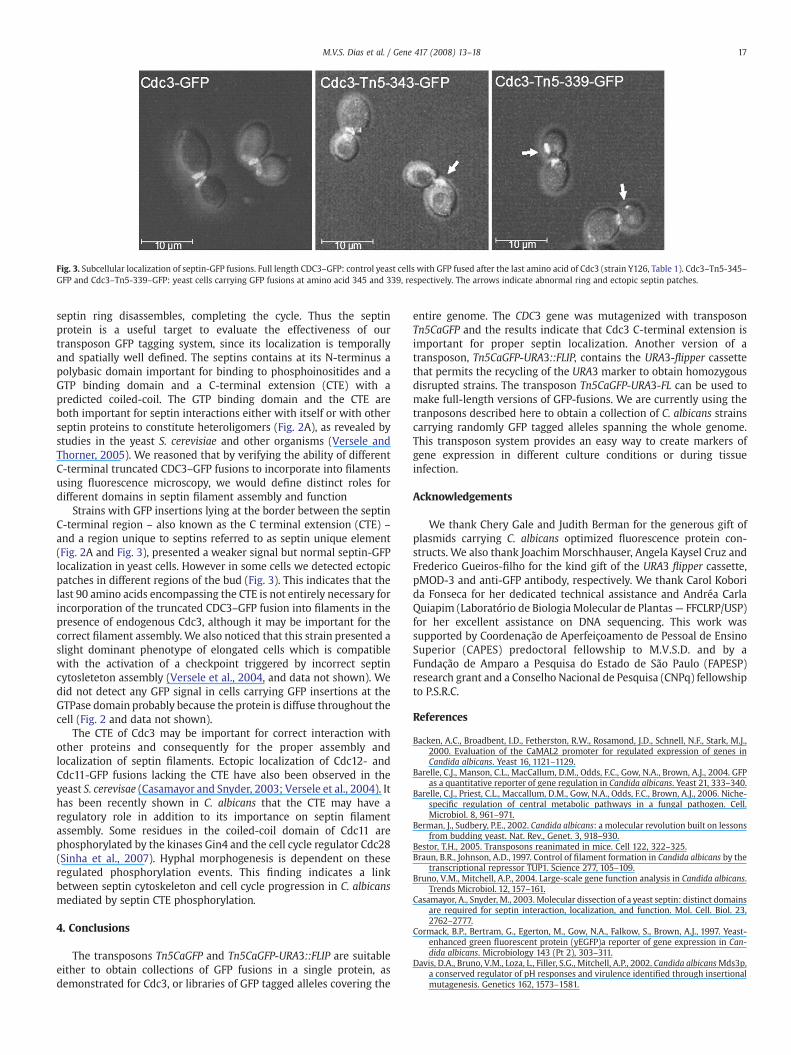
Fig. 3. Subcellular localization of septin-GFP fusions. Full length CDC3–GFP: control yeast cells with GFP fused after the last amino acid of Cdc3 (strain Y126, Table 1). Cdc3–Tn5-345–GFP and Cdc3–Tn5-339–GFP: yeast cells carrying GFP fusions at amino acid 345 and 339, respectively. The arrows indicate abnormal ring and ectopic septin patches.
17M.V.S. Dias et al. / Gene 417 (2008) 13–18
septin ring disassembles, completing the cycle. Thus the septinprotein is a useful target to evaluate the effectiveness of ourtransposon GFP tagging system, since its localization is temporallyand spatially well defined. The septins contains at its N-terminus apolybasic domain important for binding to phosphoinositides and aGTP binding domain and a C-terminal extension (CTE) with apredicted coiled-coil. The GTP binding domain and the CTE areboth important for septin interactions either with itself or with otherseptin proteins to constitute heteroligomers (Fig. 2A), as revealed bystudies in the yeast S. cerevisiae and other organisms (Versele andThorner, 2005). We reasoned that by verifying the ability of differentC-terminal truncated CDC3–GFP fusions to incorporate into filamentsusing fluorescence microscopy, we would define distinct roles fordifferent domains in septin filament assembly and function
Strains with GFP insertions lying at the border between the septinC-terminal region – also known as the C terminal extension (CTE) –and a region unique to septins referred to as septin unique element(Fig. 2A and Fig. 3), presented a weaker signal but normal septin-GFPlocalization in yeast cells. However in some cells we detected ectopicpatches in different regions of the bud (Fig. 3). This indicates that thelast 90 amino acids encompassing the CTE is not entirely necessary forincorporation of the truncated CDC3–GFP fusion into filaments in thepresence of endogenous Cdc3, although it may be important for thecorrect filament assembly. We also noticed that this strain presented aslight dominant phenotype of elongated cells which is compatiblewith the activation of a checkpoint triggered by incorrect septincytosleteton assembly (Versele et al., 2004, and data not shown). Wedid not detect any GFP signal in cells carrying GFP insertions at theGTPase domain probably because the protein is diffuse throughout thecell (Fig. 2 and data not shown).
The CTE of Cdc3 may be important for correct interaction withother proteins and consequently for the proper assembly andlocalization of septin filaments. Ectopic localization of Cdc12- andCdc11-GFP fusions lacking the CTE have also been observed in theyeast S. cerevisae (Casamayor and Snyder, 2003; Versele et al., 2004). Ithas been recently shown in C. albicans that the CTE may have aregulatory role in addition to its importance on septin filamentassembly. Some residues in the coiled-coil domain of Cdc11 arephosphorylated by the kinases Gin4 and the cell cycle regulator Cdc28(Sinha et al., 2007). Hyphal morphogenesis is dependent on theseregulated phosphorylation events. This finding indicates a linkbetween septin cytoskeleton and cell cycle progression in C. albicansmediated by septin CTE phosphorylation.
4. Conclusions
The transposons Tn5CaGFP and Tn5CaGFP-URA3::FLIP are suitableeither to obtain collections of GFP fusions in a single protein, asdemonstrated for Cdc3, or libraries of GFP tagged alleles covering the
entire genome. The CDC3 gene was mutagenized with transposonTn5CaGFP and the results indicate that Cdc3 C-terminal extension isimportant for proper septin localization. Another version of atransposon, Tn5CaGFP-URA3::FLIP, contains the URA3-flipper cassettethat permits the recycling of the URA3 marker to obtain homozygousdisrupted strains. The transposon Tn5CaGFP-URA3-FL can be used tomake full-length versions of GFP-fusions. We are currently using thetranposons described here to obtain a collection of C. albicans strainscarrying randomly GFP tagged alleles spanning the whole genome.This transposon system provides an easy way to create markers ofgene expression in different culture conditions or during tissueinfection.
Acknowledgements
We thank Chery Gale and Judith Berman for the generous gift ofplasmids carrying C. albicans optimized fluorescence protein con-structs. We also thank JoachimMorschhauser, Angela Kaysel Cruz andFrederico Gueiros-filho for the kind gift of the URA3 flipper cassette,pMOD-3 and anti-GFP antibody, respectively. We thank Carol Koborida Fonseca for her dedicated technical assistance and Andréa CarlaQuiapim (Laboratório de Biologia Molecular de Plantas— FFCLRP/USP)for her excellent assistance on DNA sequencing. This work wassupported by Coordenação de Aperfeiçoamento de Pessoal de EnsinoSuperior (CAPES) predoctoral fellowship to M.V.S.D. and by aFundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP)research grant and a Conselho Nacional de Pesquisa (CNPq) fellowshipto P.S.R.C.
References
Backen, A.C., Broadbent, I.D., Fetherston, R.W., Rosamond, J.D., Schnell, N.F., Stark, M.J.,2000. Evaluation of the CaMAL2 promoter for regulated expression of genes inCandida albicans. Yeast 16, 1121–1129.
Barelle, C.J., Manson, C.L., MacCallum, D.M., Odds, F.C., Gow, N.A., Brown, A.J., 2004. GFPas a quantitative reporter of gene regulation in Candida albicans. Yeast 21, 333–340.
Barelle, C.J., Priest, C.L., Maccallum, D.M., Gow, N.A., Odds, F.C., Brown, A.J., 2006. Niche-specific regulation of central metabolic pathways in a fungal pathogen. Cell.Microbiol. 8, 961–971.
Berman, J., Sudbery, P.E., 2002. Candida albicans: a molecular revolution built on lessonsfrom budding yeast. Nat. Rev., Genet. 3, 918–930.
Bestor, T.H., 2005. Transposons reanimated in mice. Cell 122, 322–325.Braun, B.R., Johnson, A.D., 1997. Control of filament formation in Candida albicans by the
transcriptional repressor TUP1. Science 277, 105–109.Bruno, V.M., Mitchell, A.P., 2004. Large-scale gene function analysis in Candida albicans.
Trends Microbiol. 12, 157–161.Casamayor, A., Snyder, M., 2003. Molecular dissection of a yeast septin: distinct domains
are required for septin interaction, localization, and function. Mol. Cell. Biol. 23,2762–2777.
Cormack, B.P., Bertram, G., Egerton, M., Gow, N.A., Falkow, S., Brown, A.J., 1997. Yeast-enhanced green fluorescent protein (yEGFP)a reporter of gene expression in Can-dida albicans. Microbiology 143 (Pt 2), 303–311.
Davis, D.A., Bruno, V.M., Loza, L., Filler, S.G., Mitchell, A.P., 2002. Candida albicansMds3p,a conserved regulator of pH responses and virulence identified through insertionalmutagenesis. Genetics 162, 1573–1581.

18 M.V.S. Dias et al. / Gene 417 (2008) 13–18
Douglas, L.M., Alvarez, F.J., McCreary, C., Konopka, J.B., 2005. Septin function in yeastmodel systems and pathogenic fungi. Eukaryot. Cell 4, 1503–1512.
Enloe, B., Diamond, A., Mitchell, A.P., 2000. A single-transformation gene function test indiploid Candida albicans. J. Bacteriol. 182, 5730–5736.
Firon, A., Villalba, F., Beffa, R., D'Enfert, C., 2003. Identification of essential genes in thehuman fungal pathogen Aspergillus fumigatus by transposon mutagenesis.Eukaryot. Cell 2, 247–255.
Fonzi, W.A., Irwin, M.Y., 1993. Isogenic strain construction and genemapping in Candidaalbicans. Genetics 134, 717–728.
Gerami-Nejad, M., Berman, J., Gale, C.A., 2001. Cassettes for PCR-mediated constructionof green, yellow, and cyan fluorescent protein fusions in Candida albicans. Yeast 18,859–864.
Gillum, A.M., Tsay, E.Y., Kirsch, D.R., 1984. Isolation of the Candida albicans gene fororotidine-5′-phosphate decarboxylase by complementation of S. cerevisiae ura3and E. coli pyrF mutations. Mol. Gen. Genet. 198, 179–182.
Goyard, S., Tosi, L.R., Gouzova, J., Majors, J., Beverley, S.M., 2001. New Mos1 marinertransposons suitable for the recovery of gene fusions in vivo and in vitro. Gene 280,97–105.
Green, C.B., Zhao, X., Hoyer, L.L., 2005. Use of green fluorescent protein and reversetranscription-PCR to monitor Candida albicans agglutinin-like sequence geneexpression in a murine model of disseminated candidiasis. Infect. Immun. 73,1852–1855.
Jones, T., et al., 2004. The diploid genome sequence of Candida albicans. Proc. Natl. Acad.Sci. U. S. A. 101, 7329–7334.
Judson, N., Mekalanos, J.J., 2000. Transposon-based approaches to identify essentialbacterial genes. Trends Microbiol. 8, 521–526.
Kim, H.B., Haarer, B.K., Pringle, J.R., 1991. Cellular morphogenesis in the Saccharomycescerevisiae cell cycle: localization of the CDC3 gene product and the timing of eventsat the budding site. J. Cell Biol. 112, 535–544.
Mao, Y., Zhang, Z., Wong, B., 2003. Use of green fluorescent protein fusions to analysethe N- and C-terminal signal peptides of GPI-anchored cell wall proteins in Candidaalbicans. Mol. Microbiol. 50, 1617–1628.
Merkulov, G.V., Boeke, J.D., 1998. Libraries of green fluorescent protein fusionsgenerated by transposition in vitro. Gene 222, 213–222.
Morschhauser, J., Michel, S., Hacker, J., 1998. Expression of a chromosomally integrated,single-copy GFP gene in Candida albicans, and its use as a reporter of generegulation. Mol. Gen. Genet. 257, 412–420.
Morschhauser, J., Michel, S., Staib, P., 1999. Sequential gene disruption in Candidaalbicans by FLP-mediated site-specific recombination. Mol. Microbiol. 32, 547–556.
Nobile, C.J., Mitchell, A.P., 2005. Regulation of cell-surface genes and biofilm formationby the C. albicans transcription factor Bcr1p. Curr. Biol. 15, 1150–1155.
Noble, S.M., Johnson, A.D., 2007. Genetics of Candida albicans, a diploid human fungalpathogen. Annu. Rev. Genet. 41, 193–211.
Nobile, C.J., Bruno, V.M., Richard, M.L., Davis, D.A., Mitchell, A.P., 2003. Genetic control ofchlamydospore formation in Candida albicans. Microbiology 149, 3629–3637.
Pfaller, M.A., Diekema, D.J., 2007. Epidemiology of invasive candidiasis: a persistentpublic health problem. Clin. Microbiol. Rev. 20, 133–163.
Reznikoff, W.S., 2006. Tn5 transposition: a molecular tool for studying proteinstructure-function. Biochem. Soc. Trans. 34, 320–323.
Reznikoff, W.S., Goryshin, I.Y., Jendrisak, J.J., 2004. Tn5 as a molecular genetics tool: Invitro transposition and the coupling of in vitro technologies with in vivotransposition. Methods Mol. Biol. 260, 83–96.
Ross-Macdonald, P., Sheehan, A., Roeder, G.S., Snyder, M., 1997. A multipurposetransposon system for analyzing protein production, localization, and function inSaccharomyces cerevisiae. Proc. Natl. Acad. Sci. U. S. A. 94, 190–195.
Ross-Macdonald, P., et al., 1999. Large-scale analysis of the yeast genome by transposontagging and gene disruption. Nature 402, 413–418.
Schaub, Y., Dunkler, A., Walther, A., Wendland, J., 2006. New pFA-cassettes for PCR-based gene manipulation in Candida albicans. J. Basic Microbiol. 46, 416–429.
Sinha, I., Wang, Y.M., Philp, R., Li, C.R., Yap, W.H., Wang, Y., 2007. Cyclin-dependentkinases control septin phosphorylation in Candida albicans hyphal development.Dev. Cell 13, 421–432.
Sudbery, P.E., 2001. The germ tubes of Candida albicans hyphae and pseudohyphaeshow different patterns of septin ring localization. Mol. Microbiol. 41, 19–31.
Sudbery, P., Gow, N., Berman, J., 2004. The distinct morphogenic states of Candidaalbicans. Trends Microbiol. 12, 317–324.
Uhl, M.A., Biery, M., Craig, N., Johnson, A.D., 2003. Haploinsufficiency-based large-scaleforward genetic analysis of filamentous growth in the diploid human fungalpathogen C. albicans. Embo J. 22, 2668–2678.
Versele, M., Thorner, J., 2005. Some assembly required: yeast septins provide theinstruction manual. Trends Cell Biol. 15, 414–424.
Versele, M., et al., 2004. Protein–protein interactions governing septin heteropentamerassembly and septin filament organization in Saccharomyces cerevisiae. Mol. Biol.Cell 15, 4568–4583.
Warenda, A.J., Konopka, J.B., 2002. Septin function in Candida albicans morphogenesis.Mol. Biol. Cell 13, 2732–2746.




















