
NMR petrophysical cross correlations for partially saturated reservoir ...
Upload
independentCategory
view
1download
0
Partially functional Cenpa^GFP fusion protein causes increasedchromosome missegregation and apoptosis during mouse embryogenesis
Paul Kalitsis, Kerry J. Fowler, Elizabeth Earle, Belinda Grif¢ths, Emily Howman,Ainsley J. Newson & K. H. Andy Choo*Murdoch Children’s Research Institute, Royal Children’s Hospital, Flemington Road, Parkville 3052,Melbourne, Victoria, Australia; Tel: 61-3-8341 6306; Fax: 61-3-9348 1391;E-mail: [email protected]*Correspondence
Received 31 January 2003. Received in revised form and accepted for publication by Herbert Macgregor 10 March 2003
Key words: aneuploidy, apoptosis, CENP-A, centromere, chromatin
Abstract
CENP-A is an essential histone H3-like protein that localizes to the centromeric region of eukaryoticchromosomes. Heterozygous and homozygous Cenpa^GFP fusion-protein mouse mutants, generatedthrough targeted insertion of the green £uorescent protein (GFP) gene into the mouse Cenpa gene locus,show speci¢c localized £uorescence at all the centromeres. Heterozygous mice are healthy and fertile.Cenpa^GFP homozygotes (Cenpag/g) undergo many cell divisions, giving rise to up to one million cellsthat show relatively accurate differentiation into distinct mouse embryonic tissues until day 10.5 whensigni¢cant levels of chromosome missegregation, aneuploidy and apoptosis result in death. Cenpag/g
embryos assemble functional kinetochores that bind to a host of centromere-speci¢c structural and mitoticspindle checkpoint proteins (Cenpc, BubR1, Mad2 and Zw10). Examination of the nucleosomal phasingof centromeric minor and pericentromeric major satellite sequences indicates that the formation ofCenpag/g homotypic nucleosomes is not accompanied by any overt alteration to the overall size of themonomeric nucleosomal structure or the spacing of these structures. This study provides the ¢rst exampleof an essential centromeric protein gene variant in which subtle perturbation at the centromeric nucleo-somal/chromatin level manifests in a signi¢cantly delayed lethality when compared with Cenpa null mice.
Introduction
The centromere is a DNA-protein structure that isresponsible for the accurate segregation of chro-mosomes during eukaryotic cell division. Theposition of the centromere along a chromosome isde¢ned by a group of constitutive centromereproteins (Choo 2001, Sullivan et al. 2001). Theseproteins act as a foundation layer that is present at
the centromeric locus throughout the cell cycle,commonly known as the prekinetochore. In ver-tebrate centromeres, these proteins include thehistone H3-like CENP-A (Sullivan 2001), thehighly basic CENP-C (Saitoh et al. 1992), CENP-H(Sugata et al. 2000) and the Mis6 orthologue,CENP-I (Nishihashi et al. 2002). Data gainedfrom genetic and immunodepletion experiments inmouse, Drosophila and C. elegans are starting to
Chromosome Research 11: 345^357, 2003. 345# 2003 Kluwer Academic Publishers. Printed in the Netherlands

reveal a centromere/kinetochore assembly path-way (Blower & Karpen 2001, Howman et al. 2000,Oegema et al. 2001). CENP-A most likely formsthe initial epigenetic mark upon which subsequentcentromeric proteins are assembled, including thetransient or passenger proteins. The resultingcentromere/kinetochore structure provides essen-tial functions for the accurate segregation ofchromosomes, including mitotic spindle captureand checkpoint control, sister centromere cohe-sion and release, and physical movement ofchromosomes to opposite poles.CENP-A is the most widely studied and con-
served member of the constitutive centromereproteins. This protein is thought to replace histoneH3 in its role in dictating the formation of centro-mere-speci¢c chromatin (Lo et al. 2001, Yodaet al. 2000). Genetic knockout or depletion studieshave shown that CENP-A is essential for celldivision, resulting in the impaired binding ofdownstream centromere proteins, severe chro-mosome missegregation, and rapid arrest of thecell cycle (Stoler et al. 1995, Figueroa et al. 1998,Howman et al. 2000, Blower & Karpen 2001,Oegema et al. 2001).The green £uorescent protein (GFP) from the
jelly¢sh Aequorea victoria has been widely used tovisualize proteins in the context of their naturalcellular environment, either in vivo or in vitro (forreview, see Tsien 1998). It has also been exploitedin the study of chromosome dynamics during thecell cycle. Chromosomal and centromere proteinshave been fused to GFP to allow their spatial andtemporal organization to be visualized in cellculture (Belmont 2001). To our knowledge, nocentromere-speci¢c GFP fusion proteins to datehave been expressed in whole multicellularorganisms. In this report, we have fused GFP tothe carboxyl end of CENP-A and expressed itunder the control of the natural CENP-A pro-moter in mice. We describe the e¡ects the fusionprotein has on centromere function and mitoticintegrity during mouse development.
Materials and methods
Generation of targeting construct
The mouse Cenpa gene was isolated from amouse 129/Sv genomic bacteriophage library as
described previously (Kalitsis et al. 1998b). Thetargeting construct contained 3.6 kb of 50 Cenpasequence including exons 2, 3 and 4 (up to the stopcodon), and 3.7 kb of 30 sequence including exon 5.PCR ampli¢cation was used to replace the stopcodon and place the GFP and Cenpa codingsequences in frame. An IRES^neomycin fusioncassette was introduced downstream of the GFP,replacing the segment between exon 4 and theXhoI site in exon 5 (see Figure 1A-i and -ii). Thise¡ectively placed the promoterless IRES^neomycin cassette downstream of GFP codingsequence to facilitate tissue culture selection forcorrect targeting events. The 8 amino acid peptidelinker between Cenpa and GFP is GIHRPVAT.The Cenpa^GFP fusion polypeptide is 381 aminoacids in length with a predicted molecular weightof 43.2 kD.
Gene targeting
Mouse embryonic stem (ES) cell culture andmanipulations were carried out using standardprocedures. For the creation of heterozygous celllines, 50 mg of linearized DNA was electro-porated at 0.8 kV, 3 mF, 1 O into 5� 107 mouse129/1 or W9.5 ES cells. The cells were selectedwith either G418 (Gibco-BRL) or hygromycin(Roche), at 200 mg/ml, or 120 mg/ml, respec-tively. Correctly targeted events were detectedusing Southern blot hybridization. A 400-bpSphI-BamHI fragment was used as a 50-externalprobe in the hybridization assay (Figure 1A, B).Genomic DNA was digested with SphI, gene-rating a 10-kb wild-type or 7-kb recombinantband. A 512-bp 30-external probe was generatedusing oligonucleotide primers, CENA30O (50-CCATCCTTGACTGACTGAGATTGG-30) andCENA30P (50-CATACAGACACACACACA-GAGGTGG-30). These primers were identi¢edfrom the Celera mouse genomic database.Genomic DNA was digested using KpnI, pro-ducing a 10-kb wild type or 8-kb recombinantbands (Figure 1A, C).
Mouse breeding
Targeted ES cell lines were injected into C57BL/6blastocysts by standard methods. The injectedblastocysts were then transferred into recipient
346 P. Kalitsis et al.
pseudopregnant mice HSD Ola (Gpi-1bb). Chi-meric mice were selected by coat colour andmated with C57BL/6 mice to generate hetero-zygotes. Progeny from chimeric and heterozygouscrosses were genotyped as described below.The heterozygous mice were crossed to obtainhomozygotes.
Genotyping of mice
Mouse tail DNA was extracted using standardtechniques. For PCR genotyping, a duplexstrategy was employed using the followingprimers: CENAI4G (50-GGAGCTTCACGCTT-GTAAGC-30) and CENAI4H (50- AGCACAA-
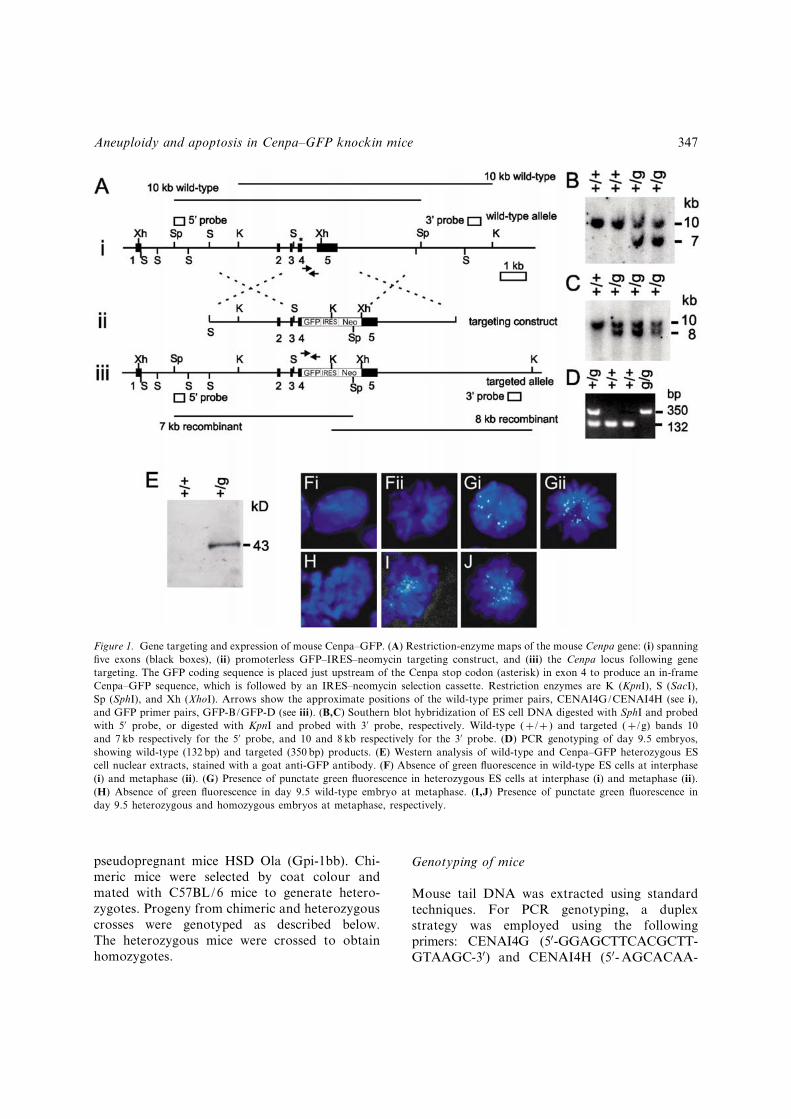
Figure 1. Gene targeting and expression of mouse Cenpa^GFP. (A) Restriction-enzyme maps of the mouse Cenpa gene: (i) spanning¢ve exons (black boxes), (ii) promoterless GFP^IRES^neomycin targeting construct, and (iii) the Cenpa locus following genetargeting. The GFP coding sequence is placed just upstream of the Cenpa stop codon (asterisk) in exon 4 to produce an in-frameCenpa^GFP sequence, which is followed by an IRES^neomycin selection cassette. Restriction enzymes are K (KpnI), S (SacI),Sp (SphI), and Xh (XhoI). Arrows show the approximate positions of the wild-type primer pairs, CENAI4G/CENAI4H (see i),and GFP primer pairs, GFP-B/GFP-D (see iii). (B,C) Southern blot hybridization of ES cell DNA digested with SphI and probedwith 50 probe, or digested with KpnI and probed with 30 probe, respectively. Wild-type (þ/þ ) and targeted (þ/g) bands 10and 7 kb respectively for the 50 probe, and 10 and 8 kb respectively for the 30 probe. (D) PCR genotyping of day 9.5 embryos,showing wild-type (132 bp) and targeted (350 bp) products. (E) Western analysis of wild-type and Cenpa^GFP heterozygous EScell nuclear extracts, stained with a goat anti-GFP antibody. (F) Absence of green £uorescence in wild-type ES cells at interphase(i) and metaphase (ii). (G) Presence of punctate green £uorescence in heterozygous ES cells at interphase (i) and metaphase (ii).(H) Absence of green £uorescence in day 9.5 wild-type embryo at metaphase. (I,J) Presence of punctate green £uorescence inday 9.5 heterozygous and homozygous embryos at metaphase, respectively.
Aneuploidy and apoptosis in Cenpa^GFP knockin mice 347

CTCATCCGTTCAC-30) for the wild-type allele(132 bp), and GFP-B (50-ATGCCGTTCTTCT-GCTTGTC-30) and GFP-D (50-AAGTTCAT-CTGCACCACCG-30) for the Cenpa-GFP allele(350 bp) (Figure 1A, D). For day 9.5 embryos,DNA from the yolk sac was extracted andresuspended in 20 ml of TE. One ml was used in aPCR reaction with FastStart DNA polymerase(Roche).
Cytogenetics
Day 9.5 embryos were mechanically disruptedusing scalpel blades and then syringed through a¢ne needle. The cells were then put into DMEMcontaining 10% fetal calf serum and incubatedwith 100 mg/ml colcemid (Gibco BRL) for 2 h.After the incubation, the cells were treated with0.56% KCl for 10min and then ¢xed in acetic^methanol (1 : 3). Chromosomes were stained withGiemsa.
Immuno£uorescence
For antibody analyses the colcemid-treated cellswere spun onto microscope slides using a cytospincentrifuge (Shandon) at 1000 rpm for 5min. Thecentrifuged cells were permeablized in KCM(120mmol/L KCl; 20mmol/L NaCl; 10mmol/LTris^HCl; 0.5mmol/L NaEDTA; 0.1% (v/v)Triton X-100) for 10min. Primary antibodieswere diluted in KCM and incubated on the cellpreparations for 1 h at 37�C and then rinsed threetimes in KCM. Fluorescently conjugated sec-ondary antibodies were used according to thesupplier’s instructions. Antibodies were thenrinsed o¡ the slides with KCM, ¢xed in 10%formalin and then mounted in Vectashield(Vector Laboratories) containing DAPI. Imageswere captured using an Axioplan2 microscope(Zeiss), with a Sensys-cooled CCD camera(Photometrics) and processed with IP Lab soft-ware (Scanalytics). Antibodies used in this studyincluded anti-mouse Cenpa (Lo et al. 2001), anti-mouse Cenpc (Sa¡ery et al. 1999), anti-humanBUBR1 (Jablonski et al. 1998), anti-XenopusMad2 (Waters et al. 1998) and anti-human ZW10(Starr et al. 1997).
Embryo histology and TUNEL staining
Whole embryos were ¢xed in 4% paraformalde-hyde and subsequently embedded in para⁄n.Embryo sections were cut at 5 mm and stained withDAPI or haematoxylin and eosin. TUNEL assayswere performed on embryos as described inGavrieli et al. 1992.
Western analysis
Equal amounts of nuclear extracts of parentalwild-type and Cenpa^GFP ES cell lines were runon 15% SDS-PAGE gels and probed with a goatanti-GFP antibody (Rockland) using standardprocedures.
Chromatin extraction for day 9.5 embryos
Day 9.5 embryos were frozen at �70�C in 10%DMSO/fetal calf serum whilst the yolk sac DNAwas genotyped. Chromatin was extracted from 10wild-type, 10 þ/g, and 25 g/g day-9.5 pooledembryos. This method was a variation of thechromatin extraction procedure in Lo et al. 2001.Brie£y, pooled embryos were rinsed three times inice-cold PBS and were incubated in TBS(0.01mol/L Tris-HCl pH 7.5, 3mmol/L CaCl2,2mmol/L MgCl2) with 0.1mmol/L phenylmethyl-sulphonyl £uoride (PMSF) and proteinase inhibi-tors (Complete, Proteinase Inhibitor CocktailTablet, Roche) with 0.25% Tween 40 at 4�C on aroller stirrer for 1 h before extruding the nucleiusing 30 strokes with the Tight or A pestle on aDounce homogenizor (Wheaton). Nuclei wereseparated from cytoplasmic debris by cen-trifugation at 1500 g for 20min at 4�C through a25%/50% discontinuous sucrose gradient. Par-tially digested nucleosomes were produced by in-cubating nuclei with micrococcal nuclease (USB)in digestion bu¡er (0.32mol/L sucrose, 50mmol/LTris-HCl at pH 7.5, 4mmol/L MgCl2, 1mmol/LCaCl2, 0.1mmol/L PMSF) at a concentration of0.5 or 0.1 units/10 mg of DNA at 37�C for 20min.The digestion mix was then stopped by adding0.5mol/L EDTA, and puri¢ed by extracting withphenol^chloroform, chloroform and ethanolprecipitation. TE resuspended samples were runon a 2% gel for Southern blot hybridization.
348 P. Kalitsis et al.

Mouse major and minor satellite probe isolation
Mouse major satellite DNAwas isolated by partialdigestion of total genomic DNA with MnlI. The235-bp monomer band was cut out of an agarosegel and cloned into pBluescript. Mouse minorsatellite DNA was puri¢ed from genomic DNA byPCR ampli¢cation using primers, MINa (50-AATGATAAAAACCACACTG-30) and MINb(50- TTCTCATTGTAACTCATTG-30) derived fromthe minor satellite consensus sequence (Wong &Rattner 1988). Multimers of the mouse minorrepeating unit were gel puri¢ed and cloned intopGEM-T (Promega). Clones were subsequentlyveri¢ed by DNA sequencing.
Results and discussion
Cenpa^GFP localizes to centromeresin heterozygous ES cells and mice
In order to achieve an accurate spatial and tem-poral expression pattern for the Cenpa^GFPfusion protein that was comparable to that ofendogenous Cenpa, we designed a gene-targetingconstruct that allowed the fusion protein to beexpressed under the control of the endogenousmouse Cenpa promoter. Twenty-three out of 80(29%) targeted heterozygous Cenpa^GFP ES-celllines were identi¢ed by Southern blot analysis(Figure 1B, C). Western blot analysis of thenuclear extracts of these cell lines using a goat anti-GFP antibody showed expression of the expected43-kDa band (Figure 1E). These cells gave distinctgreen £uorescent signals at the centromerethroughout the cell cycle (Figure 1G; comparedwith the absence of similar signals on wild-typecells in Figure 1F). The green £uorescent signalscolocalized with a CREST antiserum that recog-nizes CENP-A and CENP-B (not shown). Thepresence of the Cenpa-GFP fusion protein did notappear to exert any measurable e¡ects on thegrowth of the ES cells, since these cells continuedto divide at normal rates and contained stablekaryotypes.Standard blastocyst injection methods were
used to generate chimeric mice, which were used toproduce heterozygous Cenpa^GFP animals(Cenpaþ/g). The heterozygous mice were healthy,
fertile and normal, showing no signs of a¡ectedgrowth rates or defective cell division. The loca-lization of the GFP £uorescence in heterozygousembryos coincided with the centromeres in allstages of the cell cycle as seen in the ES cells (anexample of a cell in metaphase is shown in Figure1I; compared with the absence of similar signals ona wild-type embryo in Figure 1H).
Homozygous Cenpa^GFP mice show delayeddevelopment and survive until day 10.5
In order to study the functional e¡ects of theCenpa^GFP fusion protein in the absence of wild-type copies of Cenpa, we intercrossed hetero-zygous mice to produce homozygotes (Cenpag/g)where both copies of the wild-type Cenpa genewere replaced by Cenpa-GFP. A cohort of 88 pupsfrom heterozygous matings was genotyped byPCR (Figure 1D). Thirty-one were (þ/þ ), 57(þ/g) and 0 (g/g), indicating that the Cenpag/g
mice died during embryo development.We next investigated earlier embryonic stages to
de¢ne the time and characteristics of lethality. Atday 9.5, the normal mouse embryo has changedfrom the ‘S’-shape to the ‘C’-shape and is vas-cularized with a beating heart (Figure 2A). Incomparison, surviving Cenpag/g embryos showeddevelopmental delay and varied from approxi-mately 1/10 to 1/3 the size of a normal embryo(Figure 2B, C). Structures like the branchial arch,heart, neural tube and forebrain can be identi¢edin most of the a¡ected embryos but showabnormal morphology (Figure 2B, G). Genotypeanalysis of day 9.5 embryos taken from severallitters revealed 11 (þ/þ ), 21 (þ/g) and 12 (g/g),approximating the expected Mendelian ratio.Genotyping of day-10.5 embryos gave 14 (þ/þ ),28 (þ/g) and 6 (g/g), indicating a signi¢cantdecline in the number of surviving Cenpag/g
embryos. By day 12.5, Cenpag/g embryos were nolonger detected.Para⁄n-embedded sections of day 9.5 embryos
were stained with DAPI to allow the investigationof nuclear morphology. Normal embryos showedregularly shaped interphase nuclei with congres-sing metaphase chromosomes (Figure 3A). Incontrast, the Cenpag/g embryos contained a highproportion of irregularly shaped interphase nuclei,
Aneuploidy and apoptosis in Cenpa^GFP knockin mice 349
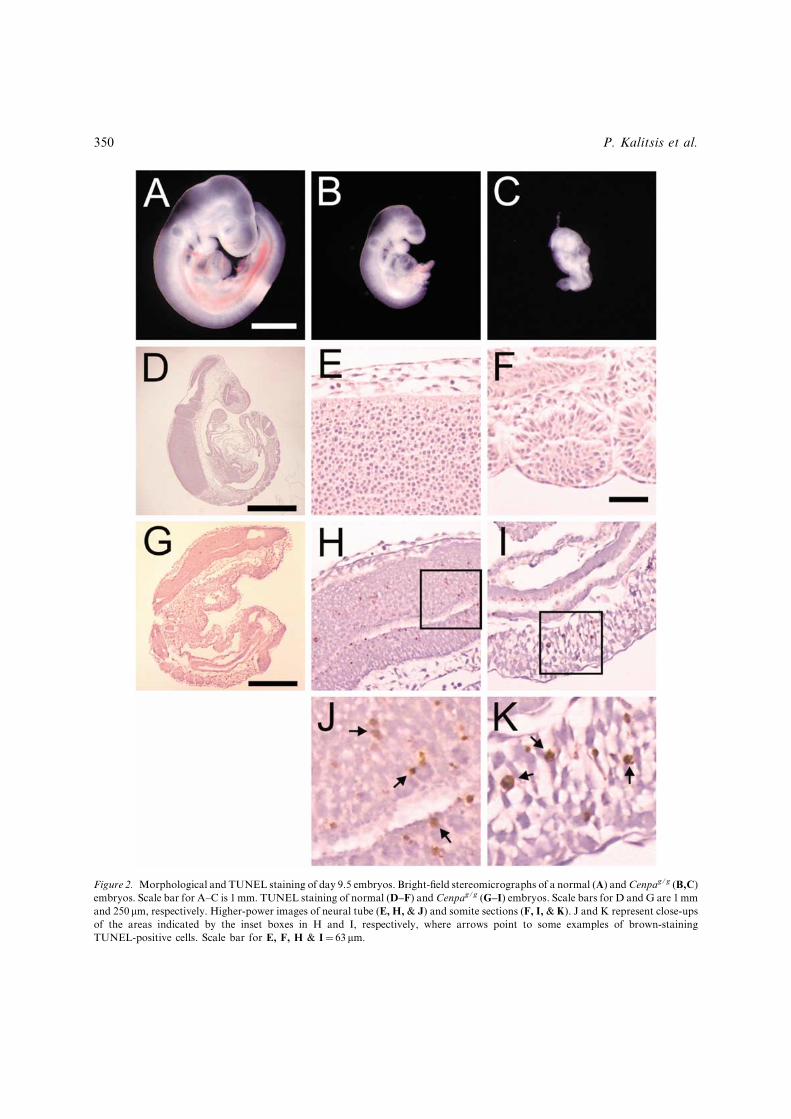
Figure 2. Morphological and TUNEL staining of day 9.5 embryos. Bright-¢eld stereomicrographs of a normal (A) andCenpag/g (B,C)embryos. Scale bar for A^C is 1mm. TUNEL staining of normal (D^F) and Cenpag/g (G^I) embryos. Scale bars for D and G are 1mmand 250 mm, respectively. Higher-power images of neural tube (E, H, & J) and somite sections (F, I, & K). J and K represent close-upsof the areas indicated by the inset boxes in H and I, respectively, where arrows point to some examples of brown-stainingTUNEL-positive cells. Scale bar for E, F, H & I¼ 63 mm.
350 P. Kalitsis et al.
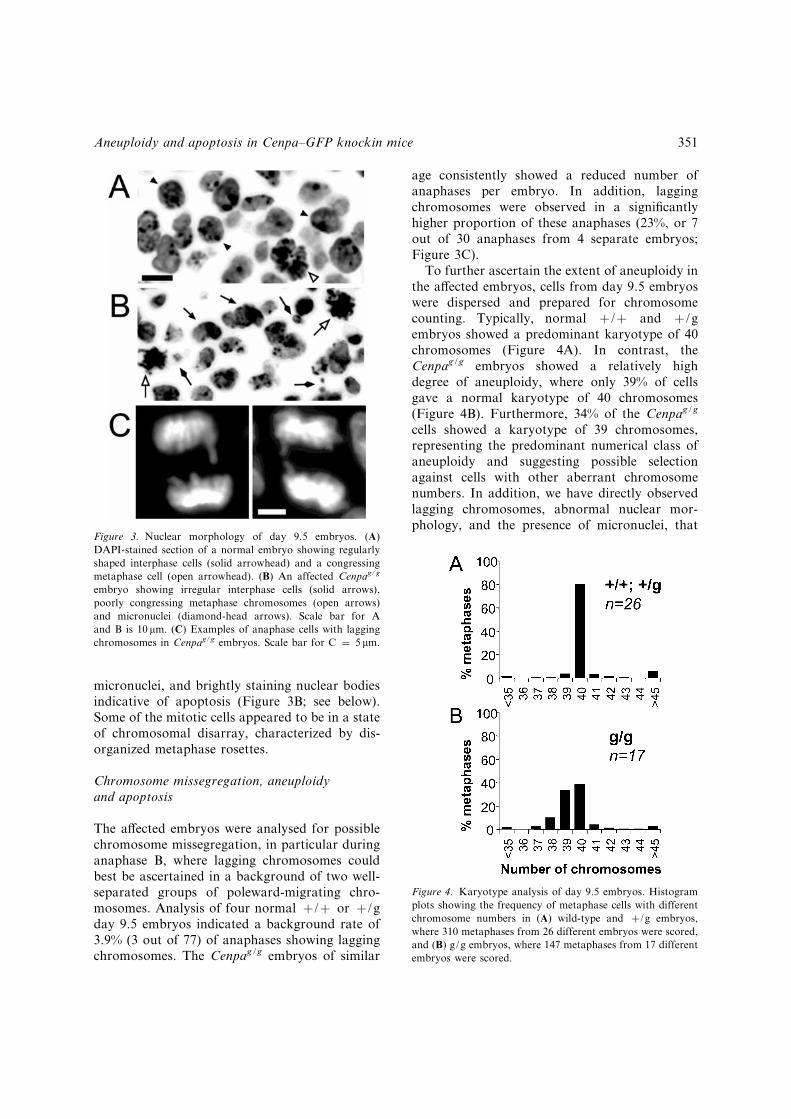
micronuclei, and brightly staining nuclear bodiesindicative of apoptosis (Figure 3B; see below).Some of the mitotic cells appeared to be in a stateof chromosomal disarray, characterized by dis-organized metaphase rosettes.
Chromosome missegregation, aneuploidyand apoptosis
The a¡ected embryos were analysed for possiblechromosome missegregation, in particular duringanaphase B, where lagging chromosomes couldbest be ascertained in a background of two well-separated groups of poleward-migrating chro-mosomes. Analysis of four normal þ/þ or þ/gday 9.5 embryos indicated a background rate of3.9% (3 out of 77) of anaphases showing laggingchromosomes. The Cenpag/g embryos of similar
age consistently showed a reduced number ofanaphases per embryo. In addition, laggingchromosomes were observed in a signi¢cantlyhigher proportion of these anaphases (23%, or 7out of 30 anaphases from 4 separate embryos;Figure 3C).
To further ascertain the extent of aneuploidy inthe a¡ected embryos, cells from day 9.5 embryoswere dispersed and prepared for chromosomecounting. Typically, normal þ/þ and þ/gembryos showed a predominant karyotype of 40chromosomes (Figure 4A). In contrast, theCenpag/g embryos showed a relatively highdegree of aneuploidy, where only 39% of cellsgave a normal karyotype of 40 chromosomes(Figure 4B). Furthermore, 34% of the Cenpag/g
cells showed a karyotype of 39 chromosomes,representing the predominant numerical class ofaneuploidy and suggesting possible selectionagainst cells with other aberrant chromosomenumbers. In addition, we have directly observedlagging chromosomes, abnormal nuclear mor-phology, and the presence of micronuclei, that
Figure 3. Nuclear morphology of day 9.5 embryos. (A)DAPI-stained section of a normal embryo showing regularlyshaped interphase cells (solid arrowhead) and a congressingmetaphase cell (open arrowhead). (B) An affected Cenpag/g
embryo showing irregular interphase cells (solid arrows),poorly congressing metaphase chromosomes (open arrows)and micronuclei (diamond-head arrows). Scale bar for Aand B is 10 mm. (C) Examples of anaphase cells with laggingchromosomes in Cenpag/g embryos. Scale bar for C ¼ 5 mm.
Figure 4. Karyotype analysis of day 9.5 embryos. Histogramplots showing the frequency of metaphase cells with differentchromosome numbers in (A) wild-type and þ/g embryos,where 310 metaphases from 26 different embryos were scored,and (B) g/g embryos, where 147 metaphases from 17 differentembryos were scored.
Aneuploidy and apoptosis in Cenpa^GFP knockin mice 351

were not typically detected in the wild-type andheterozygous embryos (Figure 3B, C). Theseresults therefore point to a compromised cen-tromere function in the a¡ected embryos leadingto an overall increased level of chromosomemissegregation.To con¢rm whether the brightly DAPI-staining
or pyknotic nuclei seen in Figure 3B wereapoptotic, we performed TUNEL staining onnormal and a¡ected embryos. Normal day 9.5embryos showed very few positively staining cells(Figure 2D^F). In contrast, the Cenpag/g embryosshowed an increase in the number of TUNEL-staining cells indicating a signi¢cant elevation inapoptosis (Figure 2G^K). The average number ofapoptotic cells observed in the Cenpag/g embryoswas 3.9%, compared with 0.1% positivelyTUNEL-stained cells for either the wild-type orheterozygous embryos. This increased level ofapoptosis was observed throughout the embryoand was not con¢ned to speci¢c tissue types(Figures 2H & J and 2I & K show examples for theneural tube and somites, respectively).Apoptosis is de¢ned as a programmed cell
death, where the cell undergoes a proteolyticcascade following genetic, developmental orchemical triggers. The relationship betweenapoptosis and chromosome missegregation withrespect to centromere function is poorly under-stood. To date, there have been no reports linkingCENP-A dysfunction with apoptosis probablybecause the null mutants result in catastrophicmitotic disarray that renders studies di⁄cult(Howman et al. 2000, Blower & Karpen, 2001,Oegema et al. 2001). We have shown that alteringthe Cenpa protein through GFP fusion results in amore subtle form of mutation that, while stillsupporting mitotic divisions, results in an eventualincrease in the level of apoptosis in the Cenpag/g
embryos. In this case, apoptosis could be caused byincreasingly defective centromeres leading toaberrant microtubule binding and prolongedmitotic arrest by spindle checkpoints. Alter-natively, cells could enter into the next cell cycleafter chromosomal missegregation, whereupon thedaughter cells with abnormal chromosome num-bers would be prevented from progressing furtherthrough the cycle by checkpoint mechanisms. Anincreased level of apoptosis has also been observedin a conditional knockout of a constitutive
centromere protein gene CENP-C in chicken and anull mutation of the Mad2 mitotic spindlecheckpoint gene in mouse (Fukagawa & Brown,1997, Dobles et al. 2000). Together, these studiesindicate that perturbation of key centromereprotein functions, whether through complete geneknockout or subtle structural changes, can induceapoptosis.
Normal binding of centromere proteinsin Cenpag=g cells
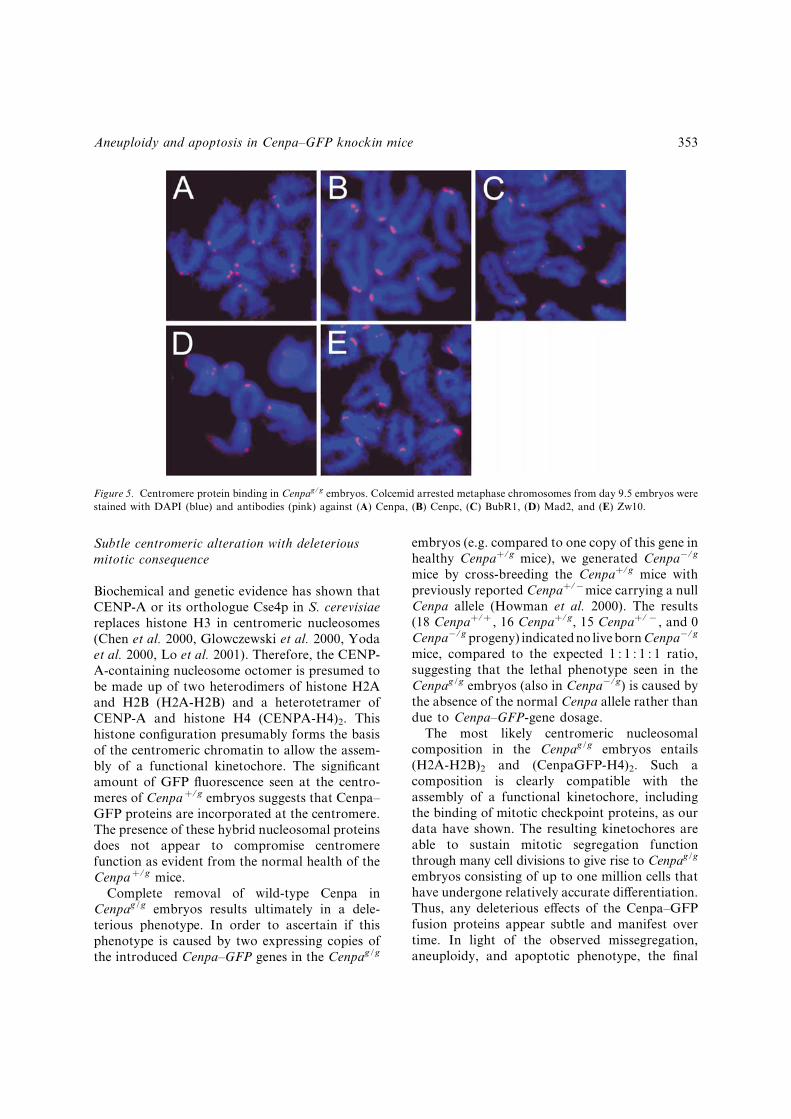
In view of the observed chromosomal mis-segregation phenotype, we were interested toknow if centromere protein binding properties inthe Cenpag/g embryos were normal. The speci¢cbinding of Cenpa^GFP proteins at the cen-tromeres of the Cenpag/g embryos throughoutthe cell cycle was demonstrated by £uorescenceof the GFP (an example is shown in Figure 1J).This binding was con¢rmed using an anti-mouse-Cenpa antibody (Figure 5A). In addition, wehave employed a panel of antibodies againstdi¡erent centromere proteins to further examinewhether the binding of other centromere proteinswas a¡ected. In previous studies, binding of theconstitutive centromere protein CENP-C hasbeen shown to be dependent on the presence ofCENP-A (Howman et al. 2000, Oegema et al.2001, Van Hooser et al. 2001). Direct testing ofcells from day 9.5 Cenpag/g embryos using ananti-Cenpc antibody showed similar levels ofimmuno£uorescence to those of wild-type orheterozygous embryos, suggesting that Cenpcbinding in the Cenpag/g embryos was normal(Figure 5B). Since previous studies have shownthat the mitotic checkpoint is active duringnormal mouse embryonic development (Dobleset al. 2000, Kalitsis et al. 2000), we furtherdetermined if binding of centromere checkpointproteins was a¡ected using antibodies againstBubR1, Mad2 and Zw10 (Starr et al. 1997,Jablonski et al. 1998, Waters et al. 1998). Allthree proteins were shown to bind with normalintensity in the day-9.5 Cenpag/g cells (Figure5C^E). These results indicate that the kine-tochore assembly pathway and at least somecomponents of the mitotic checkpoint were nota¡ected at levels detectable by light microscopy.
352 P. Kalitsis et al.
Subtle centromeric alteration with deleteriousmitotic consequence
Biochemical and genetic evidence has shown thatCENP-A or its orthologue Cse4p in S. cerevisiaereplaces histone H3 in centromeric nucleosomes(Chen et al. 2000, Glowczewski et al. 2000, Yodaet al. 2000, Lo et al. 2001). Therefore, the CENP-A-containing nucleosome octomer is presumed tobe made up of two heterodimers of histone H2Aand H2B (H2A-H2B) and a heterotetramer ofCENP-A and histone H4 (CENPA-H4)2. Thishistone con¢guration presumably forms the basisof the centromeric chromatin to allow the assem-bly of a functional kinetochore. The signi¢cantamount of GFP £uorescence seen at the centro-meres of Cenpaþ/g embryos suggests that Cenpa^GFP proteins are incorporated at the centromere.The presence of these hybrid nucleosomal proteinsdoes not appear to compromise centromerefunction as evident from the normal health of theCenpaþ/g mice.Complete removal of wild-type Cenpa in
Cenpag/g embryos results ultimately in a dele-terious phenotype. In order to ascertain if thisphenotype is caused by two expressing copies ofthe introduced Cenpa^GFP genes in the Cenpag/g
embryos (e.g. compared to one copy of this gene inhealthy Cenpaþ/g mice), we generated Cenpa�/g
mice by cross-breeding the Cenpaþ/g mice withpreviously reported Cenpaþ/�mice carrying a nullCenpa allele (Howman et al. 2000). The results(18 Cenpaþ/þ , 16 Cenpaþ/g, 15 Cenpaþ/� , and 0Cenpa�/gprogeny) indicatedno live bornCenpa�/g
mice, compared to the expected 1 : 1 : 1 : 1 ratio,suggesting that the lethal phenotype seen in theCenpag/g embryos (also in Cenpa�/g) is caused bythe absence of the normal Cenpa allele rather thandue to Cenpa^GFP-gene dosage.The most likely centromeric nucleosomal
composition in the Cenpag/g embryos entails(H2A-H2B)2 and (CenpaGFP-H4)2. Such acomposition is clearly compatible with theassembly of a functional kinetochore, includingthe binding of mitotic checkpoint proteins, as ourdata have shown. The resulting kinetochores areable to sustain mitotic segregation functionthrough many cell divisions to give rise to Cenpag/g
embryos consisting of up to one million cells thathave undergone relatively accurate di¡erentiation.Thus, any deleterious e¡ects of the Cenpa^GFPfusion proteins appear subtle and manifest overtime. In light of the observed missegregation,aneuploidy, and apoptotic phenotype, the ¢nal
Figure 5. Centromere protein binding in Cenpag/g embryos. Colcemid arrested metaphase chromosomes from day 9.5 embryos werestained with DAPI (blue) and antibodies (pink) against (A) Cenpa, (B) Cenpc, (C) BubR1, (D) Mad2, and (E) Zw10.
Aneuploidy and apoptosis in Cenpa^GFP knockin mice 353

lethal outcome in the older embryos can best beexplained by the gradual accumulation of mitoticerrors, a¡ecting embryonic development andultimately reaching a level that becomes incom-patible with life.We and others have previously described mouse
knockouts for a number of other essential con-stitutive and transient centromere-protein genesCenpc, Incenp, Mad2, Cenpa, Bub3, Survivin andCenpe (Kalitsis et al. 1998a, Cutts et al. 1999,Dobles et al. 2000, Howman et al. 2000, Kalitsiset al. 2000, Uren et al. 2000, Putkey et al. 2002). Ineach case, early embryonic lethality was observedwhere the null embryos completely failed todevelop beyond the implantation stage and dis-integrated after a period of limited mitotic cap-ability conferred presumably by the maternalRNA pools. In this respect, the Cenpag/g embryosrepresent the ¢rst example of a genetic hypomorphof Cenpa in which subtle changes at the cen-tromeric nucleosomal or chromatin level result ina signi¢cantly delayed embryonic lethality whencompared to Cenpa null embryos and other earlylethal knockout embryos.Our Cenpa^GFP fusion protein design involves
the attachment of GFP to the C-terminus ofCenpa. The resulting 43-kDa fusion protein isrelatively bulky compared with the 17-kDa nativemouse Cenpa. Subtle steric hindrance at thecentromeric nucleosomes o¡ers a likely explana-tion for the observed mitotic defects seen in theCenpag/g embryos.
Lack of a visible effect on nucleosomalorganization
To examine whether the overall organization ofCenpag/g homotypic nucleosomes was a¡ected bythe incorporation of the Cenpa^GFP carboxyl tail,we partially digested nuclear extracts from day 9.5embryos with micrococcal nuclease. Digestionproducts were run on a 2% agarose gel and stainedwith ethidium bromide. The DNA revealed acharacteristic laddering pattern typical of nucleasedigestion of the DNA linker region betweenneighbouring nucleosomes (data not shown). Thenuclease-protected region should serve as ameasure of the loop size for the DNA that wrapsaround single and multiple nucleosomes, deter-mined next.
In mouse, a class of repetitive DNA known asminor satellite resides at the centromere. ThisDNA closely overlaps with the kinetochoredomain as determined by anti-centromere sera(Wong & Rattner, 1988) or by combined anti-Cenpa immuno£uorescence and FISH analysis ofextended chromatin ¢bres (PK, EE and KHAC,unpublished data). This DNA therefore provides agood probe to allow the speci¢c detection ofCenpa-containing nucleosomal DNA. A secondclass of repetitive DNA known as the majorsatellite recognizes a much larger region juxta-posed to the kinetochore and forming a separatesubstantial domain of pericentromeric hetero-chromatin.Mouse minor and major satellite DNA probes
were used to hybridize Southern blots containingmicrococcal nuclease-digested nucleosomal pre-parations. No di¡erence in band migration wasseen in the wild-type, heterozygotes and homo-zygotes for both the probes (Figure 6). Closer
Figure 6. Micrococcal nuclease digests of day 9.5 embryos. Tenmicrograms of day 9.5 nuclear extracts were digested with 0.5or 0.1 units of micrococcal nuclease for 20min. Digests wereelectrophoresed on a 2% agarose gel, blotted and hybridisedwith either mouse minor or major satellite DNA. Horizontaldashes indicate nucleosomal ladders, beginning withmonomeric nucleosomes at the bottom of the gel.
354 P. Kalitsis et al.

examination of the banding patterns between thedi¡erent genotypes indicated identical sizes for theprotected mononucleosomal loop DNA (repre-sented by the lowest band) as well as the inter-nucleosomal linker regions (evident from theregularity of the spacing of the banding ladders).These results showed that the formation ofCenpag/g homotypic nucleosomes has not in adetectable way altered the overall size of theprimary monomeric nucleosomal structure or thesecondary spacing of these nucleosomes to resultin a signi¢cant increase or decrease in the numberof base pairs wrapping around or linking thesenucleosomes.Studies performed with the S. cerevisiae CENP-
A homolog Cse4p have demonstrated no obser-vable phenotype when GFP or other fusion tags,such as the smaller HA epitope, are placed withinthe N-terminal tail (Chen et al. 2000), whereas astrong temperature-sensitive phenotype is seenwhen the tag is placed at the C-terminus (S. Stolerand M. Fitzgerald-Hayes, personal communica-tion). This suggests that the position of the tag maybe important in in£uencing steric properties on thenucleosome or higher-order chromatin organi-zation. Another study in S. pombe has shown thata C-terminus SpCENP-A-GPF fusion proteintargets to the centromere and is functional(Takahashi et al. 2000); however, the detailede¡ects of this protein on chromosome segregationhave not been described. Other reported C-ter-minus CENPA^GFP fusion proteins in HeLa andDrosophila cell lines do not show a chromosomemissegregation phenotype (Heniko¡ et al. 2000),presumably because these proteins have beenexpressed in wild-type cells containing nativeCENP-A and are therefore more akin to thesituation in our phenotypically normal Cenpaþ/g
heterozygotes.Comparison of the N-terminal domains of
CENP-A and other histone H3-like proteinsreveals variable length and little sequence con-servation (Sullivan 2001). Additionally, theN-terminal tails of CENP-A and other histonesare characteristically £exible and protrude fromthe nucleosome where they are exposed to post-translational modi¢cations (Zeitlin et al. 2001,Zhang & Reinberg 2001). In contrast, theC-terminal segment encompasses the globularhistone fold domain that is important for the
interactions between histones (Luger et al. 1997).Any structural perturbation of this terminus, asmay be inadvertently caused by the (CenpaGFP-H4)2 heterotetramer in the Cenpa
g/g embryos, hasthe potential to compromise the bundling of thecarboxyl-Cenpa helices or the packaging of cen-tromeric chromatin. Despite our failure to detectany alteration to the organization of the nucleo-somes at the resolution used, the detailed chro-mosomal missegregation phenotype nonethelessindicates clearly that a subtle and as yet unde¢nedmode of interference to the centromeric chromatinstructure is su⁄cient to compromise the normallong-term mitotic segregation behaviour of theCenpag/g embryos containing only the hetero-tetramer (CenpaGFP-H4)2.In the Cenpaþ /g mice, assuming comparable
levels of expression of the Cenpa^GFP and nativeCenpa genes (since both are under the control ofendogenous Cenpa promoter) and similar rateof incorporation, the centromeric nucleosomes ofthese animals are predicted to contain a 1 : 2 : 1mixture of the three possible heterotetramer types(Cenpa-H4)2 : (Cenpa-H4)(CenpaGFP-H4) : (Cen-paGFP-H4)2. Thus, in view of the phenotypice¡ects seen in the Cenpag/g embryos, it is of notethat the presence of the Cenpa^GFP hetero-tetramer types in possibly up to three-quarters ofthe centromeric nucleosomes in the heterozygoteshas little or no detrimental e¡ect on centromerefunction. These observations therefore furtherhighlight the resilience and pliability of the cen-tromere chromatin structure. Further exploitationof the Cenpag/g and Cenpaþ/g fusion systemshould facilitate the understanding of the orga-nizational requirements of centromeric chromatinin general, and the role of CENP-A in particular.
Role of centromere protein gene mutationsin human abnormalities
A number of studies in mice have shown that nullmutations of most centromere protein genes resultin a severe chromosome missegregation pheno-type, where cells can only undergo a limitednumber of aberrant divisions during the ¢rst 2^3days of embryogenesis (Kalitsis et al. 1998a, Cuttset al. 1999, Dobles et al. 2000, Howman et al.2000, Kalitsis et al. 2000, Uren et al. 2000, Putkeyet al. 2002). However, mice heterozygous for
Aneuploidy and apoptosis in Cenpa^GFP knockin mice 355

the mitotic spindle checkpoint gene, Mad2, showan increased incidence of tumour formation(Michel et al. 2001). The only viable centromereprotein gene knockout described has been the non-essential centromere protein Cenpb where nullmutants are by and large mitotically and meio-tically normal (Hudson et al. 1998, Kapoor et al.1998, Perez-Castro et al. 1998, Fowler et al. 2000).To date, no constitutional mutations in any cen-tromere protein gene have been identi¢ed inhumans, due presumably to such mutations eithershowing no major e¡ects (for example CENP-B)or, as would be predicted for most of the genesbased on evidence from the published mousemodels, causing a severe phenotype that results invery early embryo loss. An example of possiblenon-constitutional centromere protein genemutation in humans has been described for themitotic spindle checkpoint protein BUB1 in whichmutation in one allele appears to exert an apparentdominant-negative e¡ect leading to colon cancer(Cahill et al. 1998). By creating a mutant Cenpafusion in mice, we have demonstrated the possi-bility that a subtle constitutional mutation of anessential centromere protein gives rise to a chro-mosome missegregation phenotype that causesdelayed embryonic lethality in mice comparedwith null mutations of the corresponding Cenpaand other essential centromere protein genes(Howman et al. 2000). It is conceivable that a hostof naturally occurring mutations in these essentialcentromere protein genes may exist in humans thatsimilarly cause a chromosome missegregation orgenome instability phenotype that can be linked tolater-stage miscarriage during pregnancy andpossibly other growth defects and cancer.
Acknowledgements
We thank T. Yen for anti-hBUBR1, E. D. Salmonand B. Howell for anti-Mad2, and B. Williams andM. L. Goldberg for anti-Zw10, B. Anderson forhistological preparations, P. Farmer for TUNELstaining, D. Newgreen and P. Farlie for discus-sions on mouse embryos, S. Gazeas and mouselaboratory sta¡ for mouse care and mouse tailDNA preparations. This work was supported bythe National Health and Medical ResearchCouncil of Australia. K. H. A. C. is a Senior
Principal Research Fellow of the council and anassociate of the Department of Paediatrics, Uni-versity of Melbourne.
References
Belmont AS (2001) Visualizing chromosome dynamics withGFP. Trends Cell Biol 11: 250^257.
Blower MD, Karpen GH (2001) The role of Drosophila CID inkinetochore formation, cell-cycle progression and hetero-chromatin interactions. Nat Cell Biol 3: 730^739.
Cahill DP, Lengauer C, Yu J et al. (1998) Mutations of mitoticcheckpoint genes in human cancers. Nature 392: 300^303.
Chen Y, Baker RE, Keith KC, Harris K, Stoler S,Fitzgerald-Hayes M (2000) The N terminus of the centro-mere H3-like protein Cse4p performs an essential functiondistinct from that of the histone fold domain. Mol Cell Biol20: 7037^7048.
Choo KHA (2001) Domain organization at the centromere andneocentromere. Dev Cell 1: 165^177.
Cutts SM, Fowler KJ, Kile BT et al. (1999) Defective chromo-some segregation, microtubule bundling and nuclearbridging in inner centromere protein gene (Incenp)-disruptedmice. Hum Mol Genet 8: 1145^1155.
Dobles M, Liberal V, Scott ML, Benezra R, Sorger PK (2000)Chromosome missegregation and apoptosis in mice lackingthe mitotic checkpoint protein Mad2. Cell 101: 635^645.
Figueroa J, Saffrich R, Ansorge W, Valdivia M (1998)Microinjection of antibodies to centromere protein CENP-Aarrests cells in interphase but does not prevent mitosis.Chromosoma 107: 397^405.
Fowler KJ, Hudson DF, Salamonsen LA et al. (2000) Uterinedysfunction and genetic modi¢ers in centromere proteinB-de¢cient mice. Genome Res 10: 30^41.
Fukagawa T, Brown WRA (1997) Ef¢cient conditionalmutation of the vertebrate CENP-C gene. Hum Mol Genet6: 2301^2308.
Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identi¢cation ofprogrammed cell death in situ via speci¢c labeling of nuclearDNA fragmentation. J Cell Biol 119: 493^501.
Glowczewski L, Yang P, Kalashnikova T, Santisteban MS,Smith MM (2000) Histone^histone interactions andcentromere function. Mol Cell Biol 20: 5700^5711.
Henikoff S, Ahmad K, Platero JS, van Steensel B (2000)Heterochromatic deposition of centromeric histone H3-likeproteins. Proc Natl Acad Sci USA 97: 716^721.
Howman EV, Fowler KJ, Newson AJ et al. (2000) Early dis-ruption of centromeric chromatin organization in centromereprotein A (Cenpa) null mice. Proc Natl Acad Sci USA 97:1148^1153.
Hudson DF, Fowler KJ, Earle E et al. (1998) Centromere pro-tein B null mice are mitotically and meiotically normalbut have lower body and testis weights. J Cell Biol 141:309^319.
Jablonski SA, Chan GK, Cooke CA, Earnshaw WC, Yen TJ(1998) The hBUB1 and hBUBR1 kinases sequentiallyassemble onto kinetochores during prophase with hBUBR1
356 P. Kalitsis et al.

concentrating at the kinetochore plates in mitosis.Chromosoma 107: 386^396.
Kalitsis P, Fowler KJ, Earle E, Hill J, Choo KHA (1998a)Targeted disruption of mouse centromere protein C geneleads to mitotic disarray and early embryo death. Proc NatlAcad Sci USA 95: 1136^1141.
Kalitsis P, MacDonald AC, Newson AJ, Hudson DF, ChooKHA (1998b) Gene structure and sequence analysis of mousecentromere proteins A and C. Genomics 47: 108^114.
Kalitsis P, Earle E, Fowler KJ, Choo KHA (2000) Bub3 genedisruption in mice reveals essential mitotic spindlecheckpoint function during early embryogenesis. GenesDev 14: 2277^2282.
Kapoor M, Montes de Oca Luna R, Liu G et al. (1998) ThecenpB gene is not essential in mice. Chromosoma 107:570^576.
Lo AWI, Craig JM, Saffery R et al. (2001) A 330 kb CENP-Abinding domain and altered replication timing at a humanneocentromere. Embo J 20: 2087^2096.
Luger K, Mader AW, Richmond RK, Sargent DF, RichmondTJ (1997) Crystal structure of the nucleosome core particleat 2.8 A resolution. Nature 389: 251^260.
Michel LS, Liberal V, Chatterjee A et al. (2001) MAD2haplo-insuf¢ciency causes premature anaphase and chromo-some instability in mammalian cells. Nature 409: 355^359.
Nishihashi A, Haraguchi T, Hiraoka Y et al. (2002) CENP-I isessential for centromere function in vertebrate cells. Dev Cell2: 463^476.
Oegema K, Desai A, Rybina S, KirkhamM, Hyman AA (2001)Functional analysis of kinetochore assembly in Caenor-habditis elegans. J Cell Biol 153: 1209^1226.
Perez-Castro AV, Shamanski FL, Meneses JJ et al. (1998)Centromeric protein B null mice are viable with no apparentabnormalities. Dev Biol 201: 135^143.
Putkey F, Cramer T, Morphew M et al. (2002) Unstablekinetochore-microtubule capture and chromosomal insta-bility following deletion of CENP-E. Dev Cell 3: 351.
Saffery R, Earle E, Irvine DV, Kalitsis P, Choo KHA (1999)Conservation of centromere protein in vertebrates. Chromo-some Res 7: 261^265.
Saitoh H, Tomkiel J, Cooke CA et al. (1992) CENP-C, an auto-antigen in scleroderma, is a component of the human innerkinetochore plate. Cell 70: 115^125.
Starr DA, Williams BC, Li Z, Etemad-Moghadam B, DaweRK, Goldberg ML (1997) Conservation of the centromere/kinetochore protein ZW10. J Cell Biol 138: 1289^1301.
Stoler S, Keith KC, Curnick KE, Fitzgerald-Hayes M (1995) Amutation in CSE4, an essential gene encoding a novelchromatin-associated protein in yeast, causes chromosomenondisjunction and cell cycle arrest at mitosis. Genes Dev.9: 573^586.
Sugata N, Li S, Earnshaw WC et al. (2000) Human CENP-Hmultimers colocalize with CENP-A and CENP-C at activecentromere^kinetochore complexes. Hum Mol Genet 9:2919^2926.
Sullivan KF (2001) A solid foundation: functional speciali-zation of centromeric chromatin. Curr Opin Genet Dev11: 182^188.
Sullivan BA, Blower MD, Karpen GH (2001) Determiningcentromere identity: cyclical stories and forking paths.Nat Rev Genet 2: 584^596.
Takahashi K, Chen ES, Yanagida M (2000) Require-ment of Mis6 centromere connector for localizing aCENP-A-like protein in ¢ssion yeast. Science 288:2215^2219.
Tsien RY (1998) The green £uorescent protein. Annu RevBiochem 67: 509^544.
Uren AG, Wong L, Pakusch M et al. (2000) Survivin and theinner centromere protein INCENP show similar cell-cyclelocalization and gene knockout phenotype. Curr Biol 10:1319^1328.
Van Hooser AA, Ouspenski II, Gregson HC et al. (2001) Speci-¢cation of kinetochore-forming chromatin by the histone H3variant CENP-A. J Cell Sci 114: 3529^3542.
Waters JC, Chen RH, Murray AW, Salmon ED (1998)Localization of Mad2 to kinetochores depends onmicrotubule attachment, not tension. J Cell Biol 141:1181^1191.
Wong AK, Rattner JB (1988) Sequence organization andcytological localization of the minor satellite of mouse.Nucleic Acids Res 16: 11645^11661.
Yoda K, Ando S, Morishita S et al. (2000) Human centromereprotein A (CENP-A) can replace histone H3 in nucleosomereconstitution in vitro. Proc Natl Acad Sci USA 97:7266^7271.
Zeitlin SG, Barber CM, Allis CD, Sullivan KF (2001)Differential regulation of CENP-A and histone H3phosphorylation in G2/M. J Cell Sci 114: 653^661.
Zhang Y, Reinberg D (2001) Transcription regulation byhistone methylation: interplay between different covalentmodi¢cations of the core histone tails. Genes Dev 15:2343^2360.
Aneuploidy and apoptosis in Cenpa^GFP knockin mice 357
Copyright © 2022 FDOKUMEN




















