
Dual Fluorescent Reporter Pig for Cre Recombination: Transgene Placement at the ROSA26 Locus
Upload
independentCategory
view
2download
0
A functional Bucky ball-GFP transgene visualizes germ plasmin living zebrafishStephan Riemer a, Franck Bontems b, Pritesh Krishnakumar a, Jasmin Gömann a,Roland Dosch a,*a Institut für Entwicklungsbiochemie, GZMB, Universitätsmedizin Göttingen, Georg-August-Universität, Göttingen, Germanyb Laboratory of Metabolism, Department of Internal Medicine Specialties, Faculty of Medicine, University of Geneva, Switzerland
A R T I C L E I N F O
Article history:Received 29 April 2015Received in revised form 8 May 2015Accepted 9 May 2015Available online 2 July 2015
Keywords:Bucky ballGermline developmentPrimordial germ cellTransgeneBalbiani bodyVasa
A B S T R A C T
In many animals, the germline is specified by maternal RNA-granules termed germ plasm. The correctlocalization of germ plasm during embryogenesis is therefore crucial for the specification of germ cells.In zebrafish, we previously identified Bucky ball (Buc) as a key regulator of germ plasm formation. Here,we used a Buc antibody to describe its continuous germ plasm localization. Moreover, we generated atransgenic Buc-GFP line for live imaging, which visualizes germ plasm from its assembly during oogen-esis up to the larval stages. Live imaging of Buc-GFP generated stunning movies, as they highlighted thedynamic details of germ plasm movements. Moreover, we discovered that Buc was still detected in pri-mordial germ cells 2 days after fertilization. Interestingly, the transgene rescued buc mutants demonstratinggenetically that the Buc-GFP fusion protein is functional. These results show that Buc-GFP exerts all bio-chemical interactions essential for germline development and highlight the potential of this line to analyzethe molecular regulation of germ plasm formation.
© 2015 Elsevier B.V. All rights reserved.
1. Introduction
Multicellular organisms separate their soma from reproductivegerm cells, which are specified in many species by the inheritanceof cell fate determinants called germ plasm (reviewed inEwen-Campen et al., 2010; Houston and King, 2000; Juliano et al.,2010; Schisa, 2012; Strome and Lehmann, 2007; Voronina et al., 2011).Germ plasm assembles during oogenesis forming a membrane-free compartment called the Balbiani body, which is observed inthe oocytes of many species including humans and zebrafish(Heasman et al., 1984; reviewed in Dosch, 2015; Kloc et al., 2004;Lindeman and Pelegri, 2010; Raz, 2003). Although germ plasm hasa critical role for germ line development, the molecular mecha-nisms by which it induces germ cell formation are still largelyunknown.
Bucky ball (Buc) is a crucial regulator of germline developmentin zebrafish, since buc−/− oocytes fail to form a Balbiani body (Bontemset al., 2009; Marlow and Mullins, 2008). Fascinatingly, overexpressionof Buc reprograms somatic cells in vivo into primordial germ cells
(PGCs), which so far has not been reported for other vertebrate genes(Bontems et al., 2009). In addition, injecting Buc-GFP mRNA labelsgerm plasm during oogenesis and embryogenesis identifying Bucas the first germ plasm organizer in vertebrates. However, injec-tion of mRNA encoding a Buc-GFP-fusion is (I) technically challengingin early oocytes, (II) creates unspecific background, (III) requires atleast an hour to develop a fluorescent signal, and (IV) influencesgermline development limiting its application as a germline re-porter. Existing germline reporters in zebrafish have the advantageof being biologically inactive i.e. they do not interfere with germlinedevelopment (Blaser et al., 2005; Krøvel and Olsen, 2002; Leu andDraper, 2010; Onichtchouk et al., 2003). Unfortunately, they do notlabel germ plasm as a subcellular compartment and thus, a differ-ent approach is required to monitor germ plasm in living zebrafish.
Here, we used the Buc protein to follow germ plasm dynamicsin zebrafish. We first characterized the endogenous localization ofBuc with a novel antibody validating its suitability as a germ plasmmarker. Then we generated a transgenic line expressing Buc-GFPunder control of its own promotor (tg(buc:buc-egfp), hereafter calledBuc-GFP). The Buc-GFP signal perfectly recapitulated germ plasmlocalization at high resolution in living oocytes and embryos. In ad-dition, we detected fluorescence in germ cells 2 days post fertilizationin the Buc-GFP line showing that Buc continuously highlights germplasm from the oocyte up to larval stages. Interestingly, the transgenerescued the mutant phenotype demonstrating genetically that Buc-GFP is also functional and performs all biochemical interactionsessential for germline development like the endogenous protein.
Abbreviations: Buc, Bucky ball; GFP, green fluorescent protein; hpf, hours postfertilization; PGC, primordial germ cell.
* Corresponding author. Universitätsmedizin Göttingen, Georg-August-Universität,Inst. Entwicklungsbiochemie, Justus-von-Liebig-Weg 11, 37077 Göttingen, Germany.Tel.: +49 551 3914607; fax: +49 551 3914614.
E-mail address: [email protected] (R. Dosch).
http://dx.doi.org/10.1016/j.gep.2015.05.0031567-133X/© 2015 Elsevier B.V. All rights reserved.
Gene Expression Patterns 18 (2015) 44–52
Contents lists available at ScienceDirect
Gene Expression Patterns
journal homepage: www.elsevier.com/ locate /gep
Thus, the Buc-GFP line provides an excellent entry point for furtherstudies dissecting the molecular regulation of germ plasm in thezebrafish. As Buc homologs are present in many vertebrate genomes,the protein might also be a versatile germline marker for these species.
2. Results
2.1. A novel Bucky ball antibody
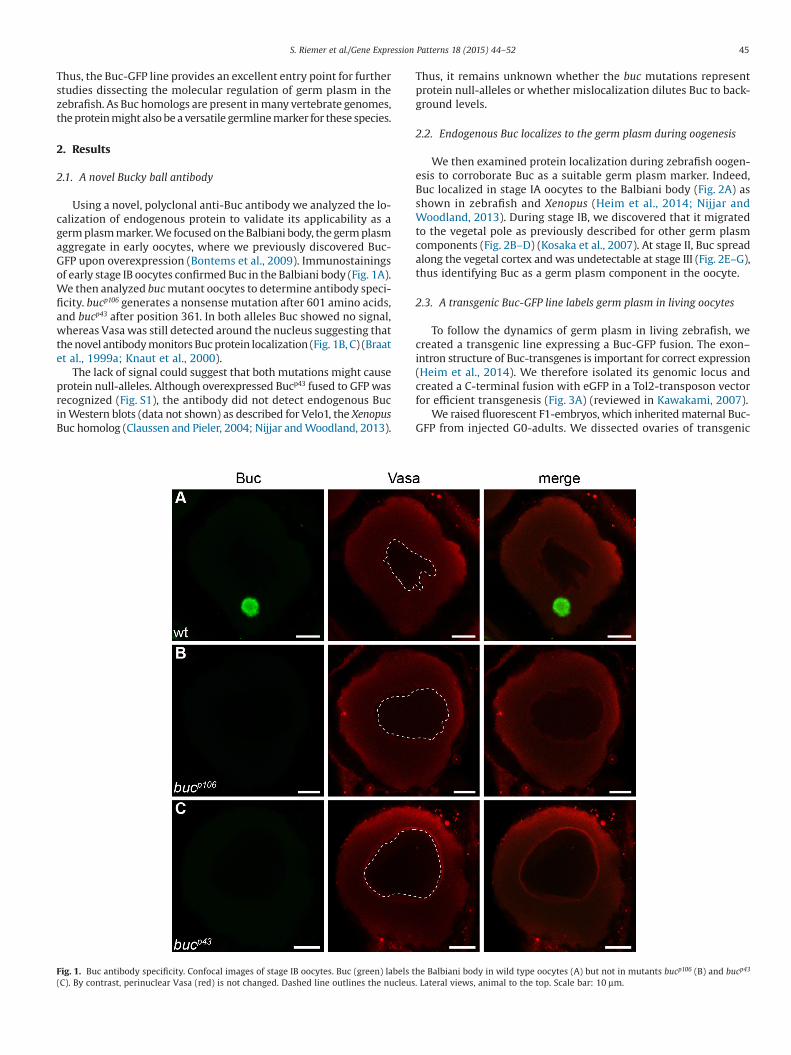
Using a novel, polyclonal anti-Buc antibody we analyzed the lo-calization of endogenous protein to validate its applicability as agerm plasm marker. We focused on the Balbiani body, the germ plasmaggregate in early oocytes, where we previously discovered Buc-GFP upon overexpression (Bontems et al., 2009). Immunostainingsof early stage IB oocytes confirmed Buc in the Balbiani body (Fig. 1A).We then analyzed buc mutant oocytes to determine antibody speci-ficity. bucp106 generates a nonsense mutation after 601 amino acids,and bucp43 after position 361. In both alleles Buc showed no signal,whereas Vasa was still detected around the nucleus suggesting thatthe novel antibody monitors Buc protein localization (Fig. 1B, C) (Braatet al., 1999a; Knaut et al., 2000).
The lack of signal could suggest that both mutations might causeprotein null-alleles. Although overexpressed Bucp43 fused to GFP wasrecognized (Fig. S1), the antibody did not detect endogenous Bucin Western blots (data not shown) as described for Velo1, the XenopusBuc homolog (Claussen and Pieler, 2004; Nijjar and Woodland, 2013).
Thus, it remains unknown whether the buc mutations representprotein null-alleles or whether mislocalization dilutes Buc to back-ground levels.
2.2. Endogenous Buc localizes to the germ plasm during oogenesis
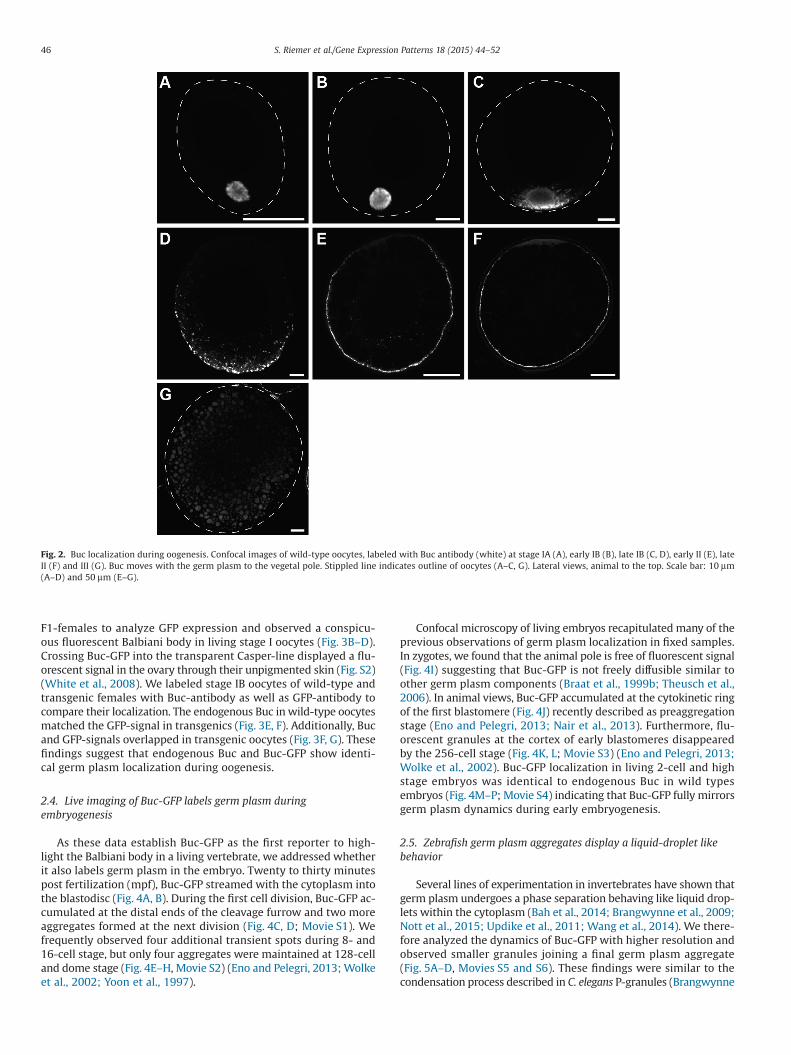
We then examined protein localization during zebrafish oogen-esis to corroborate Buc as a suitable germ plasm marker. Indeed,Buc localized in stage IA oocytes to the Balbiani body (Fig. 2A) asshown in zebrafish and Xenopus (Heim et al., 2014; Nijjar andWoodland, 2013). During stage IB, we discovered that it migratedto the vegetal pole as previously described for other germ plasmcomponents (Fig. 2B–D) (Kosaka et al., 2007). At stage II, Buc spreadalong the vegetal cortex and was undetectable at stage III (Fig. 2E–G),thus identifying Buc as a germ plasm component in the oocyte.
2.3. A transgenic Buc-GFP line labels germ plasm in living oocytes
To follow the dynamics of germ plasm in living zebrafish, wecreated a transgenic line expressing a Buc-GFP fusion. The exon–intron structure of Buc-transgenes is important for correct expression(Heim et al., 2014). We therefore isolated its genomic locus andcreated a C-terminal fusion with eGFP in a Tol2-transposon vectorfor efficient transgenesis (Fig. 3A) (reviewed in Kawakami, 2007).
We raised fluorescent F1-embryos, which inherited maternal Buc-GFP from injected G0-adults. We dissected ovaries of transgenic
Fig. 1. Buc antibody specificity. Confocal images of stage IB oocytes. Buc (green) labels the Balbiani body in wild type oocytes (A) but not in mutants bucp106 (B) and bucp43
(C). By contrast, perinuclear Vasa (red) is not changed. Dashed line outlines the nucleus. Lateral views, animal to the top. Scale bar: 10 μm.
45S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52
F1-females to analyze GFP expression and observed a conspicu-ous fluorescent Balbiani body in living stage I oocytes (Fig. 3B–D).Crossing Buc-GFP into the transparent Casper-line displayed a flu-orescent signal in the ovary through their unpigmented skin (Fig. S2)(White et al., 2008). We labeled stage IB oocytes of wild-type andtransgenic females with Buc-antibody as well as GFP-antibody tocompare their localization. The endogenous Buc in wild-type oocytesmatched the GFP-signal in transgenics (Fig. 3E, F). Additionally, Bucand GFP-signals overlapped in transgenic oocytes (Fig. 3F, G). Thesefindings suggest that endogenous Buc and Buc-GFP show identi-cal germ plasm localization during oogenesis.
2.4. Live imaging of Buc-GFP labels germ plasm duringembryogenesis
As these data establish Buc-GFP as the first reporter to high-light the Balbiani body in a living vertebrate, we addressed whetherit also labels germ plasm in the embryo. Twenty to thirty minutespost fertilization (mpf), Buc-GFP streamed with the cytoplasm intothe blastodisc (Fig. 4A, B). During the first cell division, Buc-GFP ac-cumulated at the distal ends of the cleavage furrow and two moreaggregates formed at the next division (Fig. 4C, D; Movie S1). Wefrequently observed four additional transient spots during 8- and16-cell stage, but only four aggregates were maintained at 128-celland dome stage (Fig. 4E–H, Movie S2) (Eno and Pelegri, 2013; Wolkeet al., 2002; Yoon et al., 1997).
Confocal microscopy of living embryos recapitulated many of theprevious observations of germ plasm localization in fixed samples.In zygotes, we found that the animal pole is free of fluorescent signal(Fig. 4I) suggesting that Buc-GFP is not freely diffusible similar toother germ plasm components (Braat et al., 1999b; Theusch et al.,2006). In animal views, Buc-GFP accumulated at the cytokinetic ringof the first blastomere (Fig. 4J) recently described as preaggregationstage (Eno and Pelegri, 2013; Nair et al., 2013). Furthermore, flu-orescent granules at the cortex of early blastomeres disappearedby the 256-cell stage (Fig. 4K, L; Movie S3) (Eno and Pelegri, 2013;Wolke et al., 2002). Buc-GFP localization in living 2-cell and highstage embryos was identical to endogenous Buc in wild typesembryos (Fig. 4M–P; Movie S4) indicating that Buc-GFP fully mirrorsgerm plasm dynamics during early embryogenesis.
2.5. Zebrafish germ plasm aggregates display a liquid-droplet likebehavior
Several lines of experimentation in invertebrates have shown thatgerm plasm undergoes a phase separation behaving like liquid drop-lets within the cytoplasm (Bah et al., 2014; Brangwynne et al., 2009;Nott et al., 2015; Updike et al., 2011; Wang et al., 2014). We there-fore analyzed the dynamics of Buc-GFP with higher resolution andobserved smaller granules joining a final germ plasm aggregate(Fig. 5A–D, Movies S5 and S6). These findings were similar to thecondensation process described in C. elegans P-granules (Brangwynne
Fig. 2. Buc localization during oogenesis. Confocal images of wild-type oocytes, labeled with Buc antibody (white) at stage IA (A), early IB (B), late IB (C, D), early II (E), lateII (F) and III (G). Buc moves with the germ plasm to the vegetal pole. Stippled line indicates outline of oocytes (A–C, G). Lateral views, animal to the top. Scale bar: 10 μm(A–D) and 50 μm (E–G).
46 S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52

et al., 2009). Thus, with the Buc-GFP line it is now feasible to in-vestigate the biophysical properties of germ plasm in a vertebrate.
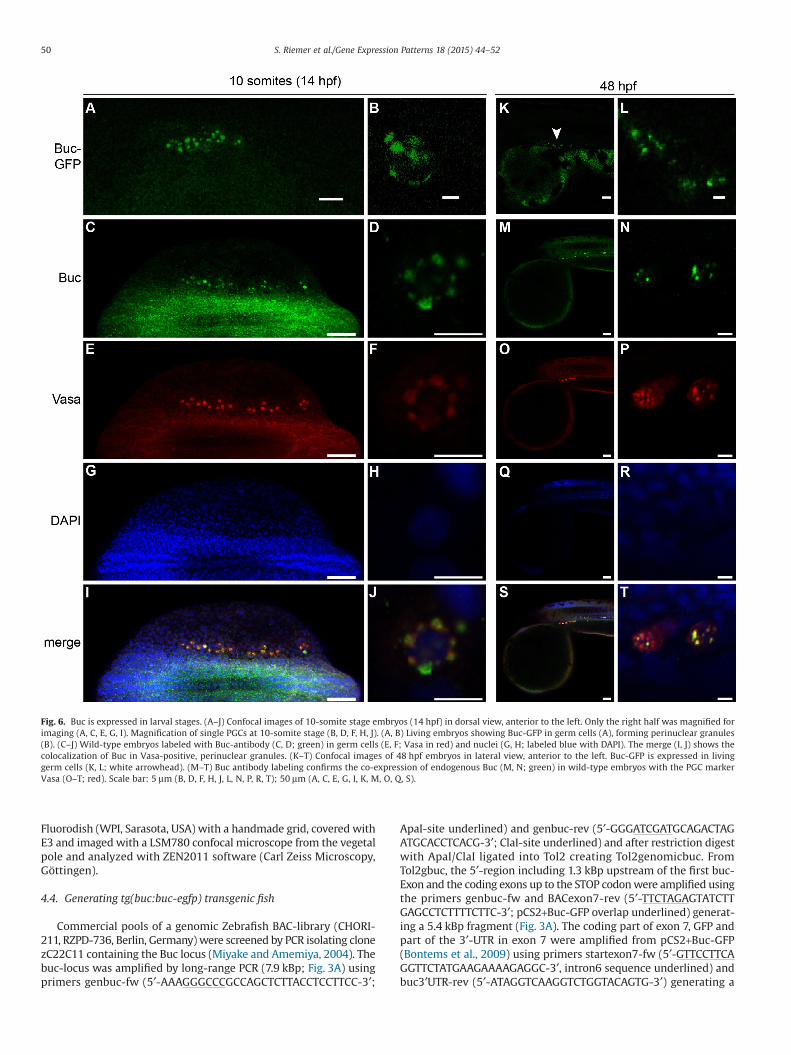
2.6. Buc is expressed in germ cells 2 days post fertilization
The fluorescence of overexpressed Buc-GFP disappears arounddome stage (4.3 hours post fertilization; hpf) (Bontems et al., 2009),which led us to assume that Buc protein is downregulated duringmid-blastula transition. Surprisingly, when we examined germ cellsof 10-somite stage or 48 hpf embryos, Buc-GFP was still visible(Fig. 6A, B, K, L). We analyzed the expression of endogenous Buc todetermine whether the GFP-fusion increases protein stability.However, endogenous Buc protein was also detected at 48 hpf inwild-type larvae (Fig. 6C, D, M, N). Vasa labeling confirmed their germcell identity (Fig. 6E–T) and colocalized with Buc in perinuclear gran-ules (Köprunner et al., 2001; Strasser et al., 2008). These results showthat maternal Buc is preserved in the germ plasm until 48 hpf, whileits mRNA is degraded at 4 hpf. Taken together, the Buc-GFP line re-capitulated germ plasm dynamics in living samples continuouslyfrom its assembly during oogenesis until larval stages.
2.7. The Buc-GFP fusion protein is functional
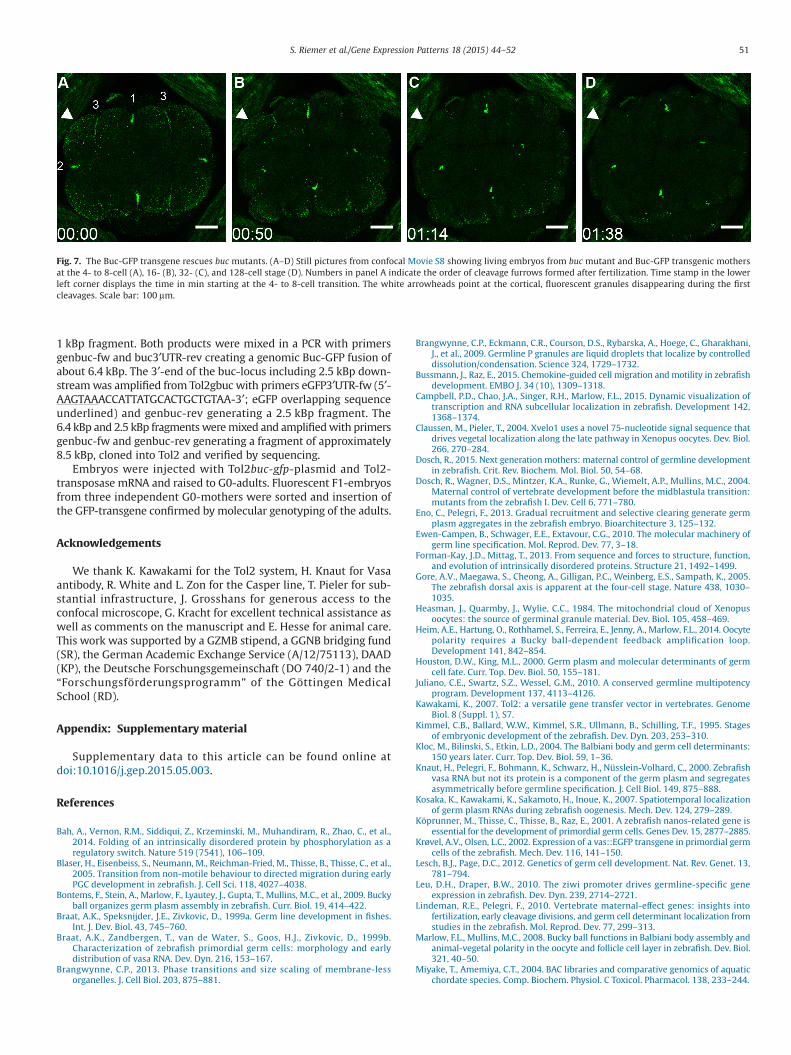
The Buc-GFP transgene mirrored germ plasm localization, butimaging experiments do not address the activity of the protein fusedto GFP. Therefore, we crossed the transgene into buc mutants and ana-lyzed whether the embryonic polarity defect is complemented by Buc-GFP (Dosch et al., 2004). Embryos from mutant mothers developednormally and showed fluorescent germ plasm (Fig. 7; Movie S7). More-over, we detected the same characteristics of germ plasm dynamics
as described above e.g. degrading fluorescent granules at the blas-tomere cortex and transient germ plasm at the third cleavage furrow(Fig. 7A–D, Movie S8) (Eno and Pelegri, 2013; Wolke et al., 2002). Wepropagated the transgene over several generations in a buc−/− mutantbackground demonstrating genetically that Buc-GFP is spatially andtemporally active, when endogenous Buc is required. Based on therescue, we also concluded that Buc-GFP interacts with all binding part-ners essential for germ line development.
3. Discussion
3.1. Buc provides a useful marker for vertebrate germ plasm
In this study, we showed that endogenous Buc is continuouslypresent in germ plasm and hence, that the protein might be a veryhelpful marker in all vertebrates with a Buc homolog (Bontems et al.,2009). This will be particularly interesting in mammalian species,since they are considered to specify germ cells via inductive signals(Ewen-Campen et al., 2010; Lesch and Page, 2012; Seervai andWessel, 2013). Although a mammalian Buc mutant will be essen-tial to judge Buc function during mammalian germ cell specification,it would be already exciting to know whether Buc localizes to theBalbiani body in mammals.
3.2. Buc-GFP permits live imaging of vertebrate germ plasm
With the Buc-GFP line, we provide the first reporter to followgerm plasm in living zebrafish from its assembly during oogenesisuntil 2 days after fertilization. The line recapitulates all describedaspects of germ line dynamics and provides a useful entry point to
Fig. 3. Buc-GFP transgenic zebrafish. (A) Scheme showing the genomic Buc locus used to generate the Buc-GFP line. Thin lines represent non-coding regions. Boxes repre-sent exons (white: untranslated RNA; red: coding RNA). The green box marks the insertion point of the GFP-protein in front of the STOP-codon. (B–D) Live images of Buc-GFP (green) in oocytes from transgenic mothers at early stage IB (B), late IB (C), and II (D). The nuclei of follicle cells were counterstained with the live-cell fluorescent dyeHoechst 33342 (blue). (E–G) Confocal images comparing endogenous Buc localization with Buc-GFP. (E) Wild-type stage IB oocyte labeled with Buc antibody (white) com-pared to transgenic Buc-GFP oocyte labeled with GFP antibody (F; green) and Buc (G; white). Dashed yellow line indicates oocyte outline (E–G). Lateral view, animal to thetop. Scale bar: 10 μm (E–G) 25 μm (B–D).
47S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52

investigate the molecular mechanism of germ plasm localizationin living zebrafish. This will facilitate studies on the interaction ofgerm plasm with the cytoskeleton or RNAs by live imaging (Campbellet al., 2015; Gore et al., 2005; Tran et al., 2012), which are other-wise incompatible due to different fixation methods (Theusch et al.,2006). Furthermore, the progressive effects of experimental ma-nipulations such as cytoskeletal drugs on germ plasm are observablevia live imaging and not just endpoint analysis. In the future, it will
be possible to follow germ plasm dynamics in living samples withoutstanding spatial and temporal resolution using STED (stimulationemission depletion) or SPIM (selective plane illumination) micros-copy (Reynaud et al., 2015; Willig et al., 2006).
Similar transgenic lines in invertebrates were critical to revealthe liquid-droplet like behavior of invertebrate germ plasm (Bah et al.,2014; Brangwynne et al., 2009; Nott et al., 2015; Updike et al., 2011;Wang et al., 2014). The results provided in this study suggest a similar
Fig. 4. Buc-GFP localization during embryogenesis. (A–H) Images of living embryos from Buc-GFP mothers at 1- (A, B), 2- (C), 4- (D), 8- (E), 16- (F), 128-cell (G), and highstage (H). (I–L) Confocal images of living embryos from transgenic mothers at the 1- (I, J), 4-(K), and 256-cell-stage (L). (I) Dashed yellow line outlines the blastomere in thezygote. Note that the animal region was free of fluorescent signal. (J) Fluorescent granules formed a ring at the preaggregation stage. (K, L) The four blastomeres accumu-lated fluorescent granules at their cortex (arrowhead), which disappeared by the 256-cell stage. (M–P) Confocal images comparing endogenous Buc (white) in wild-type(M, O) with GFP (green) in Buc-GFP embryos (N, P) at the 2-cell (M, N) or high-stage stage (O, P). (A, I) Lateral views, (B–H, J–P) animal views. Scale bar: 50 μm (N, P), 100 μm(I–M, O), 200 μm (A–H).
48 S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52

phase separation in vertebrates, but require a much more quantitativeanalysis of granule condensation, dissolution and surface wetting.Germ plasm is often used as a paradigm for membrane-free com-partments, which has emerged in recent years as an exciting researchtopic at the interface of biochemistry, cell biology, and biophysics(reviewed in Brangwynne, 2013; Forman-Kay and Mittag, 2013).Thus, the Buc-GFP line will be quite potent to determine the bio-physical properties of germ plasm and whether it shows droplet-like behavior in vertebrates.
3.3. Buc-GFP encodes a functional germ plasm-tag for biochemistry
As the transgene fully rescued buc mutants over several gen-erations, we demonstrated the functionality of the Buc-GFP fusion.Many zebrafish transgenes encoding GFP-fusions are available, butto our knowledge their activity was hitherto not examined genet-ically i.e. whether the GFP fusion complements a mutation in thesame gene. Buc-GFP has therefore great potential as an endog-enously expressed protein-tag for stage-specific affinity capture ofbiochemical interactions. This will be quite helpful to determine thefunction of Buc itself, as it encodes a protein without known domainsand thus, without any predicted biochemical function.
In conclusion, our findings highlight Buc-GFP as a supportive plat-form for researchers interested in the vertebrate germline. Furtherapplications include Buc-GFP as a germ plasm reporter in forwardgenetic screens. Furthermore, the Buc-GFP line will allow the pu-rification of germ cells to generate germline clones, but moreinterestingly, to analyze temporal changes of their transcriptome,proteome and epigenetic modifications. Correlating these data withthe in vivo behavior of germ cells, e.g. at the onset of migration(Bussmann and Raz, 2015; Richardson and Lehmann, 2010) will iden-tify novel genes and provide a greater understanding of theirregulation during germline development. Taken together, the Buc-GFP fish add a novel and powerful tool to study germ plasm in avertebrate and open new avenues for germline research.
4. Experimental procedures
4.1. Animals
Fish were maintained as described (Westerfield, 2000) in accor-dance with regulations of the Georg-August University Göttingen,Germany. Oocytes and embryos were staged as published (Kimmelet al., 1995; Selman et al., 1993). The buc alleles p106 and p43 andthe transparent casper line were previously described (Bontems et al.,2009; White et al., 2008).
4.2. Immunohistochemistry
Buc antibody was raised in guinea pig against recombinant, fulllength Buc (BioGenes, Berlin, Germany). The serum was purified andtested for specificity with recombinant Buc. Antibody dilutions:guinea pig anti-Buc (1:5000), rabbit anti-Vasa (1:500 (Knaut et al.,2000)), GFP-booster Atto 488 (1:500; ChromoTek, Planegg-Martinsried, Germany), goat-anti-guinea pig Alexa Fluor 488, goatanti-rabbit Alexa Fluor 594 (1:500, Life Technologies, Carlsbad, USA).
Zebrafish ovaries were dissected from anesthetized females, trans-ferred into OR2 buffer (5 mM HEPES (pH 7.5), 82.5 mM NaCl, 2 mMKCl, 1 mM MgCl2) and dissociated for 3 min at room temperaturein Proteinase K (0.1 M Tris (pH 7.5), 10 mM EDTA, 50 μg/ml Pro-teinase K). Ovaries were washed twice with MEM (0.1 M MOPS(pH 7.4), 2 mM EGTA, 1 mM MgSO4), fixed for 1 h at room temper-ature in MEMFA (MEM, 3.7% formaldehyde), washed three times withPBT (137 mM NaCl, 10 mM Na2HPO4, 2.7 mM KCl, 1.76 mM KH2PO4;pH 7.4), 0.1% Triton X-100, 0.2% BSA) and stored in PBT at 4 °C forup to 5 days before immunostaining.
Embryos were dechorionated with pronase (50 μg/ml) inE3-medium, fixed for 6 hrs at room temperature and subsequentlyovernight at 4 °C in 4% paraformaldehyde in PBS (137 mM NaCl,10 mM Na2HPO4, 2.7 mM KCl, 1.76 mM KH2PO4; pH 7.4). Embryoswere washed three times with PBS, dehydrated in a methanol di-lution series and stored in 100% methanol at −20 °C. Forimmunostaining, embryos were gradually rehydrated in PBT.
Samples were blocked for 2 hrs at room temperature in PBT block-ing solution (PBT, 2% BSA, 2% horse serum) and incubated overnightat 4 °C with primary antibody in PBT. After washing 3 × 15 min withPBT, samples were incubated with secondary antibody in PBT over-night at 4 °C. After washing 3 × 15 min with PBT, samples werecounterstained with DAPI (0.8 μg/ml in PBT) for 1 hr at room tem-perature. After washing 3 × 15 min with PBT, oocytes weredehydrated in a methanol dilution series and stored in 100% meth-anol at −20 °C. For imaging, oocytes were transferred to a Fluorodish(35 mm; WPI, Sarasota, USA), methanol removed and the yolk clearedwith Murray’s clearing medium (2/3 benzyl benzoate, 1/3 benzylalcohol). Embryos were manually deyolked prior to clearance of theremaining yolk.
4.3. Imaging
Living oocytes, embryos and adults were imaged with a stereo-microscope SteREO Lumar.V12 (Carl Zeiss Microscopy, Göttingen).Embryos were manually dechorionated and mounted in 1.5% agarose/E3 medium. Images were analyzed with Axio Vision Rel. 4.8 software.For confocal analysis, dechorionated embryos were placed in a
Fig. 5. Zebrafish germ plasm dynamics suggest liquid droplet-like behavior. (A–D) Still pictures from confocal Movie S5 show the condensation of two granules (white ar-rowhead) with a germ plasm aggregate. Scale bar: 20 μm.
49S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52
Fluorodish (WPI, Sarasota, USA) with a handmade grid, covered withE3 and imaged with a LSM780 confocal microscope from the vegetalpole and analyzed with ZEN2011 software (Carl Zeiss Microscopy,Göttingen).
4.4. Generating tg(buc:buc-egfp) transgenic fish
Commercial pools of a genomic Zebrafish BAC-library (CHORI-211, RZPD-736, Berlin, Germany) were screened by PCR isolating clonezC22C11 containing the Buc locus (Miyake and Amemiya, 2004). Thebuc-locus was amplified by long-range PCR (7.9 kBp; Fig. 3A) usingprimers genbuc-fw (5′-AAAGGGCCCGCCAGCTCTTACCTCCTTCC-3′;
ApaI-site underlined) and genbuc-rev (5′-GGGATCGATGCAGACTAGATGCACCTCACG-3′; ClaI-site underlined) and after restriction digestwith ApaI/ClaI ligated into Tol2 creating Tol2genomicbuc. FromTol2gbuc, the 5′-region including 1.3 kBp upstream of the first buc-Exon and the coding exons up to the STOP codon were amplified usingthe primers genbuc-fw and BACexon7-rev (5′-TTCTAGAGTATCTTGAGCCTCTTTTCTTC-3′; pCS2+Buc-GFP overlap underlined) generat-ing a 5.4 kBp fragment (Fig. 3A). The coding part of exon 7, GFP andpart of the 3′-UTR in exon 7 were amplified from pCS2+Buc-GFP(Bontems et al., 2009) using primers startexon7-fw (5′-GTTCCTTCAGGTTCTATGAAGAAAAGAGGC-3′, intron6 sequence underlined) andbuc3′UTR-rev (5′-ATAGGTCAAGGTCTGGTACAGTG-3′) generating a
Fig. 6. Buc is expressed in larval stages. (A–J) Confocal images of 10-somite stage embryos (14 hpf) in dorsal view, anterior to the left. Only the right half was magnified forimaging (A, C, E, G, I). Magnification of single PGCs at 10-somite stage (B, D, F, H, J). (A, B) Living embryos showing Buc-GFP in germ cells (A), forming perinuclear granules(B). (C–J) Wild-type embryos labeled with Buc-antibody (C, D; green) in germ cells (E, F; Vasa in red) and nuclei (G, H; labeled blue with DAPI). The merge (I, J) shows thecolocalization of Buc in Vasa-positive, perinuclear granules. (K–T) Confocal images of 48 hpf embryos in lateral view, anterior to the left. Buc-GFP is expressed in livinggerm cells (K, L; white arrowhead). (M–T) Buc antibody labeling confirms the co-expression of endogenous Buc (M, N; green) in wild-type embryos with the PGC markerVasa (O–T; red). Scale bar: 5 μm (B, D, F, H, J, L, N, P, R, T); 50 μm (A, C, E, G, I, K, M, O, Q, S).
50 S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52
1 kBp fragment. Both products were mixed in a PCR with primersgenbuc-fw and buc3′UTR-rev creating a genomic Buc-GFP fusion ofabout 6.4 kBp. The 3′-end of the buc-locus including 2.5 kBp down-stream was amplified from Tol2gbuc with primers eGFP3′UTR-fw (5′-AAGTAAACCATTATGCACTGCTGTAA-3′; eGFP overlapping sequenceunderlined) and genbuc-rev generating a 2.5 kBp fragment. The6.4 kBp and 2.5 kBp fragments were mixed and amplified with primersgenbuc-fw and genbuc-rev generating a fragment of approximately8.5 kBp, cloned into Tol2 and verified by sequencing.
Embryos were injected with Tol2buc-gfp-plasmid and Tol2-transposase mRNA and raised to G0-adults. Fluorescent F1-embryosfrom three independent G0-mothers were sorted and insertion ofthe GFP-transgene confirmed by molecular genotyping of the adults.
Acknowledgements
We thank K. Kawakami for the Tol2 system, H. Knaut for Vasaantibody, R. White and L. Zon for the Casper line, T. Pieler for sub-stantial infrastructure, J. Grosshans for generous access to theconfocal microscope, G. Kracht for excellent technical assistance aswell as comments on the manuscript and E. Hesse for animal care.This work was supported by a GZMB stipend, a GGNB bridging fund(SR), the German Academic Exchange Service (A/12/75113), DAAD(KP), the Deutsche Forschungsgemeinschaft (DO 740/2-1) and the“Forschungsförderungsprogramm” of the Göttingen MedicalSchool (RD).
Appendix: Supplementary material
Supplementary data to this article can be found online atdoi:10.1016/j.gep.2015.05.003.
References
Bah, A., Vernon, R.M., Siddiqui, Z., Krzeminski, M., Muhandiram, R., Zhao, C., et al.,2014. Folding of an intrinsically disordered protein by phosphorylation as aregulatory switch. Nature 519 (7541), 106–109.
Blaser, H., Eisenbeiss, S., Neumann, M., Reichman-Fried, M., Thisse, B., Thisse, C., et al.,2005. Transition from non-motile behaviour to directed migration during earlyPGC development in zebrafish. J. Cell Sci. 118, 4027–4038.
Bontems, F., Stein, A., Marlow, F., Lyautey, J., Gupta, T., Mullins, M.C., et al., 2009. Buckyball organizes germ plasm assembly in zebrafish. Curr. Biol. 19, 414–422.
Braat, A.K., Speksnijder, J.E., Zivkovic, D., 1999a. Germ line development in fishes.Int. J. Dev. Biol. 43, 745–760.
Braat, A.K., Zandbergen, T., van de Water, S., Goos, H.J., Zivkovic, D., 1999b.Characterization of zebrafish primordial germ cells: morphology and earlydistribution of vasa RNA. Dev. Dyn. 216, 153–167.
Brangwynne, C.P., 2013. Phase transitions and size scaling of membrane-lessorganelles. J. Cell Biol. 203, 875–881.
Brangwynne, C.P., Eckmann, C.R., Courson, D.S., Rybarska, A., Hoege, C., Gharakhani,J., et al., 2009. Germline P granules are liquid droplets that localize by controlleddissolution/condensation. Science 324, 1729–1732.
Bussmann, J., Raz, E., 2015. Chemokine-guided cell migration and motility in zebrafishdevelopment. EMBO J. 34 (10), 1309–1318.
Campbell, P.D., Chao, J.A., Singer, R.H., Marlow, F.L., 2015. Dynamic visualization oftranscription and RNA subcellular localization in zebrafish. Development 142,1368–1374.
Claussen, M., Pieler, T., 2004. Xvelo1 uses a novel 75-nucleotide signal sequence thatdrives vegetal localization along the late pathway in Xenopus oocytes. Dev. Biol.266, 270–284.
Dosch, R., 2015. Next generation mothers: maternal control of germline developmentin zebrafish. Crit. Rev. Biochem. Mol. Biol. 50, 54–68.
Dosch, R., Wagner, D.S., Mintzer, K.A., Runke, G., Wiemelt, A.P., Mullins, M.C., 2004.Maternal control of vertebrate development before the midblastula transition:mutants from the zebrafish I. Dev. Cell 6, 771–780.
Eno, C., Pelegri, F., 2013. Gradual recruitment and selective clearing generate germplasm aggregates in the zebrafish embryo. Bioarchitecture 3, 125–132.
Ewen-Campen, B., Schwager, E.E., Extavour, C.G., 2010. The molecular machinery ofgerm line specification. Mol. Reprod. Dev. 77, 3–18.
Forman-Kay, J.D., Mittag, T., 2013. From sequence and forces to structure, function,and evolution of intrinsically disordered proteins. Structure 21, 1492–1499.
Gore, A.V., Maegawa, S., Cheong, A., Gilligan, P.C., Weinberg, E.S., Sampath, K., 2005.The zebrafish dorsal axis is apparent at the four-cell stage. Nature 438, 1030–1035.
Heasman, J., Quarmby, J., Wylie, C.C., 1984. The mitochondrial cloud of Xenopusoocytes: the source of germinal granule material. Dev. Biol. 105, 458–469.
Heim, A.E., Hartung, O., Rothhamel, S., Ferreira, E., Jenny, A., Marlow, F.L., 2014. Oocytepolarity requires a Bucky ball-dependent feedback amplification loop.Development 141, 842–854.
Houston, D.W., King, M.L., 2000. Germ plasm and molecular determinants of germcell fate. Curr. Top. Dev. Biol. 50, 155–181.
Juliano, C.E., Swartz, S.Z., Wessel, G.M., 2010. A conserved germline multipotencyprogram. Development 137, 4113–4126.
Kawakami, K., 2007. Tol2: a versatile gene transfer vector in vertebrates. GenomeBiol. 8 (Suppl. 1), S7.
Kimmel, C.B., Ballard, W.W., Kimmel, S.R., Ullmann, B., Schilling, T.F., 1995. Stagesof embryonic development of the zebrafish. Dev. Dyn. 203, 253–310.
Kloc, M., Bilinski, S., Etkin, L.D., 2004. The Balbiani body and germ cell determinants:150 years later. Curr. Top. Dev. Biol. 59, 1–36.
Knaut, H., Pelegri, F., Bohmann, K., Schwarz, H., Nüsslein-Volhard, C., 2000. Zebrafishvasa RNA but not its protein is a component of the germ plasm and segregatesasymmetrically before germline specification. J. Cell Biol. 149, 875–888.
Kosaka, K., Kawakami, K., Sakamoto, H., Inoue, K., 2007. Spatiotemporal localizationof germ plasm RNAs during zebrafish oogenesis. Mech. Dev. 124, 279–289.
Köprunner, M., Thisse, C., Thisse, B., Raz, E., 2001. A zebrafish nanos-related gene isessential for the development of primordial germ cells. Genes Dev. 15, 2877–2885.
Krøvel, A.V., Olsen, L.C., 2002. Expression of a vas::EGFP transgene in primordial germcells of the zebrafish. Mech. Dev. 116, 141–150.
Lesch, B.J., Page, D.C., 2012. Genetics of germ cell development. Nat. Rev. Genet. 13,781–794.
Leu, D.H., Draper, B.W., 2010. The ziwi promoter drives germline-specific geneexpression in zebrafish. Dev. Dyn. 239, 2714–2721.
Lindeman, R.E., Pelegri, F., 2010. Vertebrate maternal-effect genes: insights intofertilization, early cleavage divisions, and germ cell determinant localization fromstudies in the zebrafish. Mol. Reprod. Dev. 77, 299–313.
Marlow, F.L., Mullins, M.C., 2008. Bucky ball functions in Balbiani body assembly andanimal-vegetal polarity in the oocyte and follicle cell layer in zebrafish. Dev. Biol.321, 40–50.
Miyake, T., Amemiya, C.T., 2004. BAC libraries and comparative genomics of aquaticchordate species. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 138, 233–244.
Fig. 7. The Buc-GFP transgene rescues buc mutants. (A–D) Still pictures from confocal Movie S8 showing living embryos from buc mutant and Buc-GFP transgenic mothersat the 4- to 8-cell (A), 16- (B), 32- (C), and 128-cell stage (D). Numbers in panel A indicate the order of cleavage furrows formed after fertilization. Time stamp in the lowerleft corner displays the time in min starting at the 4- to 8-cell transition. The white arrowheads point at the cortical, fluorescent granules disappearing during the firstcleavages. Scale bar: 100 μm.
51S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52

Nair, S., Marlow, F., Abrams, E., Kapp, L., Mullins, M.C., Pelegri, F., 2013. Thechromosomal passenger protein birc5b organizes microfilaments and germ plasmin the zebrafish embryo. PLoS Genet. 9, e1003448.
Nijjar, S., Woodland, H.R., 2013. Protein interactions in Xenopus germ plasm RNPparticles. PLoS ONE 8, e80077.
Nott, T.J., Petsalaki, E., Farber, P., Jervis, D., Fussner, E., Plochowietz, A., et al., 2015.Phase transition of a disordered nuage protein generates environmentallyresponsive membraneless organelles. Mol. Cell 57, 936–947.
Onichtchouk, D., Aduroja, K., Belting, H.G., Gnugge, L., Driever, W., 2003. Transgenedriving GFP expression from the promoter of the zona pellucida gene zpc isexpressed in oocytes and provides an early marker for gonad differentiation inzebrafish. Dev. Dyn. 228, 393–404.
Raz, E., 2003. Primordial germ-cell development: the zebrafish perspective. Nat. Rev.Genet. 4, 690–700.
Reynaud, E.G., Peychl, J., Huisken, J., Tomancak, P., 2015. Guide to light-sheetmicroscopy for adventurous biologists. Nat. Methods 12, 30–34.
Richardson, B.E., Lehmann, R., 2010. Mechanisms guiding primordial germcell migration: strategies from different organisms. Nat. Rev. Mol. Cell Biol. 11,37–49.
Schisa, J.A., 2012. New insights into the regulation of RNP granule assembly in oocytes.Int. Rev. Cell Mol. Biol. 295, 233–289.
Seervai, R.N., Wessel, G.M., 2013. Lessons for inductive germline determination. Mol.Reprod. Dev. 80, 590–609.
Selman, K., Wallace, R.A., Sarka, A., Qi, X.P., 1993. Stages of oocyte development inthe zebrafish, Brachydanio rerio. J. Morphol. 218, 203–224.
Strasser, M.J., Mackenzie, N.C., Dumstrei, K., Nakkrasae, L.I., Stebler, J., Raz, E., 2008.Control over the morphology and segregation of zebrafish germ cell granulesduring embryonic development. BMC Dev. Biol. 8, 58.
Strome, S., Lehmann, R., 2007. Germ versus soma decisions: lessons from flies andworms. Science 316, 392–393.
Theusch, E.V., Brown, K.J., Pelegri, F., 2006. Separate pathways of RNA recruitmentlead to the compartmentalization of the zebrafish germ plasm. Dev. Biol. 292,129–141.
Tran, L.D., Hino, H., Quach, H., Lim, S., Shindo, A., Mimori-Kiyosue, Y., et al., 2012.Dynamic microtubules at the vegetal cortex predict the embryonic axis inzebrafish. Development 139, 3644–3652.
Updike, D.L., Hachey, S.J., Kreher, J., Strome, S., 2011. P granules extend the nuclearpore complex environment in the C. elegans germ line. J. Cell Biol. 192, 939–948.
Voronina, E., Seydoux, G., Sassone-Corsi, P., Nagamori, I., 2011. RNA granules in germcells. Cold Spring Harb. Perspect. Biol. 3.
Wang, J.T., Smith, J., Chen, B.C., Schmidt, H., Rasoloson, D., Paix, A., et al., 2014.Regulation of RNA granule dynamics by phosphorylation of serine-rich,intrinsically disordered proteins in C. elegans. Elife 4.
Westerfield, M., 2000. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish(Danio Rerio), fourth ed. University of Oregon Press, Eugene.
White, R.M., Sessa, A., Burke, C., Bowman, T., LeBlanc, J., Ceol, C., et al., 2008.Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell StemCell 2, 183–189.
Willig, K.I., Kellner, R.R., Medda, R., Hein, B., Jakobs, S., Hell, S.W., 2006. Nanoscaleresolution in GFP-based microscopy. Nat. Methods 3, 721–723.
Wolke, U., Weidinger, G., Köprunner, M., Raz, E., 2002. Multiple levels ofposttranscriptional control lead to germ line-specific gene expression in thezebrafish. Curr. Biol. 12, 289–294.
Yoon, C., Kawakami, K., Hopkins, N., 1997. Zebrafish vasa homologue RNA is localizedto the cleavage planes of 2- and 4-cell-stage embryos and is expressed in theprimordial germ cells. Development 124, 3157–3165.
52 S. Riemer et al./Gene Expression Patterns 18 (2015) 44–52
Copyright © 2022 FDOKUMEN




















