
The presence of thiolated compounds allows the immobilization of enzymes on glyoxyl agarose at mild...
8
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of The presence of thiolated compounds allows the immobilization of enzymes on glyoxyl agarose at mild...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Enzyme and Microbial Technology 45 (2009) 477–483
Contents lists available at ScienceDirect
Enzyme and Microbial Technology
journa l homepage: www.e lsev ier .com/ locate /emt
The presence of thiolated compounds allows the immobilization of enzymes onglyoxyl agarose at mild pH values: New strategies of stabilization by multipointcovalent attachment
Juan M. Bolivara,1, Fernando López-Gallegoa,1, Cesar Godoya, Dasciana S. Rodriguesa,b,Rafael C. Rodriguesa, Pilar Batallaa, Javier Rocha-Martína, Cesar Mateoa,Raquel L.C. Giordanob,∗, José M. Guisána,∗∗
a Departamento de Biocatálisis, Instituto de Catálisis (CSIC), Campus UAM, Cantoblanco, 28049 Madrid, Spainb Chemical Engineering Department, Federal University of São Carlos, Via Washington Luiz, Km 235, São Carlos 13 565-905, Brazil
a r t i c l e i n f o
Article history:Received 24 April 2009Received in revised form 1 September 2009Accepted 2 September 2009
Keywords:Enzyme immobilizationEnzyme stabilizationMultipoint covalent attachmentGlyoxylImino bonds
a b s t r a c t
Highly activated glyoxyl-supports rapidly immobilize proteins at pH 10 (where the �-amino groups of theLys groups of the protein surface are very reactive), and stabilize them by multipoint covalent attachment.However, they do not immobilize proteins at pH 8. This paper shows that the enzyme immobilizationat this mild pH value is possible by incubation of the enzymes in the presence of different thiolatedcompounds (dithiothreitol, DTT; was selected as optimal reagent). The thiolated compounds (even the notreducing ones) stabilized the imino bonds formed at pH 8 between the aldehydes in the support and theamino groups of the protein. However, thiolated compounds are unable to reduce the imino bonds or thealdehyde groups and a final reduction step (e.g., using sodium borohydride) was always necessary. Afterenzyme immobilization through the most reactive amino group of the protein, the further incubation ofthis immobilized enzyme at pH 10 would improve the reactivity of the �-amino groups of the Lys residuesof the protein surface. Then, an intense multipoint covalent reaction of the enzyme with the dense layer ofglyoxyl groups in the support could be obtained, increasing the stability of the immobilized enzyme. Usingthree different industrially relevant enzymes (penicillin G acylase from Escherichia coli (PGA), lipase fromBacillus thermocatenulatus (BTL2) and glutaryl acylase from Pseudomonas sp. (GA)), new immobilized-stabilized biocatalysts of the enzymes were produced. After reduction, the preparations incubated at pH10 were more stable than those that were only immobilized and reduced at pH 8. In the case of the PGA,this preparation was even 4–5-fold more stable than those obtained by direct immobilization at pH 10(around 40,000–50,000-fold more stable than the soluble enzyme).
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
The immobilization of enzymes is a usual requirement touse them at industrial scale [1–3]. However, this compulsoryrequirement may be coupled to the development of strategiesto improve other enzyme properties during the immobilization(stability, selectivity, etc.) [4]. Most immobilization protocols maysuccessfully solve the problem of the solubility of the enzyme, butonly properly designed immobilization protocols may permit toimprove the enzyme properties. For example, an enzyme attached
∗ Corresponding author. Tel.: +55 16 3351 8707; fax: +55 16 3351 8266.∗∗ Corresponding author. Tel.: +34 91 585 4809; fax: +34 91 585 4760.
E-mail addresses: [email protected] (R.L.C. Giordano), [email protected](J.M. Guisán).
1 Both authors have equally contributed to this paper.
to the support by just one point should present rigidity similarto the soluble enzyme (although intermolecular process such asaggregation, proteolysis, and interaction with surfaces should beprevented) [5,6]. However, an enzyme attached to the support viamany enzyme–support bonds should increase its rigidity [7,8]. Inthis case, the relative positions of the groups involved in the immo-bilization should be maintained during any conformational changeand, that way, the enzyme should become stabilized against anydistorting agent.
The amino groups of the protein surface are the most usedgroups to immobilize proteins, because they are abundant in theprotein surface and they can react with many different groups(aldehydes, epoxydes, cyanogen bromide, etc.) without any kindof previous activation of the enzyme, when they are in a non-ionized form [9]. However, it should be considered that proteinsmay have, at least, two different kinds of primary amino groups.The �-amino group of the Lys residues is the most abundant one,
0141-0229/$ – see front matter © 2009 Elsevier Inc. All rights reserved.doi:10.1016/j.enzmictec.2009.09.001

Author's personal copy
478 J.M. Bolivar et al. / Enzyme and Microbial Technology 45 (2009) 477–483
but presents a pK relatively high (around 10.5–10.8) having poorreactivity at pH values under 9 [10]. However, these groups need tobe involved in the protein immobilization if an intense multipointcovalent attachment is desired [11].
The other amino groups are those having an especially low pK.Considering that the groups that are able to react with the supportshould be fully exposed to the medium, the most common primaryamino group having a low pK value might be the terminal aminogroup of the protein.
Most of the enzyme immobilization methods involving theamino groups in the enzymes are able to immobilize the pro-tein at pH 7–8 (via a very stable bond), in a very efficient way(glutaraldehyde, cyanogen bromide, tresyl chloride, tosyl chloride,etc.) [12–17]. This feature makes these supports very useful toimmobilize proteins under these very mild conditions. However,under these conditions, the protein immobilization should mainlyproceed by the most reactive amino group in the protein, hardlyinvolving many Lys residues [10,11]. Therefore, the impact on theenzyme stability of this kind of immobilization protocols may bequite small [4]. On the other hand, the low stability of these reactivegroups in the support at alkaline pH value did not permit to performlong immobilization nor incubation processes at alkaline pH values,where the Lys could be more reactive. Long immobilization timesare necessary to get an intense multipoint covalent attachment,because the formation of several bonds between enzyme and sup-port requires the correct alignment between groups placed in twodissimilar and rigid structures as a support and a protein surface[11,18]. Thus, the most usual orientation of the enzyme on conven-tional supports (that involving the most reactive amino) could notpermit a high stabilization of the enzyme using standard protocols.
A support activated with glyoxyl groups also reacts with theprimary amino moieties of the proteins, forming reversible iminobonds [10]. However, due to the reversibility of each individualimino bond, the proteins only become immobilized to the supportvia several attachments [10,19]. This means that the immobiliza-tion on these supports needs to be performed at alkaline pH values,and the immobilization occurs via the area of the protein havingmost Lys residues [10]. This is one of the advantages of these sup-port, immobilization is directed towards the areas of the proteinwhere there are the highest possibilities of multipoint covalentattachment and permit its stabilization [20].
However, it could be interesting to develop strategies that couldenable an intense multipoint covalent attachment by differentareas of the protein, because this may alter the enzyme catalyticproperties (e.g., specificity, inhibition, and selectivity) [4] or beingvery relevant for the enzyme stability (and therefore, a less intensemultipoint covalent attachment by this area may be more stabiliz-ing than a more intense multipoint attachment by another area ofthe other protein) [21,22]. This support is very inert and hydrophilicand therefore, very adequate to immobilize proteins for affinitychromatography even when protein stabilization is not pursued.
Furthermore, after the immobilization under mild conditions, itmay be possible to progressively increase the pH value, to achievea progressive increment in the number of enzyme–support bonds.
Here, it is presented a strategy to enable a multipoint cova-lent attachment of proteins immobilized on glyoxyl-supports bythe most reactive amino group located in the protein surface. Tothis goal, it is necessary to be able to immobilize the enzyme onglyoxyl-supports at mild pH, where only amino groups with a lowpK should be reactive. However, under these conditions glyoxylagarose did not immobilize most proteins [10,19]. This could beovercome if this unique imino bond would be stabilized some-how. If this immobilization at more neutral pH values was achieved,the already immobilized enzyme may be incubated at alkaline pHvalue to involve the �-amino groups of Lys residues located aroundthe most reactive amino group in the protein-support reaction.
This may give a multipoint immobilization and rigidify this proteinarea.
Thus, some way to stabilize the imino bonds needs to beattempted in order to permit the first immobilization at mildpH value, using a compound that did not affect the aldehydegroups to permit the further multipoint covalent attachment of theenzyme (that should be decreased if some of the aldehyde becomeinactivated by the reagent or some secondary by-product) [11].Therefore, this work investigated the possibility of using differ-ent thiol compounds to stabilize the imino bonds without risks ofreducing the aldehyde groups on the support. Some papers reporta reversible nucleophilic attack of ionized thiol groups on carbonylmoieties [23,24]. In fact one-point immobilizations on aldehydesupports have been reported by thiazolidine formation [25–27].This transient reduction of thiol groups is fully reversible underaerobic conditions when thiolated compounds are washed away. Asimilar mechanism could also promote an interesting apparent sta-bilization of Schiff’s bases improving the enzyme immobilization onglyoxyl-supports under neutral or slightly alkaline conditions.
As model enzymes, we have chosen three enzymes withindustrial relevance. Penicillin G acylase from Escherichia coli(PGA) is currently utilized for the industrial production of6-aminopenicillanic acid (6-APA) and 7-aminodesacetoxy-cephalosporanic acid (7-ADCA) [28,29]. Moreover, PGA could alsobe used for the enzymatic production of amide bond using a widerange of nucleophiles (amino acids, antibiotic nucleus, alcohols,sugars, etc.) [30–32].
Lipase BTL2 from Bacillus thermocatenulatus (BTL2) is a veryinteresting enzyme with high stability (both at high temperaturesand in the presence of organic solvents) [33–38]. This enzyme hasbeen reported to be useful as an enantioselective biocatalyst indiverse reactions [37,38].
Glutaryl-7-aminocephalosporanic acid acylase (GA) is used inthe industrial production of 7-amino cephalosporanic acid (7-ACA)from cephalosporin C (CPC) [39].
2. Materials and methods
2.1. Materials
p-Nitrophenyl butyrate (p-NPB) was from Fluka. Glutaryl 7-ACA acylase waspurchased from Roche. Glutaryl 7-ACA was kindly donated by Bioferma MurciaS.A. Semipurified penicillin G acylase from E. coli and penicillin G were kind giftsfrom Antibioticos S.A. BTL2 was produced and purified as previously described [40].DL-dithiothreitol (DTT), N-acetyl-l-cysteine, 2-mercaptoethanol, were from SigmaChem. Co. All other reagents were of analytical grade. Low molecular weight cali-bration kit for SDS electrophoresis was purchased from GE Healthcare. Crosslinked10% agarose beads were kindly donated by Hispanagar S.A. (Burgos, Spain). Gly-oxyl agarose beads with different activation degrees were prepared as previouslydescribed [20]. Protein concentration was determined using Bradford’s method [41].
2.2. Methods
2.2.1. Preparation of glutaryl 7-ACA acylaseCommercial preparation of GA was diluted (1/5) in 25 mM potassium phosphate
at pH 7.0 and then dialyzed against 3 × 100 volumes of a 5 mM potassium phosphateat pH 7.0. The dialyzed enzyme was then centrifuged (10,000 × g for 30 min at 4 ◦C)and the supernatant (containing 16 IU/ml) was used as the enzymatic preparationfor further experiments. More than 90% of initial activity was recovered after thisprocess.
2.2.2. Activity determination of GAThe activity of both immobilized and soluble enzyme was measured as pre-
viously described [42]: the initial reaction rates were measured with a pH-STATusing an automatic titrator (Crison micro TT 2050) to determine the amount of glu-taric acid formed. The assays were carried out by adding 0.1 ml of soluble enzymeor suspension of immobilized enzyme to 10 ml of 10 mM glutaryl 7-ACA in 0.1 Mpotassium phosphate at pH 7.5 at 25 ◦C under continuous stirring, and titrating thereaction mixture with 25 mM NaOH. One unit of glutaryl 7-ACA acylase activity wasdefined as the amount of enzyme that is necessary to produce 1 �mol of glutaricacid per minute in the previously described conditions. This enzymatic preparationof GA presented 1.45 IU/mg.
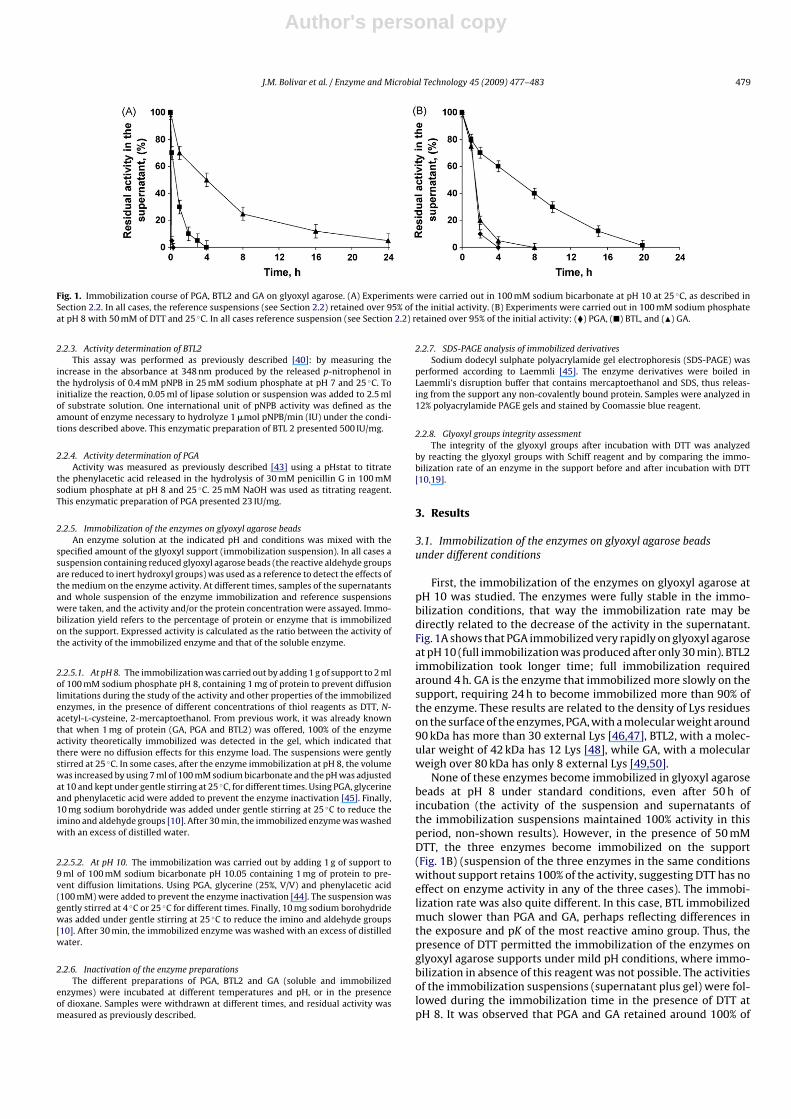
Author's personal copy
J.M. Bolivar et al. / Enzyme and Microbial Technology 45 (2009) 477–483 479
Fig. 1. Immobilization course of PGA, BTL2 and GA on glyoxyl agarose. (A) Experiments were carried out in 100 mM sodium bicarbonate at pH 10 at 25 ◦C, as described inSection 2.2. In all cases, the reference suspensions (see Section 2.2) retained over 95% of the initial activity. (B) Experiments were carried out in 100 mM sodium phosphateat pH 8 with 50 mM of DTT and 25 ◦C. In all cases reference suspension (see Section 2.2) retained over 95% of the initial activity: (�) PGA, (�) BTL, and (�) GA.
2.2.3. Activity determination of BTL2This assay was performed as previously described [40]: by measuring the
increase in the absorbance at 348 nm produced by the released p-nitrophenol inthe hydrolysis of 0.4 mM pNPB in 25 mM sodium phosphate at pH 7 and 25 ◦C. Toinitialize the reaction, 0.05 ml of lipase solution or suspension was added to 2.5 mlof substrate solution. One international unit of pNPB activity was defined as theamount of enzyme necessary to hydrolyze 1 �mol pNPB/min (IU) under the condi-tions described above. This enzymatic preparation of BTL 2 presented 500 IU/mg.
2.2.4. Activity determination of PGAActivity was measured as previously described [43] using a pHstat to titrate
the phenylacetic acid released in the hydrolysis of 30 mM penicillin G in 100 mMsodium phosphate at pH 8 and 25 ◦C. 25 mM NaOH was used as titrating reagent.This enzymatic preparation of PGA presented 23 IU/mg.
2.2.5. Immobilization of the enzymes on glyoxyl agarose beadsAn enzyme solution at the indicated pH and conditions was mixed with the
specified amount of the glyoxyl support (immobilization suspension). In all cases asuspension containing reduced glyoxyl agarose beads (the reactive aldehyde groupsare reduced to inert hydroxyl groups) was used as a reference to detect the effects ofthe medium on the enzyme activity. At different times, samples of the supernatantsand whole suspension of the enzyme immobilization and reference suspensionswere taken, and the activity and/or the protein concentration were assayed. Immo-bilization yield refers to the percentage of protein or enzyme that is immobilizedon the support. Expressed activity is calculated as the ratio between the activity ofthe activity of the immobilized enzyme and that of the soluble enzyme.
2.2.5.1. At pH 8. The immobilization was carried out by adding 1 g of support to 2 mlof 100 mM sodium phosphate pH 8, containing 1 mg of protein to prevent diffusionlimitations during the study of the activity and other properties of the immobilizedenzymes, in the presence of different concentrations of thiol reagents as DTT, N-acetyl-l-cysteine, 2-mercaptoethanol. From previous work, it was already knownthat when 1 mg of protein (GA, PGA and BTL2) was offered, 100% of the enzymeactivity theoretically immobilized was detected in the gel, which indicated thatthere were no diffusion effects for this enzyme load. The suspensions were gentlystirred at 25 ◦C. In some cases, after the enzyme immobilization at pH 8, the volumewas increased by using 7 ml of 100 mM sodium bicarbonate and the pH was adjustedat 10 and kept under gentle stirring at 25 ◦C, for different times. Using PGA, glycerineand phenylacetic acid were added to prevent the enzyme inactivation [45]. Finally,10 mg sodium borohydride was added under gentle stirring at 25 ◦C to reduce theimino and aldehyde groups [10]. After 30 min, the immobilized enzyme was washedwith an excess of distilled water.
2.2.5.2. At pH 10. The immobilization was carried out by adding 1 g of support to9 ml of 100 mM sodium bicarbonate pH 10.05 containing 1 mg of protein to pre-vent diffusion limitations. Using PGA, glycerine (25%, V/V) and phenylacetic acid(100 mM) were added to prevent the enzyme inactivation [44]. The suspension wasgently stirred at 4 ◦C or 25 ◦C for different times. Finally, 10 mg sodium borohydridewas added under gentle stirring at 25 ◦C to reduce the imino and aldehyde groups[10]. After 30 min, the immobilized enzyme was washed with an excess of distilledwater.
2.2.6. Inactivation of the enzyme preparationsThe different preparations of PGA, BTL2 and GA (soluble and immobilized
enzymes) were incubated at different temperatures and pH, or in the presenceof dioxane. Samples were withdrawn at different times, and residual activity wasmeasured as previously described.
2.2.7. SDS-PAGE analysis of immobilized derivativesSodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) was
performed according to Laemmli [45]. The enzyme derivatives were boiled inLaemmli’s disruption buffer that contains mercaptoethanol and SDS, thus releas-ing from the support any non-covalently bound protein. Samples were analyzed in12% polyacrylamide PAGE gels and stained by Coomassie blue reagent.
2.2.8. Glyoxyl groups integrity assessmentThe integrity of the glyoxyl groups after incubation with DTT was analyzed
by reacting the glyoxyl groups with Schiff reagent and by comparing the immo-bilization rate of an enzyme in the support before and after incubation with DTT[10,19].
3. Results
3.1. Immobilization of the enzymes on glyoxyl agarose beadsunder different conditions
First, the immobilization of the enzymes on glyoxyl agarose atpH 10 was studied. The enzymes were fully stable in the immo-bilization conditions, that way the immobilization rate may bedirectly related to the decrease of the activity in the supernatant.Fig. 1A shows that PGA immobilized very rapidly on glyoxyl agaroseat pH 10 (full immobilization was produced after only 30 min). BTL2immobilization took longer time; full immobilization requiredaround 4 h. GA is the enzyme that immobilized more slowly on thesupport, requiring 24 h to become immobilized more than 90% ofthe enzyme. These results are related to the density of Lys residueson the surface of the enzymes, PGA, with a molecular weight around90 kDa has more than 30 external Lys [46,47], BTL2, with a molec-ular weight of 42 kDa has 12 Lys [48], while GA, with a molecularweigh over 80 kDa has only 8 external Lys [49,50].
None of these enzymes become immobilized in glyoxyl agarosebeads at pH 8 under standard conditions, even after 50 h ofincubation (the activity of the suspension and supernatants ofthe immobilization suspensions maintained 100% activity in thisperiod, non-shown results). However, in the presence of 50 mMDTT, the three enzymes become immobilized on the support(Fig. 1B) (suspension of the three enzymes in the same conditionswithout support retains 100% of the activity, suggesting DTT has noeffect on enzyme activity in any of the three cases). The immobi-lization rate was also quite different. In this case, BTL immobilizedmuch slower than PGA and GA, perhaps reflecting differences inthe exposure and pK of the most reactive amino group. Thus, thepresence of DTT permitted the immobilization of the enzymes onglyoxyl agarose supports under mild pH conditions, where immo-bilization in absence of this reagent was not possible. The activitiesof the immobilization suspensions (supernatant plus gel) were fol-lowed during the immobilization time in the presence of DTT atpH 8. It was observed that PGA and GA retained around 100% of
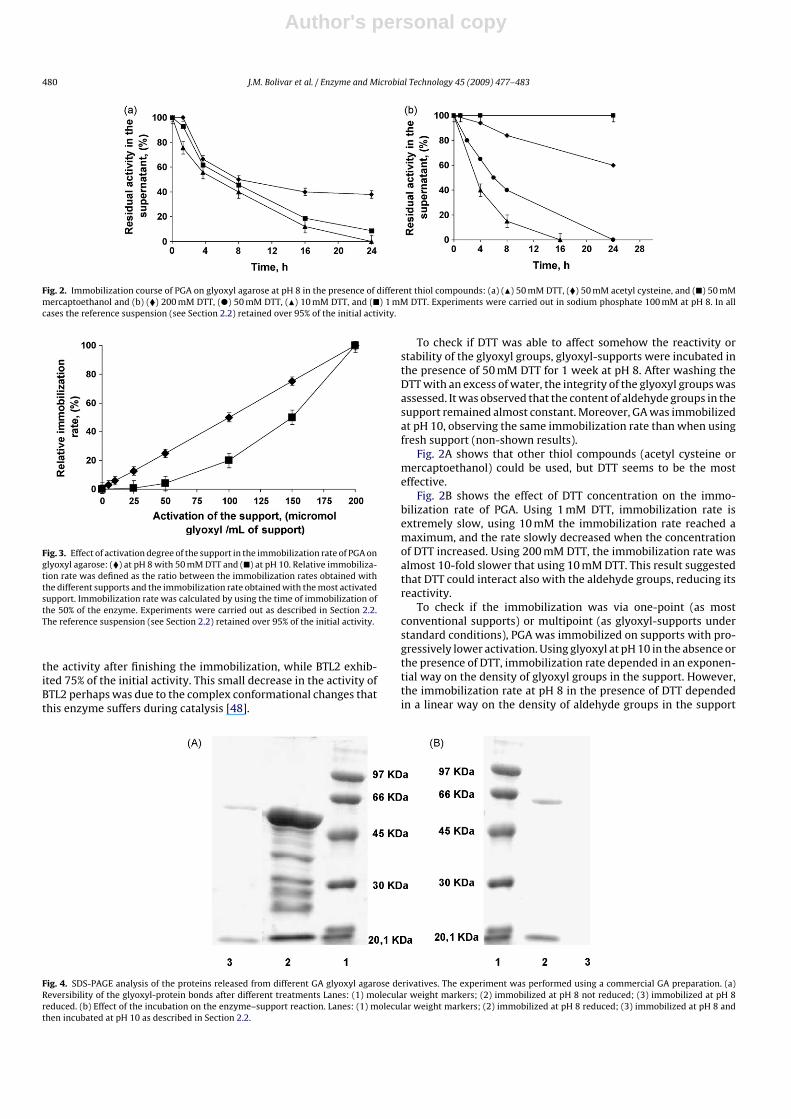
Author's personal copy
480 J.M. Bolivar et al. / Enzyme and Microbial Technology 45 (2009) 477–483
Fig. 2. Immobilization course of PGA on glyoxyl agarose at pH 8 in the presence of different thiol compounds: (a) (�) 50 mM DTT, (�) 50 mM acetyl cysteine, and (�) 50 mMmercaptoethanol and (b) (�) 200 mM DTT, (�) 50 mM DTT, (�) 10 mM DTT, and (�) 1 mM DTT. Experiments were carried out in sodium phosphate 100 mM at pH 8. In allcases the reference suspension (see Section 2.2) retained over 95% of the initial activity.
Fig. 3. Effect of activation degree of the support in the immobilization rate of PGA onglyoxyl agarose: (�) at pH 8 with 50 mM DTT and (�) at pH 10. Relative immobiliza-tion rate was defined as the ratio between the immobilization rates obtained withthe different supports and the immobilization rate obtained with the most activatedsupport. Immobilization rate was calculated by using the time of immobilization ofthe 50% of the enzyme. Experiments were carried out as described in Section 2.2.The reference suspension (see Section 2.2) retained over 95% of the initial activity.
the activity after finishing the immobilization, while BTL2 exhib-ited 75% of the initial activity. This small decrease in the activity ofBTL2 perhaps was due to the complex conformational changes thatthis enzyme suffers during catalysis [48].
To check if DTT was able to affect somehow the reactivity orstability of the glyoxyl groups, glyoxyl-supports were incubated inthe presence of 50 mM DTT for 1 week at pH 8. After washing theDTT with an excess of water, the integrity of the glyoxyl groups wasassessed. It was observed that the content of aldehyde groups in thesupport remained almost constant. Moreover, GA was immobilizedat pH 10, observing the same immobilization rate than when usingfresh support (non-shown results).
Fig. 2A shows that other thiol compounds (acetyl cysteine ormercaptoethanol) could be used, but DTT seems to be the mosteffective.
Fig. 2B shows the effect of DTT concentration on the immo-bilization rate of PGA. Using 1 mM DTT, immobilization rate isextremely slow, using 10 mM the immobilization rate reached amaximum, and the rate slowly decreased when the concentrationof DTT increased. Using 200 mM DTT, the immobilization rate wasalmost 10-fold slower that using 10 mM DTT. This result suggestedthat DTT could interact also with the aldehyde groups, reducing itsreactivity.
To check if the immobilization was via one-point (as mostconventional supports) or multipoint (as glyoxyl-supports understandard conditions), PGA was immobilized on supports with pro-gressively lower activation. Using glyoxyl at pH 10 in the absence orthe presence of DTT, immobilization rate depended in an exponen-tial way on the density of glyoxyl groups in the support. However,the immobilization rate at pH 8 in the presence of DTT dependedin a linear way on the density of aldehyde groups in the support
Fig. 4. SDS-PAGE analysis of the proteins released from different GA glyoxyl agarose derivatives. The experiment was performed using a commercial GA preparation. (a)Reversibility of the glyoxyl-protein bonds after different treatments Lanes: (1) molecular weight markers; (2) immobilized at pH 8 not reduced; (3) immobilized at pH 8reduced. (b) Effect of the incubation on the enzyme–support reaction. Lanes: (1) molecular weight markers; (2) immobilized at pH 8 reduced; (3) immobilized at pH 8 andthen incubated at pH 10 as described in Section 2.2.
Author's personal copy
J.M. Bolivar et al. / Enzyme and Microbial Technology 45 (2009) 477–483 481
(Fig. 3). Thus, immobilization in the presence of DTT seemed tobe via just one group in the support. The obtained results indi-cate that apparently DTT is not only able to stabilize the iminebonds, but also to interact with the aldehyde groups, thus reducingtheir reactivity. Therefore, it is necessary to reach a compromisewhen defining the amount of DDT to be used. It must not reducesignificantly the reactive aldehyde groups concentration, while itstabilizes the imine bonds, allowing immobilization at pH 8.0 andmultipoint attachment at pH 10.
To check the effect of DTT (stabilization of the imino bonds orreduction of these bonds to secondary amino bonds), the differ-ent immobilized preparations of GA were boiled in the presenceof mercaptoethanol and SDS (Fig. 4A). The enzyme immobilized atpH 8 (and later reduced) did release some protein subunits, butmost of the enzyme remained attached to the support. Very likely,these protein subunits desorbed from the support were the sub-units not attached to the support. At pH 8 the reactivity of the Lysgroups, even very near to the glyoxyl groups of the support, willbe low and the involvement of both subunits in the immobilizationshould be difficult. If the biocatalysts were not reduced with sodiumborohydride, the enzyme immobilized at pH 8 in the presence ofDTT released all the immobilized protein to the medium. This sug-gested that DTT was able to stabilize the imino bonds but it was notable to reduce them to stable secondary amino bonds. This effectof DTT and other thiolated compounds could be similar to the onepreviously described for thiols on aldehyde moieties: a transientreduction reversed when removing the thiolated compound underaerobic conditions [23,24]. However, the practical interest and thecomplexity of mechanism of action demand still more studies, fora complete understanding of these effects.
3.2. Incubation of the immobilized enzymes at alkaline pH values
The objective of this strategy was to rigidify (by multipoint cova-lent attachment) the enzyme surface near to the amino group/s thatwas/were reactive at pH 8, thus the enzymes immobilized at pH 8in the presence of DTT were incubated at pH 10 for 24 h [11]. Then,the enzyme derivatives were reduced with sodium borohydride[10]. The activity recoveries of the full immobilization–incubationprotocol were 100% for PGA (the incubation was performed in thepresence of 100 mM phenylacetic acid and 25% (V/V) glycerine topreserve the enzyme activity) [44], and 50–60% for GA and BTL2(during this alkaline incubation of multipoint attachment that theenzyme may be distorted and therefore loses activity). These resultswere similar to those obtained after direct immobilization of theenzymes at pH 10 on glyoxyl agarose [43,44,51].
Immobilized PGA and GA, proteins composed of two dissim-ilar subunits, were boiled in the presence of 1 M ethanolamine,
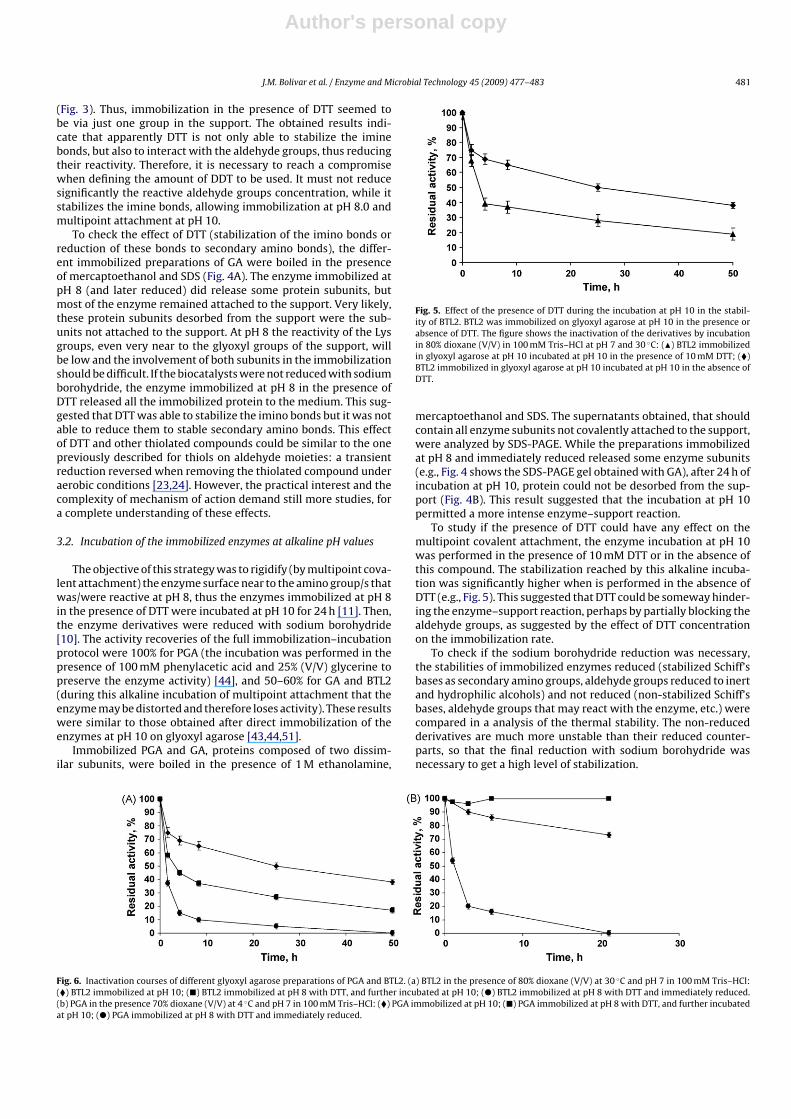
Fig. 5. Effect of the presence of DTT during the incubation at pH 10 in the stabil-ity of BTL2. BTL2 was immobilized on glyoxyl agarose at pH 10 in the presence orabsence of DTT. The figure shows the inactivation of the derivatives by incubationin 80% dioxane (V/V) in 100 mM Tris–HCl at pH 7 and 30 ◦C: (�) BTL2 immobilizedin glyoxyl agarose at pH 10 incubated at pH 10 in the presence of 10 mM DTT; (�)BTL2 immobilized in glyoxyl agarose at pH 10 incubated at pH 10 in the absence ofDTT.
mercaptoethanol and SDS. The supernatants obtained, that shouldcontain all enzyme subunits not covalently attached to the support,were analyzed by SDS-PAGE. While the preparations immobilizedat pH 8 and immediately reduced released some enzyme subunits(e.g., Fig. 4 shows the SDS-PAGE gel obtained with GA), after 24 h ofincubation at pH 10, protein could not be desorbed from the sup-port (Fig. 4B). This result suggested that the incubation at pH 10permitted a more intense enzyme–support reaction.
To study if the presence of DTT could have any effect on themultipoint covalent attachment, the enzyme incubation at pH 10was performed in the presence of 10 mM DTT or in the absence ofthis compound. The stabilization reached by this alkaline incuba-tion was significantly higher when is performed in the absence ofDTT (e.g., Fig. 5). This suggested that DTT could be someway hinder-ing the enzyme–support reaction, perhaps by partially blocking thealdehyde groups, as suggested by the effect of DTT concentrationon the immobilization rate.
To check if the sodium borohydride reduction was necessary,the stabilities of immobilized enzymes reduced (stabilized Schiff’sbases as secondary amino groups, aldehyde groups reduced to inertand hydrophilic alcohols) and not reduced (non-stabilized Schiff’sbases, aldehyde groups that may react with the enzyme, etc.) werecompared in a analysis of the thermal stability. The non-reducedderivatives are much more unstable than their reduced counter-parts, so that the final reduction with sodium borohydride wasnecessary to get a high level of stabilization.
Fig. 6. Inactivation courses of different glyoxyl agarose preparations of PGA and BTL2. (a) BTL2 in the presence of 80% dioxane (V/V) at 30 ◦C and pH 7 in 100 mM Tris–HCl:(�) BTL2 immobilized at pH 10; (�) BTL2 immobilized at pH 8 with DTT, and further incubated at pH 10; (�) BTL2 immobilized at pH 8 with DTT and immediately reduced.(b) PGA in the presence 70% dioxane (V/V) at 4 ◦C and pH 7 in 100 mM Tris–HCl: (�) PGA immobilized at pH 10; (�) PGA immobilized at pH 8 with DTT, and further incubatedat pH 10; (�) PGA immobilized at pH 8 with DTT and immediately reduced.

Author's personal copy
482 J.M. Bolivar et al. / Enzyme and Microbial Technology 45 (2009) 477–483
Fig. 7. Schematic representation of the novel strategy for multipoint covalent immobilization of industrial enzymes.
3.3. Stability of the different immobilized enzymes
Fig. 6 shows the inactivation of different PGA and BTL2 glyoxylagarose preparations: the enzymes immobilized at pH 8 in the pres-ence of DTT and then reduced, the enzymes immobilized at pH 8 andincubated at pH 10 for 24 h, and the enzymes directly immobilizedat pH 10 for 24 h. GA stability was similar for the three prepara-tions, and the stabilization factor compared to the soluble enzymewas very small [42].
For both enzymes, the incubation at pH 10 permitted to improvethe enzyme stability of the enzymes compared to that of theenzyme immobilized at pH 8. This suggested that effectively a rigid-ification of the area involved in the immobilization at pH 8 (themost reactive amine) could be achieved by incubation at a morealkaline pH. Obviously, the stabilization achieved depended on theenzyme. While PGA stability increased 1000 folds, BTL2 increasedits half life by around 50 folds. When the enzymes directly immo-bilized at pH 10 on glyoxyl-supports in the absence of DTT werecompared to those biocatalysts obtained using this new protocol,the results were not homogeneous. BTL2 directly immobilized atpH 10 was 4–5 folds more stable that the preparations obtained byusing the new protocol. However, the new PGA-glyoxyl derivativewas 4–5-fold more stable that the standard glyoxyl preparation.Considering that this PGA preparation was already around 10,000-fold more stable than the soluble enzyme [43], this means that thefinal stabilization reached by this new immobilization protocol wasaround 4–5 × 104.
4. Conclusions
Enzymes hardly immobilize on glyoxyl-supports at neutral orslightly alkaline pH values as Schiff’s bases formed between aminogroups on the protein surface and glyoxyl groups on activatedsupports are very unstable [10,19]. However, different thiolatedcompounds, even with very low reducing potential, are able togreatly stabilize these Schiff’s bases without irreversible reductionof them. This way, immobilization of enzymes on glyoxyl-supportsunder mild pHs (e.g., pH 8) is now possible with no modification ofthe remaining glyoxyl groups (Fig. 7).
Thus, a novel two step multipoint covalent attachment ofenzymes on glyoxyl-supports was designed (Fig. 7): (i) a first
enzyme immobilization at pH 8.0 which very likely is directedvia the most reactive amino group on the enzyme surface (e.g.,the amino terminal one) and (ii) an additional incubation ofthe derivative at pH 10.0 in order to promote an intense mul-tipoint covalent immobilization between Lys residues close tothe support and the very high concentration of glyoxyl groups[11]. This novel strategy was compared with a more conventionalone involving the direct immobilization at pH 10 through theregion having the highest amount of Lys residues. PGA immobi-lized through this novel protocol is 5-fold more stable than PGAdirectly immobilized at pH 10. On the contrary, the new BTL deriva-tive is now 5-fold less stable than the derivative obtained throughthe direct immobilization at pH 10. It seems that effectively afirst immobilization at pH 8.0 involves different enzyme regionsthan the direct immobilization at pH 10. The rigidification of dif-ferent regions of an enzyme surface can be useful to improveenzyme derivatives acting on macromolecular substrates as wellas to test different stabilizing effects. Very likely some enzymeregions could be more sensitive to stabilization even when therigidification occurs through a lower number of covalent attach-ments.
Similar first directed immobilizations under neutral conditionscould be performed by using cyanoborohydride (specific reducingagent of Schiff’s bases) [52–55]. However, from a practical point ofview, CNBH3 is usually unstable yielding traces of BH4 which causesthe reduction of glyoxyl groups of the support making the new twostep rigidification process more difficult.
Acknowledgments
This work has been supported by grants of code S0505/PPQ/0344 of the Comunidad Autónoma de Madrid (CAM) and projectsBIO-2005-8576 and CTQ2005-02420/PPQ from the Ministry of Edu-cation and Science (MEC). Authors thank the Brazilian researchfunding agencies CAPES and CNPq. J.M. Bolívar and J. Rocha areholders of a PhD fellowship from CAM. C. Godoy is a holder of aPhD Spanish MCI. D.S. Rodrigues is hold of a CAPES (Brazil) fel-lowship. P. Batalla has a fellowship from CSIC (cofounded withFEDER founds). Donation of PGA by Antibioticos S.A. is gratefullyrecognized.

Author's personal copy
J.M. Bolivar et al. / Enzyme and Microbial Technology 45 (2009) 477–483 483
References
[1] Cao L. Immobilised enzymes: science or art? Curr Opin Chem Biol2005;9:217–26.
[2] Cao L, van Langen L, Sheldon RA. Immobilised enzymes: carrier-bound orcarrier-free? Curr Opin Biotechnol 2003;14:387–94.
[3] Sheldon RA. Enzyme immobilization: the quest for optimum performance. AdvSynth Catal 2007;349:1289–307.
[4] Mateo C, Palomo JM, Fernandez-Lorente G, Guisan JM, Fernandez-Lafuente R.Improvement of enzyme activity, stability and selectivity via immobilizationtechniques. Enzyme Microb Technol 2007;40:1451–63.
[5] Bolivar JM, Wilson L, Ferrarotti SA, Fernandez-Lafuente R, Guisan JM, Mateo C.Stabilization of a formate dehydrogenase by covalent immobilization on highlyactivated glyoxyl-agarose supports. Biomacromolecules 2006;7:669–73.
[6] Bolivar JM, Wilson L, Ferrarotti A, Guisán SJM, Fernández-Lafuente R, Mateo C.Improvement of the stability of alcohol dehydrogenase by covalent immobi-lization on glyoxyl-agarose. J Biotechnol 2006;125:85–94.
[7] Klibanov AM. Enzyme stabilization by immobilization. Anal Biochem1979;93:1–25.
[8] Gianfreda L, Scarfi MR. Enzyme stabilization: state of the art. Mol Cell Biochem1991;100:97–128.
[9] Guisán JM. Immobilization of enzymes and cells: methods in biotechnology.Totowa, NJ, USA: Humana Press; 2006.
[10] Mateo C, Abian O, Bernedo M, Cuenca E, Fuentes M, Fernandez-Lorente G, etal. Some special features of glyoxyl supports to immobilize proteins. EnzymeMicrob Technol 2005;37:456–62.
[11] Pedroche J, Yust MM, Mateo C, Fernández-Lafuente R, Girón-Calle J, Alaiz M,et al. Effect of the support and experimental conditions in the intensity ofthe multipoint covalent attachment of proteins on glyoxyl-agarose supports:correlation between enzyme–support linkages and thermal stability. EnzymeMicrob Technol 2007;40:1160–7.
[12] Nilsson K, Mosbach K. p-Toluenesulfonyl chloride as an activating agent ofagarose for the preparation of immobilized affinity ligands and proteins. Eur JBiochem 1980;112:397–402.
[13] Albayrak N, Yang S-T. Immobilization of Aspergillus oryzae �-galactosidase ontosylated cotton cloth. Enzyme Microb Technol 2002;31:371–83.
[14] Migneault I, Dartiguenave C, Bertrand MJ, Waldron KC. Glutaraldehyde: behav-ior in aqueous solution, reaction with proteins, and application to enzymecrosslinking. BioTechniques 2004;37:790–802.
[15] Betancor L, López-Gallego F, Hidalgo A, Alonso-Morales N, Mateo GD-O-C,Fernández-Lafuente R, et al. Different mechanisms of protein immobilizationon glutaraldehyde activated supports: effect of support activation and immo-bilization conditions. Enzyme Microb Technol 2006;39:877–82.
[16] Schnapp J, Shalitin Y. Immobilization of enzymes by covalent binding to aminesupports via cyanogen bromide activation. Biochem Biophys Res Commun1976;70:8–14.
[17] Jennissen HP. Cyanogen bromide and tresyl chloride chemistry revisited: thespecial reactivity of agarose as a chromatographic and biomaterial support forimmobilizing novel chemical groups. J Mol Recognit 1995;8:116–24.
[18] Blanco RM, Calvete JJ, Guisan JM. Immobilization-stabilization of enzymes;variables that control the intensity of the trypsin (amine)-agarose (aldehyde)multipoint attachment. Enzyme Microb Technol 1989;11:353–9.
[19] Grazu V, Betancor L, Montes T, Lopez-Gallego F, Guisan JM, Fernandez-LafuenteR. Glyoxyl agarose as a new chromatographic matrix. Enzyme Microb Technol2006;38:960–6.
[20] Mateo C, Palomo JM, Fuentes M, Betancor L, Grazu V, Lopez-Gallego F, et al.Glyoxyl-agarose: a fully inert hydrophilic support for immobilization and highstabilization of proteins. Enzyme Microb Technol 2006;39:274–80.
[21] Mansfeld J, Vriend G, Vanden Burg B, Eijsink V, Ulbrich-Hofmann R. Probing theunfolding region in a thermolysin-like protease by site-specific immobilization.Biochemistry 1999;38:8240–5.
[22] Mansfeld J, Ulbrich-Hofmann R. Site-specific and random immobilizationof thermolysin-like proteases reflected in the thermal inactivation kinetics.Biotechnol Appl Biochem 2000;32:185–95.
[23] Pesek JJ, Frost JH. Decomposition of thiazolidines in acidic and basic solu-tion. Spectroscopic evidence for Schiff base intermediates. Tetrahedron1975;31:907–13.
[24] Szilágyi L, Györgydeák Z. Comments on the putative stereoselectivity incysteine-aldehyde reactions. Selective C(2) inversion and C(4) epimerizationin thiazolidine-4-carboxylic acids. J Am Chem Soc 1979;101:427–32.
[25] Bi X, Hartono D, Yang KL. Controlling orientations of immobilized oligopeptidesusing N-terminal cysteine labels. Langmuir 2008;24:5238–40.
[26] Zatsepin TS, Stetsenko DA, Arzumanov AA, Romanova EA, Gait MJ, OretskayaTS. Synthesis of peptide-oligonucleotide conjugates with single and multiplepeptides attached to 2′-aldehydes through thiazolidine, oxime, and hydrazinelinkages. Bioconjug Chem 2002;13:822–30.
[27] Shao J, Tam JP. Unprotected peptides as building blocks for the synthesis ofpeptide dendrimers with oxime, hydrazone and thiazolidine linkages. J AmChem Soc 1995;117:3893–9.
[28] Sio CF, Quax WJ. Improved �-lactam acylases and their use as industrial bio-catalysts. Curr Opin Biotechnol 2004;15:349–55.
[29] Arroyo M, De La Mata I, Acebal C, Castillón MP. Biotechnological appli-cations of penicillin acylases: state-of-the-art. Appl Microbiol Biotechnol2003;60:507–14.
[30] Fernández-Lafuente R, Rosell CM, Piatkowska B, Guisán JM. Synthesis ofantibiotics (cephaloglycin) catalyzed by penicillin G acylase: evaluation andoptimization of different synthetic approaches. Enzyme Microb Technol1996;19:9–14.
[31] Chen C-X, Wu Q, Liu B-K, Lv D-S, Lin X-F. Anhydrous tert-pentanol as a novelmedia for the efficient enzymatic synthesis of amoxicillin. Enzyme MicrobTechnol 2008;42:601–7.
[32] Rolland-Fulcrand V, Rolland M, Roumestant M-L, Martinez J. Chemoenzy-matic synthesis of enantiomerically pure (2S,3R,4S)-4-hydroxyisoleucine, aninsulinotropic amino acid isolated from fenugreek seeds. Eur J Org Chem2004;4:873–7.
[33] Schmidt-Dannert C, Rúa ML, Schmid RD. Two novel lipases from thermophileBacillus thermocatenulatus: screening, purification, cloning, overexpression,and properties. Methods Enzymol 1997;284:194–222.
[34] Schmidt-Dannert C, Rua ML, Wahl S, Schmid RD. Bacillus thermocatenulatuslipase: a thermoalkalophilic lipase with interesting properties. Biochem SocTrans 1997;25:178–82.
[35] Schmidt-Dannert C, Rúa ML, Atomi H, Schmid RD. Thermoalkalophilic lipaseof Bacillus thermocatenulatus. I. Molecular cloning, nucleotide sequence,purification and some properties. Biochim Biophys Acta, Lipids Lipid Metab1996;1301:105–14.
[36] Schmidt-Dannert C, Sztajer H, Stocklein W, Schmid RD. Screening, purifica-tion and properties of a thermophilic lipase from Bacillus thermocatenulatus.Biochim Biophys Acta, Lipids Lipid Metab 1994;1214:43–53.
[37] Palomo JM, Fernández-Lorente G, Rúa ML, Guisán JM, Fernández-Lafuente R.Evaluation of the lipase from Bacillus thermocatenulatus as an enantioselectivebiocatalyst. Tetrahedron Asymmetry 2003;14:3679–87.
[38] Martín JR, Nus M, Gago JVS, Sánchez-Montero JM. Selective esterification ofphthalic acids in two ionic liquids at high temperatures using a thermostablelipase of Bacillus thermocatenulatus: a comparative study. J Mol Catal B: Enzym2008;52–53:162–7.
[39] Fechtig B, Peter H, Bickel H, Vischer E. Concerning the preparation of 7-amino-cephalosporanic acid. Helv Chim Acta 1968;51:1108–19.
[40] Palomo JM, Segura RL, Fernández-Lorente G, Pernas M, Rua ML, Guisán JM, etal. Purification, immobilization, and stabilization of a lipase from Bacillus ther-mocatenulatus by interfacial adsorption on hydrophobic supports. BiotechnolProg 2004;20(2):630–5.
[41] Bradford MM. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein dye binding. AnalBiochem 1976;72(1–2):248–54.
[42] López-Gallego F, Betancor L, Hidalgo A, Mateo C, Guisán JM, Fernández-LafuenteR. Optimization of an industrial biocatalyst of glutaryl acylase: stabilization ofthe enzyme by multipoint covalent attachment onto new amino-epoxy sepa-beads. J Biotechnol 2004;111(2):219–27.
[43] López-Gallego F, Montes T, Fuentes M, Alonso N, Grazu V, Betancor L, et al.Improved stabilization of chemically aminated enzymes via multipoint cova-lent attachment on glyoxyl supports. J Biotechnol 2005;116:1–10.
[44] Rosell CM, Fernández-Lafuente R, Guisán JM. Modification of enzyme proper-ties by the use of inhibitors during their stabilization by multipoint covalentattachment. Biocatal Biotransform 1995;12:67–76.
[45] Laemmli UK. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature 1970;277:680–5.
[46] Kutzbach C, Rauenbusch E. Preparation and general properties of crystallinepenicillin acylase from Escherichia coli ATCC 11105. Hoppe-Seyler’s Z PhysiolChem 1974;355:45–53.
[47] Duggleby HJ, Tolley SP, Hill GP, Dodson EJ, Dodson G, Moody PC. Peni-cillin acylase has a single-amino-acid catalytic centre. Nature 1995;373:264–8.
[48] Carrasco-López C, Godoy C, de las Rivas B, Fernández-Lorente G, Palomo JM,Guisán JM, et al. Activation of bacterial thermoalkalophilic lipases is spurredby dramatic structural rearrangements. J Biol Chem 2009;284:4365–72.
[49] Matsuda A, Matsuyama K, Yamamoto K, Ichikawa S, Komatsu K-I. Cloning andcharacterization of the genes for two distinct cephalosporin acylases from aPseudomonas strain. J Bacteriol 1987;169:5815–20.
[50] Aramori I, Fukagawa M, Tsumura M, Iwami M, Isogai T, Ono H, et al. Cloningand nucleotide sequencing of new glutaryl 7-ACA and cephalosporin C acylasegenes from Pseudomonas strains. J Ferment Bioeng 1991;72:232–43.
[51] Fernandez-Lorente G, Godoy C, Lopez-Gallego F, Grazu V, de las Rivas B, PalomoJM, et al. Solid-phase chemical amination of a lipase from Bacillus thermo-catenulatus to improve its stabilization via covalent immobilization on highlyactivated glyoxyl-agarose. Biomacromolecules 2008;9:2553–61.
[52] Girelli AM, Mattei E, Messina A. Immobilized tyrosinase reactor for on-lineHPLC application. Development and characterization. Sens Actuator B-Chem2007;121:515–21.
[53] Hornsey VS, Prowse CV, Pepper DS. Reductive amination for solid-phase cou-pling of protein. A practical alternative to cyanogen bromide. J ImmunolMethods 1986;93:83–8.
[54] Weingarten S, Thiem J. Facile formation of novel carbohydrate-amino acid con-jugates by reductive amination. Synlett 2003:1052–4.
[55] Burteau N, Burton S, Crichton RR. Stabilisation and immobilisation of penicillinamidase. FEBS Lett 1989;258:185–9.




















