
The methyltransferase YfgB/RlmN is responsible for modification of adenosine 2503 in 23S rRNA
10
10.1261/rna.814408 Access the most recent version at doi: 2008 14: 98-106; originally published online Nov 19, 2007; RNA Seok-Ming Toh, Liqun Xiong, Taeok Bae and Alexander S. Mankin adenosine 2503 in 23S rRNA The methyltransferase YfgB/RlmN is responsible for modification of References http://www.rnajournal.org/cgi/content/full/14/1/98#References This article cites 55 articles, 20 of which can be accessed free at: service Email alerting click here top right corner of the article or Receive free email alerts when new articles cite this article - sign up in the box at the Notes http://www.rnajournal.org/subscriptions/ go to: RNA To subscribe to © 2008 RNA Society Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.org Downloaded from
Transcript of The methyltransferase YfgB/RlmN is responsible for modification of adenosine 2503 in 23S rRNA

10.1261/rna.814408Access the most recent version at doi: 2008 14: 98-106; originally published online Nov 19, 2007; RNA
Seok-Ming Toh, Liqun Xiong, Taeok Bae and Alexander S. Mankin
adenosine 2503 in 23S rRNAThe methyltransferase YfgB/RlmN is responsible for modification of
References
http://www.rnajournal.org/cgi/content/full/14/1/98#References
This article cites 55 articles, 20 of which can be accessed free at:
serviceEmail alerting
click heretop right corner of the article or Receive free email alerts when new articles cite this article - sign up in the box at the
Notes
http://www.rnajournal.org/subscriptions/ go to: RNATo subscribe to
© 2008 RNA Society
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

The methyltransferase YfgB/RlmN is responsible
for modification of adenosine 2503 in 23S rRNA
SEOK-MING TOH,1 LIQUN XIONG,1 TAEOK BAE,2 and ALEXANDER S. MANKIN1
1Center for Pharmaceutical Biotechnology, University of Illinois, Chicago, Illinois 60607, USA2Indiana University School of Medicine, Gary-Northwest, Gary, Indiana 46408, USA
ABSTRACT
A2503 in 23S rRNA of the Gram-negative bacterium Escherichia coli is located in a functionally important region of theribosome, at the entrance to the nascent peptide exit tunnel. In E. coli, and likely in other species, this adenosine residue is post-transcriptionally modified to m2A. The enzyme responsible for this modification was previously unknown. We identified E. coliprotein YfgB, which belongs to the radical SAM enzyme superfamily, as the methyltransferase that modifies A2503 of 23S rRNAto m2A. Inactivation of the yfgB gene in E. coli led to the loss of modification at nucleotide A2503 of 23S rRNA as revealed byprimer extension analysis and thin layer chromatography. The A2503 modification was restored when YfgB protein wasexpressed in the yfgB knockout strain. A similar protein was shown to catalyze post-transcriptional modification of A2503 in23S rRNA in Gram-positive Staphylococcus aureus. The yfgB knockout strain loses in competition with wild type in a co-growthexperiment, indicating functional importance of A2503 modification. The location of A2503 in the exit tunnel suggests itspossible involvement in interaction with the nascent peptide and raises the possibility that its post-transcriptional modificationmay influence such an interaction.
Keywords: exit tunnel; peptidyl transferase; post-transcriptional modifications; radical SAM methyl transferase; ribosome
INTRODUCTION
Ribosomal RNA plays a critical role in protein synthesis. Inthe ribosome, rRNA adopts an intricate spatial structurethat places chemical groups of rRNA nucleotides in theproper position for carrying out such elaborate functions asmonitoring codon–anticodon interactions, catalyzing pep-tide bond formation, binding ligands and cofactors, etc. Invitro transcribed rRNA can be assembled into small andlarge ribosomal subunits and can carry out protein syn-thesis reasonably well, suggesting that the main functionalproperties of rRNA are dictated solely by its nucleotidesequence and spatial arrangement (Krzyzosiak et al. 1987;Green and Noller 1999; Khaitovich et al. 1999). Yet in thecell, rRNA carries a number of post-transcriptional mod-ifications at specific nucleotide residues. The clustering ofthe modified nucleotides in functionally critical regions ofrRNA indicate their importance for ribosome activity(Ofengand and Fournier 1998). However, with a few
exceptions, the exact role of these modified nucleotidesremains largely unknown. Analyzing functional propertiesof the ribosome lacking individual modifications is themost direct way to elucidate the contribution of post-transcriptional modifications to translation. For that, oneneeds to know the enzymes that alter the chemical nature ofrRNA nucleotides.
In bacteria, rRNA modifications involve mainly methyl-ation of various rRNA residues and pseudouridylation. Inthe case of Escherichia coli 23S rRNA, 25 modified nucleo-tides have been mapped (Andersen and Douthwaite 2006).Fourteen of these modifications are associated with domainV, the main component of the peptidyl transferase center(PTC) (Fig. 1; Noller 1984). The pseudouridine synthasesresponsible for all five pseudouridine modifications in thisregion (positions 2457, 2504, 2580, 2604, and 2605) havebeen identified (Conrad et al. 1998; Huang et al. 1998; DelCampo et al. 2001). Of the 9 remaining modified nucleo-tides, 7 carry methylations (m6A2030, m7G2069, Gm2251,m2G2445, Cm2498, m2A2503, and Um2552), one is adihydrouridine (D2449), and one is a predicted 2-thio-cytidine (s2C2501) (Andersen et al. 2004). So far, only threeof the methyltransferases responsible for post-transcrip-tional methylation of nucleotides in domain V of 23S rRNA
Reprint requests to: Alexander S. Mankin, Center for PharmaceuticalBiotechnology, m/c 870, University of Illinois, 900 S. Ashland Avenue,Chicago, IL 60607, USA; e-mail: [email protected]; fax: (312) 413-9303.
Article published online ahead of print. Article and publication date are athttp://www.rnajournal.org/cgi/doi/10.1261/rna.814408.
98 RNA (2008), 14:98–106. Published by Cold Spring Harbor Laboratory Press. Copyright � 2008 RNA Society.
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

have been identified—RlmE (synonymous to FtsJ or RrmJ),which generates 29-O-methylated uridine at position 1552(Bugl et al. 2000; Caldas et al. 2000), RlmL (synonymousto YcbY), which methylates the N2 position of G2445(Lesnyak et al. 2006), and RlmB (synonymous with YifH),which generates 29-O-methyl guanine at position 2251(Lovgren and Wikstrom 2001).
One of the methylated nucleotides in the PTC is A2503.In E. coli, A2503 is post-transcriptionally modified tom2A (Kowalak et al. 1995). A2503 is located at the entrance ofthe exit tunnel (Ban et al. 1998; Harms et al. 2001; Schuwirthet al. 2005; Selmer et al. 2006); however, the precise functionalrole of m2A2503 has not been explored. This nucleotide isfairly close to the binding sites of several PTC-targetingantibiotics, including clindamycin, carbomycin, streptogra-min A, anisomycin, and linezolid (Schlunzen et al. 2001;Hansen et al. 2002; Tu et al. 2005). Mutations of A2503 conferresistance to chloramphenicol and streptogramins (Vester andGarrett 1988; Porse and Garrett 1999) and its hypermethyla-tion by Cfr methyltransferase in staphylococci or E. coli leadsto resistance against a number of PTC-targeting antibiotics(Kehrenberg et al. 2005; Long et al. 2006).
Here, we report the identification of the methyltransfer-ase, RlmN, which is responsible for generation of m2A2503in the ribosomes of Gram-positive and Gram-negativebacteria.
RESULTS
The E. coli gene yfgB encodes an enzyme responsiblefor methylation of A2503 in 23S rRNA
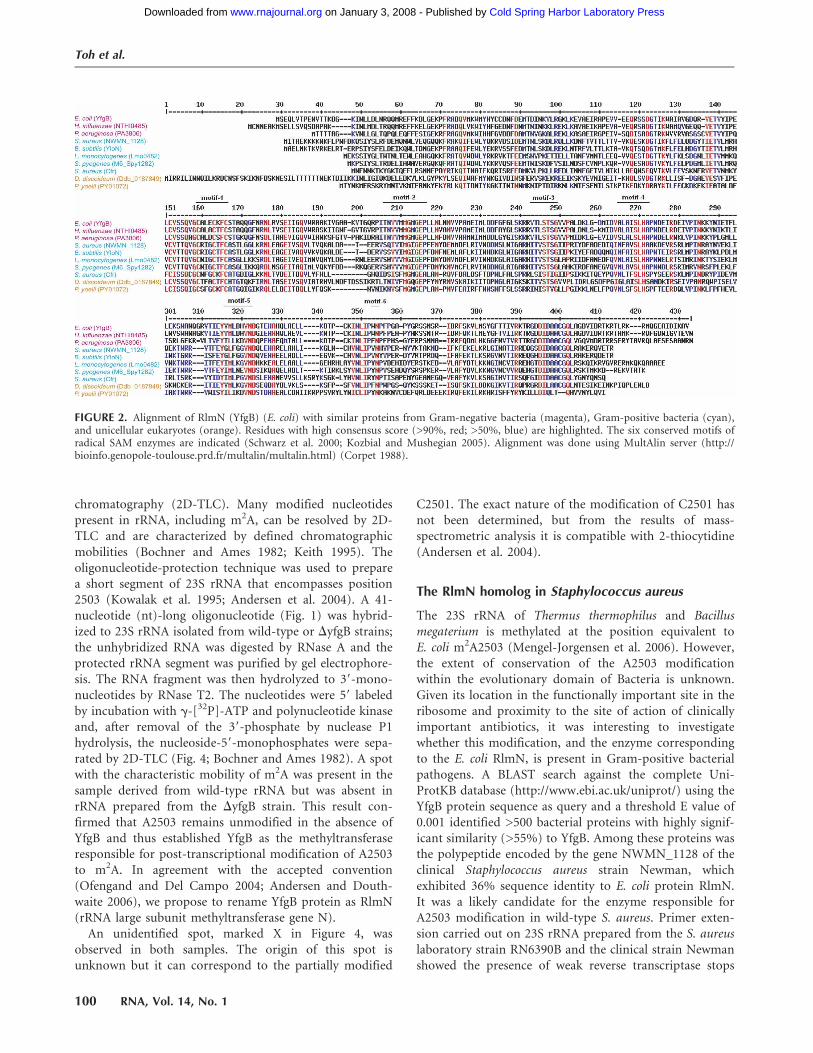
S-adenosylmethionine- (SAM) dependent methyltransfer-ase Cfr, whose gene was found in staphylococcal isolatesfrom animal sources and more recently in a clinical strainof Staphylococcus aureus, confers resistance to a number ofPTC-targeting inhibitors of protein synthesis (Kehrenberget al. 2005; Long et al. 2006; Toh et al. 2007). Cfr-mediatedantibiotic resistance results from hypermethylation ofA2503 in 23S rRNA. Sequence analysis of the staphylococ-cal cfr gene showed its significant homology (32% sequenceidentity) with an E. coli gene yfgB that encodes a proteinof unknown function (Schwarz et al. 2000; Kozbial andMushegian 2005). Similar to Cfr, YfgB contains thecysteine-rich motif C124ALECKFC, which is a characteristicsignature of the superfamily of proteins known as radicalSAM enzymes (motif 1 in Fig. 2; Schwarz et al. 2000).Importantly, homology between Cfr and YfgB extends intothe C-terminal portion of the protein, which in radicalSAM enzymes is often responsible for substrate binding,suggesting that Cfr and YfgB may operate upon the same orsimilar substrates. Therefore, we hypothesized that E. coliyfgB could encode the elusive methyltransferase responsiblefor natural modification of A2503 in the 23S rRNA ofE. coli to m2A.
To test this hypothesis we used the strain JW2501 fromthe ‘‘Keio’’ collection of E. coli clones containing single-gene knockouts of all nonessential genes in the E. coli K-12strain BW25113 (Baba et al. 2006). In strain JW2501 of thecollection, the kanamycin resistance cassette replaced theyfgB gene. Inactivation of the yfgB gene in strain JW2501was confirmed by PCR.
The modification status of A2503 in the JW2501 strainwas initially analyzed by primer extension. In wild-typeE. coli, the methylation of A2503 at C2 results in a weakreverse transcriptase stop (Fig. 3A). When 23S rRNAextracted from the DyfgB strain JW2501 was used as atemplate, the band corresponding to the reverse transcrip-tase stop at m2A2503 was not observed, indicating the lossof m2A2503 modification in this strain. When the DyfgBstrain was transformed with a plasmid overexpressing theYfgB protein (Kitagawa et al. 2005), the reverse transcrip-tase stop reappeared (Fig. 3B) establishing a causativerelationship between the presence of yfgB and post-tran-scriptional modification of A2503 in E. coli.
We further investigated the presence of m2A2503 mod-ification in the DyfgB strain by two-dimensional thin-layer
FIGURE 1. Secondary structure of domain V of E. coli 23S rRNA(Cannone et al. 2002). The sites of post-transcriptional modificationare indicated. Green: nucleotides that carry methylations; blue:pseudouridines, dihydoruridine, or 2-thiocytidine. m2A2503 is circledand in red. A DNA oligonucleotide used to prepare a fragment of 23SrRNA for the nucleotide analysis is shown by a solid line.
RlmN methylates A2503 in 23S rRNA
www.rnajournal.org 99
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from
chromatography (2D-TLC). Many modified nucleotidespresent in rRNA, including m2A, can be resolved by 2D-TLC and are characterized by defined chromatographicmobilities (Bochner and Ames 1982; Keith 1995). Theoligonucleotide-protection technique was used to preparea short segment of 23S rRNA that encompasses position2503 (Kowalak et al. 1995; Andersen et al. 2004). A 41-nucleotide (nt)-long oligonucleotide (Fig. 1) was hybrid-ized to 23S rRNA isolated from wild-type or DyfgB strains;the unhybridized RNA was digested by RNase A and theprotected rRNA segment was purified by gel electrophore-sis. The RNA fragment was then hydrolyzed to 39-mono-nucleotides by RNase T2. The nucleotides were 59 labeledby incubation with g-[32P]-ATP and polynucleotide kinaseand, after removal of the 39-phosphate by nuclease P1hydrolysis, the nucleoside-59-monophosphates were sepa-rated by 2D-TLC (Fig. 4; Bochner and Ames 1982). A spotwith the characteristic mobility of m2A was present in thesample derived from wild-type rRNA but was absent inrRNA prepared from the DyfgB strain. This result con-firmed that A2503 remains unmodified in the absence ofYfgB and thus established YfgB as the methyltransferaseresponsible for post-transcriptional modification of A2503to m2A. In agreement with the accepted convention(Ofengand and Del Campo 2004; Andersen and Douth-waite 2006), we propose to rename YfgB protein as RlmN(rRNA large subunit methyltransferase gene N).
An unidentified spot, marked X in Figure 4, wasobserved in both samples. The origin of this spot isunknown but it can correspond to the partially modified
C2501. The exact nature of the modification of C2501 hasnot been determined, but from the results of mass-spectrometric analysis it is compatible with 2-thiocytidine(Andersen et al. 2004).
The RlmN homolog in Staphylococcus aureus
The 23S rRNA of Thermus thermophilus and Bacillusmegaterium is methylated at the position equivalent toE. coli m2A2503 (Mengel-Jorgensen et al. 2006). However,the extent of conservation of the A2503 modificationwithin the evolutionary domain of Bacteria is unknown.Given its location in the functionally important site in theribosome and proximity to the site of action of clinicallyimportant antibiotics, it was interesting to investigatewhether this modification, and the enzyme correspondingto the E. coli RlmN, is present in Gram-positive bacterialpathogens. A BLAST search against the complete Uni-ProtKB database (http://www.ebi.ac.uk/uniprot/) using theYfgB protein sequence as query and a threshold E value of0.001 identified >500 bacterial proteins with highly signif-icant similarity (>55%) to YfgB. Among these proteins wasthe polypeptide encoded by the gene NWMN_1128 of theclinical Staphylococcus aureus strain Newman, whichexhibited 36% sequence identity to E. coli protein RlmN.It was a likely candidate for the enzyme responsible forA2503 modification in wild-type S. aureus. Primer exten-sion carried out on 23S rRNA prepared from the S. aureuslaboratory strain RN6390B and the clinical strain Newmanshowed the presence of weak reverse transcriptase stops
FIGURE 2. Alignment of RlmN (YfgB) (E. coli) with similar proteins from Gram-negative bacteria (magenta), Gram-positive bacteria (cyan),and unicellular eukaryotes (orange). Residues with high consensus score (>90%, red; >50%, blue) are highlighted. The six conserved motifs ofradical SAM enzymes are indicated (Schwarz et al. 2000; Kozbial and Mushegian 2005). Alignment was done using MultAlin server (http://bioinfo.genopole-toulouse.prd.fr/multalin/multalin.html) (Corpet 1988).
Toh et al.
100 RNA, Vol. 14, No. 1
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

corresponding to the possible occurrence of post-transcrip-tional modification at A2503 (Fig. 3C). However, when 23SrRNA prepared from the S. aureus Newman strain in whichthe NWMN_1128 gene was inactivated by transposoninsertion (Bae et al. 2004) was analyzed, the correspondingband was no longer present (Fig. 3C). This result indicatedthat the position equivalent to E. coli A2503 is post-transcriptionally modified in S. aureus and that the proteinencoded by the NWMN_1128 gene is responsible for thismodification. By analogy with E. coli, we expect that S.aureus carries m2A modification at position 2503 of 23SrRNA.
The lack of methylation of A2503 in E. coli 23S rRNAreduces cell fitness
Inactivation of rlmN (yfgB) did not notably affect growth ofE. coli in LB medium (data not shown). The rlmN� cellsgrew with a doubling time of 43 min, comparable to that ofwild-type BW25113 cells (41 min) or to a control strainwhere the kanamycin marker replaced the xylA gene, whichis nonessential for growth in rich medium (44 min).
However, in a co-growth competition experiment, therlmN knockout mutant showed reduced fitness comparedto wild type (Fig. 5). In a mixed culture initially containingequal proportions of wild-type and mutant cells, therepresentation of the rlmN mutant reduced to 6% aftersix dilutions (z50 doublings). Similarly, rlmN knockoutcells also lost in co-growth competition with the controlxylA knockout strain. Thus, the absence of m2A2503 modi-fication produces a subtle but significant effect on cellfitness apparently through its effect on protein synthesis orribosome assembly.
The lack of m2A2503 modification affectsantibiotic sensitivity
Acquired antibiotic resistance enzymes often target rRNAand prevent drug binding by modifying specific rRNAresidues, usually via attaching one or several methyl groups(Weisblum 1995; Mann et al. 2001; Treede et al. 2003;Kehrenberg et al. 2005). A2503 is the target of modificationby Cfr methyltransferase, which renders cells resistant tophenicols, lincosamides, oxazolidinones, pleuromutilins, andstreptogramin A antibiotics (Long et al. 2006). In somecases, the lack of natural modification at specific rRNApositions may influence susceptibility of bacterial cells toantibiotics (Helser et al. 1972; Van Buul et al. 1983; Johansenet al. 2006; Okamoto et al. 2007). Therefore, we testedsusceptibility of the rlmN knockout E. coli mutant to anarray of peptidyl transferase-targeting antibiotics. ThoughE. coli is naturally resistant to many antibiotics owing toactive efflux, high concentrations of the drugs do inhibitprotein synthesis and cell growth (Xiong et al. 2000). TheE. coli rlmN knockout strain showed a small but repro-ducible twofold increased susceptibility to tiamulin, hygro-mycin A, and sparsomycin compared to the wild-type strain(Table 1). Inactivation of rlmN in S. aureus resulted in two-fold increased susceptibility to linezolid.
FIGURE 3. Primer extension analysis of 23S rRNA. (A) rRNA from acontrol wild-type E. coli strain BW25113 (wt) and yfgB (rlmN)knockout mutant strain (DyfgB); sequencing lanes (A,G) are indi-cated. (B) rRNA from the E. coli yfgB knockout strain transformedwith the yfgB expression plasmid (DyfgB/pyfgB) or with the emptyvector (DyfgB/pCA24N); sequencing lane (A) is marked. (C) rRNAfrom S. aureus: ‘‘RN6390B,’’ control laboratory strain; ‘‘CM05’’clinical isolate CM05 that carries the cfr gene (Toh et al. 2007);‘‘1128-KO,’’ S. aureus Newman strain with the insertionally inacti-vated gene NWMN_1128, ‘‘control-KO,’’ S. aureus Newman strainwith the insertionally inactivated neutral gene NWMN-1912;‘‘NWMN,’’ wild-type Newman strain.
FIGURE 4. 2D-TLC analysis of nucleotides from the yfgB knockoutand wild-type E. coli 23S rRNA fragment. The positions of 59 mono-phosphate UV markers are indicated by dashed-line circles. Theradioactive spots used for alignment of the PhosphorImager scanswith the TLC plates are circled with a solid line.
RlmN methylates A2503 in 23S rRNA
www.rnajournal.org 101
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

DISCUSSION
We have identified the enzyme responsible for ‘‘natural’’post-transcriptional modification of A2503 to m2A inE. coli 23S rRNA. Cells lacking the gene yfgB (rlmN) carriedunmodified A at position 2503 of 23S rRNA. Reintroduc-tion of rlmN into the knockout strain restored modification.These result unequivocally established RlmN as the methyl-transferase that modifies A2503 in 23S rRNA to m2A.
There are 14 post-transcritionally modified purineresidues in rRNA of the E. coli ribosome (Andersen andDouthwaite 2006). All the post-transcriptional modifica-tions of purines in rRNA entail transfer of one or severalmethyl groups to the nitrogen base or to the ribose. In mostcases, the methyl-accepting groups (endo- or exocyclicnitrogen atoms of the purine base or 29oxygen of theribose sugar) are nucleophilic and the methylation reactionproceeds through a classic alkylation mechanism involvingthe methyl group of SAM. The only nucleotide in rRNAthat is methylated at the carbon atom of the purine base isA2503. Low nucleophilicity of the adenine’s C2 makes thisreaction much more challenging in chemical terms than the
classic methylation reaction and calls for a principallydifferent reaction mechanism (Wang and Frey 2007). It isnot surprising, therefore, that the enzyme responsible forthis reaction, RlmN, exhibits little similarity to otherknown methyltransferases that operate on rRNA.
The amino acid sequence of RlmN shows characteristicsignatures of a superfamily of proteins known as radicalSAM enzymes (marked as ‘‘motifs’’ in Fig. 2) (Sofia et al.2001; Kozbial and Mushegian 2005). All radical SAMenzymes contain the cysteine-rich motif CxxxCxxC (inRlmN: C124ALECKFC, motif 1 in Fig. 2), which is involvedin chelation of the [4Fe–4S]1+ cluster. The cluster initiates areductive cleavage of SAM, resulting in formation of 59
deoxyadenosyl radical. This radical then abstracts a hydro-gen atom from a chemically unreactive CH group therebyleading to its chemical activation. The general role of SAMin radical SAM enzymes is thus of a substrate for generationof reactive radical. In regards to RlmN, this mechanism willimply that one molecule of SAM will be consumed togenerate a 59deoxyadenosyl radical that will activate the C2aromatic carbon atom of the adenine base. The source of themethyl group that is then transferred to the activated C2carbon atom of adenine 2503 is unknown, but by analogywith some other radical SAM methyltransferases, one canexpect that a second molecule of SAM may serve as themethyl donor (Pierrel et al. 2004; Hernandez et al. 2007).
We have shown that a protein with the structure, andlikely function, similar to that of RlmN of Gram-negativeE. coli is encoded in the genome of phylogenetically distantGram-positive S. aureus. Though we have not explicitlyanalyzed the phylogenetic distribution of rlmN-like genesamong sequenced genomes, BLAST search shows that closeRlmN homologs are encoded in chromosomal DNA ofmany Gram-negative and Gram-positive bacteria (Fig. 2).The presence of methylated A2503 in 23S rRNA wasexperimentally detected in three bacterial species, E. coli,T. thermophilus, and B. megaterium (Kowalak et al. 1995;Mengel-Jorgensen et al. 2006). Proteins with considerable(30%–50%) similarity to RlmN are also encoded ingenomes of several unicellular eukaryotes (Fig. 2) indicat-ing that the adenine residue corresponding to the bacterial
TABLE 1. Antibiotic susceptibility of the RlmN knockout strains in E. coli and S. aureus compared with the control strains
Strain
MIC (mg/mL)
Tiamulin Chloramphenicol Clindamycin Linezolid Hygromycin A Sparsomycin Streptomycin
E. coli (wt)a 1024 8 512 1024 512 32 32E. coli RlmN� 512 8 512 1024 256 16 32S. aureus NWMN_1912b <1 8 >1024 4 128 32 128S. aureus NWMN_1128c <1 8 >1024 2 128 32 128
aStrain BW25113.bStrain Newman with a transposon insertion in a ‘‘neutral’’ gene NWMN_1912.cStrain Newman with a transposon insertion in a gene NWMN_1128 homologous to the E. coli RlmN.
FIGURE 5. Growth of DyfgB cells in competition with the initiallyequal number of wild-type (closed boxes) and DxylA mutant cells (openboxes). The percent of DyfgB cells in the population was determined ateach cycle by plating as described in Materials and Methods.
Toh et al.
102 RNA, Vol. 14, No. 1
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

A2503 may be post-transcriptionally modified in thesespecies. BLAST search in the archaeal genomes revealedproteins with only limited similarity to RlmN. However, atleast one report mentioned the presence of m2A2503 in 23SrRNA of an archaeon Haloarcula marismortui (cited inMengel-Jorgensen et al. 2006). Thus, it appears that themodification of A2503 to m2A is conserved among evolu-tionarily distant species.
A rather ‘‘difficult’’ chemistry involved in formation ofm2A2503 implies that there must have been a compellingreason for the cells to evolve an enzyme catalyzing thisreaction. Furthermore, broad distribution of RlmN homo-logs among species indicates that there is strong evolution-ary pressure to maintain post-transcriptional modificationof A2503 in rRNA. What could the functional purpose ofhaving adenosine at position 2503 in 23S rRNA modifiedbe? A2503 is located at the entrance to the nascent peptideexit tunnel and is exposed in the tunnel lumen (Ban et al.2000). It tops the stack of three adenines (A2058/A2059/A2503) (Fig. 6). These nucleotides, which are located at theconstriction of the nascent peptide tunnel, might be a partof the mechanism that monitors the sequence of thenascent peptide (Mankin 2006). Indeed, mutations inA2058 were shown to affect the ribosome stalling duringtranslation of the SecM nascent peptide, which depends onthe ribosome-nascent peptide interaction (Nakatogawa andIto 2002). In the crystallographic structures of the largeribosomal subunits of H. marismortui, T. thermophilus, andE. coli, the base of A2503 is in syn conformation but in the
Deinococcus radiodurans 50S subunit, A2503 was modeledin anti conformation (Ban et al. 1998; Harms et al. 2001;Schuwirth et al. 2005; Selmer et al. 2006). Since themodification status of A2503 in D. radiodurans is unknown(even though D. radiodurans genomes encodes a proteinhomologous to RlmN), it is unclear whether this differenceresults from the lack of A2503 methylation or is a merereflection of insufficient resolution. Nevertheless, a possiblesyn–anti transition of A2503 or, in general, the exactorientation of the base should be affected by the presenceof the C2 methyl group (Saenger 1984). Directly, orindirectly (via changing conformation of A2059 and/orA2058), the precise placement of A2503 base may influencethe contacts between the tunnel wall and the nascentpeptide and could be a part of the ribosome responsemechanism to specific nascent peptides in the tunnel.
RlmN was initially identified on the basis of its homol-ogy with Cfr, an enzyme that hypermethylates A2503 andrenders cells resistant to several antibiotics, inhibitors of thepeptidyl transferase activity (Schwarz et al. 2000, 2004). Incells that acquire cfr, A2503 has not one extra methylgroup, as m2A2503 in wild-type rRNA, but two. However,the exact placement of the second methyl group in the Cfr-modified A2503 is unknown (Kehrenberg et al. 2005).Given that, similar to RlmN, Cfr belongs to the radicalSAM enzyme superfamily, one would expect that the Cfr-catalyzed methylation reaction should also involve a CHcenter. The only aromatic carbon atom available formethylation in m2A is C8. Even though C8-methylatedadenine has not yet been found in natural RNAs, it ispossible that Cfr utilizes the radical mechanism to meth-ylate this chemically inert center. Another possibility is thatCfr converts m2A to 2-ethyladenosine. Though unusual,such a modification appears to be compatible with thepublished results of mass spectrometry of Cfr-modified 23SrRNA (Kehrenberg et al. 2005).
Hypermethylation of A2503 by Cfr confers resistance toseveral drugs that bind in the vicinity of this nucleotide inthe peptidyl transferase center (Long et al. 2006). The lackof A2503 natural modification in the E. coli rlmN knockoutstrain led to a modest increase in susceptibility to tiamulin,hygromycin A, and sparsomycin, confirming that themodification status of A2503 affects antibiotic sensitivity.This result is in line with a growing perception that the lossof natural post-transcriptional modifications in rRNA mayaffect susceptibility of bacterial cells to antibiotics, a notionthat may have clinical implications (Helser et al. 1972; VanBuul et al. 1983; Johansen et al. 2006; Okamoto et al. 2007).
MATERIALS AND METHODS
Strains and plasmids
The collection of gene knockout mutants of the E. coli strainBW25113 (Keio collection) was obtained from Nara Institute of
FIGURE 6. Position of the A2503 (red) in the E. coli ribosome(Schuwirth et al. 2005) relative to the tRNA substrates docked in the Psite (light green) and A site (light blue) (Leach et al. 2007). A2503 islocated within 9 A of peptidyl tRNA and its base stacks on top ofA2059 and A2058, located at the constriction of the nascent peptidetunnel. The C2 methyl group added to A2503 by RlmN is indicated bya red ball.
RlmN methylates A2503 in 23S rRNA
www.rnajournal.org 103
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

Science and Technology (Baba et al. 2006). The strain JW2501 inthe collection, which has the yfgB gene replaced with thekanamycin resistance cassette, was used in the experiments. Thegene replacement was verified by PCR analysis. The strainJW3537, in which the kanamycin resistance cassette replaces xylAgene dispensable for growth in rich media, was used as control insome experiments. Plasmid expressing yfgB was from the ASKAlibrary strain JW2501 (Kitagawa et al. 2005).
S. aureus strains used in this work include RN6390B (Novicket al. 1993), Newman (wild type), and the following transpo-son mutants: NWMN_1128 (the yfgB gene homolog) andNWMN_1912 (a prophage gene), the control mutant. Themutants were generated by transducing the transposon insertionsfrom the FNJ-0032 and FNJ-3200 strains of the Phoenixlibrary of bursa aurealis mutants (Bae et al. 2004) into Newmanwild-type strain with the staphylococcal phage F85.
Isolation of total cellular RNA and primerextension analysis
For isolation of total RNA, E. coli or S. aureus cells were grown inLB broth or tryptic soy broth, respectively, to an optical density ofA550 = 1. E. coli RNA was isolated using the TRIzol kit(Invitrogen) whereas S. aureus RNA was isolated using theUltraClean Microbial RNA Isolation Kit (Mo Bio Laboratories)according to the manufacturer’s protocols.
Primer extension analysis was carried out following the pub-lished protocol (Merryman and Noller 1998) with some mod-ifications. Ten picomoles of the DNA primer TCGCGTACCACTTTA, complementary to the positions 2563–2577 of 23SrRNA (E. coli numbering), were 32P-59-terminally labeled byincubation in the final volume of 10 mL with 10 mCi (1.7 pmol)g-[32P] ATP (6000 Ci/mmol) and 10 units of polynucleotidekinase in a 13 polynucleotide kinase buffer (Fermentas). Reactionwas incubated at 37°C for 30 min and then stopped by incubationfor 2 min at 90°C.
We combined 0.5 pmol of labeled primer with 2 mg oftotal cellular RNA in 4.5 mL of hybridization buffer (50 mMHEPES-KOH at pH 7, 100 mM KCl). Reactions were incubatedat 90°C for 1 min and then cooled over z10 min to 45°C.Annealed reactions were then centrifuged for 15 sec to collect anycondensate.
An extension reaction premix contained 0.65 mL of 103
reaction buffer (1.3 M Tris-HCl at pH 8.5, 100 mM MgCl2,100 mM DTT), 1.5 mL of dNTP mix (1 mM dGTP, 1 mM dATP,1 mM dTTP, and 1 mM dCTP), 0.75 mL of RNase-free water,and 0.1 mL (3 units) of AMV reverse transcriptase (SeikagakuAmerica) per reaction. The amount of premix was adjustedaccording to the number of samples to be analyzed. We added 3mL of the pre-mix to each reaction tube with the annealed primer/RNA complex. Samples were incubated at 42°C for 30 min.Following incubation, 120 mL of stop buffer (84 mM NaOAc atpH 5.5, 70% EtOH, 0.8 mM EDTA) were added to each tubefollowed by centrifugation at maximum speed for 10 min at roomtemperature. After complete removal of supernatant and air-drying of the tubes for 2–3 min, the pellets were resuspended in5 mL of formamide loading dye (Sambrook et al. 1989). Weloaded 2 mL onto a 5-mm- wide slot of a 20 cm 3 40 cm 3 0.25mM, 6% urea-polyacrylamide gel. The gel was run at 40 W untilthe bromophenol blue dye reached the bottom of the gel. The gel
was transferred to Whatman 3MM paper, dried, and exposed on aphosphorimager screen. The screen was scanned on a MolecularDynamics PhosphorImager, and the band intensities were quan-tified using Molecular Dynamics Image Quant software.
Preparation of an rRNA segmentencompassing A2503
E. coli cells were grown in liquid LB medium to an optical densityof A550 = 0.4 and harvested by centrifugation. Cells were washedand resuspended in Buffer A (20 mM Tris-HCl at pH 7.5, 100 mMNH4Cl, 10 mM MgCl2, 0.5 mM EDTA, 6 mM BME). The cellswere lysed by passing them through a French press, and cell debriswas removed by centrifugation (15 min at 16,000 rpm, then 10min at 24,000 rpm in the Beckman JA25 rotor, at 4°C). Ribosomeswere collected from the supernatants by centrifugation at 33,000rpm for 22 h at 4°C in a Beckman Ti70 rotor. The pellet wasresuspended in resuspension buffer (50 mM Tris-HCl at pH 7.5,150 mM NH4Cl, 5 mM MgCl2, 6 mM BME) and stored at �80°C.We diluted 330 A260 of ribosomes in extraction buffer 0.3 MNaOAc (pH 5.5) to the final volume of 1000 mL, and rRNA wasisolated by three repeated extractions with equal volumes ofphenol/chloroform followed by ethanol precipitation.
To isolate a defined rRNA sequence, 1200 pmol of a 41-nt DNAoligonucleotide, GATGTGATGAGCCGACATCGAGGTGCCAAACACCGCCGTCG, complementary to the 23S rRNA segment2480–2520 of E. coli 23S rRNA, were combined with 400 pmolof total rRNA and 60 mL hybridization buffer (250 mM HEPES,500 mM KCl at pH 7) in a total volume of 200 mL. The mixturewas incubated for 5 min at 90°C and then allowed to cool to roomtemperature over 1.5 h. We added 0.00625 mg of RNase A (Sigma-Aldrich)/pmol rRNA, followed by a 50 min incubation at 37°C.The reaction was extracted once with phenol/chloroform. Nucleicacids were ethanol precipitated, dissolved in water, and incubatedwith RQ1 DNase (Promega) at 37°C for 15 min. Following an-other phenol/chloroform extraction and ethanol precipitation, theprotected rRNA fragment was purified on a 13% polyacrylamidegel containing 7 M urea. Bands were visualized by ethidium bro-mide staining, and the RNA band was excised and eluted over-night at 24°C in 0.3 M NaOAc (pH 5.5). RNA was extracted withphenol, phenol/chloroform, chloroform and ethanol precipitated.
Analysis of nucleotide composition by 2D-TLC
The isolated rRNA fragment (0.1–0.5 mg) was denatured byincubation at 65°C for 5 min and then placed on ice. Thedenatured RNA was digested to nucleoside 39 phosphates with10 units of RNase T2 (Invitrogen) for 15 min at 37°C. The 39
mononucleotides were then 59 32P phosphorylated by incubationwith 10 mCi (1.7 pmol) g-[32P] ATP (6000 Ci/mmol) and 10 unitspolynucleotide kinase in a 13 polynucleotide kinase buffer(Fermentas). The reaction was extracted with phenol-chloro-form-isoamyl alcohol (25:24:1) and chloroform and then incu-bated with 2 mg of nuclease P1 (USBiological) for 3 h to overnightat 37°C. Prior to loading on TLC plates, the radiolabeled samplewas mixed with 1 mL of the marker comprising 1 A260 of each of59 AMP, GMP, CMP, and UMP.
Two-dimensional separation of the 59-[32P] nucleoside mono-phosphates was performed on glass-backed polyethyleneiminecellulose F plates (Merck). Plates (20 cm 3 10 cm) were developed
Toh et al.
104 RNA, Vol. 14, No. 1
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

at room temperature (first dimension: 20 cm, second dimen-sion:10 cm). The solvent systems used were the following: firstdimension: isobutyric acid: NH4OH conc: H2O=66:1:33, seconddimension: isopropanol: HCl: H2O=70:15:15. Identification ofeach radiolabeled spot on the plate was performed by comparisonwith those in published reference maps (Keith 1995).
Cell growth experiments and microbiological testing
The growth rates of wild-type BW25113, YfgB� and XylA� strainswere determined during exponential growth phase in triplicatecultures at 37°C in rich LB liquid media. Overnight cultures werediluted to an optical density of A550 = 0.01 and growth wasmonitored at A550 every 20 min.
A co-growth competition experiment was performed in LBmedia at 37°C as described (Gutgsell et al. 2000). Initially, 10 mLeach from an overnight culture of kanamycin-sensitive BW25113parental strain and kanamycin-resistant YfgB� cells were mixed in10 mL of LB media. At each 6-h interval, the culture was diluted1000-fold. At the same time, serial dilutions of the cultures wereplated on LB agar and LB agar supplemented with kanamycin(30 mg/mL). The plates were incubated at 37°C and the colonieswere counted.
A co-growth competition assay with XylA knockout strain wasperformed in a similar way except that cells were plated onMacConkey agar plates supplemented with 1% (w/v) D-xylose.On these plates, XylA� cells formed white colonies whereas YfgB�
cells formed red colonies.Antibiotic sensitivity testing was performed using CLSI-
approved protocol (National Committee for Clinical LaboratoryStandards 2003).
ACKNOWLEDGMENTS
We wish to thank Stephen Douthwaite and Birte Vester, Univer-sity of Southern Denmark, for sharing information and advice onexperiments. This work was supported by NIH grant RO1AI072445 to A.S.M.
Received September 6, 2007; accepted October 9, 2007.
REFERENCES
Andersen, N.M. and Douthwaite, S. 2006. YebU is a m5C methyl-transferase specific for 16S rRNA nucleotide 1407. J. Mol. Biol.359: 777–786.
Andersen, T.E., Porse, B.T., and Kirpekar, F. 2004. A novel partialmodification at C2501 in Escherichia coli 23S ribosomal RNA.RNA 10: 907–913.
Baba, T., Ara, T., Hasegawa, M., Takai, Y., Okumura, Y., Baba, M.,Datsenko, K.A., Tomita, M., Wanner, B.L., and Mori, H. 2006.Construction of Escherichia coli K-12 in-frame, single-gene knock-out mutants: the Keio collection. Mol. Syst. Biol. 2: 2006.0008. doi:10.1038/msb4100050.
Bae, T., Banger, A.K., Wallace, A., Glass, E.M., Aslund, F.,Schneewind, O., and Missiakas, D.M. 2004. Staphylococcus aureusvirulence genes identified by bursa aurealis mutagenesis andnematode killing. Proc. Natl. Acad. Sci. 101: 12312–12317.
Ban, N., Freeborn, B., Nissen, P., Penczek, P., Grassucci, R.A.,Sweet, R., Frank, J., Moore, P.B., and Steitz, T.A. 1998. A 9 Aresolution x-ray crystallographic map of the large ribosomalsubunit. Cell 93: 1105–1115.
Ban, N., Nissen, P., Hansen, J., Moore, P.B., and Steitz, T.A. 2000. Thecomplete atomic structure of the large ribosomal subunit at 2.4 Aresolution. Science 289: 905–920.
Bochner, B.R. and Ames, B.N. 1982. Complete analysis of cellularnucleotides by two-dimensional thin layer chromatography. J.Biol. Chem. 257: 9759–9769.
Bugl, H., Fauman, E.B., Staker, B.L., Zheng, F., Kushner, S.R.,Saper, M.A., Bardwell, J.C.A., and Jakob, U. 2000. RNA methyl-ation under heat shock control. Mol. Cell 6: 349–360.
Caldas, T., Binet, E., Bouloc, P., Costa, A., Desgres, J., andRicharme, G. 2000. The FtsJ/RrmJ heat shock protein of Escher-ichia coli is a 23S ribosomal RNA methyltransferase. J. Biol. Chem.275: 16414–16419.
Cannone, J.J., Subramanian, S., Schnare, M.N., Collett, J.R.,D’Souza, L.M., Du, Y., Feng, B., Lin, N., Madabusi, L.V.,Muller, K.M., et al. 2002. The Comparative RNA Web (CRW)Site: An online database of comparative sequence and structureinformation for ribosomal, intron, and other RNAs. BMC Bio-informatics 3: 2.
Conrad, J., Sun, D., Englund, N., and Ofengand, J. 1998. The rluCgene of Escherichia coli codes for a pseudouridine synthase thatis solely responsible for synthesis of pseudouridine at positions955, 2504, and 2580 in 23S ribosomal RNA. J. Biol. Chem. 273:18562–18566.
Corpet, F. 1988. Multiple sequence alignment with hierarchicalclustering. Nucleic Acids Res. 16: 10881–10890. doi: 10.1093/nar/16.22.10881.
Del Campo, M., Kaya, Y., and Ofengand, J. 2001. Identification andsite of action of the remaining four putative pseudouridinesynthases in Escherichia coli. RNA 7: 1603–1615.
Green, R. and Noller, H.F. 1999. Reconstitution of functional 50Sribosomes from in vitro transcripts of Bacillus stearothermophilus23S rRNA. Biochemistry 38: 1772–1779.
Gutgsell, N., Englund, N., Niu, L., Kaya, Y., Lane, B.G., andOfengand, J. 2000. Deletion of the Escherichia coli pseudouridinesynthase gene truB blocks formation of pseudouridine 55 in tRNAin vivo, does not affect exponential growth, but confers a strongselective disadvantage in competition with wild-type cells. RNA 6:1870–1881.
Hansen, J.L., Ippolito, J.A., Ban, N., Nissen, P., Moore, P.B., andSteitz, T.A. 2002. The structures of four macrolide antibioticsbound to the large ribosomal subunit. Mol. Cell 10: 117–128.
Harms, J., Schluenzen, F., Zarivach, R., Bashan, A., Gat, S., Agmon, I.,Bartels, H., Franceschi, F., and Yonath, A. 2001. High-resolutionstructure of the large ribosomal subunit from a mesophiliceubacterium. Cell 107: 679–688.
Helser, T.L., Davies, J.E., and Dahlberg, J.E. 1972. Mechanismof kasugamycin resistance in Escherichia coli. Nat. New Biol. 235:6–9.
Hernandez, H., Pierrel, F., Elleingand, E., Garcia-Serres, R.,Huynh, B.H., Johnson, M.K., Fontecave, M., and Atta, M. 2007.MiaB, a bifunctional radical-S-adenosylmethionine enzyme involvedin the thiolation and methylation of tRNA, contains two essential[4Fe–4S] clusters. Biochemistry 46: 5140–5147.
Huang, L., Ku, J., Pookanjanatavip, M., Gu, X., Wang, D.,Greene, P.J., and Santi, D.V. 1998. Identification of two Escherichiacoli pseudouridine synthases that show multisite specificity for 23SRNA. Biochemistry 37: 15951–15957.
Johansen, S.K., Maus, C.E., Plikaytis, B.B., and Douthwaite, S. 2006.Capreomycin binds across the ribosomal subunit interface usingtlyA-encoded 29-O-methylations in 16S and 23S rRNAs. Mol. Cell23: 173–182.
Kehrenberg, C., Schwarz, S., Jacobsen, L., Hansen, L.H., and Vester, B.2005. A new mechanism for chloramphenicol, florfenicol, andclindamycin resistance: Methylation of 23S ribosomal RNA atA2503. Mol. Microbiol. 57: 1064–1073.
Keith, G. 1995. Mobilities of modified ribonucleotides on two-dimensional cellulose thin-layer chromatography. Biochimie 77:142–144.
RlmN methylates A2503 in 23S rRNA
www.rnajournal.org 105
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from

Khaitovich, P., Tenson, T., Kloss, P., and Mankin, A.S. 1999.Reconstitution of functionally active Thermus aquaticus largeribosomal subunits with in vitro-transcribed rRNA. Biochemistry38: 1780–1788.
Kitagawa, M., Ara, T., Arifuzzaman, M., Ioka-Nakamichi, T.,Inamoto, E., Toyonaga, H., and Mori, H. 2005. Complete set ofORF clones of Escherichia coli ASKA library (a complete set of E.coli K-12 ORF archive): Unique resources for biological research.DNA Res. 12: 291–299.
Kowalak, J.A., Bruenger, E., and McCloskey, J.A. 1995. Posttranscrip-tional modification of the central loop of domain V in Escherichiacoli 23S ribosomal RNA. J. Biol. Chem. 270: 17758–17764.
Kozbial, P. and Mushegian, A. 2005. Natural history of S-adenosyl-methionine-binding proteins. BMC Struct. Biol. 5: 19.
Krzyzosiak, W., Denman, R., Nurse, K., Hellmann, W., Boublik, M.,Gehrke, C.W., Agris, P.F., and Ofengand, J. 1987. In vitro synthesisof 16S ribosomal RNA containing single base changes and assemblyinto a functional 30S ribosome. Biochemistry 26: 2353–2364.
Leach, K.L., Swaney, S.M., Colca, J.R., McDonald, W.G., Blinn, J.R.,Thomasco, L.M., Gadwood, R.C., Shinabarger, D., Xiong, L., andMankin, A.S. 2007. The site of action of oxazolidinone antibiotics inliving bacteria and in human mitochondria. Mol. Cell 26: 393–402.
Lesnyak, D.V., Sergiev, P.V., Bogdanov, A.A., and Dontsova, O.A.2006. Identification of Escherichia coli m2G methyltransferases: I.The ycbY gene encodes a methyltransferase specific for G2445 ofthe 23S rRNA. J. Mol. Biol. 364: 20–25.
Long, K.S., Poehlsgaard, J., Kehrenberg, C., Schwarz, S., and Vester, B.2006. The Cfr rRNA methyltransferase confers resistance to pheni-cols, lincosamides, oxazolidinones, pleuromutilins, and streptogra-min A antibiotics. Antimicrob. Agents Chemother. 50: 2500–2505.
Lovgren, J.M. and Wikstrom, P.M. 2001. The rlmB gene is essentialfor the formation of Gm2251 in 23S rRNA but not for ribosomematuration in Escherichia coli. J. Bacteriol. 183: 6957–6960.
Mankin, A.S. 2006. Nascent peptide in the ‘‘birth canal’’ of theribosome. Trends Biochem. Sci. 31: 11–13.
Mann, P.A., Xiong, L., Mankin, A.S., Chau, A.S., Mendrick, C.A.,Najarian, D.J., Cramer, C.A., Loebenberg, D., Coates, E.,Murgolo, N.J., et al. 2001. EmtA, a rRNA methyltransferase confer-ring high-level evernimicin resistance. Mol. Microbiol. 41: 1349–1356.
Mengel-Jorgensen, J., Jensen, S.S., Rasmussen, A., Poehlsgaard, J.,Iversen, J.J.L., and Kirpekar, F. 2006. Modifications in Thermusthermophilus 23S ribosomal RNA are centered in regions of RNA–RNA contact. J. Biol. Chem. 281: 22108–22117.
Merryman, C. and Noller, H.F. 1998. Footprinting and modification-interference analysis of binding sites on RNA. In RNA: Proteininteractions, a practical approach (ed. C.W.J. Smith), pp. 237–253.Oxford University Press, Oxford.
Nakatogawa, H. and Ito, K. 2002. The ribosomal exit tunnel functionsas a discriminating gate. Cell 108: 629–636.
National Committee for Clinical Laboratory Standards. 2003. Methodsfor dilution antimicrobial susceptibility tests for bacteria that growaerobically; approved standard M7-A6. Clinical Laboratory Stand-ards Institute, Wayne, PA.
Noller, H.F. 1984. Structure of ribosomal RNA. Annu. Rev. Biochem.53: 119–162.
Novick, R.P., Ross, H.F., Projan, S.J., Kornblum, J., Kreiswirth, B., andMoghazeh, S. 1993. Synthesis of staphylococcal virulence factorsis controlled by a regulatory RNA molecule. EMBO J. 12: 3967–3975.
Ofengand, J. and Del Campo, M. 2004. Modified nucleosides inEscherichia coli ribosomal RNA. In EcoSal—Escherichia coli andSalmonella: Cellular and molecular biology (ed. R. Curtiss), Chap.4.6.1. ASM Press, Washington, DC.
Ofengand, J. and Fournier, M.J. 1998. The pseudouridine residues ofrRNA: Number, location, biosynthesis, and function. In Modifi-cation and editing of RNA (eds. H. Grosjean and R. Benne), pp.229–253. ASM Press, Washington, DC.
Okamoto, S., Tamaru, A., Nakajima, C., Nishimura, K., Tanaka, Y.,Tokuyama, S., Suzuki, Y., and Ochi, K. 2007. Loss of a conserved
7-methylguanosine modification in 16S rRNA confers low-levelstreptomycin resistance in bacteria. Mol. Microbiol. 63: 1096–1106.
Pierrel, F., Douki, T., Fontecave, M., and Atta, M. 2004. MiaB proteinis a bifunctional radical-S-adenosylmethionine enzyme involvedin thiolation and methylation of tRNA. J. Biol. Chem. 279: 47555–47563.
Porse, B.T. and Garrett, R.A. 1999. Sites of interaction of strepto-gramin A and B antibiotics in the peptidyl transferase loop of 23SrRNA and the synergism of their inhibitory mechanisms. J. Mol.Biol. 286: 375–387.
Saenger, W. 1984. Principles of nucleic acid structure. Springer-Verlag,New York.
Sambrook, J., Fritsch, E.F., and Maniatis, T. 1989. Molecular cloning; alaboratory manual. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
Schlunzen, F., Zarivach, R., Harms, J., Bashan, A., Tocilj, A.,Albrecht, R., Yonath, A., and Franceschi, F. 2001. Structural basisfor the interaction of antibiotics with the peptidyl transferasecentre in eubacteria. Nature 413: 814–821.
Schuwirth, B.S., Borovinskaya, M.A., Hau, C.W., Zhang, W., Vila-Sanjurjo, A., Holton, J.M., and Cate, J.H.D. 2005. Structures of thebacterial ribosome at 3.5 A resolution. Science 310: 827–834.
Schwarz, S., Werckenthin, C., and Kehrenberg, C. 2000. Identificationof a plasmid-borne chloramphenicol-florfenicol resistance gene inStaphylococcus sciuri. Antimicrob. Agents Chemother. 44: 2530–2533.
Schwarz, S., Kehrenberg, C., Doublet, B., and Cloeckaert, A. 2004.Molecular basis of bacterial resistance to chloramphenicol andflorfenicol. FEMS Microbiol. Rev. 28: 519–542.
Selmer, M., Dunham, C.M., Murphy, F.V.I.V., Weixlbaumer, A.,Petry, S., Kelley, A.C., Weir, J.R., and Ramakrishnan, V. 2006.Structure of the 70S ribosome complexed with mRNA and tRNA.Science 313: 1935–1942.
Sofia, H.J., Chen, G., Hetzler, B.G., Reyes-Spindola, J.F., andMiller, N.E. 2001. Radical SAM, a novel protein superfamilylinking unresolved steps in familiar biosynthetic pathways withradical mechanisms: Functional characterization using new anal-ysis and information visualization methods. Nucleic Acids Res. 29:1097–1106. doi: 10.1093/nar/29.5.1097.
Toh, S.M., Xiong, L., Arias, C.A., Villegas, M.V., Lolans, K., Quinn, J.,and Mankin, A.S. 2007. Acquisition of a natural resistance generenders a clinical strain of methicillin-resistant Staphylococcusaureus resistant to the synthetic antibiotic linezolid. Mol. Micro-biol. 64: 1506–1514.
Treede, I., Jakobsen, L., Kirpekar, F., Vester, B., Weitnauer, G.,Bechthold, A., and Douthwaite, S. 2003. The avilamycin resistancedeterminants AviRa and AviRb methylate 23S rRNA at theguanosine 2535 base and the uridine 2479 ribose. Mol. Microbiol.49: 309–318.
Tu, D., Blaha, G., Moore, P.B., and Steitz, T.A. 2005. Structures ofMLSBK antibiotics bound to mutated large ribosomal subunitsprovide a structural explanation for resistance. Cell 121: 257–270.
Van Buul, C.P., Damm, J.B., and Van Knippenberg, P.H. 1983.Kasugamycin resistant mutants of Bacillus stearothermophiluslacking the enzyme for the methylation of two adjacent adenosinesin 16S ribosomal RNA. Mol. Gen. Genet. 189: 475–478.
Vester, B. and Garrett, R.A. 1988. The importance of highly conservednucleotides in the binding region of chloramphenicol at thepeptidyl transfer center of Escherichia coli 23S ribosomal RNA.EMBO J. 7: 3577–3587.
Wang, S.C. and Frey, P.A. 2007. S-adenosylmethionine as an oxidant:The radical SAM superfamily. Trends Biochem. Sci. 32: 101–110.
Weisblum, B. 1995. Erythromycin resistance by ribosome modifica-tion. Antimicrob. Agents Chemother. 39: 577–585.
Xiong, L., Kloss, P., Douthwaite, S., Andersen, N.M., Swaney, S.,Shinabarger, D.L., and Mankin, A.S. 2000. Oxazolidinone resis-tance mutations in 23S rRNA of Escherichia coli reveal the centralregion of domain V as the primary site of drug action. J. Bacteriol.182: 5325–5331.
Toh et al.
106 RNA, Vol. 14, No. 1
Cold Spring Harbor Laboratory Press on January 3, 2008 - Published by www.rnajournal.orgDownloaded from




















