The effects of dietary sodium loading on the activity and expression of Na, K-ATPase in the rectal...
16
Comparative Biochemistry and Physiology Part B 151 – 152 (2002) 185–200 1096-4959/02/$ - see front matter 2002 Elsevier Science Inc. All rights reserved. PII: S1096-4959 Ž 01 . 00493-6 The effects of dietary sodium loading on the activity and expression of Na, K-ATPase in the rectal gland of the European Dogfish (Scyliorhinus canicula) S. MacKenzie , C.P. Cutler , N. Hazon , G. Cramb * a b c b, Unitat de Fisiologia Animal, Dept. de Biol. Cellular, a Fisiologia i d’lmmunologia Facultat de Ciencies. Edifici C. Universitat Autonoma de Barcelona 08913 Bellaterra (Barcelona) ` School of Biology, University of St. Andrews, Bute Medical Buildings, St. Andrews, Fife KY16 9TS, UK b School of Biology, University of St. Andrews, Gatty Marine Laboratory, St. Andrews, Fife KY16 8LB, UK c Received 21 June 2001; received in revised form 25 September 2001; accepted 8 October 2001 Abstract cDNA fragments of both the a- and b-subunits of the Na, K-ATPase and a cDNA fragment of the secretory form of Na – K – Cl cotransporter from the European dogfish (Scyliorhinus canicula) were amplified and cloned using degenerate primers in RT-PCR. These clones were used along with a sCFTR cDNA from the related dogfish shark, Squalus acanthias to characterise the expression of mRNAs for these ion transporters in the dogfish rectal gland subsequent to an acute feeding episode. Following a single feeding event where starved dogfish were fed squid portions (20 g squid y kg fish), there was a delayed and transient 40-fold increase in the activity of Na, K-ATPase in crude rectal gland homogenates. Increases in enzyme activity were apparent 3 h after the feeding event and peaked at 9 h before returning to control values within 24 h. These increases in activity were accompanied by small and transient decreases in plasma sodium and chloride concentrations lasting up to 3 days. Significant increases in the expression of mRNAs for a- and b-subunits of the Na, K-ATPase, the Na–K–Cl cotransporter and CFTR chloride channel were detected but not until 1 –2 days after the feeding event. It is concluded that the transient increase in Na, K-ATPase activity is not attributable to increases in the abundance of a- and b-subunit mRNAs but must be associated with some, as yet unknown, post- transcriptional activation mechanism. 2002 Elsevier Science Inc. All rights reserved. Keywords: European dogfish; Scyliorhinus canicula; Dietary salt; Na; K-ATPase; CFTR; Na,K,Cl-cotransporter; Expression 1. Introduction The rectal gland is fundamental to the control of salt and water balance in marine elasmobranch fish. This small digitform organ intermittently secretes a fluid which is iso-osmotic with the plasma but composed almost entirely of NaCl (Burger, 1962). Rectal gland epithelial cells trans- port chloride ions from the blood to the lumen of * Corresponding author. Tel.: q44-1334-463530; fax: q44- 1334-463600. E-mail address: [email protected] (G. Cramb). the secretory tubules by a process known as secondary active transport. This process involves the concerted activity of a number of epithelial ion transporters including a basolaterally located, bumetanide-sensitive, secretory type Na–K–Cl cotransporter (NKCC1; Lytle and Forbrush, 1992a,b; Lytle et al., 1992; Xu et al., 1994) and an apically located chloride channel (Gogelein et al., 1987) which is the shark homologue of the mammalian cystic fibrosis transmembrane conduc- tance regulator (sCFTR; Marshall et al., 1991). The extrusion of chloride ions creates a lumen
-
Upload
st-andrews -
Category
Documents
-
view
1 -
download
0
Transcript of The effects of dietary sodium loading on the activity and expression of Na, K-ATPase in the rectal...

Comparative Biochemistry and Physiology Part B 151–152(2002) 185–200
1096-4959/02/$ - see front matter� 2002 Elsevier Science Inc. All rights reserved.PII: S1096-4959Ž01.00493-6
The effects of dietary sodium loading on the activity and expressionof Na, K-ATPase in the rectal gland of the European Dogfish
(Scyliorhinus canicula)
S. MacKenzie , C.P. Cutler , N. Hazon , G. Cramb *a b c b,
Unitat de Fisiologia Animal, Dept. de Biol. Cellular,a
Fisiologia i d’lmmunologia Facultat de Ciencies. Edifici C. Universitat Autonoma de Barcelona 08913 Bellaterra (Barcelona)`School of Biology, University of St. Andrews, Bute Medical Buildings, St. Andrews, Fife KY16 9TS, UKb
School of Biology, University of St. Andrews, Gatty Marine Laboratory, St. Andrews, Fife KY16 8LB, UKc
Received 21 June 2001; received in revised form 25 September 2001; accepted 8 October 2001
Abstract
cDNA fragments of both thea- andb-subunits of the Na, K-ATPase and a cDNA fragment of the secretory form ofNa–K–Cl cotransporter from the European dogfish(Scyliorhinus canicula) were amplified and cloned using degenerateprimers in RT-PCR. These clones were used along with a sCFTR cDNA from the related dogfish shark,Squalusacanthias to characterise the expression of mRNAs for these ion transporters in the dogfish rectal gland subsequent toan acute feeding episode. Following a single feeding event where starved dogfish were fed squid portions(20 g squidykg fish), there was a delayed and transient 40-fold increase in the activity of Na, K-ATPase in crude rectal glandhomogenates. Increases in enzyme activity were apparent 3 h after the feeding event and peaked at 9 h before returningto control values within 24 h. These increases in activity were accompanied by small and transient decreases in plasmasodium and chloride concentrations lasting up to 3 days. Significant increases in the expression of mRNAs fora- andb-subunits of the Na, K-ATPase, the Na–K–Cl cotransporter and CFTR chloride channel were detected but not until 1–2 days after the feeding event. It is concluded that the transient increase in Na, K-ATPase activity is not attributableto increases in the abundance ofa- andb-subunit mRNAs but must be associated with some, as yet unknown, post-transcriptional activation mechanism.� 2002 Elsevier Science Inc. All rights reserved.
Keywords: European dogfish; Scyliorhinus canicula; Dietary salt; Na; K-ATPase; CFTR; Na,K,Cl-cotransporter; Expression
1. Introduction
The rectal gland is fundamental to the controlof salt and water balance in marine elasmobranchfish. This small digitform organ intermittentlysecretes a fluid which is iso-osmotic with theplasma but composed almost entirely of NaCl(Burger, 1962). Rectal gland epithelial cells trans-port chloride ions from the blood to the lumen of
* Corresponding author. Tel.:q44-1334-463530; fax:q44-1334-463600.
E-mail address: [email protected](G. Cramb).
the secretory tubules by a process known assecondary active transport. This process involvesthe concerted activity of a number of epithelialion transporters including a basolaterally located,bumetanide-sensitive, secretory type Na–K–Clcotransporter (NKCC1; Lytle and Forbrush,1992a,b; Lytle et al., 1992; Xu et al., 1994) andan apically located chloride channel(Gogelein etal., 1987) which is the shark homologue of themammalian cystic fibrosis transmembrane conduc-tance regulator(sCFTR; Marshall et al., 1991).The extrusion of chloride ions creates a lumen

186 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
negative transepithelial potential which is generallyconsidered to drive the secretion of sodium via theparacellular pathway(Silva et al., 1977). Theimmediate energy supply for the overall process isderived from the maintenance of the sodium gra-dient across the basolateral membrane provided bythe Na, K-ATPase(Silva et al., 1977; Eveloff etal., 1979) which allows the cellular accumulationof chloride ions via the Na–K–Cl cotransporter.Excess accumulation of potassium is prevented bythe presence of a basolaterally located potassiumchannel which mediates the passive efflux ofpotassium and the generation of the restingmembrane potential of the cell(Greger et al.,1987).Chloride secretion by the rectal gland is consid-
ered to be primarily under hormonal control withincreases in secretion the direct result of activationof the apically located CFTR chloride channel.Both cAMPyPKA (Shuttleworth and Thompson,1980; Forrest et al., 1983; Greger et al., 1984a,b;Valentich and Forrest, 1991; Lear et al., 1992;Hanrahan et al., 1993) and cGMPyPKG (Kennedyet al., 1991; Karnaky et al., 1991, 1992) secondmessenger pathways are implicated in the regula-tion of chloride secretion where increases in apicalchloride conductance are correlated with increasesin the phosphorylation state of the CFTR protein.The perfused rectal gland secretion rate has beenreported to increase by as much as 40-fold afterthe inclusion of cyclic AMP in the perfusate(Stoffet al., 1979). CFTR has been shown to functionas a cyclic AMPyPKA-activated chloride channelin both mammals(Cheng et al., 1991; Dulhantyand Riordan, 1994) and in the dogfish shark(Squalus acanthias) rectal gland(Devor et al.,1995). In addition to direct channely transporterregulation at the protein level by the myriad ofpossible kinase cascades both transcriptional andtranslational mechanisms are also possiblyinvolved in both acute and long-term regulation ofion secretion.The secretory activity of the gland is thought to
be related to the extent of whole body NaClloading associated with feeding(Shuttleworth,1988). Despite this, very little is known about thehormonal andyor ionic mechanisms which contrib-ute to the regulation of rectal gland secretionduring, and immediately after a feeding episode.A number of hormones have been reported to alterthe activity andyor expression of various compo-
nents of the secretory pathway but their involve-ment, if any, with the feeding-induced stimulationof secretion remains to be established. Likewisethe effect of feeding on the subsequent expressionand activity of these ion transporters is alsounknown. In this initial study we have examinedthe effect of a single feeding event on the activityof Na, K-ATPase in rectal gland homogenates andon the expression of mRNAs for botha1 andb1subunits of the Na, K-ATPase, the secretory iso-form of the Na–K–Clcotransporter and the CFTRchloride channel. Degenerate oligonucleotide prim-ers were used in the polymerase chain reaction(PCR) to isolate, clone and sequence partial c-DNAs for the a1 and b1 subunits of Na, K-ATPase and the Na–K–Cl cotransporter from therectal gland of the European dogfish(Scyliorhinuscanicula). These cDNAs, along with a heterolo-gous 1.8 kb cDNA encoding a region of the shark(Squalus acanthias) CFTR sequence, were usedas probes to assess the effects if feeding had anyeffect on the expression of mRNAs for these iontransporters in the rectal gland.
2. Materials and methods
2.1. Animals
European lesser spotted dogfish,Scyliorhinuscanicula, of mixed age, sex and body weight(0.5–1.2 kg) were obtained by both long line andtrawling from coastal waters around Millport onthe Firth of Clyde in Scotland and from coastalwaters around Bangor on the north west coast ofWales. The fish were not fed and held in 150 lconstant flow-through circular tanks under a 12 hlight:12 h dark photoperiod for 3–4 weeks beforebeing used in experiments.
2.2. Tissue sampling
At timed intervals after a single feeding eventconsisting of quartered squid(20 g squidykg fish),fish were killed by a sharp blow to the headfollowed immediately by transection of the spinalcord directly behind the cranium and destructionof the central nervous system by pithing. Bloodwas sampled via direct cardiac puncture using an18-gauge syringe needle and collection into a 10ml syringe. Blood samples(5–10 ml) were mixedwith 20 ml of an anti-coagulanty inhibitor cocktail

187S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
comprising 0.5 M 1,10-phenanthroline, 0.225 MEDTA and 2500 KIUyml aprotinin and centrifuged(14 926=g) for 30 s to obtain plasma. Rectalglands (0.1–0.3 g) were removed directly aftersacrifice, washed in ice-cold phosphate bufferedshark Ringer solution comprising 240 mM NaCl,7 mM KCl, 10 mM CaCl , 4.9 mM MgCl , 9.62 2
mM NaH PO , 1.5 mM KH PO , 360 mM urea,2 4 2 4
60 mM trimethylamine oxide(pH 7.3) and hom-ogenised in the appropriate buffer using a PolytronPT-10 tissue homogeniser(Kinematica, Gmbh,Switzerland). Homogenisation solutions used weredependent on the analysis. For total RNA extrac-tion, glands were homogenized at room tempera-ture for 30 s at setting 8 in 10 vols.(wyv) of achaotropic buffer consisting of 4 M guanidiniumiso-thiocyanate, 10%(vyv) b-mercaptoethanol, 1mM EDTA, 10 mM Tris–HCl(pH 7.5). Ouabainsensitive Na, K-ATPase activity was determinedfollowing homogenisation of glands for 3=10 sbursts in 10 vol.(wyv) of an ice-cold buffercomprising 50 mM HEPES, 1 mM EDTA and0.18 mgyml phenyl methyl sulfonyl fluoride(PMSF), 0.01% sodium deoxycholate(DOC) (pH7.4). Homogenates were either used directly orfrozen aty90 8C before further processing. Freez-ing the homogenates did not significantly altersubsequent Na, K-ATPase enzyme activities orRNA yields or viabilities.
2.3. Plasma analysis
The plasma sodium ion concentrations weredetermined using flame photometry(CorningModel 450) and chloride ion concentrations weredetermined using a Chloride Analyser(Corning,Essex, UK). Plasma osmolalities were assessedusing a vapour pressure osmometer(Wescor Inc.,Logan, UT). Plasma sodium and chloride concen-trations are expressed as mmoly l and plasmaosmolality as mOsmykg. Plasma urea was deter-mined using a diagnostic kit(Cat. No. 640-A)obtained from Sigma Aldrich Ltd, Dorset, UKutilising urease as the indicator for ureaconcentration.
2.4. Na, K-ATPase assay
Samples of rectal gland homogenates(0.1 mgprotein) were incubated at room temperature(18–20 8C) for 1 h in an assay buffer(total volume
0.5 ml) comprising 120 mM NaCl, 20 mM KCl,30 mM L-histidine, 4 mM MgCl , 3 mM ATPy2
Tris (pH 7.4). The reactions were terminated bythe addition of an equal volume of ice-cold 10%(wyv) trichloroacetic acid(TCA), left on ice for15 min and then centrifuged at 2000=g for 10min to remove the precipitate. The phosphateconcentration in the supernatant fraction was deter-mined by the method of Fiske and Subbarrow(1925). Assays were carried out in the presenceand absence of ouabain(2 mM) to determine theactivity attributable to the Na, K-ATPase. Proteinconcentrations were determined by the method ofBradford(1976).
2.5. RNA extraction
Total RNA was extracted from rectal glands andmessenger RNA purified using oligo(dT) (Phar-macia) as described previously(Cutler et al.,1995).
2.6. Reverse transcriptase (RT)-PCR amplification
Single-strand cDNA was synthesised from 5mgof dogfish rectal gland mRNA using SuperscriptII reverse transcriptase as instructed by the manu-facturer(GibcoBRL Life Technologies Ltd., Pais-ley, UK) and PCR amplifications were performedusing a standard buffer for AmpliTaq DNA poly-merase(PE Applied Biosystems, Cheshire, UK),containing 200mM dNTPs and 5mM degenerateoligonucleotide primers as described previously(Cutler et al., 1995, 1997). PCR amplificationsused degenerate oligonucleotide primer pairswhich were designed to sequences exhibiting highamino acid conservation between all known spe-cies and incorporating nucleotide wobbles andyorinosine(I) bases at positions of nucleotide uncer-tainty (Cutler et al., 1995). Na, K-ATPaseb1-subunit isoform primers were as previouslydescribed(primer pair 2: Cutler et al., 1997). Asthea3 isoform was reported to be expressed bothin teleost fish(Pressley, 1992) and in the dogfishrectal gland(Benz et al., 1992) degeneratea3-specific sense and P-type ATPase antisense primerswere designed and used in PCR. Although ana3-specific fragment was expected to amplify, mis-priming of the degenerate sense oligonucleotideresulted in the amplification of a homologue of

188 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Table 1Degenerate primers for RT-PCR
Protein Primer sequence Amino acidlocation
Na, K-ATPase alphaSense GAAyGACIyCGAAyGGATyCAyCCIyCAAT yCGA 484–493
TyCAAT yCAyCGIyCTAAnti-sense gycaycaygtcaygttiycaygtyciycccaygtc 705–712
i yccciycgt
Na, K-ATPase betaSense AyCAA yGCCIyCTGTyCITI yCAyTyCTIyCAyGTIyCAA 173–182
A yGAyTyCTIyCAAT yCAyCGAnti-sense ACIyCAyGIyCIyCAyTA yTyGIyCTyCTyGIyCCCAyGTA 241–250
A yGTAI yCGGAyGAyTAA yGTA
NayKyCl co-transporterSense GA(A yG)GA(TyC)CA(TyC)GT(I yC)AA(A yG)AA(TyC)T 754–765
(A yT)(CyT)(A yC)G(I yC)CC(I yC)CA(A yG)TG(TyC)(CyT)TAnti-sense ATyCAyGTCIATI yCGTIyCTyCTyCTyCTTIyCCCTyCTG 994–1003
TyCTTTyCTTTyCTGAyGAA
The table indicates the degenerate primer sets used to amplify dogfish Na, K-ATPasea andb subunit and Na, K, Cl-cotransporter(NKCC1) cDNAs. Amino acid locations are derived from the Human Na, K-ATPasea3 (Accession Number – NCBI; NM000703),eel Na, K-ATPaseb1 and the shark Na, K, Cl-cotransporter. IsInosine.
the a1 isoform. The primers used in the RT-PCRexperiments are indicated in Table 1.
2.7. Cloning and sequencing
PCR amplified fragments were electrophoresedon standard TBE-agarose gels(Sambrook et al.,1989). Bands of the appropriate size were excisedfrom the gel and the DNA was recovered byelectroelution(UEA Analytical Electroeluter, IBI,New Haven, CT). The Na, K-ATPasea1 isoformPCR product was cloned into the plasmid vectorpCR II using a TA cloning kit(Invitrogen, Leek,The Netherlands). The Na, K-ATPaseb1 isoformand NayK yCl-cotransporter PCR fragments werecloned using a pGEM-T vector systems kit(Pro-mega, Southampton, UK) and sequenced asdescribed in Cutler et al.,(1997). Contiguous PCRfragments generated were aligned to yield cDNAfragment sequences using a GeneJockey computersoftware package(Elsevier Biosoft). Derived ami-no acid sequences were also aligned using thesame software package.
2.8. Northernydot blot analysis
Total RNA was size separated by denaturing gelelectrophoresis(Sambrook et al., 1989). Typically5–20 mg of total RNA was electrophoresed on1.2% denaturing formaldehyde agarose gels at 80
V for 4 h in a buffer comprising 20 mM 3-wN-morpholinox propane sulfonic acid, 5 mM Naacetate, 1 mM EDTA, pH 7.0(1= MOPS). Gelswere electroblotted onto nylon membranes(usinga Trans-blot cell and Zetaprobe GT membranes,both Bio-Rad Laboratories, Watford, Herts., UK)and baked under vacuum for 30 min at 808C.Filters were pre-hybridised at 428C in a solution
comprising 20 mM NaH POy Na HPO , 1 mM2 4 2 4
EDTA, 3% (wyv) SDS, 50% (vyv) deionisedformamide (pH 7.4), containing 5= Denhardt’s(0.1% polyvinylpyrrolidone, 0.1% Ficoll, 0.1%BSA) and 0.5 mgyml boiled, sonicated calf thy-mus DNA and 0.5 mgyml boiled, sonicated yeasttotal RNA. Hybridisation was carried out overnightin the presence of P-labelled transporter-specific32
cDNA probes (Megaprime DNA labelling kit,Amersham Pharmacia Biotech, UK). The probesused were the cloned cDNAs for thea1 (672 bp)and b1 (181 bp) subunits of the Na, K-ATPaseand the Na–K–Clcotransporter(685 bp). A full-length (6.4 kb) sCFTR clone fromSqualus acan-thias was kindly provided by Professor J.R.Riordan and Dr T.D. Singer, Hospital for SickChildren, University of Toronto. Following radiol-abelling the specific activities of all cDNA probeswere approximately 1=10 dpmymg. Blots were9
also probed with an 18S ribosomal RNA probe(59-GCGGTGTGTACAAAGGGCAGGGACTTA-ATC-39) to the 309 helix motif at nts 3370–3400

189S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 1. Interleaved nucleotide and amino acid sequence of the dogfish(Scyliorhinus canicula) Na, K-ATPasea1 cDNA fragment(Accession Number, EMBL; Y18920). The region of the fragment which shares high homology within individual mammalian isoformshas a solid underline. The location of the intended binding site of thea3 specific primer is indicated by a dotted underline. Numberson the left refer to the positions of nucleotides, and on the right to derived amino acids within the sequence.
of the rat 18S rRNA sequence(Dams et al., 1988).This single stranded DNA probe was end labelledwith w Px by incubation of 300 ng of probe for 132
h at 37 8C in a phosphate-exchange cocktailcomprising 50 mM imidazole–HCl, 12 mMMgCl , 1 mM b-mercaptoethanol, 70 nM ADP, 22
mM wg- PxATP(6000 Ciymmol) and 20 units T432
polynucleotide kinase(GibcoBRL Life Technolo-gies, Paisley, UK) in a total volume of 25ml. Theradiolabelled probe was then separated from radiol-abelled nucleotides by exclusion chromatographyusing Sephadex G50(Amersham Pharmacia Bio-tech, UK). After hybridisation, blots were washedin a series of NaClySDS buffers of increasingstringency(Cutler et al., 1995). Autoradiographswere obtained by exposing blots to photographicfilm (Hyperfilm-MP, Amersham Pharmacia Bio-tech, UK) for up to 24 h aty90 8C and theintensity of the photographic images quantified bydensitometry(Shimadzu CS-9000 flying-spot den-sitometer, Shimadzu Corporation, Kyoto, Japan).
For dot blot analysis, total RNA samples(0.5mg) were diluted to 12.5mgyml in 1= MOPScontaining 50% formamide and 17.5% formalde-hyde and denatured by heating to 658C for 15min. Denatured RNA samples were further dilutedby addition of 10 vols. of 10= SSC (8.8% wyvNaCl, 4.4% wyv trisodium citrate, pH 7.0) beforeapplying directly onto Hybond N Nylon filtersq
(Amersham Pharmacia Biotech, UK) under vacu-um (5 inches Hg) using a dot blotting manifold(GibcoBRL Life Technologies, Paisley, UK). Fil-ters were washed twice with equal volumes of 10= SSC before baking and subsequent processingfor hybridisation as described above.
2.9. Statistical analysis
All data obtained were analysed using factorialone-way analysis variance(ANOVA). Post-hoctests used were Bonferroni and Dunn(plasmaelectrolytes, osmolality, relative abundance m-

190 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
RNAs) and Scheffe’sF-test (for Na, K-ATPaseactivities); levels of significance, *P-0.05; **P,0.01; ***P-0.001.
3. Results
3.1. Cloning and sequencing of ion transportersin Scyliorhinus canicula
3.1.1. The Na, K-ATPase a1 isoformSequencing of the products isolated following
the use of the Na, K-ATPase alpha subunit-specificprimer set revealed that a homologue of thea1isoform had in fact been amplified. The sequenceinformation indicated that thea3-specific senseprimer had annealed to the cDNA at a site more59 than the intended hybridisation site in theputative elasmobranch Na, K-ATPasea3 isoform(see Fig. 1). Following sequencing the translatedamino acid sequence of the amplified Na, K-ATPasea cDNA fragment was found to be moresimilar to the known Na, K-ATPasea1 isoformsrather than existinga3 isoforms. Within a con-served region known to be specific to each isoformthe cloned dogfish cDNA shares seven out ofeleven amino acids with the conserveda1sequence compared with only six out of elevenconserved amino acids with the normally highlyconserveda3 sequence(Cutler et al., 1995). Thedogfish cDNA also shares high levels of aminoacid homology with the only other elasmobranchsequence available, that of the ray,Torpedo cali-fornica (87.5%), as well as other Na, K-ATPasea subunit sequences from teleost fish and mam-mals(Fig. 2).
3.1.2. The Na, K-ATPase b1 isoformA dogfish Na, K-ATPaseb1 cDNA was ampli-
fied from the rectal gland RNA using RT-PCR(Fig. 3). This cDNA fragment shares high levelsof amino acid homology with the Na, K-ATPaseb subunit sequence fromTorpedo californica(68.3%) with lower levels of homology to Na, K-ATPaseb1 isoform sequences from teleost fishand mammals(Fig. 4).
3.1.3. The NayKyCl cotransporterA dogfish Na–K–Cl cotransporter cDNA frag-
ment was also obtained using RT-PCR(Fig. 5).This cDNA fragment shares high levels of aminoacid homology with the Na–K–Cl cotransportersequence fromSqualus acanthias (95.2%) as well
with other secretory-type Na–K–Cl (NKCC1)cotransporter isoform sequences previously report-ed from mammals(Fig. 6).
3.2. Plasma ion concentrations and osmolality inthe dogfish after a single feeding event
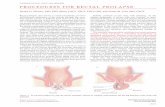
The plasma sodium and chloride concentrationsin starved dogfish were 308"11.7 mmoly l and274"14.78 mmoly l, respectively. Following a sin-gle feeding event(20 g squidykg body wt.)significant differences in both the plasma sodiumand chloride concentrations were found over thefollowing 3 days(Fig. 7). Within 6 h of feedingthe plasma sodium concentrations started todecrease, reaching a minimum of 222.5"6.2mmoly l (some 27% below control levels) 2 daysafter the feeding event before eventually recover-ing to control levels after 5 days. The decrease inplasma sodium was accompanied by a significantfall in plasma chloride concentration 12 h after thefeeding event, reaching a minimum concentrationof 224"14.0 mmoly l (some 18% below controllevels) two days after the initial feeding eventbefore recovering to control values the followingday. No significant changes in plasma urea con-centrations or plasma osmolality(Fig. 7) wereobserved up to 10 days after the single feedingevent.
3.3. Na, K-ATPase activity
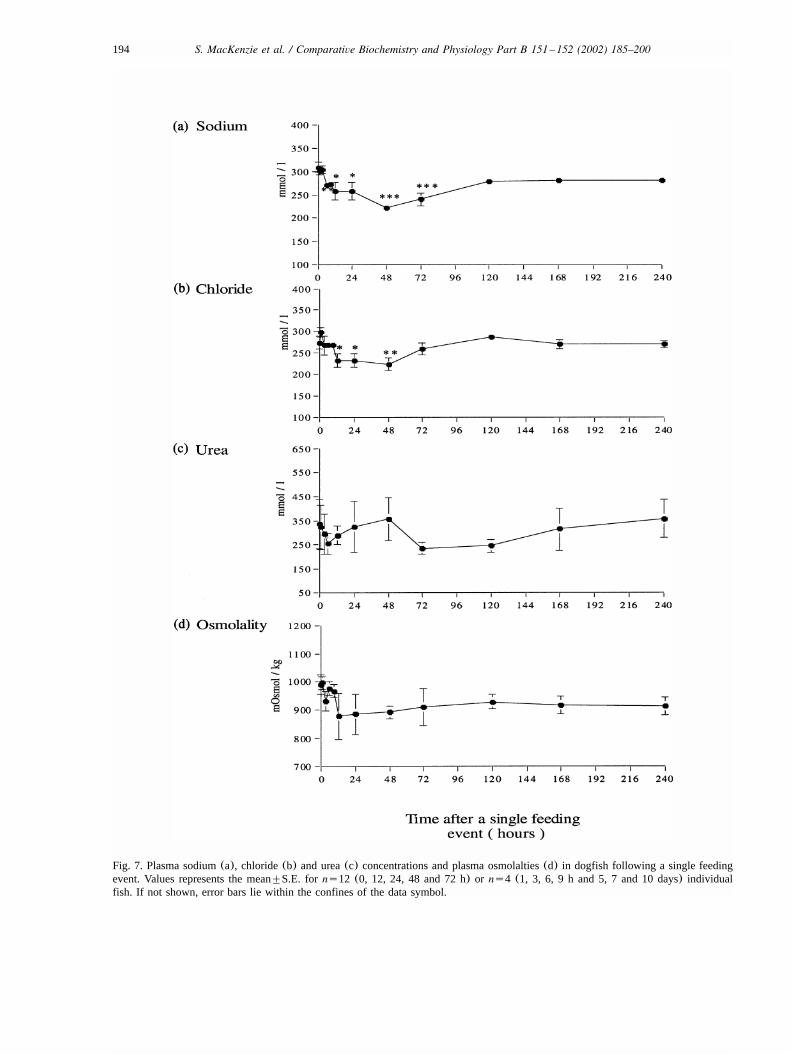
Following the single feeding event, there was adelayed and transient rise in Na, K-ATPase activityin rectal gland homogenates(Fig. 8). AlthoughNa, K-ATPase activity did not change significantlywithin the first 3 h post-feeding, by the 6 h timepoint a mean 25-fold increase in Na, K-ATPaseactivity was recorded. The Na, K-ATPase activitycontinued to increase reaching a maximum 40-foldincrease over control levels some 9 h after thefeeding event before subsequently returning backto basal activities by 24 h.
3.4. Na, K-ATPase a1 and b1 subunit, Na–K–Clcotransporter and sCFTR mRNA abundance
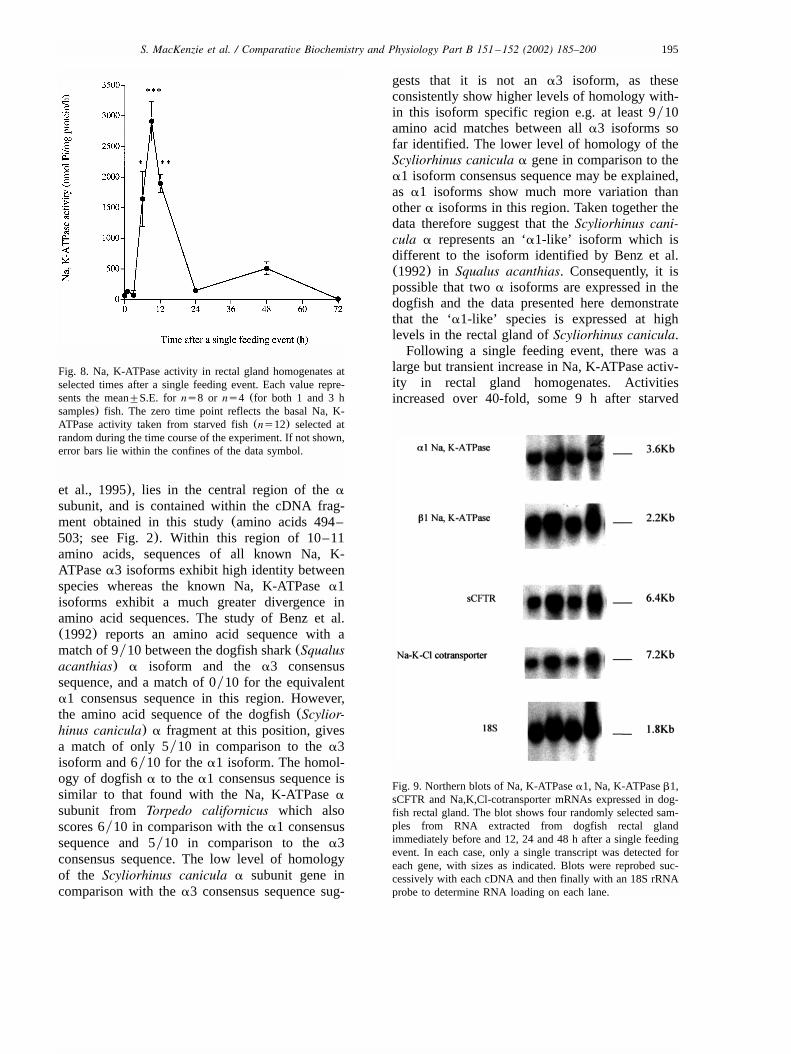
Since Northern blots determined that all probeshybridised to single mRNA species of 3.6 kb(Na,K-ATPasea1 subunit), 2.2 kb(Na, K-ATPaseb1subunit), 6.4 kb (CFTR) and 7.2 kb(Na–K–Clcotransporter) (Fig. 9), subsequent quantification

191S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 2. Alignment of the dogfish Na, K-ATPasea1 isoform derived amino acid sequence with sequences from other species. The dashes(–) represent positions within the sequence that are identical between sequences, and the dots(Ø) indicate gaps which were added toimprove the alignment of sequences. The numbers represent the position of the sequence within that ofTorpedo californica. Sequencesare from EMBLySwiss Prot sequence database, with Accession Numbers as follows:Torpedo, P05025; Eel, Q92030; White Sucker,P25489; Human, P05023.
Fig. 3. Interleaved nucleotide and derived amino acid sequence of a cDNA fragment of the dogfish Na, K-ATPaseb1 cDNA fragment(Accession Number, EMBL; Y18921). Numbers on the left refer to the positions of nucleotides and on the right to derived amino acidswithin the sequence.
of expression was conducted using dot blotting.The relative expression of rectal gland transportermRNAs were followed over a 10 day period(Fig.10) following a single feeding episode with squid
(20 gykg body wt.). Autoradiographic images ofdot blots were quantified by densitometry. Absorb-ance readings related to the hybridisation of spe-cific transporter probes were normalised to the

192 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 4. Alignment of the dogfish Na, K-ATPaseb1 isoform derived amino acid sequence with sequences from other species. The dashes(–) represent positions within the sequence that are identical between species, and the dots(Ø) indicate gaps which were added toimprove the alignment of sequences. The numbers represent the position of the sequence within that ofTorpedo californica. Sequencesare from EMBLySwiss Prot sequence database, with Accession Numbers as follows:Torpedo, P05025; Eel, P51165; Human, P050026.
Fig. 5. Interleaved nucleotide and derived amino acid sequence of a cDNA fragment of the dogfish Na–K–Cl cotransporter cDNA(Accession Number, EMBL; Y18919). Numbers on the left refer to the positions of nucleotides, and on the right, to derived aminoacids within the sequence.
total amount of RNA loaded for each sample onthe blots by re-probing the filters with an 18Sribosomal RNA probe. There was a gradualincrease in the abundance ofa1 subunit mRNA,which was significantly greater than control valuesafter 2 days (q75.6"5.6%). Although levelsappeared to fall at the 3 day point,a1 mRNAabundance increased again toq89"8.7% overthe starved fish control values at the 5 day time
point. This increase ina1 mRNA abundance wassustained for up to 10 days witha1 mRNA levelsremaining 50"15% higher than control values.The relative abundance ofb1 subunit mRNA wasalso determined in the same total RNA samplesand although exhibiting more variability, signifi-cant increases in abundance were found at boththe 2 and 5 day time points(q39.5"5.8% andq42"19% increases above starved fish control

193S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 6. Alignment of the dogfish NKCC1 secretory Na–K–Cl cotransporter isoform derived amino acid sequence with sequences fromother species. The dashes(–) represent positions within the sequence that are identical between sequences, and the dots(Ø) indicategaps which were added to improve the alignment of sequences. The numbers represent the position of the sequence within that of thesharkSqualus acanthias. Sequences are from EMBLySwiss Prot sequence database, with Accession Numbers as follows: Shark, P55013;Mouse, P55012; Human, P55011.
values, respectively) (Fig. 9b). Similar increasesin the expression of the Na–K–Cl cotransporterand sCFTR mRNAs were also found peaking atq55"11% andq65"38%, respectively, some 2–5 days after the single feeding event.
4. Discussion
Complementary DNA fragments for three iontransport genes(a1 andb1 subunits of the Na, K-ATPase and the Na–K–Cl cotransporter) wereisolated from the dogfish(Scyliorhinus canicula)rectal gland using homology cloning and RT-PCR.In addition, a CFTR mRNA has been identified inScyliorhinus canicula using a cDNA probe from aclosely related species of shark,Squalus acanthias.All genes expressed a single transcript on Northernblots with the sizes of the mRNAs identified as3.6 kb (a1), 2.2 kb (b1), 7.2 kb (cotransporter)and 6.4 kb(CFTR). All of these ion transportmRNAs were found in relatively high abundancein the rectal gland and could easily be detected inNorthern blots using 5mg total RNA.Earlier work by Benz et al.(1992) reported the
isolation and cloning of a Na, K-ATPasea subunitfrom the rectal gland of the spiny dogfish sharkSqualus acanthias. From sequence analyses of the
cloned RT-PCR fragments, the 3.8 kba subunitwas found to share the greatest homology with thea3 isoforms reported for mammals. It was there-fore concluded that thisa3-like Na, K-ATPasesubunit was the predominanta isoform expressedin the rectal gland of the dogfish shark. In mam-mals the Na, K-ATPasea3 isoform is develop-mentally regulated and only expressed to anysignificant degree in a limited number of tissueswith the highest levels of expression generallyfound within the brain(Emanuel et al., 1987). Inthis study, Northern blot analyses using the isolatedNa, K-ATPasea subunit cDNA probe isolatedfrom Scyliorhinus canicula identified a single,highly abundant, 3.6 kb mRNA species. Thisresult, together with the sequence informationobtained suggest that the predominanta isoformexpressed in the rectal gland of the lesser spotteddogfish, Scyliorhinus canicula, is more homolo-gous to thea1 subtype. Amino acid sequencedifferences between thea1 and thea3 isoformsare minor and rely upon comparisons of smallregions of amino acids which are specific to, andare conserved within each isoform’s sequencebetween all species. One such area, which haspreviously been used to determine the Na, K-ATPasea isoform subtype(Pressley, 1992; Cutler
194 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 7. Plasma sodium(a), chloride(b) and urea(c) concentrations and plasma osmolalties(d) in dogfish following a single feedingevent. Values represents the mean"S.E. for ns12 (0, 12, 24, 48 and 72 h) or ns4 (1, 3, 6, 9 h and 5, 7 and 10 days) individualfish. If not shown, error bars lie within the confines of the data symbol.
195S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 8. Na, K-ATPase activity in rectal gland homogenates atselected times after a single feeding event. Each value repre-sents the mean"S.E. for ns8 or ns4 (for both 1 and 3 hsamples) fish. The zero time point reflects the basal Na, K-ATPase activity taken from starved fish(ns12) selected atrandom during the time course of the experiment. If not shown,error bars lie within the confines of the data symbol.
Fig. 9. Northern blots of Na, K-ATPasea1, Na, K-ATPaseb1,sCFTR and Na,K,Cl-cotransporter mRNAs expressed in dog-fish rectal gland. The blot shows four randomly selected sam-ples from RNA extracted from dogfish rectal glandimmediately before and 12, 24 and 48 h after a single feedingevent. In each case, only a single transcript was detected foreach gene, with sizes as indicated. Blots were reprobed suc-cessively with each cDNA and then finally with an 18S rRNAprobe to determine RNA loading on each lane.
et al., 1995), lies in the central region of theasubunit, and is contained within the cDNA frag-ment obtained in this study(amino acids 494–503; see Fig. 2). Within this region of 10–11amino acids, sequences of all known Na, K-ATPasea3 isoforms exhibit high identity betweenspecies whereas the known Na, K-ATPasea1isoforms exhibit a much greater divergence inamino acid sequences. The study of Benz et al.(1992) reports an amino acid sequence with amatch of 9y10 between the dogfish shark(Squalusacanthias) a isoform and the a3 consensussequence, and a match of 0y10 for the equivalenta1 consensus sequence in this region. However,the amino acid sequence of the dogfish(Scylior-hinus canicula) a fragment at this position, givesa match of only 5y10 in comparison to thea3isoform and 6y10 for thea1 isoform. The homol-ogy of dogfisha to thea1 consensus sequence issimilar to that found with the Na, K-ATPaseasubunit from Torpedo californicus which alsoscores 6y10 in comparison with thea1 consensussequence and 5y10 in comparison to thea3consensus sequence. The low level of homologyof the Scyliorhinus canicula a subunit gene incomparison with thea3 consensus sequence sug-
gests that it is not ana3 isoform, as theseconsistently show higher levels of homology with-in this isoform specific region e.g. at least 9y10amino acid matches between alla3 isoforms sofar identified. The lower level of homology of theScyliorhinus canicula a gene in comparison to thea1 isoform consensus sequence may be explained,as a1 isoforms show much more variation thanothera isoforms in this region. Taken together thedata therefore suggest that theScyliorhinus cani-cula a represents an ‘a1-like’ isoform which isdifferent to the isoform identified by Benz et al.(1992) in Squalus acanthias. Consequently, it ispossible that twoa isoforms are expressed in thedogfish and the data presented here demonstratethat the ‘a1-like’ species is expressed at highlevels in the rectal gland ofScyliorhinus canicula.Following a single feeding event, there was a
large but transient increase in Na, K-ATPase activ-ity in rectal gland homogenates. Activitiesincreased over 40-fold, some 9 h after starved

196 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Fig. 10. Quantification of Na, K-ATPasea1 (a) andb1 subunit(b), CFTR (c) and Na,K,Cl-cotransporter(d) mRNA expression inextracts of dogfish rectal gland at timed intervals after a single feeding episode. Transcript abundance was determined using dot blotsas described in the Materials and methods section and expressed as the relative signal intensity to 18S rRNA. Values are means"S.E.for quadruplicate samples taken from each of four individual fish(ns4). If not shown, error bars lie within the confines of the datasymbol.
dogfish were fed with a squid diet(20 g squidykg body wt.). The increase in activity was delayed,with the first significant increases in activityappearing after 3 h, peaking between 6 and 12 hand returning to basal levels again some 24 h after
the feeding event. Thereafter there was no signif-icant change in enzyme activity over the following2–9 days(results not shown). Subsequent to thesingle feeding episode there were also transientdecreases in both plasma sodium and chloride

197S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
concentrations which were significant within thefirst 6–12 h and which were sustained for up to 3days before returning to control levels. The initialdecreases in plasma sodium and chloride coincidedwith the observed increases in rectal gland Na, K-ATPase activity suggesting that increased rectalgland activity may be responsible for the reductionin the plasma concentrations of these two ions.Despite the reduction in concentration of theseions, there were no significant changes in eitherplasma osmolalities or plasma urea concentrationsalthough it is possible that decreases in the formerwere masked by the high variabilities foundbetween fish after feeding.Despite the large increase in Na, K-ATPase
activity found over the first 24 h, there was noconcomitant increase in mRNA abundance foreither for thea1 or b1 subunits of the Na, K-ATPase indicating that an increase in transcription-al activity is not required for the increased enzymeactivity. Although small(-50%) increases ina1subunit abundance were observed during the first24 h, these were not significantly different fromthe levels of expression found in starved fish. Asenzyme activities were determined using crudehomogenates, the increase in Na, K-ATPase activ-ity was not a result of translocation of activesodium pumps to the plasma membrane, althoughtranslocation of inactive pumps with subsequentactivation remains a possibility. As transcriptionalregulation within the first 24 h of feeding seemsunlikely, translational andyor post-translationalregulatory mechanisms must account for the largeand transient increases seen in rectal gland Na, K-ATPase activity. Both phosphorylationy dephos-phorylation and vesicle uptake and recruitment tothe plasma membrane have been implicated inacute hormonal regulation of Na, K-ATPase activ-ity in a number of model systems. Phosphorylationof specific serine and threonine residues on thea-subunit by both protein kinases A(PKA) and C(PKC) have been reported(Chibalin et al., 1992,1993; Pedemonte et al., 1997; Blanco and Mercer,1997) although the physiological significance ofthese covalent modifications still remain to beestablished. More recently small molecular weightproteins (g-subunit and phospholemman) whichare known to co-localise with thea-subunit havebeen implicated as potential regulators of activity(Arystarkhova et al., 1999; Mahmmoud et al.,2000). Rapid and transient increases in rat corticalcollecting duct Na, K-ATPase activity have been
reported following the synergistic application ofaldosterone and arginine vasopressin(Blot-Cha-baud et al. 1990; Coutry et al., 1995). Theseincreases in sodium pump activity were attributedto recruitment andyor activation of existing latentNa, K-ATPase units. In rabbit cortical collectingduct, aldosterone induces the recruitment of latentNa, K-ATPase to the plasma membrane, a processwhich is inhibited by the anti-microtubular agent,colchicine (Blot-Chabaud et al., 1990). Na, K-ATPase activity in cultured dogfish rectal glandcells is also known to be down-regulated afterincubation of cells with colchicine(unpublishedobservation). Although protein kinase-mediatedregulation of Na, K-ATPase activity is a distinctpossibility, in terms of stimulation of the secretoryresponse in the rectal gland epithelia, it is likelyto be secondary to kinase mediated changes inapical chloride conductance induced by the phos-phorylation of CFTR(Greger et al., 1984b). Onething is indeed clear, the rectal gland has a sub-stantial capacity for up-regulating Na, K-ATPaseactivity by a transcriptional-independent mech-anism.The small, but significant increases(q30–
80%) in a1 andb1 mRNA abundance which weredetected between 2 and 10 days after the feedingevent were not accompanied by any measurablechange in Na, K-ATPase activity in rectal glandhomogenates(results not shown). This again sug-gests that unknown regulatory mechanisms down-stream of transcription are the major factorscontrolling Na, K-ATPase activity in the rectalgland. In addition to the small increases in Na, K-ATPase subunit expression, similar increases inthe expression of Na–K–Cl cotransporter andCFTR mRNAs were also found. The increases incotransporter mRNA expression were significant12 hours after the feeding event, with levelsreaching over 150% of controls and remainingelevated for the following 5 days. Increases in theexpression of CFTR mRNA were not so obviousalthough significant increases were found at the 3-and 5-day time points. The delayed increases inexpression of mRNAs for all three ion transporters(CFTR, Na–K–Cl cotransporter and Na, K-ATP-ase subunits) may reflect a general hypertrophyand up-regulation of many transport proteins whichfollow a transient increase in rectal gland activity.It is well accepted that increases inwNax cani
increase mRNA expression of both Na, K-ATPasegenes (Pressley, 1988; Tang and McDonough,

198 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
1992; Yamamoto et al., 1993). Changes in thehormonal status of an animal may also stimulatean increase in the expression of Na, K-ATPasegenes independent ofwNax . Increases in thei
expression of Na, K-ATPase subunits have beenreported following administration of aldosterone(Geering et al., 1982; Wiener et al., 1992; Oguchiet al., 1993; Farman et al., 1994), insulin (Russoand Sweadner, 1993; Tirupattur et al., 1993),thyroid hormones (McDonough et al., 1988;Orlowski and Lingrel, 1990; Hensley et al., 1992;Azuma et al., 1993) and corticosteroids(Orlowskiand Lingrel, 1990; Muto et al., 1996). Thereforealterations in the hormonal status in the dogfishassociated with the feeding episode may be respon-sible for the longer-term changes seen in theexpression of ion transport genes in the rectalgland. In the avian salt gland both the long- andshort-term mechanisms of regulation of secretoryactivity appear to be associated with acetylcholineacting via muscarinic type receptors(Shuttleworthand Hildebrandt, 1999). Both the rapid activationof existing ion channels and transporters and thelonger-term general up-regulation of proteinexpression and cell number are mediated by dif-ferent intracellular pathways which are activatedby a rise inwCa x . Such mechanisms of regula-2q
i
tion would be consistent with the changes in Na,K-ATPase activity and subsequent ion transporterexpression found in the dogfish rectal gland afterfeeding.The increases in mRNA abundance for the
dogfish Na, K-ATPase, Na–K–Cl cotransporterand CFTR chloride channel suggest that changesin plasma ion andyor hormonal balance inducedby feeding can induce transcriptional up-regulationof key ion transporters in the rectal gland.Although these increases in ion transporter mRNAmay serve to ‘prime’ the rectal gland for subse-quent stimulation, it appears that for the Na, K-ATPase at least, additional, and as yet unknown,regulatory systems operating downstream of tran-scriptional events are responsible for the acutecontrol of rectal gland activity. It seems highlylikely that these systems are heavily associatedwith the intermittent activity previously reportedfor the rectal gland in vivo.In conclusion, the results presented in this study
have shown that feeding activity in dogfish hassubsequent physiological effects on the rectalgland secretory epithelia resulting in the delayedbut large and transient increase in Na, K-ATPase
activity, one of the major ion transporting proteinsinvolved in transepithelial chloride secretion. Thisincrease in activity is accompanied by decreasesin both plasma sodium and chloride concentrationsindicative of enhanced rectal gland activity.Northern blot analysis identified homologousmRNAs for CFTR, Na–K–Cl cotransporter anda1yb1 subunits of the Na, K-ATPase in thedogfish (Scyliorhinus canicula) rectal gland andthe abundance of these mRNAs was observed toincrease by up to 50%, but only 1–3 days afterthe single feeding event. As the acute increases inNa, K-ATPase activity were only observed within6–24 h after the feeding episode the increases inenzyme activity are independent of increases inmRNA expression fora andb subunits. We wouldlike to postulate that the acute increases in Na, K-ATPase activity and the longer term increases inion-transporter mRNA abundance are the directresult of changes in hormonal status induced bythe feeding response where the seawater imbibedalong with the squid diet would contribute sub-stantially to the overall salt ingested. These resultssuggest that the rectal gland, in addition to itsbasal homeostatic role in general osmoregulationand NaCl-secretion, has considerable latent secre-tory activity which can be activated acutely andtransiently to remove any excess amounts of NaClabsorbed from the intestine following a feedingepisode.
Acknowledgements
These studies were supported by Project andStudentship awards from the Natural EnvironmentResearch Council.
References
Arystarkhova, E., Wetzel, R.K., Asinovski, N.K., Sweadner,K.J., 1999. Theg-subunit modulates Na and K affinitiesq q
of the renal NayK-ATPase. J. Biol. Chem. 274, 33183–33185.
Azuma, K.K., Hensley, C.B., Tang, M.J., McDonough, A.,1993. Thyroid hormone specifically regulates Na, K-ATPasea2 andb2 isoforms. Am. J. Physiol. 265, C680–C687.
Benz, E.J., Scofield, P., Appel, J.Z., et al., 1992. Cloning andcharacterisation of sequences encoding the major form ofNa, K-ATPase in shark rectal gland. Bull. Mt. Desert. Isl.Biol. Lab. 31, 103–104.
Blanco, G., Mercer, R.W., 1997. Regulation of thea2b1 anda3b1 isozymes of the NayK -ATPase by Ca , PKAq q 2q
and PKC. Ann. N.Y. Acad. Sci. 834, 572–575.Blot-Chabaud, M., Wanstok, F., Bonvalet, J-P., Farman, N.,1990. Cell sodium-induced recruitment of Na, K-ATPase

199S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
pumps in rabbit cortical collecting tubules is aldosteronedependent. J. Biol. Chem. 265, 11676–11681.
Bradford, M.M., 1976. A rapid sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72, 248–254.
Burger, J.W., 1962. Further studies on the function of the rectalgland in spiny dogfish. Physiol. Zool. 35, 205–217.
Cheng, S.H., Rich, D.P., Marshall, J., Gregory, R.J., Welsh,M.J., Smith, A.E., 1991. Phosphorylation of the R domainby cAMP dependent protein kinase regulates the CFTRchloride channel. Cell 66, 1027–1036.
Chibalin, A.V., Vasilets, L.A., Hennekes, H., Pralong, D.,Geering, K., 1992. Phosphorylation of Na, K-ATPase alpha-subunits in microsomes and in homogenates ofXenopusoocytes resulting from the stimulation of protein kinase-Aand protein-kinase-C. J. Biol. Chem. 267, 22378–22384.
Chibalin, A.V., Lopina, O.D., Petukhov, S.P., Vasilets, L.A.,1993. Phosphorylation of the NayK-ATPase by Ca y2q
phospholipid-dependent and cAMP-dependent proteinkinases. Mapping of the region phosphorylated by Cay2q
phospholipid-dependent protein kinase. J. Bioeneg. Biom-emb. 25, 61–66.
Coutry, N., Farman, N., Bonvalet, J.P., Blot-Chabaud, M.,1995. Synergistic action of vasopressin and aldosterone onbaso-lateral Na, K-ATPase in the cortical collecting duct. J.Membr. Biol. 145, 99–106.
Cutler, C.P., Sanders, I.L., Hazon, N., Cramb, G., 1995.Primary sequence, tissue specificity and expression of Na,K-ATPasea1 subunit in the European Eel(Anguilla anguil-la). Comp. Biochem. Physiol. 111B, 567–573.
Cutler, C.P., Sanders, I.L., Hazon, N., Cramb, G., 1997.Expression of Na, K-ATPasea subunit isoforms in theEuropean eel(Anguilla anguilla). Fish Physiol. Biochem.17, 371–376.
Dams, E., Hendriks, L., Van de Peer, Y., et al., 1988.Compilation of small ribosomal subunit RNA sequences.Nucl. Acids Res. 16, r87–r174.
Devor, D.C., Forrest, J.N., Suggs, W.K., Frizzell, R.A., 1995.cAMP activated Cl channels in primary cultures of spinyy
dogfish (Squalus acanthias) rectal gland. Am. J. Physiol.268, C70–C79.
Dulhanty, A.M., Riordan, J.R., 1994. A two-domain model forthe R-domain of the cystic fibrosis transmembrane conduc-tance regulator. FEBS Lett. 343, 109–114.
Emanuel, J.R., Garetz, S., Stone, L., Levenson, R., 1987.Differential expression of Na ,K -ATPasea- andb-subunitq q
mRNAs in rat tissues and cell lines. Proc. Natl. Acad. Sci.USA 84, 9093–9094.
Eveloff, J., Karnaky, K.J., Silva, P., Epstein, F.H., Kinter, W.B.,1979. Elasmobranch rectal gland cell: autoradiographiclocalization ofw HxOuabain sensitive Na, K-ATPase in rectal3
gland of dogfish,Squalus acanthias. J. Cell. Biol. 83, 16–32.
Farman, N., Bonvalet, J.P., Secki, J.R., 1994. Aldosteroneselectively increases Na, K-ATPasea3 subunit mRNAexpression in rat hippocampus. Am. J. Physiol. 266, C579–C589.
Fiske, C.H., Subbarrow, Y., 1925. Phosphate determination. J.Biol. Chem. 66, 375–400.
Forrest, J.N., Wang, F., Beyenbach, K.W., 1983. Perfusion ofisolated tubules of the shark rectal gland. J. Clin. Invest.72, 1163–1167.
Geering, K., Giradet, M., Bron, C., Kraehenbuhl, J.P., Rossier,B.C., 1982. Hormonal regulation of Na, K-ATPase biosyn-thesis in the toad bladder: effect of aldosterone and 3,5,39-thyronine. J. Biol. Chem. 257, 10338–10343.
Gogelein, H., Schlatter, E., Greger, R., 1987. The ’small’conductance chloride channel in the luminal membrane ofthe rectal gland of the dogfish(Squalus acanthias). Pflugers.Arch. 409, 122–125.
Greger, R., Schlatter, E., Gogelein, H., 1984a. Mechanism ofNaCl secretion in the rectal gland of spiny dogfish(Squalusacanthias): 1. Experiments in isolated in vitro perfusedrectal gland tubules. Pflugers. Arch. 402, 63–75.
Greger, R., Schlatter, E., Gogelein, H., 1984b. Mechanism ofNaCl secretion in the rectal gland of spiny dogfish(Squalusacanthias): 3. Effects of stimulation of secretion by cyclicAMP. Pflugers. Arch. 402, 376–384.
Greger, H., Greger, R., Schlatter, E., 1987. Potassium channelsin the basolateral membrane of the rectal gland of thedogfish(Squalus acanthias): regulation and inhibitors. Plug-ers. Arch. 409, 107–113.
Hanrahan, J.W., Duguay, F., Samson, S., Alon, N., Jensen, T.,Riordan, J.R., Grzelczak, Z., 1993. Low-conductance chlo-ride channel activated by cAMP in the rectal gland of theshark (Squalus acanthias) and in cells heterologouslyexpressing the shark CFTR gene. Bull. Mt. Desert. Isl. Biol.Lab. 32, 48–49.
Hensley, C.B., Azuma, K.K., Tang, M-J., McDonough, A.,1992. Thyroid hormone induction of rat myocardial Na, K-ATPase:a1, a2 andb1-mRNA and protein levels at steadystate. Am. J. Physiol. 262, C484–C492.
Karnaky, J.K., Valentich, J.D., Currie, M.G., Oehlenschlager,W.F., Kennedy, M.P., 1991. Atriopeptin stimulates chloridesecretion in cultured shark rectal gland cells. Am. J. Physiol.260, C1125–C1130.
Karnaky, K.J., Stidham, J.D., Nelson, D.S., et al., 1992. C-type natriuretic peptide is a potent secretagogue for thecultured shark(Squalus acanthias) rectal gland. Bull. Mt.Desert. Isl. Biol. Lab. 31, 122–123.
Kennedy, M.P., Valentich, J.D., Currie, M.G., Ochlenschlager,W.F., Karnaky, K.J., 1991. Intracellular cGMP in culturedS.acanthias rectal gland epithelium: effects of atrial natriureticpeptide andE. coli heat stable enterotoxin. Bull. Mt. Desert.Isl. Biol. Lab. 30, 102–103.
Lear, S., Choen, B.J., Silva, P., Lachene, C., Epstein, F.H.,1992. cAMP activates the sodium pump in cultured cells ofthe Elasmobranch rectal gland. J. Am. Soc. Nephrol. 2,1523–1527.
Lytle, C., Forbrush, B., 1992a. Na–K–Cl cotransport in theshark rectal gland. II. Regulation in isolated tubules. Am. J.Physiol. 262, C1009–C1017.
Lytle, C., Forbrush, B., 1992b. The Na–K–Cl cotransportprotein of shark rectal gland. III. Regulation by directphosphorylation. J. Biol. Chem. 267, 25438–25443.
Lytle, C., Xu, J-C., Biemesderfer, D., Haas, M., Forbrush,III.B., 1992. The Na–K–Cl cotransport protein of sharkrectal gland. 1. Development of monoclonal antibodies,immunoaffinity purification and partial biochemical char-acterisation. J. Biol. Chem. 267, 25428–25437.

200 S. MacKenzie et al. / Comparative Biochemistry and Physiology Part B 151 –152 (2002) 185–200
Mahmmoud, Y.A., Vorum, H., Cornelius, F., 2000. Identifica-tion of a phospholemman-like protein from shark rectalglands. J. Biol. Chem. 275, 35969–35977.
Marshall, J., Martin, A.K., Piccioto, M., Hockfield, S., Nairn,A.C., Kaczmarek, L.K., 1991. Identification and localizationof a dogfish homolog of human cystic fibrosis transmem-brane conductance regulator. J. Biol. Chem. 266, 22749–22754.
McDonough, A.A., Brown, T.A., Horowitz, B., et al., 1988.Thyroid hormone coordinately regulates Na, K-ATPasea1andb1-subunit mRNA levels in the kidney. Am. J. Physiol.254, C323–C329.
Muto, S., Nemoto, J., Watanbe, Y., et al., 1996. Differentialregulation of Na, K-ATPase gene expression by corticoste-roids in vascular smooth muscle cells. Am. J. Physiol. 270,C731–C739.
Oguchi, A., Ikeda, U., Kanabe, T., et al., 1993. Regulation ofNa, K-ATPase gene expression by aldosterone in vascularsmooth muscle. Am. J. Physiol. 265, H1167–H1172.
Orlowski, J., Lingrel, J.B., 1990. Thyroid and glucocorticoidhormones regulate the expression of multiple Na, K-ATPasegenes in cultured rat neonatal cardiocytes. J. Biol. Chem.265, 3462–3470.
Pedemonte, C.H., Pressley, T.A., Lokhandwala, M.F., Cinelli,A.R., 1997. Regulation of NayK -ATPase transport activ-q q
ity by protein kinase C. J. Memb. Biol. 155, 219–227.Pressley, T.A., 1988. Ion concentration-dependent regulationof Na, K-pump abundance. J. Membr. Biol. 105, 187–195.
Pressley, T.A., 1992. Phylogenetic conservation of iso-formspecific regions withina-subunit of Na, K-ATPase. Am. J.Physiol. 262, C743–C751.
Russo, J.J., Sweadner, K.J., 1993. Na, K-ATPase subunitisoform pattern modification by mitogenic insulin concen-tration in 3T3-L1 preadipocytes. Am. J. Physiol. 264, C311–C316.
Sambrook, J., Fritsch, E.F., Maniatus, T., second ed, 1989.Molecular Cloning: a Laboratory Manual. Cold SpringHarbor Laboratory Press, Cold Spring Harbor.
Shuttleworth, T.J., 1988. Salt and water balance: extra-renalmechanisms. In: Shuttleworth, T.J.(Ed.), The Physiologyof Elasmobranch Fishes, Springer, London, .
Shuttleworth, T.J., Hildebrandt, J-P., 1999. Vertebrate saltglands: short- and long-term regulation of function. J. Exp.Zool. 283, 689–701.
Shuttleworth, T.J., Thompson, J.L., 1980. The mechanism ofcAMP stimulation of secretion in the dogfish rectal gland.J. Comp. Physiol. 140, 209–216.
Silva, P., Stoff, J., Field, M., Fine, L., Forrest, J.N., Epstein,F.H., 1977. Mechanism of active chloride secretion by sharkrectal gland: role of Na, K-ATPase in chloride transport.Am. J. Physiol. 233, F298–F306.
Stoff, J.S., Rosa, R., Hallac, R., Silva, P., Epstein, F.H., 1979.Hormonal regulation of active chloride transport in thedogfish rectal gland. Am. J. Physiol. 237, F138–F144.
Tang, M-J., McDonough, A.A., 1992. Low K increasesq
Na -K -ATPase a- and b-subunit mRNA and proteinq q
abundance in cultured renal proximal tubule cells. Am. J.Physiol. 263, C436–C442.
Tirupattur, P.R., Ram, J.L., Standley, P.R., Sowers, J.R., 1993.Regulation of Na, K-ATPase expression by insulin in vas-cular smooth muscle cells. Am. J. Hypertens. 6, 626–629.
Valentich, J.D., Forrest, J.N., 1991. Cl secretion by culturedy
shark rectal gland cells. 1. Transepithelial transport. Am. J.Physiol. 260, C813–C823.
Wiener, H., Nielson, J.M., Klaerke, D.A., Jorgensen, P.L.,1992. Aldosterone and thyroid hormone modulation ofa1,b1-mRNA and Na, K-pump sites in rabbit distal colonepithelium. Ann. N.Y. Acad. Sci. 671, 461–463.
Xu, J.-C., Lytle, C., Zhu, T.T., Payne, J.A., Benz, E., Jr, B.,Forbrush, III, 1994. Molecular cloning and functionalexpression of the bumetanide sensitive Na–K–Cl cotrans-porter. Proc. Natl. Acad. Sci. USA. 91, 2201–2205.
Yamamoto, K., Ikeda, U., Seino, Y., Tsuruya, Y., Oguchi, A.,Okada, K., Ishikawa, S.E., Saito, T., Kawakami, K., Hara,Y., Shimada, K., 1993. Regulation of Na, K-adenosinetriphosphatase gene-expression by sodium-ions in culturedneonatal rat cardiocytes. J. Clin. Invest. 92, 1889–1895.