
Activation of the plasma membrane H+-ATPase of Saccharomyces cerevisiae by glucose is mediated by...
11
Activation of the plasma membrane H + -ATPase of Saccharomyces cerevisiae by glucose is mediated by dissociation of the H + -ATPase–acetylated tubulin complex Alexis N. Campetelli 1 , Gabriela Previtali 1 , Carlos A. Arce 2 , He ´ ctor S. Barra 2 and Ce ´ sar H. Casale 1 1 Departamento de Biologı ´a Molecular, Facultad de Ciencias Exactas, Fı ´sico-Quı ´micas y Naturales, Universidad Nacional de Rı ´o Cuarto, Co ´ rdoba, Argentina 2 Centro de Investigaciones en Quı ´mica Biolo ´ gica de Co ´ rdoba (CIQUIBIC), UNC-CONICET, Departamento de Quı ´mica Biolo ´ gica, Facultad de Ciencias Quı ´micas, Universidad Nacional de Co ´ rdoba, Argentina We recently described the interaction of Na + ⁄ K + -ATP ase with acetylated tubulin in neural [1–3] and non- neural cells [4]. Formation of such a complex inhibits ATPase activity. Conversely, dissociation of the com- plex leads to activation of the enzyme. The ATPase– acetylated tubulin complex behaves as a hydrophobic Keywords glucose; H + -ATPase; proton pump; tubulin; yeast Correspondence H. S. Barra, Departamento de Quı ´mica Biolo ´ gica, Facultad de Ciencias Quı ´micas, Universidad Nacional de Co ´ rdoba, Ciudad Universitaria, 5000-Co ´ rdoba, Argentina Fax: +54 3514334074 Tel: +54 3514334168 E-mail: [email protected] (Received 29 July 2005, revised 2 September 2005, accepted 6 September 2005) doi:10.1111/j.1742-4658.2005.04959.x In the yeast Saccharomyces cerevisiae, plasma membrane H + -ATPase is activated by d-glucose. We found that in the absence of glucose, this enzyme forms a complex with acetylated tubulin. Acetylated tubulin usu- ally displays hydrophilic properties, but behaves as a hydrophobic com- pound when complexed with H + -ATPase, and therefore partitions into a detergent phase. When cells were treated with glucose, the H + -ATP- ase–tubulin complex was disrupted, with two consequences, namely (a) the level of acetylated tubulin in the plasma membrane decreased as a function of glucose concentration and (b) the H + -ATPase activity increased as a function of glucose concentration, as measured by both ATP-hydrolyzing capacity and H + -pumping activity. The addition of 2-deoxy-d-glucose inhibited the above glucose-induced phenomena, sug- gesting the involvement of glucose transporters. Whereas total tubulin is distributed uniformly throughout the cell, acetylated tubulin is concentra- ted near the plasma membrane. Results from immunoprecipitation experiments using anti-(acetylated tubulin) and anti-(H + -ATPase) immu- noglobulins indicated a physical interaction between H + -ATPase and acetylated tubulin in the membranes of glucose-starved cells. When cells were pretreated with 1 mm glucose, this interaction was disrupted. Dou- ble immunofluorescence, observed by confocal microscopy, indicated that H + -ATPase and acetylated tubulin partially colocalize at the periphery of glucose-starved cells, with predominance at the outer and inner sides of the membrane, respectively. Colocalization was not observed when cells were pretreated with 1 mm glucose, reinforcing the idea that glucose treatment produces dissociation of the H + -ATPase–tubulin complex. Biochemical experiments using isolated membranes from yeast and purified tubulin from rat brain demonstrated inhibition of H + -ATPase activity by acetylated tubulin and concomitant increase of the H + -ATP ase–tubulin complex. Abbreviations HAT, hydrophobic acetylated tubulin. 5742 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Activation of the plasma membrane H+-ATPase of Saccharomyces cerevisiae by glucose is mediated by...

Activation of the plasma membrane H+-ATPase ofSaccharomyces cerevisiae by glucose is mediated bydissociation of the H+-ATPase–acetylated tubulin complexAlexis N. Campetelli1, Gabriela Previtali1, Carlos A. Arce2, Hector S. Barra2 and Cesar H. Casale1
1 Departamento de Biologıa Molecular, Facultad de Ciencias Exactas, Fısico-Quımicas y Naturales, Universidad Nacional de Rıo Cuarto,
Cordoba, Argentina
2 Centro de Investigaciones en Quımica Biologica de Cordoba (CIQUIBIC), UNC-CONICET, Departamento de Quımica Biologica,
Facultad de Ciencias Quımicas, Universidad Nacional de Cordoba, Argentina
We recently described the interaction of Na+ ⁄K+-ATP
ase with acetylated tubulin in neural [1–3] and non-
neural cells [4]. Formation of such a complex inhibits
ATPase activity. Conversely, dissociation of the com-
plex leads to activation of the enzyme. The ATPase–
acetylated tubulin complex behaves as a hydrophobic
Keywords
glucose; H+-ATPase; proton pump; tubulin;
yeast
Correspondence
H. S. Barra, Departamento de Quımica
Biologica, Facultad de Ciencias Quımicas,
Universidad Nacional de Cordoba, Ciudad
Universitaria, 5000-Cordoba, Argentina
Fax: +54 3514334074
Tel: +54 3514334168
E-mail: [email protected]
(Received 29 July 2005, revised 2 September
2005, accepted 6 September 2005)
doi:10.1111/j.1742-4658.2005.04959.x
In the yeast Saccharomyces cerevisiae, plasma membrane H+-ATPase is
activated by d-glucose. We found that in the absence of glucose, this
enzyme forms a complex with acetylated tubulin. Acetylated tubulin usu-
ally displays hydrophilic properties, but behaves as a hydrophobic com-
pound when complexed with H+-ATPase, and therefore partitions into
a detergent phase. When cells were treated with glucose, the H+-ATP-
ase–tubulin complex was disrupted, with two consequences, namely (a)
the level of acetylated tubulin in the plasma membrane decreased as a
function of glucose concentration and (b) the H+-ATPase activity
increased as a function of glucose concentration, as measured by both
ATP-hydrolyzing capacity and H+-pumping activity. The addition of
2-deoxy-d-glucose inhibited the above glucose-induced phenomena, sug-
gesting the involvement of glucose transporters. Whereas total tubulin is
distributed uniformly throughout the cell, acetylated tubulin is concentra-
ted near the plasma membrane. Results from immunoprecipitation
experiments using anti-(acetylated tubulin) and anti-(H+-ATPase) immu-
noglobulins indicated a physical interaction between H+-ATPase and
acetylated tubulin in the membranes of glucose-starved cells. When cells
were pretreated with 1 mm glucose, this interaction was disrupted. Dou-
ble immunofluorescence, observed by confocal microscopy, indicated that
H+-ATPase and acetylated tubulin partially colocalize at the periphery
of glucose-starved cells, with predominance at the outer and inner sides
of the membrane, respectively. Colocalization was not observed when
cells were pretreated with 1 mm glucose, reinforcing the idea that glucose
treatment produces dissociation of the H+-ATPase–tubulin complex.
Biochemical experiments using isolated membranes from yeast and
purified tubulin from rat brain demonstrated inhibition of H+-ATPase
activity by acetylated tubulin and concomitant increase of the H+-ATP
ase–tubulin complex.
Abbreviations
HAT, hydrophobic acetylated tubulin.
5742 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS
compound, whereas free tubulin is soluble in water.
This property allowed us to isolate the complex, termed
hydrophobic acetylated tubulin (HAT), which was
subsequently quantified by immunoblot with antibody
specific to acetylated tubulin.
The present study is focused on the H+-ATPase of
yeast plasma membrane, another P-type ATPase. This
enzyme is formed by several polypeptides, most prom-
inently a 100 kDa chain that is partly immersed in the
plasma membrane. It functions to hydrolyze ATP and
to transport H+ out of the cell, thereby regulating
internal pH. An important finding was that glucose
activates the plasma membrane H+-ATPase of yeast
cells [5]. This activation has been extensively investi-
gated; however, its molecular mechanism is not com-
pletely understood.
The activation of yeast H+-ATPase by glucose is
regulated at the transcriptional and post-transcrip-
tional levels [6–10]. Glucose increases mRNA synthesis
by inducing transcription of the H+-ATPase gene
(PMA1p), increases phosphorylation of the enzyme,
the Km decreases and the Vmax increases. Proteolytic
degradation of a protein that inhibits the glucose-
activation process also seems to be involved [11]. We
report here that yeast H+-ATPase interacts with tubu-
lin, that such an interaction inhibits enzyme activity,
and that glucose treatment of cells induces dissociation
of the ATPase–tubulin complex with a concomitant
increase in the amount of active enzyme.
Results
Effect of D-glucose on H+-ATPase activity and
HAT quantity
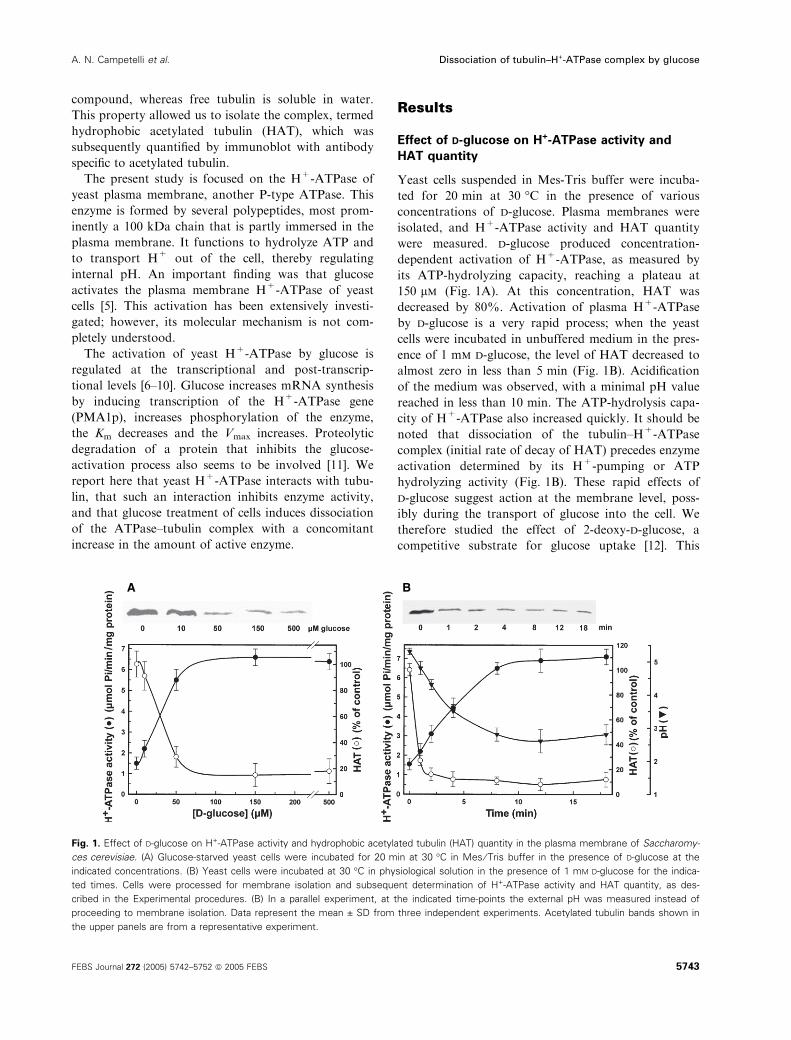
Yeast cells suspended in Mes-Tris buffer were incuba-
ted for 20 min at 30 �C in the presence of various
concentrations of d-glucose. Plasma membranes were
isolated, and H+-ATPase activity and HAT quantity
were measured. d-glucose produced concentration-
dependent activation of H+-ATPase, as measured by
its ATP-hydrolyzing capacity, reaching a plateau at
150 lm (Fig. 1A). At this concentration, HAT was
decreased by 80%. Activation of plasma H+-ATPase
by d-glucose is a very rapid process; when the yeast
cells were incubated in unbuffered medium in the pres-
ence of 1 mm d-glucose, the level of HAT decreased to
almost zero in less than 5 min (Fig. 1B). Acidification
of the medium was observed, with a minimal pH value
reached in less than 10 min. The ATP-hydrolysis capa-
city of H+-ATPase also increased quickly. It should be
noted that dissociation of the tubulin–H+-ATPase
complex (initial rate of decay of HAT) precedes enzyme
activation determined by its H+-pumping or ATP
hydrolyzing activity (Fig. 1B). These rapid effects of
d-glucose suggest action at the membrane level, poss-
ibly during the transport of glucose into the cell. We
therefore studied the effect of 2-deoxy-d-glucose, a
competitive substrate for glucose uptake [12]. This
A B
Fig. 1. Effect of D-glucose on H+-ATPase activity and hydrophobic acetylated tubulin (HAT) quantity in the plasma membrane of Saccharomy-
ces cerevisiae. (A) Glucose-starved yeast cells were incubated for 20 min at 30 �C in Mes ⁄ Tris buffer in the presence of D-glucose at the
indicated concentrations. (B) Yeast cells were incubated at 30 �C in physiological solution in the presence of 1 mM D-glucose for the indica-
ted times. Cells were processed for membrane isolation and subsequent determination of H+-ATPase activity and HAT quantity, as des-
cribed in the Experimental procedures. (B) In a parallel experiment, at the indicated time-points the external pH was measured instead of
proceeding to membrane isolation. Data represent the mean ± SD from three independent experiments. Acetylated tubulin bands shown in
the upper panels are from a representative experiment.
A. N. Campetelli et al. Dissociation of tubulin–H+-ATPase complex by glucose
FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS 5743

compound, at a concentration of 1 mm in the culture
medium, blocked the H+-ATPase activation and HAT
decrease induced by 30 min treatment of cells with
0.1 mm d-glucose (Table 1, Exp. 1); however, it had no
effect when used at 10 mm. The blocking of the ATP-
ase activating effect of glucose was not the result of a
toxic effect of 2-deoxy-d-glucose, as subsequent incuba-
tion in the presence of 10 mm glucose resulted again in
H+-ATPase activation with concomitant dissociation
of the ATPase–tubulin complex (HAT decrease)
(Table 1, Exp. 2). The blocking effect of 2-deoxy-d-
glucose is immediate as, 1 min after its addition, the
H+-pumping activity of H+-ATPase ceased (Fig. 2A)
and the HAT quantity began to increase (Fig. 2B).
When isolated plasma membranes from glucose-
starved yeast were used instead of intact cells, treat-
ment with 1 mm d-glucose for 20 min had no effect on
the ATP-hydrolyzing capacity of H+-ATPase, nor on
HAT quantity (data not shown).
Previous studies [2–4], and the findings described
below, suggest that the decrease of HAT in the plasma
membrane should be interpreted as a lower level of the
H+-ATPase–tubulin complex. We therefore presume
that the immediate uptake of glucose after its addition
induces dissociation of the H+-ATPase–acetylated tub-
ulin complex, resulting in an increased enzyme activity,
characterized by higher ATPase hydrolyzing capacity
and H+-pumping activity.
Characterization of acetylated tubulin in
Saccharomyces cerevisiae
Localization of acetylated tubulin in cells was studied
by immunofluorescence. Acetylated tubulin (Fig. 3A)
Table 1. Effect of 2-deoxy-D-glucose on H+-ATPase activation induced by D-glucose. Glucose-starved cells suspended in Mes-Tris buffer
were incubated at 37 �C under the indicated conditions for 30 min (experiment 1) or 60 min (experiment 2), then frozen in liquid nitrogen,
and hydrophobic acetylated tubulin (HAT) and H+-ATPase activities were measured. Values represent the mean ± SD from three independ-
ent experiments.
Experiment
Additions and
30 min of incubation
Additions and 30 min
of further incubation
HAT
(% of control)
H+-ATPase activity
(lmol PiÆmin)1Æmg protein)1)
1 No addition (control) 100 ± 11 1.49 ± 0.6
0.1 mM D-glucose 20 ± 7 8.42 ± 0.27
0.1 mM D-glucose plus 82 ± 9 2.21 ± 0.25
1 mM 2-deoxy-D-glucose
10 mM D-glucose plus 29 ± 13 5.72 ± 0.5
1 mM 2-deoxy-D-glucose
2 No addition (control) No addition 100 ± 9 1.39 ± 0.5
0.1 mM D-glucose plus
1 mM 2-deoxy-D-glucose
10 mM D-glucose 28 ± 12 5.60 ± 0.7
A
B
Fig. 2. Effect of 2-deoxy-D-glucose on H+-pumping activity and hydrophobic acetylated tubulin (HAT) quantity of Saccharomyces cerevisiae.
(A) Yeast cells were incubated at 30 �C in physiological solution in the presence of 1 mM D-glucose, and the external pH was measured at
the indicated time-points. At the time indicated by the arrow, 2-deoxy-D-glucose (10 mM final concentration) was added (d). A control, with-
out added 2-deoxy-D-glucose, was included (s). (B) In a parallel experiment, instead of pH measurement, HAT quantity was measured. Data
represent the mean ± SD from three independent experiments. Acetylated tubulin bands shown are from a representative experiment.
Dissociation of tubulin–H+-ATPase complex by glucose A. N. Campetelli et al.
5744 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS
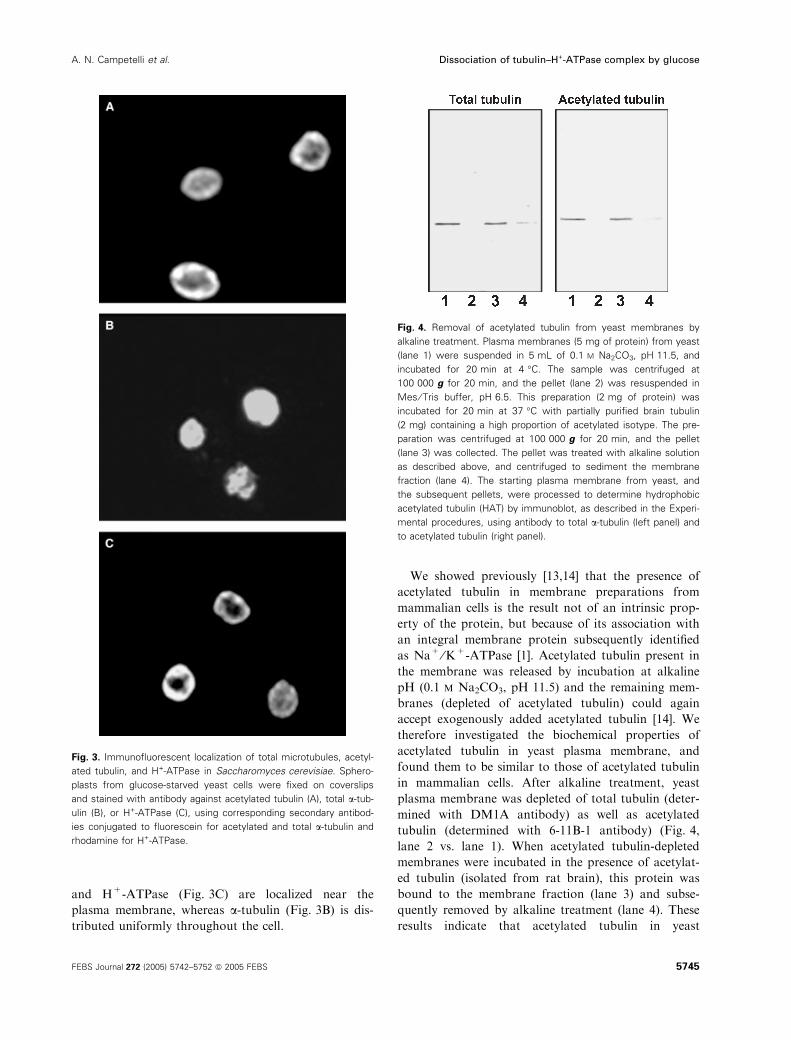
and H+-ATPase (Fig. 3C) are localized near the
plasma membrane, whereas a-tubulin (Fig. 3B) is dis-
tributed uniformly throughout the cell.
We showed previously [13,14] that the presence of
acetylated tubulin in membrane preparations from
mammalian cells is the result not of an intrinsic prop-
erty of the protein, but because of its association with
an integral membrane protein subsequently identified
as Na+ ⁄K+-ATPase [1]. Acetylated tubulin present in
the membrane was released by incubation at alkaline
pH (0.1 m Na2CO3, pH 11.5) and the remaining mem-
branes (depleted of acetylated tubulin) could again
accept exogenously added acetylated tubulin [14]. We
therefore investigated the biochemical properties of
acetylated tubulin in yeast plasma membrane, and
found them to be similar to those of acetylated tubulin
in mammalian cells. After alkaline treatment, yeast
plasma membrane was depleted of total tubulin (deter-
mined with DM1A antibody) as well as acetylated
tubulin (determined with 6-11B-1 antibody) (Fig. 4,
lane 2 vs. lane 1). When acetylated tubulin-depleted
membranes were incubated in the presence of acetylat-
ed tubulin (isolated from rat brain), this protein was
bound to the membrane fraction (lane 3) and subse-
quently removed by alkaline treatment (lane 4). These
results indicate that acetylated tubulin in yeast
Fig. 3. Immunofluorescent localization of total microtubules, acetyl-
ated tubulin, and H+-ATPase in Saccharomyces cerevisiae. Sphero-
plasts from glucose-starved yeast cells were fixed on coverslips
and stained with antibody against acetylated tubulin (A), total a-tub-
ulin (B), or H+-ATPase (C), using corresponding secondary antibod-
ies conjugated to fluorescein for acetylated and total a-tubulin and
rhodamine for H+-ATPase.
Fig. 4. Removal of acetylated tubulin from yeast membranes by
alkaline treatment. Plasma membranes (5 mg of protein) from yeast
(lane 1) were suspended in 5 mL of 0.1 M Na2CO3, pH 11.5, and
incubated for 20 min at 4 �C. The sample was centrifuged at
100 000 g for 20 min, and the pellet (lane 2) was resuspended in
Mes ⁄ Tris buffer, pH 6.5. This preparation (2 mg of protein) was
incubated for 20 min at 37 �C with partially purified brain tubulin
(2 mg) containing a high proportion of acetylated isotype. The pre-
paration was centrifuged at 100 000 g for 20 min, and the pellet
(lane 3) was collected. The pellet was treated with alkaline solution
as described above, and centrifuged to sediment the membrane
fraction (lane 4). The starting plasma membrane from yeast, and
the subsequent pellets, were processed to determine hydrophobic
acetylated tubulin (HAT) by immunoblot, as described in the Experi-
mental procedures, using antibody to total a-tubulin (left panel) and
to acetylated tubulin (right panel).
A. N. Campetelli et al. Dissociation of tubulin–H+-ATPase complex by glucose
FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS 5745

membrane is a peripheral protein that is bound to
some integral membrane protein, presumably H+-ATP-
ase, in a manner similar to its association with
Na+ ⁄K+-ATPase in the plasma membrane of mamma-
lian cells.
To confirm the interaction of acetylated tubulin with
H+-ATPase and the presence of the H+-ATPase–acet-
ylated tubulin complex in membranes of glucose-starved
yeast, we performed immunoprecipitation experiments
using either anti-(acetylated tubulin) immunoglobulin
or anti-(H+-ATPase) immunoglobulin bound to Seph-
arose beads. When detergent-solubilized plasma mem-
branes from glucose-starved yeast were treated with
anti-(acetylated tubulin)–Sepharose beads and then cen-
trifuged, in addition to tubulin, the pellet contained the
110 kDa H+-ATPase polypeptide (Fig. 5B, lane –).
When solubilized membranes were treated with anti-
H+-ATPase–Sepharose beads, the pellet contained acet-
ylated tubulin in addition to the 110 kDa H+-ATPase
polypeptide (Fig. 5C, lane –). These findings indicate
that a complex containing both acetylated tubulin and
H+-ATPase is present in solubilized yeast membranes
from glucose-starved cells. When cells were pretreated
with 1 mm glucose for 1 h and then processed for im-
munoprecipitation with anti-H+-ATPase–Sepharose
beads, the pellet contained the 110 kDa H+-ATPase
polypeptide and only a minor amount of acetylated tub-
ulin (Fig. 5C, lane +), indicating that glucose treatment
induced dissociation of the H+-ATPase–acetylated tub-
ulin complex. An identical conclusion was drawn from
immunoprecipitation experiments with anti-(acetylated
tubulin)–Sepharose beads (Fig. 5B, lane +). Figure 5D
shows that Sepharose beads without bound antibodies
practically do not adsorb tubulin or ATPase. To esti-
mate the proportion of acetylated tubulin and H+-ATP-
ase that was immunoprecipitated, we determined also
the amount of each protein present in the solubilized-
membrane preparations before immunoprecipitation
(Fig. 5A). From visual comparison of blots, it can be
seen that most of the acetylated tubulin and ATPase
were involved in the immunoprecipitation process. By
measuring the intensity of the ATPase band in Fig. 5B
(lane –) with respect to that in Fig. 5A (lane –), it was
calculated that 88 ± 15% of ATPase was associated
with acetylated tubulin. In addition, by comparing the
intensity of the ATPase band in lane (+) with respect to
that in lane (–) in Fig. 5B, it was calculated that
88 ± 12% of the complex was dissociated by glucose
treatment. These results represent the mean ± SD val-
ues from three independent experiments.
Further evidence for the occurrence of the H+-ATP-
ase–acetylated tubulin complex in the plasma mem-
brane of S. cerevisiae was the co-localization of the
two components in confocal immunofluorescence ana-
lysis. The image obtained with anti-(acetylated tubulin)
immunoglobulin partially overlapped that obtained
with anti-(H+-ATPase) immunoglobulin (Fig. 6).
Enlargement of a portion of the merge image (Fig. 6D)
shows that H+-ATPase (red) is localized on the
plasma membrane without extending into the cyto-
plasm, while acetylated tubulin (green) is localized near
the membrane, overlapping (yellow) with the inner side
of the region occupied by H+-ATPase. The more
external region of the membrane H+-ATPase is not in
contact with acetylated tubulin. When cells were pre-
treated with 1 mm glucose for 1 h and then processed
for immunofluorescence, acetylated tubulin was distri-
buted uniformly throughout the cytoplasm and did not
Fig. 5. Physical interaction between acetylated tubulin and H+-ATP-
ase in Saccharomyces cerevisiae membrane. (A) To estimate the
total amount of both acetylated tubulin and ATPase present in the
detergent-solubilized yeast plasma membrane from cells previously
treated (lanes +) or not treated (lanes –) with 1 mM glucose, 23 lL
aliquots were subjected to SDS ⁄ PAGE and simultaneous immuno-
blot staining with anti-(acetylated tubulin) and anti-(H+-ATPase)
immunoglobulin. (B) and (C) An aliquot of 0.7 mL of detergent-solubi-
lized yeast plasma membrane from cells previously treated (lanes +)
or not treated (lanes –) with 1 mM glucose was mixed with 0.3 mL
of packed anti-acetylated tubulin (6-11-B-1)–Sepharose beads (B), or
anti-H+-ATPase (Pma1p)–Sepharose beads (C), and incubated at
20 �C for 30 min. The samples were then centrifuged, and the preci-
pitated material was washed five times with 50 mM Tris ⁄HCl buffer,pH 7.4, containing 150 mM NaCl (TBS)-Triton. A fraction of 50 lL of
packed beads was resuspended in 50 lL of Laemmli sample buffer
[13], treated at 50 �C for 15 min, and centrifuged. Twenty microlitre
aliquots of the soluble fractions were subjected to SDS ⁄ PAGE and
then to simultaneous immunoblot staining with anti-(acetylated tubu-
lin) and anti-(H+-ATPase) immunoglobulin. (D) A control was run in
parallel using glycine–Sepharose instead of antibodies–Sepharose.
Note that the protein bands shown arise from equal amounts of
membrane preparations.
Dissociation of tubulin–H+-ATPase complex by glucose A. N. Campetelli et al.
5746 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS
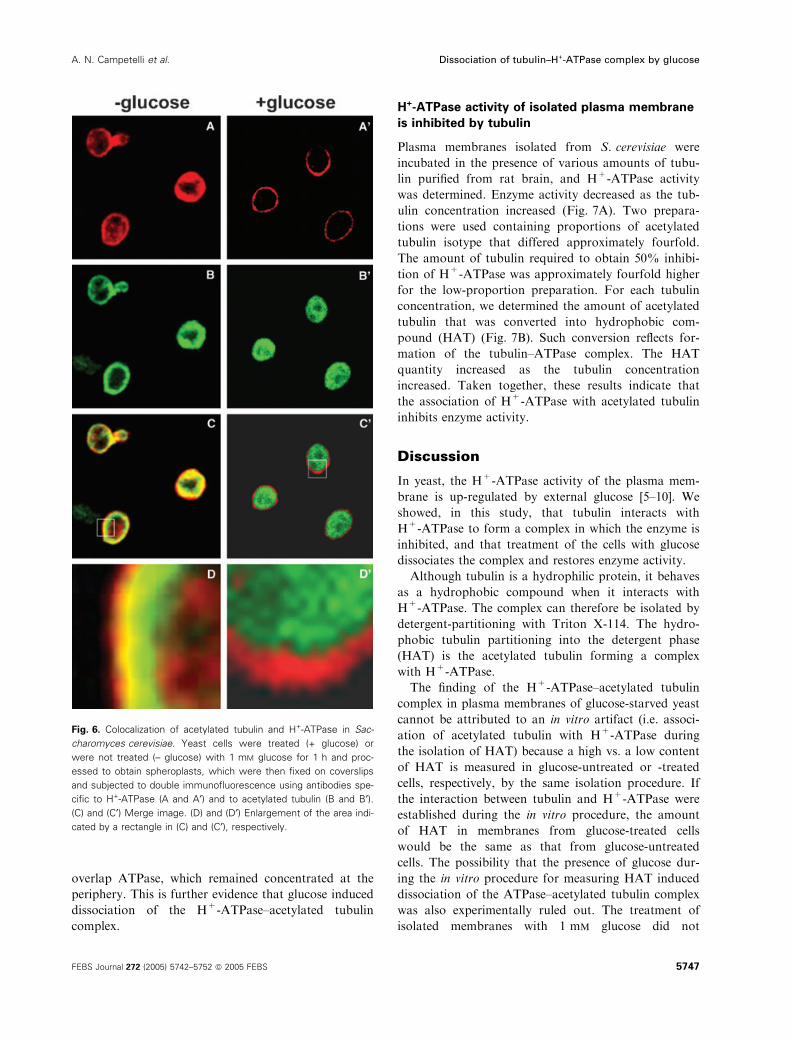
overlap ATPase, which remained concentrated at the
periphery. This is further evidence that glucose induced
dissociation of the H+-ATPase–acetylated tubulin
complex.
H+-ATPase activity of isolated plasma membrane
is inhibited by tubulin
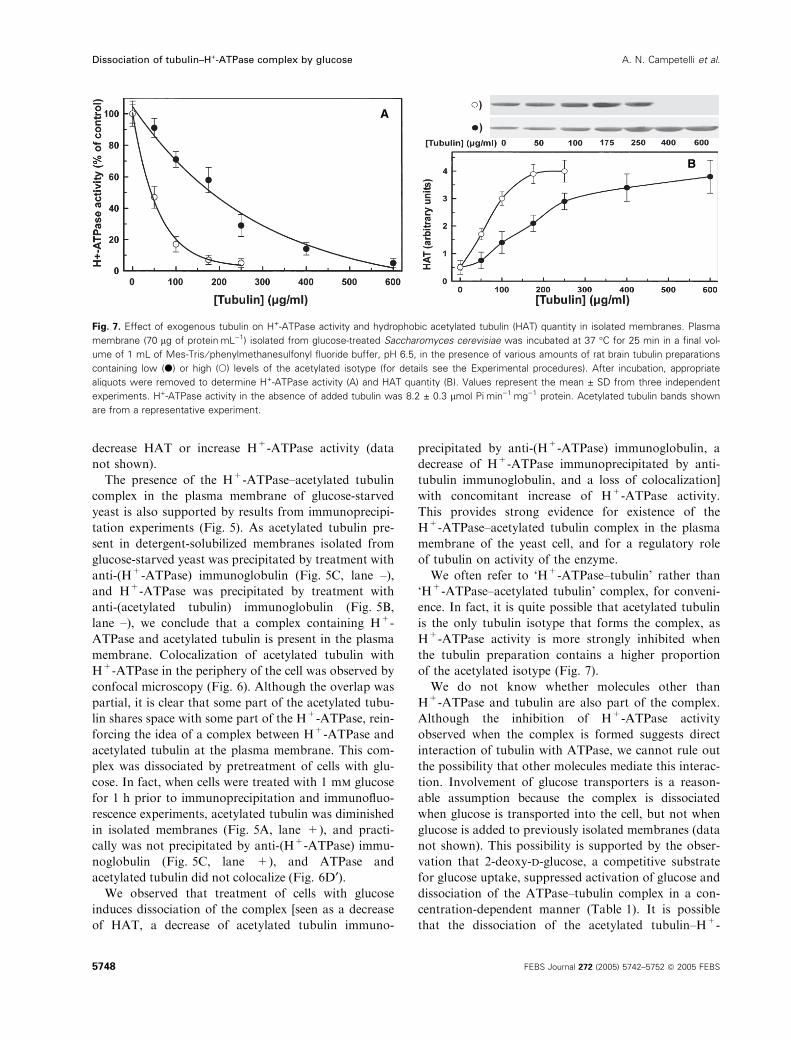
Plasma membranes isolated from S. cerevisiae were
incubated in the presence of various amounts of tubu-
lin purified from rat brain, and H+-ATPase activity
was determined. Enzyme activity decreased as the tub-
ulin concentration increased (Fig. 7A). Two prepara-
tions were used containing proportions of acetylated
tubulin isotype that differed approximately fourfold.
The amount of tubulin required to obtain 50% inhibi-
tion of H+-ATPase was approximately fourfold higher
for the low-proportion preparation. For each tubulin
concentration, we determined the amount of acetylated
tubulin that was converted into hydrophobic com-
pound (HAT) (Fig. 7B). Such conversion reflects for-
mation of the tubulin–ATPase complex. The HAT
quantity increased as the tubulin concentration
increased. Taken together, these results indicate that
the association of H+-ATPase with acetylated tubulin
inhibits enzyme activity.
Discussion
In yeast, the H+-ATPase activity of the plasma mem-
brane is up-regulated by external glucose [5–10]. We
showed, in this study, that tubulin interacts with
H+-ATPase to form a complex in which the enzyme is
inhibited, and that treatment of the cells with glucose
dissociates the complex and restores enzyme activity.
Although tubulin is a hydrophilic protein, it behaves
as a hydrophobic compound when it interacts with
H+-ATPase. The complex can therefore be isolated by
detergent-partitioning with Triton X-114. The hydro-
phobic tubulin partitioning into the detergent phase
(HAT) is the acetylated tubulin forming a complex
with H+-ATPase.
The finding of the H+-ATPase–acetylated tubulin
complex in plasma membranes of glucose-starved yeast
cannot be attributed to an in vitro artifact (i.e. associ-
ation of acetylated tubulin with H+-ATPase during
the isolation of HAT) because a high vs. a low content
of HAT is measured in glucose-untreated or -treated
cells, respectively, by the same isolation procedure. If
the interaction between tubulin and H+-ATPase were
established during the in vitro procedure, the amount
of HAT in membranes from glucose-treated cells
would be the same as that from glucose-untreated
cells. The possibility that the presence of glucose dur-
ing the in vitro procedure for measuring HAT induced
dissociation of the ATPase–acetylated tubulin complex
was also experimentally ruled out. The treatment of
isolated membranes with 1 mm glucose did not
Fig. 6. Colocalization of acetylated tubulin and H+-ATPase in Sac-
charomyces cerevisiae. Yeast cells were treated (+ glucose) or
were not treated (– glucose) with 1 mM glucose for 1 h and proc-
essed to obtain spheroplasts, which were then fixed on coverslips
and subjected to double immunofluorescence using antibodies spe-
cific to H+-ATPase (A and A¢) and to acetylated tubulin (B and B¢).(C) and (C¢) Merge image. (D) and (D¢) Enlargement of the area indi-
cated by a rectangle in (C) and (C¢), respectively.
A. N. Campetelli et al. Dissociation of tubulin–H+-ATPase complex by glucose
FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS 5747
decrease HAT or increase H+-ATPase activity (data
not shown).
The presence of the H+-ATPase–acetylated tubulin
complex in the plasma membrane of glucose-starved
yeast is also supported by results from immunoprecipi-
tation experiments (Fig. 5). As acetylated tubulin pre-
sent in detergent-solubilized membranes isolated from
glucose-starved yeast was precipitated by treatment with
anti-(H+-ATPase) immunoglobulin (Fig. 5C, lane –),
and H+-ATPase was precipitated by treatment with
anti-(acetylated tubulin) immunoglobulin (Fig. 5B,
lane –), we conclude that a complex containing H+-
ATPase and acetylated tubulin is present in the plasma
membrane. Colocalization of acetylated tubulin with
H+-ATPase in the periphery of the cell was observed by
confocal microscopy (Fig. 6). Although the overlap was
partial, it is clear that some part of the acetylated tubu-
lin shares space with some part of the H+-ATPase, rein-
forcing the idea of a complex between H+-ATPase and
acetylated tubulin at the plasma membrane. This com-
plex was dissociated by pretreatment of cells with glu-
cose. In fact, when cells were treated with 1 mm glucose
for 1 h prior to immunoprecipitation and immunofluo-
rescence experiments, acetylated tubulin was diminished
in isolated membranes (Fig. 5A, lane +), and practi-
cally was not precipitated by anti-(H+-ATPase) immu-
noglobulin (Fig. 5C, lane +), and ATPase and
acetylated tubulin did not colocalize (Fig. 6D¢).We observed that treatment of cells with glucose
induces dissociation of the complex [seen as a decrease
of HAT, a decrease of acetylated tubulin immuno-
precipitated by anti-(H+-ATPase) immunoglobulin, a
decrease of H+-ATPase immunoprecipitated by anti-
tubulin immunoglobulin, and a loss of colocalization]
with concomitant increase of H+-ATPase activity.
This provides strong evidence for existence of the
H+-ATPase–acetylated tubulin complex in the plasma
membrane of the yeast cell, and for a regulatory role
of tubulin on activity of the enzyme.
We often refer to ‘H+-ATPase–tubulin’ rather than
‘H+-ATPase–acetylated tubulin’ complex, for conveni-
ence. In fact, it is quite possible that acetylated tubulin
is the only tubulin isotype that forms the complex, as
H+-ATPase activity is more strongly inhibited when
the tubulin preparation contains a higher proportion
of the acetylated isotype (Fig. 7).
We do not know whether molecules other than
H+-ATPase and tubulin are also part of the complex.
Although the inhibition of H+-ATPase activity
observed when the complex is formed suggests direct
interaction of tubulin with ATPase, we cannot rule out
the possibility that other molecules mediate this interac-
tion. Involvement of glucose transporters is a reason-
able assumption because the complex is dissociated
when glucose is transported into the cell, but not when
glucose is added to previously isolated membranes (data
not shown). This possibility is supported by the obser-
vation that 2-deoxy-d-glucose, a competitive substrate
for glucose uptake, suppressed activation of glucose and
dissociation of the ATPase–tubulin complex in a con-
centration-dependent manner (Table 1). It is possible
that the dissociation of the acetylated tubulin–H+-
A
B
Fig. 7. Effect of exogenous tubulin on H+-ATPase activity and hydrophobic acetylated tubulin (HAT) quantity in isolated membranes. Plasma
membrane (70 lg of proteinÆmL)1) isolated from glucose-treated Saccharomyces cerevisiae was incubated at 37 �C for 25 min in a final vol-
ume of 1 mL of Mes-Tris ⁄ phenylmethanesulfonyl fluoride buffer, pH 6.5, in the presence of various amounts of rat brain tubulin preparations
containing low (d) or high (s) levels of the acetylated isotype (for details see the Experimental procedures). After incubation, appropriate
aliquots were removed to determine H+-ATPase activity (A) and HAT quantity (B). Values represent the mean ± SD from three independent
experiments. H+-ATPase activity in the absence of added tubulin was 8.2 ± 0.3 lmol PiÆmin)1Æmg)1 protein. Acetylated tubulin bands shown
are from a representative experiment.
Dissociation of tubulin–H+-ATPase complex by glucose A. N. Campetelli et al.
5748 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS

ATPase complex requires the presence of Snf3p (a glu-
cose sensor), Gpa2 protein (a G protein) [10] and
protein kinases [15], which were demonstrated to parti-
cipate in the glucose-induced activation of the plasma
membrane ATPase.
Our results in the present work show that a glucose-
sensitive association ⁄dissociation of acetylated tubulin
and H+-ATPase participates in an early stage of the
mechanism that leads to, respectively, inhibition and
activation of the enzyme. The increase of enzyme
activity (determined by ATP-hydrolyzing capacity or
by H+-pumping activity) starts immediately after glu-
cose addition and reaches a maximum in � 10 min,
whereas dissociation of the ATPase–tubulin complex
(decrease of HAT quantity) is completed within the
first 2 min (Fig. 1B, and Fig. 2A,B). The reason for
this temporal difference is not clear. It is possible that
besides dissociation of the complex, the enzyme
requires some additional, time-consuming process for
its activation. Anyway, the important conclusion from
these experiments is that dissociation of the ATPase–
tubulin complex is at least part of the reason for the
increased enzyme activity induced by external glucose.
In agreement with this view, exogenously added tubu-
lin was bound to membrane H+-ATPase and inhibited
enzyme activity (Fig. 7), indicating that H+-ATPase
activation by glucose is caused by an increased concen-
tration of active enzyme. Similar effects of tubulin
were demonstrated for Na+ ⁄K+-ATPase in our previ-
ous studies [2–4].
Interestingly, for both H+-ATPase in yeast and
Na+ ⁄K+-ATPase in mammalian cells [3,4], the
ATPase–acetylated tubulin complex can be dissociated
during the uptake of d-glucose and l-glutamate,
respectively. In either case, corresponding enzyme
activity increases upon dissociation of the complex.
We suspect that these events are part of a signal trans-
duction cascade and are accordingly investigating the
nature of membranous and cytoplasmic components of
the system, interactions between components and
modulation of these interactions.
Experimental procedures
Materials
Triton X-114, ATP, d-glucose, 2-deoxy-d-glucose, mouse
monoclonal antibody DM1A specific to a-tubulin, mouse
monoclonal antibody 6-11B-1 specific to acetylated tubulin,
anti-mouse and anti-rabbit IgG conjugated to peroxidase
were from Sigma Chemical Co. (St Louis, MO, USA).
[32P]ATP[cP] was from Perkin-Elmer (Wellesley, MA,
USA). Rabbit polyclonal antibody Pma1p, specific to
H+-ATPase, was provided by R. Serrano (Instituto de Bio-
logıa Molecular y Celular de Plantas, Valencia, Spain).
Yeast strain and growth conditions
S. cerevisiae, strain CECT 1891 (Spanish Type Culture
Collection, University of Valencia, Valencia, Spain), was
used. Cells were grown on synthetic medium YP [0.5%
(w ⁄ v) yeast extract and 0.5% (w ⁄ v) peptone] containing
4% (w ⁄ v) glucose. Cells were grown in a rotary incubator
(New Brunswick model G24; 200 r.p.m.; NJ, USA) at
30 �C until the end of the exponential phase. Cells were
then harvested (centrifugation at 1000 g for 10 min), sus-
pended in Mes ⁄Tris buffer (100 mm Mes ⁄Tris, pH 6.5)
and magnetically stirred for 60 min to eliminate glucose
activation [5]. These cells are referred to as ‘glucose-
starved cells’.
Yeast plasma membrane preparation
Plasma membranes were isolated by a modification of the
method of Villalba et al. [16]. Briefly, yeast cells were suspen-
ded in Mes ⁄Tris buffer supplemented with 1 mm phenyl-
methanesulfonyl fluoride (Mes ⁄Tris ⁄ phenylmethanesulfonyl
fluoride buffer), with or without d-glucose, according to the
conditions of each experiment, homogenized by vigorous
shaking with glass beads, and centrifuged at 1000 g for
10 min. The resulting supernatant was centrifuged at
70 000 g for 60 min to obtain the total membrane fraction.
The total membrane fraction from 10 g of cells was suspen-
ded in 3 mL of Mes ⁄Tris ⁄ phenylmethanesulfonyl fluoride
buffer. The suspension was applied to a discontinuous gradi-
ent – from 5.0 mL of 60% (w ⁄ v) sucrose to 5.0 mL of 40%
(w ⁄ v) sucrose – in 10 mm Tris ⁄HCl buffer (pH 7.6) contain-
ing 1 mm EDTA and 1 mm dithiothreitol. Plasma mem-
branes were centrifuged for 3 h at 100 000 g, collected
from the 40 ⁄ 60% sucrose interface, diluted 10-fold with
Mes ⁄Tris ⁄ phenylmethanesulfonyl fluoride buffer and centri-
fuged at 100 000 g for 1 h. The resulting pellet was resus-
pended in Mes ⁄Tris ⁄ phenylmethanesulfonyl fluoride buffer
and stored at )70 �C until use (maximum storage time
3 months).
Plasma membrane H+-ATPase activity assay
We used the [32P]ATP[cP] hydrolysis method of Malpartida
& Serrano [6]. The incubation mixture (0.5 mL final vol-
ume) contained Mes ⁄Tris ⁄phenylmethanesulfonyl fluoride
buffer, 10 mm MgCl2, 2 mm [32P]ATP[cP] (450 d.p.m.Ænmol)1) and 7 lgÆmL)1 plasma membrane protein. After
20 min at 30 �C, the reaction was stopped by adding 50 lLof 66% (w ⁄ v) trichloroacetic acid per mL of incubation
mixture, and the released 32Pi was quantified. Plasma mem-
brane H+-ATPase activity was calculated as the difference
A. N. Campetelli et al. Dissociation of tubulin–H+-ATPase complex by glucose
FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS 5749

of ATP hydrolysis in the presence vs. absence of 100 lmsodium orthovanadate.
Isolation and determination of HAT
HAT was isolated into the Triton X-114 phase, as des-
cribed previously [2], except that plasma membranes were
solubilized with Triton X-100, as described below, before
adding Triton X-114 and partitioning. Membranes from
S. cerevisiae (28 lg of protein) were washed once with
50 mm Tris ⁄HCl buffer, pH 7.4, containing 150 mm NaCl
(TBS) and immediately solubilized in 1 mL of TBS con-
taining 1% (v ⁄ v) Triton X-100. After 30 min at 0 �C, thepreparation was centrifuged at 100 000 g for 15 min, and
the supernatant fraction was processed for HAT isolation
by partitioning in Triton X-114 (1% final detergent con-
centration). For phase separation, the preparation was
warmed at 37 �C for 5 min and centrifuged at 600 g for
5 min. The aqueous upper phase and detergent-rich
lower phase were carefully separated, and the detergent-
rich phase (which contains HAT) was washed once
with TBS. Aliquots were subjected to electrophoresis
and immunoblotting to determine acetylated and total
tubulin.
Electrophoresis and immunoblotting
Proteins were separated by SDS ⁄PAGE on 10% (w ⁄ v)polyacrylamide slab gels [17], transferred to nitrocellulose,
and reacted with mouse monoclonal antibody 6-11B-1
(dilution 1 : 1000) to determine acetylated tubulin [18],
mouse monoclonal antibody DM1A (dilution 1 : 1000) to
determine total a-tubulin, or rabbit polyclonal antibody
Pma1p (dilution 1 : 1000) to determine plasma membrane
H+-ATPase [15]. The nitrocellulose sheet was reacted with
anti-mouse (for 6-11B-1 and DM1A antibodies) or anti-
rabbit (for Pma1p antibody) IgG conjugated with peroxi-
dase. Intensities of tubulin bands were quantified by Scion
imaging software.
Measurement of H+-pumping activity
Samples of cells (� 50 mg) were washed twice, suspended
in 15 mL of 0.9% (w ⁄ v) NaCl, and stirred gently at 30 �Cin a beaker. Incubation conditions were as indicated in each
experiment. External pH was measured using a pH meter
with a calomel electrode.
Preparation of spheroplasts from yeast
S. cerevisiae cells (50 mg fresh weight) were washed twice
in 2 mL of 0.1 m Tris ⁄HCl, pH 7.2, containing 5 mm
EGTA and 5 mm dithiothreitol, incubated with stirring
for 10 min, washed with water (containing 1 mm dithio-
threitol), and resuspended in 2 mL of 0.1 m Tris ⁄HCl,
pH 7.2, containing 1 m sorbitol and 1 mm dithiothreitol.
Zymolyase was added to the suspension (final concentra-
tion 0.1 mgÆmL)1), gently stirred for 30 min, and cells
were harvested by centrifugation. The pellet was resus-
pended in 2 mL of 0.1 m Tris ⁄HCl, pH 7.2, containing
5 mm EGTA and 5 mm dithiothreitol, and used immedi-
ately. All procedures were carried out at room tempera-
ture.
Immunofluorescence
Spheroplasts were fixed with anhydrous methanol at
)20 �C on coverslips. Samples were washed, incubated
with 2% (w ⁄ v) BSA in NaCl ⁄Pi (PBS) for 30 min, and
stained by indirect immunofluorescence, as described by
DeWitt et al. [19]. Two primary antibodies were used:
mouse 6-11B-1 monoclonal antibody (diluted 1 : 200) to
visualize acetylated tubulin, and rabbit Pma1p polyclonal
antibody (diluted 1 : 200) to determine plasma membrane
H+-ATPase. Fluorescein-conjugated anti-mouse IgG and
Rhodamine-conjugated anti-rabbit immunoglobulin, at a
1 : 400 dilution, were used as secondary antibodies,
respectively. Coverslips were mounted in FluorSave and
observed with an LSM 5 Pascal confocal microscope
(Zeiss, Jena, Germany) using dual channel filters for sim-
ultaneous viewing of rhodamine and fluorescein isothio-
cyanate fluorochromes.
Tubulin preparations containing different
proportions of acetylated tubulin
The procedure used to isolate rat brain tubulin prepara-
tions containing different proportions of the acetylated iso-
type has been described previously [2]. These preparations
contained low and high acetylated tubulin proportions dif-
fering by a factor of � 4.
Preparation of 6-11B-1–Sepharose and Pma1p–
Sepharose
6-11B-1 and Pma1p antibodies were covalently bound to
cyanogen bromide-activated Sepharose 4B following the
procedure of Hubert et al. [20] with slight modifications.
Sepharose beads were washed with a 100-fold volume
excess of 0.001 m HCl at 21 �C. The resulting packed beads
(0.3 mL) were mixed with 2 mg of 6-11B-1 antibody (or
2 mg of Pma1p antibody) in 1 mL of coupling buffer
(0.5 m NaCl containing 0.2 m NaHCO3, pH 8.2). The mix-
ture was agitated on a platform rocker for 4 h at 21 �C,and loaded into a small chromatographic column.
Unbound 6-11B-1 antibody (or Pma1p antibody) was
removed by filtration and by washing with 10 mL of coup-
ling buffer. The 6-11B-1-Sepharose (or Pma1p-Sepharose)
Dissociation of tubulin–H+-ATPase complex by glucose A. N. Campetelli et al.
5750 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS

beads were loaded into a beaker, and added with 2 mL of
coupling buffer containing 0.2 m glycine to block unreacted
Sepharose sites. The mixture was agitated for 2 h at 21 �C,and unbound glycine was removed by washing the beads
with 20 mL of coupling buffer. The resulting affinity
adsorbant was washed with 30 mL of 0.01 m Tris ⁄HCl,
pH 8, containing 0.14 m NaCl and 0.025% NaN3, and
stored at 4 �C until use (maximum 2 days).
Protein determination
Protein concentration was determined by the Bradford
method [21].
Acknowledgements
We thank Dr J. A. Curtino for critical reading of
the manuscript, Dr R. Serrano (Valencia, Spain) for
the generous gift of anti-(H+-ATPase), and Dr S. An-
derson for editing. This work was supported by
grants from Agencia Nacional de Promocion Cientıfi-
ca y Tecnologica de la Secretarıa de Ciencia y Tec-
nologıa del Ministerio de Cultura y Educacion en el
marco del Programa de Modernizacion Tecnologica
(BID 802 OC ⁄AR), Fundacion Antorchas, Secretarıa
de Ciencia y Tecnica de la Universidad Nacional de
Cordoba, Secretarıa de Ciencias de la Universidad
Nacional de Rıo Cuarto, y Agencia Cordoba Ciencia
del Gobierno de la Provincia de Cordoba,
Argentina.
References
1 Alonso AC, Nunez-Fernandez M, Beltramo DM,
Casale CH & Barra HS (1998) Na+,K+-ATPase was
found to be the membrane component responsible for
the hydrophobic behaviour of the brain membrane tub-
ulin. Biochem Biophys Res Commun 253, 824–927 [note:
erratum published in Biochem. Biophys Res Commun
257 (1999), 642].
2 Casale CH, Alonso A del C & Barra HS (2001) Brain
plasma membrane Na+,K+-ATPase is inhibited by
acetylated tubulin. Mol Cell Biochem 216, 85–92.
3 Casale CH, Previtali G & Barra HS (2003) Involvement of
acetylated tubulin in the regulation of Na+,K+-ATPase
activity in cultured astrocytes. FEBS Lett 534, 115–118.
4 Casale CH, Previtali G, Serafino JJ, Arce CA & Barra
HS (2005) Regulation of acetylated tubulin ⁄Na+,K+-
ATPase interaction by l-glutamate in non-neural cells:
involvement of microtubules. Biochim Biophys Acta
1721, 185–192.
5 Serrano R (1983) In vivo glucose activation of the yeast
plasma membrane ATPase. FEBS Lett 156, 11–14.
6 Malpartida F & Serrano R (1981) Phosphorylated inter-
mediate of the ATPase from the plasma membrane of
yeast. Eur J Biochem 116, 413–417.
7 Capieaux E, Vignais M-L, Sentenac A & Goffeau A
(1989) The yeast H+-ATPase gene is controlled by the
promoter binding factor TUF. J Biol Chem 264, 7437–
7446.
8 Eraso P & Portillo F (1994) Molecular mechanism of
regulation of yeast plasma membrane H(+)-ATPase
by glucose. Interaction between domains and identifi-
cation of new regulatory sites. J Biol Chem 269,
10393–10399.
9 Garcia-Arranz M, Maldonado AM, Mazon MJ &
Portillo F (1994) Transcriptional control of yeast
plasma membrane H(+)-ATPase by glucose. Cloning
and characterization of a new gene involved in this
regulation. J Biol Chem 269, 18076–18082.
10 Souza MAA, Tropia MJ & Brandao RL (2001) New
aspects of the glucose activation of the H+-ATPase in
the yeast Saccharomyces cerevisiae. Microbiology 147,
2849–2855.
11 de la Fuente N, Maldonado AM & Portillo F (1997)
Glucose activation of the plasma membrane H+-ATP
ase requires the ubiquitin-proteosome proteolytic path-
way. FEBS Lett 411, 308–312.
12 Coons DM, Boulton RB & Bisson LF (1995) Compu-
ter-assisted nonlinear regression analysis of the multi-
component glucose uptake kinetics. J Bacteriol 177,
3251–3258.
13 Nunez-Fernandez M, Beltramo DM, Alonso A del C
& Barra HS (1997) Conversion of hydrophilic tubulin
into a hydrophobic compound. Evidence for the invol-
vement of membrane proteins. Mol Cell Biochem 170,
91–98.
14 Beltramo DM, Nunez M, Alonso A del C & Barra HS
(1994) The relationship of hydrophobic tubulin with
membranes in neural tissue. Mol Cell Biochem 141,
57–63.
15 Goossens A, de La Fuente N, Forment J, Serrano R &
Portillo F (2000) Regulation of yeast H+-ATPase by
protein kinases belonging to a family dedicated to acti-
vation of plasma membrane transporters. Mol Cell Biol
20, 7654–7661.
16 Villalba JM, Palmgren MG, Berberian GE, Ferguson C
& Serrano R (1992) Functional expression of plant
plasma membrane H(+)-ATPase in yeast endoplasmic
reticulum. J Biol Chem 267, 12341–12349.
17 Laemmli UK (1970) Cleavage of structural proteins
during the assembly of the head of bacteriophage T4.
Nature 227, 680–685.
18 Piperno G & Fuller MT (1985) Monoclonal antibodies
specific for an acetylated form of alpha-tubulin recog-
nize the antigen in cilia and flagella from a variety of
organisms. J Cell Biol 101, 2085–2094.
A. N. Campetelli et al. Dissociation of tubulin–H+-ATPase complex by glucose
FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS 5751

19 DeWitt ND, dos Santos CF, Allen KE & Slayman CW
(1998) Phosphorylation region of the yeast plasma-
membrane H+-ATPase. Role in protein folding and
biogenesis. J Biol Chem 273, 21744–21751.
20 Hubert JJ, Schenk DB, Skelly H & Leffert HL (1986) Rat
hepatic (Na+,K+)-ATPase: alpha-subunit isolation by
immunoaffinity chromatography and structural analysis
by peptide mapping. Biochemistry 5, 4156–4163.
21 Bradford MM (1976) A rapid and sensitive method for
the quantitation of microgram quantities of protein util-
izing the principle of protein-dye binding. Anal Biochem
72, 248–254.
5752 FEBS Journal 272 (2005) 5742–5752 ª 2005 FEBS
Dissociation of tubulin–H+-ATPase complex by glucose A. N. Campetelli et al.




















