
Vacuolar Pathology in the Median Eminence of the Hypothalamus After Hyponatremia
Upload
independentCategory
view
0download
0
A Common Genetic Variant in the 3�-UTR of VacuolarH�-ATPase ATP6V0A1 Creates a Micro-RNA Motif to Alter
Chromogranin A Processing and Hypertension RiskZhiyun Wei, BS; Nilima Biswas, PhD; Lei Wang, PhD; Maite Courel, PhD; Kuixing Zhang, MD, PhD;
Alex Soler-Jover, PhD; Laurent Taupenot, PhD; Daniel T. O’Connor, MD
Background—The catecholamine release-inhibitor catestatin and its precursor chromogranin A (CHGA) may constitute“intermediate phenotypes” in the analysis of genetic risk for cardiovascular disease such as hypertension. Previously,the vacuolar H�-ATPase subunit gene ATP6V0A1 was found within the confidence interval for linkage with catestatinsecretion in a genome-wide study, and its 3�-UTR polymorphism T�3246C (rs938671) was associated with bothcatestatin processing from CHGA and population blood pressure. We explored the molecular mechanism of this effectby experiments with transfected chimeric photoproteins in chromaffin cells.
Methods and Results—Placing the ATP6V0A1 3�-UTR downstream of a luciferase reporter, we found that the C (variant)allele decreased overall gene expression. The 3�-UTR effect was verified by coupled in vitro transcription/translationof the entire/intact human ATP6V0A1 mRNA. Chromaffin granule pH, monitored by fluorescence of CHGA/EGFPchimera during vesicular H�-ATPase inhibition by bafilomycin A1, was more easily perturbed during coexpression ofthe ATP6V0A1 3�-UTR C-allele than the T-allele. After bafilomycin A1 treatment, the ratio of CHGA precursor to itscatestatin fragments in PC12 cells was substantially diminished, though the qualitative composition of such fragmentswas not affected (on immunoblot or matrix-assisted laser desorption ionization (MALDI) mass spectrometry).Bafilomycin A1 treatment also decreased exocytotic secretion from the regulated pathway, monitored by a CHGAchimera tagged with embryonic alkaline phosphatase. 3�-UTR T�3246C created a binding motif for micro-RNAhsa-miR-637; cotransfection of hsa-miR-637 precursor or antagomir/inhibitor oligonucleotides yielded the predictedchanges in expression of luciferase reporter/ATP6V0A1-3�-UTR plasmids varying at T�3246C.
Conclusions—The results suggest a series of events whereby ATP6V0A1 3�-UTR variant T�3246C functioned:ATP6V0A1 expression probably was affected through differential micro-RNA effects, altering vacuolar pH andconsequently CHGA processing and exocytotic secretion. (Circ Cardiovasc Genet. 2011;4:381-389.)
Key Words: chromaffin � catecholamine � adrenal � hypertension � vacuolar pH
Chromogranin A (CHGA), a member of the chromogra-nin/secretogranin family of neuroendocrine secretory
proteins, is the precursor to several bioactive peptides includ-ing the catecholamine release-inhibitory catestatin (humanCHGA352–372).1,2 Catestatin secretion may be an “intermedi-ate phenotype” in the analysis of genetic risk for cardiovas-cular disease3 because it is diminished not only in hyperten-sive individuals but also in the still-normotensive offspring ofpatients with hypertension,4 and replacement of catestatin can“rescue” the severe hypertension observed in mice withtargeted ablation of the Chga gene.5 The plasma ratio ofCHGA/catestatin (precursor/product) was significantlyhigher in a hypertensive population, suggesting an impair-ment of CHGA processing in this disorder.6
Editorial see p 337Clinical Perspective on p 389
Previously, we studied catestatin secretion in a large series oftwin and sibling pairs from North America and Australia, enablingestimation of its heritability, genome-wide linkage (positional clon-ing), and marker-on-trait association.6 We found that theATP6V0A1 gene (ATP6N1, ATP6N1A, VPP1; OMIM No. 192130;human chromosome 17q21) was a positional candidate locus withinthe confidence interval for catestatin linkage. Furthermore, allelicvariation at ATP6V0A1 (3�-UTR single nucleotide polymorphism(SNP) T�3246C; rs938671; minor allele frequency, 9% to 13%) isassociated with catestatin concentration, the CHGA/catestatin ratio,and basal blood pressure (BP) in the population.6
Received August 23, 2010; accepted April 26, 2011.From the Departments of Medicine and Pharmacology, Institute for Genomic Medicine, University of California at San Diego, the VA San Diego
Healthcare System, La Jolla, CA.Zhiyun Wei is a visiting scholar from Bio-X Institutes, Shanghai Jiao Tong University, China.The online-only Data Supplement is available at http://circgenetics.ahajournals.org/cgi/content/full/CIRCGENETICS.111.959767/DC1.Correspondence to Daniel T. O’Connor, MD, Department of Medicine (0838), UCSD School of Medicine and VASDHS, 9500 Gilman Dr, La Jolla,
CA 92093-0838. E-mail [email protected]© 2011 American Heart Association, Inc.
Circ Cardiovasc Genet is available at http://circgenetics.ahajournals.org DOI: 10.1161/CIRCGENETICS.111.959767
381

ATP6V0A1 (NC_000017), initially isolated in 1995,7 en-codes the �1 subunit of the vacuolar (V) H�-translocatingATPase heteromultimeric complex, which mediates acidifi-cation of eukaryotic intracellular organelles; the �1 subunit isa 116-kDa integral membrane protein that participates di-rectly in H� translocation. The pH of organelles along thesecretory pathway decreases progressively from the endo-plasmic reticulum to the secretory granule,8–11 and chemical(bafilomycin A1) inhibition of the vacuolar H�-ATPaseimpairs chromaffin granule formation and catecholaminestorage and secretory protein trafficking into the regulatedpathway.12
In the present study, we explore the molecular mechanismwhereby ATP6V0A1 T�3246C influences CHGA and cates-tatin concentrations and their ratios. It was plausible tohypothesize that changes in control of vacuolar pH wouldinfluence such traits through the effect of the secretorypathway pH to modulate either precursor proteolytic process-ing or exocytotic secretion.13 Through the use of transfectedchimeric photoprotein reporters in chromaffin cells, ourresults reveal that ATP6V0A1 3�-UTR variant T�3246Calters gene expression through differential binding to aparticular micro-RNA, thereby altering vacuolar pH andconsequently the processing of CHGA to catestatin.
MethodsSee the online-only Data Supplement Methods section for construc-tion of human expression plasmids and cell culture and transfection.
Luciferase Reporter Activity AssayAfter transfection and cell growth over an 8- to 24-hour time course,cells were lysed with passive lysis buffer (Promega) for sequentialmeasurement of luciferase enzymatic activity and total proteinconcentration. Luciferase enzymatic activity was measured using theLuciferase Assay System (Promega) on a Luminometer Autolumat953 (EG&G Berthold, Bad Wildbad, Germany). Total proteinconcentration was measured using a dye-binding protein assay(Bio-Rad) on a SmartSpec Plus spectrophotometer (Bio-Rad). Re-sults are expressed as the ratio of luciferase activity/protein concen-tration to normalize luciferase activity.
In Vitro Transcription/Translation ofHuman ATP6V0A1SP6 promoter–driven, full-length human ATP6V0A1 cDNA (includ-ing the 3�-UTR in 2 versions, T�3246C) was transcribed/translatedin vitro by use of the TNT Quick Coupled Transcription/TranslationSystem (Promega L5080), using a rabbit reticulocyte lysate. Newlysynthesized proteins were labeled by incorporation of biotinylated-lysine and then detected by chemiluminescence with streptavidin–horseradish peroxidase in the Transcend Non-Radioactive Transla-tion Detection System (Promega).
Measurement of Intravacuolar (CatecholamineStorage Vesicle) pHWe previously described the use of transfected/expressed humanCHGA/EGFP chimeras and their trafficking to chromaffin granules12
as well as the use of an SCG2/EGFP chimera to monitor pH withinchromaffin granules.12 We used a CHGA/EGFP chimera to monitorpH changes within living rat pheochromocytoma cell line (PC12)cells during coexpression of human ATP6V0A1 with alternative3�-UTR alleles. PC12 cells grown on poly-L-lysine–coated andcollagen-coated 25-mm round glass coverslips (Warner Instruments)were transfected with pCMV-CHGA-EGFP and transferred into aperfusion chamber (Quick Exchange, Warner Instruments). Moni-
toring of single-cell fluorescence was achieved with an ImageMasterIM-2103 to 6-HQ imaging system (Photon Technology Interna-tional). Correlation between GFP fluorescence and pH value wasobtained in situ by incubating the cells for 3 minutes in KCl-richmedia buffered to pH values ranging from 4.5 to 8.5 (at a 0.5 pH unitinterval). In cells cotransfected with CHGA/EGFP and full-lengthATP6V0A1 (with 3�-UTR), pH was followed with GFP fluorescenceat baseline and after bafilomycin was added to a 100 nmol/L finalconcentration. A detailed protocol was described previously.14
Chemiluminescence Detection of SecretoryProtein/Embryonic Alkaline Phosphatase ChimeraSecretion by Chromaffin CellsDetection of EAP enzymatic activity release from CHGA–embry-onic alkaline phosphatase (EAP) chimera-expressing PC12 cells wasachieved using the chemiluminescent substrate 3-(4-methoxyspiro[1,2-dioxetane-3,2�-(5�chloro)tricyclo[3.3.1.1(3,7)]decan]-4-yl) phe-nyl phosphate (CSPD) (Phospha-Light, Applied Biosystems, Fostercity, California) on a Luminometer Autolumat 953 (EG&GBerthold). EAP activities are measured in the culture supernatant andthe cell lysate. Exocytotic secretion of the transfected/expressed EAPchimeras was provoked by the potent regulated pathway stimulusBa2� (2 mmol/L), which acts by blocking efflux of cellular K� andhence cell membrane repolarization.15 A detailed protocol wasdescribed previously.15 The secretion rate is calculated as superna-tant EAP activity as a percentage of total enzymatic activity (cellplus supernatant). The “sorting index” is calculated as a function ofincrease in secretion rate after stimulation of the regulated exocytoticpathway by Ba2�: (stimulated minus basal)/basal.
Immunoblot AnalysisProteins from PC12 cell lysates were separated in a 10% SDS-PAGE(Novex precast gel; Invitrogen) gel and electrophoretically trans-ferred to nitrocellulose membranes (Protran, BA85; Whatman Inc,Florham Park, NJ). The membrane was blocked with 5% (wt/vol)powdered/dry milk in Tris-buffered saline with 0.1% Tween-20.After incubation with primary antibody (rabbit anti-rat catestatin),the membrane was washed and incubated with secondary antibody(horseradish peroxidase–conjugated donkey anti-rabbit). The mem-brane was then developed by the Supersignal West pico-chemiluminescent substrate (Pierce, Rockford, IL). Antiactin wasused as internal control. Immunoreactive band quantification wasdone on the software Quantity One (Bio-Rad).
Matrix Assisted Laser Desorption Ionization-TimeOf Flight Mass SpectrometryPC12 cells were treated with vehicle (DMSO) or bafilomycin A1(100 nmol/L; from Streptomyces griseus; Sigma B1793), which is aspecific inhibitor of the vacuolar-type ATPase (V-ATPase)16 andwas shown to inhibit vacuolar acidification17 as well as dense coregranule formation and secretory protein trafficking12 effectively.After exposure for 22 hours, cells were lysed, precleared with normalrabbit serum, and then subjected to immunoprecipitation with anti-rat catestatin antibody. Antigen-antibody complexes were thenisolated using protein G plus/protein A agarose; the complexes werewashed several times, and the bound peptides were eluted withacetonitrile/water/trifluoroacetic acid. Eluted peptides were concen-trated by lyophilization and subjected to matrix-assisted laser de-sorption ionization-time of flight (MALDI-TOF) analysis: reflectronmode to scan the 1000 to 2100 Da range and linear mode to scan the2000 to 25 000 Da range, on a Voyager De STR MALDI-TOF massspectrometer (Applied Biosystems) at a proteomics core facility(http://massspec.ucsd.edu/bioms/). Resulting peptide masses wereanalyzed in the program Protein Prospector (http://prospector.ucsf.edu/) to identify the fragments of rat Chga.
Evaluation of Micro-RNA Function
ComputationalInterspecies sequence alignments were done by Clustal-W. 3�-UTRmicro-RNA (miRNA) motifs were predicted at RegRNA (http://
382 Circ Cardiovasc Genet August 2011
regrna.mbc.nctu.edu.tw/). The effects of the variant on RNA hybridstructures and predicted minimum folding energies were analyzedusing BiBiServ (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/submission.html).
ExperimentalStrategies of both overexpression and knockdown were adopted toexplore miRNA function in cella. For overexpression, Ambion(Austin, TX) PremiR 24-mer specific precursor for miRNA humanmicro-RNA 637 (hsa-miR-637; PM11545) was used to increase thatmiRNA signal, and a predesigned negative control precursor (No. 1;AM17110) was used to minimize off-target effects. For knockdown,an Exiqon (Vedbaek, Denmark) miRCURY LNA (“locked,” stabi-lized nucleic acid) 18-mer inhibitor for the specific miRNA hsa-miR-637 (412259-00) was used in an attempt to decrease the signal fromthat miRNA, and a predesigned 22-mer negative control inhibitor(“Scramble-miR”; Exiqon 199002-00) was used in the control group.The identity of each synthetic oligonucleotide was verified bymass spectrometry.
StatisticsValues are given as mean�SEM. Numbers of experimental repli-cates are given in the figure legends. Data shown are representativeof typical experiments. Statistical analyses were performed withSPSS-17 (SPSS Inc, Chicago, IL) by t test or 2-way ANOVA, asappropriate, after inspection of the data distribution. Differenceswere considered significant at P�0.05.
ResultsATP6V0A1 3�-UTR Variant T�3246C InfluencesGene Expression
Evidence From Isolated 3�-UTR Segments on Transfected3�-UTR/Luciferase Reporter PlasmidsWild-type (T�3246) versus variant (�3246C) 1458 bpATP6V0A1 3�-UTRs were ligated into the reporter plasmid
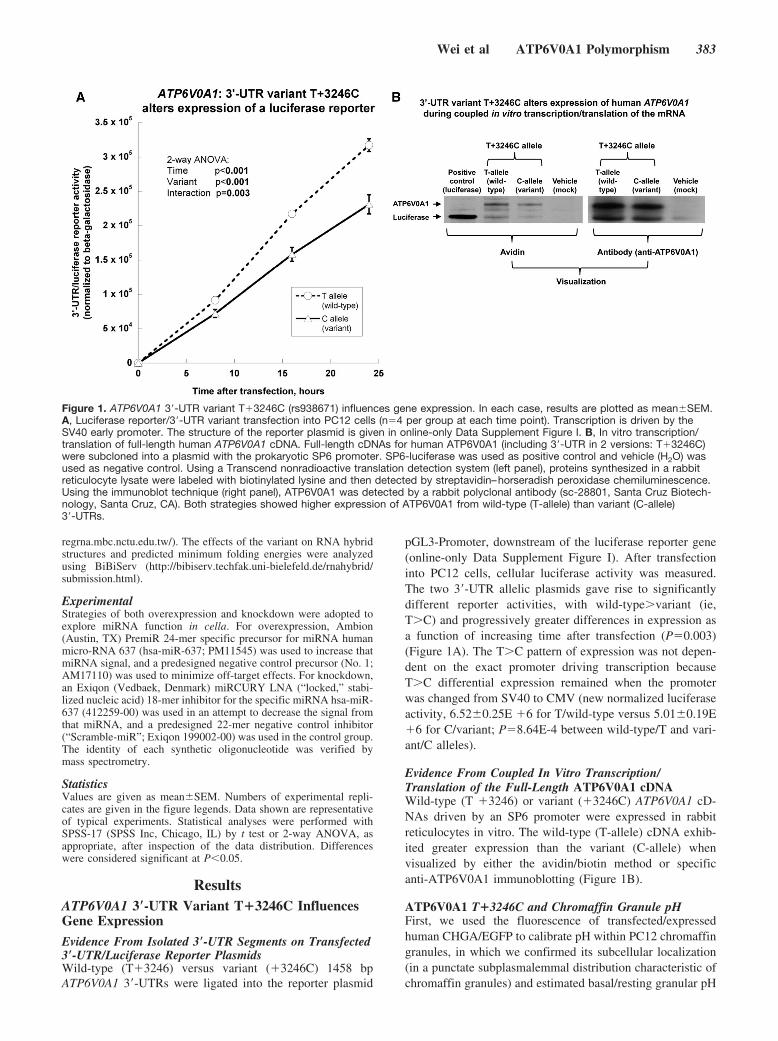
pGL3-Promoter, downstream of the luciferase reporter gene(online-only Data Supplement Figure I). After transfectioninto PC12 cells, cellular luciferase activity was measured.The two 3�-UTR allelic plasmids gave rise to significantlydifferent reporter activities, with wild-type�variant (ie,T�C) and progressively greater differences in expression asa function of increasing time after transfection (P�0.003)(Figure 1A). The T�C pattern of expression was not depen-dent on the exact promoter driving transcription becauseT�C differential expression remained when the promoterwas changed from SV40 to CMV (new normalized luciferaseactivity, 6.52�0.25E �6 for T/wild-type versus 5.01�0.19E�6 for C/variant; P�8.64E-4 between wild-type/T and vari-ant/C alleles).
Evidence From Coupled In Vitro Transcription/Translation of the Full-Length ATP6V0A1 cDNAWild-type (T �3246) or variant (�3246C) ATP6V0A1 cD-NAs driven by an SP6 promoter were expressed in rabbitreticulocytes in vitro. The wild-type (T-allele) cDNA exhib-ited greater expression than the variant (C-allele) whenvisualized by either the avidin/biotin method or specificanti-ATP6V0A1 immunoblotting (Figure 1B).
ATP6V0A1 T�3246C and Chromaffin Granule pHFirst, we used the fluorescence of transfected/expressedhuman CHGA/EGFP to calibrate pH within PC12 chromaffingranules, in which we confirmed its subcellular localization(in a punctate subplasmalemmal distribution characteristic ofchromaffin granules) and estimated basal/resting granular pH
Figure 1. ATP6V0A1 3�-UTR variant T�3246C (rs938671) influences gene expression. In each case, results are plotted as mean�SEM.A, Luciferase reporter/3�-UTR variant transfection into PC12 cells (n�4 per group at each time point). Transcription is driven by theSV40 early promoter. The structure of the reporter plasmid is given in online-only Data Supplement Figure I. B, In vitro transcription/translation of full-length human ATP6V0A1 cDNA. Full-length cDNAs for human ATP6V0A1 (including 3�-UTR in 2 versions: T�3246C)were subcloned into a plasmid with the prokaryotic SP6 promoter. SP6-luciferase was used as positive control and vehicle (H2O) wasused as negative control. Using a Transcend nonradioactive translation detection system (left panel), proteins synthesized in a rabbitreticulocyte lysate were labeled with biotinylated lysine and then detected by streptavidin–horseradish peroxidase chemiluminescence.Using the immunoblot technique (right panel), ATP6V0A1 was detected by a rabbit polyclonal antibody (sc-28801, Santa Cruz Biotech-nology, Santa Cruz, CA). Both strategies showed higher expression of ATP6V0A1 from wild-type (T-allele) than variant (C-allele)3�-UTRs.
Wei et al ATP6V0A1 Polymorphism 383
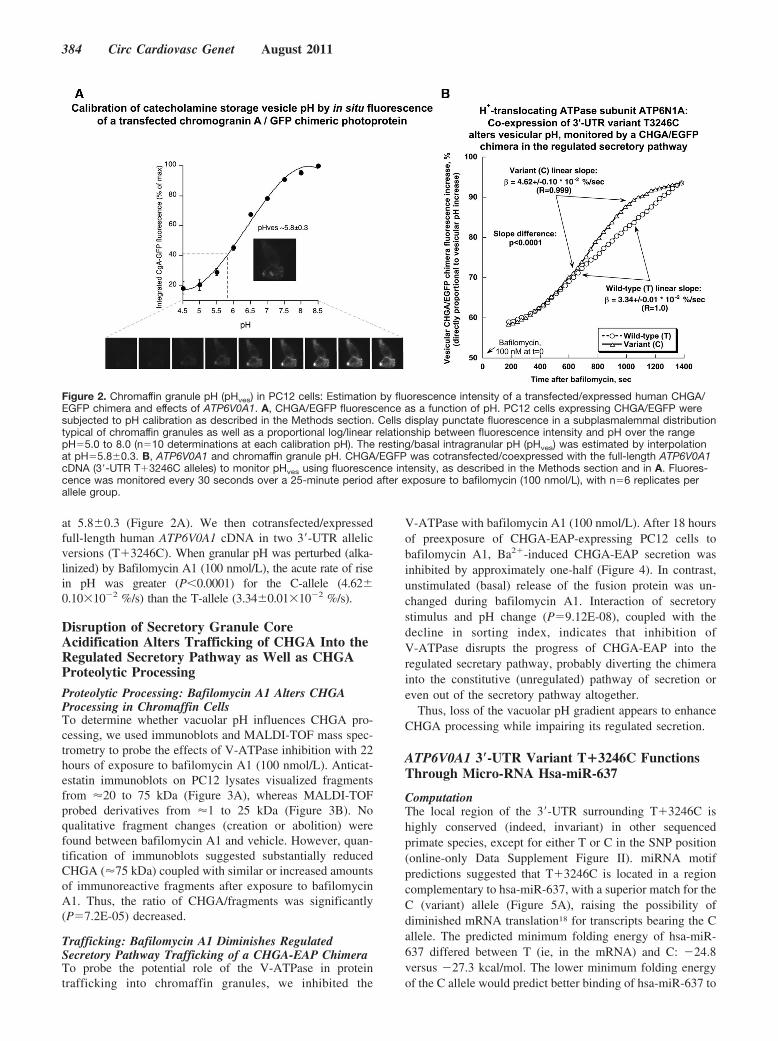
at 5.8�0.3 (Figure 2A). We then cotransfected/expressedfull-length human ATP6V0A1 cDNA in two 3�-UTR allelicversions (T�3246C). When granular pH was perturbed (alka-linized) by Bafilomycin A1 (100 nmol/L), the acute rate of risein pH was greater (P�0.0001) for the C-allele (4.62�0.10�10�2 %/s) than the T-allele (3.34�0.01�10�2 %/s).
Disruption of Secretory Granule CoreAcidification Alters Trafficking of CHGA Into theRegulated Secretory Pathway as Well as CHGAProteolytic Processing
Proteolytic Processing: Bafilomycin A1 Alters CHGAProcessing in Chromaffin CellsTo determine whether vacuolar pH influences CHGA pro-cessing, we used immunoblots and MALDI-TOF mass spec-trometry to probe the effects of V-ATPase inhibition with 22hours of exposure to bafilomycin A1 (100 nmol/L). Anticat-estatin immunoblots on PC12 lysates visualized fragmentsfrom 20 to 75 kDa (Figure 3A), whereas MALDI-TOFprobed derivatives from 1 to 25 kDa (Figure 3B). Noqualitative fragment changes (creation or abolition) werefound between bafilomycin A1 and vehicle. However, quan-tification of immunoblots suggested substantially reducedCHGA (75 kDa) coupled with similar or increased amountsof immunoreactive fragments after exposure to bafilomycinA1. Thus, the ratio of CHGA/fragments was significantly(P�7.2E-05) decreased.
Trafficking: Bafilomycin A1 Diminishes RegulatedSecretory Pathway Trafficking of a CHGA-EAP ChimeraTo probe the potential role of the V-ATPase in proteintrafficking into chromaffin granules, we inhibited the
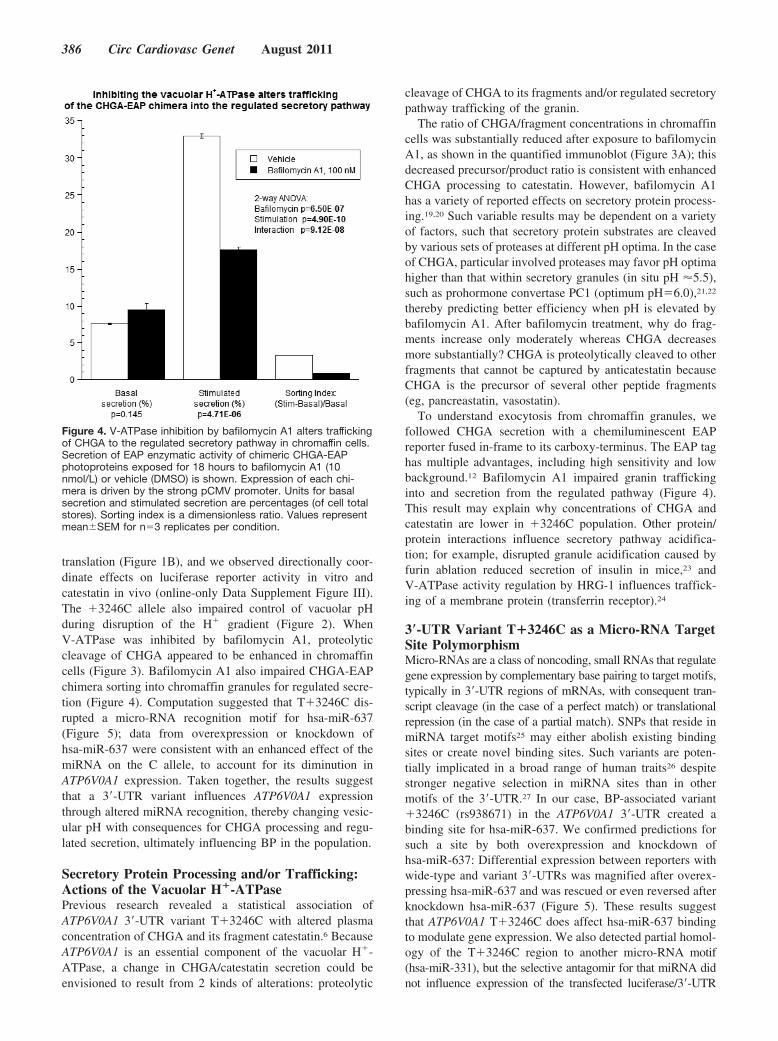
V-ATPase with bafilomycin A1 (100 nmol/L). After 18 hoursof preexposure of CHGA-EAP-expressing PC12 cells tobafilomycin A1, Ba2�-induced CHGA-EAP secretion wasinhibited by approximately one-half (Figure 4). In contrast,unstimulated (basal) release of the fusion protein was un-changed during bafilomycin A1. Interaction of secretorystimulus and pH change (P�9.12E-08), coupled with thedecline in sorting index, indicates that inhibition ofV-ATPase disrupts the progress of CHGA-EAP into theregulated secretary pathway, probably diverting the chimerainto the constitutive (unregulated) pathway of secretion oreven out of the secretory pathway altogether.
Thus, loss of the vacuolar pH gradient appears to enhanceCHGA processing while impairing its regulated secretion.
ATP6V0A1 3�-UTR Variant T�3246C FunctionsThrough Micro-RNA Hsa-miR-637
ComputationThe local region of the 3�-UTR surrounding T�3246C ishighly conserved (indeed, invariant) in other sequencedprimate species, except for either T or C in the SNP position(online-only Data Supplement Figure II). miRNA motifpredictions suggested that T�3246C is located in a regioncomplementary to hsa-miR-637, with a superior match for theC (variant) allele (Figure 5A), raising the possibility ofdiminished mRNA translation18 for transcripts bearing the Callele. The predicted minimum folding energy of hsa-miR-637 differed between T (ie, in the mRNA) and C: �24.8versus �27.3 kcal/mol. The lower minimum folding energyof the C allele would predict better binding of hsa-miR-637 to
Figure 2. Chromaffin granule pH (pHves) in PC12 cells: Estimation by fluorescence intensity of a transfected/expressed human CHGA/EGFP chimera and effects of ATP6V0A1. A, CHGA/EGFP fluorescence as a function of pH. PC12 cells expressing CHGA/EGFP weresubjected to pH calibration as described in the Methods section. Cells display punctate fluorescence in a subplasmalemmal distributiontypical of chromaffin granules as well as a proportional log/linear relationship between fluorescence intensity and pH over the rangepH�5.0 to 8.0 (n�10 determinations at each calibration pH). The resting/basal intragranular pH (pHves) was estimated by interpolationat pH�5.8�0.3. B, ATP6V0A1 and chromaffin granule pH. CHGA/EGFP was cotransfected/coexpressed with the full-length ATP6V0A1cDNA (3�-UTR T�3246C alleles) to monitor pHves using fluorescence intensity, as described in the Methods section and in A. Fluores-cence was monitored every 30 seconds over a 25-minute period after exposure to bafilomycin (100 nmol/L), with n�6 replicates perallele group.
384 Circ Cardiovasc Genet August 2011

the �3246C mRNA and hence more efficient translationalrepression (or perhaps even degradation) of the variant/CmRNA, a finding consistent with decreased reporter expres-sion by the variant/C luciferase/3�-UTR plasmid (Figure 1).
Hsa-miR-637 MimicryTo validate a differential hsa-miR-637 effect on T�3246C, aspecific 24-mer miRNA precursor was cotransfected with theluciferase/3�-UTR plasmid into PC12 cells. Exogenous/cotransfected hsa-miR-637 precursor decreased reporter ex-pression (Figure 5B). The expression difference between Tand C was maintained after additional/exogenous hsa-miR-637, and the % difference between 2 genotypes (T3C) wasamplified appreciably (32.8% in the negative controls versus41.6% during hsa-miR-637 treatment). By 2-way ANOVA,P�0.001 for variant, P�1.45E-05 for precursor, andP�0.757 for interaction.
Hsa-miR-637 AntagonismInhibition of hsa-miR-637 by a specific “antagomir” in-creased reporter expression (Figure 5C). In the baseline state(negative control group), the expression pattern was C�T,
consistent with the initial transfection experiments (Figure 1).After knockdown of hsa-miR-637, however, this differencewas reversed to C�T.
Both enhancement and inhibition results for hsa-miR-637were consistent with the hypothesis that the C�T expressionpattern occurs because of higher affinity of the C allele forhsa-miR-637, with consequent translational repression. By2-way ANOVA, P�6.67E-04 for variant, P�3.07E-13 forinhibitor, and P�4.25E-06 for interaction.
Discussion
OverviewIn the present study, we aimed to clarify the mechanismwhereby 3�-UTR (T�3246C; rs938671) genetic variation atthe vacuolar H�-ATPase subunit ATP6V0A1 influencesCHGA/catestatin secretion and consequently BP in the pop-ulation.6 We began by isolating the ATP6V0A1 3�-UTR ontoa luciferase reporter gene, revealing that �3246C decreasedreporter expression (Figure 1A); the effects on gene expres-sion were documented by coupled in vitro transcription/
Figure 3. CHGA proteolytic processing to fragments during inhibition of the vacuolar pH gradient in chromaffin cells. A, Immunoblotafter SDS-PAGE. Rat pheochromocytoma PC12 cells (n�3 replicate cultures) were treated with bafilomycin A1 (100 nmol/L) or vehicle(DMSO) for 22 hours, then lysed, subjected to SDS-PAGE, and immunoblotted with anti-rat catestatin antibody (top and middle panels).The upper panel (Mr 25 to 75 kDa) was exposed for 30 seconds. The middle panel (relative molecular mass [Mr] 15 to 37 kDa) shows adarker exposure (45 minutes) of the lower part of the same immunoblot. A parallel blot was stained with antiactin antibody (bottompanel) to indicate comparable protein load/lane. B, Mass spectrometry. Cell lysates from the vehicle and bafilomycin A1–treated cellswere subjected to immunoprecipitation with anti-rat catestatin antibody followed by MALDI-TOF analysis. Calculated mass (M/Z) valuesfor fragments of the parent molecule (rat Chga) are listed in the table.
Wei et al ATP6V0A1 Polymorphism 385
translation (Figure 1B), and we observed directionally coor-dinate effects on luciferase reporter activity in vitro andcatestatin in vivo (online-only Data Supplement Figure III).The �3246C allele also impaired control of vacuolar pHduring disruption of the H� gradient (Figure 2). WhenV-ATPase was inhibited by bafilomycin A1, proteolyticcleavage of CHGA appeared to be enhanced in chromaffincells (Figure 3). Bafilomycin A1 also impaired CHGA-EAPchimera sorting into chromaffin granules for regulated secre-tion (Figure 4). Computation suggested that T�3246C dis-rupted a micro-RNA recognition motif for hsa-miR-637(Figure 5); data from overexpression or knockdown ofhsa-miR-637 were consistent with an enhanced effect of themiRNA on the C allele, to account for its diminution inATP6V0A1 expression. Taken together, the results suggestthat a 3�-UTR variant influences ATP6V0A1 expressionthrough altered miRNA recognition, thereby changing vesic-ular pH with consequences for CHGA processing and regu-lated secretion, ultimately influencing BP in the population.
Secretory Protein Processing and/or Trafficking:Actions of the Vacuolar H�-ATPasePrevious research revealed a statistical association ofATP6V0A1 3�-UTR variant T�3246C with altered plasmaconcentration of CHGA and its fragment catestatin.6 BecauseATP6V0A1 is an essential component of the vacuolar H�-ATPase, a change in CHGA/catestatin secretion could beenvisioned to result from 2 kinds of alterations: proteolytic
cleavage of CHGA to its fragments and/or regulated secretorypathway trafficking of the granin.
The ratio of CHGA/fragment concentrations in chromaffincells was substantially reduced after exposure to bafilomycinA1, as shown in the quantified immunoblot (Figure 3A); thisdecreased precursor/product ratio is consistent with enhancedCHGA processing to catestatin. However, bafilomycin A1has a variety of reported effects on secretory protein process-ing.19,20 Such variable results may be dependent on a varietyof factors, such that secretory protein substrates are cleavedby various sets of proteases at different pH optima. In the caseof CHGA, particular involved proteases may favor pH optimahigher than that within secretory granules (in situ pH 5.5),such as prohormone convertase PC1 (optimum pH�6.0),21,22
thereby predicting better efficiency when pH is elevated bybafilomycin A1. After bafilomycin treatment, why do frag-ments increase only moderately whereas CHGA decreasesmore substantially? CHGA is proteolytically cleaved to otherfragments that cannot be captured by anticatestatin becauseCHGA is the precursor of several other peptide fragments(eg, pancreastatin, vasostatin).
To understand exocytosis from chromaffin granules, wefollowed CHGA secretion with a chemiluminescent EAPreporter fused in-frame to its carboxy-terminus. The EAP taghas multiple advantages, including high sensitivity and lowbackground.12 Bafilomycin A1 impaired granin traffickinginto and secretion from the regulated pathway (Figure 4).This result may explain why concentrations of CHGA andcatestatin are lower in �3246C population. Other protein/protein interactions influence secretory pathway acidifica-tion; for example, disrupted granule acidification caused byfurin ablation reduced secretion of insulin in mice,23 andV-ATPase activity regulation by HRG-1 influences traffick-ing of a membrane protein (transferrin receptor).24
3�-UTR Variant T�3246C as a Micro-RNA TargetSite PolymorphismMicro-RNAs are a class of noncoding, small RNAs that regulategene expression by complementary base pairing to target motifs,typically in 3�-UTR regions of mRNAs, with consequent tran-script cleavage (in the case of a perfect match) or translationalrepression (in the case of a partial match). SNPs that reside inmiRNA target motifs25 may either abolish existing bindingsites or create novel binding sites. Such variants are poten-tially implicated in a broad range of human traits26 despitestronger negative selection in miRNA sites than in othermotifs of the 3�-UTR.27 In our case, BP-associated variant�3246C (rs938671) in the ATP6V0A1 3�-UTR created abinding site for hsa-miR-637. We confirmed predictions forsuch a site by both overexpression and knockdown ofhsa-miR-637: Differential expression between reporters withwide-type and variant 3�-UTRs was magnified after overex-pressing hsa-miR-637 and was rescued or even reversed afterknockdown hsa-miR-637 (Figure 5). These results suggestthat ATP6V0A1 T�3246C does affect hsa-miR-637 bindingto modulate gene expression. We also detected partial homol-ogy of the T�3246C region to another micro-RNA motif(hsa-miR-331), but the selective antagomir for that miRNA didnot influence expression of the transfected luciferase/3�-UTR
Figure 4. V-ATPase inhibition by bafilomycin A1 alters traffickingof CHGA to the regulated secretory pathway in chromaffin cells.Secretion of EAP enzymatic activity of chimeric CHGA-EAPphotoproteins exposed for 18 hours to bafilomycin A1 (10nmol/L) or vehicle (DMSO) is shown. Expression of each chi-mera is driven by the strong pCMV promoter. Units for basalsecretion and stimulated secretion are percentages (of cell totalstores). Sorting index is a dimensionless ratio. Values representmean�SEM for n�3 replicates per condition.
386 Circ Cardiovasc Genet August 2011

reporter plasmids (data not shown). Inspection of NCBI GEOdata sets reveals abundant endogenous ATP6V0A1 expres-sion in the adrenal gland (GDS1464)28 as well as clonalchromaffin cells (GDS2555).29
Vacuolar H�-ATPase FunctionsThe vacuolar H�-ATPase (V-ATPase) is a multisubunitcomplex that has important roles in the acidification of avariety of several intracellular compartments as well asextracellular milieus. The ATP6V0A1 that we investigate inthis work encodes the �1 isoform of the “�” subunit in the V0(H� translocation, integral membrane) domain. There are 4known isoforms of the “�” subunit (�1 through �4) that havedifferential cellular localizations.30 Even the �1 subunit itselfmay have several alternative splice forms that are targeted todifferent membrane compartments of the cell.31 Such diver-sification may lend far-ranging functions to the H�-ATPase.
In Drosophila, a functional role for the V-ATPase V0domain �1 subunit occurs in late stages of synaptic vesicleexocytosis32; mutation in this subunit did not affect neu-rotransmitter content but impaired evoked synaptic transmis-sion. In mice, the �3 isoform may have a regulatory functionin exocytotic secretion of insulin but not insulin processing.33
In addition to secretion, the V-ATPase participates in proteindegradation and membrane fusion.34,35 Thus the vacuolarH�-ATPase is involved in a variety of physiological andanatomic systems,36 including the heart,37 kidney,38 andskeletal muscle.39
Advantages and Limitations of The StudyWe undertook functional studies at a positional candidategenetic locus on chromosome 17q, ATP6V0A1 to documentbiological mechanisms underlying the statistical genetic as-sociation of marker and trait. Thus, ATP6V0A1 appears to bea trans-QTL (quantitative trait locus) for CHGA/catestatinsecretion and BP, whereas T�3246C (rs938671) may repre-sent a trans-QTN (Quantitative Trait Nucleotide) at thatlocus. The use of luciferase and EAP as reporter genesallowed us to quantify expression and secretion and sodocument effects of the variant as well as the transactingfactor hsa-miR-637, thereby permitting quantitative verifica-tion to augment the plausibility of our conclusions.
Although T�3246C/rs938671 is a human variant, thefunctional studies conducted were performed in rodent (ratPC12 cell) pheochromocytoma cells because the PC12 line iswell established for studies of secretory protein traffic.15
Figure 5. 3�-UTR variant T�3246C influences miRNA hsa-miR-637 regulation of ATP6V0A1 gene expression. A, Computation. The cal-culated miRNA:mRNA interaction is shown, with the SNP highlighted. B and C, Luciferase reporter activity of luciferase/ATP6V0A13�-UTR plasmids cotransfected with hsa-miR-637 precursor (B), inhibitor (C), or their negative control; values represent mean�SEM inn�6 replicates for each condition.
Wei et al ATP6V0A1 Polymorphism 387

Human MIR637, the gene encoding hsa-miR-637, is locatedon human chromosome 19p in the fifth intron of DAPK3.Although miR-637 is not yet well characterized in the rat, thehomologous intronic region of rat Dapk3 displays 82%sequence identity with human miR637. Intriguingly, ratDapk3 is also within a QTL region for BP.40
Catecholamine storage depends on the vacuolar H�-ATPase,12 but our transient transfection studies typicallytarget only a minor percentage of plated cells and thus did notpermit a more global analysis of catecholamine metabolism.Finally, we did not examine the effect of other geneticvariants at the positional candidate locus, ATP6V0A1, al-though T�3246C (rs938671) was selected for study becauseit is the only common (minor allele frequency �5%) variantobserved thus far in the transcript region (exons) of Cauca-sians (CEU), the source population for the original linkageand association result.6 Finally, future studies with appropri-ate animal models may assist in confirming the H�-ATPasemechanism in vivo.
Conclusions and PerspectivesThe ATP6V0A1 3�-UTR common polymorphism T�3246C(rs938671) is associated with CHGA/catestatin secretion andsystemic BP in the population. We explored precisely howsuch genetic variation affected sympathochromaffin exocyto-sis. As shown in online-only Data Supplement Figure IV, ourresults support the viewpoint that T�3246C is located in abinding motif for micro-RNA hsa-miR-637, at which the Callele may impair translation of the ATP6V0A1 mRNA.Because ATP6V0A1 is a necessary subunit of the vacuolarH�-ATPase, its reduced expression may thus impair thephysiological acidification of intracellular organelles such aschromaffin granules. Consequently, the processing of CHGAmay be enhanced and the secretion of proteins/peptides suchas CHGA/catestatin may be impaired. The results shed newlight on the role of chromaffin granule acidification inprocessing and trafficking of secretory peptides/proteins,pointing to new molecular strategies for probing autonomiccontrol of the circulation and ultimately the susceptibility toand pathogenesis of cardiovascular disease states such ashypertension.
We explored molecular mechanisms of how a polymor-phism (T�3246C) in the 3�-UTR of the proton translocatingATPase (V-ATPase) subunit ATP6V0A1 influences humancardiovascular traits, including chromogranin A (CHGA) andBP. First, with the use of a luciferase reporter plasmid, wefound that the C (variant) allele of T�3246C (rs938671)decreased overall gene expression. The 3�-UTR effect wasverified by coupled in vitro transcription/translation of theentire/intact human ATP6V0A1 mRNA. Second, by overex-pression and knockdown of a specific miRNA, we found thatT�3246C disrupted a miRNA recognition motif for hsa-miR-637. Third, since ATP6V0A1 is an important component ofthe V-ATPase, chromatin granule pH was monitored byfluorescence of a CHGA/EGFP chimera: The �3246C alleleimpaired control of vacuolar pH. Fourth, increased pH(caused by the C allele) was then mimicked by the V-ATPaseinhibitor bafilomycin A1. After bafilomycin A1, the ratio ofCHGA precursor to its catestatin fragment was diminished.
Bafilomycin A1 also decreased exocytotic secretion fromthe regulated pathway. The results point to new molecularand mRNA translational strategies for probing autonomiccontrol of the circulation and ultimately the susceptibilityto and pathogenesis of cardiovascular disease states suchas hypertension.
Sources of FundingThis study was supported by the National Institutes of Health[HL58120; RR00827 (UCSD General Clinical Research Center);MD000220 (UCSD Comprehensive Research Center in HealthDisparities, CRCHD)], Department of Veterans Affairs.
DisclosuresNone.
References1. Mahata SK, O’Connor DT, Mahata M, Yoo SH, Taupenot L, Wu H, Gill
BM, Parmer RJ. Novel autocrine feedback control of catecholaminerelease: a discrete chromogranin a fragment is a noncompetitive nicotiniccholinergic antagonist. J Clin Invest. 1997;100:1623–1633.
2. Taupenot L, Harper KL, O’Connor DT. The chromogranin-secretograninfamily. N Engl J Med. 2003;348:1134–1149.
3. Mahata SK, Mahata M, Fung MM, O’Connor DT. Catestatin: a multi-functional peptide from chromogranin a. Regul Pept. 2010;162:33–43.
4. O’Connor DT, Kailasam MT, Kennedy BP, Ziegler MG, Yanaihara N,Parmer RJ. Early decline in the catecholamine release-inhibitory peptidecatestatin in humans at genetic risk of hypertension. J Hypertens. 2002;20:1335–1345.
5. Mahapatra NR, O’Connor DT, Vaingankar SM, Hikim AP, Mahata M,Ray S, Staite E, Wu H, Gu Y, Dalton N, Kennedy BP, Ziegler MG, RossJ, Mahata SK. Hypertension from targeted ablation of chromogranin a canbe rescued by the human ortholog. J Clin Invest. 2005;115:1942–1952.
6. O’Connor DT, Zhu G, Rao F, Taupenot L, Fung MM, Das M, Mahata SK,Mahata M, Wang L, Zhang K, Greenwood TA, Shih PA, Cockburn MG,Ziegler MG, Stridsberg M, Martin NG, Whitfield JB. Heritability andgenome-wide linkage in US and Australian twins identify novel genomicregions controlling chromogranin a: implications for secretion and bloodpressure. Circulation. 2008;118:247–257.
7. Brody LC, Abel KJ, Castilla LH, Couch FJ, McKinley DR, Yin G, Ho PP,Merajver S, Chandrasekharappa SC, Xu J, Cole JL, Struewing JP, ValdesJM, Collins FS, Weber BL. Construction of a transcription map sur-rounding the brca1 locus of human chromosome 17. Genomics. 1995;25:238–247.
8. Kim JH, Johannes L, Goud B, Antony C, Lingwood CA, Daneman R,Grinstein S. Noninvasive measurement of the pH of the endoplasmicreticulum at rest and during calcium release. Proc Natl Acad Sci U S A.1998;95:2997–3002.
9. Miesenbock G, De Angelis DA, Rothman JE. Visualizing secretion andsynaptic transmission with pH-sensitive green fluorescent proteins.Nature. 1998;394:192–195.
10. Wu MM, Grabe M, Adams S, Tsien RY, Moore HP, Machen TE.Mechanisms of pH regulation in the regulated secretory pathway. J BiolChem. 2001;276:33027–33035.
11. Wu MM, Llopis J, Adams S, McCaffery JM, Kulomaa MS, Machen TE,Moore HP, Tsien RY. Organelle pH studies using targeted avidin andfluorescein-biotin. Chem Biol. 2000;7:197–209.
12. Taupenot L, Harper KL, O’Connor DT. Role of H�-ATPase-mediatedacidification in sorting and release of the regulated secretory proteinchromogranin A: evidence for a vesiculogenic function. J Biol Chem.2005;280:3885–3897.
13. Nishi T, Forgac M. The vacuolar (H�)-ATPases: nature’s most versatileproton pumps. Nat Rev Mol Cell Biol. 2002;3:94–103.
14. Courel M, Soler-Jover A, Rodriguez-Flores JL, Mahata SK, Elias S,Montero-Hadjadje M, Anouar Y, Giuly RJ, O’Connor DT, Taupenot L.Pro-hormone secretogranin II regulates dense core secretory granulebiogenesis in catecholaminergic cells. J Biol Chem. 2010;285:10030–10043.
15. Taupenot L. Analysis of regulated secretion using pc12 cells. Curr ProtocCell Biol. 2007;15:1512.
388 Circ Cardiovasc Genet August 2011

16. Bowman EJ, Siebers A, Altendorf K. Bafilomycins: a class of inhibitorsof membrane ATPases from microorganisms, animal cells, and plantcells. Proc Natl Acad Sci U S A. 1988;85:7972–7976.
17. Yoshimori T, Yamamoto A, Moriyama Y, Futai M, Tashiro Y. Bafi-lomycin a1, a specific inhibitor of vacuolar-type H�-ATPase, inhibitsacidification and protein degradation in lysosomes of cultured cells. J BiolChem. 1991;266:17707–17712.
18. Pillai RS, Bhattacharyya SN, Filipowicz W. Repression of protein syn-thesis by miRNAs: how many mechanisms? Trends Cell Biol. 2007;17:118–126.
19. Van Horssen AM, Martens GJ. Biosynthesis of secretogranin II inxenopus intermediate pituitary. Mol Cell Endocrinol. 1999;147:57–64.
20. Oda K, Nishimura Y, Ikehara Y, Kato K. Bafilomycin a1 inhibits thetargeting of lysosomal acid hydrolases in cultured hepatocytes. BiochemBiophys Res Commun. 1991;178:369–377.
21. Zhou Y, Lindberg I. Purification and characterization of the prohormoneconvertase pc1(pc3). J Biol Chem. 1993;268:5615–5623.
22. Jean F, Basak A, Rondeau N, Benjannet S, Hendy GN, Seidah NG,Chretien M, Lazure C. Enzymic characterization of murine and humanprohormone convertase-1 (mpc1 and hpc1) expressed in mammaliangh4c1 cells. Biochem J. 1993;292:891–900.
23. Louagie E, Taylor NA, Flamez D, Roebroek AJ, Bright NA, MeulemansS, Quintens R, Herrera PL, Schuit F, Van de Ven WJ, Creemers JW. Roleof furin in granular acidification in the endocrine pancreas: identificationof the v-ATPase subunit ac45 as a candidate substrate. Proc Natl Acad SciU S A. 2008;105:12319–12324.
24. O’Callaghan KM, Ayllon V, O’Keeffe J, Wang Y, Cox OT, Loughran G,Forgac M, O’Connor R. Heme-binding protein hrg-1 is induced byinsulin-like growth factor I and associates with the vacuolar H�-ATPaseto control endosomal pH and receptor trafficking. J Biol Chem. 2010;285:381–391.
25. Mishra PJ, Humeniuk R, Longo-Sorbello GSA, Banerjee D, Bertino JR.A Mir-24 microRNA binding-site polymorphism in dihydrofolatereductase gene leads to methotrexate resistance. Proc Natl Acad SciU S A. 2007;104:13513–13518.
26. Sethupathy P, Collins FS. MicroRNA target site polymorphisms andhuman disease. Trends Genet. 2008;24:489–497.
27. Chen K, Rajewsky N. Natural selection on human microRNA bindingsites inferred from SNP data. Nat Genet. 2006;38:1452–1456.
28. Friese RS, Mahboubi P, Mahapatra NR, Mahata SK, Schork NJ, Schmid-Schonbein GW, O’Connor DT. Common genetic mechanisms of bloodpressure elevation in two independent rodent models of human essentialhypertension. Am J Hypertens. 2005;18:633–652.
29. Lattanzi W, Bernardini C, Gangitano C, Michetti F. Hypoxia-like tran-scriptional activation in TMT-induced degeneration: microarrayexpression analysis on pc12 cells. J Neurochem. 2007;100:1688–1702.
30. Schulz N, Dave MH, Stehberger PA, Chau T, Wagner CA. Differentiallocalization of vacuolar h-ATPases containing a1, a2, a3, or a4(atp6v0a1-4) subunit isoforms along the nephron. Cell Physiol Biochem.2007;20:109–120.
31. Poea-Guyon S, Amar M, Fossier P, Morel N. Alternative splicing controlsneuronal expression of v-ATPase subunit a1 and sorting to nerve ter-minals. J Biol Chem. 2006;281:17164–17172.
32. Hiesinger PR, Fayyazuddin A, Mehta SQ, Rosenmund T, Schulze KL,Zhai RG, Verstreken P, Cao Y, Zhou Y, Kunz J, Bellen HJ. Thev-ATPase v0 subunit a1 is required for a late step in synaptic vesicleexocytosis in drosophila. Cell. 2005;121:607–620.
33. Sun-wada GH, Toyomura T, Murata Y, Yamamoto A, Futai M, Wada Y.The a3 isoform of v-ATPase regulates insulin secretion from pancreatic�-cells. J Cell Sci. 2006;119:4531–4540.
34. Hurtado-Lorenzo A, Skinner M, El Annan J, Futai M, Sun-Wada GH,Bourgoin S, Casanova J, Wildeman A, Bechoua S, Ausiello DA, BrownD, Marshansky V. V-ATPase interacts with arno and arf6 in earlyendosomes and regulates the protein degradative pathway. Nat Cell Biol.2006;8:124–136.
35. Williamson WR, Wang D, Haberman AS, Hiesinger PR. A dual functionof V0-ATPase a1 provides an endolysosomal degradation mechanism indrosophila melanogaster photoreceptors. J Cell Biol. 2010;189:885–899.
36. Forgac M. Vacuolar ATPases: rotary proton pumps in physiology andpathophysiology. Nat Rev Mol Cell Biol. 2007;8:917–929.
37. Gottlieb RA, Gruol DL, Zhu JY, Engler RL. Preconditioning in rabbitcardiomyocytes: role of pH, vacuolar proton ATPase, and apoptosis.J Clin Invest. 1996;97:2391–2398.
38. Smith AN, Skaug J, Choate KA, Nayir A, Bakkaloglu A, Ozen S, HultonSA, Sanjad SA, Al-Sabban EA, Lifton RP, Scherer SW, Karet FE.Mutations in atp6n1b, encoding a new kidney vacuolar proton pump116-kd subunit, cause recessive distal renal tubular acidosis with pre-served hearing. Nat Genet. 2000;26:71–75.
39. Ramachandran N, Munteanu I, Wang P, Aubourg P, Rilstone JJ, IsraelianN, Naranian T, Paroutis P, Guo R, Ren ZP, Nishino I, Chabrol B,Pellissier JF, Minetti C, Udd B, Fardeau M, Tailor CS, Mahuran DJ,Kissel JT, Kalimo H, Levy N, Manolson MF, Ackerley CA, MinassianBA. Vma21 deficiency causes an autophagic myopathy by compromisingv-ATPase activity and lysosomal acidification. Cell. 2009;137:235–246.
40. Rapp JP. Genetic analysis of inherited hypertension in the rat. PhysiolRev. 2000;80:135–172.
CLINICAL PERSPECTIVEHere we explored molecular mechanisms of how a polymorphism (T�3246C) in the 3�-UTR of the proton translocatingATPase (V-ATPase) subunit ATP6V0A1 influences human cardiovascular traits, including chromogranin A (CHGA) andBP. First, using a luciferase reporter plasmid, we found that the C (variant) allele of T�3246C (rs938671) decreased overallgene expression. The 3�-UTR effect was verified by coupled in vitro transcription/translation of the entire/intact humanATP6V0A1 mRNA. Second, by over-expression and knockdown of a specific microRNA, we found that T�3246Cdisrupted a microRNA recognition motif for hsa-miR-637. Third, since ATP6V0A1 is an important component of theV-ATPase, chromatin granule pH was monitored by fluorescence of a CHGA/EGFP chimera: the �3246C allele impairedcontrol of vacuolar pH. Fourth, increased pH (caused by the C allele) was then mimicked by the V-ATPase inhibitorbafilomycin A1. After bafilomycin A1, the ratio of CHGA precursor to its catestatin fragment was diminished. BafilomycinA1 also decreased exocytotic secretion from the regulated pathway. The results point to new molecular and mRNAtranslational strategies for probing autonomic control of the circulation, and ultimately the susceptibility to andpathogenesis of cardiovascular disease states such as hypertension.
Wei et al ATP6V0A1 Polymorphism 389
Copyright © 2022 FDOKUMEN




















