
Preparation of lipoproteins containing cation-dependent ATPase
11
BIOCHIMICA ET BIOPHYSICA ACTA 603 BBA 25153 PREPARATION OF LIPOPROTEINS CONTAINING CATION-DEPENDENT ATPase K. AHMED AND J. D. JUDAH Division of Metabolic Research, The Chicago Medical School, Institute for Medical Research, Chicago, Ill. (U.S.A.) (Received March I6th, 1964) SUMMARY I. Methods for the isolation of Na +, K + and Mga+ activated ATPases from a variety of tissues are described. The use of mannitol rather than of sucrose appears to be of decisive importance in some instances such as that of liver. 2. Preparations obtained by this method show extremely high degrees of stimu- lation in the presence of Na+ and K +, together with Mg2+, when compared to activity in the presence of Mg2+ alone. Stimulations of between 5oo and 7o0 % are commonly obtained. 3. Upon floatation of the preparations in high-density sucrose solutions, activity was recovered in material at the top of the tubes, implying that the ATPase system is a lipoprotein. 4. Preparations from different tissues exhibit differing sensitivities to strophan- thin-G, in agreement with sensitivities of the parent tissues. In every instance, it can be shown that inhibition by strophanthin-G varies inversely as the K + concen- tration of the system. 5. It can be shown that the brain lipoprotein splits p-nitrophenyl phosphate disodium. In the presence of K +, but not of Na+ or choline, the rate of splitting is accelerated 5-fold. It is inhibited by strophanthin-G. This supports the view that K + takes part in a phosphatase reaction. 6. Labelling of the ATPase systems of brain and liver shows that 32p passes from E32p!ATP into phosphoproteins in the system. Na+ accelerates this passage. In the presence of Na+ and K+ together, the radioaetLdty of the phosphoprotein is greatly reduced. INTRODUCTION Enzyme systems which split ATP in the presence of Na +, K + and Mg2+ are now regarded as playing an important role in the transport of Na + in many tissues. SKou 1 was the first to demonstrate this system using crab nerve; more recently SKou z has obtained preparations from kidney and brain. The enzyme system appears to reside in the microsomal fraction of many cellsa,4; but in human erythrocytes the activity has been recovered from membrane fragments 5. In kidney (and perhaps in other tissues) powerful Na+, I¢+ and Mg/+ activated ATPases are also found in the Biochim. Biophys. Acta, 93 (1964) 6o3k-613
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Preparation of lipoproteins containing cation-dependent ATPase

BIOCHIMICA ET BIOPHYSICA ACTA 603
BBA 25153
P R E P A R A T I O N OF L I P O P R O T E I N S C O N T A I N I N G
C A T I O N - D E P E N D E N T ATPase
K. A H M E D AND J. D. J U D A H
Division of Metabolic Research, The Chicago Medical School, Institute for Medical Research, Chicago, Ill. (U.S.A.)
(Received March I6th , 1964)
S U M M A R Y
I. Methods for the isolation of Na +, K + and Mg a+ activated ATPases from a variety of tissues are described. The use of mannitol rather than of sucrose appears to be of decisive importance in some instances such as that of liver.
2. Preparations obtained by this method show extremely high degrees of stimu- lation in the presence of Na+ and K +, together with Mg 2+, when compared to activity in the presence of Mg 2+ alone. Stimulations of between 5oo and 7o0 % are commonly obtained.
3. Upon floatation of the preparations in high-density sucrose solutions, activity was recovered in material at the top of the tubes, implying that the ATPase system is a lipoprotein.
4. Preparations from different tissues exhibit differing sensitivities to strophan- thin-G, in agreement with sensitivities of the parent tissues. In every instance, it can be shown that inhibition by strophanthin-G varies inversely as the K + concen- tration of the system.
5. I t can be shown that the brain lipoprotein splits p-nitrophenyl phosphate disodium. In the presence of K +, but not of Na+ or choline, the rate of splitting is accelerated 5-fold. It is inhibited by strophanthin-G. This supports the view that K + takes part in a phosphatase reaction.
6. Labelling of the ATPase systems of brain and liver shows that 32p passes from E32p!ATP into phosphoproteins in the system. Na+ accelerates this passage. In the presence of Na+ and K+ together, the radioaetLdty of the phosphoprotein is greatly reduced.
INTRODUCTION
Enzyme systems which split ATP in the presence of Na +, K + and Mg 2+ are now regarded as playing an important role in the transport of Na + in many tissues. SKou 1 was the first to demonstrate this system using crab nerve; more recently SKou z has obtained preparations from kidney and brain. The enzyme system appears to reside in the microsomal fraction of many cellsa,4; but in human erythrocytes the activity has been recovered from membrane fragments 5. In kidney (and perhaps in other tissues) powerful Na+, I¢+ and Mg/+ activated ATPases are also found in the
Biochim. Biophys. Acta, 93 (1964) 6o3k-613

604 K. AHMED, J. D. JUDAH
"nuclear" fraction 6 which also contains much debris and probably fragments of the cell membrane.
In the present work it is shown that Na +, K+ and Mg 2+ activated ATPases occur in liver cells, both in the "nuclear" and in the microsomal fractions. Previous com- parative studies23 have indicated that no such system occurs in liver microsomes, while others have demonstrated rather poorly stimulated systems s. Our own prepa- rations exhibit 5-7-fold stimulation of ATP breakdown in the presence of Na +, K + and Mg 2+ as compared to Mg 2+ alone. The methods are applicable to other tissues to yield active preparations which show similar degrees of activation by Na + and K+. The response of these systems to cardiac glycosides, the effect of labelling them with the aid of [a2P!ATP and the demonstration of a phosphatase highly dependent upon the presence c~f K + are also discussed.
METHODS AND MATERIALS
Animals
Male Sprague-Dawley rats (body wt. 250-300 g) were used. They were kept on a pellet diet, supplemented by c~-tocopherol (20 mg by mouth) thrice weekly as de- scribed previously 9. Nucleotides were obtained from Sigma Chemical Co., St. Louis, Mo. ATP was rendered alkali metal free by passage through Dowex-5o resin which had first been converted to the Tris form. KC1, NaC1 and choline were recrystallised before use. Sucrose of "enzyme-quality" was obtained from Mann Biochemicals Co., New York. Mannitol was of the highest purity available. EDTA was obtained from Sigma Chemical Company and neutralised with Tris before use. Strophanthin-G was obtained from Mann Biochemicals, New York. Inorganic phosphate (Pl) was estimated either by the method of FISKE AND SUBBARow 1° or by the method of BERENBLUM AND CHAIN 11. Protein was determined by the method of LOWRY et al. 12.
The system always used for the assay of ATPase activity was: ATP (Tris salt), 5 raM; MgSO4, 5 raM; Tris buffer (pH 7.4 o, glass electrode, measured at 2o°), 30 raM; NaC1, 115 mM and KC1, IO mM in a final volume of 2.0 ml. Where either NaC1 or KC1 was left out, equimolar amounts of choline chloride were substituted. The hydrolysis of p-nitrophenyl phosphate disodium (obtained from Sigma Chemical Company, St. Louis, Mo.) was measured in the following system: MgSO4, 5 mM;Tris buffer (pH 7.4o), IOO mM and p-nitrophenyl phosphate disodium, 5 raM. Final volume, I.O ml. Ions were added as indicated in the text. The reaction was stopped by addition of trichloroacetic acid (final conc. 5 %, w/v).
A o.5-ml aliquot of the clear supernatant after centrifugation was added to a measured volume of I M Tris (pH about IO.4), final volume 3.0 ml, and the yellow solution was read at 400 m/~ in the Zeiss PMQ spectrophotometer against a suitable blank to correct for the small amount of free phenol present. [3~PIATP was prepared as described previously 9. It was freed from ammonium and other ions by first adsorption on charcoal, elution by ammonia-ethanol (ammonium hydroxide-ethanol-water , IO:25:65, v/v) and subsequent passage through Dowex-5o (Tris form). The ATP solution was then concentrated to a suitable volume by lyophilisation.
Phosphoprotein was assayed as described previously 9.
Preparation of the A TPases
A IO % homogenate of rat liver was prepared in a medium consisting of 0.25 M
Biochim. Biophys. dcta, 93 (1964) 6o3-613

CATION STIMULATED ATPASES 605
mannitol, 5 mM EDTA (Tris salt), 3o mM histidine-HC1 (pH 6.5) and deoxycholate (o.I %, w/v, dissolved as a Tris salt). The final pH was 6.8 (glass electrode, o°). A Dounce homogeniser was used, with loose fitting pestle (these may be obtained from the Blaessig Glass Co., Rochester, N.Y.). The homogenate was centrifuged at 8000 × g for 20 rain in a refrigerated centrifuge, and the supernatant fluid (fraction SI) was carefully decanted and frozen. I t was stored at - - IO °. After storage for 24-48 h, SI was thawed and centrifuged for 35 rain at IOOOOO x g (av.) in the Spinco Model L preparative centrifuge, using the No. 4 ° or No. 50 rotor. The supernatant from this step showed little ATPase activity and was discarded. The pellet (fraction RI) was suspended in a medium containing o.25 M mannitol, I.O mM EDTA (Tris salt) and 30 mM Tris-HC1 (pH 7.4, glass electrode, o°), which was kept ice-cold. The suspension was gently homogenised (Dounce homogeniser) and centrifuged at 20000 x g for 50 rain at o °. The residue (fraction R2) was suspended in the same medium. The supernatant was discarded. Fraction R2 was perfectly suitable for studies of the ATPase reaction. I t could be stored frozen for 3 weeks or more without appreciable loss of activity. After this time, activity fell gradually but unpredictably.
Isolation of lipoprotein fraction
Fraction R2 was suspended in a minimum volume of 0.25 M mannitol, containing I.O mM EDTA and 30 mM Tris-HC1 (pH 7.40). To this suspension was added slowly and with constant stirring a solution containing sucrose, 2.1 M; EDTA (Tris salt), 5 mM and 2o mM Tris- HC1 (pH 7.40). I t was gently homogenised to obtain a uniform suspension and the density of the suspension was adjusted to 1.22 by addition of above mentioned medium (mannitol) as necessary. This suspension was centrifuged for 5-6 h in the swinging bucket rotor (No. SW 25.1) at 61500 × g (av.) in the Spinco Model L centrifuge. A layer formed at the top of the sucrose and was skimmed off with a spatula. Tile No. 4 ° or No. 50 rotors of the Spinco may also be used for this step, applying a force of IOOOOO × g (av.) for 3 h. These angle heads are not so con- venient for harvesting the lipoprotein.
The fraction so obtained (lipoprotein) was suspended in m a n n i t o l - E D T A - T r i s as described above and stored frozen at - - IO °. I t was stable for several weeks. I t is advisable to store in small aliquots so that frequent freezing and thawing is avoided; however, preparations have survived two freezings and thawings without loss of activity. The procedure described above has been applied to rat brain and kidney, and also to the smooth muscle of guinea-pig ileum. For the last named, the ileum was washed free of contents by the use of 0.25 M mannitol, and the mucosa was gently scraped off with the edge of a spatula. The remainder of the tissue was finely minced with scissors under ice-cold homogenising medium and then it was homogenised and put through the procedure described above. In each case, active lipoprotein fractions were obtained. A detailed flow sheet for the liver system is given in Table II .
Preparation of the A TPase system from microsomes
Microsomes isolated from rat liver are also suitable for the preparation of Na + and K+ activated ATPase. The microsomes can be homogenised in the deoxycholate medium described above and frozen as usual. A fully active lipoprotein fraction can be isolated by following the procedure described above. The activity of this prepa- ration is the same as that described in the previous section.
Biochim. Biophys. Acta, 93 (1964) 6o3-613

606 K. AHMED, J. D. JUDAH
R E S U L T S
The Na +, K + and Mg ~+ stimulated A TPase of liver homogenates
Pre l iminary exper iments showed tha t 0.25 M sucrose homogenates of rat liver conta ined an ATPase system which was s t imula ted by Mg ~+, and further s t imula ted by the s imul taneous presence of Na + (115 mM) and K + (IO raM). Ei ther of these cations was ineffective by itself. Differential centr i fugat ion showed tha t almost all the ac t iv i ty of the homogenate could be recovered in the "nuclear" fraction, which also contains much debris, and is sedimented by centr i fugat ion at 800 × g for IO min. At tempts at separat ing the ac t iv i ty from the crude fraction were unsuccessful, and were complicated by the rapid loss of ac t iv i ty with time. The loss was observed only in the Na +- and K+-s t imula ted fraction of the ATPase ac t iv i ty ; and it was accom- panied b y the s imul taneous increase in ac t iv i ty in the presence of Mg 2+ alone. I t was decided tha t a l though considerable ac t iv i ty could be demonstra ted, the properties of the fraction made it useless for our purpose. Table I shows some results obtained
TABLE [
D I S T R I B U T I O N O F Mg 2+ P L U S N a + P L U S tX + S T I M U L A T E D ATPases I N F R E S H R A T - L I V E R H O M O G E N A T B I N 0 . 2 5 M S U C R O S E
Rat-liver homogenate made in 0.25 M sucrose was fractionated as described in the text. Standard ATPase assay as described in METHODS AND MATERIALS WaS used. Temperature 38°, incubation
time io rain, tissue equivalent to 20 mg fresh liver was used in every case.
, 4 T P split (/~moles/g/h) in presence of IP Expt. Tissue jraction Drug - - - ( M g 2+ plus Na + No. Mg2+ Mg~+ :'~[g2+ plus plus K +)
3lgZ~ plus K + plus N a + N a + plus K + minus Mg ~
I Homogenate - I35 135 135 600 4()5 "Nuclear" debris 15o 288 i6o 5io 36o Mitochondria -- 90 9o 9 o t 20 3 ° Supernatant - 15 ° 15 ° 15 ° 165 ~ 5
2 Homogenate - - 280 280 280 6o0 32o Homogenate Strophantbin-G
( I O 4 M) 280 280 280 390 IiO
with this crude ATPase system. I t is seen tha t it behaves much as expected, being dependent upon the presence of Na+, K + and Mg 2+ together, and being inhibi ted by s t rophanth in-G in concentra t ions comparable to those inhibi t ing K + uptake by liver slices. The other fractions of the liver homogenate in sucrose were near ly devoid of Na +, K + and Mg 2+ act ivated ATPase act ivi ty, and all a t t empts to show any signi- ficant separat ion were failures. We had observed during our exper iments with the "nuclear" fraction tha t subs t i tu t ion of mann i to l for sucrose as homogenising medium gave somewhat stabler preparat ions and we decided to invest igate this sugar in the preparat ion of microsomal fractions from liver.
The Na +, K+ and Mg 2+ activated A TPase of liver microsomes
Freshly prepared microsomes prepared from either 0.25 M sucrose or 0.25 M manni to l homogenates possess a Mg2+-stimulated ATPase. Addit ion of Na+ and K+ to such systems results in little or no increase in ac t iv i ty over tha t observed in the
Biockim. Biophys. Acta, 93 (~964) 6o3-613

CATION S T I M U L A T E D A T P A S E S 607
T A B L E I I
PURIFICATION OF N a +, I ( + AND Mg 2+ ACTIVATED A T P a s e OF LIVER MICROSOMES
T h e a s s a y and prepara t ion of f rac t ions are as descr ibed in METHODS AND MATERIALS. The figures in parenthese s af ter those in the " 'AP" c o l u m n are the rat ios of a c t i v i t y in the presence of Mg e+
p lus N a + p lus K + / a c t i v i t y in presence of Mg ~+ alone and are a m e a s u r e of s t i m u l a t i o n .
A T P split (amoles/mg protein[h) in presence of A P Recovered (Mg *+ plus Na + Total A TPase
Fraction Mg 2+ Mg*+ MgZ* - minus (o ) Mg 2+ plus plus K ÷) in fraction activity plus Na + plus K + Na~p+lus Mg 2+
Fresh S 1 3.0 3.0 3.0 4.0 i .o ( i .25) 300 i o o
R I I . o4 1.o4 1-o4 3.34 2.30 (3 -2) 27 9
R2 i . oo i . oo i . oo 4.7 ° 3.7 ° (4.7) 14 4-7
Lipopro te in 2.0 2.0 2.0 11.4o 9.4 (5.7) 14 4.7
presence of Mg 2+ alone. It was decided that a test of deoxycholate on these prepa- rations was worthwhile, since SKOU 2 had shown that the use of this agent improved the proportion of Na 2+ and K+ stimulated ATPase to unstimulated. Preliminary tests with sucrose and mannitol homogenates showed that even with deoxycholate in the medium, preparations obtained from homogenates made in sucrose were useless, since the total of ATPase acti~'ity rose after deoxycholate treatment, but the effect of Na + and K + was either absent or minimal. Mannitol homogenates yielded micro- somal preparations which, on the contrary, showed lower basic activity, with con- siderable stimulation in the presence of Na+ and K +. Table II shows a flow sheet for the preparation. A few additional remarks are necessary. The basic activity of the enzyme system toward ATP (obtained in presence of Mg 2+ alone) declines as the preparations are aged. The decline being greater than that of the total activity in the presence of Na +, K + and Mg 2+, the proportion of stimulated to unstimulated activity rises steadily. Ageing of the preparation may be conducted at 5 ° , but is hastened by freezing the preparation. We state that a period of 24-48 h is adequate for the ageing of the frozen material, but we have observed often that freezing for
T A B L E I I I
ACTIVITY OF N a +, K + AND l~g 2+ ACTIVATED A r P a s e s FROM SEVERAL TISSUES
The a s s a y and preparat ion of fract ions are as described in METHODS aND MATERIALS. T h e r e su l t s for brain l ipoprote in are r e p r e s e n t a t i v e of i o s u c h preparat ions . T w o prepara t ions were m a d e
f r o m rat k i d n e y and one from s m o o t h musc l e der ived from g u i n e a - p i g i l eum.
Tissue Fraction
A T P s p l i t (Bmoles/mg protein/h) in presence of
Mg ~+ Mg2+ plus K + Mg ~+ plus K + Mg *+ plus Na + plus Na +
Ratio A P Mg2+plus Na +
plus K+/Mg 2+
Brain L ipopro te in 12. 5 ± 2. 5 12. 5 ± 2. 5 12. 5 ~ 2. 5 81.o ~- 5 68.5
K i d n e y R2 7.8 7.8 7.8 24.0 16.2 R2 5.1 5.1 5.1 22. 4 17.3
S n l o o t h m u s c l e R2 3.1 3.1 3.1 14. 7 11.6
6.5
3.1 4.4
4.7
B i o c h i m . B i o p h y s . A c t a , 93 (1964) 6 o 3 - 6 1 3

6 0 8 K. AHMED, J . D. J U D A H
a week or more causes still further decline in basic activity to very low levels. It is noted that the procedure we employ results in considerable loss of total activity, only, a small fraction of the system being recovered. On the other hand, the basic activity in the presence of Mg 2+ alone is very low indeed, and excellent activity is observed upon addition of Na + and K + to the system. The general applicability of the method is illustrated in Table I I I , which gives results for several other tissues. It is to be noted that extremely high activities are obtained for the rat brain preparation.
Cation requirements for the enzwne system
The basic activity of the system was unaltered by the presence of Na + or K + alone at any concentration. Stimulation depended upon the presence of both Na+ and K + together (Fig. IA). When the concentration of Na + is fixed (I15 raM), and the concentration of K + increased, the activity of the ATPase rises to an optimum and then falls. The optimum K + concentration at Na + concentration of 115 mM is about IO mM (Fig. IB). Neither Li + nor choline would substitute for either Na+ or K +. The presence of Mg 2+ is essential for the effect of Na+ and I£+
A
ck <3
i
0 19 39 77 IL5 135 154 154 r35 115 77 39 19 0
Fig . I . S t a n d a r d A T P a s e a s s a y s y s t e m
x .
L A N O (raM) 0 5 I0 t5 20 25 30 35
K(mM) mM K in medium
as d e s c r i b e d in ~ I E T H O D S A N D M A T E R I A L S ~,vas used . h i F ig . IA , c o n c e n t r a t i o n s of b o t h N a + a n d K + w e r e v a r i e d as s h o w n . I n F ig . IB , N a + c o n c e n t r a t i o n w a s f ixed a t 115 m M a n d K + c o n c e n t r a t i o n w a s v a r i e d as s h o w n . LIP re fe r s to a c t i v i t y w i t h Mg 2+,
N a + a n d K + m i n u s a c t i v i t y w i t h Mg 2+ a lone . L i v e r l i p o p r o t e i n f r a c t i o n w a s t e s t e d .
pH optimum for A TPase activity
The liver system was investigated, and shows a pH optimum at pH 7.20-7.40. On either side of this range, activity falls rapidly. The loss of activity is due to loss of stimulated activity only. The basic ATPase in presence of Mg 2+ alone is relatively insensitive to pH change.
Effect of strophanthin-G and effect of K+ ions
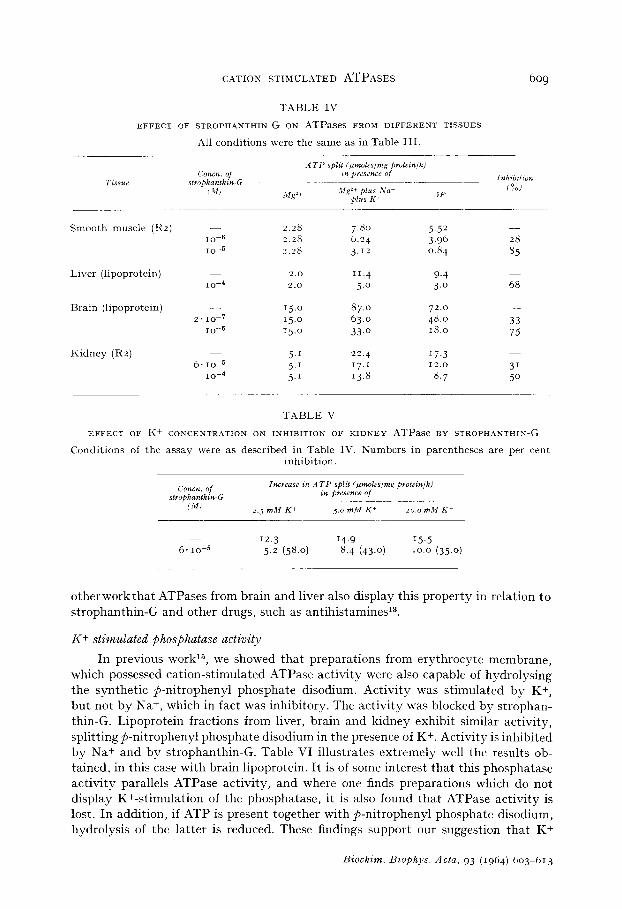
The Na +- and K+-stimulated portion of the ATPase activity, but not the basic activity in the presence of Mg 2+ alone (or in combination with any one of the alkali metals), is inhibited by strophanthin-G. The concentration of strophanthin-G required to inhibit varies from tissue to tissue, and is closely related to the concentration of inhibitor required to block ion transport in the parent tissue. Table IV illustrates this point. The inhibitory effect of strophanthin-G also varies with the K+ concentration of the medium. This is shown in Table V for the kidney system. We have shown in
Biochim. Biophys. Acla, 93 (1964) 603 613
CATION STIMULATED ATPASES 609
TABLE IV
EFFECT OF STROPHANTHIN-G ON ATPases FROM DIFFERENT TISSUES
All conditions were the same as in Table III.
Concn. of slrophanthmG
i.lfi
Smooth muscle (112) I o-6
10-S
Liver (lipoprotein) 10-d
Brain (lipoprotein) 2.10-T
10-E
Kidney (R2) 6. IO+
10-a
2.0
2.0
15.0 87.0 IS.0 63.0 rg.0 33.0
5.1 22.4 5.1 17.1 j.1 13.8
11.4 j.0
j.52
3.96 0.84
9-4 3.0
72.0 48.0 18.0
17.3 12.0 8.7
28
85
68
33 7j
31 50
TABLE V
EFFECT OF Ii+ CONCENTRATION ON INHIBITION OF KIDNEY ATPase BY STROPHANTHIN-G
Conditions of the assay were as described in Table IV. Numbers in parentheses are per cent inhibition.
12.3 14.9 15.5 6.10-5 5.2 (58.0) 8.4 (43.0) 10.0 (35.0)
otherworkthat ATPases from brain and liver also display this property in relation to strophanthin-G and other drugs, such as antihistamines13.
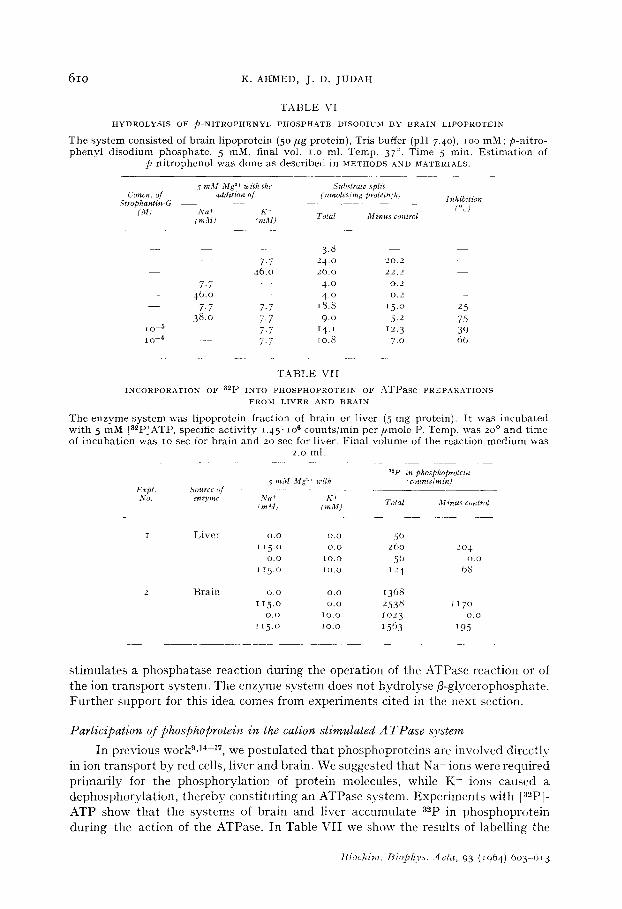
Kf stimulated $hosphatase activity
In previous work15, we showed that preparations from erythrocyte membrane, which possessed cation-stimulated ATPase activity were also capable of hydrolysing the synthetic $-nitrophenyl phosphate disodium. Activity was stimulated by K+, but not by Na+, which in fact was inhibitory. The activity was blocked by strophan- thin-G. Lipoprotein fractions from liver, brain and kidney exhibit similar activity, splitting@nitrophenyl phosphate disodium in the presence of Kf. Activity is inhibited by Na+ and by strophanthin-G. Table VI illustrates extremely well the results ob- tained, in this case with brain lipoprotein. It is of some interest that this phosphatase activity parallels ATPase activity, and where one finds preparations which do not display K+-stimulation of the phosphatase, it is also found that ATPase activity is lost. In addition, if ATP is present together with p-nitrophenyl phosphate disodium, hydrolysis of the latter is reduced. These findings support our suggestion that K+
Biochim. Biophys. Acta, 93 (1964) 603-613
610 K. AI-IMED, J. D. JUDAH
T A B L E VI
H Y D R O L Y S I S O F p - N I T R O P H E N Y L P H O S P H A T E D I S O D I U M B Y B R A I N L I P O P R O T E I N
T h e s y s t e m cons is ted of b ra in l ipoprote in (5 o / , g protein) , Tris buffer (pH 7.4o), lOO mM; p-ni t ro- p h e n y l d i sod ium phospha t e , 5 raM, final voI. I.o ml. Temp. 37 °. T ime 5 rain. E s t i m a t i o n of
p -n i t ropheno l was done as descr ibed in METHODS .aND MATF2RIALS.
5 m M Mg 2+ with the Substrate split Conch. of addition of (l,moles~rag protein~h)
Strophantin-G (M) Na + K +
Total Minus control ImM) (raM)
- - - 3.8 - 7.7 24-0 20.2
- - - 46.0 26.0 22.2 7.7 - - 4.0 0.2
-- 46.0 4.0 0.2 7.7 7-7 18.8 i5.o
38.o 7.7 9.o 5.2 lO 5 _ _ 7.7 14.t 12.3 IO 4 _ 7.7 lO.8 7.o
Inhibition (%)
2.5
75 39 6o
T A B L E VII
I N C O R P O R A T I O N O F a2p I N T O P H O S P H O P R O T E I N O F AWPase P R E P A R A T I O N S
F R O M L I V E R A N D B R A I N
The e n z y m e s y s t e m was l ipoprote in f ract ion of bra in or l iver (5 m g protein) . I t was i ncuba t ed wi th 5 m M [a=pZATP, specific ac t iv i ty i .45" IO6 coun t s / r a in p e r / , m o l e P. Temp. was 20 ° a n d t ime of i n c u b a t i o n was to sec for b ra in and 20 sec for liver. F ina l vo lume of the reac t ion m e d i u m was
2.o ml.
a2p in phosphoprotein .5 raM Mg ~-+ with (counts~rain)
Expt . Source of No. ¢~az3,me N a + K +
Total Minus control fra3l) traM)
I Liver O.O o.o 50 115.o o.o 26o 2o 4
o.o [o.o 56 o.o i 1 5 . o ~ o . o 124 68
o.o o.o 1368 1 1 5 . o O.O 2 5 3 8 1 1 7 0
0 . 0 1 0 . o 1 0 2 3 0 . 0
1 1 5 . 0 1o.o 15(> 3 195
2 Bra in
s t i m u l a t e s a p h o s p h a t a s e r e a c t i o n d u r i n g t h e o p e r a t i o n o f t h e A T P a s e r e a c t i o n o r o f
t h e i o n t r a n s p o r t s y s t e m . T h e e n z y m e s y s t e m d o e s n o t h y d r o l y s e f l - g l y c e r o p h o s p h a t e .
F u r t h e r s u p p o r t f o r t h i s i d e a c o m e s f r o m e x p e r i m e n t s c i t e d in t h e n e x t s e c t i o n .
Participation of phosphoprotein in the cation stimulated A TPase system
I n p r e v i o u s workg , 1~-17, w e p o s t u l a t e d t h a t p h o s p h o p r o t e i n s a r e i n v o l v e d d i r e c t l y
i n i o n t r a n s p o r t b y r e d ce l l s , l i v e r a n d b r a i n . W e s u g g e s t e d t h a t N a + i o n s w e r e r e q u i r e d
p r i m a r i l y f o r t h e p h o s p h o r y l a t i o n o f p r o t e i n m o l e c u l e s , w h i l e K + i o n s c a u s e d a
d e p h o s p h o r y l a t i o n , t h e r e b y c o n s t i t u t i n g a n A T P a s e s y s t e m . E x p e r i m e n t s w i t h Ia2p] -
A T P s h o w t h a t t h e s y s t e m s of b r a i n a n d l i v e r a c e m n u l a t e a2p in p h o s p h o p r o t e i n
d u r i n g t h e a c t i o n o f t h e A T P a s e . I n T a b l e V I I we s h o w t h e r e s u l t s o f l a b e l l i n g t h e
t3iochim. Biopkys..1eta, 93 (1964) 603 613

CATION STIMULATED A T P A S E S 611
liver lipoprotein with [32P~ATP. The presence of Mg ~t alone is sufficient to bring about some transfer of label. This is greatly increased by the addition of Na t , while K+ alone has little or no effect. When K + is added in the presence of Na t, the increase in labelling due to Na t is abolished or reduced greatly. Similar results are obtained with brain lipoprotein. Further studies on the mechanism of the enzyme are being carried out.
c 5.0
~ 40
~' 5.0
"~ 2.0 q
1.0
<~ 0 ~ _ _ t i i i t
6.2,,__66 _ _ 7 0 7.2j 74 7.8 82. v-
I m } d o z o l e 2 0 m M / v
Tris 20rnM
pH of the medium
Fig. 2. All cond i t ions for the &TPase assay were as descr ibed in METHODS AND MATERIALS, excep t t h a t the buffers of different p H as shown were used. I m i d a z o l e - H C I buffers were used for p H 6 .2 - 7. 2 and T r i s - H C 1 for p H 7.o-8.2, buffer c o n c e n t r a t i o n in the m e d i u m in each case be ing 20 mM. / ]P refers to a c t i v i t y w i t h Mg ~+, Na + a nd K + minus a c t i v i t y w i t h Mg 2+ alone. L ive r l i pop ro t e in
f rac t ion was tes ted.
DISCUSSION
From a practical point of view, the present work affords a simple method for the preparation of Na +, K + and Mg 2+ activated ATPases. This can be seen from the results. Even liver, which has not so far been used as a source of active preparations, yields enzymes capable of considerable activation by the monovalent cations. A serious drawback is the considerable loss of activity during the procedure. This is probably due to the use of deoxycholate in the preparation and so far we have not been able to escape this. On the other hand, the method yields preparations which show very large stimulations of activity in the presence of Na+ and K + from smooth muscle, brain and kidney as well as from liver. For our present purpose, which is the demonstration of the mechanism of action of the system this advantage outweighs the low yields of enzyme.
The demonstration that the ATPases may be floated on sucrose strongly suggests that they exist in the form of lipoproteins and opens the way for further purification, and possibly to solubilisation of these systems.
The use of mannitol in this preparation must be emphasised. We were unable to obtain active preparations from liver with the use of sucrose alone. We have not tested other polyhydric substances, but it is clear that their use might be advan- tageous, especially in obtaining better yields and possibly higher activity.
Two interesting points emerge from the use of cardiac glycosides as inhibitors: One is that there is close agreement between the sensitivity of the whole cells and
~iochim. Bioph3,s. Acta, 93 (1964) 6o3-613

6 1 2 K. AHMED, J . D, J U D A H
the isolated Na +, K + and Mg 2+ activated ATPase derived from that tissue to a given dose of strophanthin-G. Thus liver slices are relatively insensitive to the glycoside, and so is the ATPase derived from liver. On the contrary, brain is quite sensitive to strophanthin-G, and so is the ATPase. The difference between brain and liver enzymes is nearly 5oo-fold. We have discussed this problem elsewhere ~3 with reference to the action of antihistamine and other drugs, which also show remarkable differences in activity toward preparations derived from different tissues.
The second point follows from the first, it is that these enzyme systems contain iso-enzymes. The systems from brain, liver, kidney and other tissues must be different in detail, while being similar in general.
Our demonstration that the effect of strophanthin-G is reversed when the K + concentration of the medium is raised agrees well with the findings of DUNI-IAM AND GLYNN ~s, who used red-cell membranes. Similar findings are obtained with other drugs known to inhibit ion transport and the ATPases la.
Finally, the present work affords some support for our previous suggestion that phosphoproteins are intimately concerned in Na + and K + transport t:. We put forward the view that Na + was concerned in a phosphorylating reaction, while K + took part in a reaction by which phosphate groups are split from the phosphoprotein. We demonstrated such effects in red cells and liver slices. We now find that the ATPase system derived from liver and brain may be labelled with [a~P!ATP, and that label passes rapidly into phosphoprotein. In the presence of Na+ alone, phosphate rapidly accumulates in the phosphoserine groups, while in the presence of Na + and K + together, radioactivity is reduced The existence of a K+-dependent phosphatase reaction is further supported by the demonstration that p-nitrophenyl phosphate di- sodium is split by the brain, the red-cell and other ATPase preparations, the reaction being greatly accelerated in the presence of K +, (but not of Li+ or of Na+) and in- hibited by strophanthin-G.
ACKNOWLEDGEMENTS
We wish to thank Dr. L. A. MANSON of the Wistar Institute, Philadelphia, Pa., for his advice on the separation of lipoprotein.
This work was supported by Grant Number AM 7226-oi, from U.S. Public Health Service and also by grants from the Burroughs-Wellcome Fund, the Life Insurance Medical Research Fund and the Otho Sprague Memorial Foundation of Chicago.
Miss S. ELSEY and Mr. W. GARRETT gave skilled technical assistance throughout the work.
R E F E R E N C E S
1 j . C. SKOU, Biochim. Biophys. Acla, 23 (1957) 394. 2 j . C. SKOU, Bioehim. Biophys. Acta, 58 (1962) 314 .
D. H. DEUL a ~ n H. MClLWAIN, J. Neurochem., 8 (1961) 246. 4 j . JARNJ~FELT, Biochim. Bioph)'s. Acla, 59 (1962) 643, 655.
I¢.. L. PosT, C. R . MERRIT, C. R. IX~INSOLVING AND C. D. ALBRIGHT, J. Biol. Chem., 235 (196o) 796.
6 ]{. WHITTAI',t AND K. P. \¥HEELER, Biochim. Biophys. Acta, 5 t (1961) 622. v S. L. BONTING, L. C. CARAVAGGIO AND N. M. HAVCKINS, Arch. Biochem. Biophys., 88 (1962) 413 . s A. SCHWARTZ, Biochem. Biophys. Res. Commun., 9 (1962) 3 o l .
Bioahim. Biophys. Acta, 93 (1964) 603 -6J 3

CATION STIMULATED A T P A s E S 613
9 K. AHMED AND J. D. JUDAH, Bioehim. Biophys. Acla, 71 (1963) 295. 10 C. H. FISKE AND Y. SUBBAt{OW, J. Biol. Chem., 66 (1925) 375. 11 I. BERENBLUM AND E. CHAIN, Biochem. J., 32 (1938) 295. 12 O. H. LOWRY, N. J. ]~OSENBROUGH, A. L. FARR AND I{. J. RANDALL, .]. Biol. Chem., 193 (I95t)
265. 13 j . D. JUDAH AND I£. AHMED, J. Cell. Comp. Physiol., 64 (1964). 14 K. AHMED AND J. D. JUDAH, Biochim. Biophys. Acta, 57 (1962) 245. 15 j. D. JUDAH, t(. AHMED AND A. E. M. McLEAN, Biochim. Biophys. Acla, 65 (1962) 472. 16 ~ . AHMED, J. D. JUDAH AND H. WALLC, REN, Biochim. Biophys. Acla, 69 (1963) 42S. 17 j . D. JUDAH AND K. AHMED, Biol. Revs., 39 (1964) 16o. is E. T. DUNHAM AND I. M. GLYNN, J. _Physiol., 156 (1961) 274.
Bioehim. Biophvs. ~4cla, 93 (1964) 603 -6t3




















