
Classi¢cation and phylogeny of hydrogenases1
47
Classi¢cation and phylogeny of hydrogenases 1 Paulette M. Vignais a; *, Bernard Billoud b , Jacques Meyer c ; 2 a CEA/Grenoble, Laboratoire de Biochimie et Biophysique des Syste 'mes Inte ¤gre ¤s (UMR CEA/CNRS/UJF no. 5092), De ¤partement de Biologie Mole ¤culaire et Structurale, 17 Avenue des Martyrs, 38054 Grenoble Cedex 9, France b Atelier de Bioinformatique, Universite ¤ Paris 6, 12 rue Cuvier, 75006 Paris, France c CEA/Grenoble, Laboratoire de Bioe ¤nerge ¤tique Cellulaire et Pathologique, De ¤partement de Biologie Mole ¤culaire et Structurale, 17 Avenue des Martyrs, 38054 Grenoble Cedex 9, France Received 1 February 2001; accepted 28 May 2001 First published online 22 June 2001 Abstract Hydrogenases (H 2 ases) catalyze the reversible oxidation of molecular hydrogen and play a central role in microbial energy metabolism. Most of these enzymes are found in Archaea and Bacteria, but a few are present in Eucarya as well. They can be distributed into three classes: the [Fe]-H 2 ases, the [NiFe]-H 2 ases, and the metal-free H 2 ases. The vast majority of known H 2 ases belong to the first two classes, and over 100 of these enzymes have been characterized genetically and/or biochemically. Compelling evidence from sequences and structures indicates that the [NiFe]- and [Fe]-H 2 ases are phylogenetically distinct classes of proteins. The catalytic core of the [NiFe]-H 2 ases is a heterodimeric protein, although additional subunits are present in many of these enzymes. Functional classes of [NiFe]-H 2 ases have been defined, and they are consistent with categories defined by sequence similarity of the catalytic subunits. The catalytic core of the [Fe]-H 2 ases is a ca. 350-residue domain that accommodates the active site (H-cluster). A few monomeric [Fe]-H 2 ases are barely larger than the H-cluster domain. Many others are monomeric as well, but possess additional domains that contain redox centers, mostly iron^sulfur. Some [Fe]- H 2 ases are oligomeric. The modular structure of H 2 ases is strikingly illustrated in recently unveiled sequences and structures. It is also remarkable that most of the accessory domains and subunits of H 2 ases have counterparts in other redox complexes, in particular NADH- ubiquinone oxidoreductase (Complex I) of respiratory chains. Microbial genome sequences are bringing forth a significant body of additional H 2 ase sequence data and contribute to the understanding of H 2 ase distribution and evolution. Altogether, the available data suggest that [Fe]-H 2 ases are restricted to Bacteria and Eucarya, while [NiFe]-H 2 ases, with one possible exception, seem to be present only in Archaea and Bacteria. H 2 ase processing and maturation involve the products of several genes which have been identified and are currently being characterized in the case of the [NiFe]-H 2 ases. In contrast, near to nothing is known regarding the maturation of the [Fe]-H 2 ases. Inspection of the currently available genome sequences suggests that the [NiFe]-H 2 ase maturation proteins have no similar counterparts in the genomes of organisms possessing [Fe]-H 2 ases only. This observation, if confirmed, would be consistent with the phylogenetic distinctiveness of the two classes of H 2 ases. Sequence alignments of catalytic subunits of H 2 ases have been implemented to construct phylogenetic trees that were found to be consistent, in the main, with trees derived from other data. On the basis of the comparisons performed and discussed here, proposals are made to simplify and rationalize the nomenclature of H 2 ase-encoding genes. ß 2001 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved. Keywords : [NiFe]-hydrogenase ; [Fe]-hydrogenase ; Phylogeny ; Respiratory chain ; Complex I ; Ferredoxin Contents 1. Introduction .......................................................... 456 2. The three phylogenetically distinct classes of hydrogenases ........................ 457 3. Biosynthesis and localization of hydrogenases ................................. 459 3.1. Cellular functions of hydrogenases ...................................... 459 0168-6445 / 01 / $20.00 ß 2001 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved. PII:S0168-6445(01)00063-8 * Corresponding author. Tel.: +33 (4) 38 78 33 99; Fax: +33 (4) 38 78 51 85. E-mail address : [email protected] (P.M. Vignais). 1 The sequence alignments and phylogenetic trees are available at the web site : http ://wwwabi.snv.jussieu.fr/research/hydrogenases/index.html. 2 Also corresponding author. Tel.: +33 (4) 38 78 44 23; Fax: +33 (4) 38 78 58 72; E-mail address : [email protected]. FEMS Microbiology Reviews 25 (2001) 455^501 www.fems-microbiology.org Downloaded from https://academic.oup.com/femsre/article/25/4/455/525649 by guest on 14 March 2022
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Classi¢cation and phylogeny of hydrogenases1

Classi¢cation and phylogeny of hydrogenases1
Paulette M. Vignais a;*, Bernard Billoud b, Jacques Meyer c;2
a CEA/Grenoble, Laboratoire de Biochimie et Biophysique des Syste©mes Integres (UMR CEA/CNRS/UJF no. 5092),Departement de Biologie Moleculaire et Structurale, 17 Avenue des Martyrs, 38054 Grenoble Cedex 9, France
b Atelier de Bioinformatique, Universite Paris 6, 12 rue Cuvier, 75006 Paris, Francec CEA/Grenoble, Laboratoire de Bioenergetique Cellulaire et Pathologique, Departement de Biologie Moleculaire et Structurale, 17 Avenue des Martyrs,
38054 Grenoble Cedex 9, France
Received 1 February 2001; accepted 28 May 2001
First published online 22 June 2001
Abstract
Hydrogenases (H2ases) catalyze the reversible oxidation of molecular hydrogen and play a central role in microbial energy metabolism.Most of these enzymes are found in Archaea and Bacteria, but a few are present in Eucarya as well. They can be distributed into threeclasses: the [Fe]-H2ases, the [NiFe]-H2ases, and the metal-free H2ases. The vast majority of known H2ases belong to the first two classes,and over 100 of these enzymes have been characterized genetically and/or biochemically. Compelling evidence from sequences and structuresindicates that the [NiFe]- and [Fe]-H2ases are phylogenetically distinct classes of proteins. The catalytic core of the [NiFe]-H2ases is aheterodimeric protein, although additional subunits are present in many of these enzymes. Functional classes of [NiFe]-H2ases have beendefined, and they are consistent with categories defined by sequence similarity of the catalytic subunits. The catalytic core of the [Fe]-H2asesis a ca. 350-residue domain that accommodates the active site (H-cluster). A few monomeric [Fe]-H2ases are barely larger than the H-clusterdomain. Many others are monomeric as well, but possess additional domains that contain redox centers, mostly iron^sulfur. Some [Fe]-H2ases are oligomeric. The modular structure of H2ases is strikingly illustrated in recently unveiled sequences and structures. It is alsoremarkable that most of the accessory domains and subunits of H2ases have counterparts in other redox complexes, in particular NADH-ubiquinone oxidoreductase (Complex I) of respiratory chains. Microbial genome sequences are bringing forth a significant body ofadditional H2ase sequence data and contribute to the understanding of H2ase distribution and evolution. Altogether, the available datasuggest that [Fe]-H2ases are restricted to Bacteria and Eucarya, while [NiFe]-H2ases, with one possible exception, seem to be present only inArchaea and Bacteria. H2ase processing and maturation involve the products of several genes which have been identified and are currentlybeing characterized in the case of the [NiFe]-H2ases. In contrast, near to nothing is known regarding the maturation of the [Fe]-H2ases.Inspection of the currently available genome sequences suggests that the [NiFe]-H2ase maturation proteins have no similar counterparts inthe genomes of organisms possessing [Fe]-H2ases only. This observation, if confirmed, would be consistent with the phylogeneticdistinctiveness of the two classes of H2ases. Sequence alignments of catalytic subunits of H2ases have been implemented to constructphylogenetic trees that were found to be consistent, in the main, with trees derived from other data. On the basis of the comparisonsperformed and discussed here, proposals are made to simplify and rationalize the nomenclature of H2ase-encoding genes. ß 2001Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: [NiFe]-hydrogenase; [Fe]-hydrogenase; Phylogeny; Respiratory chain; Complex I; Ferredoxin
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4562. The three phylogenetically distinct classes of hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . 4573. Biosynthesis and localization of hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 459
3.1. Cellular functions of hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 459
0168-6445 / 01 / $20.00 ß 2001 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 1 6 8 - 6 4 4 5 ( 0 1 ) 0 0 0 6 3 - 8
* Corresponding author. Tel. : +33 (4) 38 78 33 99; Fax: +33 (4) 38 78 51 85.E-mail address: [email protected] (P.M. Vignais).
1 The sequence alignments and phylogenetic trees are available at the web site : http://wwwabi.snv.jussieu.fr/research/hydrogenases/index.html.2 Also corresponding author. Tel. : +33 (4) 38 78 44 23; Fax: +33 (4) 38 78 58 72; E-mail address: [email protected].
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
FEMS Microbiology Reviews 25 (2001) 455^501
www.fems-microbiology.org
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

3.2. Biosynthesis of [NiFe] active sites. Accessory genes . . . . . . . . . . . . . . . . . . . . . . . . . . 4603.3. Biosynthesis of [Fe] active sites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4613.4. Biosynthesis of Fe^S clusters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4613.5. Other accessory genes and regulatory genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4613.6. Signal peptides and subcellular localization of hydrogenases . . . . . . . . . . . . . . . . . . . . 462
4. [NiFe]-hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4664.1. The basic enzyme is a heterodimer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4664.2. Biodiversity of [NiFe]-H2ases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 467
5. [Fe]-hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4755.1. The catalytic subunit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4755.2. Additional subunits . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4765.3. Redox partners of [Fe]-H2ases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4775.4. Putative hydrogenases and hydrogenase-related proteins . . . . . . . . . . . . . . . . . . . . . . . 478
6. Hydrogenase-related genes in fully sequenced genomes . . . . . . . . . . . . . . . . . . . . . . . . . . . 4787. Phylogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 480
7.1. [NiFe]-hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4807.2. [Fe]-hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4847.3. Similarities between hydrogenases and NADH-ubiquinone oxidoreductases (Complex I)
of respiratory chains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4857.4. Distribution of hydrogenases in the domains of life . . . . . . . . . . . . . . . . . . . . . . . . . . 488
8. Nomenclature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4898.1. [NiFe]-hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4898.2. [Fe]-hydrogenases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 489
9. Concluding remarks and perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 490
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 490
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 491
1. Introduction
Hydrogenases (H2ases) catalyze the interconversion ofmolecular hydrogen and protons and electrons accordingto the reaction: H2I2H�+2e3. Although most, if not all,known H2ases can catalyze the reaction in either directionin vitro, they are usually committed to catalyze either hy-drogen uptake or evolution in vivo, depending on thedemands of the host organism.
Research in the ¢eld of H2ases is now at a turningpoint. Previously, traditional physiological and biochemi-cal studies have provided information on the cellular func-tion of H2ases. Accordingly, a number of early reviewshave been focused on the physiology of speci¢c organismssuch as aerobic hydrogen oxidizing bacteria [1,2], cyano-bacteria [3,4], photosynthetic bacteria [5^7], rhizobia [8^10], sulfate reducers [11,12], anaerobic fermenters [13^15],and extremophiles [16,17]. A shifted emphasis was appar-ent in subsequent reviews as a result of the widespread useof molecular biology and genetics [18^28]. The soaring ofgenome sequencing is resulting in a rapidly increasing £owof new data that provide new insights into the biodiversityand evolution of H2ases.
The overwhelming majority of H2ases are metalloen-zymes, and their metal sites belong to two main categories.One of these consists of the classical [2Fe^2S], [3Fe^4S],and [4Fe^4S] iron^sulfur clusters. These ubiquitous metalsites shuttle electrons between the H2-activating site and
the redox partners of H2ases. The general properties ofFe^S clusters are well known [29,30]. In contrast, theirbiosynthesis is not so well understood but is actively in-vestigated [31,32]. The second type of metal clusters arethe H2-activating sites, which are idiosyncratic to H2ases.They come in two varieties, the [NiFe] [33] and [Fe] [34,35]active sites.
Crystal structures have unveiled the general fold anddetails of the catalytic sites of several H2ases of the[NiFe] [33,36^39] and [Fe] [34,35,40^42] classes. Thesestructures have given a new stimulus to the implementa-tion of sophisticated spectroscopic techniques [43^48],chemical modeling [49^51], calculations ([52] and referen-ces therein) and analysis of enzyme kinetics [53]. Suchcombined e¡orts are providing novel insights into the cat-alytic mechanism of H2ases. Since surveys and discussionsof these aspects have appeared recently [42,51^53], theyhave not been dwelt upon in detail here. Crystal structuresof H2ases have also shed light on a number of other pen-dent questions. Among these, the modular structure ofH2ases, which had previously been inferred from primarystructures, has been con¢rmed. Also, the absence of aphylogenic relationship between the [NiFe]- and [Fe]-H2ases [21] has been put on ¢rm ground by the 3D struc-tures [33^42].
The compilation of sequence data demonstrates thatH2ases are related to other redox proteins and enzymes.Fe^S-containing subunits and modules of H2ases are
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501456
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

clearly homologous to soluble ferredoxins and parts ofrespiratory chain complexes, in particular NADH-ubiqui-none oxidoreductase (Complex I) [54^60]. Furthermore,such homologies appear to exist also in the case of theH2-activating domain of [NiFe]-H2ases [56,58,61]. Theserelationships con¢rm at the molecular level the existenceof extensive phylogenic connections between anaerobicand aerobic bioenergetic machines.
H2ases were ¢rst isolated from prokaryotes of the do-main Bacteria [13] then from Archaea [62]. The latter do-main comprises two kingdoms, the Euryarchaeota and theCrenarchaeota [63], many members of which rely on H2 asa main energy source. Finally, H2ases have also beenfound in subcellular organelles of eukaryotes, namely hy-drogenosomes of protozoa [64,65] and chloroplasts ofgreen algae [25,66^72]. These enzymes are thus presentin all major domains of life, and in a considerable varietyof physiological contexts.
The isolation of novel microorganisms and the increas-ingly fast pace of genomic sequencing are providing uswith a continuous supply of new H2ase sequences whichneed to be annotated, ordered, and which are likely,thanks to their ubiquity, to be very useful for the deriva-tion of evolutionary patterns. As the tools for retrievaland analysis of these data are being developed, informa-tion on the structure, function, and subcellular localizationof the encoded proteins can also be obtained at an unpre-cedented pace. We therefore intend to center this reviewon these novel aspects of hydrogenase research, with theexpectation that it will be helpful for the retrieval andanalysis of new data.
2. The three phylogenetically distinct classes ofhydrogenases
The ¢rst isolated and characterized H2ases were mono-mers or dimers and were found to be iron^sulfur proteins[13,73]. The classi¢cation was initially based on the iden-tity of speci¢c electron donors and acceptors, namelyNAD (hydrogenases of EC class 1.12.1.12), cytochromes(class 1.12.2.1), coenzyme F420 (class 1.12.99.1) or ferre-doxins (class 1.18.99.1).
Nickel was later found in the H2ase from Methanobac-terium thermoautotrophicum [62] then in many otherH2ases, and [Fe]-H2ases turned out to be fewer in num-bers. A subgroup of the [NiFe]-H2ases also contains sele-nium, as selenocysteine coordinated to the nickel[39,74,75]. From the comparison of the conserved se-quence elements in the nickel-binding regions of 16[NiFe]-H2ases, Voordouw [20] divided up these enzymesinto ¢ve subgroups. Subsequently, Wu and Mandrand [21]proposed a more elaborated classi¢cation based on theamino acid sequences of 30 microbial H2ases. In this re-view we report the analysis of a number of H2ase sequen-ces more than 3-fold higher, and have found that a few
readjustments are required. While the main features of the[NiFe]-H2ase classi¢cation [21] remains valid (Section 4),the [Fe]- and [NiFe]-H2ases are phylogenetically distinct,and their separation is to be placed at a level qualitativelydi¡erent from the separations among the various groupsof [NiFe]-H2ases. Also, one of the previously proposedgroups (Class VI) [21] consisted of putative H2ase sequen-ces [369] which have subsequently been shown to beclosely related to aminotransferases [370]. This group istherefore to be dismissed.
A metal-free H2ase has been discovered in some meth-anogens [76^78]. This enzyme is a homodimer encoded bya monocistronic gene [79^81], and is described as an H2-forming methylenetetrahydromethanopterin dehydroge-nase (Hmd) (EC 1.12.99.4). Hmd catalyzes the reversiblereduction of N5,N10-methenyltetrahydromethanopterin(methenyl-H4MPT�) with H2 to N5,N10-methylenetetrahy-dromethanopterin (methylene-H4MPT). The mechanismof H2 formation has been analyzed using hydrogen iso-topes [82,83] and the stereoselective hydride transfer by 2DNMR spectroscopy [84]. Together with the F420-dependentmethylenetetrahydromethanopterin dehydrogenase (Mtd),Hmd catalyzes the reduction of F420 with H2 [80,81,85].Both enzymes are induced during growth under conditionsof nickel deprivation [86]. Recently, it has been demon-strated that under Ni-limited growth, Hmd synthesis isincreased 6-fold in Methanothermobacter marburgensis(formerly M. thermoautotrophicum strain Marburg) [371]while the synthesis of F420-reducing hydrogenase (Frh) is20-fold lower than in cells grown on nickel replete medium[86]. The presence of an organic cofactor in these metal-free H2ases has recently been reported [87].
At this time, the sequences of altogether over 100H2ases are available, and the genomes of well over 40microorganisms have been sequenced. These data con¢rmthat, despite their increasingly conspicuous diversity inmany respects (size, quaternary structure, electron donorsand acceptors) H2ases consist of three phylogeneticallydistinct classes, the [Fe]-H2ases, the [NiFe]-H2ases, andthe metal-free H2ases, each characterized by a distinctivefunctional core which is conserved within each class. Thiscore consists of the subunits or domains that accommo-date the catalytic site and that are minimally required forstructure and function. Metal content as well as sequencesimilarity thus is a reliable classi¢cation criterion. The ex-istence of these phylogenetically independent classes ofH2ases has recently received additional and compellingsupport from X-ray crystallography, at least in the casesof [NiFe]- and [Fe]-H2ases [33^35,37]. Interestingly, theactive sites of the latter two classes of H2ases displaysome striking similarities in their structural frameworksand chemistry, despite the absence of any resemblancebetween the polypeptide folds that surround them[40,42,52]. This is deemed to be a well-supported case ofconvergent evolution. On the grounds of sequence data, itis most likely that the structure of the metal-free H2ases is
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 457
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

Tab
le1
Hom
olog
ous
hydr
ogen
ase
gene
clus
ters
enco
ding
[NiF
e]-h
ydro
gena
ses
inP
rote
obac
teri
a
R.
caps
ulat
usR
.sp
haer
oide
sR
.le
gum
inos
arum
B.
japo
nicu
mR
.eu
trop
haA
.vi
nela
ndii
A.
chro
ococ
cum
E.
coli
Thi
ocap
saro
seop
ersi
cina
Pse
udom
onas
hydr
ogen
ovor
aF
eatu
res
orfu
ncti
on
AB
CD
EF
GH
IJ
MB
HSH
H2as
e1
H2as
e2
H2as
e3
H2as
e2
H2as
e1
hupT
hupT
hupT
e(h
oxJ)
fhi
stid
ine
kina
sehu
pUhu
pUhu
pU(h
oxB
)fH
2se
nsor
,sm
all
subu
nit
hupV
hupV
hupV
(hox
C)f
subu
nit
H2
sens
or,
larg
esu
buni
thu
pShu
pShu
pShu
pSho
xKho
xYh
hoxK
hupS
hyaA
hybO
hycG
jhu
pShy
dSo
hupS
H2as
esm
all
subu
nit
hupL
hoxH
chu
pLhu
pLhu
pLho
xGho
xHh
hoxG
hupL
hyaB
hybC
jhy
cEj
hupL
hydL
ohu
pLH
2as
ela
rge
subu
nit
hupC
hupC
hupC
hupC
hoxZ
hoxZ
hupZ
hyaC
hybB
jhu
pChu
pCcy
toch
rom
eb
hupD
hupD
hupD
hupD
hoxM
hoxW
hho
xMhu
pMhy
aDhy
bDk
hycI
j;khu
pDhu
pDm
atur
atio
npr
otea
sek
hupE
dhu
pEhu
pFhu
pFhu
pFhu
pFho
xLho
xLhu
pNhu
pFhu
pGhu
pGhu
pGhu
pGho
xOho
xOhu
pOhy
aEhu
pGhu
pHhu
pHhu
pHhu
pHho
xQho
xQhu
pQhy
aFhu
pHhu
pHhu
pJ(5P)
hupJ
(5P)
hupI
hupI
hoxR
hoxR
hupR
hupI
hupI
rubr
edox
in-l
ike
hupJ
(3P)
hupJ
(3P)
hupJ
hupJ
hoxT
hoxT
hupT
hybE
hupJ
hupK
hupK
hupK
hupK
hoxV
hoxV
hupV
hupK
hypA
hypA
hypA
hypA
hypA
1hy
pA2h
hypA
hupA
hybF
hypA
Ni
inse
rtio
n,m
atur
atio
nhy
pBhy
pBhy
pBhy
pBhy
pB1
hypB
2hhy
pBhu
pBhy
pBG
TP
ase,
Ni
dono
ran
dst
orag
ehy
pFa
hypF
ahy
pFhy
pFhy
pF1
hypF
2hhy
pFhu
pYhy
pFm
CO
and
CN
synt
hesi
sp
hypC
hypC
hypC
hypC
hypC
hypC
hupC
hybG
lhy
pCl
chap
eron
e,m
atur
atio
nl
hypD
hypD
hypD
hypD
hypD
hypD
hupD
hypD
Fe/
Spr
otei
n,m
atur
atio
nq
hypE
hypE
hypE
hypE
hypE
hupE
hypE
mat
urat
ion
hypX
hoxX
hypX
mat
urat
ion
hupR
bhu
pRb
8ho
xAho
xAho
xAg
hydG
(zra
R)i
fhlA
hupR
nre
spon
sere
gula
tor
hupT
hoxB
g
hoxC
g
hoxJ
ghy
dH(z
raS
)i
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501458
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

unlike any of those of the two metal-containing classes ofH2ases. Structural data are expected to con¢rm these in-ferences in the near future.
Only the [NiFe]- and [Fe]-H2ases will be considered insome detail in this review. Indeed, given the paucity ofsequence data on metal-free H2ases, their phylogeny can-not be adequately discussed, and they therefore fall out-side the scope of this paper.
3. Biosynthesis and localization of hydrogenases
3.1. Cellular functions of hydrogenases
Although H2ases catalyze a very simple reaction, theydo so in many di¡erent metabolic contexts and thus as-
sume very diverse functions. In fermentative bacteria ofthe clostridial type, the reduction of protons into dihydro-gen by H2ases is a means of disposing of excess reducingequivalents [13]. Various microorganisms can use H2 as anelectron source either aerobically [1,2] or anaerobically(e.g. the methanogens [80,81,85], the sulfate reducers [12]and the photosynthetic bacteria [5,6]). Nitrogen ¢xers usu-ally contain uptake H2ases that recycle the H2 producedby nitrogenase [4^6,9,88^90]. These various functions areoften associated with di¡erent cellular localizations, e.g.hydrogen evolution is most often cytosolic, whereas hy-drogen uptake is usually periplasmic or membrane-local-ized. However, cytoplasmic bidirectional H2ases also me-diate H2 uptake. Some bacteria contain two or moredi¡erent H2ases, localized in di¡erent cell compartments.The multiplicity of H2ases in such organisms re£ects the
6Identi¢ed gene clusters are shown in the table. Homologous genes are found in other species but either they are scattered in the chromosome or theyhave not been completely sequenced yet. In each column the genes appear in the same order as in the hydrogenase gene clusters. Exceptions, which areindicated in footnotes, have been introduced by the constraint that homologous genes occur on the same line. The references are related either to thegenes (in italics) or to the properties of the gene products (written with a capital letter). (A) R. capsulatus H2ase gene cluster: hoxHORF2hupTUVhypF-hupSLCDFGHJKhypABhupRhypCDE. NB: hupT and hupR were initially termed hupR2 and hupR1, respectively. Ref. hupSL [93], hupC/ORFX [94] (thehupC gene initially termed ORFX was named hupM [95,96], then renamed hupC [23]), hupDFGHJK,hypABDE, hypF [96], C-terminal hupJ/ORF1, hupK/ORF2, hypB/ORF4, N-ter hupR/ORF5P [97], hupR/hupR1 [98], hupTU [99], hupTUV [100], HupT [101], HupUV [102], HupR [103], HupR, HupT [104],hypF, HypF, HupUV, HupSL [105], hoxH, HupUV, HupSL hypD [106]. (B) R. sphaeroides H2ase gene cluster: hupTUVhypFhupSLEC---HJKhypAB-hupRhypCD. Ref. hupTUV, hupS [107], hupTUVhypFhupSLEC (accession Y14197), hupHJKhypABhupRhypCD (accession AF214145), hupT (accessionAJ277115), hupR (accession AJ243734), hupDFG (http://www.jgi.doe.gov/tempweb/JGI_microbial/html/rhodobacter). (C) R. leguminosarum H2ase genecluster : hupSLCDEFGHIJKhypABFCDEXiihoxA. Ref. hupSL [108,109], hupCDEF [110], hupGHIJK [111], hypABFCDE [112], HupK [113], HypB [114],hypBFCDE [115], 8hoxA [116], hypX [117], hypA [118], see also accession X52974. (D) B. japonicum H2ase gene cluster: hupUVSLCDFGHIJKhyp-ABFCDEXhoxXAhupT. Ref. hupSL [119], hupCDFG [120], hypDP/incomplete sequence, hypE, hoxXA [121], hupUV [122], hupSLCDF operon [123], hup-GHIJK [124], hypA, hypB [125], HypB [126], HoxA [127], HoxX, HoxA [128], HypB [129], hypFCD [130], HupT [131], HypB/nickelin [132]. (E) R. eu-tropha MBH operon: hoxKGZMLOQRTVhypA1B1F1CDEXhoxABCJ ; SH operon: hoxFUYHWIhypA2B2F2. Ref. hoxFUYH [133], hoxA [134],hoxKGZMLOQRTV [135], hypABCDE [136], hypB1F1 [137] HoxHFUY [137], hoxX [138], HoxA [139], hoxKGZMLOQRTV [140], HoxZ [141], hoxW,HoxH, HoxW [142], hypA1B1F1, hypA2B2F2 [143], hoxABCJ [144],HoxFUYH [145], HoxA, HoxB, HoxC, HoxJ [146], hoxX/hypX [147], HoxH[148,149], hoxA, HoxA [150,151], HoxBC/RH [152]. (F) A. vinelandii H2ase gene cluster: hoxKGZMLOQRTVhypABFCDE. Ref. hoxKG [153], hoxZM-LOQ [154], ORF1-2/hoxRT [155] ; ORF3-8/hoxVhypABFCD [156], HoxZ [157], HoxG [158,159], HoxK [160], hypE [161], hypB, hoxVhypABFCD [162],HoxZ/cyt.b [163]. (G) A. chroococcum H2ase gene cluster: hupSLZMNOQRTVhupABYCDE. Ref. hupSL [164], hupDE [165], hupABYC [166], hupZM-NOQRTV [167]. (H) E. coli H2ase 1 gene cluster (hya operon): hyaABCDEF ; H2ase 2 gene cluster (hyb operon): hybOABCDEFG ; H2ase 3 gene cluster(hyc operon): hypABCDEfhlAhycABCDEFGHI; hypF. Ref. hydG, hydH [168] (it has recently been shown [372] that the two-component regulatory sys-tem encoded by the hydHG genes is not a speci¢c activator of H2ase 3 synthesis but rather regulates the synthesis of ZraP, a periplasmic protein in-volved in the tolerance to high zinc concentrations; accordingly, the authors [372] have proposed to replace hydH by zraS and hydG by zraR), hyaABC-DEF [169], hya operon [170], hydA/hypF [171], hypABCDE [172,173], hyb operon [174], hyc operon [175], hyc operon [176], HypB [177], HypB [178],HycE [179], hycI [180,187], hypF [181], HycE [182], hybO [183], HypC, HycE [184,185], HybD [186,187], HycI [187,188], HypF [194]. (I) T. roseopersici-na H2ase 2 gene cluster: hupSLCDHIR ; H2ase 1 structural genes: hydSL Ref. hupSLCDHI [189], hupR (accession L22980) [189], hydSL, HydSL [190],HydSL [373]. (J) P. hydrogenovora H2ase gene cluster: hupSLCDFGHIJK. Ref. hupSL [191,192], hupC [193].ahypF is located between hupV and hupS.bhupR is located between hypB and hypC.choxH is located upstream from hupT.dhupE is located between hupL and hupC.ehupT is located downstream from hoxA.f Located downstream from hoxA.gCorrect location of the gene in the cluster.hBelongs to the hoxFUYHWIhypA2B2F2 operon.iLocated elsewhere on the chromosome, actually not involved in H2ase synthesis, renamed zraR and zraS [372].jNot in that order in the gene cluster.kThe HybD and HycI maturation proteases have been biochemically characterized. The crystal structure of HybD is known [186].lHybG, chaperone for H2ase 1 and H2ase 2 (A. Bo«ck, private communication), HypC, speci¢c chaperone for H2ase3 maturation [184,185].mLocated elsewhere on the chromosome.nhupR is located immediately downstream from hupI.oThe hydS and hydL genes are separated by IS [190].p[194].qA. Bo«ck, private communication.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 459
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

importance of H2 in their metabolism and also ensures arapid and e¤cient response to variations in energetic needsunder changing growth conditions. Transmembrane hy-drogen redox cycling leading to formation of transmem-brane proton gradients has been suggested in sulfate re-ducers [91], but redundancy of H2ases is possible in somecases. The latter hypothesis is bolstered by the presence ofmultiple copies of H2ase-encoding genes in some genomes(see below).
Considering this complexity in function and localiza-tion, in addition to the structural sophistication of H2asesand their active sites, it is not surprising that many genesare required for the biosynthesis of these enzymes. In thecase of the [Fe]-H2ases such putative genes remain to beidenti¢ed. In contrast, the frequent genomic clustering ofthe genes involved in [NiFe]-H2ase synthesis and matura-tion has facilitated their identi¢cation [22,23,92].
3.2. Biosynthesis of [NiFe] active sites. Accessory genes
In Proteobacteria, the genes encoding the H2 uptakeH2ases are clustered, either on the chromosome or on amegaplasmid (cf. references in Table 1). The genes haveusually been labeled alphabetically in accordance withtheir order in the cluster (Table 1). The structural geneencoding the small subunit (28^35 kDa) has been namedS or K (`small' or `klein') (e.g. hupS, hoxK, hydS,) whilethe large subunit (45^70 kDa) has been designated L or G(`large' or `gross', e.g. hupL, hoxG, hydL). Downstreamfrom the structural genes are several genes (hup or hox),the products of which are involved in the maturation ofthe heterodimeric enzyme. Another set of proteins, en-coded by the hyp (`p' for pleiotropic) genes, is involvedin the insertion of Ni, Fe, CO and CN into the active site.Finally, these H2ase gene clusters also comprise regulatorygenes that control expression of the structural genes. Thefunctions of some of the numerous genes implicated in thebiosynthesis of [NiFe]-H2ases (reviewed in [92]) are brie£yrecalled hereafter, taking as reference the assembly systemassociated with H2ase 3 of Escherichia coli.
The synthesis of the NiFe active center in E. coli H2ase3 has been intensively studied by the group of A. Bo«ck inMunich, Germany. It was found that mutations mappingin the 58^59 min region of the E. coli chromosome a¡ectedthe synthesis of all three H2ases of this organism. Sequenc-ing that region revealed the presence of an operon with¢ve genes designated hypABCDE (for genes a¡ecting hy-drogenases pleiotropically) [172]. A sixth hyp gene, hypF(initially termed hydA [171]), was found in another loca-tion of the chromosome. Similar sets of genes were thenidenti¢ed in aerobic H2-oxidizing bacteria [22,23,28,92](Table 1), and later in fully sequenced genomes (Section6). The same hyp nomenclature was used for homologousgenes except for Azotobacter chroococcum in which the hyphomologs have been designated hupABYCDE [165^167].
Other genes not belonging to the hyp group are also
involved in H2ase maturation. For instance, the C-termi-nal cleavage of the large subunit of each of the threeH2ases from E. coli is performed by a di¡erent protease(Table 1). The recent determination of the crystal structureof HybD, the speci¢c endopeptidase involved in matura-tion of H2ase 2 from E. coli, has demonstrated that HybDis a metal-binding protein that uses nickel present in thelarge subunit precursor as a recognition and binding motif[186].
The roles of accessory gene products in the assembly of[NiFe]-H2ases are not yet fully elucidated, but consider-able progress has been made recently. The maturation ofthe [NiFe]-H2ase 3 of E. coli is initiated by the binding ofthe active site iron and its diatomic ligands. As describedbelow, proteolytic processing (endoproteolytic cleavage atthe C-terminus) of the large subunit is achieved only aftercorrect incorporation of Fe(CO)(CN)2 and Ni. It has beenshown [181] that such a proteolytic processing does notoccur in an E. coli mutant having a deletion in the hypFgene. Starting from the observation that the HypF proteincontains a sequence motif characteristic of o-carbamoyl-transferases, the group of Bo«ck has recently demonstratedthat carbamoylphosphate is required for synthesis of theactive center of [NiFe]-H2ases and suggested that it is thesource of the CO and CN ligands to the Fe atom [194]. Itis not yet known whether insertion of the diatomic ligandstakes place in the large subunit or on some sca¡old pro-tein ¢rst, with subsequent transfer to the large subunit[194]. HypX (HoxX), found in Rhizobium leguminosarum[117], Bradyrhizobium japonicum [128], and Ralstonia eu-tropha [147], which bears some similarity to formyl trans-ferase enzymes [117,195], has been considered a possiblealternative protein involved in the insertion of the dia-tomic ligands. No HypX homolog has been found in otherproteobacteria, including E. coli. Insertion of theFe(CO)(CN)2 unit is then followed by Ni insertion intothe partially unfolded protein. The precursor form of thelarge subunit (pre-HycE) containing both metals is subse-quently cleaved C-terminally by a speci¢c protease, HycIfor E. coli H2ase 3 [180,182,187,188,196]. This cleavagetriggers a conformation change resulting in the closureof the bridge between the two metals by the most C-ter-minally located cysteine residue (Cys534). It has been dem-onstrated that pre-HycE incorporates nickel while forminga complex with the chaperone-like protein HypC in theabsence of the small subunit. HypC has then to leavethe complex for the C-terminal processing of pre-HycEto occur, since only the HypC-free, nickel-containingform of pre-HycE is a substrate for the maturation endo-peptidase [184,185,197]. The cleavage position in the largesubunit, as determined biochemically, was found to occurC-terminal to a conserved histidine residue in Azotobactervinelandii [158,159], R. eutropha [142] and H2ases 1 and2 of E. coli [142,158,159,187], and C-terminal to similarlypositioned glutamine and arginine residues in Methano-coccus voltae FrhA and E. coli H2ase 3, respectively
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501460
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

[179,198]. HypD would play a role in maturation at a laterstage.
HypB is a GDP- and GTP-binding protein that exhibitsa low intrinsic GTPase activity [132,177] and binds Niatoms [114,129]. It has been hypothesized that HypB inthe GTP-bound state interacts with nickel-free pre-HycEand donates the nickel to it [199]. B. japonicum HypBbinds 18 nickel ions (per HypB dimer) and hence hasbeen termed nickelin for its role in nickel storage[129,132]. The dual roles of nickelin in nickel storageand GTP-dependent Ni mobilization can be separatedfunctionally and structurally and the two functions as-signed to di¡erent domains of the protein [132]. WhileHypB, HypD, HypE and HypF appear to act on the mat-uration of all three E. coli H2ases, the products of hypAand hypC would be involved only in the maturation ofH2ase 3 [173]. However, genes homologous to E. colihypA and hypC are present in other organisms (Table 1).In Helicobacter pylori, which contains a membrane-bounduptake H2ase coupled to a cytochrome-dependent respira-tory chain, mutations of the hypA or hypB genes a¡ectboth hydrogenase and urease activities. The deleteriouse¡ects of these mutations can partially be overcome byadding nickel to the culture medium [132,200].
Additional genes (not shown in Table 1) are requiredfor the transport of Ni ions necessary for Ni-con-taining enzymes [201]. The Ni2� transporters activelystudied in connection with H2ase biosynthesis are theNik system of E. coli and the high a¤nity Ni2�-speci¢cpermease encoded by hoxN of R. eutropha (reviewed in[202]).
3.3. Biosynthesis of [Fe] active sites
No accessory genes involved in the biosynthesis of [Fe]-H2ases have been formally identi¢ed yet. The main di¤-culty is that, unlike in the case of most [NiFe]-H2ases, the[Fe]-H2ase-encoding operons consist of structural genesonly. A recently discovered exception occurs in the ge-nome of Thermotoga maritima [203], where the genes en-coding the trimeric H2ase [204] are part of an operonincluding eight open reading frames (ORFs, Section 6).However, the roles of these additional genes remain tobe elucidated. Biochemical and, if possible, genetic inves-tigations of this system are warranted. Some idiosyncrasiescommon to the [NiFe]- and [Fe]-H2ases may also providevaluable information. For instance, since both types ofactive sites contain CO and CN metal ligands, it mightbe expected that similar proteins be involved in the pro-duction and insertion of these ligands in both cases. Onthe other hand, given the apparently independent evolu-tion of the [Fe]- and [NiFe]-H2ases, these two classes ofenzymes may well possess quite di¡erent maturationmechanisms. Indeed, analysis of the genome from T. ma-ritima, which appears to encode [Fe]-H2ases only, has sofar failed to reveal any genes similar to those known to
participate in [NiFe]-H2ase maturation in other organisms(our unpublished results, see Section 6). In any event, un-like the [NiFe] enzymes, [Fe]-H2ases do not require thenumerous genes committed to Ni transport, transfer, andinsertion. Neither do they undergo C-terminal cleavage oftheir catalytic subunit, except for some periplasmic en-zymes. The latter process has been evidenced in Desulfovi-brio desulfuricans [35,205] and may be associated withperiplasmic export (see Section 3.6) rather than assemblyof the catalytic metal site [35]. Thus, despite their uniquerequirement for a dithiolate bridging ligand [35], [Fe]-H2ases might require a lesser number of genes for theirmaturation than the [NiFe] ones.
3.4. Biosynthesis of Fe^S clusters
Pioneering studies have been carried out on the geneproducts involved in nitrogenase biosynthesis, whichhave proved to be of general relevance to Fe^S proteins([206], [32] and references therein). While up to 10 proteinsmay be required, the best known ones are the NifU-likeand NifS-like proteins, which are involved in Fe(S) andsul¢de mobilization, respectively [32,207]. The subcellularlocalization of these processes in eukarya is currently thefocus of active research [207,208]. It is likely, but not dem-onstrated, that accessory Fe^S centers in H2ases are as-sembled in similar ways. Whether assembly of the catalyticsites of H2ases also involves the general Fe^S pathway,together with the speci¢c hyp gene products, remains tobe established.
3.5. Other accessory genes and regulatory genes
The H2ase gene clusters of Proteobacteria contain othergenes besides the structural ones and those discussed inSection 3.2 (Table 1). These include several regulatorygenes, such as hupUV [100,122]/hoxBC [144] which encodean H2-sensing H2ase [102,106,146] also called regulatoryH2ase [152], as well as genes encoding two-componentregulatory systems. One of the components is a histidinekinase (termed HupT [99,101,104,131] or HoxJ [144,146]),the other one is an NtrC-like transcription factor (termedHupR [98,103,104] or HoxA [127,128,139]). BesidesRhodobacter capsulatus, R. eutropha, B. japonicum andE. coli (Table 1), the hyperthermophile Aquifex aeolicusalso appears to contain such sets of regulatory genes([209] and Section 6). HupE is more narrowly distributed(Table 1) and is an homolog of ureJ which encodesa nickel transport protein involved in the assembly ofurease [210]. The hupE gene has been identi¢ed in thehup cluster of R. leguminosarum and Rhodobacter sphaer-oides (Table 1) and seems also to be located downstreamfrom the hupSL genes of Rubrivivax gelatinosus [211](formerly Rhodopseudomonas gelatinosa and Rhodocyclusgelatinosus) (GenBank accession X52522, our unpublishedresults).
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 461
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

3.6. Signal peptides and subcellular localization ofhydrogenases
The distinction between soluble intracytoplasmic H2asesand membrane-bound or periplasmic or organellar H2asesis made at the genetic level. The protein to be exported tothe periplasm or imported into an organelle is tagged by asignal peptide at the N-terminus of one of the subunits.
The [NiFe]-H2ases are minimally heterodimers com-posed of a small and a large subunit, of which the encod-ing genes follow each other in that order (Table 1). Themembrane-targeted H2ases are characterized by the pres-ence of a long signal peptide (30^70 amino acid residues)at the N-terminus of the small subunit. This signal peptidecontains a conserved RRxFxK motif [20,21] recognizableby a speci¢c protein translocation pathway known as themembrane targeting and translocation (Mtt) [212] or twin-arginine translocation (Tat) [213] pathway, by which thecorrectly folded and fully active dimer can cross the mem-brane [214^216]. E. coli H2ase 1 [213,217] and H2ase 2[213,217^219], as well as the membrane-bound H2ases ofWolinella succinogenes [220] and R. eutropha [221], havebeen demonstrated to use this so-called hitchhiker modelof cotranslocation of the two subunits. This should be truefor all the hydrogenases listed in Fig. 1, since all the or-
ganisms of Fig. 1 are expected to contain the tat genes.The R. capsulatus genome indeed contains the tat genes(L.-F. Wu, unpublished results). Thus, the presence of thetwin-arginine signature at the N-terminus of the precursorform of the small subunit is an additional characteristic ofthis family of H2ases (Group 1 in our classi¢cation, seeSection 4). Some of the H2ases having the leader peptideand belonging to this family have not been annotated assuch in the databases. The hupS gene from Clostridiumacetobutylicum borne on plasmid pSol1 (Table 2) is theonly sequence of Group 1 lacking the twin-arginine motif.This may have to do with the fact that in C. acetobutyli-cum the HupSL H2ase is cytoplasmic.
It is inferred that [NiFe]-H2ases missing the signal pep-tide at the N-terminus of the small subunit remain in thesoluble cytoplasmic compartment. This has been demon-strated to be the case for the dimeric H2-sensing H2asesHupUV of R. capsulatus [106] and HoxBC of R. eutropha[152], and for multimeric bidirectional H2ases (see below).
A number of [Fe]-H2ases are cytosolic and are thereforedevoid of signal sequences. These include enzymes fromclostridia, e.g. Clostridium pasteurianum [272], and Mega-sphaera elsdenii [273], as well as the tetrameric NADP-reducing H2ase from Desulfovibrio fructosovorans [274].H2ases from anaerobic eukaryotes not containing hydro-
Fig. 1. Twin-arginine motif in signal peptides that function in [NiFe] and [Fe] hydrogenase transport. The signal peptides at the N-terminus of smallsubunits of [NiFe]-H2ases belonging to Group 1 are aligned up to the proteolytic cleavage site. The predicted length of the signal peptide indicated atthe right includes the ¢rst translated methionine residue. When species names are followed by `P', the genes are plasmid-encoded. The Twin-argininemodule is shown in reverse shading.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501462
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

Table 2Structural genes encoding the catalytic subunits of [NiFe]-hydrogenases
Organism Group Gene name GenBankaccession(protein)a
Totalsubunitsb
Locali-zationc
Accession andprotein name inSwissProta
Reference
Acetomicrobium £avidum DSM20663 3d hydS CAA56463 4 U Q59113 Cytc3S [222]hydL CAA56464 Q59114 Cytc3L
Alcaligenes hydrogenophilus strain M50 2b hoxB AAB49360 C P94154 HoxB [144]hoxC AAB49361 2 P94155 HoxC
Alcaligenes hydrogenophilus strain M50 1 hupS AAB25779 3 M P33375 HupS [223]hupL AAB25780 P33374 HupL
Anabaena sp. PCC7120 2a hupS AAC79877 2 M Q44215 HupS [224]hupL AAC79878 Q44216 HupL
Anabaena sp. PCC7120 3d hoxY CyanoBased
hoxHAnabaena variabilis ATCC 29413 2a hupS CAA73658 2 M Q9ZAK3 HupS [225]
hupL CAA73659 Q9ZAK2 HupLAnabaena variabilis ATCC 29413 3d hoxY CAA55876 4 C Q44515 HoxY [226]
hoxH CAA55878 Q44517 HoxHAnabaena variabilis strain IAM M58 3d hoxY BAB39386 unpublishedAquifex aeolicus 1 mbhS1 AAC06862 U O66894 MbhS1 [209]
mbhL1 AAC06861 O66895 MbhL1Aquifex aeolicus 1 mbhS2 AAC07047 U O67095 MbhS2 [209]
mbhL2 AAC07046 O67092MbhL2Aquifex aeolicus 2 mbhS3 AAC06946 U O66987MbhS3 [209]
mbhL3 AAC06045 O66988MbhL3Archaeoglobus fulgidus strain VC-16 1 vhtG AAB89863 U O28890 VhtG [227]
vhtA AAB89864 O28891 VhtAArchaeoglobus fulgidus strain VC-16 3c vhuG AAB89871 U O28898 VhuG [227]
vhuA AAB89872 O28899 VhuAAzotobacter chroococcum strain MCD1 1 hupS CAA37133 3 M P18190 HupS [164]
hupL CAA37134 P18191 HupLAzotobacter vinelandii strain OP 1 hoxK AAA82505 3 M P21950 HoxK [153]
hoxG AAA82506 P21949 HoxGBradyrhizobium japonicum strainUSDA 110
2b hupU AAA62627 2 C Q45254 HupU [122]hupV AAA62628 Q45255 HupV
Bradyrhizobium japonicum strainUSDA 122
1 hupS AAA26218 3 M P12635 HupS [119,123]hupL AAA26219 P12636 HupL
Campylobacter jejuni NCTC 11168 1 hydA CAB73521 U Q9PN31 HydA [228]hydB CAB73520 Q9PN32 HydB
Campylobacter jejuni NCTC 11168 1 hydA2e CAB73823 U Q9PMQ8 HydA2e [228]Citrobacter freundii 1 hyaA BAA05929 U Q46045 HyaA unpublished
hyaB BAA05930 Q46046 HyaB Acc D28594Clostridium acetobutylicum ATCC824(pSOL1 megaplasmid)
1 hupS 2 U unpublishedf
hupLDesul¢tobacterium dehalogenanshalorespiration-de¢cient mutant HRD6
1 hydA AAF13046 U Q9RP13 HRD4-1 [229]hydB AAF13047 Q9RP12 PITH
Desulfomicrobium baculatus DSM1743(Desulfovibrio baculatus)
1 PhsS (hysB)g AAA23376 2 P P13063 PhsS [230,231]PhsL (hysA)g AAA23375 P13065 PhsL
Desulfovibrio desulfuricans ATCC 27774 1 hydA 2 P13061 [232](small subunit) AAF43137 Q9L869 unpublished(large subunit) AAF43138 Q9L868
Desulfovibrio fructosovorans 1 hynB AAA23371 2 P P18187h HydA [233]hynA AAA23372 P18188 HydB
Desulfovibrio gigas 1 hynB AAA23377i 2 P P12943 HydA [33,230]hynA AAA23378i P12944 HydB [234]
Desulfovibrio vulgaris Miyazaki F 1 hynB AAA23369 2 P P21853 HydA [235]hynA AAA23370 P21852 HydB
Escherichia coli strain K12 1 hyaA AAA23997 3 M P19928 HyaA [169]hyaB AAA23998 P19927 HyaB [170,236](H2ase 1)
Escherichia coli strain K12 1 hybO AAC76033 3 M Q46847 HybO [174,183]hybC AAA21591 P37181 HybC [236](H2ase 2)
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 463
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

Table 2 (continued)
Organism Group Gene name GenBankaccession(protein)a
Totalsubunitsb
Locali-zationc
Accession andprotein name inSwissProta
Reference
Escherichia coli strain K12 4 hycG CAA35552 6 M/C P16433 HycG [175,176]hycE CAA35550 P16431 HycE [179,236](H2ase 3)
Escherichia coli strain K12 4 hyfI AAB88571 U P77668 HyfI [236,237]hyfG AAB88569 P77329 HyfG(H2ase 4)
Helicobacter pylori strain J99(Campylobacter pylori)
1 hyaA AAD06157 U Q9ZLK5 HyaA [239]hyaB AAD06147 Q9ZLK4 HyaB
Helicobacter pylori ATCC 700392 = strain26695 (C. pylori)
1 hydA AAD07691 U O25348 HydA [238]hydB AAD07692 O25349 HydB
Methanobacterium thermoautotrophicumstrain vH
3a frhG AAA72189 3 C P19498 FrhG [240,241]frhA AAA72187 P19496 FrhA
Methanobacterium thermoautotrophicumstrain vH
3c mvhG1 AAB02350 3 C Q50782 MvhG1 [241,242]mvhA1 AAB02351 Q50783 MvhA1
Methanobacterium thermoautotrophicumstrain vH
3c mvhG2 AAB85624 U O27207 MvhG2 [241]mvhA2 AAB85623 O27206 MvhA2
Methanothermobacter marburgensis(M. thermoautotrophicum strain Marburg)DSM 2133
4 ehaN CAB52769 s 6 M Q9UXP5 EhaN [243]ehaO CAB52770 Q9UXP4 EhaO
Methanothermobacter marburgensis(M. thermoautotrophicum strain Marburg)DSM 2133
4 ehbM CAB52790 s 6 M Q9V2X8 EhbM [243]ehbN CAB52791 Q9V2X7 EhbN
Methanococcus jannaschii DSM2661 4 ehaN (CooL) AAB98505 s 6 M Q57936 [243,244]ehaO (CooH) AAB98504 Q57935
Methanococcus jannaschii DSM2661 4 ehbM AAB99371 s 6 M Q58758 [243,244]ehbN AAB99031 Q58433
Methanococcus jannaschii DSM2661 3a frhG AAB98010 4 U Q60340 FrhG [244]frhA Q60338 FrhA
Methanococcus jannaschii DSM2661 3c vhuG AAB99193 U Q58591 VhuG [244]vhuA AAB99194 Q58592 VhuA
Methanococcus voltae strain PS(DSM1537)
3a frcG CAA43498 3 Q00393 FRHG [245,246]frcA CAA43496 Q00390 FRHA
Methanococcus voltae strain PS(DSM1537)
3a fruG CAA43502 3 M Q00397 FruG [245,246]fruA CAA43500 Q00394 FruA [247]
Methanococcus voltae strain PS(DSM1537)
3c vhcG CAA43505 3 Q00406 VhcG [245,246]vhcA CAA43506 Q00404 VhcA
Methanococcus voltae strain PS(DSM1537)
3c vhuG CAA43509 3 Q00409 VhuG [245,246]vhuA CAA43510 Q00407 VhuA
Methanosarcina barkeri strain Fusaro(DSM804)
3a frhG1 CAA74096 4 O33163 FrhG [248,249]frhA1 CAA74094 O33161 FrhA [250]
Methanosarcina barkeri strain Fusaro(DSM804)
3a frhG2 CAA74092 4 O33167 FrhG [249]frhA2 CAA74090 033165 FrhA
Methanosarcina barkeri strain Fusaro(DSM804)
4 echC CAA76119 6 M O59654 EchC [59,360]echE CAA76121 O59656 EchE
Methanosarcina mazei strain Go«1 1 vhoG CAA58113 3 M Q50248 VhoG [251]vhoA CAA58114 Q50249 VhoA
Methanosarcina mazei strain Go«1 1 vhtG CAA58176 3 M Q50225 VhtG [251]vhtA CAA58177 Q50226 VhtA
Methanothermus fervidus 3c mvhG AAA72831 3 Q49178 MvhGj [252]mvhA AAA72832 Q49179 MvhA
Nostoc muscorum hoxY not sequenced 4 C [253]hoxH
Nostoc punctiforme. (Nostoc sp.PCC 73102)
2a hupS AAC16276 2 C O68307 HupLhk [254]hupL AAC16277 O68306 HupShk
Oligotropha carboxidovorans DSM 1227(plasmid pHCG3) (Pseudomonascarboxydovorans)
1 hoxS CAA72673 3 M O33405 HoxS [255]hoxL CAA72674 O33406 HoxL
Prochlorothrix hollandicastrain ACC 15-2
3d hoxY AAB53703 4 C O05930 HoxY unpublishedhoxH AAB53705 O05932 HoxH Acc U88400
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501464
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

Table 2 (continued)
Organism Group Gene name GenBankaccession(protein)a
Totalsubunitsb
Locali-zationc
Accession andprotein name inSwissProta
Reference
Pseudomonas hydrogenovora strain 9-5 1 hupS BAA13222 3 M Q51860 HupS [191]hupL BAA13223 Q51862 HupL
Pyrococcus abyssi strain Orsay 3b none CAB49780 U Q9V0C4 Cytc3D unpublishedl
none CAB49779 Q9V0C5Cytc3APyrococcus abyssi strain Orsay 3b hydD2 CAB49862 U Q9V044 Cytc3D unpublishedl
hydA2 CAB49863 Q9V043 Cytc3APyrococcus abyssi strain Orsay 4 cooL-like CAB49637 U Q9V0R6 CooL-like unpublishedl
cooH-like CAB49635 Q9V0R8 CooH-likePyrococcus furiosus DSM 3638 4 mbhJ (mbh10)m;n s 6 M [256,257]
mbhL (mbh12)m;o
Pyrococcus furiosus DSM 3638 4 mbxJ p s 6 M [257]mbxLq
Pyrococcus furiosus DSM 3638 3b hydD CAA53036 4 C Q59669 cytc3 H2ase [258]hydA CAA53037 Q59670 cytc3 H2ase(sulfhydrogenase I)
Pyrococcus furiosus DSM 3638 3b shyD2 AAF61853 4 C Q9P9M5 ShyD [259]shyA2 AAF61854 Q9P9M4 ShyA(sulfhydrogenase II)
Pyrococcus horikoshii strain OT3 3b PH1292 BAA30395 U O59013 Cytc3D [260]PH1294 BAA30397 O59011 Cytc3A
Ralstonia eutropha strain H16(ATCC 17699) (pHG1 megaplasmid)(A. eutrophus)
2b hoxB AAB49364 2 C P95603 HoxB [146,134]hoxC AAB49365 P95604 HoxC
Ralstonia eutropha strain H16(ATCC 17699) (pHG1 megaplasmid)(A. eutrophus)
1 hoxK AAA16461 3 M P31892 HoxK [135]hoxG AAA16462 P31891 HoxG
Ralstonia eutropha strain H16(ATCC 17699) (pHG1 megaplasmid)(A. eutrophus)
3d hoxY AAC06142 4 C P22319 HoxY [133]hoxH P22320 HoxH
Rhizobium leguminosarum strainsUPM791 and B10 (pSym megaplasmid)
1 hupS (hupA) CAA37148,CAA85430
3 M P18637 HupS(HupA)
[108,109]
hupL (hupB) CAA37149,CAA85431
P18636 HupL(HupB)
[261]
Rhodobacter capsulatus strain B10(Rhodopseudomonas capsulata)
3d hoxH AAD38065 Q9XBW8 HoxH [106]
Rhodobacter capsulatus strain B10(R. capsulata)
2b hupU AAC32032 2 C Q52695 HupU [100]hupV AAC32033 O86457 HupV
Rhodobacter capsulatus strain B10(R. capsulata)
1 hupS CAA31869 3 M P15283 HupS [93]hupL CAA31870 P15284 HupL
Rhodobacter sphaeroides strain 2.4.1.(Rhodopseudomonas sphaeroides)
2b hupU1 AAA99490 2 C Q53163 HupU1 [107]Q53164 HupU2or HupV
Rhodobacter sphaeroides strain RV(R. sphaeroides)
2b hupU CAA74584 2 C O86466 HupU unpublishedhupV CAA74585 Acc. Y14197
Rhodobacter sphaeroides strain RV(R. sphaeroides)
1 hupS CAA74586 3 M O86467 HupS [107], unpublishedhupL CAA74587 O86468 HupL Acc. Y14197
Rhodococcus opacus MR11(plasmid pHG201) (N. opaca)
3d hoxY AAB57891 4 C P72306 HoxY [262,263]hoxH AAB57892 P72307 HoxH
Rubrivivax gelatinosus (R. gelatinosus) 1 hupS CAA36754 2r M HupS [211]hupL CAA36755 HupL
Rhodospirillum rubrum strains UR1and UR2
4 cooL AAC45118 6 M P72317 CooL [264,265]cooH AAC45121 P31895 CooH
Synechococcus sp. PCC6301(A. nidulans)
3d hoxY CAA66382 4 C P94158 HoxY [226,266]hoxH CAA66383 P94159 HoxH [267]
Synechocystis sp. PCC6803 3d hoxY CAA66211 4 C P74021 HoxY [268,269]BAA18094
hoxH CAA66212 P74018 HoxHBAA18091
Thermococcus litoralis DSM 4573 3b hydD AAB94935 4 C Q9UWQ7 HydD [270]hydA AAB94936 Q9UWQ HydA
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 465
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

genosomes, e.g. Entamoeba histolytica, are also cytosolic[275]. On the other hand, in those organisms that containhydrogenosomes, as in Trichomonas vaginalis, H2ases ap-pear to be located in these organelles. For instance, twogenes (hydA and hydB) from T. vaginalis encoding twoshort monomeric (ca. 50 kDa) [Fe]-H2ases have been se-quenced (see Section 5) and found to include a hydroge-nosomal-targeting presequence [276]. Likewise, a gene en-coding a 64-kDa monomeric H2ase in the same organismdisplays an alanine- and serine-rich N-terminal extensionwith the hydrogenosomal-targeting RN motif [277] at po-sitions 14^15 [275]. The [Fe]-H2ase-encoding genes fromthe green algae Scenedesmus obliquus [72] Chlamydomonasreinhardtii, and Chlorella fusca (T. Happe, personal com-munication; see Section 5) comprise signal sequences con-sistent with chloroplastic localization. The N-terminal se-quence of these H2ases is similar to stroma-targettingtransit peptides (rich in basic and hydroxylated residues),and is cleaved by a stromal peptidase at a conserved Val^X^Ala site [375,376]. In the case of the dimeric periplasmic[Fe]-H2ases, e.g. D. vulgaris, the gene encoding the largesubunit precedes the one encoding the small subunit andthe latter possesses a tat-type signal sequence [216,278](Fig. 1). Sequence and 3D structure comparisons haveshown that the small subunit of periplasmic [Fe]-H2asesis homologous to the C-terminal region of monomeric
cytoplasmic H2ases, and that the splitting of the geneand introduction of the signal sequence have been con-comitant [35]. In these enzymes the large subunit includesa C-terminal extension that is cleaved o¡ post-translation-ally [42,205] (see also Section 3.3). Whether this cleavage isconnected with export to the periplasm has remained anopen question so far.
4. [NiFe]-hydrogenases
4.1. The basic enzyme is a heterodimer
The ¢rst Ni-containing H2ases were isolated as KL het-erodimers with the large (K) and small (L) subunits havingaverage masses of 60 and 30 kDa, respectively[12,18,22,23,57]. The X-ray structure of the [NiFe]-H2asesfrom Desulfovibrio gigas [33] and Desulfovibrio vulgaris[37] showed that the two H2ase subunits interact very ex-tensively through a large contact surface and form a glob-ular heterodimer. The bimetallic NiFe center of the activesite is located in the large subunit and is deeply buriedinside the protein. The small subunit contains up to threeFe^S clusters, which conduct electrons between the H2-activating center and the physiological electron acceptoror donor of H2ase. The [4Fe^4S] cluster that is proximal
Table 2 (continued)
Organism Group Gene name GenBankaccession(protein)a
Totalsubunitsb
Locali-zationc
Accession andprotein name inSwissProta
Reference
Thiocapsa roseopersicina strain BBS 1 hydS AAC38281 2 P O51820 HydS [190,373]hydL (H2ase 1)s AAC38282 O51823 HydL
Thiocapsa roseopersicina strain BBS 1 hupS AAA27409 3 M Q56359 HupS [189]hupL (H2ase 2) AAA27410 Q56360HupL
Wolinella succinogenes 1 hydA CAA46302 3 M P31884 HydA [271]hydB CAA46303 P31883 HydB
Present nomenclature as found in databases and literature.aThe protein sequences in the database entries are usually those of the precursors.bDeduced from operon sequence or protein characterization.cDeduced from signal sequences or biochemical studies. C: cytoplasm; P: periplasm; M: membrane; U: unknown.dCyanoBase: http://www.kazusa.or.jp/cyano/anabaena/.ePutative small subunit of second hydrogenase.f The hupSL genes are located on the pSOL1 megaplasmid (L. Fontaine personal communication) which has been fully sequenced (accessionNC_001988).gThe names given in parentheses are those used in the literature. hys is for selenocysteine-containing hydrogenase.hP18187 gives the protein sequence without the signal peptide.iThis entry contains an erroneous sequence [234] that has been corrected later [33,230].jPutative small subunit.kHupLh, HupSh: HupL, HupS homologs.lGenoscope: http://www.genoscope.cns.fr.mSubunits named MbhJ and MbhL in [257] and Mbh10 and MBh12 in [256].nEntry PF_1342770 in genome sequence (http://comb5-156.umbi.umd.edu/).oEntry PF_1344050 in genome sequence (http://comb5-156.umbi.umd.edu/).pEntry PF_1352206 in genome sequence (http://comb5-156.umbi.umd.edu/).qEntry PF_1351020 in genome sequence (http://comb5-156.umbi.umd.edu/).rThe gene encoding the putative third subunit (HupC) was not found immediately downstream from hupL [211].sThis enzyme has been isolated in a dimeric soluble form [373]. However, the presence of two additional ORF [190] one of which encodes a putativetransmembrane protein suggests that this H2ase might be tetrameric and membrane-associated in vivo. These remarks are likewise applicable to theMbhS2L2 H2ase from A. aeolicus (see Table 6).
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501466
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

to the active site is `essential' to H2 activation in [NiFe]-H2ases [33,57,279]. Hydrophobic channels expandingthrough both subunits have been identi¢ed by crystallo-graphic analysis of xenon binding and molecular dynamicssimulations of xenon and H2 di¡usion within the interiorof the enzyme [280]. Those channels linking the active siteto the surface of the molecule were suggested to facilitategas access to the active site [279,280]. Finally, phylogeneticanalyses have shown that the two subunits of [NiFe]-H2ases have evolved conjointly (see Section 7.1)
[NiFe]-H2ases are encoded by multicistronic operonsalso comprising genes encoding polypeptides involved inelectron transfer, but the basic structural unit is the KLheterodimer. The original nomenclature of this KL cata-lytic unit as identi¢ed in Archaea and Bacteria is given inTable 2. In order to eliminate confusing designations andbe more consistent with amino acid sequence similaritiesand phylogenetic relationships (see Sections 7 and 8), newnames are proposed in Table 3 for some of the genes listedin Table 2. We propose to reserve hyd for the [Fe]-H2asesand more speci¢cally to use hydA exclusively for genesencoding catalytic subunits of [Fe]-H2ases (see Sections 5and 8), and to use preferentially the letters S and L for thesmall and the large subunits, respectively, of [NiFe]-H2ases. Thus, we suggest to replace hydA by hynS andhydB by hynL for the [NiFe]-H2ases from Campylobacterjejuni, D. desulfuricans, Desul¢tobacterium dehalogenans,Thiocapasa roseopersicina and W. succinogenes (Table 3).We propose to rename HynSL the [NiFe]-H2ase from D.desulfuricans, in keeping with the designation of the ho-mologous enzyme from D. vulgaris, and in order to clearly
distinguish it from the [Fe]-H2ase present in the same or-ganism [35]. For the two H. pylori strains (Table 2), wesuggest to replace hyaAB and hydAB by hynSL. The[NiFe]-H2ase of Acetomicrobium £avidum should betermed HoxYH, for it clearly belongs to this family (Sec-tion 7.1). The two cytoplasmic H2ases (I and II) fromPyrococcus furiosus can reduce So in vitro [259,281] andwere called sulfhydrogenases (Shy) [258,281]. However,this appears not to be a physiological reaction as bothenzymes are repressed when cells are grown in the pres-ence of sulfur [374]. In addition, these enzymes are likelyto be regulated by other metabolites (M.W.W. Adams,personal communication), so the Shy name is confusingand out of date. It is therefore proposed to rename Hyh(`h' for hyperthermophile) the enzymes formerly known asShy (Table 3).
4.2. Biodiversity of [NiFe]-H2ases
In their review, Wu and Mandrand [21] divided the[NiFe]-H2ases into four classes. Since then the numberof known sequences has increased 3-fold but the classi¢-cation of [NiFe]-H2ases as proposed here remains roughlythe same, in spite of the addition of recently identi¢edH2ases such as the H2-sensing regulatory H2ases[100,102,106,122,144,146,152] and the so-called sulfhydro-genases [258,259,281].
Sequence alignments of large subunits of [NiFe]-H2asesreveal two very conserved regions surrounding the twopairs of cysteine ligands of the NiFe site, near the N-and C-terminus of the sequence [18,20,21]. These L1 and
Table 3List of new gene names used in the phylogenetic trees of [NiFe]-H2ases
Species New proposed gene names Former gene namesa
small subunit large subunit small subunit large subunit
A. £avidum hoxY hoxH hydS hydLA. aeolicus hupS hupL mbhS1 mbhL1A. aeolicus hynS hynL mbhS2 mbhL2C. jejuni hynS hynL hydA hydBC. jejuni hynS2 hydA2D. desulfuricans hynS hynL hydA hydBD. dehalogenans hynS hynL hydA hydBD. fructosovorans hynS hynL hynB hynAD. gigas hynS hynL hynB hynAD. vulgaris hynS hynL hynB hynAH. pylori-J99 hynS hynL hyaA hyaBH. pylori-26695 hynS hynL hydA hydBP. abyssi echC echE cooL-like cooH-likeP. abyssi hyhS1 hyhL1 cytc3D cytc3AP. abyssi hyhS2 hyhL2 hydD2 hydA2P. horikoshii hyhS hyhL cytc3N cytc3KP. furiosus hyhS1 hyhL1 hydN hydKP. furiosus hyhS2 hyhL2 shyD2 shyA2R. sphaeroides hupU hupV hupU1 hupU2T. litoralis hyhS hyhL hydD hydAT. roseopersicina hynS hynL hydS hydLW. succinogenes hynS hynL hydA hydB
aSee Table 2 for references.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 467
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

L2 patterns [282] have been retrieved and aligned for all ofthe sequences now available. These patterns de¢ne groupsof [NiFe]-H2ases (Fig. 2) which are in very good agree-ment with the groups derived from cellular functions (Sec-tion 3) and full sequence alignments (Section 7). In mostcases the L2 signature ends at a histidine residue which isthe endopeptidase cleavage site at the C-terminus of thelarge subunit. A typical feature of Group 4 H2ases is that,as in HycE [187], there is an arginine residue at the posi-tion of the conserved histidine (Fig. 2).
4.2.1. Membrane-associated respiratory uptake[NiFe]-hydrogenases (Group 1)
The so-called respiratory uptake H2ases are those en-zymes capable of supporting growth with H2 as an energysource. The respiratory H2ases of Group 1 transfer elec-trons from H2 to a cytochrome that is bound to a mem-brane-located complex coupling electron transfer to trans-membrane proton translocation.
4.2.1.1. Membrane-bound, periplasmically oriented hy-drogenases. The H2-uptake [NiFe]-H2ases are the most
numerous and best documented. They are attached tothe membrane and connected to the electron transportchain. H2 oxidation is linked to reduction of various elec-tron acceptors such as O2, NO3
3 , SO234 , fumarate or CO2.
The electrons are transferred from H2 to the quinone poolof the membrane via a cytochrome b encoded by the thirdgene of the structural operon (Table 1), e.g. to fumarate orinorganic oxidants in W. succinogenes [271] or to oxygenin R. eutropha [141]. Electron transfer in the respiratorychain is coupled to vectorial proton translocation acrossthe membrane with the establishment of an electrochem-ical proton gradient (vWH� ) as a means of energy recovery[284^287].
The third subunit (cytochrome b) is not merely a redoxcarrier but also an anchor for the binding of the uptakehydrogenase to the membrane. Directed by the signal pep-tide present at the N-terminus of the small subunit, whichcontains the conserved ^RRxFxK^ motif (twin-arginineelement, Fig. 1), the completely folded KL heterodimer istranslocated through the cytoplasmic membrane by theMtt/Tat translocation pathway (cf. Section 3.6). It remainsattached to the periplasmic side of the membrane by the
Fig. 2. Characterization of [NiFe]-H2ase groups. L1 and L2 signatures. Inferred [NiFe]-H2ase amino acid sequence of each group shown in the evolu-tionary trees (Fig. 6) were aligned using Clustal W [283]. The sequences including the cysteine residues liganding the Ni atom in the large subunit formthe so-called L1 and L2 signatures [282]. The sequences which shared less than 20% identity with the other members of the group or were incompleteor too short (e.g. D. dehalogenans HydL) were excluded from the pattern determination (Section 7.1). Brackets include the residues occurring at a singleposition in the set of sequences. X was used when more than three di¡erent residues were found.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501468
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

cytochrome b anchor protein (HupC/HoxZ/HupZ/HyaC inTable 1) and a C-terminal hydrophobic extension of thesmall subunit [95,141,287]. Thus, the oxidation of H2 re-sults in a release of protons in the periplasmic compartmentof Gram-negative bacteria. The integral membrane cyto-chrome b (third subunit) was also shown to be necessaryfor growth on H2 of R. capsulatus cells [95] and for fullheterodimer-catalyzed H2 oxidation in A. vinelandii [163].
In nitrogen-¢xing bacteria, uptake [NiFe]-H2ases are in-duced when nitrogenase is synthesized and produces mo-lecular hydrogen (an intrinsic property of nitrogenase). Byrecycling the hydrogen produced by nitrogenase, H2aserecoups energy which otherwise would be lost by thecell. This reaction was observed more than 20 years agoin A. chroococcum [90], in R. capsulatus [5] and in cyano-bacteria [88,89]. Uptake [NiFe]-H2ases are induced togeth-er with nitrogenase in Proteobacteria (e.g. in R. legumino-sarum [116], in T. roseopersicina [189], in R. capsulatus[289]) and in cyanobacteria (e.g. in Nostoc [253,290] andin Anabaena variabilis [291]). The control is exerted at thetranscriptional level [116,253,289]. In R. leguminosarum,both the H2ase- and nitrogenase-encoding genes are con-trolled by the nitrogen ¢xation regulatory protein NifA[116]. In R. capsulatus, the global RegB/RegA regulatorysystem, which responds to the redox status of the cell,controls the transcription of the hydrogenase hupSL genesand the regulatory nifA gene [289]. R. capsulatus also usesan additional regulatory system involving the speci¢c H2-sensing HupUV hydrogenase to regulate the synthesis ofthe respiratory HupSL H2ase (see below).
These membrane-bound uptake [NiFe]-H2ases are tri-meric respiratory enzymes; they typically transfer elec-trons from H2 to the quinone pool of electron transportchains via a b-type cytochrome and are linked to a chem-iosmotic mechanism for energy conservation.
4.2.1.2. Periplasmic soluble hydrogenases of sulfate re-ducers. Anaerobic sulfate-reducing bacteria of the genusDesulfovibrio contain three types of H2ases, namely[NiFe]-, [NiFe(Se)]- and [Fe]-H2ases, localized either inthe periplasm, in the membrane or in the cytoplasm [12].Several Desulfovibrio species contain periplasmic low-po-tential c-type cytochromes, in particular the most abun-dant tetraheme cytochrome c3, and a high-molecular masscytochrome with 16 c-type hemes (HmcA). The latter ispart of the transmembrane Hmc complex that comprisesfour integral membrane proteins containing Fe^S clustersor b-type heme. Cytochrome c3 was suggested to be theredox partner of periplasmic H2ases [292] and to mediatereduction of exogenous metallic cations [293]. The Hmccomplex was proposed to catalyze electron transfer linkingperiplasmic H2 oxidation to the cytoplasmic sulfate reduc-tion pathway [294]. Complexes between c-type cyto-chromes and periplasmic Desulfovibrio H2ases have beenisolated [295,296]. Studies of electron transfer between hy-
drogenases and mono- and multiheme cytochromes in De-sulfovibrio species [297], including Desulfomicrobium norve-gicum [296], have shown that catalytic amounts ofcytochrome c3 increase the rate of electron transfer from[Fe]- or [NiFe]-H2ases to HmcA. It has been proposedthat the shuttling of electrons by cytochrome c3 fromH2ases to various other polyheme cytochromes (Hmc, anine-heme cytochrome c, a new member of the Hmc fam-ily [298], and an octaheme cytochrome c3) is a generalmechanism in Desulfovibrio species [296,297]. However, acytochrome c3 mutant of D. desulfuricans strain G20 wasimpaired in the oxidation of pyruvate by sulfate but couldgrow normally on H2 and sulfate [299]. The use of a mu-tant deleted in the hmc operon has recently demonstratedthe importance of the Hmc complex in electron transportfrom hydrogen to sulfate in D. vulgaris subsp. vulgarisHildenborough [300]. However, Hmc does not appear tobe the only complex capable of coupling electron transportto proton pumping since the cell yield per mol of sulfatereduced was similar for the wild-type strain and for theHmc3 mutant [300]. A complementary mode of energyconservation might consist in cycling hydrogen betweena cytoplasmic H2-evolving H2ase and a periplasmic H2-consuming H2ase, thus leading to the build up of a pro-ton-motive force [11,91].
In Desulfovibrio, periplasmic H2ases transfer electronsfrom H2 to low-potential c-type cytochromes. H2 oxida-tion by these H2ases establishes a proton gradient acrossthe membrane for energy conservation but all the redoxpartners involved have not yet been identi¢ed.
4.2.1.3. Membrane-bound archaeal uptake hydrogena-ses. Group 1 also includes archaeal enzymes with tat-containing signal peptide. For the reduction of CO2 tomethane, the membrane-bound hydrogenase VhoGA ofthe methanogenic archeon Methanosarcina mazei Go«1transfers electrons from H2 to a cytochrome b (encodedby vhoC) ; the electrons are then channeled through meth-anophenazine to heterodisul¢de reductase. The latter re-duces the CoenzymeM^S^S^CoenzymeB heterodisul¢de.CoenzymeB^SH is the reductant for the formation ofmethane from methyl^S^CoM. This electron transfer sys-tem, called H2 : heterodisul¢de oxidoreductase, is coupledto the creation of a proton-motive force [288], an energy-conserving system analogous to the respiration-linked oxi-dative phosphorylation occurring in Proteobacteria ofGroup 1.
4.2.2. Cytoplasmic heterodimeric[NiFe]-hydrogenases(Group 2)
These [NiFe]-H2ases include the cyanobacterial uptakeH2ases (Group 2a) and the cytoplasmic soluble H2asesinvolved in H2 sensing (Group 2b). They all lack the mem-brane-targeting signal peptide at the N-terminus of thesmall subunit (see Section 3.6).
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 469
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

4.2.2.1. H2-uptake hydrogenases of cyanobacteria(Group 2a). Uptake HupSL H2ases have been found inall N2-¢xing unicellular and ¢lamentous cyanobacteria ex-amined so far [3,4,25,27]. The hupSL genes of Nostoc andAnabaena are induced under N2-¢xing conditions[224,225,254,291] and in Anabaena sp. strain PCC7120are expressed only in N2-¢xing heterocysts [224]. In Nostocmuscorum, hupSL transcription is followed by the appear-ance of an in vivo light-dependent H2 uptake activity[253]. Mutational analyses have shown that the HupSLuptake H2ase of A. variabilis reoxidizes at high rate thehydrogen produced by nitrogenase [225].
The sequences of cyanobacterial hupSL genes are highlyconserved (s 90% identity) and therefore the L1 and L2signatures for Group 2a include only totally conservedresidues (Fig. 2). They are nevertheless provided in Fig.2 to point out the di¡erences with the enzymes of Group2b. In particular, a histidine residue present at the end ofL1 in uptake H2ases (Group 2a) is replaced by a glutamineresidue in H2-sensing H2ases (Group 2b). Although theyful¢l the same function as other uptake H2ases of Group1, the enzymes of Group 2a appear to be more closelyrelated to the H2-sensing H2ases of Group 2b. They alsodi¡er from the enzymes of Group 1 by deletions through-out the sequences of the hupL genes. The deletions occur-ring in the genes of Group 2a are also present in the genesof Group 2b. The latter display additional deletions thatmake them even shorter. The enzymes of both Group 2aand Group 2b are characterized by the absence of a signalpeptide at the N-terminus of the small subunit, in keepingwith their cytoplasmic localization. It is commonly ac-cepted that cyanobacterial uptake H2ases are bound to amembrane. They are probably localized on the cytoplas-mic side of the cytoplasmic membrane or of the thylakoidmembrane [25,27].
4.2.2.2. H2-signaling hydrogenases (Group 2b). TheH2-sensing H2ases have been shown in B. japonicum[122], R. capsulatus [100] and R. eutropha [144] to be in-volved in H2ase gene regulation in response to H2, togeth-er with other regulatory proteins expressed from theH2ase gene clusters (Table 1). Molecular hydrogen is de-tected by the H2-sensing HupUV/HoxBC H2ase whichtransduces the signal to a histidine kinase (HupT/HoxJ)[99,101,131,146] which, together with a response regulator(HupR/HoxA) [103,104,127,150,151] forms a two-compo-nent regulatory system required for induction by H2. Thepeculiarity of this system is that the response regulator(transcription factor) activates hupSL/hoxKG gene tran-scription in the non-phosphorylated form [104,131,146]. Thus the histidine kinase exerts a negative con-trol in the absence of H2 [99,101,131,146], and the re-sponse regulator exerts a positive control in the presenceof H2 [103,104,127,150,151]. They act as antagonistic part-ners of a two-component H2ase-synthesis regulatory sys-tem.
In the aerobic bacteria R. eutropha and B. japonicum,the HoxBC/HupUV H2 sensor is necessary for the syn-thesis of uptake H2ase. Deletions in hoxB or hoxC of R.eutropha [146] or inactivation of hupUV in B. japonicum[122] prevent H2ase synthesis. In the presence of H2, theH2-sensing H2ase (HoxBC) releases the negative e¡ect ofHoxJ [146], possibly by preventing HoxJ phosphorylationand hence allowing the response regulator (HoxA) to re-main non-phosphorylated and active. In the facultativeanaerobic bacterium R. capsulatus, deletion of the hupUVgenes leads to constitutively increased hupSL gene expres-sion [100], which indicates that the HupUV H2 sensor,besides HupT, exerts a negative control on H2ase geneexpression in absence of H2. Under these conditions, theR. capsulatus H2 sensor appears to promote autophos-phorylation of the histidine kinase HupT and conse-quently the inactivation of HupR by phosphorylation. Inthe presence of H2, in all three bacteria the response reg-ulator (HupR/HoxA) remains in the non-phosphorylatedactive form and activates hupSL/hoxKG gene transcription[104,131,146]. The above-mentioned dissimilarities in thefunctioning of the H2 sensors may re£ect di¡erences ininteractions with the cognate histidine kinases. The mech-anism of transcription activation may also di¡er since inR. capsulatus HupR binds to a c70-dependent promoter[104] while in R. eutropha HoxA binds to a c54-dependentpromoter [150].
The H2-sensing H2ases (Group 2b) are characterized byspeci¢c sequence features. The small subunit of these H2-sensing HupUV/HoxBC regulatory H2ases lacks the signalpeptide for export across the membrane [122,100,144] (seeSection 3.6) and the C-terminal hydrophobic motif [100]capable of anchoring the protein to the membrane[21,141,153,287]. Accordingly, the regulatory H2ase is lo-cated in the cytoplasm [106,152]. The large subunit(HupV/HoxC) is characterized by an L1 motif ending bya glutamine instead of a histidine (Fig. 2). It lacks the C-terminal extension which is cleaved after Ni insertion (andthus ends at the last His residue of the L2 motif), butnevertheless contains a NiFe bimetallic center with theCO and CN ligands [47,152,301]. The hypF gene productis required for the formation of the CO and CN ligands[194]. The lack of HupSL H2ase activity and of HupUVH2ase activity in HypF-minus mutants of R. capsulatus[105] suggests that HypF has a function in R. capsulatussimilar to that in E. coli. The R. capsulatus HypF3 mu-tants are hence relieved from the negative control exertedby HupUV on hupSL gene expression (the expression of ahupS: :lacZ gene fusion was increased in the HypF3 mu-tants and independent from H2) [96,105]. These resultswere reported in an article [105] of which the title hasunfortunately been misinterpreted. This has resulted inthe erroneous annotation of HypF as a transcriptionfactor in several genome sequences (Table 4). It is stressedhere that HypF is not a transcription factor but maycontribute to the transcriptional control of hydrogenase
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501470
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

biosynthesis by acting on H2-sensors of the HupUV-type.
In keeping with its regulatory function, the speci¢c ac-tivity of this regulatory HupUV H2ase is very low. It isinsensitive to pH, as measured by the hydrogen^deuterium(H/D) exchange reaction. These results would suggest arestricted access of the protons to the active site [106]. Ithas also been observed that the H/D exchange reactioncatalyzed by HupUV is not inhibited by oxygen or acety-lene, in contrast with the HupSL-catalyzed H/D exchangereaction. This indicates that gas access to the NiFe centerof HupUV may be restricted as well [106].
The putative H2ase 3 (MbhS3L3) from A. aeolicus [209]displays the characteristic sequence features of Group 2(see Sections 6 and 7.1), but cannot be clearly assignedto either one of the subgroups 2a and 2b. However, themere fact that two other [NiFe]-H2ases are encoded bythis genome suggests that mbhS3L3 encodes an H2-sensingH2ase.
The H2-signaling H2ases are cytoplasmic heterodimericenzymes that do not provide energy to the cell. Instead,they detect H2 and together with HupT/HoxJ histidinekinases act as H2-signal transducers by controlling thephosphorylation state of HupR/HoxA transcription acti-vators. It remains to be understood how the H2-signalingH2ase communicates with the cognate histidine kinaseand, in the presence of H2, prevents the HupR/HoxA re-sponse regulator from being inactivated by phosphoryla-tion.
4.2.3. Cytoplasmic heteromultimeric reversible [NiFe]-hydrogenases (Group 3)
The [NiFe]-H2ases of Group 3 include subunits able tobind a soluble cofactor, e.g. F420, NAD or NADP andfunction reversibly. It will be interesting to see whetherthis is also the case for the Vhc/Vhu archaeal H2ases ofGroup 3c once their physiological electron acceptor(s)/do-nor(s) are identi¢ed.
4.2.3.1. F420-reducing hydrogenases of methanogens(Group 3a). Methanogenic archaea are strictly anaerobicorganisms. They can be divided into two major groups onthe basis of the substrates utilized: the hydrogenotrophicmethanogens, which use only H2+CO2 or formate as sub-strates and the methylotrophic methanogens of the orderMethanosarcinales, which utilize simple C1 componentssuch as methanol and methylamines (some can alsogrow on H2+CO2 or acetate). The major di¡erence be-tween the two groups is that the former organisms donot contain cytochromes (cf. [302] for a review). The re-duction of CO2 to methane involves a central cytosolicelectron carrier known as cofactor 420 (F420, 8-hydroxy-5-deaza£avin) [80]. The H2ases from methanogens that areable to reduce F420 are encoded by the frh, frc or fru genes.Those that do not reduce F420 can instead deliver electronsto arti¢cial acceptors such as viologens, hence the mvh, vhuand vhc names (Table 2).
In Methanosarcinales, reducing equivalents are pro-vided by membrane-bound electron transport systems us-ing H2 or F420H2 as electron donors. Electron transferinvolves H2ases, b-type cytochromes and F420H2 dehydro-genases, and is coupled to proton translocation across thecytoplasmic membrane, which results in the formation of aproton-motive force. The F420H2 dehydrogenase of M.mazei, encoded by the fpo genes, is a redox-driven protonpump functionally, and to some extent structurally, ho-mologous to the proton-translocating NADH:quinoneoxidoreductase of respiratory chains ([303] ; reviewed in[302]; Section 7.3).
In the hydrogenotrophic methanogen M. voltae growingon formate, F420H2 generated by formate dehydrogenaseis the only electron source for the reduction of CO2. Bro-dersen et al. [247] have recently demonstrated that in M.voltae, F420H2 oxidation is catalyzed by a membrane-asso-ciated H2ase practically identical to the selenium-contain-ing F420-reducing H2ase encoded by the fruADGB operon[245,246]. A distinct F420H2 dehydrogenase similar to the
Table 4Erroneous annotations of HypF
Functional annotationsa Accession/organism
Transcriptional regulatory protein AAC75754/E. coli ; AAC06863/A. aeolicus ; AAC06379/M. mazei ;CAB75258/C. jejuni ; BAA29991/P. horikoshii ; BAA10154/Synechocystis sp.A71079/P. horikoshii ; O66902b/A. aeolicus ; Q9PHP5b/C. jejuni
H2ase expression regulatory protein HypF AAB98708/M. jannaschii ; AAB89876/A. fulgidusTranscriptional regulator HypF homolog AAB85769/M. thermoautotrophicum
C69038/M. thermoautotrophicum vH; B70359/A. aeolicusDNA binding, transcription regulation:Superfamily: probable transcription regulator HypF S32948/R. capsulatusInvolved in the regulation of H2ase (by similarity) Q60190b/M. mazeiHydrogenase expression/formation regulatory protein CAB50145/P. abyssi ; AAB98706/A. fulgidus
D75031/P. abyssi ; E69420/A. fulgidus ; Q58123b/M. jannaschii
aThe correct annotation should be: `H2ase maturation protein HypF', as for P30131/E. coli.bSwissProt entries ; all others are GenBank entries.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 471
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

one present in methylotrophic methanogens was notfound. Genes capable of encoding a F420H2 dehydroge-nase appear to be absent in Methanococcus jannaschii, aclosely related organism [247].
The F420-reducing H2ases are trimeric enzymes. ThefruADGB operon encodes the large (FruA) and the small(FruG) subunit of the basic catalytic unit. The FruB sub-unit is only found in these H2ases, it probably binds F420.The product of fruD is a protease [246].
4.2.3.2. Tetrameric bifunctional hydrogenases of hyper-thermophiles (Group 3b). The cytoplasmic H2ase of theanaerobic hyperthermophilic archaeon P. furiosus was ini-tially found to have sulfur reductase activity [281]. Both So
and polysul¢de could serve as substrates for H2S produc-tion. The So-reducing and the H2-oxidizing activitiesshowed di¡erent responses to pH, temperature and inhib-itors. This bifunctional enzyme able to dispose of theexcess reductant generated during fermentation, usingeither polysul¢des or protons as electron acceptors, wastherefore called `sulfhydrogenase' [258,281]. CytoplasmicH2ases from other fermentative, So-reducing hyperthermo-philes such as the archaeon Thermococcus litoralis, thebacterium T. maritima, were also found to be able to re-duce So to H2S [281]. It was then suggested by Ma et al.[281] that all such H2ases could share that ability. Thephysiological electron donor to P. furiosus sulfhydroge-nase is NADPH [304].
It is remarkable that the enzymes of Group 3b are allfrom hyperthermophiles, i.e. organisms able to grow opti-mally above 80³C [16,377].
The enzymes of Group 3b are tetrameric. Their K and Nsubunits constitute the dimeric [NiFe]-H2ase moiety. Thetwo other subunits (L and Q) are also well conserved. Theenzyme from P. furiosus can indeed reduce So in vitro[259,281] but appears to be repressed in cells grown inthe presence of sulfur [374]. It is now likely that metabo-lites other than sulfur can regulate this H2ase (M.W.W.Adams, personal communication). In view of the fact thatthe regulations of this type of enzyme are still under in-vestigation, it is preferable to use names unrelated to func-tion. It is therefore proposed to replace the name Shy bythe more general term Hyh (`h' for hyperthermophile, sug-gestion of M.W.W. Adams), with L and S for the largeand small subunits, for the homogeneous Group 3b ofarcheal enzymes (Table 3, Fig. 6).
4.2.3.3. The methyl viologen-reducing hydrogenases(Group 3c). Nothing is known presently on the functionof these H2ases that are merely characterized by their abil-ity to reduce methyl viologen.
4.2.3.4. The bidirectional NAD-linked hydrogenases(Group 3d). A soluble tetrameric H2ase able to revers-ibly reduce NAD with H2 was ¢rst isolated from R. eutro-pha (formerly Alcaligenes eutrophus) [305]. The NAD-
linked H2ase is a heterotetramer composed of two dimericmoieties. One of these, encoded by the hoxU and hoxFgenes, has diaphorase activity, is homologous to two sub-units of Complex I (see Section 7.3) and contains NAD,FMN and Fe^S-binding sites. The second moiety is en-coded by the hoxY and hoxH genes and is a [NiFe]-H2ase[133]. Genes encoding a homologous NAD-reducingH2ase have been isolated from Rhodococcus opacus (for-merly Nocardia opaca) [262]. Site-directed mutagenesis andin vitro reconstitution of subforms showed that the twomoieties can function independently and that the diaphor-ase part contributes to the stabilization of the R. eutrophaenzyme [145].
Cyanobacteria contain two types of H2ases [3,4], anuptake H2ase induced under N2-¢xing conditions and areversible or bidirectional H2ase able to either produceor oxidize H2. The latter is believed to be constitutivelysynthesized [306,307] and hence present in N2-¢xing andnon-N2-¢xing cyanobacteria. The hoxFUYH genes, encod-ing the bidirectional H2ase in the ¢lamentous heterocys-tous A. variabilis, were isolated and sequenced in 1995[226]. Similar genes were identi¢ed in the unicellular cya-nobacteria Synechocystis sp. strain PCC 6803 [268] andAnacystis nidulans ( = Synechococcus PCC 6301) [266,308]. Very recently an hoxH gene has also been detectedin Chroococcidiopsis thermalis CALU 758 [309]. The geneproducts are homologous to the soluble NAD-linkedH2ase of R. eutropha.
In cyanobacteria, the bidirectional H2ase catalyzes H2
uptake in a strongly light-stimulated way. The enzyme ofthe unicellular A. nidulans seems to be associated with thecytoplasmic membrane [306] whereas in A. variabilis anassociation with the thylakoid region was reported [307].Since the respiratory Complex I of cyanobacteria lacks thesubunits homologous to the diaphorase moiety of the bi-directional H2ase, it was suggested that the bidirectionalH2ase could be linked to Complex I [25,267,268,310].However, a survey of H2ase distribution in cyanobacteriashowed that the bidirectional enzyme is absent in a signi¢-cant set of strains [311,312]. This dismisses the hypothesisof a common use of the diaphorase subunits by the bidi-rectional H2ase and Complex I in cyanobacteria [312,313].Appel et al. [314] have shown recently that the bidirec-tional H2ase of Synechocystis sp. PCC 6803 works as anelectron valve for the disposal of low-potential electronsgenerated at the onset of illumination. Indeed, an hoxHmutant was impaired in the oxidation of photosystem I(PS I) upon illumination exciting either PS I alone or bothphotosystems. Isolation of membranes from Synechocystisrevealed an association of the H2ase large subunit with thethylakoid membrane [314]. If the bidirectional H2ase de-pends on photosynthesis, then there would be no need forsuch an enzyme in symbiotic cyanobacteria like Nostocpunctiforme (Nostoc sp. PCC 73102) [312], which live inassociation with roots [315].
Bidirectional [NiFe]-H2ases are tetrameric enzymes,
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501472
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

soluble or loosely attached to membranes. They can dis-sociate into two dimeric moieties, a diaphorase encoded byhoxFU with NAD, FMN and Fe^S-binding sites, and aheterodimeric hydrogenase encoded by hoxYH.
4.2.4. The membrane-associated H2-evolving respiratory[NiFe]-hydrogenases (Group 4)
H2ase 3 from E. coli was the ¢rst extensively studiedenzyme of this group. It is part of the formate^hydrogenlyase complex (FHL) expressed only under strictly anaer-obic conditions. This complex couples formate oxidationby formate dehydrogenase H (Fdh-H) to the reduction ofprotons by H2ase 3. E. coli H2ase 3 is a [NiFe]-H2asewhich was considered until recently to produce molecularhydrogen without energy recovery (in contrast to E. coliH2ases 1 and 2).
The eight genes encoding formate^hydrogen lyase be-long to the hyc operon. The catalytic H2ase subunits (Hyc-GE) are extrinsic membrane proteins (cytoplasmic), whilethe other subunits are intrinsic membrane proteins thatfunction as membrane anchors and electron carriers be-tween formate dehydrogenase and H2ase 3. The productsof ¢ve of the hyc genes share similarities with subunits ofNADH-ubiquinone oxidoreductase (Complex I, see Sec-tion 7.3). The large H2ase subunit, HycE, and the NuoDsubunit of Complex I have several sequence motifs incommon. The N-terminal part (ca. 170 residues) of thesmall subunit HycG is homologous to NuoB, whileHycF and HycC are homologous to NuoI and NuoL,respectively (Section 7.3).
In 1997, Andrews et al. [237] reported on the occurrencein E. coli of a 12-gene operon, hyf, capable of encoding aputative ten-subunit H2ase complex (hydrogenase four orHyf), homologous to H2ase 3. Two gene products of thehyf operon, HyfC and HyfF, are related to two transmem-brane subunits of Complex I, NuoH (ND1) and NuoL(ND5), respectively. These subunits have a crucial role inproton translocation and energy coupling in Complex I(Section 7.3). The authors proposed that Hyf, in conjunc-tion with Fdh-H, forms a respiration-linked proton trans-locating FHL pathway (FHL-2), which di¡ers from thenon-energy transducing hyc-encoded FHL [237].
Another member of Group 4 is the CO-induced H2aseof Rhodospirillum rubrum, which is encoded by the cooLHgenes [264,265]. This enzyme is a component of the CO-oxidizing system that allows R. rubrum to grow in a CO-dependent manner in the dark. CO dehydrogenase togeth-er with CooLH H2ase catalyze oxidation of CO to CO2
with H2 production. The CO oxidation/H2 evolutionenzyme complex is membrane bound. As R. rubrum canuse CO as the sole energy source, the oxidation of COmust be coupled to energy conservation. Since CO dehy-drogenase is a peripheral membrane protein, the CooLHH2ase has been proposed to be the energy coupling site[264,265].
E. coli H2ase 3 and R. rubrum CooLH H2ase are labile
enzymes, and the exact number of their subunits remainsto be determined.
Recently, [NiFe]-H2ases of the E. coli H2ase 3-type havebeen discovered in Archaea, ¢rst in Methanosarcina bar-keri [59] and M. thermoautotrophicum [243], then in thehyperthermophilic archeon P. furiosus [256,257]. Theyare hexameric enzymes and function under strict anaerobicconditions.
The Ech H2ase from M. barkeri is encoded by theechABCDEF operon [59]. It has been puri¢ed from ace-tate-grown cells, and has been proposed to oxidize thecarbonyl group of acetate to form CO2 and H2 [59,360].
M. thermoautotrophicum is a methanogenic archaeonthat can only grow on CO2/H2 as carbon and energysource [81,85]. Besides F420-reducing and F420-non-reduc-ing H2ases (Table 2), this organism contains two genegroups designated `energy converting H2ase A' (eha) and`energy converting H2ase B' (ehb), which encode putativemultisubunit membrane-bound H2ases similar to theabove-mentioned H2ases of Group 4 [243].
The anaerobic archaeon P. furiosus is an obligate orga-notroph of which the fermentative metabolism producesmainly acetate, H2 and CO2. This organism contains twocytoplasmic H2-evolving H2ases (I and II) [258,259] andwas recently shown to contain a membrane-bound H2ase.The genes proposed to encode the small (mbh10/mbhJ) andthe large (mbh12/mbhL) H2ase subunits belong to a puta-tive 14-gene operon (termed mbh, either mbh1-14 [256] ormbhA-N [257]). Besides the catalytic H2ase subunits, theoperon can encode four proteins that resemble products ofthe ech operon from M. barkeri [59,360], the coo operonfrom R. rubrum [264], the hyc [176] and the hyf [237] op-erons from E. coli, and share similarities with subunits ofComplex I [256,257]. The P. furiosus genome contains an-other putative operon, mbhxABCDFGHHPMJKLN, en-coding a multisubunit transmembrane complex homolo-gous to the mbh operon [257]. The MbhJL H2asecatalyzes H2 evolution rather than H2 uptake (the ratioof H2 evolution to H2 uptake is 250:1 at pH 7.4) in aDCCD-sensitive reaction [257]. The MbhM subunit is sim-ilar to NuoH/ND1 and has been proposed to hold theDCCD-binding site [257]. The last ORF of the operon(mbh14/mbhN) encodes a 2[4Fe^4S] protein which maybe involved in electron transfer to and from the H2ase[256,257].
The sequence features of Group 4 H2ases have beensummarized by Tersteegen and Hedderich [243]. TheH2ase small subunit of this group of enzymes is consider-ably smaller than that of other [NiFe]-H2ases (Fig. 2) andcontains only the cysteine ligands for the proximal [4Fe^4S] cluster. Two additional Fe^S clusters may be providedby a soluble 2[4Fe^4S] protein found in all multimericH2ase complexes. The large subunit is characterized bythe presence of an arginine residue, and not a histidineresidue, at the endopeptidase cleavage site, as ¢rst shownfor HycE [187]. Arginine is present at this position even
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 473
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

Tab
le5
Stru
ctur
alge
nes
enco
ding
cata
lyti
csu
buni
tsof
[Fe]
-hyd
roge
nase
s
Org
anis
mG
ene
nam
eP
ropo
sed
chan
gesa
Gen
Ban
kac
cess
ion
(pro
tein
)Si
zeb
Tot
alsu
buni
tsc
Loc
aliz
atio
ndR
efer
ence
[Fe]
-H2as
esC
lost
ridi
umac
etob
utyl
icum
P26
2hy
dAA
AA
8578
557
41
C[3
16]
Clo
stri
dium
acet
obut
ylic
umA
TC
C82
4hy
dAA
AB
0372
358
21
C[3
17]
Clo
stri
dium
di¤
cile
AT
CC
stra
in63
0no
nehn
dAno
nee
593
(p)
3C
unpu
blis
hed
Clo
stri
dium
di¤
cile
AT
CC
stra
in63
0no
nehy
dA1
none
e45
8(p
)1
Cun
publ
ishe
dC
lost
ridi
umdi
¤ci
leA
TC
Cst
rain
630
none
hydA
2no
nee
496
(p)
1C
unpu
blis
hed
Clo
stri
dium
di¤
cile
AT
CC
stra
in63
0no
nehy
dA3
none
e49
8(p
)1
Cun
publ
ishe
dC
lost
ridi
umdi
¤ci
leA
TC
Cst
rain
630
none
hydA
4no
nee
478
(p)
1C
unpu
blis
hed
Clo
stri
dium
past
euri
anum
AT
CC
6013
hydA
AA
A23
248
574
1C
[272
]C
lost
ridi
umpe
rfri
ngen
sN
CT
C82
37hy
dAB
AA
7472
657
21
C[3
18]
Clo
stri
dium
perf
ring
ens
stra
in13
hydA
BA
A74
738
572
1C
unpu
blis
hed
Clo
stri
dium
ther
moc
ellu
mA
TC
C27
405
hydA
AA
D33
071
579
1C
unpu
blis
hedf
Meg
asph
aera
elsd
enii
AT
CC
2594
0hy
dAA
AF
2211
448
41
C[2
73]
Des
ulfo
vibr
iode
sulf
uric
ans
AT
CC
7757
hydA
none
g42
1*2
Ph
[35]
Des
ulfo
vibr
iode
sulf
uric
ans
G20
hydA
AA
K11
625
421*
2P
hun
publ
ishe
dD
esul
fovi
brio
fruc
toso
vora
nsD
SM36
04hn
dDhn
dAi
AA
A87
057
585
4C
[274
]D
esul
fovi
brio
fruc
toso
vora
nsD
SM36
04hy
dAC
AA
7242
342
1*2
Ph
[319
]D
esul
fovi
brio
vulg
aris
Hild
enbo
roug
hhy
dAhy
dA1
CA
A26
266
421*
2P
h[2
78]
Des
ulfo
vibr
iovu
lgar
isH
ilden
boro
ugh
hydC
hydA
2C
AA
4097
060
6(p
)1
C[3
20]
Des
ulfo
vibr
iovu
lgar
isox
amic
usM
onti
cello
hydA
AA
A23
373
421*
2P
h[3
21]
The
rmot
oga
mar
itim
aM
SB8
hndD
hndA
2A
AD
3529
360
8(p
)4
C[2
03]
The
rmot
oga
mar
itim
aM
SB8
and
DSM
3109
hydA
hndA
1A
AD
3649
664
53
C[2
03,2
04]
The
rmot
oga
mar
itim
aM
SB8
TM
1421
jA
AD
3649
130
1(p
)n.
d.k
n.d.
k[2
03]
Chl
amyd
omon
asre
inha
rdti
iC
9hy
d1hy
dAA
AG
0059
149
7**l
1C
HL
unpu
blis
hed
Chl
orel
lafu
sca
UT
EX
251
hydA
AJ2
9822
743
6**m
1C
HL
unpu
blis
hed
Sce
nede
smus
obliq
uus
Ku «t
zing
276-
6hy
dAA
J271
546n
448*
*o1
CH
L[7
2]E
ntam
oeba
hist
olyt
ica
HM
-1:I
MSS
none
hydA
AA
G09
783p
468
1n.
d.k
unpu
blis
hed
Nyc
toth
erus
oval
isno
nehn
dAC
AA
7637
312
06(p
)**
1H
[322
]S
piro
nucl
eus
bark
hanu
sA
TC
C50
380
hydA
AA
G31
038
467
(p)
1n.
d.k
[275
]T
rich
omon
asva
gina
lisA
TC
C30
01T
vhyd
Ahy
dA1
AA
C47
159
468*
*1
H[2
76]
Tri
chom
onas
vagi
nalis
AT
CC
3001
Tvh
ydB
hydA
2A
AC
4716
044
9**
1H
[276
]T
rich
omon
asva
gina
lisno
nehy
dA3
AA
G31
037
589
(p)*
*1
H[2
75]
[Fe]
-H2a
se-l
ikeq
Klu
yver
omyc
esla
ctis
2359
/152
LE
T1
CA
A49
833
469
(p)
unpu
blis
hed
Sac
char
omyc
esce
revi
siae
S288
CN
ar1
NP
_014
159
491
(p)
[323
]S
chiz
osac
char
omyc
espo
mbe
972h
hyd
CA
B40
177
538
(p)
unpu
blis
hed
Cae
norh
abdi
tis
eleg
ans
Bri
stol
N2
hyd
AA
F60
782
569
(p)
unpu
blis
hed
Dro
soph
ilam
elan
ogas
ter
hyd
AA
F45
404r
241
(p)
[324
]H
omo
sapi
ens
(kid
ney)
hydK
BA
B15
261r
374
(p)
unpu
blis
hed
Hom
osa
pien
s(h
epat
oma)
hydH
BA
B15
199
476
(p)
unpu
blis
hed
Hom
osa
pien
s(H
eLa)
Nar
fA
AD
5144
645
6[3
25]
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501474
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

when the C-terminal peptide is missing, as in EchE of M.barkeri [59] or in CooH of R. rubrum [265] (Fig. 2). Thesemultimeric membrane-bound H2ase complexes are re-markable by their similarities with subunits of ComplexI and in particular with transmembrane subunits involvedin proton pumping and energy coupling (Section 7.3).These membrane-bound H2ases appear to be able to cou-ple the oxidation of a carbonyl group (originating fromformate, acetate or carbon monoxide) with the reductionof protons to H2. This results in energy recovery by thecreation of a proton-motive force across the membrane.The acronym Ech, which was initially meant for E. coliH2ase 3-type [59], may thus be accepted with the moregeneral meaning of `energy-converting hydrogenase' (R.Thauer, personal communication).
5. [Fe]-hydrogenases
5.1. The catalytic subunit
Unlike [NiFe]-H2ases, which are composed of at leasttwo subunits, many [Fe]-H2ases are monomeric and con-sist of the catalytic subunit only. However, dimeric, tri-meric and tetrameric enzymes are also known (see Section5.2). The catalytic subunits of [Fe]-H2ases (Table 5), incontrast to those of the [NiFe] enzymes, display consider-able variations in size. Indeed, in addition to a conserveddomain of ca. 350 residues containing the active site(H-cluster [14,34,35]), they most often comprise addi-tional domains which accommodate Fe^S clusters and
which may altogether consist of more than 800 residues(Fig. 3).
5.1.1. The H-cluster domainThe ¢rst [Fe]-H2ase sequences [272,278,320] revealed
modular structures composed of previously known do-mains and of a unique and very conserved C-terminaldomain. The latter was therefore assumed to contain theH-cluster, and was subsequently found to be present in all[Fe]-H2ase sequences ([72,273] and references therein). 3Dstructures have con¢rmed the modular structure of [Fe]-H2ases, and revealed the unique protein fold of the H-cluster domain, its high conservation, and its unprece-dented metal cluster. The latter consists of a binucleariron site bound to a [4Fe^4S] cluster by a bridging cysteine[34,35]. Totally conserved amino acids include the fourcysteine ligands of the active site, a few residues liningthe active site cavity, as well as those suspected to beinvolved in the H2 and H� channels connecting the buriedactive site to the surface of the protein [35,42]. Sequencealignments show that the most conserved parts of the H-cluster domains are three segments encompassing the fourcysteine ligands of the metal site (Fig. 4). The smallest[Fe]-H2ases (ca. 45^48 kDa) have been found in greenalgae [66,67]. The recently disclosed sequences of the en-zymes from S. obliquus [72], C. reinhardtii, and C. fusca(Table 5) consist of an H-cluster domain only. Accordingto the available structural models [34,35], the absence ofaccessory domains would signi¢cantly reduce the distancefrom the H-cluster to the protein surface. However, the C.reinhardtii sequence includes an insertion which is absent
6aIt is suggested that all catalytic subunits be named hydA, with numbers in addition for multiple copies in the same organism. hydB genes encode thesmall subunits of dimeric enzymes. The hnd denomination, which has been introduced for the NADP-dependent H2ase from D. fructosovorans [274], isproposed to be applied to all NAD(P)(H)-dependent enzymes (see Section 8).bNumber of encoded amino acid residues in precursor. p: putative protein. *C-terminal processing results in the removal of 24 residues [35,205].**N-terminal processing.cDeduced from operon sequence or protein characterization.dDeduced from signal sequences or biochemical studies. C: cytoplasm; P: periplasm; H: hydrogenosome; CHL: chloroplast.eUnpublished sequences retrieved from the Sanger Institute web site (http://www.sanger.ac.uk/Projects/C_di¤cile) using the TBLASTN [341] programwith H-domain sequences as baits.f Although an entry corresponding to this sequence (AF148212) has been annotated as `partial', the translated protein sequence possesses all the featuresof a full-length clostridial H2ase sequence. The only signi¢cant discrepancy is the presence of a `SCSPGW' segment instead of the consensus `SCCPGW'in the L1Fe signature (Fig. 4). This would result in the replacement of one of the cysteine ligands of the H-cluster by a serine.gUnpublished sequence. Reported in [35] to be identical to the amino acid sequence of D. vulgaris H2ase [278].hThe signal sequence is N-terminal to the small subunit. The gene encoding the latter (hydB) follows the gene encoding the large subunit (hydA).iThese genes were initially named according to their ordering in the operon, thus the catalytic subunit was HndD [274]. Using HndA instead would re-sult in a consistent notation for all [Fe]-H2ases (see Section 8).jThis putative gene (locus name TM1421 in [203]) would encode a [Fe]-H2ase with large deletions in the H-domain (see Fig. 5 and Section 6).kn.d.: not determined.lMaturation of the protein results in the removal of 56 N-terminal residues (T. Happe, personal communication).mMaturation of the protein results in the removal of 21 N-terminal residues (T. Happe, personal communication).nEntry for nucleotide sequence. Another entry, AJ012098, is to be released shortly (T. Happe, personal communication). The recently reported sequenceof strain `wild-type D3' (AAG59621) appears to miss 17 residues at the N-terminus of the processed protein.oMaturation of the protein results in the removal of 35 N-terminal residues [72].pThe sequence in entry AAG31036 [275] seems to be a truncated version (nine N-terminal residues missing or uncertain) of this one.qMany sequences of this type have been found in the genomes of aerobic eukaryotes. Only a selection is given here. Some may be truncated. The fulllength ones consist of the 2[4Fe^4S] and H-cluster domains (see [275,325] and Section 5.3).rPossibly truncated.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 475
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

in the other sequences and which might compensate forthe missing domains in their H-cluster-sequestering role(T. Happe, personal communication). The previously re-ported presence of [NiFe]-H2ases in green algae [68,70,368]has not been supported by gene cloning and sequencing,and has been questioned in a recent publication describingin detail the [Fe]-H2ase from S. obliquus [72].
5.1.2. Accessory domainsMany [Fe]-H2ases possess, in addition to the H-cluster
domain, an N-terminal domain homologous to the 2[4Fe^4S] bacterial ferredoxins. Signi¢cant examples include thetwo short (ca. 50 kDa) monomeric T. vaginalis H2ases[276] and the M. elsdenii H2ase [273]. Some enzymes hav-ing the same structural framework are dimeric (Section5.2). The clostridial-type [Fe]-H2ases (ca. 64 kDa) are sig-ni¢cantly larger [14,272] (see however Section 6 for possi-ble exceptions), and contain three domains in addition tothe H domain: starting from the N-terminus, a [2Fe^2S]plant ferredoxin-like domain [34,326,327], a unique [4Fe^4S]-containing fold, and a 2[4Fe^4S] domain [34]. A re-cently sequenced gene from T. vaginalis [275] encodes aclostridial-type monomeric H2ase of 64 kDa [329]. Thecatalytic subunit of the tetrameric NADP-reducing H2asefrom D. fructosovorans also has the same size and domaincomposition as clostridial H2ases [274]. A further increasein size is observed in the catalytic subunit (73 kDa, Fig. 3)of the trimeric T. maritima H2ase [204]: it is clostridial-like, but has a C-terminal extension homologous to theNuoE subunit of NADH-ubiquinone oxidoreductases(Complex I, see Section 7.3) and bacterial thioredoxin-like[2Fe^2S] ferredoxins [328,330]. The largest catalytic subunitknown so far (ca. 130 kDa) is the putative monomericH2ase from the anaerobic eukaryote Nyctotherus ovalis[322]: it possesses all the domains present in the T. maritimaenzyme, and yet an additional C-terminal domain homolo-gous to the NuoF subunit of Complex I (Fig. 3).
5.2. Additional subunits
As mentioned above (Section 3.6), the small subunit ofthe dimeric [Fe]-H2ases is a counterpart of the C-terminusof the monomeric enzymes, with regard to sequence aswell as structure; it contains no prosthetic groups andembraces the large subunit in a circular fashion [35]. Thesmall subunit also carries the signal sequence allowing ex-port of the whole enzyme to the bacterial periplasm inmembers of the genus Desulfovibrio [216,278].
The tetrameric HndABCD H2ase from D. fructosovo-rans [274] and the trimeric HydABC H2ase from T. mari-tima [204] are closely related [204]. The catalytic subunitsare in both cases encoded by the last (in the order ofappearance in the operons) of the structural genes (hndDand hydA, respectively) and have been described above(Section 5.1 and Fig. 3). The ¢rst of the structural genes(hndA and hydC, respectively) encode the smallest subunits
(19 kDa) which contain a [2Fe^2S] cluster and are homol-ogous to the NuoE subunit of Complex I (Section 7.3).The middle subunit (HydB) of T. maritima resembles afusion of the subunits HndB and HndC of D. fructosovo-rans (our unpublished observations). Thus the dissimilaritybetween the quaternary structures of these two H2asesdoes not re£ect a signi¢cant structural di¡erence. The sim-ilarity of the T. maritima and D. fructosovorans H2ases isfurther underscored by the presence of an operon encod-ing a putative Hnd-like trimeric H2ase in the currentlysequenced genome of Clostridium di¤cile (http://www.
Fig. 3. Schematic representation of the modular structure of [Fe]-H2ases. The sequences (also listed in Table 5) are from S. obliquus [72],M. elsdenii [273], C. pasteurianum [272], D. fructosovorans [274], T. mari-tima [204], N. ovalis [322]. The domains are inferred from comparisonsof sequences and structures [34,35]. They are not drawn to scale andtheir sizes are as follows: H-cluster domain, ca. 350 residues [34,35] ;[2Fe^2S] plant ferredoxin-like domain, ca. 75 residues [34,326,327]; thio-redoxin- and NuoE-like [2Fe^2S] domain, ca. 100^150 residues[204,322,328]; [4Fe^4S] cluster with (Cys)3His ligation, ca. 60 residues[34] ; 2[4Fe^4S] ferredoxin-like domain, ca. 80 residues [34,35] ; NuoF-like domain, ca. 450 residues [204,322]. The masses (in kDa) are givenbelow each subunit. The catalytic subunits of the dimeric periplasmicenzymes from the genus Desulfovibrio (Table 5) are similar to the onefrom M. elsdenii. Numbers inside the symbols of H2ases 4 and 5 indi-cate the order of the encoding genes in the operon. Subunit 2 of T. ma-ritima H2ase is homologous to a fusion of subunits 2 and 3 of D. fruc-tosovorans H2ase (Section 5.2). H2ases 1, 2 and 3 interact withferredoxins, £avodoxins or low-potential cytochromes. H2ases 4 and 5interact with NAD(P), presumably through the NuoE- and NuoF-likedomains. Due to the presence of these latter domains in the H2ase fromN. ovalis, it is also presumed to implement NAD(P) as a redox partner(Section 5.3). Thus, H2ases 4, 5 and 6 are of the `Hnd' subgroup (Sec-tion 8).
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501476
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

sanger.ac.uk/Projects/C_di¤cile). This putative H2ase ismost similar to that of T. maritima, and its second subunitalso resembles a fusion of the HndB and HndC subunitsof D. fructosovorans (our unpublished observations). Be-cause of these similarities, we propose the designation Hndfor all these hydrogenases (Section 8 and Table 5), whichare remarkable by the extensive similarities of their acces-sory subunits with subunits of Complex I (Section 7.3).The number and patterns of cysteine residues in Hnd-type H2ases suggest unusually high numbers of Fe^S clus-ters. For instance, the trimeric T. maritima H2ase maycontain, according to its sequence, altogether four [2Fe^2S] and six [4Fe^4S] clusters in addition to the H-cluster.This inference is well supported by the published analyt-ical and spectroscopic data [204]. The T. maritima H2asealso stands out by its containing one NuoE-like, or thio-redoxin-like [2Fe^2S] domain [328,330] in each of its threesubunits [204].
5.3. Redox partners of [Fe]-H2ases
In keeping with their occurrence in very diverse organ-isms endowed with a wide range of metabolic capabilities,[Fe]-H2ases are very versatile with respect to electron do-nors and acceptors.
Clostridial H2ases are reduced in vivo by a 2[4Fe^4S]ferredoxin under iron replete conditions, but under ironstarvation they use £avodoxin which the cells synthesize asa substitute of the iron demanding ferredoxin [331]. Invitro, clostridial H2ases can use a variety of oxidants orreductants [332]. Many investigations ([332] and referencestherein), a number of which were aimed at the develop-ment of biological H2 production [333,334], have demon-strated that [2Fe^2S] plant-type ferredoxins are very e¤-cient electron donors to clostridial H2ases. Kinetic studiesimplementing site-directed variants of the C. pasteurianum2[4Fe^4S] ferredoxin have indicated that electrostaticforces play only a minor role, if any, in the interactionsof this electron carrier with H2ase [335]. Further investi-gations have been stimulated by the availability of a struc-tural model for C. pasteurianum H2ase [34]. Indeed, di¡er-ent interaction sites have been proposed for ferredoxin onthe basis of charge [40] or surface [336] complementarity,respectively. The three accessory domains (Section 5.1 andFig. 3), each of which contains Fe^S sites, o¡er a largeand diverse surface which is likely to allow e¤cient inter-actions with numerous redox partners, including those notindigenous to clostridial cells, e.g. plant-type [2Fe^2S] fer-redoxins. It is to be mentioned here that the metabolism ofM. elsdenii is closely related to the clostridial one and thatthis bacterium likewise uses a 2[4Fe^4S] ferredoxin and a£avodoxin as electron carriers ([273] and references there-in). Nonetheless, its H2ase di¡ers from the clostridial onesby the absence of the two N-terminal accessory domains([273] and Fig. 3).
The small H2ases from the green algae S. obliquus, C.
reinhardtii and C. fusca are devoid of accessory domains([72] and Table 5). In these organisms the electrons for H2
evolution are provided by the fermentative metabolism,fed via the plastoquinone pool into PS I, which in turnreduces the [2Fe^2S] ferredoxin. The latter protein hasbeen shown by inhibition studies to be the physiologicalelectron donor to H2ase [72]. The periplasmic dimeric Fe-[H2ase] from Desulfovibrio reduces cytochrome c3. A struc-tural model for this interaction has been derived fromNMR experiments and docking calculations [295]. Thetetrameric cytosolic [Fe]-H2ase from D. fructosovorans re-duces NADP [274].
The phosphoroclastic reaction followed by H2 evolutionis an essential feature of the energy metabolism in manyanaerobic eukaryotes [64,337]. Unlike clostridia, however,these organisms implement a plant- and mammalian-type[2Fe^2S] ferredoxin to transfer electrons from pyruvate-ferredoxin oxidoreductase (PFO) to H2ase [338]. Some an-aerobic chytrids have recently been reported to use pyru-vate-formate lyase instead of PFO for the oxidation ofpyruvate [339]. In this case H2ase has been hypothesizedto interact with the hydrogenosomal membrane and to bereduced by NADPH [339].
In summary, [Fe]-H2ases are e¤cient users of a widerange of electron donors or acceptors. It has also beenshown above (Section 5.1, Fig. 3) that they di¡er greatlyin size and number of accessory domains. This might sug-gest that these domains determine the selectivity of theenzymes towards redox partners. Several observations,however, are in contradiction with this inference. For in-stance, H2ases in clostridia and M. elsdenii have di¡erentaccessory domains and nevertheless implement similar re-dox partners [273]. Also, it is well established that [2Fe^2S] plant-type ferredoxins are very e¤cient electron do-nors to a wide range of H2ases having di¡erent sizes andaccessory domains [72,332^334]. At present, the only cor-relation between accessory domains and redox partnerspeci¢city appears to be the presence of NuoE- and
Fig. 4. Characteristic sequence signatures within the H-cluster domainof [Fe]-H2ases. The L1Fe, L2Fe and L3Fe signatures have been derivedfrom alignments of 24 of the H2ase sequences listed in Table 5. TheH2ase-like sequences from aerobic eukaryotes, ORF TM1421 from T.maritima, and H2ases from C. di¤cile and C. fusca have been excluded.The edges of the three segments have been numbered according to theC. pasteurianum sequence [34,272]. Cysteine ligands of the H-cluster arestarred. Residues in underlined bold lettering are fully conserved. Thebold non-underlined residues are conserved in all but one sequence:S298 is T in T. vaginalis HydA3; C300 is S in C. thermocellum ; M353is T in M. elsdenii ; A498 is V in S. barkhanus ; G507 is A in E. histoly-tica. Brackets include residues found at that position in the aligned se-quences. Underlined residues within brackets occur in more than 80%of the sequences. X indicates that more than four di¡erent residues arefound at that position.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 477
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

NuoF-like domains (Section 7.3) in [Fe]-H2ases interactingwith NAD(P)(H) [204,274].
5.4. Putative hydrogenases and hydrogenase-relatedproteins
A gene from D. vulgaris, hydQ, has been reported toencode a [Fe]-H2ase that was deemed putative becauseof the absence of detectable protein product [320]. Itdoes nevertheless possess all the conserved features of gen-uine H2ases, and has a length comparable to that of theclostridial enzymes [320,272] (Table 5).
An unusual sequence has been uncovered in the genomeof T. maritima (ORF #Tm1421, [203]). This very shortORF (ca. 300 codons) would encode an H domain only,with several deletions. Some of the latter are extensive (upto 40 codons [273]) but nevertheless involve mostly loopsor secondary structure elements that are near the surfaceof the molecule (Fig. 5). It should therefore not be ruledout that this gene might encode a stable H2ase or H2ase-like protein. Similar genes are present in C. di¤cile andC. acetobutylicum (Section 6).
The genomes of yeasts and several higher eukaryoteshave recently been found to contain sequences homolo-gous to [Fe]-H2ases [325] (Table 5). These sequences aremost similar to those of H2ases having an H domain and a2[4Fe^4S] domain, e.g. M. elsdenii (Section 5.1.2 and Fig.3), but they lack the counterparts of the cysteine ligands of
the [4Fe^4S] cluster distal from the H-cluster. Whereas itis unlikely that any of the putative products of these geneswill turn out to be endowed with H2ase activity, the trans-lated sequences display most of the conserved featuresknown to be essential for the activity of genuine H2ases[34,35]. The product of such a gene from Homo sapienshas been shown to be located in the nucleus and to inter-act with prelamin A (hence its name Narf) [325]. In viewof the sequence similarities it is almost certain that thesegenes share a common ancestor with [Fe]-H2ases (Section7.2), but the pathways of this evolution and the presentfunction of the gene products remain to be elucidated.
6. Hydrogenase-related genes in fully sequenced genomes
Well over 40 bacterial and archaeal genomes have beensequenced over the last 6 years (http://www.tigr.org). Hy-drogen metabolism plays an important role in several ofthese organisms, and it is no surprise that their genomesencode a number of H2ases or accessory proteins thereof.For those organisms in which H2ases had been well char-acterized genetically and biochemically, genome sequenceshave provided mainly con¢rmatory information. For in-stance, the complete genome of E. coli [236] contains thegenes encoding the three previously known H2ases andtheir accessory proteins as well as the set of genes encod-ing the fourth H2ase (Hyf) discovered by DNA sequencing[237].
In contrast, a wealth of information has been extractedfrom genome sequences of bacteria for which biochemicaland genetic data were limited or altogether absent. Thegenomes of the archaeon Archaeoglobus fulgidus [227]and the bacterium A. aeolicus [209] (Table 6) contain sev-eral sets of genes encoding putative [NiFe]-H2ases. Thegenome sequence of the bacterium T. maritima [203] hascon¢rmed the presence of genes encoding the previouslysequenced and biochemically characterized trimeric [Fe]-H2ase [204] (Sections 5 and 8). However, it shows in ad-dition that these structural genes are part of a putativeoperon consisting of eight ORFs (Table 7). This is the ¢rstobservation of an [Fe]-H2ase-encoding operon includinggenes other than the structural ones. The T. maritima ge-nome also contains other hnd-like genes that might encodeat least one additional [Fe]-H2ase (Table 7) related to thetrimeric [Fe]-H2ase isolated from the same organism [204]and the Hnd enzyme from D. fructosovorans [274].
Clostridial [Fe]-H2ases appear to be a very homogene-ous group of enzymes. Cloning experiments by classicalmethods [272,316^318] have so far suggested the presenceof a single [Fe]-H2ase-encoding gene in each of the inves-tigated species. The only known exception was the isola-tion of a second [Fe]-H2ase from C. pasteurianum [14,340].Then, the genome sequence of C. di¤cile, now in the ¢n-ishing stages (http://www.sanger.ac.uk/Projects/C_di¤-cile), seems to reveal a far more complex picture. It en-
Fig. 5. Hypothetical model of the putative product of ORF TM1421from T. maritima ([203], see also Table 7). The model is based on thecrystal structure of the [Fe]-H2ase from C. pasteurianum [34], and de-rived from sequence alignments [273]. The protein fold of the H-clusterdomain (residues 210^574) of C. pasteurianum H2ase is shown here. Theregions corresponding to deletions in the putative H2ase from T. mariti-ma [273] are highlighted in a darker shade of gray. Atoms of the H-cluster are represented in space ¢lling (CPK). The molecule is orientedapproximately as in ¢gure 1A of [34]. The ¢gure was prepared using theprogram RASMOL [378].
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501478
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

codes a putative Hnd-like trimeric enzyme (Sections 5.2and 8, Table 5) similar to the trimeric enzyme fromT. maritima [204], a premiere among clostridia. In addi-tion, four genes encoding putative monomeric [Fe]-H2asesare also present (Table 5). These [Fe]-H2ases would beshorter than the C. pasteurianum HydA, and show unex-pectedly diverse features in several regions of their se-quence. HydA1 (458 codons) resembles the M. elsdenii se-quence, but exhibits a di¡erent distribution of cysteineligands in the N-terminal 2[4Fe^4S] domain. HydA1 isthe only one among the four hyd genes of C. di¤cilethat appears to have a complete H domain. Indeed,hydA2 (496 codons), hydA3 (498 codons) and hydA4(478 codons) all display large deletions in their H-domainswhich strongly relate them to ORF Tm1421 from T. ma-ritima (Fig. 5, Section 7.2). Whether hydA1-A4 are truegenes remains to be demonstrated, but all of them arepreceded by consensus ribosome-binding sequences. Ge-
nomes of other clostridia are being sequenced. For in-stance, the genome of C. acetobutylicum (http://www.ncbi.nlm.nih.gov/Microb_blast/un¢nishedgenome.html),contains an [Fe]-H2ase-encoding gene identical to one thathas already been cloned and sequenced [317], as well as anadditional one that is related to hydA2-4 from C. di¤cileand ORF Tm1421 from T. maritima.
The numerous genes required for the biosynthesis/mat-uration of [NiFe]-H2ases, in particular hypA-F and occa-sionally hupE, are present in annotated genomes as ex-pected. No accessory genes other than those previouslyevidenced have been detected in genome sequences. It isnoted that the hypA-F genes are clustered in A. fulgidus[227] as in most other organisms, but in contrast they aredistributed over three loci in A. aeolicus (Table 6). Regard-ing the maturation of [Fe]-H2ases, genome sequences arenot very informative, despite the discovery of a putativemulticistronic operon in T. maritima (Table 7). It is never-
Table 6Genes encoding hydrogenase-related proteins in the genome of A. aeolicusa
Aq # Strandb Original annotationc Proposed annotationd Function of encoded protein
660 mbhL1 hupS NiFe small subunit662 mbhL1 hupL NiFe large subunit (neighbors #660)665 hoxZ hupC cytochrome b-containing membrane subunit666 hupE Ni transport (putative, similar to ureJe)667 hupD protease (large subunit C-terminal maturation)f
668 none hupG unknown669 none hupH unknown671 hypB Ni insertion (neighbors #669)672 hypF hypF CO/CN synthesisg
802 mbhS3 NiFe small subunith
804 mbhL3 NiFe large subunith
960 compl mbhL2 hynL NiFe large subuniti
961 compl hdrD putative transmembrane electron transferi;j
963 compl none putative transmembrane electron transferi
965 compl mbhS2 hynS NiFe small subuniti
1019 hypE unknown1021 hypA Ni insertion (putative, cysteine rich)1156 compl hoxX hypX maturationk
1157 compl hypD maturation, putative FeS protein1158l compl none hypC maturation
aGenes encoding H2ases and related proteins occur in six clusters : 660^672, 802^804, 960^965, 1019^1021, and 1156^1158. Only [NiFe]-H2ases appearto be present. Five genes encoding NtrC-like transcription regulators are not included in this table. An ORF (#1591) was initially annotated shyS for`soluble hydrogenase small subunit' [209]. New searches and comparisons have shown that it is in fact homologous to aminotransferases (our unpub-lished data), and has therefore not been included in the table.bcompl: complementary strand; encoded by direct strand when void.cReference [209].dWhen di¡ering from the original one.eReference [210].f Function inferred by homology with the well-characterized HybD and HycI proteins (Table 1).gReference [194].hPossess features characteristic of H2-sensing H2ases (Tables 2 and 8, Section 4.2.2.2) but the function of the putative products of these genes remainsto be elucidated. Therefore, no revision of the current annotation is proposed.iORFs #960^963 display the highest similarity to four consecutive ORFs of the hyd (Table 2) operon from T. roseopersicina [190], and they appear inthe same order on the chromosome. Similar functions may therefore be predicted for these enzymes. ORFs #961 and #963 are homologous to the inser-tion sequences isp2 and isp1, respectively, that occur between the hydS and hydL genes of T. roseopersicina [190]. They also resemble two ORFs fromthe hmc operon which encodes a transmembrane electron transfer complex in D. vulgaris (GenBank L16784, [294]).jHomologous to heterodisul¢de reductases present in a large number of organisms. Neighbours #963.kLocal sequence similarities with formyl transfer catalysts [117] and in particular methionyl-tRNA formyl transferase, of which the 3D structure isknown [195].lNot detected in [209]. Number attributed because it neighbors ORF #1157.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 479
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

theless signi¢cant that the latter bacterium, which appearsto contain only [Fe]-H2ases, is altogether devoid of genesbearing resemblance to any of those involved in the mat-uration of [NiFe]-H2ases (our unpublished results,searches carried out using TBLASTN [341,342]). Thiswould suggest that the two classes of metal-containingH2ases rely upon phylogenetically distinct proteins fortheir maturation.
7. Phylogeny
As detailed in Sections 4 and 5, H2ases display highlymodular structures, an amazing diversity in quaternarystructure and, at least in the case of [Fe]-H2ases, in thesize of the catalytic subunit (Fig. 3). Furthermore, most ofthe subunits and domains other than the H-cluster domainof [Fe]-H2ases have counterparts in other proteins andcomplexes, for instance ferredoxins and NADH-ubiqui-none oxidoreductase (Section 7.3). These observations re-£ect a widespread swapping of redox protein modulesamong energy conservation systems, and they will doubt-less be instrumental in improving our understanding of theevolution of bioenergetics at large. However, within theframework of hydrogenase biodiversity and evolution, itappears more relevant to focus on those features that areidiosyncratic to H2ases, or at least very conserved withineither the [Fe]- or [NiFe]-H2ases. Sequence comparisonswere therefore restricted to the catalytic subunits (oreven H-cluster domains) for the [Fe]-enzymes, and to thelarge and small subunits for the [NiFe] enzymes.
7.1. [NiFe]-hydrogenases
Phylogenetic trees of [NiFe]-H2ases have been derivedfrom amino acid sequence alignments (Fig. 6). OnlyH2ases for which complete sequences of both subunitswere available have been included in the trees. Incompleteones are listed in Table 8. The trees show groupings con-sistent with the functional classes de¢ned in Section 4. Theposition of these groups relative to the outgroup con¢rmthe monophyletic origin of the [NiFe]-H2ases. Moreover,within each H2ase class, the trees derived from the sequen-ces of the large and small subunits are nearly superimpos-able. This indicates that these enzymes have consisted oftwo tightly associated subunits for most, and probably all,of their evolutionary history. Within each of the classes,H2ase phylogeny is essentially consistent with speciestaxonomy derived from other data [63,348,349]. In Group1 for instance, H2ases of N- and O-proteobacteria are clus-tered on a deeper branch than those of the K-, L- and Q-proteobacteria, in keeping with the proposal that they ap-peared earlier than the other proteobacteria [379] (Fig.6B). Some discrepancies are nevertheless observed. Forexample, in Group 1, the mixing of H2ases from Q-proteo-bacteria with those from K-proteobacteria is in contradic-tion with the proposal that the former bacteria emergedlater [379]. These contradictions, as well as others to bementioned below, may be accounted for by various events,e.g. lateral gene transfer [203,350,351]. Another unex-pected occurrence is the presence of the uptake H2asesfrom cyanobacteria in Group 2, together with the H2-sens-ing proteins. The proteins of subgroups 2a and 2b have
Table 7Genes encoding H2ases and related proteins in the genome of T. maritima MSB8a
Tm #b Strandc Annotationb Proposed annotationd Function
10 compl hndC-1 hndBC2 NADP-reducing [Fe]-H2ase (subunit C)e
11 compl hndB hndBN2 NADP-reducing [Fe]-H2ase (subunit B)e
12 compl hndA-1 hndC2 NADP-reducing [Fe]-H2ase (subunit A)e
201 none hndA2 NADP-reducing [Fe]-H2ase (subunit D, catalytic)e
227 hndA-2 hndC3 NADP-reducing [Fe]-H2ase (subunit A, sequence contains aframeshift)
228 hndC-2 hndB3 NADP-reducing [Fe]-H2ase (subunit C)1420 none putative [2Fe^2S] ferredoxinf
1421 none putative [Fe]-H2aseg
1422 none rnfB-related (FeS, electron transfer)1423 none unknown, putative transcription regulator1424 hydC hndC1 [Fe]-H2ase (subunit C)h
1425 hydB hndB1 [Fe]-H2ase (subunit B)h
1426 hydA hndA1 [Fe]-H2ase (subunit A, catalytic)h
1427 none similar to YdiH, function unknown
aGenes encoding H2ases and related proteins occur in four clusters: 10^12, 201, 227^228, and 1420^1427. Only [Fe]-H2ases appear to be present.bReference [203].ccompl: complementary strand; encoded by direct strand when void.dSee Section 8.eThis set of four genes might encode an NADP-reducing [Fe]-H2ase homologous to the one isolated from D. fructosovorans [274].f Similar to, but truncated version of, thioredoxin-like [2Fe^2S] ferredoxins from A. aeolicus and several other bacteria [328]. Also homologous to hndC1(Tm1424).gShort version (ca. 300 codons) of the H-domain of [Fe]-H2ases, with deletions in surface exposed regions [273] (Section 5.3 and Fig. 5).hThe H2ase encoded by these genes has been characterized biochemically [204].
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501480
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

most probably evolved from a common ancestor and theirfunctions have diverged (H2ase and H2 sensing) dependingon the host organism.
While representatives of the four groups of enzymes arefound in Bacteria, the Archaea are missing (for the timebeing) representatives of Group 2, which suggests that theemergence of Group 2 H2ases occurred within Bacteriaafter the divergence of the domains Archaea and Bacteria.These lineages are very unequally represented in the fourH2ase groups. Indeed, most bacterial sequences map with-in Groups 1 and 2, whereas archaeal ones (emphasized bylight gray shading in Fig. 6) belong to Groups 3 and 4(except for VhtGA from A. fulgidus and M. mazei, andVhoGA from M. mazei, which are associated to Group 1).Thus, the earliest divergence event inferred from compar-ison of [NiFe]-H2ases is the branching o¡ of the Bacteriafrom the Archaea, in agreement with the generally ac-cepted phylogeny.
The presence of H2ases 1 and 2 from A. aeolicus amongH2ases from Proteobacteria in Group 1 would seem to beinconsistent with the view that Aqui¢cales are the deepestbranching organisms [380] in the domain Bacteria. How-ever, recent studies in comparative genomics [381] indicatethat Aqui¢cales are closer to the Proteobacteria than pre-viously reported. The latter conclusion would then be in
better agreement with the positions of H2ases 1 and 2from A. aeolicus in the trees of Fig. 6.
In Archaea, early duplication events seem to have led tothe presence in the same genome of genes belonging todi¡erent subgroups of Group 3 (M. voltae, M. thermoau-totrophicum). Other duplications have probably occurredin a more recent past since closely related genes arepresent in some organisms (e.g. frhG1A1 and frhG2A2 inM. barkeri, mvhG1A1 and mvhG2A2 in M. thermoautotro-phicum, hynSL and hupSL in A. aeolicus and T. roseoper-sicina). An interesting phenomenon is the presence of cy-anobacterial HoxYH H2ases in Group 3 where no otherbacterial [NiFe]-H2ases are found (except the plasmid-en-coded HoxYH enzymes of R. eutropha and R. opacus).This may result from various causes of which the mostlikely could be a loss of HoxYH in a common ancestorof the Proteobacteria.
The H2ases of Group 4 belong to multisubunit proteincomplexes and share marked similarities with several sub-units of Complex I and F420H2 :quinone oxidoreductase(Section 7.3). In this Group, the nesting of bacterial se-quences (E. coli and R. rubrum) within the archaeal onesappears to contradict the general phylogenetic tree. Itmight be a remnant of early stages in the evolutionaryprocess [351].
Table 8Group assignment of genes not included in phylogenetic trees
Species Accessionnumber
Gene product Closest relative Large subunitpatterns
Proposedgroup
identity (%) (best scores with) L1 L2
Anabaena sp. CyanoBasea HoxY 69.0 P. hollandica HoxYPCC7120 CyanoBasea HoxH 73.0 P. hollandica HoxY 3d 3d 3dA. variabilis Q9ZAK3b HupS 98.1 Anabaena sp. HupSATCC 29413 Q9ZAK2b HupL 98.5 Anabaena sp. HupL 2a 2a 2aA. variabilis BAB39386 HoxYc 3dIAM M58A. aeolicus O66987 MbhS3d 46.8 Anabaena sp. HupS close to close to
O66988 MbhL3d 36.0 R. capsulatus HupV 2b 2b 2bA. fulgidus O28898 VhuGd 38.9 M. jannaschii VhuG close to close to
O28899 VhuAd 40.6 M. voltae VhuA 3c 3c 3cC. jejuni Q9PMQ8 HynS2c;e 47.8 C. jejuni HynSe 1 1 1
Q9L869 HynSe;f 70.9 D. vulgaris_Mi HynSD. desulfuricans Q9L868 HynLe;f 67.8 D. vulgaris_Mi HynL 1 1 1
P13061 HynSe;f 71.3 D. vulgaris_Mi HynSM. fervidus Q49178 MvhGf 80.3 M. thermoautotrophicum MvhG2
Q49179 MvhA 78.6 M. thermoautotrophicum MvhA2 3c 3c 3cP. furiosus Pf1352205 MbxJ 28.5 P. furiosus MbhJ
Pf1351019 MbxL 34.8 P. abyssi EchEe g g 4close to
R. capsulatus Q9XBW8 HoxHc 40.4 A. variabilis HoxH 3d 3d 3d
aCyanoBase: http://www.kazusa.or.jp/cyano/anabaena/.bSequences completed and updated too recently to be included in trees.cSecond subunit missing.dThe two subunits do not map in the same subfamily.eNew proposed gene name (see Table 3).f Incomplete sequence.gAssigned to Group 4, notwithstanding some pattern alterations.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 481
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

Fig. 6. Phylogenetic trees of the [NiFe]-H2ases small (A) and large (B) subunits. Only the complete sequences of the two subunits listed in Table 2 wereanalyzed. The sequences not included in the tree are listed in Table 8. The trees were obtained by the neighbor-joining method [343] using the BIONJprogram [344]. Distance matrices were computed with the PROTDIST program of the PHYLIP package [345] on sequences aligned with Clustal W[283]. Trees were displayed and printed with NJPLOT [346]. Distances along the horizontal axis re£ect the degree of relatedness of the sequences.NuoB (subunit of Complex I) and HydA (catalytic subunits of [Fe]-H2ases) sequences were used as outgroups for the tree of the small and the largesubunits, respectively. When species names are followed by `P', the genes are plasmid-encoded. Bootstrap support values (1000 replicates) [347] areshown above the relevant nodes. Numbers below the nodes are those of H2ase Groups (Table 2 and Section 4.2). Archaeal enzymes are shaded in lightgray, cyanobacterial enzymes in darker gray. The subdivisions of Proteobacteria are indicated on the right in B.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501482
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022
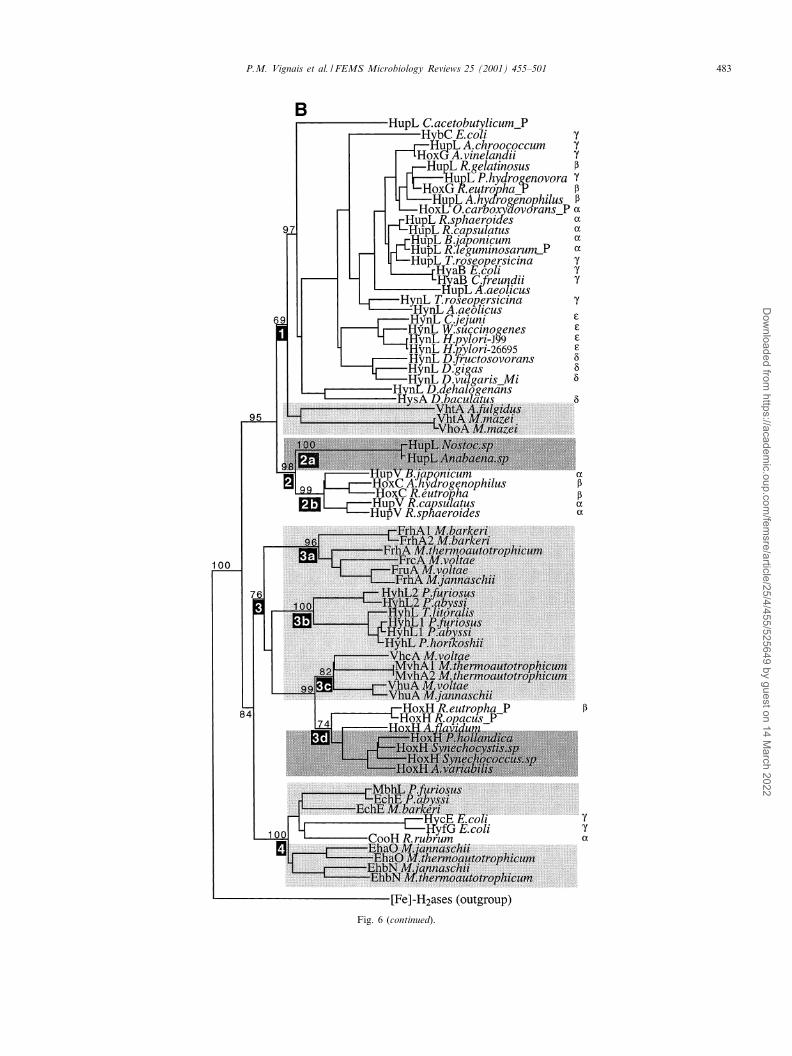
Fig. 6 (continued).
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 483
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

7.2. [Fe]-hydrogenases
Neither the quaternary structures, nor the sizes of thecatalytic subunits of [Fe]-H2ases are correlated with thephylogenetic status of the host organisms. Indeed, Bacteriacontain monomeric [272,273], dimeric [278], trimeric [204],and tetrameric [274] [Fe]-H2ases, whereas only mono-meric enzymes have so far been found in Eucarya[72,275,276,322]. Likewise, the smallest [72] and largest[322] catalytic subunits are found among Eucarya, and awide range of intermediate sizes are found in Bacteria aswell as in Eucarya (Section 5.1 and Fig. 3). It is thenarguable that the occurrence of subunit and domain swap-ping has blurred the phylogenetic information containedin whole molecules of [Fe]-H2ases, or even in full-size cat-alytic subunits. This suggests two main options for se-
quence alignments of [Fe]-H2ases. In the ¢rst one, full-length catalytic subunits would be implemented becausethey are the biologically functional molecules, notwith-standing biases arising from discrepancies in length anddomain composition. Alternatively, in view of the lackof knowledge concerning the history of accessory domainswapping, it might be sensible to perform sequence align-ments only with the catalytic core that is common to all ofthese enzymes. The latter approach has been used recentlywith sequences edited to keep only the H-cluster domainand neighboring 2[4Fe^4S] domain [275]. Considering therecent discovery of [Fe]-H2ases consisting of the H-clusterdomain only ([72] ; Table 5), alignments of edited sequen-ces should now include only this part of the sequence. Wehave carried out sequence alignments of full-length cata-lytic subunits (not shown), and also of H-cluster domains(Fig. 7). The phylogenetic trees derived from the two setsof sequences were found to be altogether consistent witheach other. Both trees indicate similar relationships, whichare also in keeping with the previously published ones[275]. As in the case of the [NiFe]-H2ases, the positionof the outgroup sequences con¢rms that all [Fe]-H2asesare monophyletic. The main features that emerge fromthe trees are the following. As expected, the H2ase-likeproteins of the aerobic eukaryotes belong to a singlebranch. In contrast, the sequences of anaerobic eukaryotes(T. vaginalis, N. ovalis, E. histolytica and Spironucleusbarkhanus) appear to be distributed on several branchesas reported previously [275]. Whether this re£ects the ac-tual evolutionary history of these organisms or resultsfrom gene transfer events remains an open question[64,275,337], and particularly so in view of non-optimalbootstrap values at some important nodes of the availabletrees ([275] and Fig. 7). Among bacteria, the sequencesfrom di¡erent genera are not well segregated, which maybe due in part to the presence of paralogous genes inseveral species.
The observations listed above suggest that phylogeneticinferences based on [Fe]-H2ases should be made with cau-tion. It is nevertheless noteworthy that the overwhelmingmajority of the catalytic subunits are either of the two-domain (2[4Fe^4S] and H) -type or of the four-domain(clostridial-like) -type (Table 5 and Fig. 3). This is madeeven clearer in the dendrogram of Fig. 8, which includesseveral sequences from C. di¤cile (Table 5), in addition tothose implemented in Fig. 7. It is signi¢cant that, regard-less of the host organism, the deepest branches of the treeas rooted by the outgroup include almost exclusively se-quences of the two-domain-type (Fig. 8). These observa-tions suggest that this relatively simple [Fe]-H2ase struc-tural framework might be a good candidate for anancestral type from which other varieties of [Fe]-H2aseshave been derived (Fig. 9). It should be stressed that thishypothesis is put forward only regarding the [Fe]-H2ases,and that it is not meant to draw conclusions concerningthe host organisms.
Fig. 7. Phylogenetic tree derived from sequence alignments of H-clusterdomains of [Fe]-H2ases. The H-cluster domain of C. pasteurianum H2aseconsists of residues 210^574 [34]. Other sequences (listed in Table 5)were edited accordingly, on the basis of the full-length sequence align-ments. Sequences were aligned as described in the legend of Fig. 6.Bootstrap values indicated on nodes were determined as in Fig. 6.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501484
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

7.3. Similarities between hydrogenases and NADH-ubiquinone oxidoreductases (Complex I) of respiratorychains
NADH-ubiquinone oxidoreductase (Complex I) is bestknown as found in the inner membrane of mitochondriaand in the cytoplasmic membrane of bacteria. The formerconsists of at least 42 subunits whereas the latter is com-posed of only 14 subunits, all of which have counterpartsin the mitochondrial enzyme. The present discussion willbe restricted to the bacterial enzyme that contains thesame prosthetic groups (FMN, 8-9 Fe^S clusters and anundetermined number of quinones) and has the same cat-alytic properties as the mitochondrial enzyme [58,61,352^354]. The subunits are distributed nearly equally between amembrane and a hydrophilic domain [355,356]. Most, if
not all, of the metal sites are located in the latter. SubunitsNuoE, F, and G are the outermost ones in the hydrophilicdomain, while NuoB, C, D, and I are nearer to the mem-brane domain [357]. The subunits of Complex I that arerelevant to the present discussion, their localizations, andFe^S contents are listed in Fig. 10.
Sequence similarities between H2ases and Complex Iwere ¢rst reported by Bo«hm et al. [175] and then by Pil-kington et al. [54]. It is now well established that subunitsNuoE, NuoF, NuoI, and the N-terminal Fe^S-bindingdomain (ca. 220 residues) of NuoG have homologouscounterparts in accessory subunits and domains of boththe [NiFe]- and [Fe]-H2ases [58] (Fig. 10). The similarity ofthe N-terminal domain of NuoG with clostridial [Fe]-H2ases suggests that it might also contain one [2Fe^2S]and three [4Fe^4S] clusters [58]. However, in most cases
Fig. 8. Dendrogram derived from sequence alignments of H-cluster domains of [Fe]-H2ases, with emphasis on the modular structure of the catalyticsubunits. Unpublished sequences of C. di¤cile (Table 5) have been included in addition to those implemented in Fig. 7. The main purpose of this den-drogram is to highlight that the deepest branches are occupied by sequences consisting of two domains, the 2[4Fe^4S] domain and the H-cluster domain(see Fig. 9). These sequences are on a darker gray background. Clostridial-type (four domains) sequences are on a lighter (checkered) gray background.The four other sequences are on a white background, and their sizes are indicated by the neighboring icons. Symbols for domains are as in Fig. 3.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 485
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

(A. aeolicus is one of the exceptions [209]) NuoG missesthe counterparts of the cysteines that ligate the [4Fe^4S]cluster proximal to the H-cluster [34].
Additional similarities with Complex I concern mainly[NiFe]-H2ases. The NuoB and NuoD subunits of ComplexI bear clear similarities with the small and large subunits,respectively, of [NiFe]-H2ases [56^58,61]. These similaritieshave recently been reexamined, put on ¢rmer ground, anddiscussed with respect to £avin content and energy cou-pling mechanisms [61]. It is worth noticing that some ofthe highest sequence similarities between NuoD and thelarge subunit occur in regions which, in the H2ase struc-ture [33,61], surround the NiFe metal site. While NuoDmisses the counterparts of the cysteine ligands of the nick-el [56,58,61], it has been hypothesized that this subunit ofComplex I might accommodate a quinone site in a posi-
tion that approximately corresponds to the NiFe site inH2ase [363].
Altogether, it appears that nearly all subunits of soluble[Fe]- and [NiFe]-H2ases have homologs in the hydrophilicdomain of Complex I, and vice versa. The H domain of[Fe]-H2ases and the C-terminal domain of NuoG appearto be exceptions. However, NuoG and clostridial-type[Fe]-H2ases have similar sizes (600^700 residues) and ho-mologous N-terminal (220 residues) domains (see above,and Fig. 10). It is therefore not totally unlikely that theirC-terminal domains might assume similar 3D folds, de-spite the absence of detectable sequence similarities. Struc-tural data on NuoG are awaited to answer this question.
It is remarkable that [Fe]- and [NiFe]-H2ases, as well asComplex I, contain Fe^S domains that also exist as smallsoluble ferredoxins of three di¡erent types (Fig. 10). Crys-
Fig. 9. Tentative evolutionary scheme for the modular structures of the [Fe]-H2ase catalytic subunits. For illustrative purposes cartoons of the structureshave been drawn using the coordinate ¢les of C. pasteurianum H2ase ([34], pdb entry 1feh) and the thioredoxin- and NuoE-like ferredoxin from A. aeo-licus ([328], pdb entry 1F37). The only actual structure is that of C. pasteurianum H2ase (2), others are composite schemes drawn using RASMOL [378]with appropriately edited coordinate ¢les. Polypeptide chains are represented as ribbons, metal sites are in space ¢lling. No structural model is availablefor the NuoF-like domain of N. ovalis H2ase (4), which has therefore been represented by an icon as in Fig. 3. The proposed domain composition ofthe putative ancestral form (1) has been inferred from the widespread occurrence of H2ases having that size, particularly in the deepest branches of thetrees derived from the sequence alignments (Figs. 7 and 8). The larger enzymes (2^4) may be derived there from by domain acquisition. In contrast, thephotosynthesis-linked enzymes (5) are assumed to have lost the 2[4Fe^4S] domain. These very short H2ases are considered as unlikely primitive forms,because they occur only in photosynthetic eukarya. Periplasmic enzymes from Desulfovibrio (6) have the same domain composition as the putative an-cestral form, but their small subunit corresponds to the C-terminus (purple) of the monomeric enzymes [35]. In this scheme no assumptions are maderegarding the host organisms, their relative ancestry and evolution.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501486
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

tal structures are available for all of these protein folds:2[4Fe^4S] [362], plant- and mammalian-type [2Fe^2S][361], and thioredoxin-like [2Fe^2S] [328]. Together withthe crystal structure of C. pasteurianum [Fe]-H2ase [34], asigni¢cant body of structural models is thus assembled.These data, considering the sequence homologies indicatedabove (Fig. 10), provide useful models for the proteinfolds around most of the Fe^S clusters in both types of
H2ases and Complex I. Indeed, despite the absence of ¢rst-hand high resolution structural data on Complex I, theprotein folds around six at least of its Fe^S clusters canbe derived from experimental data on ferredoxins andH2ases (Fig. 10). It is anticipated that any further struc-tural data on H2ases, Complex I, or ferredoxins, will pro-vide useful information concerning all these proteins, andpresumably electron transfer complexes at large.
Fig. 10. Schematic representation of homologies between hydrogenases and Complex I. The symbols for domains are as in Fig. 3. `3D' means that acrystallographic model exists for that protein fold. Crystal structures have been elucidated only for proteins in the left column (H2ases and ferredoxins),not for any of the Complex I subunits. The ¢gure thus shows that 3D models can be derived from other proteins for at least ¢ve or six subunits ofComplex I. These include the protein folds around six or seven Fe^S clusters (in NuoE, NuoG, and NuoI).a The subunits of Complex I are designated using the Nuo nomenclature implemented for E. coli and R. capsulatus [58]. Other nomenclatures may befound in [58,60,61].b Homologous to the N-terminal part of the small subunit. NuoB may contain a [4Fe^4S] cluster located similarly to the proximal cluster in [NiFe]-H2ases [58,61].c NuoC and NuoD are fused in some bacteria [209,352], but clear similarities with the large H2ase subunit have been reported for NuoD only [61].NuoC and NuoD contain no conspicuous metal-binding site. The presence of a quinone site has been proposed [358].d Homologies with H2ases are restricted to the N-terminal V220 residues. All NuoG subunits have binding sites for one [2Fe^2S] and two [4Fe^4S]clusters [54,58]. One or two additional Fe^S clusters may be present in some bacteria [60].e NuoI is larger (ca. 20 kDa) than most 2[4Fe^4S] bacterial ferredoxins (ca. 6 kDa). The N- and C-terminal extensions present in NuoI presumably an-chor it to other subunits [359].f Approximate masses (kDa) of subunits obtained by compiling data from several bacteria [58,60,352,353,354].g Several crystal structures are available [33,37,39].h Subunits HoxF and HoxU from R. eutropha [133], HoxF and HoxU from A. variabilis [226], and HoxE, HoxF, and HoxU from Synechocystis [268].See footnotes e and n.i NuoH and NuoL are named ND1 and ND5, respectively, in mitochondrial Complex I [59,243,360].j See Fig. 3 for accessory domains in catalytic subunits. Two crystal structures are known [34,35]. The H-cluster domain is not included here because ithas no homolog in Complex I.k Trimeric and tetrameric enzymes from T. maritima [204] and D. fructosovorans [274], respectively. See Fig. 3 and footnotes e and n.l Crystal structures: [361] and references therein.m Crystal structures : [362] and references therein.n Crystal structure: [328]. These ferredoxins are homodimers of a V100-residue polypeptide chain ([328,330] and references therein).In the NuoE subunits and their homologs in H2ases, the polypeptide chains have additional N-terminal extensions of ca. 80 residues [58,328]. In someH2ases this domain does not have an adequate set of cysteine ligands for the accommodation of a [2Fe^2S] cluster. For instance, only two cysteines oc-cur in HoxF from R. eutropha [133]. In the NuoE-like domain of the [Fe]-H2ase from N. ovalis, the cysteine pattern might suggest the presence of a[4Fe^4S] cluster ([322] and Hackstein, personal communication), but this protein fold is known to allow extensive swapping of cysteine ligands whilestill accommodating a [2Fe^2S] cluster ([328] and references therein).
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 487
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

The homologies discussed above concern primarily solu-ble H2ases and the hydrophilic domain of Complex I.Recently, striking similarities have been evidenced amongthe membrane domains as well (reviewed in [60,61]). Itshould ¢rst be recalled that extensive similarities exist be-tween the membrane domains of Complex I andF420H2 :quinone (or phenazine) oxidoreductase from M.mazei [303] and A. fulgidus [227,364,365]. Sequences ofoperons encoding multimeric membrane-bound [NiFe]-H2ases from M. thermoautotrophicum (eha and ehb,[243]) and M. barkeri (ech, [360]) have con¢rmed the sim-ilarities among the soluble subunits that were describedabove. They have disclosed that in addition two trans-membrane subunits of the Eha, Ehb and Ech enzymesare homologous to subunits NuoH (ND1) and NuoL(ND5) of Complex I. H2ases 3 and 4 from E. coli[176,237], as well as the CO-inducible H2ase from R. ru-brum [265] likewise display such homologies (see Section4.2.4).
Altogether, the phylogenetic relationships between
H2ases and Complex I are more extensive among the met-al-containing redox subunits, but they also encompassmembrane subunits that are likely to be involved in protontransfer and energy conversion. This suggests that notonly electron transfer complexes but also energy-transduc-ing machineries have been transposed and adapted fromthe anaerobic into the aerobic lifestyle [60,61]. Phyloge-netic studies implementing sequences and structures ofH2ases and Complex I are therefore expected to shed lighton the evolution of bioenergetics. Furthermore, the evolu-tion of these redox complexes, either soluble or mem-brane-bound, is tightly intertwined with early symbioticevents that led to the emergence of the eukaryotic nucleus[366] and subcellular organelles, i.e. mitochondria, hydro-genosomes [367] and chloroplasts. In this context, the phy-logeny of H2ases may be anticipated to become a valuablesource of information. Indeed, [Fe]-H2ases are already im-plemented to unravel the evolution of anaerobic eukary-otes [275,322,337,367].
7.4. Distribution of hydrogenases in the domains of life
The availability of numerous H2ase sequences from awide range of organisms (Sections 4.2 and 5, Tables 2and 5) opens new opportunities for analyzing the distribu-tion of these enzymes in all domains of life. Of particularinterest is the unveiling of an increasing number of com-plete genome sequences, which allow full inventories ofH2ase-encoding genes to be made in a sizeable numberof organisms. A general observation is that, as for manyother genes, H2ase-encoding ones, when present, often oc-cur in numbers (Tables 2 and 5). H2ases are most numer-ous and diverse in Bacteria, where both [Fe]- and [NiFe]-H2ases are found. Furthermore, it is noteworthy that somebacteria possess [NiFe] enzymes only (E. coli, A. aeolicus),others only [Fe] enzymes (T. maritima). In others yet, bothtypes of H2ases coexist within the same cell (genus Desul-fovibrio). Regarding T. maritima, it was recently suggestedthat it might contain genes encoding a [NiFe]-H2ase [257].Upon closer examination, however, the relevant ORF(TM1216) turns out to assume a much closer relationshipwith NuoD-like (Section 7.3) sequences than with [NiFe]-H2ases (our unpublished results).
This raises the point of detecting and identifying H2asesin sequence searches. H2ases consist of domains that arefor the most part homologous to enzymes and complexesfunctionally alien to H2ases (Section 7.3). In addition, anumber of accessory domains or subunits are common tothe [NiFe]- and [Fe]-H2ases (Fig. 10). Therefore, searchesimplementing full-length H2ase sequences will often yieldvarious kinds of false positives. It is thus necessary toperform searches with portions of sequences that are spe-ci¢c of either one of the two classes of H2ases. These arethe H-cluster domain for the [Fe]-H2ases, and the largeand small subunits (e.g. HupL and HupS) for the[NiFe]-H2ases. Even then, the latter ones have to be im-
Fig. 11. Scheme illustrating the proposed nomenclature of [Fe]-H2ases.The subunits appear from left to right in the same order as the encod-ing genes on the operons. Numbers refer to organisms in which the cor-responding H2ases types are found. 1: C. rheinhardtii [72]; 2: M. elsde-nii [273] ; 3: D. desulfuricans [35] ; 4: C. pasteurianum [34,272] ; 5: N.ovalis [322] ; 6. D. fructosovorans [274] ; 7: C. di¤cile (unpublished, seeTable 5); 8: T. maritima [204]. See Table 5 and Section 8.2 for details.Symbols for domains are as in Fig. 3. Hnd H2ases are characterized bythe presence of NuoE-like (rectangles with vertical stripes) and NuoF-like (hexagons with horizontal stripes) domains.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501488
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

plemented with care because of detectable similarities withsubunits of Complex I (Fig. 10), as shown recently in thecase of T. maritima (see above, and [257]).
With these caveats in mind, it is feasible to retrievesigni¢cant information from the large body of sequencedata. For instance, it is noted that no [Fe]-H2ase-encodinggenes have yet been found in Archaea. Likewise, [NiFe]-H2ase-encoding genes have so far remained undetected inEucarya. Evidence for the presence of [NiFe]-H2ases inEucarya is restricted to biochemical and immunologicaldata on the green alga S. obliquus [68,70,368]. There isno further support from gene cloning and sequencing. Incontrast, four di¡erent [Fe]-H2ase sequences from thegreen algae S. obliquus [72], C. reinhardtii and C. fusca(T. Happe, personal communication) have been deter-mined by three di¡erent groups (Table 5). In addition,compelling biochemical data have been published on a[Fe]-H2ase from S. obliquus [72]. It may there from beinferred that, for the time being, [Fe]-H2ases are restrictedto Bacteria and Eucarya, while [NiFe]-H2ases are onlyoccurring in Bacteria and Archaea.
8. Nomenclature
As noted earlier in this review, over 100 sets of H2ase-encoding genes have been sequenced. These enzymes arefound in a wide range of organisms where they assumevery diverse structures and cellular functions. An ex-tremely heterogeneous and occasionally inconsistent no-menclature of H2ase-encoding genes has resulted therefrom. We are well aware that the present naming systemshave been extensively implemented in the literature overmore than a decade, and have in addition been engravedin hundreds of database entries. A reversal of this situa-tion will therefore require a protracted e¡ort of the scien-ti¢c community, which the proposals made here aim atinitiating.
8.1. [NiFe]-hydrogenases
As this is merely a proposal for initiating a reorganiza-tion of the nomenclature of H2ase-encoding genes, onlylimited changes have been made (Table 3, Sections 4 and7.1). Our main objective at this stage was to suppressinterference with the nomenclature of [Fe]-H2ases. Thus,the name hyd, which should be reserved for [Fe]-H2ases(Section 8.2), has been systematically replaced. For thesame reason, the letters A and B for the [NiFe]-H2asesubunits have been replaced in most cases by S and L(Table 3). When proposing new names, we have takencare to put forward designations that had been previouslyused for enzymes belonging to the same group, e.g. hox,hyn, ech, hup (Table 3), to avoid increasing the existingconfusion. The only exception is Hyh, which is used toavoid the now outdated term Shy (Section 4.2.3.2). In
keeping with the trend to use S and L for the small andlarge subunits of [NiFe]-H2ases, the K and N subunits ofShy are now called HyhL and HyhS, respectively. Theother two conserved subunits (beta and gamma) of thattetrameric enzyme should be termed HyhB and HyhG. Itis noted that the proposals summarized in Table 3 wouldresult in fewer H2ase names than previously. Finally, theattribution of new names has been based on phylogeneticproximity (Fig. 6 and Table 3). For instance hyn has beengiven to several sequences of a subgroup of Group 1which are closely related to Desulfovibrio sequences thathad been designated hyn previously (Tables 2 and 3). Weare aware that these proposals are still unsatisfactory, andshould be taken as no more than a starting point for arevision of the nomenclature.
8.2. [Fe]-hydrogenases
Clearly distinct groups of [Fe]-H2ases are di¤cult toidentify on the basis of sequences, structures and func-tions. It might therefore be sensible to apply the ¢rstused nomenclature, i.e. hyd [278], to all of these enzymes.
A closer inspection nevertheless reveals a subset ofmostly monomeric [Fe]-H2ases that interact with ferredox-ins, £avodoxins, or low-potential cytochromes. Some [Fe]-H2ases are dimeric and periplasmic, but structural datashow that they are very closely related to the monomericones [35]. Indeed, the small subunit has arisen by cleavageof the C-terminal part of monomeric cytoplasmic enzymesas a means of facilitating export to the periplasm [35]. Thegenes encoding these H2ases have in most cases beennamed hydA (large subunit) and hydB (small subunit). Itis proposed to retain this naming system and extend it toall genes of this subgroup of [Fe]-H2ases (Table 5, Fig.11).
A second category of [Fe]-H2ases is composed of mostlyoligomeric enzymes that interact with NAD(P), and con-tain domains homologous to the NuoE and NuoF sub-units of Complex I [204,274] (Figs. 3, 10 and 11, Section7.3). In the case of D. fructosovorans the encoding geneshave been named hnd (for H2ase NAD(P)-dependent)[274], a denomination that we propose to extend to allof these enzymes, including the monomeric one from N.ovalis, which also contains NuoE- and NuoF-like domains[322] and has been hypothesized to interact withNADP(H) [339]. Likewise, the putative trimeric Hnd-likeenzyme from C. di¤cile should belong to this group (Sec-tion 6 and Table 5). The catalytic subunits should benamed HndA, in keeping with the HydA ones (see above).The only existing discrepancy is the current use of HndDfor the catalytic subunit of D. fructosovorans tetramericH2ase [274]. The HndB and HndC subunits of that H2aseare fused in the other known cases (T. maritima and C.di¤cile). In D. fructosovorans, we therefore propose to re-place HndB by HndBN and HndC by HndBC (for N- andC-terminal), so that HndC would then be used for the
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 489
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

small subunit in all cases. In D. fructosovorans HndCwould replace HndA (Fig. 11).
The proposed nomenclature, which is summarized inFig. 11, encompasses all known [Fe]-H2ase-encodinggenes. For multiple isoenzymes in a single species, it isproposed to use numbers (Tables 2, 3, 5, 6 and 7). Genesencoding maturation proteins have not yet been identi¢edin the case of [Fe]-H2ases. When they are, letters startingwith E should be recommended, A to D being saved forstructural genes, in case an enzyme with an additional, yetunknown, structural subunit turns out. As it is most likelythat maturation proteins of the Hyd and Hnd enzymes arehomologous, care should be taken to use the same lettersfor the expectedly matching pairs of genes.
As shown in Figs. 7 and 8, the HydA and HndA se-quences are not phylogenetically distinct, but are nested inthe [Fe]-H2ase evolutionary trees. We nevertheless proposedistinct names because the Hnd H2ase can be distin-guished by function (interaction with NAD(P)) as wellas by sequence (presence of NuoE- and NuoF-like mod-ules, Fig. 11).
9. Concluding remarks and perspectives
H2ase research has made considerable progress in sev-eral directions over the last few years. The contributions ofX-ray crystallography and the consequent stimulation ofmany other investigations (spectroscopic studies, chemicalmodeling, calculations) to the ¢eld of H2ase is consider-able and cannot be overestimated. However, it is beyondthe scope of this review, and the reader is referred torecent reviews and discussions of these questions [42,51^53].
A considerable number of primary structures of H2aseshave been determined and compared over the last 15years. It has there from been inferred that the [Fe] and[NiFe] enzymes are phylogenetically distinct. This hasbeen con¢rmed by crystal structures showing that the pro-tein folds of the active site domains are completely di¡er-ent, notwithstanding some structural and chemical similar-ities between the metal sites [51,52]. Thus, the phylogeniesof the two types of H2ases are to be analyzed separately.
The structural complexity of H2ases and their catalyticsites is demanding in terms of biosynthesis. Several geneproducts are involved in the maturation of the [NiFe] en-zymes [92]. Elucidation of the structure and function ofeach of these proteins is now on its way. In this regard, theunderstanding of [Fe]-H2ases is considerably less ad-vanced, mainly because the genes in that case appear notto be clustered on the chromosomes. An additional di¤-culty in the identi¢cation of the [Fe]-H2ase maturing pro-teins arises from the fact that they appear to bear nosimilarity to the [NiFe]-H2ase maturing proteins. Thiswould be in keeping with the absence of phylogenic rela-
tionship between the [Fe] and [NiFe] enzymes. Con¢rma-tion of this observation by future investigations wouldindicate that the [Fe]- and [NiFe]-H2ases are even moredi¡erent than previously estimated.
Whole genome sequences are increasing signi¢cantly theoutput of H2ase sequence data, but more importantly,they are providing new insights into the distribution ofthese enzymes. Indeed, complete genome sequences, to-gether with other genetic and biochemical data, indicatethat [Fe]-H2ases occur only in Bacteria and Eucarya while[NiFe]-H2ases appear to be present only in Archaea andBacteria. Regarding the latter point, some uncertaintiesremain concerning green algae ([68,70,368,72], Section 6).If these trends are con¢rmed, the data might turn out tobecome valuable touchstones for the elucidation of earlysymbiotic events that led to the emergence of Eucarya. Infact, [Fe]-H2ase sequences from anaerobic eukaryotes arealready being analyzed with the aim of elucidating theevolution of these organisms and of hydrogenosomes[275,337,367].
Another emerging track in H2ase research is linked tothe discovery of H2ase-like sequences in the genomes ofaerobic eukaryotes, including mammals. The encoded pro-teins are clearly related to [Fe]-H2ases, but appear to havelost their nominal catalytic activity. Their present functionis in the process of being elucidated [325], and the evolu-tionary pathways that led to it are likely to be revealed inthe near future.
Yet another fascinating aspect of H2ase structure andevolution is the modular structure of these enzymes. Whilethe protein folds surrounding the catalytic H2-activatingsites are idiosyncrasies of H2ases, these enzymes containadditional domains and subunits that have counterparts inother redox proteins and complexes. The latter includeferredoxins, NADH-ubiquinone oxidoreductase (ComplexI) of respiratory chains, and related membrane-boundcomplexes [60,61]. The relevant protein modules, most ofwhich contain Fe^S clusters, are proof of extensive rela-tionships among all extant energy-transducing complexes.This record contains a wealth of structural and phylogenicinformation, of which the exploitation will be acceleratedby the development of structural genomics.
Acknowledgements
We thank our colleagues Drs. M.W.W. Adams, A.Bo«ck, S. Desai, A. Dupuis, L. Fontaine, B. Friedrich, L.Girbal, M.L. Guerinot, J. Imperial, J. Hackstein, T.Happe, D. Horner, K. Kovacs, O. Lenz, P. Lindblad, G.Perrie©re, G. Rakhely, M. Rousset, T. Ruiz-Argu«eso, R.Thauer, G. Voordouw, and L.-F. Wu for discussions, ad-vice, and communication of unpublished data. We alsoacknowledge the help of Dr. G. Klein for the preparationof Fig. 6.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501490
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

References
[1] Bowien, B. and Schlegel, H.G. (1981) Physiology and biochemistry ofaerobic hydrogen-oxidizing bacteria. Annu. Rev. Microbiol. 350,405^452.
[2] Vignais, P.M., Henry, M.-F., Sim, E. and Kell, D.B. (1981) Theelectron transport system and hydrogenase of Paracoccus denitri¢-cans. Curr. Top. Bioenerg. 12, 115^196.
[3] Lambert, G.R. and Smith, G.D. (1981) The hydrogen metabolism ofcyanobacteria (blue-green algae). Biol. Rev. 56, 589^660.
[4] Houchins, J.P. (1984) The physiology and biochemistry of hydrogenmetabolism in cyanobacteria. Biochim. Biophys. Acta 768, 227^255.
[5] Meyer, J., Kelley, B. and Vignais, P.M. (1978) Nitrogen ¢xation andhydrogen metabolism in photosynthetic bacteria. Biochimie 60, 245^260.
[6] Vignais, P.M., Colbeau, A., Willison, J.C. and Jouanneau, Y. (1985)Hydrogenase, nitrogenase, and hydrogen metabolism in the photo-synthetic bacteria. Adv. Microb. Physiol. 26, 155^234.
[7] Sasikala, K., Ramana, Ch.V., Raghuveer Rao, P. and Kovacs, K.L.(1993) Anoxygenic phototrophic bacteria: physiology and advancesin hydrogen production technology. Adv. Appl. Microbiol. 38, 211^295.
[8] Maier, R.J. (1986) Biochemistry, regulation, and genetics of hydrogenoxidation in Rhizobium. CRC Crit. Rev. Biotechnol. 3, 17^38.
[9] Evans, H.J., Harker, A.R., Papen, H., Russell, S.A., Hanus, F.J. andZuber, M. (1987) Physiology, biochemistry, and genetics of the up-take hydrogenase in Rhizobia. Annu. Rev. Microbiol. 41, 335^361.
[10] Van Soom, C., Rumjanek, N., Vanderleyden, J. and Neves, M.C.P.(1993) Hydrogenase in Bradyrhizobium japonicum : genetics, regula-tion and e¡ect on plant growth. World J. Microbiol. Biotechnol. 9,615^624.
[11] Odom, J.M. and Peck, H.D. (1984) Hydrogenase, electron-transferproteins, and energy coupling in the sulphate-reducing bacteria De-sulfovibrio. Annu. Rev. Microbiol. 38, 551^592.
[12] Fauque, G., Peck Jr., H.D., Moura, J.J.G., Huynh, B.H., Berlier, Y.,DerVertanian, D.V., Teixeira, M., Przybyla, A.E., Lespinat, P.A.,Moura, I. and Le Gall, J. (1988) The three classes of hydrogenasesfrom sulfate-reducing bacteria of the genus Desulfovibrio. FEMSMicrobiol. Rev. 54, 299^344.
[13] Adams, M.W.W., Mortenson, L.E. and Chen, J.-S. (1981) Hydro-genase. Biochim. Biophys. Acta 594, 105^176.
[14] Adams, M.W.W. (1990) The structure and mechanism of iron-hydro-genases. Biochim. Biophys. Acta 1020, 115^145.
[15] Meyer, J. (2000) Clostridial iron^sulphur proteins. J. Mol. Microbiol.Biotechnol. 2, 9^14.
[16] Adams, M.W.W. (1990) The metabolism of hydrogen by extremelythermophilic sulfur-dependent bacteria. FEMS Microbiol. Rev. 75,219^238.
[17] Maier, R.J. (1996) Respiratory metabolism in hyperthermophilic or-ganisms: hydrogenases, sulfur reductases, and electron transport fac-tors that function at temperatures exceeding 100³C. Adv. ProteinChem. 48, 35^99.
[18] Przybyla, A.E., Robbins, J., Menon, N. and Peck Jr., H.D. (1992)Structure^function relationships among the nickel-containing hydro-genases. FEMS Microbiol. Rev. 88, 109^135.
[19] Reeve, J.N. and Becker, G.S. (1990) Conservation of primary struc-ture in prokaryotic hydrogenases. FEMS Microbiol. Rev. 87, 419^424.
[20] Voordouw, G. (1992) Evolution of hydrogenase genes. Adv. Inorg.Chem. 38, 397^422.
[21] Wu, L.-F. and Mandrand, M.A. (1993) Microbial hydrogenases : Pri-mary structure, classi¢cation, signatures and phylogeny. FEMSMicrobiol. Rev. 104, 243^270.
[22] Friedrich, B. and Schwartz, E. (1993) Molecular biology of hydrogenutilization in aerobic chemolithotrophs. Annu. Rev. Microbiol. 47,351^383.
[23] Vignais, P.M. and Toussaint, B. (1994) Molecular Biology of mem-brane bound H2 uptake hydrogenases (Erratum Arch. Microbiol.161, 196). Arch. Microbiol. 161, 1^10.
[24] Hahn, D. and Ku«ck, U. (1994) Biochemical and molecular geneticbasis of hydrogenases. Process. Biochem. Oxf. 29, 633^641.
[25] Appel, J. and Schulz, R. (1998) Hydrogen metabolism in organismswith oxygenic photosynthesis : hydrogenases as important regulatorydevices for a proper redox poising? J. Photochem. Photobiol. 47, 1^11.
[26] Hansel, A. and Lindblad, P. (1998) Towards optimization of cyano-bacteria as biotechnologically relevant producers of molecular hydro-gen, a clean and renewable energy source. Appl. Microbiol. Biotech-nol. 50, 153^160.
[27] Lindblad, P. and Tamagnini, P. (2000) Cyanobacterial hydrogenasesand biohydrogen: Present status and future potential. In: Biohydro-gen II (Miyake, J., Matsunaga, T. and San Pietro, T., Eds.), pp 135^162. Elsevier, Amsterdam.
[28] Robson, R.L. (2001) Biodiversity of hydrogenases. In: Hydrogen asFuel : Learning from Nature (Cammack, R., Frey, M. and Robson,R.L., Eds.), Taylor and Francis, London, in press.
[29] Beinert, H., Holm, R.H. and Mu«nck, E. (1997) Iron^sulfur clusters:nature's modular, multipurpose structures. Science 277, 653^659.
[30] Beinert, H. (2000) Iron^sulfur proteins: ancient structures, still full ofsurprises. J. Biol. Inorg. Chem. 5, 2^15.
[31] Mu«hlenho¡, U. and Lill, R. (2000) Biogenesis of iron^sulfur proteinsin eukaryotes: a novel task of mitochondria that is inherited frombacteria. Biochim. Biophys. Acta 1459, 370^382.
[32] Yuvaniyama, P., Agar, J.N., Cash, V.L., Johnson, M.K. and Dean,D.R. (2000) NifS-directed assembly of a transient [2Fe^2S]cluster within the NifU protein. Proc. Natl. Acad. Sci. USA 97,599^604.
[33] Volbeda, A., Charon, M.H., Piras, C., Hatchikian, E.C., Frey, M.and Fontecilla-Camps, J.C. (1995) Crystal structure of the nickel^iron hydrogenase from Desulfovibrio gigas. Nature 373, 580^587.
[34] Peters, J.W., Lanzilotta, W.N., Lemon, B.J. and Seefeldt, L.C. (1998)X-ray crystal structure of the Fe-only hydrogenase (CpI) from Clos-tridium pasteurianum to 1.8 angstrom resolution. Science 282, 1853^1858.
[35] Nicolet, Y., Piras, C., Legrand, P., Hatchikian, C.E. and Fontecilla-Camps, J.C. (1999) Desulfovibrio desulfuricans iron hydrogenase: thestructure shows unusual coordination to an active site Fe binuclearcenter. Structure 7, 13^23.
[36] Volbeda, A., Garcin, E., Piras, C., de Lacey, A.L., Fernandez, V.M.,Hatchikian, E.C., Frey, M. and Fontecilla-Camps, J.C. (1996) Struc-ture of the [NiFe] hydrogenase active site: Evidence for biologicallyuncommon Fe ligands. J. Am. Chem. Soc. 118, 12989^12996.
[37] Higuchi, Y., Yagi, T. and Yasuoka, N. (1997) Unusual ligand struc-ture in Ni^Fe active center and an additional Mg site in hydrogenaserevealed by high resolution X-ray structure analysis. Structure 5,1671^1680.
[38] Higuchi, Y., Ogata, H., Miki, K., Yasuoka, N. and Yagi, T. (1999)Removal of the bridging ligand atom at the Ni^Fe active site of[NiFe] hydrogenase upon reduction with H2, as revealed by X-raystructure analysis at 1.4 Aî resolution. Structure 7, 549^556.
[39] Garcin, E., Vernede, X., Hatchikian, E.C., Volbeda, A., Frey, M. andFontecilla-Camps, J.C. (1999) The crystal structure of a reduced[NiFeSe] hydrogenase provides an image of the activated catalyticcenter. Structure 7, 557^566.
[40] Peters, J.W. (1999) Structure and mechanism of iron-only hydroge-nases. Curr. Opin. Struct. Biol. 9, 670^676.
[41] Adams, M.W.W. and Stiefel, E.I. (1998) Biological hydrogen produc-tion: not so elementary. Science 282, 1842^1843.
[42] Nicolet, Y., Lemon, B.J., Fontecilla-Camps, J.C. and Peters, J.W.(2000) A novel FeS cluster in Fe-only hydrogenases. Trends Biochem.Sci. 25, 138^143.
[43] Trofanchuk, O., Stein, M., Gessner, C., Lendzian, F., Higuchi, Y.and Lubitz, W. (2000) Single crystal EPR studies of the oxidized
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 491
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

active site of [NiFe] hydrogenase from Desulfovibrio vulgaris Miyaza-ki F. J. Biol. Inorg. Chem. 5, 36^44.
[44] Popescu, C.V. and Mu«nck, E. (1999) Electronic structure of the Hcluster in [Fe]-hydrogenases. J. Am. Chem. Soc. 121, 7877^7884.
[45] De Lacey, A.L., Stadler, C., Cavazza, C., Hatchikian, E.C. and Fer-nandez, V.M. (2000) FTIR characterization of the active site of theFe-hydrogenase from Desulfovobrio desulfuricans. J. Am. Chem. Soc.122, 11232^11233.
[46] Pierik, A.J., Hulstein, M., Hagen, W.R. and Albracht, S.P.J. (1998)A low-spin iron with CN and CO as intrinsic ligands forms the coreof the active site in [Fe]-hydrogenases. Eur. J. Biochem. 258, 572^578.
[47] Pierik, A.J., Roseboom, W., Happe, R.P., Bagley, K.A. and Al-bracht, S.P.J. (1999) Carbon monoxide and cyanide as intrinsic li-gands to iron in the active site of [NiFe]-hydrogenases. Ni-Fe(CN)2CO, Biology's way to activate H2. J. Biol. Chem. 274,3331^3337.
[48] Davidson, G., Choudhury, S.B., Gu, Z., Bose, K., Roseboom, W.,Albracht, S.P.J. and Maroney, M.J. (2000) Structural examination ofthe nickel site in Chromatium vinosum hydrogenase: redox state os-cillations and structural changes accompanying reductive activationand CO binding. Biochemistry 39, 7468^7479.
[49] Kaasjager, V.E., Henderson, R.K., Bouwman, E., Lutz, M., Spek,A.L. and Reedijk, J. (1998) A structural model for [Fe]-only hydro-genases. Angew. Chem. Int. Edn. 37, 1668^1670.
[50] Schmidt, M., Contakes, S.M. and Rauchfuss, T.B. (1999) First gen-eration analogues of the binuclear site in the Fe-only hydrogenases:Fe2(W-SR)2(CO)4(CN)23
2 . J. Am. Chem. Soc. 121, 9736^9737.[51] Darensbourg, M.Y., Lyon, E.J. and Smee, J.J. (2000) The bio-orga-
nometallic chemistry of active site iron in hydrogenases. Coord.Chem. Rev. 206-207, 533^561.
[52] Adams, M.W.W. and Stiefel, E.I. (2000) Organometallic iron: thekey to biological hydrogen metabolism. Curr. Opin. Chem. Biol. 4,214^220.
[53] Bertrand, P., Dole, F., Asso, M. and Guigliarelli, B. (2000) Is there arate-limiting step in the catalytic cycle of [NiFe] hydrogenases? J. Biol.Inorg. Chem. 5, 682^691.
[54] Pilkington, S.J., Skehel, J.M., Gennis, R.B. and Walker, J.E. (1991)Relationship between mitochondrial NADH-ubiquinone reductaseand a bacterial NAD-reducing hydrogenase. Biochemistry 30, 2166^2175.
[55] Fearnley, I.M. and Walker, J.E. (1992) Conservation of sequences ofsubunits of mitochondrial complex I and their relationships withother proteins. Biochim. Biophys. Acta 1140, 105^134.
[56] Albracht, S.P.J. (1993) Intimate relationships of the large and thesmall subunits of all nickel hydrogenases with two nuclear-encodedsubunits of mitochondrial NADH:ubiquinone oxidoreductase. Bio-chim. Biophys. Acta 1144, 221^224.
[57] Albracht, S.P.J. (1994) Nickel hydrogenases: in search of the activesite. Biochim. Biophys. Acta 1188, 167^204.
[58] Albracht, S.P.J. and de Jong, A.M.Ph. (1997) Bovine-heart NADH-ubiquinone oxidoreductase is a monomer with 8 Fe^S clusters and 2FMN groups. Biochim. Biophys. Acta 1318, 92^106.
[59] Ku«nkel, A., Vorholt, J.A., Thauer, R.K. and Hedderich, R. (1998)An Escherichia coli hydrogenase-3-type hydrogenase in methanogenicarchaea. Eur. J. Biochem. 252, 467^476.
[60] Friedrich, T. and Scheide, D. (2000) The respiratory complex I ofbacteria, archaea and eukarya and its module common with mem-brane-bound multisubunit hydrogenases. FEBS Lett. 479, 1^5.
[61] Albracht, S.P.J. and Hedderich, R. (2000) Learning from hydroge-nases: location of a proton pump and of a second FMN in bovineNADH-ubiquinone oxidoreductase (Complex I). FEBS Lett. 485,1^6.
[62] Graf, E.-G. and Thauer, R.K. (1981) Hydrogenase from Methano-bacterium thermoautotrophicum, a nickel-containing enzyme. FEBSLett. 136, 165^169.
[63] Woese, C.R. (1994) There must be a prokaryote somewhere. Micro-biology's search for itself. Microbiol. Rev. 58, 1^9.
[64] Mu«ller, M. (1993) The hydrogenosome. J. Gen. Microbiol. 139,2879^2889.
[65] Page-Sharp, M., Behm, C.A. and Smith, G.D. (1996) Tritrichomonasfoetus and Trichomonas vaginalis : the pattern of inactivation of hy-drogenase activity by oxygen and activities of catalase and ascorbateperoxidase. Microbiology 142, 207^211.
[66] Happe, T. and Naber, J.D. (1993) Isolation, characterization and N-terminal amino acid sequence of hydrogenase from the green algaChlamydomonas reinhardtii. Eur. J. Biochem. 214, 475^481.
[67] Happe, T., Mosler, B. and Naber, J.D. (1994) Induction, localizationand metal content of hydrogenase in the green alga Chlamydomonasreinhardtii. Eur. J. Biochem. 222, 769^774.
[68] Schnackenberg, J., Schulz, R. and Senger, H. (1993) Characterizationand puri¢cation of a hydrogenase from the eukaryotic green algaScenedesmus obliquus. FEBS Lett. 327, 21^24.
[69] Urbig, T., Schulz, R. and Senger, H. (1993) Inactivation and reacti-vation of the hydrogenases of the green algae Scenedesmus obliquusand Chlamydomonas reinhardtii. Z. Naturforsch. 48c, 41^45.
[70] Zinn, T., Schnackenberg, J., Haak, D., Roemer, S., Schulz, R. andSenger, H. (1994) Evidence for nickel in the soluble hydrogenasefrom the unicellular green alga Scenedesmus obliquus. Z. Naturforsch.49c, 33^38.
[71] Ueno, Y., Kurano, N. and Miyachi, S. (1999) Puri¢cation and char-acterization of hydrogenase from the marine green alga, Chlorococ-cum littorale. FEBS Lett. 443, 144^148.
[72] Florin, L., Tsokoglou, A. and Happe, T. (2000) A novel type of Fe-hydrogenase in the green alga Scenedesmus obliquus is linked to thephotosynthetic electron transport chain. J. Biol. Chem. 276, 6125^6132.
[73] Chen, J.-S. and Mortenson, L.E. (1974) Puri¢cation and properties ofhydrogenase from Clostridium pasteurianum W5. Biochim. Biophys.Acta 371, 283^298.
[74] He, S.H., Teixeira, M., LeGall, J., Patil, D.S., Moura, I., Moura, J.J.,DerVartanian, D.V., Huynh, B.H. and Peck Jr., H.D. (1989) EPRstudies with 77Se-enriched (NiFeSe) hydrogenase of Desulfovibrio ba-culatus. Evidence for a selenium ligand to the active site nickel. J. Biol.Chem. 264, 2678^2682.
[75] Sorgenfrei, O., Klein, A. and Albracht, S.P.J. (1993) In£uence ofillumination on the electronic interaction between 77Se and nickelin active F420-non-reducing hydrogenase from Methanococcus voltae.FEBS Lett. 332, 291^297.
[76] Zirngibl, C., Hedderich, R. and Thauer, R.K. (1990) N5, N10-Meth-ylenetetrahydromethanopterin dehydrogenase from Methanobacte-rium thermoautotrophicum has hydrogenase activity. FEBS Lett.261, 112^116.
[77] Hartmann, G.C., Klein, A.R., Linder, M. and Thauer, R.K. (1996)Puri¢cation, properties and primary structure of H2-forming N5,N10-methylenetetrahydromethanopterin dehydrogenase from Metha-nococcus thermolithotrophicus. Arch. Microbiol. 165, 187^193.
[78] Hagemeier, C.H., Chistoserdova, L., Lidstrom, M.E., Thauer, R.K.and Vorholt, J.A. (2000) Characterization of a second methylenetetrahydromethanopterin dehydrogenase from Methylobacterium ex-torquens AM1. Eur. J. Biochem. 267, 3762^3769.
[79] Berkessel, A. and Thauer, R.K. (1995) On the mechanism of catalysisby a metal-free hydrogenase from methanogenic archaea: enzymictransformation of H2 without a metal and its analogy to the chem-istry of alkanes in superacidic solution. Angew. Chem. Int. Edn.Engl. 34, 2247^2250.
[80] Thauer, R.K., Klein, A.R. and Hartmann, G.C. (1996) Reactionswith molecular hydrogen in microorganisms: evidence for a purelyorganic hydrogenation catalyst. Chem. Rev. 96, 3031^3042.
[81] Thauer, R.K. (1998) Biochemistry of methanogenesis: a tribute toMarjory Stephenson. 1998 Marjory Stephenson Prize Lecture. Micro-biology 144, 2377^2406.
[82] Schwoerer, B., Fernandez, V.M., Zirngibl, C. and Thauer, R.K.(1993) Hydrogen-forming N5,N10-methylenetetrahydromethanopterindehydrogenase from Methanobacterium thermoautotrophicum. Studies
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501492
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

of the catalytic mechanism of hydrogen formation using hydrogenisotopes. Eur. J. Biochem. 212, 255^261.
[83] Klein, A.R., Fernandez, V.M. and Thauer, R.K. (1995) H2-formingN5,N10-methylenetetrahydromethanopterin dehydrogenase: mecha-nism of H2 formation analyzed using hydrogen isotopes. FEBSLett. 368, 203^206.
[84] Schleucher, J., Griesinger, C., Schwoerer, B. and Thauer, R.K.(1994) H2-forming N5,N10-methylenetetrahydromethanopterin de-hydrogenase from Methanobacterium thermoautotrophicum catalyzesa stereoselective hydride transfer as determined by two-dimensionalNMR spectroscopy. Biochemistry 33, 3986^3993.
[85] Reeve, J.N., No«lling, J., Morgan, R.M. and Smith, D.R. (1997)Methanogenesis: genes, genomes, and who's on ¢rst? J. Bacteriol.179, 5975^5986.
[86] Afting, C., Hochheimer, A. and Thauer, R.K. (1998) Function ofH2-forming methylenetetrahydromethanopterin dehydrogenase fromMethanobacterium thermoautotrophicum in coenzyme F420 reductionwith H2. Arch. Microbiol. 169, 206^210.
[87] Buurman, G., Shima, S. and Thauer, R.K. (2000) The metal-freehydrogenase from methanogenic archae: evidence for a bound co-factor. FEBS Lett. 485, 200^204.
[88] Benemann, J.R. and Weare, N.M. (1974) Nitrogen ¢xation by Ana-baena cylindrica. III. Hydrogen-supported nitrogenase activity.Arch. Microbiol. 101, 401^408.
[89] Bothe, H., Distler, E. and Eisbrenner, G. (1978) Hydrogen metab-olism in blue-green algae. Biochimie 60, 277^289.
[90] Walker, C.C. and Yates, M.G. (1978) The hydrogen cycle in nitro-gen-¢xing Azotobacter chroococcum. Biochimie 60, 225^231.
[91] Odom, J.M. and Peck Jr., H.D (1981) Hydrogen cycling as a generalmechanism for energy coupling in the sulfate-reducing bacteria De-sulfovibrio sp. FEMS Lett. 12, 47^50.
[92] Casalot, L. and Rousset, M. (2001) Maturation of the [NiFe] hydro-genases. Trends Microbiol. 9, 228^237.
[93] Leclerc, M., Colbeau, A., Cauvin, B. and Vignais, P.M. (1988)Cloning and sequencing of the genes encoding the large and thesmall subunits of the H2 uptake hydrogenase (hup) of Rhodobactercapsulatus (Erratum (1989) 215, 368). Mol. Gen. Genet. 214, 97^107.
[94] Richaud, P., Vignais, P.M., Colbeau, A., U¡en, R.L. and Cauvin,B. (1990) Molecular biology of the uptake hydrogenase of Rhodo-bacter capsulatus and Rhodocyclus gelatinosus. FEMS Microbiol.Rev. 87, 413^418.
[95] Cauvin, B., Colbeau, A. and Vignais, P.M. (1991) The hydrogenasestructural operon in Rhodobacter capsulatus contains a third gene,hupM, necessary for the formation of a physiologically competenthydrogenase. Mol. Microbiol. 5, 2519^2527.
[96] Colbeau, A., Richaud, P., Toussaint, B., Caballero, J., Elster, C.,Delphin, C., Smith, R.L., Chabert, J. and Vignais, P.M. (1993)Organization of the genes necessary for hydrogenase expression inRhodobacter capsulatus. Sequence analysis and identi¢cation of twohyp regulatory mutants. Mol. Microbiol. 8, 15^29.
[97] Xu, H.W. and Wall, J.D. (1991) Clustering of genes necessary forhydrogen oxidation in Rhodobacter capsulatus. J. Bacteriol. 173,2401^2405.
[98] Richaud, P., Colbeau, A., Toussaint, B. and Vignais, P.M. (1991)Identi¢cation and sequence analysis of the hupR1 gene, which en-codes a response regulator of the NtrC family required for hydro-genase expression in Rhodobacter capsulatus. J. Bacteriol. 173, 5928^5932.
[99] Elsen, S., Richaud, P., Colbeau, A. and Vignais, P.M. (1993) Se-quence analysis and interposon mutagenesis of the hupT gene, whichencodes a sensor protein involved in repression of hydrogenase syn-thesis in Rhodobacter capsulatus. J. Bacteriol. 175, 7404^7412.
[100] Elsen, S., Colbeau, A., Chabert, J. and Vignais, P.M. (1996) ThehupTUV operon is involved in negative control of hydrogenase syn-thesis in Rhodobacter capsulatus. J. Bacteriol. 178, 5174^5181.
[101] Elsen, S., Colbeau, A. and Vignais, P.M. (1997) Puri¢cation and invitro phosphorylation of HupT, a regulatory protein controlling
hydrogenase gene expression in Rhodobacter capsulatus. J. Bacteriol.179, 968^971.
[102] Vignais, P.M., Dimon, B., Zorin, N.A., Colbeau, A. and Elsen, S.(1997) HupUV proteins of Rhodobacter capsulatus can bind H2 :evidence from the H-D exchange reaction. J. Bacteriol. 179, 290^292.
[103] Toussaint, B., de Sury d'Aspremont, R., Delic-Attree, I., Berchet,V., Elsen, S., Colbeau, A., Dischert, W., Lazzaroni, Y. and Vignais,P.M. (1997) The Rhodobacter capsulatus hupSLC promoter: identi-¢cation of cis-regulatory elements and of trans-activating factorsinvolved in H2 activation of hupSLC transcription. Mol. Microbiol.26, 927^937.
[104] Dischert, W., Vignais, P.M. and Colbeau, A. (1999) The synthesis ofRhodobacter capsulatus HupSL hydrogenase is regulated by the two-component HupT/HupR system. Mol. Microbiol. 34, 995^1006.
[105] Colbeau, A., Elsen, S., Tomiyama, M., Zorin, N.A., Dimon, B. andVignais, P.M. (1998) Rhodobacter capsulatus HypF is involved inregulation of hydrogenase synthesis through the HupUV proteins.Eur. J. Biochem. 251, 65^71.
[106] Vignais, P.M., Dimon, B., Zorin, N.A., Tomiyama, M. and Col-beau, A. (2000) Characterization of the hydrogen-deuterium ex-change activities of the energy-transducing HupSL hydrogenaseand the H2-signaling HupUV hydrogenase in Rhodobacter capsula-tus. J. Bacteriol. 182, 5997^6004.
[107] Gomelsky, M. and Kaplan, S. (1995) Isolation of regulatory mu-tants in photosynthesis gene expression in Rhodobacter sphaeroides2.4.1 and partial complementation of a PrrB mutant by the HupThistidine-kinase. Microbiology 141, 1805^1819.
[108] Hidalgo, E., Leyva, A. and Ruiz-Argu«eso, T. (1990) Nucleotide se-quence of the hydrogenase structural genes from Rhizobium legumi-nosarum. Plant Mol. Biol. 15, 367^370.
[109] Schneider, C.G., Schmitt, H.J., Schild, C., Tichy, H.V. and Lotz, W.(1990) DNA sequence encoding the two structural genes for theuptake hydrogenase of Rhizobium leguminosarum bv. viciae B10.Nucleic Acids Res. 18, 5285.
[110] Hidalgo, E., Palacios, J.M., Murillo, J. and Ruiz-Argu«eso, T. (1992)Nucleotide sequence and characterization of four additional genesof the hydrogenase structural operon from Rhizobium leguminosa-rum bv. viciae. J. Bacteriol. 174, 4130^4139.
[111] Rey, L., Hidalgo, E., Palacios, J. and Ruiz-Argu«eso, T. (1992) Nu-cleotide sequence and organization of an H2-uptake gene clusterfrom Rhizobium leguminosarum bv. viciae containing a rubredoxin-like gene and four additional open reading frames. J. Mol. Biol. 228,998^1002.
[112] Rey, L., Murillo, J., Hernando, Y., Hidalgo, E., Cabrera, E., Im-perial, J. and Ruiz-Argu«eso, T. (1993) Molecular analysis of amicroaerobically induced operon required for hydrogenase synthesisin Rhizobium leguminosarum biovar viciae. Mol. Microbiol. 8, 471^481.
[113] Imperial, J., Rey, L., Palacios, J.M. and Ruiz-Argu«eso, T. (1993)HupK, a hydrogenase-ancillary protein from Rhizobium leguminosa-rum, shares structural motifs with the large subunit of NiFe hydro-genases and could be a sca¡olding protein for hydrogenase metalcofactor assembly. Mol. Microbiol. 9, 1305^1306.
[114] Rey, L., Imperial, J., Palacios, J.M. and Ruiz-Argu«eso, T. (1994)Puri¢cation of Rhizobium leguminosarum HypB, a nickel-bindingprotein required for hydrogenase synthesis. J. Bacteriol. 176,6066^6073.
[115] Hernando, Y., Palacios, J.M., Imperial, J. and Ruiz-Argu«eso, T.(1995) The hypBFCDE operon from Rhizobium leguminosarum bio-var viciae is expressed from an Fnr-type promoter that escapesmutagenesis of the fnrN gene. J. Bacteriol. 177, 5661^5669.
[116] Brito, B., Mart|nez, M., Fernandez, D., Rey, L., Cabrera, E., Pal-acios, J.M., Imperial, J. and Ruiz-Argu«eso, T. (1997) Hydrogenasegenes from Rhizobium leguminosarum bv. viciae are controlled bythe nitrogen ¢xation regulatory protein NifA. Proc. Natl. Acad. Sci.USA 94, 6019^6024.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 493
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

[117] Rey, L., Fernandez, D., Brito, B., Hernando, Y., Palacios, J.M.,Imperial, J. and Ruiz-Argu«eso, T. (1996) The hydrogenase genecluster of Rhizobium leguminosarum bv. viciae contains an additionalgene (hypX), which encodes a protein with sequence similarity to theN10-formyltetrahydrofolate-dependent enzyme family and is re-quired for nickel-dependent hydrogenase processing and activity.Mol. Gen. Genet. 252, 237^248.
[118] Hernando, Y., Palacios, J., Imperial, J. and Ruiz-Argu«eso, T. (1998)Rhizobium leguminosarum bv. viciae hypA gene is speci¢cally ex-pressed in pea (Pisum sativum) bacteroids and required for hydro-genase activity and processing. FEMS Microbiol. Lett. 169, 295^302.
[119] Sayavedra-Soto, L.A., Powell, G.K., Evans, H.J. and Morris, R.O.(1988) Nucleotide sequence of the genetic loci encoding subunits ofBradyrhizobium japonicum uptake hydrogenase. Proc. Natl. Acad.Sci. USA 85, 8395^8399.
[120] Van Soom, C., Browaeys, J., Verreth, C. and Vanderleyden, J.(1993) Nucleotide sequence analysis of four genes, hupC, hupD,hupF and hupG, downstream of the hydrogenase structural genesin Bradyrhizobium japonicum. J. Mol. Biol. 234, 508^512.
[121] Van Soom, C., Verreth, C., Sampaio, M.J. and Vanderleyden, J.(1993) Identi¢cation of a potential transcriptional regulator of hy-drogenase activity in free-living Bradyrhizobium japonicum strains.Mol. Gen. Genet. 239, 235^240.
[122] Black, L.K., Fu, C. and Maier, R.J. (1994) Sequences and character-ization of hupU and hupV genes of Bradyrhizobium japonicum encod-ing a possible nickel-sensing complex involved in hydrogenase ex-pression. J. Bacteriol. 176, 7102^7106.
[123] Fu, C. and Maier, R.J. (1994) Sequence and characterization ofthree genes within the hydrogenase gene cluster of Bradyrhizobiumjaponicum. Gene 141, 47^52.
[124] Fu, C. and Maier, R.J. (1994) Organization of the hydrogenase genecluster from Bradyrhizobium japonicum : sequences and analysis of¢ve more hydrogenase-related genes. Gene 145, 91^96.
[125] Fu, C. and Maier, R.J. (1994) Nucleotide sequences of two hydro-genase-related genes (hypA and hypB) from Bradyrhizobium japoni-cum, one of which (hypB) encodes an extremely histidine-rich regionand guanine nucleotide-binding domains. Biochim. Biophys. Acta1184, 135^138.
[126] Fu, C., Olson, J.W. and Maier, R.J. (1995) HypB protein of Bra-dyrhizobium japonicum is a metal-binding GTPase capable of bind-ing 18 divalent nickel ions per dimer. Proc. Natl. Acad. Sci. USA 92,2333^2337.
[127] Van Soom, C., de Wilde, P. and Vanderleyden, J. (1997) HoxA is atranscriptional regulator for expression of the hup structural genesin free-living Bradyrhizobium japonicum. Mol. Microbiol. 23, 967^977.
[128] Durmowicz, M.C. and Maier, R.J. (1997) Roles of HoxX and HoxAin biosynthesis of hydrogenase in Bradyrhizobium japonicum. J. Bac-teriol. 179, 3676^3682.
[129] Olson, J.W., Fu, C. and Maier, R.J. (1997) The HypB protein fromBradyrhizobium japonicum can store nickel and is required for thenickel-dependent transcriptional regulation of hydrogenase. Mol.Microbiol. 24, 119^128.
[130] Olson, J.W. and Maier, R.J. (1997) The sequences of hypF, hypCand hypD complete the hyp gene cluster required for hydrogenaseactivity in Bradyrhizobium japonicum. Gene 199, 93^99.
[131] Van Soom, C., Lerouge, I., Vanderleyden, J., Ruiz-Argu«eso, T. andPalacios, J.M. (1999) Identi¢cation and characterization of hupT, agene involved in negative regulation of hydrogen oxidation in Bra-dyrhizobium japonicum. J. Bacteriol. 181, 5085^5089.
[132] Olson, J.W. and Maier, R.J. (2000) Dual roles of Bradyrhizobiumjaponicum nickelin protein in nickel storage and GTP-dependent Nimobilization. J. Bacteriol. 182, 1702^1705.
[133] Tran-Betcke, A., Warnecke, U., Boecker, C., Zaborosch, C. andFriedrich, B. (1990) Cloning and nucleotide sequences of the genesfor the subunits of NAD-reducing hydrogenase of Alcaligenes eutro-phus H16. J. Bacteriol. 172, 2920^2929.
[134] Eberz, G. and Friedrich, B. (1991) Three trans-acting regulatoryfunctions control hydrogenase synthesis in Alcaligenes eutrophus.J. Bacteriol. 173, 1845^1854.
[135] Kortlu«ke, C., Horstmann, K., Schwartz, E., Rohde, M., Binsack, R.and Friedrich, B. (1992) A gene complex coding for the membrane-bound hydrogenase of Alcaligenes eutrophus H16. J. Bacteriol. 174,6277^6289.
[136] Dernedde, J., Eitinger, M. and Friedrich, B. (1993) Analysis of apleiotropic gene region involved in formation of catalytically activehydrogenases in Alcaligenes eutrophus H16. Arch. Microbiol. 159,545^553.
[137] Dernedde, J., Eitinger, T., Patenge, N. and Friedrich, B. (1996) hypgene products in Alcaligenes eutrophus are part of a hydrogenase-maturation system. Eur. J. Biochem. 235, 351^358.
[138] Lenz, O., Schwartz, E., Dernedde, J., Eitinger, M. and Friedrich, B.(1994) The Alcaligenes eutrophus H16 hoxX gene participates inhydrogenase regulation. J. Bacteriol. 176, 4385^4393.
[139] Zimmer, D., Schwartz, E., Tran-Betcke, A., Gewinner, P. and Frie-drich, B. (1995) Temperature tolerance of hydrogenase expression inAlcaligenes eutrophus is conferred by a single amino acid exchangein the transcriptional activator HoxA. J. Bacteriol. 177, 2373^2380.
[140] Bernhard, M., Schwartz, E., Rietdorf, J. and Friedrich, B. (1996)The Alcaligenes eutrophus membrane-bound hydrogenase gene locusencodes functions involved in maturation and electron transportcoupling. J. Bacteriol. 178, 4522^4529.
[141] Bernhard, M., Benelli, B., Hochkoeppler, A., Zannoni, D. and Frie-drich, B. (1997) Functional and structural role of the cytochrome bsubunit of the membrane-bound hydrogenase complex of Alcali-genes eutrophus H16. Eur. J. Biochem. 248, 179^186.
[142] Thiemermann, S., Dernedde, J., Bernhard, M., Schroeder, W., Mas-sanz, C. and Friedrich, B. (1996) Carboxyl-terminal processing ofthe cytoplasmic NAD-reducing hydrogenase of Alcaligenes eutro-phus requires the hoxW gene product. J. Bacteriol. 178, 2368^2374.
[143] Wolf, I., Buhrke, T., Dernedde, J., Pohlmann, A. and Friedrich, B.(1998) Duplication of hyp genes involved in maturation of [NiFe]hydrogenases in Alcaligenes eutrophus H16. Arch. Microbiol. 170,451^459.
[144] Lenz, O., Strack, A., Tran-Betcke, A. and Friedrich, B. (1997) Ahydrogen-sensing system in transcriptional regulation of hydroge-nase gene expression in Alcaligenes species. J. Bacteriol. 179,1655^1663.
[145] Massanz, C., Schmidt, S. and Friedrich, B. (1998) Subforms and invivo reconstitution of the NAD-reducing hydrogenase of Alcaligeneseutrophus. J. Bacteriol. 180, 1023^1029.
[146] Lenz, O. and Friedrich, B. (1998) A novel multicomponent regula-tory system mediates H2 sensing in Alcaligenes eutrophus. Proc.Natl. Acad. Sci. USA 95, 12474^12479.
[147] Buhrke, T. and Friedrich, B. (1998) hoxX (hypX) is a functionalmember of the Alcaligenes eutrophus hyp gene cluster. Arch. Micro-biol. 170, 460^463.
[148] Massanz, C., Fernandez, V.M. and Friedrich, B. (1997) C-terminalextension of the H2-activating subunit, HoxH, directs maturation ofthe NAD-reducing hydrogenase in Alcaligenes eutrophus. Eur. J.Biochem. 245, 441^448.
[149] Massanz, C. and Friedrich, B. (1999) Amino acid replacements atthe H2-activating site of the NAD-reducing hydrogenase from Alca-ligenes eutrophus. Biochemistry 38, 14330^14337.
[150] Schwartz, E., Gerischer, U. and Friedrich, B. (1998) Transcriptionalregulation of Alcaligenes eutrophus hydrogenase genes. J. Bacteriol.180, 3197^3204.
[151] Schwartz, E., Buhrke, T., Gerischer, U. and Friedrich, B. (1999)Positive transcriptional feedback controls hydrogenase expressionin Alcaligenes eutrophus H16. J. Bacteriol. 181, 5684^5692.
[152] Kleihues, L., Lenz, O., Bernhard, M., Buhrke, T. and Friedrich, B.(2000) The H2 sensor of Ralstonia eutropha is a member of thesubclass of regulatory [NiFe] hydrogenases. J. Bacteriol. 182,2716^2724.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501494
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

[153] Menon, A.L., Stults, L.W., Robson, R.L. and Mortenson, L.E.(1990) Cloning, sequencing and characterization of the [NiFe] hy-drogenase-encoding structural genes (hoxK and hoxG) from Azoto-bacter vinelandii. Gene 96, 67^74.
[154] Menon, A.L., Mortenson, L.E. and Robson, R.L. (1992) Nucleotidesequences and genetic analysis of hydrogen oxidation (hox) genes inAzotobacter vinelandii. J. Bacteriol. 174, 4549^4557.
[155] Chen, J.C. and Mortenson, L.E. (1992) Two open reading frames(ORFs) identi¢ed near the hydrogenase structural genes in Azoto-bacter vinelandii, the ¢rst ORF may encode for a polypeptide similarto rubredoxins. Biochim. Biophys. Acta 1131, 122^124.
[156] Chen, J.C. and Mortenson, L.E. (1992) Identi¢cation of six openreading frames from a region of the Azotobacter vinelandii genomelikely involved in dihydrogen metabolism. Biochim. Biophys. Acta1131, 199^202.
[157] Sayavedra-Soto, L.A. and Arp, D.J. (1992) The hoxZ gene of theAzotobacter vinelandii hydrogenase operon is required for activationof hydrogenase. J. Bacteriol. 174, 5295^5301.
[158] Gollin, D.J., Mortenson, L.E. and Robson, R.L. (1992) Carboxyl-terminal processing may be essential for production of activeNiFe hydrogenase in Azotobacter vinelandii. FEBS Lett. 309, 371^375.
[159] Menon, A.L. and Robson, R.L. (1994) In vivo and in vitro nickel-dependent processing of the [NiFe] hydrogenase in Azotobacter vine-landii. J. Bacteriol. 176, 291^295.
[160] Sayavedra-Soto, L.A. and Arp, D.J. (1993) In Azotobacter vinelandiihydrogenase, substitution of serine for the cysteine residues at posi-tions 62, 65, 294, and 297 in the small (HoxK) subunit a¡ects H2
oxidation. J. Bacteriol. 175, 3414^3421.[161] Garg, R.P., Menon, A.L., Jacobs, K., Robson, R.M. and Robson,
R.L. (1994) The hypE gene completes the gene cluster for H2-oxi-dation in Azotobacter vinelandii. J. Mol. Biol. 236, 390^396.
[162] Chen, J.C., Mortenson, L.E. and Seefeldt, L.C. (1995) Analysis of agene region required for dihydrogen oxidation in Azotobacter vine-landii. Curr. Microbiol. 30, 351^355.
[163] Meek, L. and Arp, D.J. (2000) The hydrogenase cytochrome b hemeligands of Azotobacter vinelandii are required for full H2 oxidationcapability. J. Bacteriol. 182, 3429^3436.
[164] Ford, C.M., Garg, N., Garg, R.P., Tibelius, K.H., Yates, M.G.,Arp, D.J. and Seefeldt, L.C. (1990) The identi¢cation, characteriza-tion, sequencing and mutagenesis of the genes (hupSL) encoding thesmall and large subunits of the H2-uptake hydrogenase of Azoto-bacter chroococcum. Mol. Microbiol. 4, 999^1008.
[165] Du, L., Stejskal, F. and Tibelius, K.H. (1992) Characterization oftwo genes (hupD and hupE) required for hydrogenase activity inAzotobacter chroococcum. FEMS Microbiol. Lett. 75, 93^101.
[166] Tibelius, K.H., Du, L., Tito, D. and Stejskal, F. (1993) The Azoto-bacter chroococcum hydrogenase gene cluster : sequences and geneticanalysis of four accessory genes, hupA, hupB, hupY and hupC.Gene 127, 53^61.
[167] Du, L., Tibelius, K.H., Souza, E.M., Garg, R.P. and Yates, M.G.(1994) Sequences, organization and analysis of the hupZM-NOQRTV genes from the Azotobacter chroococcum hydrogenasegene cluster. J. Mol. Biol. 243, 549^557.
[168] Stoker, K., Reijnders, W.N.M., Oltmann, L.F. and Stouthamer,A.H. (1989) Initial cloning and sequencing of hydHG, an operonhomologous to ntrB and regulating the labile hydrogenase activityin Escherichia coli K-12. J. Bacteriol. 171, 4448^4456.
[169] Menon, N.K., Robbins, J., Peck Jr., H.D., Chatelus, C.Y., Choi,E.S. and Przybyla, A.E. (1990) Cloning and sequencing of a putativeEscherichia coli [NiFe] hydrogenase-1 operon containing six openreading frames. J. Bacteriol. 172, 1969^1977.
[170] Menon, N.K., Robbins, J., Wendt, J.C., Shanmugam, K.T. andPrzybyla, A.E. (1991) Mutational analysis and characterization ofthe Escherichia coli hya operon, which encodes [NiFe] hydrogenase1. J. Bacteriol. 173, 4851^4861.
[171] Tomiyama, M., Shiotani, M., Sode, K., Tamiya, E. and Karube, I.
(1991) Nucleotide sequence analysis and expression control of hydAin Escherichia coli. Abstr. 3rd Int. Conf. Mol. Biol. Hydrogenases,Troia, Portugal, pp. 24^25.
[172] Lutz, S., Jacobi, A., Schlensog, V., Bo«hm, R., Sawers, G. and Bo«ck,A. (1991) Molecular characterisation of an operon (hyp) necessaryfor the activity of the three hydrogenase isoenzymes in Escherichiacoli. Mol. Microbiol. 5, 123^135.
[173] Jacobi, A., Rossmann, R. and Bo«ck, A. (1992) The hyp operon geneproducts are required for the maturation of catalytically active hy-drogenase isoenzymes in Escherichia coli. Arch. Microbiol. 158, 444^451.
[174] Menon, N.K., Chatelus, C.Y., Dervartanian, M., Wendt, J.C.,Shanmugam, K.T., Peck Jr., H.D. and Przybyla, A.E. (1994) Clon-ing, sequencing, and mutational analysis of the hyb operon encodingEscherichia coli hydrogenase 2. J. Bacteriol. 176, 4416^4423.
[175] Bo«hm, R., Sauter, M. and Bo«ck, A. (1990) Nucleotide sequence andexpression of an operon in Escherichia coli coding for formate hy-drogenlyase components. Mol. Microbiol. 4, 231^243.
[176] Sauter, M., Bo«hm, R. and Bo«ck, A. (1992) Mutational analysis ofthe operon (hyc) determining hydrogenase 3 formation in Escheri-chia coli. Mol. Microbiol. 6, 1523^1532.
[177] Maier, T., Jacobi, A., Sauter, M. and Bo«ck, A. (1993) The productof the hypB gene, which is required for nickel incorporation intohydrogenases, is a novel guanine nucleotide-binding protein. J. Bac-teriol. 175, 630^635.
[178] Maier, T., Lottspeich, F. and Bo«ck, A. (1995) GTP hydrolysis byHypB is essential for nickel insertion into hydrogenases of Escheri-chia coli. Eur. J. Biochem. 230, 133^138.
[179] Rossmann, R., Sauter, M., Lottspeich, F. and Bo«ck, A. (1994) Mat-uration of the large subunit (HYCE) of Escherichia coli hydrogenase3 requires nickel incorporation followed by C-terminal processing atArg537. Eur. J. Biochem. 220, 377^384.
[180] Rossmann, R., Maier, T., Lottspeich, F. and Bo«ck, A. (1995) Char-acterisation of a protease from Escherichia coli involved in hydro-genase maturation. Eur. J. Biochem. 227, 545^550.
[181] Maier, T., Binder, U. and Bo«ck, A. (1996) Analysis of the hydAlocus of Escherichia coli : two genes (hydN and hypF) involved informate and hydrogen metabolism. Arch. Microbiol. 165, 333^341.
[182] Binder, U., Maier, T. and Bo«ck, A. (1996) Nickel incorporation intohydrogenase 3 from Escherichia coli requires the precursor form ofthe large subunit. Arch. Microbiol. 165, 69^72.
[183] Sargent, F., Ballantine, S.P., Rugman, P.A., Palmer, T. and Boxer,D.H. (1998) Reassignment of the gene encoding the Escherichia colihydrogenase 2 small subunit. Identi¢cation of a soluble precursorof the small subunit in a hypB mutant. Eur. J. Biochem. 255, 746^754.
[184] Magalon, A. and Bo«ck, A. (2000) Dissection of the maturationreactions of the [NiFe] hydrogenase 3 from Escherichia coli takingplace after nickel incorporation. FEBS Lett. 473, 254^258.
[185] Magalon, A. and Bo«ck, A. (2000) Analysis of the HypC-HycE com-plex, a key intermediate in the assembly of the metal center of theEscherichia coli hydrogenase 3. J. Biol. Chem. 275, 21114^21120.
[186] Fritsche, E., Paschos, A., Beisel, H.G., Bo«ck, A. and Huber, R.(1999) Crystal structure of the hydrogenase maturating endopepti-dase HYBD from Escherichia coli. J. Mol. Biol. 288, 989^998.
[187] Theodoratou, E., Paschos, A., Mintz-Weber, S. and Bo«ck, A. (2000)Analysis of the cleavage site speci¢city of the endopeptidase in-volved in the maturation of the large subunit of hydrogenase 3from Escherichia coli. Arch. Microbiol. 173, 110^116.
[188] Theodoratou, E., Paschos, A., Magalon, A., Fritsche, E., Huber, R.and Bo«ck, A. (2000) Nickel serves as a substrate recognition motiffor the endopeptidase involved in hydrogenase maturation. Eur. J.Biochem. 267, 1995^1999.
[189] Colbeau, A., Kovacs, K.L., Chabert, J. and Vignais, P.M. (1994)Cloning and sequences of the structural (hupSLC) and accessory(hupDHI) genes for hydrogenase biosynthesis in Thiocapsa roseoper-sicina. Gene 140, 25^31.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 495
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

[190] Rakhely, G., Colbeau, A., Garin, J., Vignais, P.M. and Kovacs,K.L. (1998) Unusual organization of the genes coding for HydSL,the stable [NiFe] hydrogenase in the photosynthetic bacterium Thi-ocapsa roseopersicina BBS. J. Bacteriol. 180, 1460^1465.
[191] Ohtsuki, T., Okazaki, T., Endo, Y., Kita, Y., Shimosaka, M. andOkazaki, M. (1995) Cloning and sequencing of the membrane-bound hydrogenase-encoding genes (hupS and hupL) from Pseudo-monas hydrogenovora. Gene 166, 89^93.
[192] Ohtsuki, T., Shimosaka, M. and Okazaki, M. (1997) Expression ofgenes encoding membrane-bound hydrogenase in Pseudomonas hy-drogenovora under autotrophic condition is dependent on two di¡er-ent promoters. Biosci. Biotechnol. Biochem. 61, 1986^1990.
[193] Ohtsuki, T., Kita, Y., Fujioka, T., Hashimoto, D., Shimosaka, M.and Okazaki, M. (1997) The hupC gene product is a component ofthe electron transport system for hydrogen oxidation in Pseudomo-nas hydrogenovora. FEMS Microbiol. Lett. 150, 127^133.
[194] Paschos, A., Glass, R.S. and Bo«ck, A. (2001) Carbamoylphosphaterequirement for synthesis of the active center of [NiFe]-hydroge-nases. FEBS Lett. 488, 9^12.
[195] Schmitt, E., Blanquet, S. and Mechulam, Y. (1996) Structure ofcrystalline Escherichia coli methionyl-tRNAMet formyltransferase:comparison with glycinamide ribonucleotide formyltransferase.EMBO J. 15, 4749^4758.
[196] Maier, T. and Bo«ck, A. (1996) Generation of active [NiFe] hydro-genase in vitro from a nickel-free precursor form. Biochemistry 35,10089^10093.
[197] Drapal, N. and Bo«ck, A. (1998) Interaction of the hydrogenaseaccessory protein HypC with HycE, the large subunit of Escherichiacoli hydrogenase 3 during enzyme maturation. Biochemistry 37,2941^2948.
[198] Sorgenfrei, O., Linder, D., Karas, M. and Klein, A. (1993) A novelvery small subunit of a selenium containing [nickel^iron] hydroge-nase of Methanococcus voltae is postranslationally processed bycleavage at a de¢ned position. Eur. J. Biochem. 213, 1355^1358.
[199] Maier, T. and Bo«ck, A. (1996) Nickel incorporation into hydroge-nases. In: Advances in Inorganic Biochemistry: Mechanisms ofMetallocenter Assembly (Hausinger, R.P., Eichhorn, G.L. and Mar-zilli, L.G., Eds.), pp 173^192. VCH, New York.
[200] Olson, J.W., Mehta, N.S. and Maier, R.J. (2001) Requirement ofnickel metabolism proteins HypA and HypB for full activity of bothhydrogenase and urease in Helicobacter pylori. Mol. Microbiol. 39,176^182.
[201] Hausinger, R.P. (1997) Metallocenter assembly in nickel-containingenzymes. J. Biol. Inorg. Chem. 2, 279^286.
[202] Eitinger, T. and Mandrand-Berthelot, M.-A. (2000) Nickel transportsystems in microorganisms. Arch. Microbiol. 173, 1^9.
[203] Nelson, K.E., Clayton, R.A., Gill, S.R., Gwinn, M.L., Dodson,R.J., Haft, D.H., Hickey, E.K., Peterson, J.D., Nelson, W.C.,Ketchum, K.A., McDonald, L., Utterback, T.R., Malek, J.A.,Linher, K.D., Garrett, M.M., Stewart, A.M., Cotton, M.D., Pratt,M.S., Phillips, C.A., Richardson, D., Heidelberg, J., Sutton, G.G.,Fleischmann, R.D., Eisen, J.A., White, O., Salzberg, S.L., Smith,H.O., Venter, J.C. and Fraser, C.M. (1997) Evidence for lateral genetransfer between Archaea and Bacteria from genome sequence ofThermotoga maritima. Nature 399, 323^329.
[204] Verhagen, M.F., O'Rourke, T. and Adams, M.W. (1999) The hyper-thermophilic bacterium, Thermotoga maritima, contains an unusu-ally complex iron-hydrogenase : amino acid sequence analyses versusbiochemical characterization. Biochim. Biophys. Acta 1412, 212^229.
[205] Hatchikian, E.C., Magro, V., Forget, N., Nicolet, Y. and Fontecilla-Camps, J.C. (1999) Carboxy-terminal processing of the large subunitof [Fe] hydrogenase from Desulfovibrio desulfuricans ATCC 7757.J. Bacteriol. 181, 2947^2952.
[206] Dean, D.R., Bolin, J.T. and Zheng, L. (1993) Nitrogenase metal-loclusters: structures, organization, and synthesis. J. Bacteriol. 175,6737^6744.
[207] Lill, R. and Kispal, G. (2000) Maturation of cellular Fe^S proteins:an essential function of mitochondria. Trends Biochem. Sci. 25,352^356.
[208] Tong, W.-H. and Rouault, T. (2000) Distinct iron^sulfur assemblycomplexes exist in the cytosol and mitochondria of human cells.EMBO J. 19, 5692^5700.
[209] Deckert, G., Warren, P.V., Gaasterland, T., Young, W.G., Lenox,A.L., Graham, D.E., Overbeek, R., Snead, M.A., Keller, M., Aujay,M., Huber, R., Feldman, R.A., Short, J.M., Olsen, G.J. and Swan-son, R.V. (1998) The complete genome of the hyperthermophilicbacterium Aquifex aeolicus. Nature 392, 353^358.
[210] McMillan, D.J., Mau, M. and Walker, M.J. (1998) Characterisationof the urease gene cluster in Bordetella bronchiseptica. Gene 208,243^251.
[211] U¡en, R.L., Colbeau, A., Richaud, P. and Vignais, P.M. (1990)Cloning and sequencing the genes encoding uptake-hydrogenasesubunits of Rhodocyclus gelatinosus. Mol. Gen. Genet. 221, 49^58.
[212] Weiner, J.H., Bilous, P.T., Shaw, G.M., Lubitz, S.P., Frost, L.,Thomas, G.H., Cole, J.A. and Turner, R.J. (1998) A novel andubiquitous system for membrane targeting and secretion of cofac-tor-containing proteins. Cell 93, 93^101.
[213] Sargent, F., Bogsch, E.G., Stanley, N.R., Wexler, M., Robinson, C.,Berks, B.C. and Palmer, T. (1998) Overlapping functions of compo-nents of a bacterial Sec-independent protein export pathways.EMBO J. 17, 3640^3650.
[214] Wu, L.-F., Chanal, A. and Rodrigue, A. (2000) Membrane targetingand translocation of bacterial hydrogenases. Arch. Microbiol. 173,319^324.
[215] Berks, B.C., Sargent, F. and Palmer, T. (2000) The Tat proteinexport pathway. Mol. Microbiol. 35, 260^274.
[216] Voordouw, G. (2000) A universal system for the transport of redoxproteins: early roots and latest developments. Biophys. Chem. 86,131^140.
[217] Bogsch, E., Sargent, F., Stanley, N.R., Berks, B.C., Robinson, C.and Palmer, T. (1998) An essential component of a novel bacterialprotein export system with homologues in plastids and mitochon-dria. J. Biol. Chem. 273, 18003^18006.
[218] Chanal, A., Santini, C.-L. and Wu, L.-F. (1998) Potential receptorfunction of three homologous components, TatA, TatB and TatE,of the twin-arginine signal sequence-dependent metalloenzymetranslocation pathway in Escherichia coli. Mol. Microbiol. 30,674^676.
[219] Rodrigue, A., Chanal, A., Beck, K., Mu«ller, M. and Wu, L.-F.(1999) Co-translocation of a periplasmic enzyme complex by ahitchhiker mechanism through the bacterial Tat pathway. J. Biol.Chem. 274, 13223^13228.
[220] Gross, R., Simon, J. and Kro«ger, A. (1999) The role of the twin-arginine motif in the signal peptide encoded by the hydA gene of thehydrogenase from Wolinella succinogenes. Arch. Microbiol. 172,227^232.
[221] Bernhard, M., Friedrich, B. and Siddiqui, R.A. (2000) Ralstoniaeutropha TF93 is blocked in Tat-mediated protein export. J. Bacter-iol. 182, 581^588.
[222] Mura, G.M., Pedroni, P., Pratesi, C., Galli, G., Serbolisca, L. andGrandi, G. (1996) The [Ni^Fe] hydrogenase from the thermophilicbacterium Acetomicrobium £avidum. Microbiology 142, 829^836.
[223] Yagi, K., Seto, T., Terakado, M., Umeda, F., Doi, T., Imanishi, T.and Miura, Y. (1992) Nucleotide sequences of membrane-boundhydrogenase gene in Alcaligenes hydrogenophilus. Chem. Pharm.Bull. 40, 3292^3296.
[224] Carrasco, C.D., Buettner, J.A. and Golden, J.W. (1995) Pro-grammed DNA rearrangement of a cyanobacterial hupL gene inheterocysts. Proc. Natl. Acad. Sci. USA 92, 791^795.
[225] Happe, T., Schu«tz, K. and Bo«hme, H. (2000) Transcriptional andmutational analysis of the uptake hydrogenase of the ¢lamentouscyanobacterium Anabaena variabilis ATCC 29413. J. Bacteriol. 182,1624^1631.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501496
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

[226] Schmitz, O., Boison, G., Hilscher, R., Hundeshagen, B., Zimmer,W., Lottspeich, F. and Bothe, H. (1995) Molecular biological anal-ysis of a bidirectional hydrogenase from cyanobacteria. Eur. J. Bio-chem. 233, 266^276.
[227] Klenk, H.-P., Clayton, R.A., Tomb, J.-F., White, O., Nelson, K.E.,Ketchum, K.A., Dodson, R.J., Gwinn, M., Hickey, E.K., Peterson,J.D., Richardson, D.L., Kerlavage, A.R., Graham, D.E., Kyrpides,N.C., Fleischmann, R.D., Quackenbush, J., Lee, N.H., Sutton,G.G., Gill, S., Kirkness, E.F., Dougherty, B.A., McKenney, K.,Adams, M.D., Loftus, B., Peterson, S., Reich, C.I., McNeil, L.K.,Badger, J.H., Glodek, A., Zhou, L., Overbeek, R., Gocayne, J.D.,Weidman, J.F., McDonald, L., Utterback, T., Cotton, M.D.,Spriggs, T., Artiach, P., Kaine, B.P., Sykes, S.M., Sadow, P.W.,D'Andrea, K.P., Bowman, C., Fujii, C., Garland, S.A., Mason,T.M., Olsen, G.J., Fraser, C.M., Smith, H.O., Woese, C.R. andVenter, J.C. (1997) The complete genome sequence of the hyper-thermophilic, sulphate-reducing archaeon Archaeoglobus fulgidus.Nature 390, 364^370.
[228] Parkhill, J., Wren, B.W., Mungall, K., Ketley, J.M., Churcher, C.,Basham, D., Chillingworth, T., Davies, R.M., Feltwell, T., Holroyd,S., Jagels, K., Karleyshev, A., Moule, S., Pallen, M.J., Penn, C.W.,Quail, M., Rajandream, M.A., Rutherford, K.M., VanVliet, A.,Whitehead, S. and Barrell, B.G. (2000) The genome sequence ofthe food-borne pathogen Campylobacter jejuni reveals hypervariablesequences. Nature 403, 665^668.
[229] Smidt, H., Song, D., van der Oost, J. and de Vos, W.M. (1999)Random transposition by Tn916 in Desul¢tobacterium dehalogenansallows for isolation and characterization of halorespiration mutants.J. Bacteriol. 181, 6882^6888.
[230] Voordouw, G., Menon, N.K., LeGall, J., Choi, E.S., Peck Jr., H.D.and Przybyla, A.E. (1989) Analysis and comparison of nucleotidesequences encoding the genes for [NiFe] and [NiFeSe] hydrogenasesfrom Desulfovibrio gigas and Desulfovibrio baculatus. J. Bacteriol.171, 2894^2899.
[231] Menon, N.K., Peck Jr., H.D., Le Gall, J. and Przybyla, A.E. (1987)Cloning and sequencing of the genes encoding the large and smallsubunits of the periplasmic (NiFeSe) hydrogenase of Desulfovibriobaculatus (Erratum (1988) J. Bacteriol. 170, 4429). J. Bacteriol. 169,5401^5407.
[232] Franco, R., Calvete, J.J., Thole, H.H., Raida, M., Moura, I. andMoura, J.J.G. (1997) The primary structure of the beta subunit ofDesulfovibrio desulfuricans (ATCC 27774) [NiFe] hydrogenase. Pro-tein Pept. Lett. 4, 131^138.
[233] Rousset, M., Dermoun, Z., Hatchikian, C.E. and Belaich, J.P.(1990) Cloning and sequencing of the locus encoding the largeand small subunit genes of the periplasmic [NiFe]hydrogenasefrom Desulfovibrio fructosovorans. Gene 94, 95^101.
[234] Li, C., Peck Jr., H.D., LeGall, J. and Przybyla, A.E. (1987) Cloning,characterization, and sequencing of the genes encoding the large andsmall subunits of the periplasmic [NiFe]hydrogenase of Desulfovibriogigas. DNA 6, 539^551.
[235] Deckers, H.M., Wilson, F.R. and Voordouw, G. (1990) Cloning andsequencing of a [NiFe] hydrogenase operon from Desulfovibrio vul-garis Miyazaki F. J. Gen. Microbiol. 136, 2021^2028.
[236] Blattner, F.R., Plunkett III, G., Bloch, C.A., Perna, N.T., Burland,V., Riley, M., Collado-Vides, J., Glasner, J.D., Rode, C.K., May-hew, G.F., Gregor, J., Davis, N.W., Kirkpatrick, H.A., Goeden,M.A., Rose, D.J., Mau, B. and Shao, Y. (1997) The complete ge-nome sequence of Escherichia coli K-12. Science 277, 1453^1474.
[237] Andrews, S.C., Berks, B.C., Mcclay, J., Ambler, A., Quail, M.A.,Golby, P. and Guest, J.R. (1997) A 12-cistron Escherichia coli oper-on (hyf) encoding a putative proton-translocating formate hydro-genlyase system. Microbiology 143, 3633^3647.
[238] Tomb, J.-F., White, O., Kerlavage, A.R., Clayton, R.A., Sutton,G.G., Fleischmann, R.D., Ketchum, K.A., Klenk, H.P., Gill, S.,Dougherty, B.A., Nelson, K., Quackenbush, J., Zhou, L., Kirkness,E.F., Peterson, S., Loftus, B., Richardson, D., Dodson, R., Khalak,
H.G., Glodek, A., McKenney, K., Fitzegerald, L.M., Lee, N.,Adams, M.D., Hickey, E.K., Berg, D.E., Gocayne, J.D., Utterback,T.R., Peterson, J.D., Kelley, J.M., Karp, P.D., Smith, H.O., Fraser,C.M. and Venter, J.C. (1997) The complete genome sequence of thegastric pathogen Helicobacter pylori. Nature 388, 539^547.
[239] Alm, R.A., Ling, L.S., Moir, D.T., King, B.L., Brown, E.D., Doig,P.C., Smith, D.R., Noonan, B., Guild, B.C., de Jonge, B.L., Carmel,G., Tummino, P.J., Caruso, A., Uria-Nickelsen, M., Mills, D.M.,Ives, C., Gibson, R., Merberg, D., Mills, S.D., Jiang, Q., Taylor,D.E., Vovis, G.F. and Trust, T.J. (1999) Genomic-sequence compar-ison of two unrelated isolates of the human gastric pathogen Heli-cobacter pylori (Erratum (1999) 397, 719). Nature 397, 176^180.
[240] Alex, L.A., Reeve, J.N., Orme-Johnson, W.H. and Walsh, C.T.(1990) Cloning, sequence determination, and expression of the genesencoding the subunits of the nickel-containing 8-hydroxy-5-deaza-£avin reducing hydrogenase from Methanobacterium thermoautotro-phicum delta H. Biochemistry 29, 7237^7244.
[241] Smith, D.R., Doucette-Stamm, L.A., Deloughery, C., Lee, H., Du-bois, J., Aldredge, T., Bashirzadeh, R., Blakely, D., Cook, R., Gil-bert, K., Harrison, D., Hoang, L., Keagle, P., Lumm, W., Pothier,B., Qiu, D., Spadafora, R., Vicaire, R., Wang, Y., Wierzbowski, J.,Gibson, R., Jiwani, N., Caruso, A., Bush, D., Safer, H., Patwell, D.,Prabhakar, S., McDougall, S., Shimer, G., Goyal, A., Pietrokovski,S., Church, G.M., Daniels, C.J., Mao, J.-I., Rice, P., No«lling, J. andReeve, J.N. (1997) Complete genome sequence of Methanobacteriumthermoautotrophicum delta H: functional analysis and comparativegenomics. J. Bacteriol. 179, 7135^7155.
[242] Reeve, J.N., Beckler, G.S., Cram, D.S., Hamilton, P.T., Brown,J.W., Krzycki, J.A., Kolodziej, A.F., Alex, L., Orme-Johnson,W.H. and Walsh, C.T. (1989) A hydrogenase-linked gene in Meth-anobacterium thermoautotrophicum strain delta H encodes a polyfer-redoxin. Proc. Natl. Acad. Sci. USA 86, 3031^3035.
[243] Tersteegen, A. and Hedderich, R. (1999) Methanobacterium ther-moautotrophicum encodes two multisubunit membrane-bound [Ni-Fe]hydrogenases. Transcription of the operons and sequence analy-sis of the deduced proteins. Eur. J. Biochem. 264, 930^943.
[244] Bult, C.J., White, O., Olsen, G.J., Zhou, L., Fleischmann, R.D.,Sutton, G.G., Blake, J.A., FitzGerald, L.M., Clayton, R.A., Go-cayne, J.D., Kerlavage, A.R., Dougherty, B.A., Tomb, J.F., Adams,M.D., Reich, C.I., Overbeek, R., Kirkness, E.F., Weinstock, K.G.,Merrick, J.M., Glodek, A., Scott, J.L., Geoghagen, N.S.M., Weid-man, J.F., Fuhrmann, J.L., Nguyen, D., Utterback, T.R., Kelley,J.M., Peterson, J.D., Sadow, P.W., Hanna, M.C., Cotton, M.D.,Roberts, K.M., Hurst, M.A., Kaine, B.P., Borodovsky, M., Klenk,H.P., Fraser, C.M., Smith, H.O., Woese, C.R. and Venter, J.C.(1996) Complete genome sequence of the methanogenic archaeon,Methanococcus jannaschii. Science 273, 1058^1073.
[245] Halboth, S. and Klein, A. (1992) Methanococcus voltae harbors fourgene clusters potentially encoding two [NiFe] and two [NiFeSe]hydrogenases, each of the cofactor F420-reducing or F420-non-reduc-ing types. Mol. Gen. Genet. 233, 217^224.
[246] Sorgenfrei, O., Muller, S., Pfei¡er, M., Sniezko, I. and Klein, A.(1997) The [NiFe] hydrogenases of Methanococcus voltae. Genes,enzymes and regulation. Arch. Microbiol. 167, 189^195.
[247] Brodersen, J., Gottschalk, G. and Deppenmeier, U. (1999) Mem-brane-bound F420H2-dependent heterodisul¢de reduction in Metha-nococcus voltae. Arch. Microbiol. 171, 115^121.
[248] Fiebig, K. and Friedrich, B. (1989) Puri¢cation of the F420-reducinghydrogenase from Methanosarcina barkeri. Eur. J. Biochem. 184,79^88.
[249] Vaupel, M. and Thauer, R.K. (1998) Two F420-reducing hydroge-nases in Methanosarcina barkeri. Arch. Microbiol. 169, 201^205.
[250] Michel, R., Massanz, C., Kostka, S., Richter, M. and Fiebig, K.(1995) Biochemical characterization of the 8-hydroxy-5-deaza£avin-reactive hydrogenase from Methanosarcina barkeri Fusaro. Eur. J.Biochem. 233, 727^735.
[251] Deppenmeier, U., Blaut, M., Lentes, S., Herzberg, C. and Gott-
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 497
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

schalk, G. (1995) Analysis of the vhoGAC and vhtGAC operonsfrom Methanosarcina mazei strain Goe1, both encoding a mem-brane-bound hydrogenase and a cytochrome b. Eur. J. Biochem.227, 261^269.
[252] Steigerwald, V.J., Beckler, G.S. and Reeve, J.N. (1990) Conserva-tion of hydrogenase and polyferredoxin structures in the hyperther-mophilic archaebacterium Methanothermus fervidus. J. Bacteriol.172, 4715^4718.
[253] Axelsson, R., Oxelfelt, F. and Lindblad, P. (1999) Transcriptionalregulation of Nostoc uptake hydrogenase. FEMS Microbiol. Lett.170, 77^81.
[254] Oxelfelt, F., Tamagnini, P. and Lindblad, P. (1998) Hydrogen up-take in Nostoc sp. strain PCC 73102. Cloning and characterizationof a hupSL homolog. Arch. Microbiol. 169, 267^274.
[255] Santiago, B. and Meyer, O. (1997) Puri¢cation and molecular char-acterization of the H2 uptake membrane-bound NiFe-hydrogenasefrom the carboxidotrophic bacterium Oligotropha carboxidovorans.J. Bacteriol. 179, 6053^6060.
[256] Sapra, R., Verhagen, M.F. and Adams, M.W.W. (2000) Puri¢cationand characterization of a membrane-bound hydrogenase from thehyperthermophilic archaeon Pyrococcus furiosus. J. Bacteriol. 182,3423^3428.
[257] Silva, P.J., van den Ban, E.C.D., Wassink, H., Haaker, H., de Cas-tro, B., Robb, F.T. and Hagen, W.R. (2000) Enzymes of hydrogenmetabolism in Pyrococcus furiosus. Eur. J. Biochem. 267, 6541^6551.
[258] Pedroni, P., Della Volpe, A., Galli, G., Mura, G.M., Pratesi, C. andGrandi, G. (1995) Characterization of the locus encoding the [Ni^Fe] sulfhydrogenase from the archaeon Pyrococcus furiosus : evi-dence for a relationship to bacterial sul¢te reductases. Microbiology141, 449^458.
[259] Ma, K., Weiss, R. and Adams, M.W.W. (2000) Characterization ofhydrogenase II from the hyperthermophilic archaeon Pyrococcusfuriosus and assessment of its role in sulfur reduction. J. Bacteriol.182, 1864^1871.
[260] Kawarabayasi, Y., Sawada, M., Horikawa, H., Haikawa, Y., Hino,Y., Yamamoto, S., Sekine, M., Baba, S., Kosugi, H., Hosoyama,A., Nagai, Y., Sakai, M., Ogura, K., Otsuka, R., Nakazawa, H.,Takamiya, M., Ohfuku, Y., Funahashi, T., Tanaka, T., Kudoh, Y.,Yamazaki, J., Kushida, N., Oguchi, A., Aoki, K., Yoshizawa, T.,Nakamura, Y., Robb, F.T., Horikoshi, K., Masuchi, Y., Shizuya,H. and Kikuchi, H. (1998) Complete sequence and gene organiza-tion of the genome of a hyper-thermophilic archaebacterium, Pyro-coccus horikoshii. DNA Res. 5, 55^76.
[261] Brito, B., Palacios, J.M., Imperial, J., Ruiz-Argu«eso, T., Yang,W.C., Bisseling, T., Schmitt, H., Kerl, V., Bauer, T., Kokotek, W.and Lotz, W. (1997) Temporal and spatial co-expression of hydro-genase and nitrogenase genes from Rhizobium leguminosarum bv.viciae in pea (Pisum sativum L.) root nodules. Mol. Plant MicrobeInteract. 8, 235^240.
[262] Grzeszik, C., Lu«bbers, M., Reh, M. and Schlegel, H.G. (1997)Genes encoding the NAD-reducing hydrogenase of Rhodococcusopacus MR11. Microbiology 143, 1271^1286.
[263] Grzeszik, C., Ross, K., Schneider, K., Reh, M. and Schlegel, H.G.(1997) Location, catalytic activity, and subunit composition ofNAD-reducing hydrogenases of some Alcaligenes strains and Rho-dococcus opacus MR22. Arch. Microbiol. 167, 172^176.
[264] Fox, J.D., He, Y., Shelver, D., Roberts, G.P. and Ludden, P.W.(1996) Characterization of the region encoding the CO-induced hy-drogenase of Rhodospirillum rubrum. J. Bacteriol. 178, 6200^6208.
[265] Fox, J.D., Kerby, R.L., Roberts, G.P. and Ludden, P.W. (1996)Characterization of the CO-induced, CO-tolerant hydrogenasefrom Rhodospirillum rubrum and the gene encoding the large subunitof the enzyme. J. Bacteriol. 178, 1515^1524.
[266] Boison, G., Schmitz, O., Mikheeva, L., Shestakov, S. and Bothe, H.(1996) Cloning, molecular analysis and insertional mutagenesis ofthe bidirectional hydrogenase genes from the cyanobacterium Ana-cystis nidulans. FEBS Lett. 394, 153^158.
[267] Boison, G., Schmitz, O., Schmitz, B. and Bothe, H. (1998) Unusualgene arrangement of the bidirectional hydrogenase and functionalanalysis of its diaphorase subunit HoxU in respiration of the uni-cellular cyanobacterium Anacystis nidulans. Curr. Microbiol. 36,253^258.
[268] Appel, J. and Schulz, R. (1996) Sequence analysis of an operon of aNAD(P)-reducing nickel hydrogenase from the cyanobacteriumSynechocystis sp. PCC 6803 gives additional evidence for direct cou-pling of the enzyme to NAD(P)H-dehydrogenase (complex I). Bio-chim. Biophys. Acta 1298, 141^147.
[269] Kaneko, T., Sato, S., Kotani, H., Tanaka, A., Asamizu, E., Naka-mura, Y., Miyajima, N., Hirosawa, M., Sugiura, M., Sasamoto, S.,Kimura, T., Hosouchi, T., Matsuno, A., Muraki, A., Nakazaki, N.,Naruo, K., Okumura, S., Shimpo, S., Takeuchi, C., Wada, T., Wa-tanabe, A., Yamada, M., Yasuda, M. and Tabata, S. (1996) Se-quence analysis of the genome of the unicellular cyanobacteriumSynechocystis sp. strain PCC6803. II. Sequence determination ofthe entire genome and assignment of potential protein-coding re-gions. DNA Res. 3, 109^136.
[270] Rakhely, G., Zhou, Z.H., Adams, M.W.W. and Kovacs, K.L.(1999) Biochemical and molecular characterization of the [NiFe]hydrogenase from the hyperthermophilic archaeon, Thermococcuslitoralis. Eur. J. Biochem. 266, 1158^1165.
[271] Dross, F., Geisler, V., Lenger, R., Theis, F., Kra¡t, T., Fahrenholz,F., Kojro, E., Ducheªne, A., Tripier, D., Juvenal, K. and Kro«ger, A.(1992) The quinone-reactive Ni/Fe-hydrogenase of Wolinella succi-nogenes (Erratum (1993) 214, 949^950). Eur. J. Biochem. 206, 93^102.
[272] Meyer, J. and Gagnon, J. (1991) Primary structure of hydrogenase Ifrom Clostridium pasteurianum. Biochemistry 30, 9697^9704.
[273] Atta, M. and Meyer, J. (2000) Characterization of the gene encodingthe [Fe]-hydrogenase from Megasphaera elsdenii. Biochim. Biophys.Acta 1476, 368^371.
[274] Malki, S., Saimmaime, I., De Luca, G., Rousset, M., Dermoun, Z.and Belaich, J.-P. (1995) Characterization of an operon encoding anNADP-reducing hydrogenase in Desulfovibrio fructosovorans.J. Bacteriol. 177, 2628^2636.
[275] Horner, D.S., Foster, P.G. and Embley, T.M. (2000) Iron hydro-genases and the evolution of anaerobic eukaryotes. Mol. Biol. Evol.17, 1695^1709.
[276] Bui, E.T.N. and Johnson, P.J. (1996) Identi¢cation and character-ization of [Fe]-hydrogenases in the hydrogenosome of Trichomonasvaginalis. Mol. Biochem. Parasitol. 76, 305^310.
[277] Plu«mper, E.P., Bradley, P.J. and Johnson, P.J. (1998) Implicationsof protein import on the origin of hydrogenosomes. Protist 149,303^311.
[278] Voordouw, G. and Brenner, S. (1985) Nucleotide sequence of thegene encoding the hydrogenase from Desulfovibrio vulgaris (Hilden-borough). Eur. J. Biochem. 148, 515^520.
[279] Fontecilla-Camps, J.C., Frey, M., Garcin, E., Hatchikian, C., Mon-tet, Y., Piras, C., Vernede, X. and Volbeda, A. (1997) Hydrogenase:a hydrogen-metabolizing enzyme. What do the crystal structures tellus about its mode of action? Biochimie 79, 661^666.
[280] Montet, Y., Amara, P., Volbeda, A., Vernede, X., Hatchikian,E.C., Field, M.J., Frey, M. and Fontecilla-Camps, J.C. (1997) Gasaccess to the active site of Ni^Fe hydrogenases probed by X-raycrystallography and molecular dynamics. Nat. Struct. Biol. 4, 523^526.
[281] Ma, K., Schicho, R.N., Kelly, R.M. and Adams, M.W.W. (1993)Hydrogenase of the hyperthermophile Pyrococcus furiosus is an el-emental sulfur reductase or sulfhydrogenase: Evidence for a sulfur-reducing hydrogenase ancestor. Proc. Natl. Acad. Sci. USA 90,5341^5344.
[282] Bairoch, A. and Bucher, P. (1994) PROSITE: recent developments.Nucleic Acids Res. 22, 3583^3589.
[283] Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTALW: improving the sensitivity of progressive multiple sequence align-
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501498
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

ment through sequence weighting, position-speci¢c gap penaltiesand weight matrix choice. Nucleic Acids Res. 22, 4673^4680.
[284] Porte, F. and Vignais, P.M. (1980) Electron transport chain andenergy transduction in Paracoccus denitri¢cans under autotrophicgrowth conditions. Arch. Microbiol. 127, 1^10.
[285] Paul, F., Colbeau, A. and Vignais, P.M. (1979) Phosphorylationcoupled to H2 oxidation by chromatophores from Rhodopseudomo-nas capsulata. FEBS Lett. 106, 29^33.
[286] Komen, R., Schmidt, K. and Zannoni, D. (1996) Hydrogen oxida-tion by membranes from autotrophically grown Alcaligenes eutro-phus H16: role of the cyanide-resistant pathway in energy transduc-tion. Arch. Microbiol. 165, 418^420.
[287] Gross, R., Simon, J., Theis, F. and Kro«ger, A. (1998) Two mem-brane anchors of Wolinella succinogenes hydrogenase and their func-tion in fumarate and polysul¢de respiration. Arch. Microbiol. 170,50^58.
[288] Ide, T., Baumer, S. and Deppenmeier, U. (1999) Energy conserva-tion by the H2 :heterodisul¢de oxidoreductase from Methanosarcinamazei Go«1: identi¢cation of two proton-translocating segments.J. Bacteriol. 181, 4076^4080.
[289] Elsen, S., Dischert, W., Colbeau, A. and Bauer, C.E. (2000) Expres-sion of uptake hydrogenase and molybdenum nitrogenase in Rho-dobacter capsulatus is coregulated by the RegB-RegA two-compo-nent regulatory system. J. Bacteriol. 182, 2831^2837.
[290] Oxelfelt, F., Tamagnini, P., Salema, R. and Lindblad, P. (1995)Hydrogen uptake in Nostoc sp. strain PCC73102 e¡ects of nickel,hydrogen, carbon and nitrogen. Plant Physiol. Biochem. 33, 617^623.
[291] Troshina, O.Y., Serebryakova, L.T. and Lindblad, P. (1996) Induc-tion of H2-uptake and nitrogenase activities in the cyanobacteriumAnabaena variabilis ATCC 29413: e¡ects of hydrogen and organicsubstrate. Curr. Microbiol. 33, 11^15.
[292] Bianco, P., Haladjian, J., Bruschi, M. and Guerlesquin, F. (1992)Reactivity of [Fe] and [Ni^Fe^Se] hydrogenases with their oxido-reduction partner, the tetraheme cytochrome c3. Biochem. Biophys.Res. Commun. 189, 633^639.
[293] Lovley, D.R. (1993) Metal-reducing microorganisms. Annu. Rev.Microbiol. 47, 263^290.
[294] Rossi, M., Pollock, W.B., Reij, M.W., Keon, R.G., Fu, R. andVoordouw, G. (1993) The hmc operon of Desulfovibrio vulgarissubsp. vulgaris Hildenborough encodes a potential transmembraneredox protein complex. J. Bacteriol. 175, 4699^4711.
[295] Morelli, X., Czjzek, M., Hatchikian, C.E., Bornet, O., Fontecilla-Camps, J.C., Palma, N.P., Moura, J.J.G. and Guerlesquin, F. (2000)Structural model of the Fe-hydrogenase/cytochrome c553 complexcombining transverse relaxation-optimized spectroscopy experi-ments and soft docking calculations. J. Biol. Chem. 275, 23204^23210.
[296] Aubert, C., Brugna, M., Dolla, A., Bruschi, M. and Giudici-Orti-coni, M.T. (2000) A sequential electron transfer from hydrogenasesto cytochromes in sulfate-reducing bacteria. Biochim. Biophys. Acta1476, 85^92.
[297] Pereira, I.A.C., Romao, C.V., Xavier, A.V., LeGall, J. and Teixiera,M. (1998) Electron transfer between hydrogenases and mono- andmultiheme cytochromes in Desulfovibrio ssp. J. Biol. Inorg. Chem. 3,494^498.
[298] Matias, P.M., Coelho, R., Pereira, I.A., Coelho, A.V., Thompson,A.W., Sieker, L.C., LeGall, J. and Carrondo, M.A. (1999) Theprimary and three-dimensional structures of a nine-haem cyto-chrome c from Desulfovibrio desulfuricans ATCC 27774 reveal anew member of the Hmc family. Structure 7, 119^130.
[299] Rapp-Giles, B.J., Casalot, L., English, R.S., Ringbauer Jr., J.A.,Dolla, A. and Wall, J.D. (2000) Cytochrome c3 mutants of Desul-fovibrio desulfuricans. Appl. Environ. Microbiol. 66, 671^677.
[300] Dolla, A., Pohorelic, B.K., Voordouw, J.K. and Voordouw, G.(2000) Deletion of the hmc operon of Desulfovibrio vulgarissubsp. vulgaris Hildenborough hampers hydrogen metabolism and
low-redox-potential niche establishment. Arch. Microbiol. 174,143^151.
[301] Pierik, A.J., Schmelz, M., Lenz, O., Friedrich, B. and Albracht,S.P.J. (1998) Characterization of the active site of a hydrogen sensorfrom Alcaligenes eutrophus. FEBS Lett. 438, 231^235.
[302] Deppenmeier, U., Lienard, T. and Gottschalk, G. (1999) Novel re-actions involved in energy conservation by methanogenic archaea.FEBS Lett. 457, 291^297.
[303] Ba«umer, S., Ide, T., Jacobi, C., Johann, A., Gottschalk, G. andDeppenmeier, U. (2000) The F420H2 dehydrogenase from Methano-sarcina mazei is a redox-driven proton pump closely related toNADH dehydrogenase. J. Biol. Chem. 275, 17968^17973.
[304] Ma, K., Zhou, Z.H. and Adams, M.W.W. (1994) Hydrogen pro-duction from pyruvate by enzymes puri¢ed from the hyperthermo-philic archaeon, Pyrococcus furiosus : A key role for NADPH.FEMS Microbiol. Lett. 122, 245^250.
[305] Schneider, K. and Schlegel, H.G. (1976) Puri¢cation and propertiesof the soluble hydrogenase from Alcaligenes eutrophus H16. Bio-chim. Biophys. Acta 452, 66^80.
[306] Kentemich, T., Bahnweg, M., Mayer, F. and Bothe, H. (1989) Lo-calization of the reversible hydrogenase in cyanobacteria. Z. Natur-forsch. 44c, 384^391.
[307] Serebryakova, L.T., Zorin, N.A. and Lindblad, P. (1994) Reversiblehydrogenase in Anabaena variabilis ATCC 29413. Presence and lo-calization in non N2-¢xing cells. Arch. Microbiol. 161, 140^144.
[308] Schmitz, O. and Bothe, H. (1996) NAD(P)�-dependent hydrogenaseactivity in extracts from the cyanobacterium Anacystis nidulans.FEMS Microbiol. Lett. 135, 97^101.
[309] Serebryakova, L.T., Sheremetieva, M.E. and Lindblad, P. (2000)H2-uptake and evolution in the unicellular cyanobacterium Chroo-coccidiopsis thermalis CALU 758. Plant Physiol. Biochem. 38, 525^530.
[310] Schmitz, O. and Bothe, H. (1996) The diaphorase subunit HoxU ofthe bidirectional hydrogenase as electron transferring protein in cy-anobacterial respiration? Naturwissenschaften 83, 525^527.
[311] Tamagnini, P., Troshina, O., Oxelfelt, F., Salema, R. and Lindblad,P. (1997) Hydrogenases in Nostoc sp. strain PCC 73102, a strainlacking a bidirectional enzyme. Appl. Environ. Microbiol. 63, 1801^1807.
[312] Tamagnini, P., Costa, J.L., Almeida, L., Oliveira, M.J., Salema, R.and Lindblad, P. (2000) Diversity of cyanobacterial hydrogenases, amolecular approach. Curr. Microbiol. 40, 356^361.
[313] Boison, G., Bothe, H., Hansel, A. and Lindblad, P. (1999) Evidenceagainst a common use of the diaphorase subunits by the bidirec-tional hydrogenase and by the respiratory complex I in cyanobac-teria. FEMS Microbiol. Lett. 174, 159^165.
[314] Appel, J., Phunpruch, S., Steinmu«ller, K. and Schulz, R. (2000) Thebidirectional hydrogenase of Synechocystis sp. PCC 6803 works asan electron valve during photosynthesis. Arch. Microbiol. 173, 333^338.
[315] Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M. and Sta-nier, R.Y. (1979) Generic assignments, strain histories and proper-ties of pure cultures of cyanobacteria. J. Gen. Microbiol. 111, 1^61.
[316] Santangelo, J.D., Du«rre, P. and Woods, D.R. (1995) Characteriza-tion and expression of the hydrogenase-encoding gene from Clostri-dium acetobutylicum P262. Microbiology 141, 171^180.
[317] Gorwa, M.-F., Croux, C. and Soucaille, P. (1996) Molecular char-acterization and transcriptional analysis of the putative hydrogenasegene of Clostridium acetobutylicum ATCC 824. J. Bacteriol. 178,2668^2675.
[318] Kaji, M., Taniguchi, Y., Matsushita, O., Katayama, S., Miyata, S.,Morita, S. and Okabe, A. (1999) The hydA gene encoding the H2-evolving hydrogenase of Clostridium perfringens : molecular charac-terization and expression of the gene. FEMS Microbiol. Lett. 181,329^336.
[319] Casalot, L., Hatchikian, C.E., Forget, N., De Philip, P., Dermoun,Z., Belaich, J.-P. and Rousset, M. (1998) Molecular study and par-
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 499
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

tial characterization of iron-only hydrogenase in Desulfovibrio fruc-tosovorans. Anaerobe 4, 45^55.
[320] Stokkermans, J., van Dongen, W., Kaan, A., van den Berg, W. andVeeger, C. (1989) hyd Q, a gene from Desulfovibrio vulgaris (Hilden-borough) encodes a polypeptide homologous to the periplasmic hy-drogenase. FEMS Microbiol. Lett. 58, 217^222.
[321] Voordouw, G., Strang, J.D. and Wilson, F.R. (1989) Organizationof the genes encoding [Fe] hydrogenase in Desulfovibrio vulgarissubsp. oxamicus Monticello. J. Bacteriol. 171, 3881^3889.
[322] Akhmanova, A., Voncken, F., van Alen, T., van Hoek, A., Boxma,B., Vogels, G., Veenhuis, M. and Hackstein, J.H.P. (1998) A hydro-genosome with a genome. Nature 396, 527^528.
[323] Philippsen, P., Kleine, K., Pohlmann, R., Dusterhoft, A., Hamberg,K., Hegemann, J.H., Obermaier, B., Urrestarazu, L.A., Aert, R.,Albermann, K., Altmann, R., Andre, B., Baladron, V., Ballesta,J.P., Becam, A.M., Beinhauer, J., Boskovic, J., Buitrago, M.J., Bus-sereau, F., Coster, F., Crouzet, M., D'Angelo, M., Dal Pero, F., DeAntoni, A. and Hani, J. et al. (1997) The nucleotide sequence ofSaccharomyces cerevisiae chromosome XIV and its evolutionary im-plications. Nature 387, 93^98.
[324] Adams, M.D. et al. (2000) The genome sequence of Drosophilamelanogaster. Science 287, 2185^2195.
[325] Barton, R.M. and Worman, H.J. (1999) Prenylated prelamin Ainteracts with Narf, a novel nuclear protein. J. Biol. Chem. 274,30008^30018.
[326] Atta, M., La¡erty, M.E., Johnson, M.K., Gaillard, J. and Meyer, J.(1998) Heterologous biosynthesis and characterization of the [2Fe^2S]-containing N-terminal domain of Clostridium pasteurianum hy-drogenase. Biochemistry 37, 15974^15980.
[327] Ku«mmerle, R., Atta, M., Scuiller, J., Gaillard, J. and Meyer, J.(1999) Structural similarities between the N-terminal domain ofClostridium pasteurianum hydrogenase and plant-type ferredoxins.Biochemistry 38, 1938^1943.
[328] Yeh, A.P., Chatelet, C., Soltis, S.M., Kuhn, P., Meyer, J. and Rees,D.C. (2000) Structure of a thioredoxin-like [2Fe^2S] ferredoxin fromAquifex aeolicus. J. Mol. Biol. 300, 587^595.
[329] Payne, M.J., Chapman, A. and Cammack, R. (1993) Evidence foran [Fe]-type hydrogenase in the parasitic protozoan Trichomonasvaginalis. FEBS Lett. 317, 101^104.
[330] Chatelet, C., Gaillard, J., Petillot, Y., Louwagie, M. and Meyer, J.(1999) A [2Fe^2S] protein from the hyperthermophilic bacteriumAquifex aeolicus. Biochem. Biophys. Res. Comm. 261, 885^889.
[331] Knight Jr., E. and Hardy, R.W.F. (1966) Isolation and character-istics of £avodoxin from nitrogen-¢xing Clostridium pasteurianum.J. Biol. Chem. 241, 2752^2756.
[332] Fitzgerald, M.P., Rogers, L.J., Rao, K.K. and Hall, D.O. (1980)E¤ciency of ferredoxins and £avodoxins as mediators in systemsfor hydrogen evolution. Biochem. J. 192, 665^672.
[333] Benemann, J.R., Berenson, J.A., Kaplan, N.O. and Kamen, M.D.(1973) Hydrogen evolution by a chloroplast-ferredoxin-hydrogenasesystem. Proc. Natl. Acad. Sci. USA 70, 2317^2320.
[334] Rao, K.K., Rosa, L. and Hall, D.O. (1976) Prolonged production ofhydrogen gas by a chloroplast biocatalytic system. Biochem. Bio-phys. Res. Commun. 68, 21^28.
[335] Moulis, J.-M. and Davasse, V. (1995) Probing the role of electro-static forces in the interaction of Clostridium pasteurianum ferredox-in with its redox partners. Biochemistry 34, 16781^16788.
[336] Ku«mmerle, R., Kyritsis, P., Gaillard, J. and Moulis, J.-M. (2000)Electron transfer properties of iron^sulfur proteins. J. Inorg. Bio-chem. 79, 83^91.
[337] Hackstein, J.H.P., Akhmanova, A., Boxma, B., Harhangi, H.R. andVoncken, G.J. (1999) Hydrogenosomes: eukaryotic adaptations toanaerobic environments. Trends Microbiol. 7, 441^447.
[338] Johnson, P.J.., d'Oliveira, C.E., Gorrell, T.E. and Mu«ller, M. (1990)Molecular analysis of the hydrogenosomal ferredoxin of the anaer-obic protist Trichomonas vaginalis. Proc. Natl. Acad. Sci. USA 87,6097^6101.
[339] Akhmanova, A., Voncken, F.G.J., Hosea, K.M., Harhangi, H.,Keltjens, J.T., op den Camp, H.J.M., Vogels, G.D. and Hackstein,J.H.P. (1999) A hydrogenosome with pyruvate formate-lyase: an-aerobic chytrid fungi use an alternative route for pyruvate catabo-lism. Mol. Microbiol. 32, 1103^1114.
[340] Chen, J.-S. and Blanchard, D.K. (1984) Puri¢cation and propertiesof the H2-oxidizing (uptake) hydrogenase of the N2-¢xing anaerobeClostridium pasteurianum W5. Biochem. Biophys. Res. Commun.122, 9^16.
[341] Altschul, S.F., Madden, T.L., Scha¡er, A.A., Zhang, J., Zhang, Z.,Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.Nucleic Acids Res. 25, 3389^3402.
[342] Tatusova, T.A. and Madden, T.L. (1999) BLAST2 sequences, a newtool for comparing protein and nucleotide sequences. FEMS Micro-biol. Lett. 174, 247^250.
[343] Saitou, N. and Nei, M. (1987) The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Mol. Biol. Evol. 4,406^425.
[344] Gascuel, O. (1997) BIONJ: an improved version of the NJ algo-rithm based on a simple model of sequence data. Mol. Biol. Evol.14, 685^695.
[345] Feselstein, J. (1993) PHYLIP (Phylogeny Inference Package) version3.5c. http://evolution.genetics.washington.edu/phylip.html.Distrib-uted by the author. Department of Genetics, University of Wash-ington, Seattle, WA.
[346] Perrie©re, G. and Gouy, M. (1996) WWW-Query: an on-line retrievalsystem for biological sequence banks. Biochimie 78, 364^369.
[347] Feselstein, J. (1995) Con¢dence limits on phylogenies: an approachusing the bootstrap. Evolution 39, 783^791.
[348] Wheeler, D.L., Chappey, C., Lash, A.E., Leippe, D.D., Madden,T.L., Schuler, G.D., Tatusova, T.A. and Rapp, B.A. (2000) Data-base resources of the National Center for Biotechnology Informa-tion. Nucleic Acids Res. 28, 10^14.
[349] Benson, D.A., Karsch-Mizrachi, I., Lipman, D.J., Ostell, J., Rapp,B.A. and Wheeler, D.L (2000) GenBank. Nucleic Acids Res. 28, 15^18.
[350] Doolittle, W.F. (1999) Phylogenetic classi¢cation and the universaltree. Science 284, 2124^2128.
[351] Woese, C.R. (2000) Interpreting the universal phylogenetic tree.Proc. Natl. Acad. Sci. USA 97, 8392^8396.
[352] Yagi, T., Yano, T., Di Bernardo, S. and Matsuno-Yagi, A. (1998)Procaryotic complex I (NDH-1), an overview. Biochim. Biophys.Acta 1364, 125^133.
[353] Friedrich, T. (1998) The NADH:ubiquinone oxidoreductase (com-plex I) from Escherichia coli. Biochim. Biophys. Acta 1364, 134^146.
[354] Dupuis, A., Chevallet, M., Darrouzet, E., Duborjal, H., Lunardi, J.and Issartel, J.P. (1998) The Complex I from Rhodobacter capsula-tus. Biochim. Biophys. Acta 1364, 147^165.
[355] Duborjal, H., Dupuis, A., Chapel, A., Kie¡er, S., Lunardi, J. andIssartel, J.P. (1997) Immuno-puri¢cation of a dimeric subcomplex ofthe respiratory NADH-CoQ reductase of Rhodobacter capsulatusequivalent to the FP fraction of the mitochondrial complex I.FEBS Lett. 405, 345^350.
[356] Sazanov, L.A., Peak-Chew, S.Y., Fearnley, I.M. and Walker, J.E.(2000) Resolution of the membrane domain of bovine complex Iinto subcomplexes : implications for the structural organization ofthe enzyme. Biochemistry 39, 7229^7235.
[357] Yano, T. and Yagi, T. (1999) H�-translocating NADH-quinoneoxidoreductase (NDH-1) of Paracoccus denitri¢cans. Studies ontopology and stoichiometry of the peripheral subunits. J. Biol.Chem. 274, 28606^28611.
[358] Prieur, I., Lunardi, J. and Dupuis, A. (2001) Evidence for a quinonebinding site close to the interface between NUOD and NUOB sub-units of Complex I. Biochim. Biophys. Acta 1504, 173^178.
[359] Yano, T., Magnitsky, S., Sled, V.D., Ohnishi, T. and Yagi, T. (1999)Characterization of the putative 2x[4Fe^4S]-binding NQO9 subunit
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501500
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022

of the proton-translocating NADH-quinone oxidoreductase (NDH-1) of Paracoccus denitri¢cans. J. Biol. Chem. 274, 28598^28605.
[360] Meuer, J., Bartoschek, S., Koch, J., Ku«nkel, A. and Hedderich, R.(1999) Puri¢cation and catalytic properties of Ech hydrogenase fromMethanosarcina barkeri. Eur. J. Biochem. 265, 325^335.
[361] Morales, R., Charon, M.-H., Hudry-Clergeon, G., Petillot, Y., Nor-ager, S., Medina, M. and Frey, M. (1999) Re¢ned X-ray structuresof the oxidized, at 1.3 Aî , and reduced, at 1.17 Aî , [2Fe^2S] ferredox-in from the cyanobacterium Anabaena PCC7119 show redox-linkedconformational changes. Biochemistry 38, 15764^15773.
[362] Dauter, Z., Wilson, K.S., Sieker, L.C., Meyer, J. and Moulis, J.-M.(1997) Atomic resolution (0.94 Aî ) structure of Clostridium aciduriciferredoxin. Detailed geometry of [4Fe^4S] clusters in a protein. Bio-chemistry 36, 16065^16073.
[363] Dupuis, A., Prieur, I. and Lunardi, J. (2001) Toward a character-ization of the connecting module of Complex I. J. Bioenerg. Bio-membr. Rev. 33, 161^170.
[364] Bru«ggemann, H., Falinski, F. and Deppenmeier, U. (2000) Structureof the F420H2 :quinone oxidoreductase of Archaeoglobus fulgidus.Identi¢cation and overproduction of the F420H2-oxidizing subunit.Eur. J. Biochem. 267, 5810^5814.
[365] Kunow, J., Linder, D., Stetter, K.O. and Thauer, R.K. (1994)F420H2 :quinone oxidoreductase from Archaeoglobus fulgidus. Char-acterization of a membrane-bound multisubunit complex containingFAD and iron^sulfur clusters. Eur. J. Biochem. 223, 503^511.
[366] Margulis, L., Dolan, M.F. and Guerrero, R. (2000) The chimericeukaryote: Origin of the nucleus from the karyomastigont in ami-tochondriate protists. Proc. Natl. Acad. Sci. USA 97, 6954^6959.
[367] Martin, W. and Mu«ller, M. (1998) The hydrogen hypothesis for the¢rst eukaryote. Nature 392, 37^41.
[368] Wu«nschiers, R., Senger, H. and Schulz, R. (2001) Electron pathwaysinvolved in H2-metabolism in the green alga Scenedesmus obliquus.Biochim. Biophys. Acta 1503, 271^278.
[369] Ewart, G.D., Reed, K.C. and Smith, G.D. (1990) Soluble hydro-genase of Anabaena cylindrica. Cloning and sequencing of a poten-tial gene encoding the tritium exchange subunit. Eur. J. Biochem.187, 215^223.
[370] de Zoysa, P.A. and Danpure, C.J. (1993) Possible evolutionary re-lationship between mammalian alanine:glyoxylate aminotransferase1 and the 42-kD subunit of cyanobacterial soluble hydrogenase.Mol. Biol. Evol. 10, 704^706.
[371] Afting, C., Kremmer, E., Brucker, C., Hochheimer, A. and Thauer,R.K. (2000) Regulation of the synthesis of H2-forming methylenete-trahydromethanopterin dehydrogenase (Hmd) and of HmdII andHmdIII in Methanothermobacter marburgensis. Arch. Microbiol.174, 225^232.
[372] Leonhartsberger, S., Huber, A., Lottspeich, F. and Bo«ck, A. (2001)The hydH/G genes from Escherichia coli code for a zinc and leadresponsive two-component regulatory system. J. Mol. Biol. 307, 93^105.
[373] Zorin, N.A., Dimon, B., Gagnon, J., Gaillard, J., Carrier, P. andVignais, P.M. (1996) Inhibition by iodoacetamide and acetylene ofthe H-D-exchange reaction catalyzed by Thiocapsa roseopersicinahydrogenase. Eur. J. Biochem. 241, 675^681.
[374] Adams, M.W., Holden, J.F., Menon, A.L., Schut, G.J., Grunden,A.M., Hou, C., Hutchins, A.M., Jenney Jr., F.E., Kim, C., Ma, K.,Pan, G., Roy, R., Sapra, R., Story, S.V. and Verhagen, M.F. (2001)Key role for sulfur in peptide metabolism and in regulation of threehydrogenases in the hyperthermophilic archaeon Pyrococcus furio-sus. J. Bacteriol. 183, 716^724.
[375] Franzen, L.G., Rochaix, J.D. and von Heijne, G. (1990) Chloroplasttransit peptides from the green alga Chlamydomonas reinhardtiishare features with both mitochondrial and higher plant chloroplastpresequences. FEBS Lett. 260, 165^168.
[376] Settles, A.M. and Martienssen, R. (1998) Old and new pathways ofprotein export in chloroplasts and bacteria. Trends Cell Biol. 8, 494^501.
[377] Huber, R. and Stetter, K.O. (2001) Discovery of hyperthermophilicmicroorganisms. Methods Enzymol. 330, 11^24.
[378] Sayle, R.A. and Milner-White, E.J. (1995) RasMol: biomoleculargraphics for all. Trends Biochem. Sci. 20, 374^376.
[379] Gupta, R.S. (2000) The phylogeny of proteobacteria: relationshipsto other eubacterial phyla and eukaryotes. FEMS Microbiol. Rev.24, 367^402.
[380] Burggraf, S., Olsen, G.J., Stetter, K.O. and Woese, C.R. (1992) Aphylogenetic analysis of Aquifex pyrophilus. Syst. Appl. Microbiol.15, 352^356.
[381] Daubin, V., Perrie©re, G. and Gouy, M. (2001) A phylogenomicapproach using super-trees to reconstruct prokaryotic history. In:Actes du Colloque Journees Ouvertes de Biologie, Informatique etMathematiques (JOBIM 2001), Toulouse.
FEMSRE 723 20-8-01 Cyaan Magenta Geel Zwart
P.M. Vignais et al. / FEMS Microbiology Reviews 25 (2001) 455^501 501
Dow
nloaded from https://academ
ic.oup.com/fem
sre/article/25/4/455/525649 by guest on 14 March 2022




















