Influence of the extraction time on macromolecular parameters of galactomannans
Upload
independentCategory
view
0download
0
Absorption of macromolecular proteins by therectal epithelium of the Antarctic fishNotothenia neglecta
Francisco Javier Hernandez-Blazquez and José Roberto Machado Cunha daSilva
Abstract: The absorption of intact proteins byNotothenia neglectaenterocytes was studied by both light and electronmicroscopy after administration of ferritin by the oral and anal routes. Intense pinocytosis of ferritin occurs in thesecond half of the preanal segment, located distal to the rectal valve, which is equivalent to 5% of the intestine length.The ferritin was identified after 48 h by light microscopy and was also observed in the interior of the electron-densecytoplasmic vesicles of the rectal epithelium. Uptake and transport of protein macromolecules by the oral route are lessmarked than in other fishes studied. As ferritin uptake was more intense by the anal route, we conclude that therectum ofN. neglectais the intestinal segment where endogenous or exogenous proteins are normally absorbed in greatquantity by pinocytosis.
Résumé: L’absorption des macromolécules intactes par l’épithélium intestinal du poissonNotothenia neglectaa étéétudiée aux microscopes électronique et photonique après l’administration de ferritine par voies orale et anale. Uneactivité intense de pinocytose de la ferritine a été observée dans la seconde moitié du segment préanal, postérieurementà la valve rectale. Cette région représente 5% de la longueur de l’intestin. La ferritine a été observée après 48 h parmicroscopie photonique et a été décelée aussi à l’intérieur de vésicules cytoplasmiques, opaques aux électrons, dans lesenterocytes rectaux. La capture et le transport des protéines macromoléculaires par voie orale se sont avéré moinsintenses que chez d’autres poissons. Comme l’absorption de ferritine a été plus intense par la voie anale que par lavoie orale, on peut conclure que le rectum deN. neglectaest le segment intestinal où les protéines d’origine endogèneou exogène sont normalement absorbées en grand quantité par pinocytose.
Hernandez-Blazquez and Silva 1253
The transport of intact macromolecules by enterocytes iswidespread in vertebrates. In ungulates such transportationtakes place by means of a process that ceases 2 days afterbirth, termed “gut closure” (Brandtzaeg and Krajci 1995). Infishes the transepithelial transport system remains functionalthroughout adulthood. In the grass carp,Ctenopharyngodonidella, around 70% of dietary protein is digested and ab-sorbed in the proximal segment of the intestine (Strobandand Van der Veen 1981). The remaining 30% is transportedto the middle and distal intestinal segments in whose epithe-lium markers of pinocytosis can be demonstrated.Peroxidase (Gauthier and Landis 1972; Iida and Yamamoto1985; Georgopoulou et al. 1985; Rombout and van den Berg1989), ferritin (Stroband and Van der Veen 1981; Romboutet al. 1985), and the bacteriumVibrio anguillarum(Romboutand van den Berg 1989) are usually employed as markers ofpinocytosis. Immunolocalization methods are also used to
demonstrate enterocyte uptake of proteins in fishes (Sire andVernier 1992; Dorin et al. 1994). The physiological signifi-cance of intestinal pinocytosis of proteins is controversial,although the process has been observed in several species offishes. Three functions have been attributed to thepinocytosis of proteins in fish enterocytes. The presence ofintraepithelial leukocytes, macrophages, and lymphocytesindicates that the second intestinal segment can play an im-portant immunological role in intestinal function (Romboutet al. 1989b; Dorin et al. 1993a, 1993b; Sire and Vernier1995). Georgopoulou and Vernier (1986) demonstrated anantibody response in trout following intestinal administra-tion of a soluble antigen. The enterocytes from such seg-ments in fishes are frequently compared, functionally, tomammalian M cells (Rombout et al. 1985, 1989a; Walkerand Sanderson 1995). It has also been suggested that thepinocytotic process acts as an auxiliary mechanism for pro-tein absorption in fishes (Georgopoulou et al. 1986). A“standby” function, whose importance increases when agreat quantity of food becomes available, is attributed toboth segments (Noaillac-Depeyre and Gas 1973a, 1976;Stroband and Van der Veen 1981; Rombout et al. 1985). Itwas also observed that proteolytic activity of the entericfluid diminishes throughout the intestine (Hofer 1982). Thisreduction in enzymatic activity was attributed to pinocytosisof enzymes in the middle and distal regions of the intestine.Antarctic fishes are one of the best examples of adjustmentto low temperature found in the fauna of the region. Adapta-
Can. J. Zool.76: 1247–1253 (1998) © 1998 NRC Canada
1247
Received May 13, 1997. Accepted February 12, 1998.
F.J. Hernandez-Blazquez.Faculdade de Zootecnia eEngenharia de Alimentos, Universidade de São Paulo,Avenida Duque de Caxias Norte, 225/ZAB/USP, 13.630–000- Pirassununga (SP), Brazil (e-mail: [email protected]).J.R.M.C. Silva. Instituto de Ciências Biomédicas,Universidade de São Paulo, Avenida Duque de Caxias Norte,225/ZAB/USP, 13.630–000 - Pirassununga (SP), Brazil.
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:22 PM
Color profile: DisabledComposite Default screen
tion has taken place at several levels of biological organiza-tion (Clarke 1987; di Prisco et al. 1991; Eastman 1993). Thefeeding habits ofNotothenia neglectahave been well de-scribed (Johnston et al. 1991; Johnston and Battram 1993).It is a voracious carnivorous fish, consuming 1.4–2.3% of itsbody mass in food rich in proteins (Daniels 1982; Luxmore1985; Kock 1985). Although proteins are of great impor-tance in the energy strategy of fishes (Pandian andVivekanandan 1985), there have been no studies on how lowtemperature influences the process of intestinal uptake of in-tact macromolecules in Antarctic fishes. In this paper are de-scribed morphological aspects of the capture and transport ingreat quantity of a macromolecule (ferritin) byN. neglectaenterocytes and the location in the intestine where this pro-cess occurs.
Fifty-five N. neglectawith a mean mass of 885.37 g and meanbody length of 39.5 cm were collected in February 1996 and usedin this experiment. The fishes were captured at Admiralty Bay,King George Island, south Shetland Islands. The experiment wascarried out in fiberglass tanks containing seawater in a climate-controlled room at 0ºC in the biology laboratories of the BrazilianAntarctic Station “Comandante Ferraz.” After a 7-day fast, the ani-mals were divided into two groups. The first group (group I, oralroute) received 1 mL of a solution of 50% horse spleen ferritinin 0.9% saline (Ferritin Type I from horse spleen, Sigma,100 mg/mL) directly into the stomach by oral intubation with apolyethylene tube. These animals were subdivided into three sub-groups of 5 animals each that were killed 12, 24, and 48 h after
intubation. A fourth subgroup consisted of 10 animals killed 72 hlater. Five of these received a second dose of 1 mL of the sameferritin solution 24 h after the first injection, i.e., 48 h before thesamples were collected. All the animals were killed by immersionin 50 ppm benzocaine. Tissue samples were collected from thefirst, middle, and distal third of each intestinal loop as shown inFig. 1. The rectum was subdivided into 4 identical parts. An injec-tion of 0.5 mL of the same solution of ferritin was made into therectum of 20 fishes in a second group (group II, anal route) andsamples were taken after 6, 12, 24, and 48 h. Groups I and II eachhad a control group of 5 fishes, not starved, that received only0.9% saline solution and were killed after 72 h. For the light-microscopy studies, fixation was carried out in McDowell’s fixa-tive (McDowell and Trump 1976). Afterwards, the material wasdehydrated in 95% alcohol and embedded in Historesin Leica. Sec-tions 2 µm thick were prepared in the biology laboratories of theBrazilian Antarctic Station. Perls’ Prussian blue counterstainedwith a 1% solution of safranine O was used for detecting ferritin inthe intestine (Bancroft and Stevens 1982).
Samples from each segment were also collected forultrastructural study of the intestinal mucosa. The material wasfixed in a solution of 2.5% glutaraldehyde in 0.1 M phosphatebuffer, pH 7.2, at 4ºC, postfixed in 1% osmium tetroxide in thesame buffer for 2 h and in 0.5% uranyl acetate for 12 h, dehydratedthrough a graded series of ethanol solutions, and embedded inSpurr’s resin (Sigma). Ultrathin sections 70 nm thick were gath-ered in copper grids and stained with 2% uranyl acetate in distilledwater (Watson 1958) for 1 h, washed in distilled water, and thenstained in 0.5% lead citrate in distilled water for 1 h (Reynolds1963). For observing ferritin particles in the electron microscope,the bismuth subnitrate method as described by Ainsworth andKarnovsky (1971) was used. Ultrastructure was studied in a Jeol100 CX-II electron microscope at the Institute of Biomedical Sci-ences and a Philips EM-201C electron microscope at the Facultyof Veterinary Medicine and Zootechny at the University of SãoPaulo.
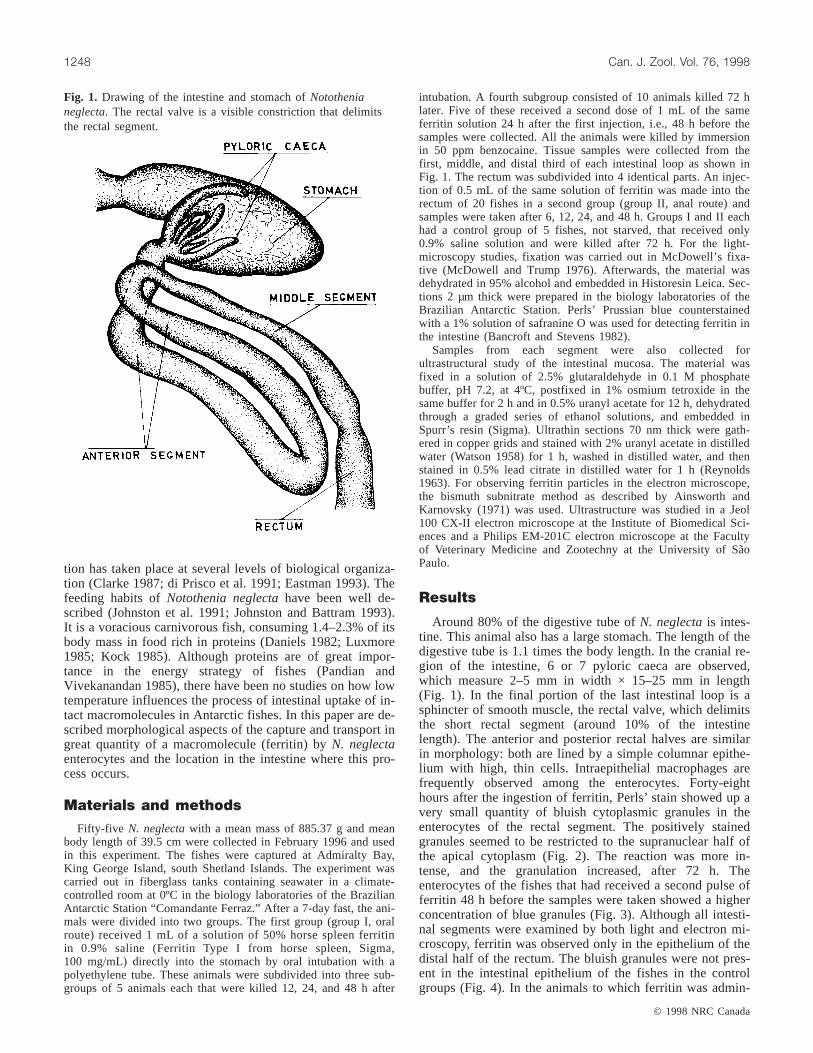
Around 80% of the digestive tube ofN. neglectais intes-tine. This animal also has a large stomach. The length of thedigestive tube is 1.1 times the body length. In the cranial re-gion of the intestine, 6 or 7 pyloric caeca are observed,which measure 2–5 mm in width × 15–25 mm in length(Fig. 1). In the final portion of the last intestinal loop is asphincter of smooth muscle, the rectal valve, which delimitsthe short rectal segment (around 10% of the intestinelength). The anterior and posterior rectal halves are similarin morphology: both are lined by a simple columnar epithe-lium with high, thin cells. Intraepithelial macrophages arefrequently observed among the enterocytes. Forty-eighthours after the ingestion of ferritin, Perls’ stain showed up avery small quantity of bluish cytoplasmic granules in theenterocytes of the rectal segment. The positively stainedgranules seemed to be restricted to the supranuclear half ofthe apical cytoplasm (Fig. 2). The reaction was more in-tense, and the granulation increased, after 72 h. Theenterocytes of the fishes that had received a second pulse offerritin 48 h before the samples were taken showed a higherconcentration of blue granules (Fig. 3). Although all intesti-nal segments were examined by both light and electron mi-croscopy, ferritin was observed only in the epithelium of thedistal half of the rectum. The bluish granules were not pres-ent in the intestinal epithelium of the fishes in the controlgroups (Fig. 4). In the animals to which ferritin was admin-
© 1998 NRC Canada
1248 Can. J. Zool. Vol. 76, 1998
Fig. 1. Drawing of the intestine and stomach ofNotothenianeglecta. The rectal valve is a visible constriction that delimitsthe rectal segment.
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:26 PM
Color profile: DisabledComposite Default screen

istered by the anal route (group II), the granules of theenterocytes appeared slightly bluish after 6 h. After 12 h, theintensity of the reaction and the quantity of reactive granulesin the animals in group II surpassed those observed at anygiven moment in the animals that received ferritin by theoral route. The more intense reaction and granulation tookplace in the animals in the 24-h subgroup of group II. Theimages observed in the 48-h subgroup were similar to thosein the 24-h one (Fig. 5).
In the electron micrographs, the enterocytes in the secondhalf of the rectal segment exhibit long microvilli. The sur-face membrane between the microvilli forms deepinvaginations that detach themselves from the surface, form-ing clear vesicles. The size of the vesicles increases towardthe nucleus (Fig. 6). The cytoplasmic region where thesehuge, clear vesicles appear coincides with the ferritin-freezone observed using light microscopy. The electron densityof the cytoplasmic vesicles also increases toward the nu-
© 1998 NRC Canada
Hernandez-Blazquez and Silva 1249
Fig. 2. Rectal segment 48 h after oral administration of ferritin. The solid line shows the absorptive zone of the apical cytoplasm,from the well-defined striated border (E) to the supranuclear region. The arrow shows the boundary between the region of clearvesicles and the region where the ferritin is stored in the form of bluish granules. Perls’ stain and safranine O. Scale bar = 10 µm.Fig. 3. Rectal segment 72 h after administration of two oral pulses of ferritin. Perls’ stain and safranine O. Scale bar = 10 µm.Fig. 4.Rectal segment of a control animal. No bluish granules are observed in the enterocytes of the fish that received saline without ferritin.Perls’ stain and safranine O. Scale bar = 50 µm.Fig. 5. Rectal enterocytes 24 h after anal administration of ferritin. Perls’ stain andsafranine O. Scale bar = 10 µm.
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:29 PM
Color profile: DisabledComposite Default screen
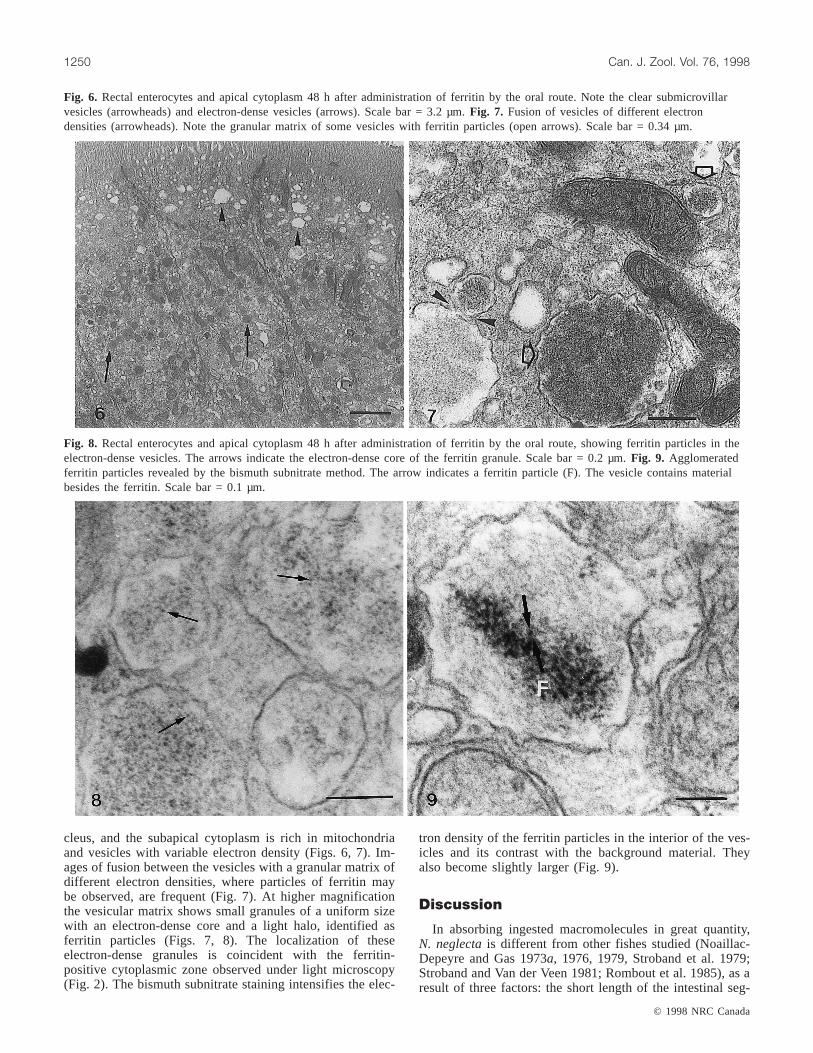
cleus, and the subapical cytoplasm is rich in mitochondriaand vesicles with variable electron density (Figs. 6, 7). Im-ages of fusion between the vesicles with a granular matrix ofdifferent electron densities, where particles of ferritin maybe observed, are frequent (Fig. 7). At higher magnificationthe vesicular matrix shows small granules of a uniform sizewith an electron-dense core and a light halo, identified asferritin particles (Figs. 7, 8). The localization of theseelectron-dense granules is coincident with the ferritin-positive cytoplasmic zone observed under light microscopy(Fig. 2). The bismuth subnitrate staining intensifies the elec-
tron density of the ferritin particles in the interior of the ves-icles and its contrast with the background material. Theyalso become slightly larger (Fig. 9).
In absorbing ingested macromolecules in great quantity,N. neglectais different from other fishes studied (Noaillac-Depeyre and Gas 1973a, 1976, 1979, Stroband et al. 1979;Stroband and Van der Veen 1981; Rombout et al. 1985), as aresult of three factors: the short length of the intestinal seg-
© 1998 NRC Canada
1250 Can. J. Zool. Vol. 76, 1998
Fig. 6. Rectal enterocytes and apical cytoplasm 48 h after administration of ferritin by the oral route. Note the clear submicrovillarvesicles (arrowheads) and electron-dense vesicles (arrows). Scale bar = 3.2 µm.Fig. 7. Fusion of vesicles of different electrondensities (arrowheads). Note the granular matrix of some vesicles with ferritin particles (open arrows). Scale bar = 0.34 µm.
Fig. 8. Rectal enterocytes and apical cytoplasm 48 h after administration of ferritin by the oral route, showing ferritin particles in theelectron-dense vesicles. The arrows indicate the electron-dense core of the ferritin granule. Scale bar = 0.2 µm.Fig. 9. Agglomeratedferritin particles revealed by the bismuth subnitrate method. The arrow indicates a ferritin particle (F). The vesicle contains materialbesides the ferritin. Scale bar = 0.1 µm.
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:32 PM
Color profile: DisabledComposite Default screen

ment where the process occurs; the long time (48 h) neededto observe the marker in the enterocytes; and the extremelydistal position of the intestinal segment, which is preanal. Inother fishes the preanal segment appears to be specializedfor the transport of water or ions.
In N. neglecta, the uptake and intracellular storage of pro-tein macromolecules administered by the oral and analroutes occur in the short preanal region that corresponds tothe last half of the postvalvular or rectal segment, whichconstitutes only 5% of the total length of the intestine. Theintestine ofN. neglectacan be considered short and typicalof carnivorous fishes. It can be concluded that the relativesuperficial area available for the uptake and accumulation ofmacromolecules is small, only that of the end of the rectum.In other species this process is an absorptive one, quantita-tively more meaningful because the length of the intestinalsegment where the process occurs is greater and the inten-sity is higher (Rombout et al. 1985). In stomachless speciesand in fish larvae, uptake of macromolecules occurs in asegment that is 12–35% of the intestinal length, usually themiddle one (Noaillac-Depeyre and Gas 1973a, 1976;Stroband et al. 1979; Stroband and Van der Veen 1981;Rombout et al. 1985). Uptake of proteins may also occur, al-though to a lesser extent, in the proximal intestine of fishes,as described in carp by Rombout et al. (1985). The caudal orrectal enterocytes of other fishes, such as the carpCyprinuscarpio (Noaillac-Depeyre and Gas 1973b), grass carp(Stroband et al. 1979), tench,Tinca tinca(Noaillac-Depeyreand Gas 1976), perch,Perca fluviatus (Noaillac-Depeyreand Gas 1979), and catfish,Ameiurus nebulosus(Noaillac-Depeyre and Gas 1983), show the ultrastructural features ofcells that actively transport water and ions, but these charac-teristics are not found in the rectal epithelium ofN. neglecta.
After an oral dose of ferritin, the reaction to Perls’ stainby the rectal enterocytes ofN. neglectawas weak after 48 hand more intense after 72 h. The reaction increased slightlyin strength when a second oral dose of ferritin was adminis-tered 48 h before samples were collected. The data obtainedduring this research clearly show that the uptake and accu-mulation of oral ferritin inN. neglectaconstitute a continu-ous, slow, and not extensive process. In fishes from warmerclimates, intestinal uptake of similar doses of ferritin startswithin a period ranging from 2 to 5 h, depending on the ad-ministration route (Stroband et al. 1979; Stroband and Kroon1981; Noaillac-Depeyre and Gas 1973a, 1976, 1983; Georgo-poulou et al. 1988; Rombout and van den Berg 1989). How-ever, anal administration of ferritin showed that the rectalenterocytes ofN. neglectaare able to capture large quanti-ties of high molecular weight proteins as quickly as those ofother fishes, since by the anal route, absorption started after6 h and it reached its peak at 24 h.
The potential for intense absorption by the rectal cellsmay be inferred from their ultrastructure, which is typical ofactive absorptive cells. They have long microvilli, and thereis a great endocytotic activity at the base of these structures,with a large number of transport vesicles. Large clear vesi-cles and a subapical tubulovesicular network are evident. Aferritin granule appears inside the vesicle as a dark pointwith a halo of light around it. Ferritin uptake seems to occurin a nonselective way, not linked to the membranes, since itis found distributed as agglomerates or scattered in small
vesicles. Rombout et al. (1985) have also observednonselective absorption of ferritin in the intestinal epithe-lium of C. carpio. Sire and Vernier (1992) demonstrated inother species that the cytoplasmic vesicles containing ferritinare phagolysosomes, since lysosomal enzymes are also pres-ent. The form of the phagolysosomes also varies accordingto the species. They can take the form of a large vesicle,called the supranuclear vacuole, as in trout, carp, and perch(Noaillac-Depeyre and Gas 1979; Georgopoulou et al. 1985;Rombout and van den Berg 1989), or may be small and nu-merous, as inN. neglecta.
Although the quantity of ferritin in cytoplasmic vesiclesof N. neglectais small, the vesicular matrix contains a lesselectron-dense material, indicating that besides the ferritinmolecules, other substances are being absorbed from the rec-tal lumen. This material is probably of endogenous origin,since the fish had been fasting for 7 days. Several moleculesare secreted into the fish intestine and may be reabsorbedenterocytes. In mammals the recycling of pancreatic en-zymes through the intestinal epithelium was stressed by Dia-mond (1976). Molecules that are present in great quantitiesexclusively in the intestinal fluid of Antarctic fishes, whereit probably arrives with the bile, are antifreeze glyco-peptides, or AFGPs (O’Grady et al. 1982a, 1982b, 1983; DeVries 1988). In Antarctic fishes, such substances have mo-lecular weights ranging from 2600 to 34 000 (De Vries1988). According to DeVries (1988), the presence of AFGPsin the intestine is necessary to avoid ice formation. With thereabsorption of water by the intestine, AFGP concentrationincreases in the distal extremity of this organ, being lostwith the faeces (O’Grady et al. 1983). In DeVries’ (1988)words, the continuous faecal loss of AFGPs is “energeticallywasteful” because of low metabolic rates at low tempera-tures. The demonstration of a rectal epithelial transport sys-tem that is able to capture and store large protein molecules,exactly where the concentration of AFGPs must be greater,leads to the unavoidable question of whether the AFGPs fol-low the same route as these macromolecules. This questioncannot be answered by our results; more studies on this sub-ject are necessary. The presence of a rectal sphincter trans-forms the rectum into a separate compartment where thefaeces may be retained for some time. The location of an ab-sorptive segment in a such distal position, instead of an ion-and water-transporting segment, would have the advantagefor this Antarctic fish of avoiding excessive loss of proteinswith the faeces.
Financial support from the Fundaçao de Amparo àPesquise do Estado de São Paulo (grant No. 94/6255.8),Conselho Nacional de Desenvolvimento Científico eTechnológico (grant Nos. 510581/93-5 and 48.0735/94-8),and PROANTAR/SECIRM (grant No. 67.0004/94) and lo-gistic support from Marinha do Brasil are acknowledged.
Ainsworth, S.K., and Karnovsky, M.J. 1971. An ultrastructuralstaining method for enhancing the size and electron opacity offerritin in thin sections. J. Histochem. Cytochem.20: 225–229.
© 1998 NRC Canada
Hernandez-Blazquez and Silva 1251
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:36 PM
Color profile: DisabledComposite Default screen

Bancroft, J.D., and Stevens, A. 1982. Theory and practice ofhistological techniques. Churchill Livingstone, Oxford.
Brandtzaeg, P., and Krajci, P. 1995. The enterocyte and immuno-globulin transport.In Mucosal immunity and the gut epithelium.Edited byS. Auricchio, A. Ferguson, and R. Troncone. Karger,Basel and Berlin. pp. 1–17.
Clarke, A. 1987. The adaptation of aquatic animals to low temper-atures.In The effects of low temperature on biological systems.Edited byB.W.W. Grout and G.J. Morris. Edward Arnold, Ltd.,London. pp. 315–348.
Daniels, R.A. 1982. Feeding ecology of some fishes of the antarc-tic peninsula. Fish. Bull.80: 575–588.
DeVries, A.L. 1988. The role of antifreeze glycopeptides and pep-tides in the freezing avoidance of antartic fishes. Comp.Biochem. Physiol. B,90: 611–621.
Diamond, J.M. 1976. Reabsorption of digestive enzymes: playingwith poison. Nature (Lond.),211: 111–112.
di Prisco, G., Maresca, B., and Tota, B. 1991. Biology of antarcticfish. Springer-Verlag, New York.
Dorin, D., Martin, P., Sire, M.F., Smal, J., and Vernier, J.M. 1993a.Protein uptake by intestinal macrophages and eosinophilicgranulocytes in trout: an in vivo study. Biol. Cell,79: 37–44.
Dorin, D., Sire, M.F., and Vernier, J.M. 1993b. Endocytosis andintracellular degradation of heterologous protein by eosinophilicgranulocytes isolated from rainbow trout (Oncorhyncus mykiss)posterior intestine. Biol. Cell,79: 219–224.
Dorin, D., Sire M.F., and Vernier, J.M. 1994. Demonstration of anantibody response of the anterior kidney following intestinal ad-ministration of a soluble protein antigen in trout. Comp.Biochem. Physiol. B,109: 499–509.
Eastman, J.T. 1993. Antartic fish biology: evolution in a uniqueenvironment. Academic Press, London.
Gauthier, G.G., and Landis, S.C. 1972. The relationship ofultrastructural and cytochemical features to absorptive activityin the goldfish intestine. Anat. Rec.172: 675–701.
Georgopoulou, U., and Vernier, J.M. 1986. Local immunologicalresponse in the posterior intestinal segment of the rainbow troutafter oral administration of macromolecules. Dev. Comp.Immunol. 10: 529–537.
Georgopoulou, U., Sire, M.F., and Vernier, J.M. 1985.Macromolecular absorption of proteins by epithelial cells of theposterior intestine segment and their intracellular digestion inthe rainbow trout. Biol. Cell,53: 269–282.
Georgopoulou, U., Sire, M.F., and Vernier J.M. 1986. Immunologi-cal demonstration of intestinal absorption and digestion of pro-tein macromolecules in the trout (Salmo gairdneri). Cell TissueRes.245: 387–395.
Georgopoulou, U., Dabrowski, K., Sire, M.F., and Vernier, J.M.1988. Absorption of intact proteins by the intestinal epitheliumof trout Salmo gairdneri: a luminescence enzyme immunoassayand cytochemical study. Cell Tissue Res.251: 145–152.
Hofer, R. 1982. Protein digestion and proteolytic activity in the di-gestive tract of an omnivorous cyprinid. Comp. Biochem.Physiol. A, 72: 55–63.
Iida, H., and Yamamoto, T. 1985. Intracellular transport of horse-radish peroxidase in the absorptive cells of goldfish hindgut invitro with special reference to the cytoplasmic tubules. Cell Tis-sue Res.240: 553–560.
Johnston, I.A., and Battram, J. 1993. Feeding energetics and me-tabolism in demersal fish species from Antarctic, temperate andtropical environments. Mar. Biol. (Berl.),115: 7–14.
Johnston, I.A., Clarke, A., and Ward, P. 1991. Temperature andmetabolic rate in sedentary fish from Antarctic, North Sea andIndo-West Pacific Ocean. Mar. Biol. (Berl.),109: 191–195.
Kock, K.H. 1985. Krill consumption by AntarticNototheniafish.In Antartic nutrient cycles and food webs.Edited by W.R.Sigfried, R.R. Condy, and R.M. Laws. Springer-Verlag, Berlin.pp. 437–444.
Luxmore, R.A. 1985. The energy budget of a population of theAntartic isopodSerolis polita. In Antartic nutrient cycles andfood webs.Edited by W.R. Sigfried, R.R. Condy, and R.M.Laws. Springer-Verlag, Berlin. pp. 389–396.
McDowell, E.M., and Trump, B.F. 1976. Histologic fixatives suit-able for diagnostic light and electron microscopy. Arch. Pathol.Lab. Med.100: 405–414.
Noaillac-Depeyre, J., and Gas, N. 1973a. Absorption of proteinmacromolecules by the enterocytes of the carp (Cyprinuscarpio L.): ultrastructural and cytochemical study. Z. Zellforsch.Mikrosk. Anat. 146: 525–541.
Noaillac-Depeyre, J., and Gas, N. 1973b. Mise en évidence d’unezone adaptée au transport des ions dans l’intestin de la Carpecommune (Cyprinus carpioL.). C. R. Acad. Sci. Paris,276:773–776.
Noaillac-Depeyre, J., and Gas, N. 1976. Electron microscopicstudy on the gut epithelium of the tench (Tinca tincaL.) with re-spect to its absorptive functions. Tissue Cell,8:11–30.
Noaillac-Depeyre, J., and Gas, N. 1979. Structure and function ofintestinal epithelial cells in the perch (Perca fluviatusL.). Anat.Rec.15: 621–640.
Noaillac-Depeyre, J., and Gas, N. 1983. Étude cytophysiologiquede l’épithélium intestinal du poisson-chat (AmeiurusnebulosusL.). Can. J. Zool.61: 2556–2573.
O’Grady, S.M., Clarke A., and DeVries, A.L. 1982a. Characteriza-tion of glycoprotein antifreeze biosynthesis in isolatedhepatocytes fromPagothenia borchgrevinsky. J. Exp. Zool.220:179–189.
O’Grady, S.M., Ellory, J.C., and DeVries, A.L. 1982b. Protein andglycoprotein antifreezes in the intestinal fluid of polar fishes. J.Exp. Biol. 98: 429–438.
O’Grady, S.M., Ellory, J.C., and DeVries, A.L. 1983. The role oflow molecular weight antifreeze glycopeptides in the bile andintestinal fluid of antartic fishes. J. Exp. Biol.104: 149–162.
Pandian, T.J., and Vivekanandan, E. 1985. Energetics of feedingand digestion.In Fish energetics: new perspectives.Edited byP.Tyler and P. Calows. The Johns Hopkins University Press, Balti-more. pp. 99–124.
Reynolds, E.S. 1963. The use of lead citrate at high pH as an elec-tron opaque stain in electron microscopy. J. Cell Biol.17: 208–213.
Rombout, J.H.W.M., and van den Berg, A.A. 1989. Immunologicalimportance of the second gut segment of carp. I. Uptake andprocessing of antigens by epithelial cells and macrophages. J.Fish Biol. 35: 13–22.
Rombout, J.H.W.M., Lamers, C.H.J., Helfrich, M.H., Dekker, A.,and Taverne-Thiele, J.J. 1985. Uptake and transport of intactmacromolecules in the intestinal epithelium of carp (Cyprinuscarpio L.) and the possible immunological implications. CellTissue Res.239: 519–530.
Rombout, J.H.W.M., Bot, H.E., and Taverne-Thiele, J.J. 1989a.Immunological importance of the second gut segment of carp.II. Characterization of mucosal leukocytes. J. Fish Biol.35:167–178.
Rombout, J.H.W.M., van den Berg, A.A., van den Berg, C.T.G.A.,Witte, P., and Egberts, E. 1989b. Immunological importance ofthe second gut segment of carp. III. Systemic and mucosal im-mune responses after immunization with soluble or particulateantigen. J. Fish Biol.35: 179–86.
© 1998 NRC Canada
1252 Can. J. Zool. Vol. 76, 1998
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:40 PM
Color profile: DisabledComposite Default screen

© 1998 NRC Canada
Hernandez-Blazquez and Silva 1253
Sire, M.F., and Vernier, J.M. 1992. Intestinal absorption of proteinin teleost fish. Comp. Biochem. Physiol. A,103: 771–781.
Sire M.F., and Vernier, J.M. 1995. Partial characterization ofeosinophilic granule cells (ECGs) and identification of mastcells of the intestinal lamina propria in rainbow trout(Oncorhynchys mykiss): biochemical and cytochemical study.Biol. Cell, 85: 35–41.
Stroband, H.W.J., and Kroon, A.G. 1981. The development of thestomach inClarias lazeraand the intestinal absorption of pro-tein macromolecules. Cell Tissue Res.215: 397–415.
Stroband, H.W.J., and Van der Veen, F.H. 1981. Localization ofprotein absorption during transport of food in the intestine of
grasscarpCtenopharyngodon idella(Val.). J. Exp. Zool.218:249–256.
Stroband, H.W.J., Meer, H.V.D., and Timmermans, L.P.M. 1979.Regional functional differentiation in the gut of the grasscarpCtenopharyngodon idella. Cell Tissue Res.64: 235–249.
Walker, W.A., and Sanderson, I.R. 1995. The enterocyte and anti-gen transport.In Mucosal immunity and the gut epithelium.Edited byS. Auricchio, A. Ferguson, and R. Troncone. Karger,Basel. pp. 18–31.
Watson, M.L. 1958. Staining of tissue sections for electron micros-copy with heavy metals. J. Biophys. Biochem. Cytol.4: 475–478.
I:\cjz\cjz76\cjz-07\ZooJuly(A).vpFriday, December 04, 1998 4:36:41 PM
Color profile: DisabledComposite Default screen
Copyright © 2022 FDOKUMEN