
The Ocular Lens Epithelium
27
Bioscience Reports, Vol. 21, No. 4, August 2001 ( 2002) MINI REVIEW The Ocular Lens Epithelium Suraj P. Bhat 1 Receiûed April 12, 2001 An adult lens contains two easily discernible, morphologically distinct compartments, the epithelium and the fiber-cell mass. The fiber-cell mass provides the lens with its functional phenotype, transparency. Metabolically, in comparison to the fiber cells the epithelium is the more active compartment of the ocular lens. For the purposes of this review we will only discuss the surface epithelium that covers the anterior face of the adult ocular lens. This single layer of cells, in addition to acting as a metabolic engine that sustains the physiological health of this tissue, also works as a source of stem cells, providing precursor cells, which through molecular and morphological differentiation give rise to fiber cells. Morphological simplicity, defined developmental history and easy access to the experi- menter make this epithelium a choice starting material for investigations that seek to address universal questions of cell growth, development, epithelial function, cancer and aging. There are two important aspects of the lens epithelium that make it highly relevant to the modern biologist. Firstly, there are no known clinically recognizable cancers of the ocular lens. Considering that most of the known malignancies are epithelial in origin this observation is more than an academic curiosity. The lack of vasculature in the lens may explain the absence of tumors in this tissue, but this provides only a teleological basis to a very important question for which the answers must reside in the molecular make-up and physiology of the lens epithelial cells. Secondly, lens epithelium as a morphological entity in the human lens is first recognizable in the 5th–6th week of gestation. It stays in this morphological state as the anterior epithelium of the lens for the rest of the life, making it an attractive paradigm for the study of the effects of aging on epithelial function. What follows is a brief overview of the present status and lacunae in our understanding of the biology of the lens epithelium. KEY WORDS: Eye lens; epithelial cells; fiber cells; programmed cell death; cataract. THE EPITHELIUM AND ITS CELLS Developmentally, all of the lens can be considered as an asymmetrically folded epi- thelium (Zamphigi et al., 2000) with region-specific morphological and physiological specialization. A single layer of cells, the lens epithelium covers the anterior face of the lens that faces the cornea and the outside world. The lens epithelium ends on the rims of the anterior surface (oval or circular in shape, depending on the species) 1 Vision Molecular Biology Laboratory, Jules Stein Eye Institute, 100 Stein Plaza, BH 623, UCLA School of Medicine, Los Angeles, CA 90095-0000; e-mail: [email protected] 537 0144-8463010800-05370 2002 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Ocular Lens Epithelium

Bioscience Reports, Vol. 21, No. 4, August 2001 ( 2002)
MINI REVIEW
The Ocular Lens Epithelium
Suraj P. Bhat1
Receiûed April 12, 2001
An adult lens contains two easily discernible, morphologically distinct compartments, theepithelium and the fiber-cell mass. The fiber-cell mass provides the lens with its functionalphenotype, transparency. Metabolically, in comparison to the fiber cells the epithelium isthe more active compartment of the ocular lens. For the purposes of this review we willonly discuss the surface epithelium that covers the anterior face of the adult ocular lens.This single layer of cells, in addition to acting as a metabolic engine that sustains thephysiological health of this tissue, also works as a source of stem cells, providing precursorcells, which through molecular and morphological differentiation give rise to fiber cells.Morphological simplicity, defined developmental history and easy access to the experi-menter make this epithelium a choice starting material for investigations that seek toaddress universal questions of cell growth, development, epithelial function, cancer andaging. There are two important aspects of the lens epithelium that make it highly relevantto the modern biologist. Firstly, there are no known clinically recognizable cancers of theocular lens. Considering that most of the known malignancies are epithelial in origin thisobservation is more than an academic curiosity. The lack of vasculature in the lens mayexplain the absence of tumors in this tissue, but this provides only a teleological basis to avery important question for which the answers must reside in the molecular make-up andphysiology of the lens epithelial cells. Secondly, lens epithelium as a morphological entityin the human lens is first recognizable in the 5th–6th week of gestation. It stays in thismorphological state as the anterior epithelium of the lens for the rest of the life, making itan attractive paradigm for the study of the effects of aging on epithelial function. Whatfollows is a brief overview of the present status and lacunae in our understanding of thebiology of the lens epithelium.
KEY WORDS: Eye lens; epithelial cells; fiber cells; programmed cell death; cataract.
THE EPITHELIUM AND ITS CELLS
Developmentally, all of the lens can be considered as an asymmetrically folded epi-thelium (Zamphigi et al., 2000) with region-specific morphological and physiologicalspecialization. A single layer of cells, the lens epithelium covers the anterior face ofthe lens that faces the cornea and the outside world. The lens epithelium ends onthe rims of the anterior surface (oval or circular in shape, depending on the species)
1Vision Molecular Biology Laboratory, Jules Stein Eye Institute, 100 Stein Plaza, BH 623, UCLA Schoolof Medicine, Los Angeles, CA 90095-0000; e-mail: [email protected]
537
0144-8463010800-05370 2002 Plenum Publishing Corporation

538 Bhat
Fig. 1. Simplified drawing of the ocular lens. Note that the epithelium has three regionsof cells in non-dividing phase (central epithelium), in dividing phase (germinative) anddifferentiating phase (equatorial). The anterior and posterior sutures are formed by themeeting of the elongating fiber cells that make the bulk of the lens mass. The surround-ing capsule (shaded area around the lens) indicates that the basal surface of the epi-thelium and the fiber cells is on the outside, while the apical surface faces the inside ofthe lens. ‘‘Apical interface’’, an area of contact between the apical surfaces of the epi-thelial cells and the fiber cells is not indicated. It is the area just below the epitheliallayer.
of the lens (Fig. 1). It contains cells in the central region that do not divide and areessentially ‘‘quiescent’’, surrounded by a germinativedividing zone of cells, followed(at the equatorial fringe) by the dividing cells that differentiate into fiber cells. Inthe newborn rat lens, as assessed by tritiated thymidine labeling in the intact animal,the epithelium contains about 4.7% of dividing cells. This decreases to 1.7% by day6 and then 0.8% by day 35 (Mikulicih and Young, 1963). A remarkable feature ofthis epithelium is its capacity to divide and differentiate almost all through the lifeof an individual. This feature of sustained growth is very much similar to its closestembryological sibling, the cells in the skin. The difference between the two howeveris noteworthy. The pattern of growth that results from the division and differen-tiation of the lens epithelial cells at the equatorial rim gives lens its unique physiog-nomy. The new (fiber) cells made from the lens epithelium, at its equatorial rim,keep adding on top of the older fiber cells in such a way so that the oldest cells areburied deep inside the lens. Thus, by peeling away, one fiber at a time one couldtheoretically unveil a temporal catalog of the molecular changes starting from theoutside (the youngest or the most recent fibers) to the center (the oldest, the‘‘nucleus’’ of the lens, which contains fibers laid in 5th6th week of gestation inhumans).
The cells in the lens epithelium represent typical epithelial morphology. Theyare cuboidal, presenting a cobble-stone-like appearance in their native state and inûitro, if cultured without excessive passaging (Fig. 2). These cells in the epithelium

Ocular Lens Epithelium 539
Fig. 2. Photomicrographs of the native human lens epithelium (A) and cultured human fetal lensepithelial cells (B). A, taken, with permission of the copyright holder, Association for Research inVision and Ophthalmology, from Mercantonio et al. (2000) Inûest. Ophthalmol. and Vis. Sci.41:1130–1141. B, taken, with permission of the copyright holder, Academic Press, Orlandlo,Florida, from Nagineni and Bhat (1988) Deûelopmental Biology 130:402–405.
are packed with very little intercellular space. Francois and Rabaey (Francois andRabaey, 1951) observed lens epithelium under a phase-contrast microscope. Theyreported the presence of ‘‘pale polyhedral’’ and ‘‘dark star-shaped’’ cell types. Arecent in ûiûo study (Balaram et al., 2000) using the non-contact specular microscopyrecognized four morphological features of the live human lens epithelium. Thesewere categorized as ‘‘linear furrows’’, ‘‘columnar organization’’, ‘‘puffy clouds’’ and‘‘black holes’’. These morphological features remain without a biochemical or mol-ecular definition at this time. The diameter of the human lens epithelial cells rangesfrom 9–17 mm (Brown and Bron, 1987). The cell size has been reported to increasewith age (Perry et al., 1979; Robinson et al., 1990), which suggests a change in thecell density. Females have been reported to have higher cell density in the humanlens epithelium than in males (Konofsky et al., 1987; Guggenmoos-Holtzmann etal., 1989). It must be recognized however, that both the cell size and cell density isspecific to a particular region or area of the lens epithelium. For example, in themonkey lens, in the germinative zone, cells increase in size by about 30% from age7 years to age 24.5 years, while the central zone epithelial cells (in the same period)increase by 150%. Notably, within the same period the cells in the germinative zoneincrease in number, while cells in the central epithelium seem to decrease in number(by about 17%) (Kuszak, 1997).
The relationship between cell density and age is interesting, although contro-versial. An earlier report (Guggenmoos-Holtzmann et al., 1989) that there is an age-related decrease in the cell density in the lens epithelium has been recently confirmed(Balaram et al., 2000). This recent study calculated loss of 675 cellmm2 in a 75-yearlife span (that amounts to a loss of 14% of the cells). This estimate is based on theunproven assumption that the rate of loss is linear with age, however it is not verydifferent from that reported in the aging monkey lens central epithelium (Kuszak,1997). Others have found no such relationship (Fagerholm and Philipson, 1981;Karin et al., 1987). Karim et al. (1987) reported a decrease in the mitotic index oflens epithelial cells under normal as well as cataractous conditions. Harocopos et

540 Bhat
al. (1998) working with capsulotomy specimens from cataract surgeries and non-cataractous lenses from eye banks concluded that there was no relationship betweencell density and the severity of cataract or between cell density and age. They how-ever reported that in cataractous lenses the epithelium directly over the opaque (cat-aractous) had higher cell density when compared to that overlaying the transparentregions. It is possible that a loss of a patch of cells overlying a cataractous fiber cellarea may lead to the activation of cell division and therefore, higher cell density.This observation, however, needs to be confirmed.
The relationship between cell proliferation and cataracts is difficult to establishparticularly in a causal context. A recent study with sugar cataracts makes a goodexample: A secondary complication of diabetes is cataractogenesis due to accumula-tion of sugarsugar alcohols in the lens causing pathological water uptake. Strucket al. (2000) make an assessment of lens epithelial cell densities in type-II diabetics(3691C346 cellsmm2) vs. non-diabetes (4162C504 cellsmm2). Among cataractouslenses, the nuclear cataract had the highest cell density (4250C513 cellmm2). Inter-estingly, they found no significant differences in cell number between the females(4036C525 cellsmm2) and the males (3788C421 cellmm2) in disagreement withearlier studies cited above. These authors also reported that the lowest mean celldensity in the lens epithelium was associated with posterior subcapsular cataracts(3620C333 cellsmm2). This is quite interesting because it suggests an aberration inthe mechanism that regulates (inhibits?) migration of cells to the posterior subscap-sular area. Whether decreased cell density in the anterior epithelium is also relatedto this aberration can only be speculated.
If it is accepted that there is a decrease in cell density either with age or withcataracts we must ponder the question as to how does this happen? Do cells in thelens epithelium die by apoptosisnecrosis? If so what triggers these processes? It isa common observation that cells when cultured at low densities do not grow well.This is true of lens epithelial cells also. It has been shown that the medium fromhigh-density lens epithelial cell cultures promotes the survival of the cells in the low-density cultures suggesting that these cells secrete growthsurvival factors (Ishizakiet al., 1993). At lower cell densities these factors are diluted and are, therefore, notavailable to cells for their survival. This leads to the questions: Are there lens-specificphysiological factors that maintain a minimal cell density in the lens epithelium toavoid intervention by apoptosis? How much decrease in cell density is toleratedbefore it becomes pathological? Recently proteinsfactors that seem to enhance thesurvival of lens epithelial cells in culture have been identified (Davidson et al., 1998;Singh et al., 2000).
DEVELOPMENTAL HISTORY
Emergence of a transparent lens is the culmination of morphogeneticremodeling, cell differentiation and development from one cell type, the surface(head) ectoderm (Spemann, 1901; Lewis, 1904; Jacob and Sater, 1988; Saha et al.1989; Grainger, 1992). Lens epithelium represents the region of the surface ectodermthat is part of the lens ‘‘field’’ in the developing embryo. The lens morphogenesiscommences with the appearance of a thickening in the pre-determined area of the

Ocular Lens Epithelium 541
Fig. 3. Early stages in the formation of the lens vesicle (A–D) and the lens. Stage E representsasymmetric morphogenesis in the lens vesicle. Posterior epithelial cells elongate (primary fiber cells)and fill the lumen of lens vesicle (not completely filled in the drawing), resulting in the lens havingno epithelium on the posterior surface. Differentiating secondary fiber cells (shown in the illus-tration) from the anterior epithelium are the main contributor to the growth of the postnatal lens.
surface ectoderm (Spemann, 1962; Li et al., 1994), the future ‘‘lens placode’’, a pointwhere the lateral protuberance from the fore brain invaginates to form what becomesthe ‘‘optic vesicle’’ or ‘‘the optic cup’’, the surface ectoderm (lens placode) alsoinvaginates and pinches off as a lens vesicle. Studies done with excised surface ecto-derm and the optic vesicle have shown that a physical contact between the twosurfaces is not essential for the formation of the lens vesicle (Jacob and Sater, 1988).Molecular cues that emanate, possibly from both the interactive surfaces, are there-fore diffusible. These inductive messengers have not yet been unequivocally ident-ified. Gene products including growth factors and transcription factors associatedwith early morphogenesis of the lens have been detailed (Ogini and Yasuda, 1998;Jean et al., 1998) and will not be discussed here. It is however interesting to followthe emergence of the incipient lens, complete with fiber cells so that a relationship(albeit morphologicalembryological) between the adult lens epithelium and the restof the lens (fiber cells) can be envisioned. This process is polar (with respect to theepithelium), in its progression and is briefly discussed below (Fig. 3).
The cells that line the lens vesicle have their apical surfaces pointing inwardsinto the lumen of the vesicle and the basal surfaces outwards contacting the base-ment membrane that makes the ‘‘capsule’’ that surrounds the vesicle (and eventuallythe lens). The transformation of this vesicle into a structure that is recognizable asa lens starts with the differentiation of the posterior epithelial cells into fiber cells(Fig. 3E). Differentiation involves elongation of these cells in an anterio-posteriorobliterating the lumen of the lens vesicle. These fiber cells are known as the ‘‘primaryfiber cells’’. This process brings the apical surface (ends) of the elongated fiber cells

542 Bhat
(posterior epithelial cells) in direct contact with the apical surfaces of the anteriorepithelial cells, this area of contact is known as the ‘‘apical interface’’ (Zamphigi etal., 2000). The posterior surface of the lens thus lacks any epithelium having beentransformed into fiber cells. The anterior epithelium, which by dint of its distancefrom the bottom of the optic cup presumably escapes the molecular influences ofthe optic cup (future retina) remains undifferentiated. The proximity of embryonicretina to the posterior lens vesicle has been suggested to promote lens fiber celldifferentiation and morphogenesis (McAvovy and Fernon, 1984). A large numberof growth factors have been implicated in this process but none unequivocally(McAvovy and Chamberlain, 1990; Lang, 1999).
The asymmetric morphogenesis within the lens vesicle (Fig. 3) results in thepresence of an epithelial layer of cells on the anterior face of the lens covering amass of fiber cells. The future growth of the lens is dependent on this anterior epi-thelium. It is the cells at the equatorial zone of this epithelium that elongate in ananterio-posterior fashion into ribbon-like long fiber cells that are responsible for thegrowth of the lens beyond the 6-8th week of gestation. These fiber cells that areproduced from the edges of the anterior epithelium are known as ‘‘secondary fibercells’’ as opposed to ‘‘primary fiber cells’’ that were made from the posterior epi-thelial cells of the lens vesicle (Fig. 3).
It is reasonable to expect that the physiology and function of the lens epitheliumwill be influenced by the aqueous humor in the front, by vitreous humor in the backand by the ciliary body at the equatorial zone. The ciliary body, which is derivedfrom the anterior lip of the optic cup, interestingly is (at least morphologically)reminiscent of the ‘‘undifferentiated’’ neuroretina, which has been suggested toinfluence the differentiation of the posterior epithelial cells in the lens vesicle (seeJacob and Sater, 1988; Saha et al., 1989; Grainger, 1992). Young and associates(Mikulicih and Young, 1963) reasoned that the surrounding ciliary body must havea regulatory influence on cell division and DNA synthesis in the equatorial regionof the lens epithelium. Whether the surrounding ciliary body has any molecularinfluence on the growth and differentiation of the fiber cells in the equatorial regionof the lens has not been directly investigated. Nagineni and Bhat (1992), showedthat a co-culture of the ciliary body fibroblasts and the lens epithelial cells (bothisolated from the human fetal eyes) results in the appearance of differentiating fibersat the interface of the fibroblasts and the lens epithelial cells. This investigation didnot address the question as to which cell type ûiz., the epithelial or the fibroblastsdifferentiated to cells that expressed crystallins, although embryological history ofthe two would suggest that it was the epithelial cells. This in ûitro culture system hasa promise of providing molecular identification of communicating influencesbetween these two cell types. A discussion on what growth factors and agents maystimulate differentiation in the lens epithelial cells (Jean et al., 1998; Lang, 1999) isbeyond the scope of this review. However it is appropriate to comment on the pos-itional integrity of the epithelium and the environment around it. This is elucidatedin two publications motivated by two different investigations about three decadesapart. Coulombre and Coulombre (1963) rotated the developing lens 180° therebyexposing the epithelium to the vitreous humor and the retinal influences, this startedsecondary fiber differentiation of the epithelial cells. Hightower and McReady (1991)

Ocular Lens Epithelium 543
discovered that only when the anterior surface of the lens was exposed to selinitedid the cataract develop. Exposure of the posterior face that lacks the epithelial layerhad no effect. These observations directly point to a very important conclusion thatan alteration in the environment around the lens cannot only alter the developmentalstatus of the overlying epithelium but it can lead to physiological changes in the restof the fiber mass.
EPITHELIUM IS THE MAJOR SITE OF TRANSPORT/METABOLISM/DETOXIFICATION
The overall metabolic status of the fiber cells in the absence of endoplasmicreticulum, mitochondria and a nucleus is comparatively very low (Lieska et al.,1992). There is no vascular system as we know it that would take nutrients to thefiber cells and remove metabolicphysiologic waste replenish the intra—and inter-cellular milieu of the lens. Mere diffusion as a process to sustain the slow but sub-stantial physiology of the ocular lens will be insufficient to accomplish thisefficiently. A study of relative rates of transport across the anterior and posteriorsurfaces of the lens had led to the model of the ‘‘pump-leak’’ system (Kinsey andReddy, 1965; Becker and Cotlier, 1962; Harris and Becker, 1965). The dynamics ofthe active transport across the lens epithelium from the aqueous humor creates agradient of NaC and KC ions in the lens; high KC in the anterior and high NaC inthe posterior of the lens (Paterson, 1972). Epithelium does most of the work inmaintaining a low sodium concentration in the lens and an active resting potential,pumping in KC and extruding NaC, which enters the lens from the posterior surfaceby diffusion. There is however very high specific resistance (−1 MΩ cm2) that helpsreduce passive diffusion of ions into fiber cells. There are three isoforms of NaCKC
ATPase in the epithelium while there is only one in the fiber cells (Mosley et al.,1996). Based on immunoblotting there are almost equal number of NaCKC-ATPase molecules in the epithelium and fiber cells per mg membrane protein, how-ever for some unknown reasons the enzyme in the fiber cells is not highly active.
There are three carrier systems in the lens epithelium for the transport of basic,neutral and acidic amino acids (see Reddy, 2000 for references). Interestingly lensaccumulates very high concentrations of taurine (2-aminoethanesulfonic acid). Whilethe significance of the presence of high concentrations of taurine in the lens epi-thelium remains speculative, changes in its concentrations are indicative of thechanges in the permeability characteristics of the epithelium, particularly in the cat-aractous lens when the concentration of taurine decreases (Reddy et al., 1976). Pres-ence of carrier systems for the transport of taurine into the lens have beendemonstrated but not characterized at the molecular level. The taurine transporterin retinal pigment epithelium seems to be regulated by calmodulin indicating thatCa2C levels may thus determine the activity of the taurine transporter (Ramamoor-thy et al., 1994). It is also possible that one of the functions of having taurine athigh concentrations may be regulation of the intracellular Ca2C levels (Huxtable,1992). Ocular lens maintains a large gradient of Ca2+ and the lens epithelium playsan important role in maintaining these levels (Jacob, 1983; Duncan and Jacob,1984). Free Ca2C in the cells can either be increased by release from stored Ca2C

544 Bhat
complexes or in its absence, by transport of Ca2C from extracellular space. It isbelieved that store-activated Ca2Centry is the major route of Ca2C influx into cells.The presence of electrical current that flows across the plasma membrane followingthis influx has been demonstrated and the existence of a ubiquitously distributedstore-operated Ca2C channel has been predicted (Parekh and Penner, 1997). Thesehave not yet been studied in the ocular lens epithelium. Interestingly, lens containshigh concentrations of myo-inositol (see Reddy, 2000). A NaCmyo-inositol con-transporter gene has been characterized in the bovine lens epithelium (Zhou andCammarata, 1997).
Human lens epithelial cells accumulate large amounts of polyol when exposedto high galactose in culture (Lin et al., 1990). Aqueous humor contains glucose atthe same levels as in the blood (Chylack, 1971). It was observed very clearly thatglucose uptake occurs in the lenses even after the removal of the capsule and theepithelium (Giles and Harris, 1959). Also, the capacity to transport glucose is com-parable at both the anterior as well as posterior faces of the lens (Kern and Ho,1973a). Oxidative phosphorylation consumed glucose transported via glucose trans-porters (GLUT) into the epithelium (Goodenough et al., 1980). In the rat lensGLUT1 is predominantly expressed in the epithelium and GLUT3 in the corticalfiber cells (Merriam-Smith et al., 1999). It is of interest to note that GLUT3, whichhas a lower Km for glucose than GLUT1 is widely expressed in cells exposed toeasily accessible levels of glucose. Delineating the relationship of the expression ofdifferent isoforms of GLUT transporters and sugar uptake by epithelium would beimportant in any attempt of regulating the accumulation of glucose in the diabeticlens.
The effect of glucocorticoids on the physiology of the lens epithelium is ofpractical significance since administration of steroid hormones is part of the thera-peutic schemes in treating glaucoma and other related inflammatory pathologies ofthe eye. Cortisol and aldosterone can alter the physiological equilibrium within thelens by altering the ionic composition of the lens and aqueous humor (Starka et al.,1986). Appreciable expression of glucocorticoid and mineralocorticoid receptors andtheir 11-β-hydroxysteroid dehydrogenases that regulate their activity at the pre-receptor level in the lens epithelium was recently reported (Stokes et al., 2000).
Lens epithelium is also the major site of detoxification and defense against oxi-dative insults (Reddy et al., 1980; Giblin, 2000). It contains active preteosome com-plexes (Andersson et al., 1998) and an active ubiquitin-dependent proteolysispathway (Shang et al., 1997) that degrades oxidized proteins such as α -crystallins(Huang et al., 1995). The lens epithelium contains the highest concentration ofglutathione (GSH) and NADPH (Giblin et al., 1981). A concentration above 20 nMhas been reported. Glutathione is synthesized within the epithelium (Reddy et al.,1966; Kern and Ho, 1973b). Presence of a sodium independent glutathione trans-porter has also been demonstrated in the lens (Kannan et al., 1995). The lens epi-thelium contains an active glutathione redox cycle (Glutathione reductase. NADPH,hexose monophosphate shunt) that keeps GSSG reduced to GSH ensuring detoxi-fication of oxidizing molecules. Epithelium is able to detoxify physiological concen-trations of H2O2 enzymatically involving glutathione reductase, glutathioneperoxidase and the hexose monophosphate shunt (see review by Giblin, 2000). NaC

Ocular Lens Epithelium 545
KC-ATPase, the main transport pump, is inhibited by peroxide thus inhibiting activecation transport (Garner et al., 1983; Giblin et al., 1987). It was discovered veryearly that peroxide is formed via the light-catalyzed oxidation of ascorbic acid inthe aqueous humor (Pirie, 1965). Since reactive oxygen species can lead to mutationsand carcinogenesis (Fridovich, 1986), it is to be expected that the lens epitheliummay have evolved an elaborate defense against oxidative (Hightower et al., 1994).
ELECTRICAL PROPERTIES
Direct electrical measurements have shown that the resting potential of the lens(−69.6 mV for the rat lens) (Lucas et al., 1987), is mostly because of the KC gradient.Cells control their volume by maintaining a negative resting potential. This isachieved by keeping the concentration of intracellular NaC low and KC high. Theepithelial cells maintain their volume by restricting the inflow of the anions. ActiveNaCKC transport, keeps the NaCKC ratio low. Both NaCKC ATPase and anumber of KC channels are synthesized in the epithelial cells. Because there is com-paratively very little functional NaCKC ATPase and none or few KC channels inthe fiber cells the resting membrane potential of the lens must be considered to beprimarily due to activities in the epithelial layer andor superficial cortical layers offiber cells. NaCKC pump exchanges three NaC ions for two KC ions thus generat-ing an outward current. Regional differences in the predominance of one or theother isoform in the lens epithelium have been reported (Garner and Horwitz, 1994).α 3 isoform is the predominant protein of the central epithelium, while α 1 isoformis more predominant in cells nearer the equator. Functionally different isoformsof NaCKC ATPase could have specific physiological consequences but regionaldifferences in the expression and activity of this pump have not been clearly corre-lated. A number of potassium channels have been cloned (Chandy and Gutman,1995). In the lens presence of inward rectifiers (they conduct inward KC currentmuch faster than they do the outward current), outward rectifiers (delayed rectifiers)and calcium-activated large-conductance KC channels have been found (Mathias etal., 1997; Rae and Shephard, 1998; Rae and Shephard, 2000). A complete molecularcatalog of all the KC channels that exist in the lens epithelium will allow a regionallocalization of specific types and their functional import.
Ocular lens has been called a syncytium (Goodenough, 1992; Mathias et al.,1997) based on the fact that the entire lens is connected, the epithelial cells to epi-thelial cells, epithelial cells to fiber cells and fiber cells to fiber cells. The way lens isplaced, it is natural to think that the epithelium becomes the link that transportsnutrients between the aqueous and the vitreous humor compartments. However, itwas recognized very early that the electric current flux at the anterior as well as atthe posterior poles was directed inward into the lens. This current followed outwarddirection only at the equatorial poles (Robinson and Patterson, 1983). This suggeststhat regionally operated circulatory systems are followed by cellular fluids (seereview by Mathias et al., 1997). The inward transport being intercellular, and theoutward intracellular. Whether there exists a molecularbiochemical link betweenpumps, channels and gap junctions and these regional electrical conductances (circu-latory systems) remains to be investigated. However, if the lens is treated as a single

546 Bhat
Fig. 4. A hypothetical schematic of the flow of currentfluids in the ocular lens. Based on Mathias et al.(1997) and Zamphigi et al. (2000). The thick dotted arrows from the anterior and the posterior followthe orientation of the apical and the basal surfaces. It is presumed that the orientation of the cellsfibersat the equatorial region is such that facilitates the flow as shown (thick dotted arrows). The active NaCKC transport is indicated by double headed open arrows on the anterior. Small dotted arrows in theposterior indicate passive and facilitated diffusion. Thin arrows at the equator suggest some activity inthe direction of the arrow before the fiber cell differentiation is complete.
folded epithelium it is easier to relate the direction of the current (and therefore thetransport of the fluids) to the anatomy of the system (Fig. 4). Significantly then, thedirection of the inward current from the anterior and the posterior pole follows theorientation of the apical and the basal surfaces of this epithelium (Zampighi, 2000).Importantly, fluid transport in cultured lens epithelia and organ cultured rabbit lenswas recently shown to follow the path from basal to apical surfaces. This transportwas blocked by the inhibitors of KC channels and aquaporins (Fischbarg et al.,1999). It is clear that transport within the fiber cells in the posterior of the lens andin the epithelial cell layer in the anterior of the lens is dictated by different morpho-logical and molecular parameters. Physically, the fiber cells present a much largersurface than epithelial cells thus facilitating diffusion and molecularly, by the pres-ence of gap junctions predominantly represented by MIP26 (major intrinsic mem-brane protein 26 kD, also known as aquaporin 0) (Gorin et al., 1984; Hamann etal., 1998). Although existence of coupling between the epithelial cells and the under-lying fibers has been questioned (Brown et al., 1990; Bassnett et al., 1994) thereseems to be no sound scientific evidence that suggests about 10% of the epithelialcells and apical surfaces of the fibers are dye coupled. This coupling is as good asbetween the epithelial cells themselves (Rae et al., 1996). The proximity of the apicalsurfaces of the fiber cells and the epithelial cells (the apical interface) allows thiscommunication (Fig. 5). Using the uptake of the lipophilic cation as an indicator ofthe resting potential in the rat lens, Cheng et al. (2000) suggested the presence oftwo compartments, one that is fast and highly sensitive to the concentration ofexternal KC, while the other is less sensitive to external KC concentrations and slow.Immunocytochemical localization of the channel proteins also suggests the existence
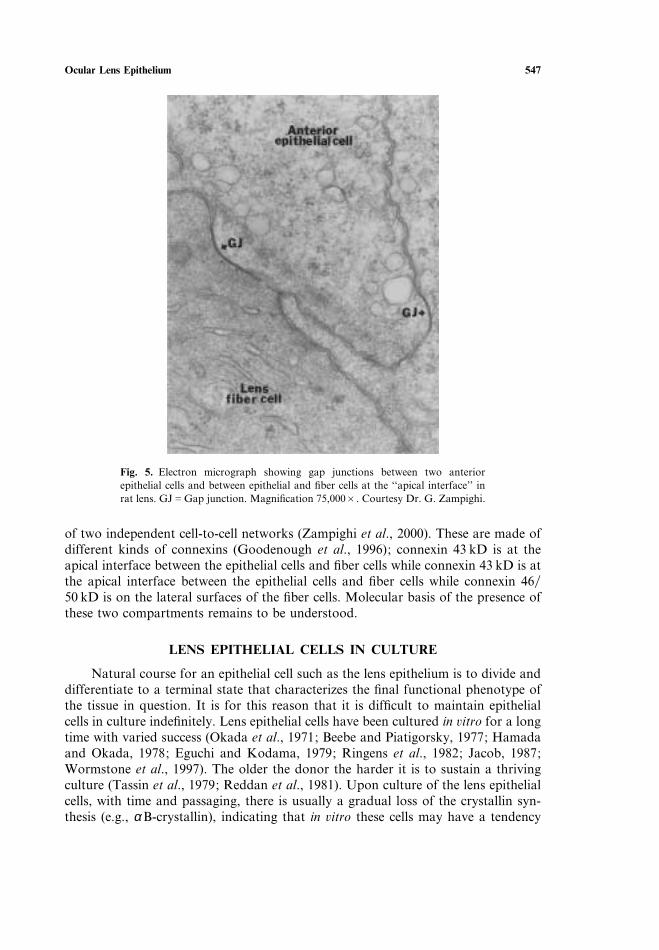
Ocular Lens Epithelium 547
Fig. 5. Electron micrograph showing gap junctions between two anteriorepithelial cells and between epithelial and fiber cells at the ‘‘apical interface’’ inrat lens. GJGGap junction. Magnification 75,000B. Courtesy Dr. G. Zampighi.
of two independent cell-to-cell networks (Zampighi et al., 2000). These are made ofdifferent kinds of connexins (Goodenough et al., 1996); connexin 43 kD is at theapical interface between the epithelial cells and fiber cells while connexin 43 kD is atthe apical interface between the epithelial cells and fiber cells while connexin 4650 kD is on the lateral surfaces of the fiber cells. Molecular basis of the presence ofthese two compartments remains to be understood.
LENS EPITHELIAL CELLS IN CULTURE
Natural course for an epithelial cell such as the lens epithelium is to divide anddifferentiate to a terminal state that characterizes the final functional phenotype ofthe tissue in question. It is for this reason that it is difficult to maintain epithelialcells in culture indefinitely. Lens epithelial cells have been cultured in ûitro for a longtime with varied success (Okada et al., 1971; Beebe and Piatigorsky, 1977; Hamadaand Okada, 1978; Eguchi and Kodama, 1979; Ringens et al., 1982; Jacob, 1987;Wormstone et al., 1997). The older the donor the harder it is to sustain a thrivingculture (Tassin et al., 1979; Reddan et al., 1981). Upon culture of the lens epithelialcells, with time and passaging, there is usually a gradual loss of the crystallin syn-thesis (e.g., α B-crystallin), indicating that in ûitro these cells may have a tendency

548 Bhat
to stray from the pleuripotent state that must otherwise lead to fiber cell differen-tiation (van Venrooij et al., 1974; Vermorken et al., 1977; Hamada et al., 1979;Ringens et al., 1982; Simonneau et al., 1983; Ibaraki et al., 1996). However, whenhuman fetal lens epithelial cells are cultured with deliberate slow passaging they canbe maintained fairly well in culture (see Fig. 2B). Not only do these cultures showa sustained synthesis of α B-crystallin, but a progressive increase in the expressionof βB2-crystallin a marker indicative of fiber cell differentiation is also seen (Nagi-neni and Bhat, 1988; Nagineni and Bhat, 1989a). Arita et al. (1990) used nonproteinbinding membranes as substrates for the culture of human lens epithelial cells andshowed enhanced fiber cell differentiation as assessed by the appearance of ‘‘lentoid’’bodies. This technique has allowed the culture of epithelial cells from older donorlenses. Lens epithelial cells secrete their own capsular materials in culture (Grant etal., 1972; Arita et al., 1993). When cultured on the lens capsule, lens epithelial cellsretain their ability to synthesize α B-crystallin (Vermorken et al., 1977).
In order to overcome the naturally restricted growth of native epithelial cells inûitro immortalized cell lines have been generated from primary lens epithelial cellcultures, (Bloemendal et al., 1980). Two cell lines were generated recently fromhuman lens epithelial cells: One by transformation of the primary cultures with avirus containing the simian virus-40 transforming gene (SV-40 T antigen) (Andleyet al., 1994) and the other by direct transfection with SV-40 large T-antigen gene(within a plasmid) (Ibaraki et al., 1998). The later cell line seems to retain theexpression of α A-crystallin and βB2-crystallin. About fifty other lens epithelial celllines have been generated and investigated for the presence of different proteins(markers?) (Sax et al., 1995; Krausz et al., 1996).
Do lens epithelial cells have a specific molecular marker? At this time we donot know of any such markers, although different investigators have at times usedα -crystallins as lens cell markers, the fact is that a number of other non-lens cells inculture express these proteins (Nagineni and Bhat, 1989b), albeit at very low levels.This brings us to the unanswered questions: Does the loss of crystallin synthesisindicate loss of lens-specific characteristics? How is a lens epithelial cell, which doesnot express presumed markers (like α -crystallins) different from any other epithelialcell? If α -crystallins are indicators of lens characteristics, how much and which α -crystallin defines that characteristic? In the absence of any knowledge as to whatdefines a lens-specific characteristic (morphologic or molecular), in an epithelial cell,these are different questions to answer.
The next question is how representative are the cultured cells of the physiologyof the native epithelium? The answer to this question depends on the origin andphysiological characteristics of each cell line. However established transformed cells(spontaneously or otherwise) must be used with discretion, particularly in the studiesof cell growth, differentiation, aging and apoptosis. The SV-40 transformed cells ofthe hamster lens epithelial cells produce tumors in homologous hosts (Albert et al.,1969). Long-term cultures of bovine epithelial cells produce tumors upon injectioninto nude mice (Courtois et al., 1978). Transformed cell lines may present more thanone stable morphological and biochemical phenotype (Bloemendal et al., 1997).Most of the work with lens epithelial cells thus far has been done with cell cultures

Ocular Lens Epithelium 549
(lines) that have not been deliberately immortalized with the introduction of trans-forming (immortalizing) genes. A large number of experiments have utilized thechicken embryonic lens epithelium system for studying cell-culture control and dif-ferentiation (Piatigorsky, 1981; Zelenka et al., 1997). Rat explant cultures have beenused for studying differentiation of fiber cells from epithelial cells in ûitro (McAvovyand Fernon, 1984). Many investigations for metabolic studies have been done withbovine and rabbit lens epithelial cells.
The study of lens-specific metabolism or molecular processes must always bedone with the knowledge that the very act of in ûitro culture may have changed themolecular makeup of the cells. A few of these examples are discussed here. In theprimary cultures produced from the bovine lenses, the mitotically inactive cells showcalcium dependent spreading followed by DNA synthesis. These cells are very sensi-tive to C6-substituted purine derivatives that inhibit cell flattening as well as growth,but this sensitivity to purine derivatives is lost as the cells are sub-cultured (Glaesseret al., 1979). Culturing of the lens epithelial cells has been reported to result insignificant changes to energy metabolism of the cells (Piper et al., 1990). A calciumdependent ATPase activity present on native bovine epithelial cells is lost upon cul-ture. (Bergner and Glaesser, 1979) A recent comparative study of the muscarinicreceptors in the native lens epithelium vs. the transformed cell line, HLE-B3 (Andleyet al., 1994) is very informative. Both these cells in culture express muscarinic recep-tors, however the pattern of the subtypes that we expressed are very different. Nativelens epithelial cells almost exclusively express the MI subtype of the muscarinicreceptors. The transformed cell line HLE-B3 expresses M3 subtypes predominantly(Collison et al., 2000). Recently, a cell line developed from the human fetal lensepithelium (HFL124) with extended life span (without transformation, spontaneousor deliberate) has been shown to maintain the expression of c-met receptorexpression similar to that in the native epithelium (Wormstone et al., 2000). Investi-gations on cultured cells have contributed extensively to our understanding of differ-ent biological processes and shall continue to be utilized for such defined purposes.
Lens epithelial cells have been studied to culture for a number of metabolicand growth studies (Harding and Crabbe, 1984; McAvovy and Chamberlain, 1990;Spector et al., 2000). Among the growth factors one that stands out is FGF. FGFat concentrations of 0.15 ngml supports proliferation while a concentration of40 ngml supports differentiation in rat lens explant culture system (McAvovy andChamberlain, 1989). Effect of FGF on the lens epithelial cells have also been demon-strated in transgenic animals where recombinant FGF1 controlled by fiber cell-specific α A-crystallin promoter, was synthesized and secreted from fiber cells. Adja-cent epithelial cells were stimulated to form fiber cells (Robinson et al., 1995). Atthis time no epithelial cell-specific promoter has been characterized that would directexpression to lens epithelial cells only. It is interesting that lens epithelial cells retaintissue-specificity when assayed for reporter gene assays with different crystallin pro-moters (Kondoh et al., 1983; Sax and Piatigorsky, 1994).
Another growth factor worthy of attention in the context of lens epithelialbiology is TGF-β. Exposure of intact lenses in ûitro to TGF-β leads to the appear-ance of spindle-shaped cells that contain α -smooth muscle actin (Hales et al., 1994).However, in ûiûo, intravitreal injection of TGF-β, results in the phenotype of

550 Bhat
nucleated fiber cells in the cortex and the migration of the cells posteriorly (Hales etal., 1999) indicating disruption in fiber cell differentiation. These data suggest thatTGF-β initiates an entirely different developmental program, namely, the transform-ation of the epithelial cell to the mesenchymal cell type. Mesenchymal transform-ation has been reported to be associated with anterior as well as posteriorsubcapsular cataracts (see Marcantonio et al., 2000). Recently it was suggested thatexpression of α -smooth muscle actin, an indicator of mesenchymal transformation,is a common feature of lens epithelial cells in culture (Nagamoto et al., 2000). Epi-thelialmesenchymal transformation has developmental antecedents (Hay and Zuk,1995) and points to the tenuous nature of the pleuripotency of the lens epithelium,which when compromised may lead to secondary opacifications after cataract sur-geries. This interesting aspect of lens epithelial biology deserves more attention.
PROGRAMMED CELL DEATH AND THE LENS EPITHELIUM
The interest in the study of programmed cell death in the lens epithelium wasgenerated recently by investigators probing the role of epithelium in cataractogenesis(Li and Spector, 1996). These studies are based on the hypothesis that integrity of thelens epithelium is essential for the normal functioning of the lens and that a decreasein the cell number of the epithelium may lead to changes in homoeostasis that may inturn lead to cataractogenesis. The role that apoptosis plays in the development andtissue morphogenesis is well established (Steller, 1995). Apoptosis or cell death hasbeen morphologically documented in the very early stages of the lens vesicle formationduring development of the eye (Garcia-Porrero et al., 1984). This ‘‘morphogenetic’’cell death (Gluksmann, 1951; Ishizaki et al., 1993) is possibly related to the loss of thelens stalk, considering that at this time the lens vesicle is preparing to pinch off thesurface ectoderm. It is not clear whether apoptosis is essential to lens differentiationand development after the formation of the lens vesicle? There is no experimental dataavailable to suggest that ‘‘apoptosis’’ is a major factor either in ‘‘primary fiber cell’’ or‘‘secondary fiber cell’’ differentiation. This does not negate the role that apoptosismay come to play in the final maturation of the differentiated fiber cell (Applebyand Modak, 1977; Wride and Sanders, 1998). It is possible that during primary lensfiber cell formation not all cells differentiate into fibers and a subset of posteriorepithelial cells may go through apoptosis. Another possible scenario would have animportant bearing on the phenotype of transparency is the migration of the differen-tiating cells to the posterior capsule. Under normal, healthy conditions cells that donot stop at the equator may be triggered to die before they reach the posteriorcapsule, which if they did, would nullify the phenotype of transparency by contribu-ting to the development of the pathology known as ‘‘posterior subcapsular opaci-fication’’. The fact that apoptosis may in some form attend fiber cell differentiationis indicated by the reported involvement of caspases (cysteine-asparate proteasesknown to initiate the cascade that leads to apoptosis) in the differentiation of thefiber cells in the rat lens (Ishizaki et al., 1998).
It remains to be established if indeed there is a basal rate of apoptosis in thenormal adult lens epithelium. Ishizaki et al. (1993) found evidence of cell death(phagocytosis) in the adult rat lens epithelium in the region of the ‘‘anterior suture’’

Ocular Lens Epithelium 551
(the area where the anterior ends of the lens fibers converge). The reasons as to whythis happens remain unclear. These investigators reported ∼ 5% pyknotic cells in theyoung rats and about 1% in older rats. Cell division in the epithelium decreases withage and that may have to do with the species-specific (Kuszak, 1995) and that thereis a (positional) relationship between sutures and the apoptotic cells in the epi-thelium then it must be assumed that contribution from apoptosis to the lens devel-opmentphysiology may be species-specific. As to why there should be anassociation between the location of sutures and apoptosis remains unexplored.Whether apoptosis has a definitive role in the maintenance of a specific number ofepithelial cells in the lens is debatable. Nonetheless, a role for apoptosis in regulatingthe size of the lens by controlling the number of cells that reach terminal differen-tiation into fiber cells remains a possibility.
ULTRAVIOLET RADIATION (UVR) AND THE LENS EPITHELIUM
The single layer of epithelium is the first physical and cellular (biological)defense against electromagnetic radiation in the ocular lens (Dillon, 1991). Some ofthe direct effects of UVR exposure on cultured cells have been reviewed in detail(Hightower, 1995). The consequences of UVR exposure on the epithelium must beconsidered both in terms of mutagenic as well as cytotoxic effects (Worgul et al.,1989, 1991). UVR exposure results in unscheduled DNA synthesis and repair(Brenner and Grabner, 1982; Kleiman et al., 1990; Sidjanin et al., 1993). The actionspectrum for the rat lens in ûiûo shows that 305 nm, which reaches the lens muchmore than the 290 nm band (part of UVB, 280–315320 nm), is harmful to the ratlens. It is not possible to extrapolate directly from the rat lens to human condition,a composite of location, intensity of exposure, genetics, diet and age. However it isconceivable that the human lens epithelium accumulates insults due to UVR expo-sure in its genome over a period of time that are manifest in the aged lens (Merriamet al., 2000). There is of course an age-dependence of UVR damage to differentmolecular species including enzymes such as hexokinase, phosphofructokinase, isoc-itrate dehydrogenase and malate dehydrogenase (Reddy and Bhat, 1998). Loss ofhexokinase (Tung et al., 1988) would result in the inability of the lens to produceNADPH and downstream antioxidants. It is conceivable that proteins (such asNaCKC) ATPase, cytoskeletal elements, membrane proteins), which dependenton –SH function will be damaged by exposure to increased oxidants.
Ultraviolet radiation (300–320 nm) exposure leads to an alteration in the mor-phology of the epithelium. The morphological effects of UVR exposure encompassgeneral cell swelling including mitochondrial swelling, chromatin condensation andfragmentation of the nuclear DNA. One of the more significant early effects is the lossof positional identity of the epithelial cells. The epithelial cells from the bow regionmigrate to the posterior capsule. A molecular or biochemical basis of the loss of thispositional integrity has not been investigated. It has been suggested that a disturbancein calcium homeostasis interferes with the differentiation pathway of the lens epi-thelium into fiber cells, which leads to the migration of the bow region cells to theposterior capsule. Many investigations suggest compromise of the calcium homeo-stasis possibly starts in the epithelium and propagates to the rest of the lens. Levels

552 Bhat
of Ca2C-increase have been shown to be dose-dependent in rabbit lenses exposed toUVR. It is possible that this is because of the photo-oxidation of the Ca2C-ATPasethat contains approximately 30 thiol (SH) groups. In this context it is interesting tonote that Ca2C-ATPase activity increases with age (Brochman et al., 1989).
Different metabolites (Inoue et al., 2000) and drugs (Li et al., 1995b) have beenreported to initiate apoptosis in lens epithelial cells in culture. For the native lensepithelium, however, environmental factors such as ultraviolet radiation (UVR)become highly relevant in this context (Zigman, 2000) and Li and Spector (1996),irradiated whole rat lenses in culture with 8 Jsm2 of UVB (285–315 nm) for 60min. As expected the cells in the equatorial zone were the first to show signs ofapoptosis followed by the central epithelial cells. They reported that more than 50%of the cells were apoptotic by the 5th hour after irradiation and by 24 hr all theepithelial cells in the irradiated lenses had died. This seems to be an extreme case ofexposure to UVR that leads to 50% cell death, but it underscores the fact that it isthe cells in the bow region that are first effected followed by those in the centralepithelium. As expected the dividingdifferentiating epithelial cells are more suscep-tible to UVR damage than the resting central epithelial cells. Based on morphologi-cal grounds, in the intact animal, the equatorial regions may not be exposed to asmuch UV as the central epithelium. Recently, Michael et al. (1998) unilaterallyexposed Sprague–Dawley rat eyes to UVB (5 kJm2, 300 nm) for 15 min and thenfollowed the fate of the lens epithelium at various time-points. TUNEL (terminaldeoxynucleotidyl transferase–deoxyuridine triphosphate nick end labeling)-positivecells (indicative of cell-death) were detected only at the 24 hr time point and not at1 hr or 6 hr or 1 week after exposure. Although the type of UVR used (long, 340–400 nm vs. short 200–320 nm) may lead to temporal differences in the induction ofapoptosis, the fact that apoptotic cells and their phagocytosis by the neighboringcells were only detectable at 24 hr (not before or after that) is interesting. That thephagocytosis is completed quickly within hours is confirmed in an in ûitro study also(Shui et al., 2000). These observations suggest that apoptosis may occur discretelyin the lens epithelium and only in isolated cells that become susceptible. Such athesis connotes two interesting corollaries: (1) Any isolated apoptotic cell may bequickly eliminated by the surrounding cells(s) as a protective response against spreadof cell death. Importantly, therefore, it points to the existence of a potent mechanismthat strictly controls and inhibits cell death from spreading to the neighboring cells.Thus in order to detect any apoptotic cells after an insult, in the normal lens epi-thelium a large number of cells and time points may have to be analyzed. (2) Pres-ence or absence of apoptosis in the lens epithelium can be interpreted optimisticallyas a process that eliminates the ‘‘dysfunctional’’ cells to keep the rest of the epi-thelium healthy. A pathological state may precipitate when this ability to remove‘‘dysfunctional’’ cells is compromised, for example by aging or by exposure to harm-ful metabolites or environmental insults. The existence of a molecular mechanismthat keeps apoptosis discrete and rare in the lens epithelium remains to be identified.
APOPTOSIS AND CATARACT
It is clear that physical andor chemical insult may initiate apoptotic programin lens cells, but whether apoptosis contributes to cataractogenesis remains

Ocular Lens Epithelium 553
unresolved. Li et al. (1995a) reported a significant number of apoptotic cells in cap-sular epithelia surgical samples from cataractous patients in comparison to thoseobtained from normal donors. Haracopos et al. (1998) attributed presence of anyTUNEL-positive cells in casulotomy specimens most likely to be due to necrosiscaused by post-surgery handling of the specimen, thereby challenging the claim thatapoptosis may lead to cataractogenesis.
An interesting aspect of a relationship between cell density and apoptosisdeserves mention. Based on the ‘‘death by default’’ thesis (Raff et al., 1993; Ishizakiet al., 1993), a decrease in cell density with age could lead to paucity of the presumedsurvival factors that could in turn lead to further deterioration of the integrity ofthe lens epithelium by initiating apoptosis. But before such conclusions are reachedit needs to be established firmly if (a) there is a decrease in cell density with age orcataract and (b) this decrease in cell density is significant enough to trigger ‘‘celldeath’’. Based on the need for survival factors produced by neighboring cells in adense culture it will require large denuded areas to trigger ‘‘cell death’’ in the lensepithelium. While such denuded ‘‘black holes’’ have been recently described (Bala-ram et al., 2000), their origin is unknown and their relationship to cataracts is unpro-ven. Also, there remains the puzzle as to how the ‘‘black holes’’ are created in thefirst place?
CATARACT AND CANCER
Cataractogenesis (loss of transparency) is the culmination of biochemicalmol-ecular events that lead to the compromise of the structural integrity of the fiber cellsand their proteins. The question of whether the initiating event for the developmentof this pathology indeed lies in the molecularmetabolic compromise of the epithelialcell layer remains unsolved. Cataractous lenses and in particular the capsuleepi-thelia have morphological features, typical of aging cells such as altered hexagonalcellular arrays, changes in endoplasmic reticulum, Golgi apparatus and mitochond-ria (Straatsma et al., 1991). The lens epitheliumlens is not known to become neo-plastic except under experimental conditions when transforming genes areintroduced into the lens of transgenic animals (Mahon et al., 1987). The proliferativestatus of epithelial cells in the cataractous lens is unknown. It is not clear if the cellsin the epithelium of the cataractous lens divide at the same rate as those in thenormal lens. The maintenance of the pleuripotent state of the lens epithelium duringcataractogenesis (posterior subcapsular opacification) is in question. Does epithelialto mesenchymal transformation (discussed previously) represent an obligatory pro-cess attendant with cataractogenesis? Does increased length of telomeres in catar-actous epithelia (see below) suggest a relationship to increased proliferation?Harocopos et al. (1998) report that cataractous areas in the human lens are associ-ated with areas of higher cell density in the lens epithelium. The phenotype of thesecells with respect to their oncogenic or benign is uninvestigated. Mitotically activecells have been seen superimposed on the central epithelial cells in very advancedcataracts (Vasavada et al., 1991). Colitz et al. (1999) found ‘‘fibrous metaplasia’’ incanine hyper mature cataracts. Similarities between the causative agents in catar-actogenesis and carcinogenesis have been pointed out previously (see Bloemendal,

554 Bhat
1991). However, the thesis that a molecular compromise of the genetic integrity ofthe lens epithelium leads to a dysfunctional lens (cataract or otherwise) has actuallynever been established or investigated directly. Cataract as a problem of uncon-trolled proliferation has not been experimentally investigated. A general question asto why the normal ocular lenslens epithelium remains non-cancerous has alsoremained unexplored.
The remarkable ability of the cells in the germinative zone of the lens epitheliumto keep dividing (Harding et al., 1971) beckons the question if this is related to thepresence of telomerase in these cells. Telomerase is an enzyme complex of ribonuc-leoprotein that replenishes the shortening 3′-ends of chromosomal DNA ends (con-taining 5′-TTAGGG-3′ repeats) after each replication, thereby helping maintain thelength of the chromosome ends (telomeres) in the dividing cells. Telomerase ensuresminimal telomere length in dividing cells thus preventing cell senescence or aging.Telomerase is highly active in proliferating embryonic cells; stem cells and germcells, but is mostly absent in most normal adult somatic cells. The association oftelomerase with replicative ability is substantiated by the demonstration that cells inculture attain unlimited proliferative potential upon transfection with telomeraseexpressing vectors (Ferber, 1999; Bodnar et al., 1998; Vaziri et al., 1999).
The presence of telomerase in the lens epithelium raises interesting questionsabout the relationship of telomerase to proliferation. Quite interesting is the findingof increased telomere length in the cataractous epithelia as opposed to that in thenormal epithelium (Colitz et al., 1999). This study also reported that there was nodifferences in telomere lengths in relation to age. These findings deserve attentionbecause telomere length would be expected to show a gradual decrease similar tothat seen in culture, as the cells get closer to senescence with each passage. Interest-ingly all regions of the lens epithelium, the quiescent, the germinative and the differ-entiating cells, all had same amount of telomerase activity as assayed by TRAP-ELISA method that measures the length of the telomeres with polymerase chainreaction (Colitz et al., 1999). If the assay is quantitative it is a case of discordancebetween the amount of telomerase activity and cell division. Consistent, althoughnot very convincing is the observation that the exposure of the lens epithelium toradiation (including UVR) may physiologically predispose the cells to higher telo-merase activity as a defense against UVR-induced damage to the cell DNA (Kleimanet al., 1990). Lack of detectable telomerase activity, even in the limbal cells thatretain the proliferative potential in the cornea, a tissue also exposed to UVR (Eganet al., 1998) weakens this explanation. At this time it is not possible to ascertainif the increase of telomere length in the cataractous epithelial DNA is related tohyperproliferation and if hyperproliferation is the result or the precipitating eventin the pathogenesis of the loss of transparency?
CONCLUSION
Table 1 is a summary of the alien features of the lens epithelium. We have notconsidered a number of other interesting and important features of the lensepithelium. These include but are not restricted to ultrastructural details (Ireland etal., 1983; Rafferty et al., 1990; Kuszak, 1995) the basement membrane and its

Ocular Lens Epithelium 555
Table 1. Some Salient Features of the Lens Epithelium
Derived from surface ectodermCell density is higher in females than malesContains quiescent, non-dividing cells, dividing cells and differentiating cellsProduces cells that divide and differentiateContains cells that apoptosizeContains epithelial cell :epithelial cell junctionsContains epithelial cell :fiber cell junctions at the apical interfaceHas ability for phagocytosis and endocytosisProduces the basement membrane (collagen IV) that makes the lens capsuleTransports water, ions and nutrients into the lensPumps out NaC and transports in KC
Contains KC channelsHas a direct role in the maintenance of the resting potential of the ocular lensPolarized, with apical surface facing the lumen of the lens and the basal surface facing the outside world
(aqueous humor)Cells exposed to radiation (UVR)Synthesised glutathioneExpresses glutathione transportersContains high concentration of taurine
components that make the lens capsule (Sawhney, 1995; Menko et al., 1988), theprocess of endocytosis (Gorthy et al., 1971; Lo and Zhang, 1989) and its role intransport of macromolecules and the aquaporins (Wintour, 1997). In addition thereare a large number of gene products that are constantly being discovered, whichhave a role in the optimal functioning of this epithelium (Wen et al., 1995; Srivastavaet al., 1996; Padgaonkar et al., 1997; Andley et al., 1988; Dahm et al., 1999; Yan etal., 2000). One of the best known promoters of the lens genes is that of α A-crystallin,which directs expression specifically to the lens fiber cells (Sax and Piatigorsky,1994). It has been used to study effects on lens epithelium (rather indirectly) bytargeting genes to the fiber cells. There is a need to search for lens epithelial cellspecific genes and a characterization of their promoters so that gene products may betargeted to the lens epithelium specifically for functional studies and for therapeuticpossibilities.
A loss in the functional physiology in the lens epithelial cells with age may makethis epithelium susceptible to external (e.g., UVR) or internal physiological signals(e.g., H2O2). In the theory of the oxidative stress as the initiating event for catar-actogenesis of the lens, the initial and the primary target are the epithelial cells(Spector, 1995; Sasaki et al., 1998). This can be investigated by examining the pat-terns of gene expression in the epithelium. One of the important questions that needto be resolved is, does the lens accumulate DNA damage with age? Is it clear thatextensive analyses of the proliferative capabilities of the lens epithelium need to betaken up in order to understand any special aspects of the genetic control of cellproliferation in the lens epithelium. In this regard, lens epithelium, as a cell type thatseems to escape oncogenesis becomes a very relevant paradigm for understandingreplicative senescence as a tumor suppression strategy vs. replicative senescence as acausative mechanism for tissue dysfunction and age-related pathologies (Campisi,1999).

556 Bhat
It is only recently that investigators have started characterizing gene activity inthe epithelium under disease conditions (Kantrow et al., 1998; Sun et al., 2000). Thelens epithelium is exposed to a large number of physiological signals that comethrough aqueous humor. These signals must create differential responses at the gen-etic level that define the physiology of the ocular lens in relation to the generalmetabolism. One of the well-known signals is glucose. Glucose not only regulatesthe glucose transporters but other glucose responsive genes either directly orindirectly from metabolites derived from it (Vaulont et al., 2000). There may be aplethora of genes that are modulated in an aging lens epithelium, those patterns ofgene activity remain to be discovered. With rapidly advancing technology of geneexpression profiling (Vishwanath et al., 1999) this ignorance may be very short-livedand some of the peculiarities or lack thereof may be elaborated.
ACKNOWLEDGMENTS
I thank Dr. Guido Zampighi for providing the electron micrograph shown inFig. 5 and for his discussions about the anatomy of the lens. Thanks are due toSherry Yafai, Wendy Lam and Rick Gambino for their help with the manuscriptediting and illustrations. Thanks are due to Association for Research in Vision andOphthalmology, Rockville, MD and Academic Press, Orlando, FL for permissionto reproduce data presented in Fig. 2. Work in the laboratory of the author issupported by grants from NIH, S.P.B. is a recipient of the Research to PreventBlindness Wassermann Merit Award.
REFERENCES
Albert, D. M., Rabson, A. S., Grimes, P. A., and von Sallmann, L. (1969) Neoplastic transformation inûitro of hamster lens epithelium by Simian Virus 40, Science 164:1077–1078.
Andersson, M., Sjostrand, J., and Karlsson, J. (1998) Proteolytic cleavage of N-Succ-Leu-Leu-Val-Tyr-AMC by the proteosome in lens epithelium from clear and cataractous human lens. Exp. Eye Res.67:231–236.
Andley, U. P., Rhim, J. S., Chylack, L. T., and Fleming, T. P. (1994) Propagation and immortalizationof human lens epithelial cells in culture. Inûest. Ophthalmol. and Vis. Sci. 35:3094–3102.
Andley, U. P., Song, Z., Wawrousek, E. F., and Bassnett, S. (1998) The molecular chaperone α A-crys-tallin enhances lens epithelial cell growth and resistance to UVA stress. J. Biol. Chem. 273:31252–31261.
Appleby, D. W. and Modak, S. P. (1977) DNA degradation in terminally differentiating lens fiber cellsfrom chick embryos. Proc. Natl. Acad. Sci. U.S.A. 74:5579–5583.
Arita, T., Lin, L. R., Susan, S. R., and Reddy, V. N. (1990) Enhancement of differentiation of humanlens epithelium in tissue culture by changes in cell-substrate adhesion. Inûest. Ophthalmol. and Vis.Sci. 31:2395–2404.
Arita, T., Murata, Y., Lin, L. R., Tsuji, T., and Reddy, V. N. (1993) Synthesis of lens capsule in long-term culture of human lens epithelial cells. Inûest. Ophthalmol. and Vis. Sci. 34:355–362.
Balaram, M., Tung, W. H., Kuszak, J. R., Ayaki, M., Shinohara, T., and Chylack, Jr., L. T. (2000)Noncontact specular microscopy of human lens epithelium. Inûest. Ophthalmol. and Vis. Sci. 41:474–481.
Bassnett, S., Kuszak, J. R., Reinisch, L., Brown, H. G., and Beebe, D. C. (1994) Intercellular communi-cation between epithelial and fiber cells of the eye lens. J. Cell Sci. 107:799–811.

Ocular Lens Epithelium 557
Beebe, D. and Piatigorsky, J. (1977) The control of δ-crystallin gene expression during lens cell develop-ment: dissociation of cell elongation, cell division, δ-crystallin synthesis and δ-crystallin mRNA accu-mulation. Deû. Biol. 59:174–182.
Becker, B. and Cotlier, E. (1962) Distribution of rubidium-86 accumulated in the rabbit lens. Inûest.Ophthalmol. and Vis. Sci. 1:642–645.
Bergner, A. and Glaesser, D. (1979) Demonstration of a magnesium- and calcium-dependent ATPase onthe outer surface of bovine lens epithelial cells. Ophthalmic Res. 11:322–323.
Bloemendal, H. et al. (1980) SV40-transformed hamster lens epithelial cells: A novel system for theisolation of cytoskeletal messenger RNAs and their translation products. Exp. Eye Res. 31:513–525.
Bloemendal, H., Enzlin, J. H., Van Rijk, A. A., and Jansen, H. J. (1997) Biochemical differences betweenthree subcell-lines derived from SV40-transformed hamster lens cells. Exp. Eye Res. 64:1037–1041.
Bloemendal, H. (1991) Disorganization of membranes and abnormal intermediate filament assembly leadto cataract. Inûest. Ophthalmol. and Vis. Sci. 32:445–455.
Bodnar, A. G. et al. (1998) Extensions of life span by introduction of telomerase into normal humancells. Science 279:349–352.
Brochman, D., Delamere, N. A., and Paterson, C. A. (1989) Ca2C-ATPase activity in the human lens.Curr. Eye Res. 8:1049–1054.
Brenner, W. and Grabner, G. (1982) Unscheduled DNA repair in human lens epithelium following inûiûo and in ûitro UV irradiation. Ophthalmic Res. 14:160–167.
Brown, H. G., Passas, G. D., Ireland, M. E., and Kuszak, J. R. (1990) Ultrastructural, biochemical andimmunologic evidence of receptor-mediated endocytosis in the crystalline lens. Inûest. Ophthalmol.and Vis. Sci. 31:2579–2592.
Brown, N. A. P. and Bron, A. J. (1987) An estimate of the human cell size in ûiûo. Exp. Eye Res. 44:899–906.
Campisi, J. (1999) Replicative senescence and immortalization. In: The Molecular Basis of Cell Cycle andGrowth Control (Stein, G. S., Baserga, R., Giordano, A., and Denhardt, D. T., eds.), New York,Wiley–Liss, Inc, pp. 348–373.
Chandy, K. G. and Gutman, G. A. (1995) Voltage-gated potassium channel genes. In: Ligand andVoltage-gated Ion Channels (Norh, N. A., ed.), CRC, Boca Raton, FL, pp. 1–71.
Cheng, Q., Lichtstein, D., Russel, P., and Zigle, S. (2000) Use of lipophilic cation to monitor electricalmembrane potential in the intact rat lens. Inûest. Ophthalmol. and Vis. Sci. 41:482–487.
Chylack, L. T. (1971) Control of glycolysis in the lens. Exp. Eye Res. 11:280–293.Colitz, C. M. H., Davidson, M. G., and McGahan, M. C. (1999) Telomerase activity in lens epithelial
cells of normal and cataractous lenses. Exp. Eye Res. 69:641–649.Colitz, C. M., Malarkey, D., Dykstra, M. J., McGahan, M. C., and Davidson, M. G. (2000) Histologic
and immunohistochemical characterization of lens capsular plaques in dogs with cataracts. Am. J.Vet. Res. 61:139–143.
Collison, D. J., Coleman, R. A., James, R. S., Carey, J., and Duncan, G. (2000) Characterization ofMuscarinic receptors in human lens cells by pharmacologic and molecular techniques. Inûest.Ophthalmol. and Vis. Sci. 41:2633–2641.
Coulombre, J. L. and Coulombre, A. J. (1963) Lens development: fiber elongation and lens orientation.Science 142:1489–1490.
Courtois, Y., Simonneau, L., Tassin, J., Laurent, M. V., and Malaise, E. (1978) Spontaneous transform-ation of bovine lens epithelial cells. Differentiation 10:23–30.
Davidson, M. G., Harned, J., Grimes, A. M., Duncan, G., Wirmstone, I. M., and McGahan, M. C.(1998) Transferrin in after-cataract and as a survival factor for lens epithelium. Exp. Eye Res. 66:207–215.
Dahm, R., Marle, J. V., Prescott, A. R., and Quinlan, R. A. (1999) Gap junctions containing α 8-connexin(MP70) in the adult mammalian lens epithelium suggests a reevaluation of its role in the lens. Exp.Eye Res. 69:45–56.
Dillon, J. (1991) The photophysics and photobiology of the eye. J. Photophys. Photobiol. 10:23–40.Duncan, G. and Jacob, T. J. C. (1984) Influence of external calcium and glucose on internal total and
ionized calcium in rat lens. J. Physiol. 357:485–493.Egan, C. A., Savre-Train, I., Shay, J. W., Wilson, S. E., and Bourne, W. M. (1998) Analysis of telomere
lengths in human corneal endothelial cells from donors of different ages. Inûest. Ophthalmol. and Vis.Sci. 39:648–653.

558 Bhat
Eguchi, G. and Kodama, R. (1979) A study of human senile cataract: growth and differentiation of lensepithelial cell in in ûitro cell culture. Ophthalmic Res. 11:308–315.
Fagerholm, P. P. and Philpsin, B. T. (1981) Human lens epithelium in normal and cataractous lenses.Inûest. Ophthalmol. and Vis. Sci. 21:408–414.
Ferber, D. (1999) Immortalized cells seem cancer-free so far. Science 283:154–155.Fischbarg, J. et al. (1999) Transport of fluid by lens epithelium. Am. J. Physiol. 276:C548–557.Francois, J. and Rabaey, M. (1951) Examination of the lens by phase-contrast microscopy. Br. J.
Ophthalmol. 35:352–355.Fridovich, I. (1986) Biological effects of superoxide radical. Arch. Biochem. Biophys. 247:1–11.Garcia-Porrero, J. A., Colvee, E., and Ojeda, J. L. (1984) The mechanism of cell death and phagocytosis
in the early chick lens morphogenesis: a scanning electron microscopy and cytochemical approach.Anat. Rec. 208:123–136.
Garner, W., Garner, M., and Spector, A. (1983) H2O2-induced uncoupling of bovine lens NaK ATPase.Proc. Natl. Acad. Sci. U.S.A. 80:2044–2048.
Garner, M. and Horwitz, J. (1994) Catalytic subunit isoforms of mammalian lens NaK ATPase. Curr.Eye Res. 13:65–77.
Giblin, F. (2000) Glutathione. A vital lens antioxidant. J. Ocular Pharmacol. Therapeutics 16:121–135.Giblin, F., McReady, J., Schrimscher, L., and Reddy, V. (1987) Peroxide-induced effects on lens cation
transport following inhibition of glutathione reductase activity in ûitro. Exp. Eye Res. 45:77–91.Giblin, F. J., Nies, D. E., and Reddy, V. N. (1981) Stimulation of the hexose monophosphate shunt in
rabbit lens in response to oxidation of glutathione. Exp. Eye Res. 33:289–298.Giles, K. M. and Harris, J. E. (1959) The accumulation of 14C from uniformly labeled glucose by the
normal and diabetic rabbit lens. Am. J. Ophthalmol. 48:508–551.Glaesser, D., Rattke, W., and Iwig, M. (1979) Bovine lens epithelium: A suitable model for studying
growth control mechanisms. C6-substituted purines inhibit cell flattening and growth stimulation ofG0 cells. Exp. Cell Res. 122:281–292.
Glucksmann, A. (1951) Cell death in normal vertebrate ontogeny. Biol. Reû. 26:59–86.Goodenough, D. A., Dick, J. S. B., and Lyons, J. E. (1980) Lens metabolic cooperation: a study
of mouse lens transport and permeability visualized with freeze-substitution autoradiography andelectron microscopy. J. Cell Biol. 86:576–689.
Goodenough, D. A., Goliger, J. A., and Paul, D. L. (1996) Connexins, connexons and intercellularcommunication. Ann. Reûs. Biochem. 65:475–502.
Goodenough, D. A. (1992) The crystalline lens. A system networked by gap junctional intercellularcommunication. Semin. Cell Biol. 3:49–58.
Gorin, M. B., Yancey, S. B., Cline, J., Revel, J.-P., and Horwitz, J. (1984) The major intrinsic protein(MIP) of the bovine lens fiber membrane: characterization and structure based on cDNA cloning.Cell 39:49–59.
Gyorothy, W. C., Snavely, M. R., and Gerrong, N. D. (1971) Some aspects of transport and digestionin the lens of the normal young adult rat. Exp. Eye Res. 12:112–119.
Grainger, R. M. (1992) Embryonic lens induction: shedding light on vertebrate tissue determination.Trends Genet. 8:349–355.
Grant, M. E., Kefalides, N. A., and Prockop, D. J. (1972) The biosynthesis of basement membranecollagen in embryonic chick lens. 1. Delay between the synthesis of polypeptide chains and thesecretion of collagen by matrix-free cells. J. Biol. Chem. 247:3539–3544.
Guggenmoos-Holtzmann, I., Engel, B., Henke, V., and Naumann, G. O. H. (1989) Cell density of humanlens epithelium in women higher than in men. Inûest. Ophthalmol. and Vis. Sci. 30:330–332.
Hales, A. M., Schultz, M. W., Chamberlain, C. G., and McAvoy, J. W. (1994) TGF-β1 induces lens-cells to accumulate a smooth muscle actin, a marker for subcapsular cataracts. Curr. Eye Res. 13:885–890.
Hales, A. M., Chamberlain, C. G., Dreber, B., and McAvoy, J. W. (1990) Intravitreal injection ofTGF-b induces cataracts in rats. Inûest. Ophthalmol. and Vis. Sci. 40:3231–3236.
Hamada, Y. and Okada, T. S. (1978) In ûitro differentiation of cells of the lens epithelium of humanfetus. Exp. Eye Res. 26:91–97.
Hamada, Y., Watanabe, K., Aoyama, H., and Okada, T. S. (1979) Differentiation and dedifferentiationof rat lens epithelial cells in short- and long-term cultures. Deû. Growth Differ. 21:205–220.

Ocular Lens Epithelium 559
Hamann, S. et al. (1998) Aquaporins in complex tissues: distribution of aquaporins 1–5 in human andrat eye. Am. J. Physiol. 274:C1332–1345.
Harding, C. V., Reddan, J. R., Unakar, N. J., and Bagchi, M. (1971) The control of cell division in theocular lens. Int. Reû. Cytol. 31:215–230.
Harding, J. J. and Crabbe, J. C. (1984) The lens: development, proteins, metabolism and cataract. In:The Eye (Davson, H., ed.), New York, Academic Press, pp. 207–492.
Harocopos, G. J., Alvares, K. M., Kolker, A. E., and Beebe, D. C. (1998) Human age-related cataractand lens epithelial cell death. Inûest. Ophthalmol. and Vis. Sci. 39:2696–2706.
Harris, J. E. and Becker, B. (1965) Cation transport of the lens. Inûest. Ophthalmol. and Vis. Sci. 4:709–722.
Hay, E. D. and Zuk, A. (1995) Transformations between epithelium and mesenchyme: Normal, patho-logical and experimentally induced. Am. J. Kidney Disease 26:678–690.
Hightower, K. R. and McReady, J. P. (1991) Effect of selenite on epithelium of cultured rabbit lens.Inûest. Ophthalmol. and Vis. Sci. 32:406–409.
Hightower, K. R., Reddan, J. R., McReady, J. P., and Dziedzic, D. C. (1994) Lens epithelium: a primarytarget of UVG irradiation. Exp. Eye Res. 59:557–564.
Hightower, K. R. (1995) The role of the lens epithelium in development of UV cataract. Curr. Eye Res.14:71–78.
Huang, L. L., Shang, F., Novell Jr., T. R., and Taylor, A. (1995) Degradation of differential oxidizedalpha-crystallins in bovine lens epithelial cells. Exp. Eye Res. 61:45–54.
Huxtable, R. J. (1992) Physiological actions of taurine. Physiol. Reû. 72:101–163.Ibaraki, N., Chen, S. C., Lin, L. R., Okamoto, H., Pipas, J. M., and Reddy, V. N. (1998) Human lens
epithelial cell line. Exp. Eye Res. 67:577–585.Ibaraki, N., Lin, L. R., and Reddy, V. N. (1996) A study of growth factor receptors in human lens
epithelial cells and their relationship to fiber differentiation. Exp. Eye Res. 63:683–692.Inoue, K., Kubota, S., Tsuru, T., Araie, M., and Seyama, Y. (2000) Cholesterol induces apoptosis of
corneal endothelial and lens epithelial cells. Inûest. Ophthalmol. and Vis. Sci. 41:991–997.Ireland, M., Lieska, N., and Maisel, H. (1983) Lens actin: purification and localization. Exp. Eye Res.
37:393–408.Ishizaki, Y., Jacobson, M. D., and Raff, M. (1998) A role for caspases in lens filter differentiation. J.
Cell Biol. 140:153–158.Ishizaki, Y., Voyvodic, J. T., Burne, J. F., and Raff, M. C. (1993) Control of lens epithelial cell survival.
J. Cell Biol. 121:899–908.Jacob, T. J. C. (1983) A direct measurement of intracellular free calcium within the lens. Exp. Eye Res.
36:451–453.Jacob, T. J. C. (1987) Human lens epithelial cells in culture: A quantitative evaluation of growth rate
and proliferative capacity. Exp. Eye Res. 45:93–104.Jacob, A. G. and Sater, A. K. (1988) Features of embryonic induction. Deûelopment 104:341–359.Jean, D., Ewan, K., and Gruss, P. (1998) Molecular regulators involved in vertebrate eye development.
Mechanisms of Deûelopment 76:3–18.Kannan, R., Yi, J. R., Zlokovic, B. V., and Kaplowitz, N. (1995) Molecular characterization of a reduced
glutathione transporter in the lens. Inûest. Ophthalmol. and Vis. Sci. 36:1785–1792.Kantrow, M. et al. (1998) Differential display detects altered gene expression between cataractous and
normal human lenses. Inûest. Ophthalmol. and Vis. Sci. 39:2344–2354.Karim, A. J. A., Jacob, T. J., and Thompson, G. M. (1987) Cell density, morphology and mitotic index
in normal and cataractous lenses. Exp. Eye Res. 45:865–874.Kern, H. L. and Ho, C. K. (1973a) Localization and specificity of the transport system for sugars in the
calf lens. Exp. Eye Res. 15:751–765.Kern, H. L. and Ho, C. K. (1973b) Transport of L-glutamic acid and L-glutamine and their incorporation
into lenticular glutathione. Exp. Eye Res. 17:455–462.Kinsey, V. E. and Reddy, D. V. N. (1965) Studies on the crystalline lens. XI. The relative role of the
epithelium and capsule in transport. Inûest. Ophthalmol. and Vis. Sci. 4:104–116.Kleiman, N. J., Wang, R., and Spector, A. (1990) Ultraviolet light induced DNA damage and repair in
bovine lens epithelial cells. Curr. Eye Res. 9:1185–1193.

560 Bhat
Kondoh, H., Yasuda, K., and Okada, T. S. (1983) Tissue specific expression of a cloned chick δ-crystallingene in mouse cells. Nature 301:440–442.
Konofsky, K., Naumann, G. O. H., and Guggenmoos-Holtzmann, I. (1987) Cell density and sex chroma-tin in lens epithelium of human cataracts. Ophthalmol. 94:875–880.
Krausz, E. et al. (1996) Expression of crystallins pax 6, filensin, CP49, MIP and MP20 in lens derivedcell lines. Inûest. Ophthalmol. and Vis. Sci. 37:2120–2128.
Kuszak, J. R. (1995) The ultrastructure of epithelial and fiber cells in the crystalline lens. Intern. Reû.Cytol. 163:305–350.
Kuszak, J. R. (1997) A re-examination of primate lens epithelial cell size, density and structure as afunction of development, growth and age. Noûa Acta Leopoldina 57:45–66.
Lang, R. A. (1999) Which factors stimulate lens fiber cell differentiation in ûiûo? Inûest. Ophthalmol. Vis.Res. 40:3075–3078.
Lewis, W. H. (1904) Experimental studies on the development of the eye in amphibia. I. Origin of thelens Renu Palustris. Am. J. Anat. 3:505–536.
Li, H. C., Yang, J. M., Jacobson, R., Pasko, D., and Sundin, O. (1994) Pax-6 is first expressed in a regionof ectoderm anterior to the early neural plate: implications for stepwise determination of the lens.Deû. Biol. 162:181–194.
Li, W. and Spector, A. (1996) Lens epithelial cell apoptosis is an early event in the development of UVB-induced cataract. Free Radical Biol. Med. 20:301–311.
Li, W. C. et al. (1995a) Lens epithelial cell apoptosis appears to be a common cellular basis for non-congenital cataract development in humans and animals. J. Cell Biol. 130:169–181.
Li, W. C., Kuszak, J. R., Wang, G. M., Wu, Z. Q., and Spector, A. (1995b) Calcimycin induced lensepithelial cell apoptosis contributes to cataract formation. Exp. Eye Res. 61:91–98.
Lieska, N., Krotzer, K., and Yang, H. Y. (1992) A reassessment of protein synthesis by lens nuclear fibercells. Exp. Eye Res. 54:807–811.
Lin, L. R., Reddy, V. N., Giblin, F. J., Kador, P. F., and Kinoshita, J. H. (1990) Polyol accumulationin cultured human lens epithelial cells. Exp. Eye Res. 51:93–100.
Lo, W. K. and Zhang, W. (1989) Endocytosis of macromolecules in the lenses of guinea pig and rabbit.Lens Eye Toxicity Res. 6:603–612.
Lucas, V., Bassnet, S., Duncan, G., Stewart, S., and Gorghan, P. C. (1987) Membrane conductance andpotassium permeability of the rat lens. Q. J. Exp. Physiol. 72:81–93.
Mahon, K. A., Chepelinsky, A. B., Khillan, J. S., Overbeek, P. A., Piatigorsky, J., and Westphal, H.(1987) Oncogenesis of the lens in transgenic mice. Science 235:1622–1628.
Marcantonio, J. M., Rakic, J. M., Vrensen, G. F. J. M., and Duncan, G. (2000) Lens cell populationsstudied in human donor capsular bags with implanted intraocular lenses. Inûest. Ophthalmol. and Vis.Sci. 41:1130–1141.
Mathias, R. T., Rae, J. L., and Baldo, G. J. (1997) Physiological properties of the normal lens. Physiol.Reû. 77:21–50.
McAvovy, J. W. and Chamberlain, C. G. (1989) Fibroblast growth factor (FGF) induces differentresponses in lens epithelial cells depending on its concentration. Deûelopment 107:221–228.
McAvovy, J. W. and Fernon, V. T. (1984) Neutral retains promote cell division and fibre differentiationin lens epithelial explants. Curr. Eye Res. 3:827–834.
McAvovy, J. W. and Chamberlain, C. G. (1990) Growth factors in the eye. Progr. Growth Factor Res.2:29–43.
Menko, S., Philip, N., Veneziale, B., and Walker, J. (1988) Integrins and development: how might thesereceptors regulate differentiation of the lens. Ann. NY Acad. Sci. 15:36–41.
Merriam, J. C. et al. (2000) An action spectrum for UV-B radiation and the rat lens. Inûest. Ophthalmol.and Vis. Sci. 14:2642–2647.
Merriam-Smith, R., Donaldson, P., and Kistler, J. (1999) Differential expression of facilitative glucosetransporters GLUT1 and GLUT3 in the lens. Inûest. Ophthalmol. and Vis. Sci. 40:3224–3230.
Michael, R., Vrensen, G. F. J. M., van Marle, J., Gan, L., and Soderberg, P. G. (1998) Apoptosis in therat lens after in ûiûo threshold dose ultraviolet irradiation. Inûest. Ophthalmol. and Vis. Sci. 39:2681–2687.
Mikulicih, A. G. and Young, R. W. (1963) Cell proliferation and displacement in the lens epithelium ofyoung rats injected with tritiated thymidine. Inûest. Ophthalmol. and Vis. Sci. 2:344–354.

Ocular Lens Epithelium 561
Mosley, A. E., Dean, W. L., and Delamere, N. A. (1996) K-ATPase in rat lens epithelium and fiber cell.Inûest. Ophthalmol. and Vis. Sci. 37:1502–1508.
Nagamoto, T., Eguchi, G., and Beebe, D. C. (2000) Alpha-smooth muscle actin expression in culturedlens epithelial cells. Inûest. Ophthalmol. and Vis. Sci. 41:1122–1129.
Nagineni, C. N. and Bhat, S. P. (1988) Maintenance of the synthesis of α B-crystallin and progressiveexpression of βBp-crystallin in human fetal lens epithelial cells in culture. Deû. Biol. 130:402–405.
Nagineni, C. N. and Bhat, S. P. (1989a) Human fetal lens epithelial cells in culture: an in ûitro model forthe study of crystallin expression and lens differentiation. Curr. Eye Res. 8:285–291.
Nagineni, C. N. and Bhat, S. P. (1989b) α B-crystallin is expressed in kidney epithelial cell lines and notin fibroblasts. FEBS Lett. 249:89–94.
Nagineni, C. N. and Bhat, S. P. (1992) Lens fiber cell differentiation and expression of crystallins in co-cultures of human fetal lens epithelial cells and fibroblasts. Exp. Eye Res. 54:193–200.
Okada, T. S., Eguchi, G., and Takeichi, M. (1971) The expression of differentiation of chicken lensepithelium in in ûitro cell culture. Deû. Growth Differentiation 13:323–336.
Ogini, H. and Yasuda, K. (1998) Induction of lens differentiation by activation of a βZIP transcriptionfactor. L-Maf. Science 280:115–118.
Padgoankar, V. A., Giblin, F. J., Fowler, K., Leverenz, V. R., Reddan, J. R., and Dziedzic, D. C. (1997)Heme oxygenase synthesis is induced in cultured lens epithelium by hypobaric oxygen or puromycin.Exp. Eye Res. 65:435–443.
Parekh, A. B. and Penner, R. (1997) Store depletion and calcium influx. Physiol. Reû. 77:901–930.Paterson, C. A. (1972) Distribution and movement of ions in the ocular lens. Document. Ophthalmol.
31:1–28.Perry, M. M., Tassin, J., and Courtois, Y. A. (1979) Comparison of human lens epithelial cells in situ
and in ûitro in relation to aging: an ultra structural study. Exp. Eye Res. 28:327–341.Piatigorsky, J. (1981) Lens differentiation in vertebrates: A review of cellular and molecular features.
Differentiation 19:134–152.Piper, H. M., Spahr, R., Krutzfeldt, A., Siegmund, B., Schwartz, P., and Pau, H. (1990) Changes in the
energy metabolism of cultured lens epithelial cells in comparison with the fresh lens. Exp. Eye Res.51:131–138.
Pirie, A. (1965) Glutathione peroxidase in lens and a source of hydrogen peroxide in the aqueous humor.Biochem. J. 96:244–253.
Rae, J. L., Bartling, C., Rae, J., and Mathias, R. T. (1996) Dye transfer between cells of the lens.Membrane Biol. 150:89–103.
Rae, J. L. and Shephard, A. R. (2000) Kv3.3 potassium, channels in lens epithelium and corneal endo-thelium. Exp. Eye Res. 70:339–348.
Rae, J. and Shephard, A. R. (1996) Molecular biology and electrophysiology of calcium-activatedchannels from lens epithelium. Curr. Eye Res. 17:264–275.
Raff, M. C., Barres, B. A., Burne, J. F., Coles, H. S., Ishizaki, Y., and Jacobson, M. D. (1993) Pro-grammed cell death and the control of cell survival: Lessons from the nervous system. Science262:695–700.
Rafferty, N. S., Scholtz, D. I., Goldberg, M., and Lewckyj, M. (1990) Immunocytochemical evidence foran actin-myosin system in the lens epithelial cells. Exp. Eye Res. 51:591–600.
Ramamoorthy, S., Del Monte, M. A., Leibach, F. H., and Ganapathy, V. (1994) Molecular identity andcalmodulin-mediated regulation of the taurine transporter in a human retinal pigment epithelial cellline. Curr. Eye Res. 13:523–529.
Reddan, J. R. et al. (1981) Donor age influences the growth of rabbit lens epithelial cells in ûitro. VisionRes. 21:11–23.
Reddy, V. N. (2000) A forty-two year voyage through vision research. J. Ocular Pharmacol. Therapeut.16:97–107.
Reddy, D. V. N., Klethi, J., and Kinsey, V. E. (1966) Studies on the crystalline lens XII. Turnover ofglycine and glutamic acid in glutathione and ophthalmic acid in the rabbit. Inûest. Ophthalmol. andVis. Sci. 5:594–600.
Reddy, G. B. and Bhat, K. S. (1998) UV-B irradiation alters the activities and kinetic properties of theenzymes of energy metabolism in rat lens during aging. J. Photochem. Photobiol. B42:40–46.

562 Bhat
Reddy, V. N., Giblin, F. J., and Matsuda, H. (1980) Defense systems of the lens against oxidative damage.In: Red Blood Cell and Lens Metabolism (Srivastava, S., ed.), Elsevier, North Holland, pp. 139–154.
Reddy, V. N., Schwass, D., Chakrapani, B., and Lim, C. P. (1976) Biochemical changes associated withthe development and reversal of galactose cataracts. Exp. Eye Res. 23:483–493.
Ringens, P., Mungyer, G., Jap, P., Ramaekers, F., Hoenders, H., and Bloemendal, H. (1982) Exp. EyeRes. 35:313–324.
Robinson, K. R. and Patterson, J. W. (1983) Localization of steady currents in the lens. Curr. Eye Res.2:843–847.
Robinson Jr., W. G., Holder, N., and Kinoshita, J. H. (1990) Role of lens epithelium in sugar cataractformation. Exp. Eye Res. 50:641–646.
Robinson, M. L., Overbeek, P. A., and Verran, D. J. (1995) Extracellular FGF-1 acts as a lens differen-tiation factor in transgenic mice. Deûelopment 121:505–514.
Saha, M. S. et al. (1989) Embryonic lens induction: More than meets the optic vesicle. Cell Diff. Deû.28:153–171.
Sasaki, H., Lin, L. R., Yokoyama, T., Sevilla, M. D., Reddy, V. N., and Giblin, F. J. (1998) TEMPOLprotects against lens DNA strand breaks and cataract in the X-rayed rabbit. Inûest. Ophthalmol. andVis. Sci. 39:544–552.
Sawhney, R. S. (1995) Identification of SPARC in the anterior lens capsule and its expression by lensepithelial cells. Exp. Eye Res. 61:645–648.
Sax, C. M., Dziedzic, D. C., Piatigorsky, J., and Reddan, J. R. (1995) Analysis of a-crystallin expressionin cultured mouse and rabbit lens cells. Exp. Eye Res. 61:125–127.
Sax, C. M. and Piatigorsky, J. (1994) Expression of the alpha-crystallinsmall heat-shock proteinmol-ecular chaperone genes in the lens and other tissues. Adû. Enzymol. Related Areas of Mol. Biol.69:155–201.
Shang, F., Gong, X., and Taylor, A. (1997) Activity of ubiquitin-dependent pathway in response tooxidative stress. Ubiquitin-activating enzyme is transiently up regulated. J. Biol. Chem. 272:23086–23093.
Shui, Y. B. et al. (2000) Morphological observations on cell death and phagocytosis induced by ultravioletirradiation in a cultured human lens epithelial cell line. Exp. Eye Res. 71:609–618.
Sidjanin, D., Zigman, S., and Reddan, J. (1993) DNA damage and repair in rabbit lens epithelial cellsfollowing UVA irradiation. Curr. Eye Res. 12:773–781.
Simonneau, L., Herve, B., Jacquemin, E., and Courtois, Y. (1983) State of differentiation of bovineepithelial lens cells in ûitro. Relationship between the variation of the cell shape and the synthesis ofcrystallins. Cell Differ. 13:185–190.
Singh, D. P. et al. (2000) Lens epithelium derived growth factor LEDGF: Effects on growth and survivalfactor of lens epithelial cell, keratinocytes and fibroblasts. Biochem. Biophys. Res. Commun. 267:371–381.
Spector, A. (1995) Oxidative stress-induced cataract: mechanism of action. FASEB J. 9:1173–1182.Spector, A., Wang, R. R., Ma, W., and Kleiman, N. J. (2000) Development and characterization of and
H2O2-resistant immortal lens epithelial cell line. Inûest. Ophthalmol. and Vis. Sci. 41:832–843.Spemann, H. (1901) Uber korrelation in der entwickling des auges. Verh. Anat. Ges. 15:61–79.Spemann, H. (ed.) (1962) Embryonic Deûelopment and Induction. New York, Hafner Publishing.Srivastava, S. K., Singhal, S. S., Awasthi, S., Pikuka, S., Ansari, N. H., and Awasthi, Y. C. (1996) A
glutathione S-transferase isozyme (bGST5.8) involved in the metabolism of 4-hydroxy-2-trans-nonenal is localized in bovine lens epithelium. Exp. Eye Res. 63:329–337.
Starka, L., Hampl, R., Obenberger, J., and Doskocil, M. (1986) The role of corticosteroids in thehomeostasis of the eye. J. Steroid Biochem. 24:199–205.
Stellar, H. (1995) Mechanisms and genes of cellular suicide. Science 267:1445–1449.Stokes, J. et al. (2000) Distribution of glucocorticoid and mineralcorticoid receptors and 11β-hydroxy-
steroid dehydrogenases in human and rat ocular tissues. Inûest. Ophthalmol. and Vis. Sci. 41:1629–1638.
Straatsma, B. R., Lightfoot, D. O., Brake, R. M., Horwits, J. (1991) Lens capsule and epithelium in age-related cataract. Am. J. Ophthalmol. 112:283–296.
Struck, H. G., Hieder, C., and Lautenschlager, C. (2000) Veranderungen des linsenepithels bei diabetikernund nichtdiabetikern mit verschiedenen trubungsformen einer altersassoziierten katarakt. Klin.Monatsbl. Augenheilkd 216:204–209.

Ocular Lens Epithelium 563
Sun, K. J., Iwata, T., Zigler Jr., J. S., and Carper, D. A. (2000) Differential gene expression in male andfemale rat lenses undergoing cataract by transforming growth factor-beta (TGF-beta). Exp. Eye Res.70:169–181.
Tassin, J., Malaise, E., and Courtois, Y. (1979) Human lens cells have in ûitro proliferation capacityinversely proportional to the donor age. Exp. Cell Res. 123:388–392.
Tung, W., Chylack, L., and Andley, U. (1988) Lens hexokinase deactivation by near-UV irradiation.Curr. Eye Res. 7:257–263.
Vasavada, A. R., Cherian, M., Yadav, S., and Rawal, U. M. (1991) Lens epithelial cell density andhistomorphological study in cataractous lenses. J. Cataract Refract. Surg. 17:798–804.
Van Venrooji, W. J., Groenevald, A. A., Bloemendal, H., and Beneditti, E. L. (1974) Cultured calf lensepithelium. 1. Methods of cultivation and characteristics of the cultures. Exp. Eye Res. 18:517–526.
Vaziri, H. et al. (1999) Analysis of genomic integrity and p53-dependent G1 checkpoint in Telomerase-induced extended-life span human fibroblasts. Mol. Cell Biol. 19:2373–2379.
Vaulont, S., Vasseur-Cognet, M., and Kahn, A. (2000) Glucose regulation of gene transcription. J. Biol.Chem. 275:31555–31558.
Vermorken, A. J., Groenevald, A. A., Hilderink, J. M. H. C., de Waal, R., and Bloemendal, H. (1977)Dedifferentiation of lens epithelial cells in tissue culture. Mol. Biol. Rep. 3:371–378.
Vishwanath, R. I. et al. (1999) The transcriptional program in the response of human fibroblasts toserum. Science 283:83–867.
Wen, Y., Li, G. W., Chen, P., Wong, E., and Bekhor, I. (1995) Lens epithelial cell mRNA. II. Expressionof a mRNA encoding a lipid-binding protein in rat lens epithelial cells. Gene 158:269–274.
Wintour, E. M. (1997) Water channels and urea transporters. Clin. Exp. Pharmacol. Physiol. 24:1–9.Worgul, B. V. et al. (1991) Evidence of genotoxic damage in human cataractous lenses. Mutagenesis
6:495–499.Worgul, B. V., Merriam Jr., G. R., and Medvedovsky, C. (1989) Cortical cataract development—an
expression of primary damage to the lens epithelium. Lens Eye Toxic Res. 6:559–571.Wormstone, I. M., Liu, C. S. C., Rakic, J. M., Mercantonio, J. M., Vrensen, G. F. J. M., and Duncan,
G. (1997) Human lens epithelial cell proliferation in a protein-free medium. Inûest. Ophthalmol. andVis. Sci. 38:396–404.
Wormstone, I. M., Tamiya, S., Mercantonio, J. M., and Reddan, J. R. (2000) Hepatocyte growth factorfunction and c-Met expression in human lens epithelial cells. Inûest. Ophthalmol. and Vis. Sci.41:4216–4222.
Wride, M. A. and Saunders, E. J. (1998) Nuclear degeneration in the developing lens and its regulationby TNF-α. Exp. Eye Res. 66:371–383.
Yan Q., Clark, J. I., and Sage, E. H. (2000) Expression and characterization of SPARC in human lensand in the aqueous and vitreous humors. Exp. Eye Res. 71:81–90.
Zampighi, G. A., Eskandari, S., and Kremen, M. (2000) Epithelial organization of the mammalian lens.Exp. Eye Res. 71:415–435.
Zelenka, P. S., Gao, C. Y., Rampalli, A., Arora, J., Chauthaiwale, V., and He, H. Y. (1997) Cell cycleregulation in the lens: Proliferation, quiescence, apoptosis and differentiation. Prog. Ret. Eye Res.16:303–322.
Zhou, C. and Cammarata, P. R. (1997) Cloning the bovine NaCmyoinositol cotransporter gene andcharacterization of an osmotic responsive promoter. Exp. Eye Res. 65:349–363.
Zigman, S. (2000) Lens UV photobiology. J. Ocular Pharmacol. Therapeutics 16:161–165.




















