
Synthesis of Lens Capsule in Long-Term Culture of Human Lens Epithelial Cells
8
Synthesis of Lens Capsule in Long-Term Culture of Human Lens Epithelial Cells Tatsuo Arila, * Yasuhiro Murata, Li-Ren Lin, Toshiaki Tsuji, and Venkat N. Reddy Purpose. This study examined the extent to which human lens epithelial (HLE) cells in tissue culture retain the potential for differentiation, expression of lens-specific marker proteins, and the synthesis of lens capsule, the major characteristics of lens epithelium in vivo. Methods. Primary cultures of HLE cells were maintained for up to 450 days. Transmission and immunoelectron microscopy were used to study the thickness of the synthesized capsule and the formation of type IV collagen and laminin, two major protein components of the basement membrane of lens capsule in vivo. Results. In a long-term HLE culture system, without subcloning, lens fiber differentiation and capsular synthesis were maintained over a period of 450 days. In these cultures, the cell sheet showed three distinct zones: (1) a central zone with tight monolayer; (2) a mid peripheral zone with irregularly aggregated multilayer; and (3) a peripheral zone with loose monolayer. The basement membrane-like material was synthesized in the central zone and lentoids, which serve as a model for fiber differentiation, developed primarily in the mid peripheral zone. No capsu- lar material or lentoids were observed in the peripheral zone. The capsule-like material was 2 to 2.5 M m thick and showed the presence of type IV collagen and laminin, as detected by antibody reaction. Conclusion. This study demonstrates for the first time that HLE cells in long-term cultures synthesize a continuous sheet of capsule-like material. The findings also suggest that reforma- tion of a tight cell-to-cell relationship or generation of high cell density similar to that found in vivo may be an important factor for the synthesis of lens capsule. Invest Ophthalmol Vis Sci. 1993;34:355-362. A n ideal system for lens epithelial cells in culture maintains lens specific characteristics for a long time. The cells in vitro should retain the potential for differ- Frovi the Eye Research Institute, Oakland University, Rochester, Michigan. * Present Address: Department, of Ophthalmology, Kumamolo University School of Medicine, Kumamoto 860, Japan. This study was supported in part by the NEI Grants EY 00484 and EY 05025 (Core Grant for Vision Research). A preliminary report of these findings was presented at the annual meeting of the Association for Research in Vision and Ophthalmology, Sarasola, Florida, May, 1992. Submitted for publication: June 25, 1992; accepted September 14, 1992. Proprietary interest category: None. Reprint requests: Venkat, N. Reddy, Eye Research Institute, Oakland University, Rochester, Ml 48309-4401. entiation with concomitant expression of lens-specific marker proteins and the synthesis of the lens capsule. Recently, we have shown that human lens epithelial (HLE) cells express a and j3 crystallins for a limited time. 12 Spontaneous lentoid formation in monolayer of cultured cells is seen after 40-50 days. 3 " 5 A system in which lentoid bodies can be obtained predictably over a short time has been described 6 ' 7 and provides an important model for studying lens cell dif- ferentiation. These lentoids have been shown to ex- press 7 crystallin and MP 26 , proteins that are charac- teristic of mammalian lens fiber differentiation. 8 - 9 In addition, they continue to express a and fi crystallins analogous to lens fibers in vivo. Another characteristic of lens epithelial cells in vivo is the synthesis of a lens Investigative Ophthalmology & Visual Science, February 1993, Vol. 34, No. 2 Copyright © Association for Research in Vision and Ophthalmology 355
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Synthesis of Lens Capsule in Long-Term Culture of Human Lens Epithelial Cells

Synthesis of Lens Capsule in Long-Term Cultureof Human Lens Epithelial Cells
Tatsuo Arila, * Yasuhiro Murata, Li-Ren Lin, Toshiaki Tsuji, and Venkat N. Reddy
Purpose. This study examined the extent to which human lens epithelial (HLE) cells in tissueculture retain the potential for differentiation, expression of lens-specific marker proteins,and the synthesis of lens capsule, the major characteristics of lens epithelium in vivo.
Methods. Primary cultures of HLE cells were maintained for up to 450 days. Transmission andimmunoelectron microscopy were used to study the thickness of the synthesized capsule andthe formation of type IV collagen and laminin, two major protein components of the basementmembrane of lens capsule in vivo.
Results. In a long-term HLE culture system, without subcloning, lens fiber differentiation andcapsular synthesis were maintained over a period of 450 days. In these cultures, the cell sheetshowed three distinct zones: (1) a central zone with tight monolayer; (2) a mid peripheral zonewith irregularly aggregated multilayer; and (3) a peripheral zone with loose monolayer. Thebasement membrane-like material was synthesized in the central zone and lentoids, which serveas a model for fiber differentiation, developed primarily in the mid peripheral zone. No capsu-lar material or lentoids were observed in the peripheral zone. The capsule-like material was 2to 2.5 Mm thick and showed the presence of type IV collagen and laminin, as detected byantibody reaction.
Conclusion. This study demonstrates for the first time that HLE cells in long-term culturessynthesize a continuous sheet of capsule-like material. The findings also suggest that reforma-tion of a tight cell-to-cell relationship or generation of high cell density similar to that found invivo may be an important factor for the synthesis of lens capsule. Invest Ophthalmol Vis Sci.1993;34:355-362.
A n ideal system for lens epithelial cells in culturemaintains lens specific characteristics for a long time.The cells in vitro should retain the potential for differ-
Frovi the Eye Research Institute, Oakland University, Rochester,Michigan.* Present Address: Department, of Ophthalmology, Kumamolo UniversitySchool of Medicine, Kumamoto 860, Japan.This study was supported in part by the NEI Grants EY 00484 and EY05025 (Core Grant for Vision Research).A preliminary report of these findings was presented at the annualmeeting of the Association for Research in Vision and Ophthalmology,Sarasola, Florida, May, 1992.Submitted for publication: June 25, 1992; accepted September 14, 1992.Proprietary interest category: None.Reprint requests: Venkat, N. Reddy, Eye Research Institute, OaklandUniversity, Rochester, Ml 48309-4401.
entiation with concomitant expression of lens-specificmarker proteins and the synthesis of the lens capsule.Recently, we have shown that human lens epithelial(HLE) cells express a and j3 crystallins for a limitedtime.12 Spontaneous lentoid formation in monolayerof cultured cells is seen after 40-50 days.3"5
A system in which lentoid bodies can be obtainedpredictably over a short time has been described6'7 andprovides an important model for studying lens cell dif-ferentiation. These lentoids have been shown to ex-press 7 crystallin and MP26, proteins that are charac-teristic of mammalian lens fiber differentiation.8-9 Inaddition, they continue to express a and fi crystallinsanalogous to lens fibers in vivo. Another characteristicof lens epithelial cells in vivo is the synthesis of a lens
Investigative Ophthalmology & Visual Science, February 1993, Vol. 34, No. 2Copyright © Association for Research in Vision and Ophthalmology 355

356 Investigative Ophthalmology & Visual Science, February 1993, Vol. 34, No. 2
capsule.10 This has been demonstrated for the rat byauto radiography.1112 There have been several at-tempts to study the synthesis of capsular substance incell culture. Extracellular periodic acid-Schiff-posi-tive, diastase-resistant fibrous substance was producedby cultured epithelial cells of trout lens.13 In bovinelens cells, thin sheets and strands of capsule-like sub-stance were observed in floating cell aggregates.14 Im-munoreaction to anti-type IV collagen antibodies wasdetected in the extracellular material of multilayer ofbovine lens epithelial cell culture15 and in clonedmouse lens epithelial cells.16
To our knowledge, no in vitro studies have indi-cated expression of capsular material by HLE cells.Therefore, it is important to discover whether HLEcells in culture are capable of forming the lens capsule.Although most HLE cell cultures seem to retain thecapacity to form lentoid bodies synchronously whenthey are grown on a nonhaptotactic surface,6-7 sponta-neous lentoids do not arise evenly in monolayer cul-ture.l>3>4 Is there a prerequisite condition for the ex-pression of lens-specific characteristics for culturedHLE cells?
These questions were examined in a long-termculture of HLE cells in vitro. The morphologic andhistochemical evidence presented here demonstratesfor the first time that HLE cells in tissue culture syn-thesize lens capsule-like material as a continuoussheet. The presence of type IV collagen and laminin asdemonstrated by antibody reaction suggests that thebasement membrane-like material synthesized by HLEcells is similar to the lens capsule. This report alsodescribes some of the characteristics of HLE cellsmaintained for more than 1 yr in tissue culture.
MATERIALS AND METHODS
Cell Culture
Specimens of HLE cells were obtained from infants 5to 12 mo old who underwent surgery for retinopathyof prematurity. The method for establishing primarycultures has been described previously.1 The researchfollowed the tenets of the Declaration of Helsinki. In-formed consent was obtained and Oakland Univer-sity's institutional human experimentation committeeapproval was granted. Each primary or subculture wasestablished independently in a culture dish (Falcon;Becton Dickinson, Oxnard, CA) and maintained with-out further passage for 20 to 450 days. Subculturedcell sheets for a long-term culture were establishedwith high cell density inoculation to the center of adish—approximately 1800 cells/mm2—to eliminatethe de-differentiation of monolayer cultured cells, asdescribed previously.617 Cultures were incubated in a5% CO2 atmosphere at 36.5°C. Antibiotic-free Eagle's
minimal essential medium (Gibco, Grand Island, NY)supplemented with 20% fetal bovine serum (Gibco)was replaced twice a week.4'7
Ultrastructure of Cultured Cells and BasementMembrane
The cultures were fixed in half Karnovsky's solutionfor 2-3 hr after OsO4 vapor prefixation for 30 minand post-fixed in 1% OsO4 solution for 60 min.6'7
Compact monolayer cell sheets with a thick basementmembrane were detached and peeled off from culturedishes with fine forceps under a binocular microscopeduring the rinse with phosphate buffered saline (PBS).This epithelial-membrane tissue was dehydrated in agraded series of ethanol and infiltrated with propyleneoxide. Ultrathin sections of materials embedded inEpon 812 were stained with uranyl acetate and Rey-nold's lead solution and examined with an ISI-LEM(Pleasanton, CA) 2000 or a Phillips (Mahwah, NJ) 410transmission electron microscope (TEM).
Tissue Preparation for Immunohistochemistry
The in situ human lens was obtained from a 55-yr-oldman who underwent cataract surgery by intracapsularcataract extraction. The lens was fixed in periodate-ly-sine-paraformaldehyde (PLP) fixative for 4 hr at 4°C,washed with 0.01 mol/1 PBS that contained a gradientconcentration of sucrose (10%, 15%, 20%, and 20%+ 5% glycerol). The lens then was embedded in Tis-sue-Tek OCT compound (Miles Laboratories, Elkhart,IN), quick-frozen in liquid nitrogen, and stored at—70°C for cryostat sectioning.
The monolayer HLE cultures with basementmembranes were fixed in PLP fixative for 20 min at4°C and washed. Some cultures were fixed after 90days, whereas others were used after 450 days. Themonolayer cell sheets with thick basement membraneswere scraped off from the culture dish with a razorblade. The cell sheets were embedded, frozen, andstored as described above.
Immunoelectron MicroscopyCryosections (6 /xm) cut with cryotome (2800 Frigo-cut-E; Reichert-Jung, Nuloch, Germany) at — 20°Cwere first incubated with 5% normal rabbit serum and5% normal goat serum in PBS to prevent nonspecificbinding. They then were treated with anti-type IV col-lagen antibody (diluted 1:50; Accurate, Westbury, NY)and anti-laminin antibody (diluted 1:50; Gibco), re-spectively, overnight at 4°C. All sections were washedthree times with PBS containing 10% sucrose, fol-lowed by an overnight incubation with horseradishperoxidase-conjugated 4 F(ab')2 rabbit anti-goat IgG(diluted 1:50; Cappel, Durham, NC) and F(ab')2 goatanti-rabbit IgG (diluted 1:50; Cappel) for anti-type IVcollagen antibody and anti-laminin antibody staining,

Lens Capsule Synthesis in Human Lens Epithelium 357
respectively. After several washes, the sections werefixed in 1% glutaraldehyde in PBS for 10 min. Theperoxidase reaction then was initiated by adding0.005% H2O2 for 10 min. The sections were post-fixedin 1% OsO4 for 1 hr at room temperature, dehydratedin ethanol, and embedded in Epon 812. Unstainedultrathin sections were examined with a TEM. In thecontrol sections, nonimmune goat serum and nonim-mune rabbit serum were used in place of anti-type IVcollagen antibody and anti-laminin antibody, respec-tively.
RESULTS
Cell Culture
Cultures could be maintained for more than a yearwith no microbial contamination with the antibiotic-free culture medium. Both cell sheets, established byprimary explants and by high cell density inoculation,showed no significant differences in their growth pat-tern and appearance under the phase contrast micro-scope. Mitosis of HLE cells was most frequently seenin the mid peripheral zone, where the cell densitieswere higher than in the peripheral zone but lower thanin the central areas.
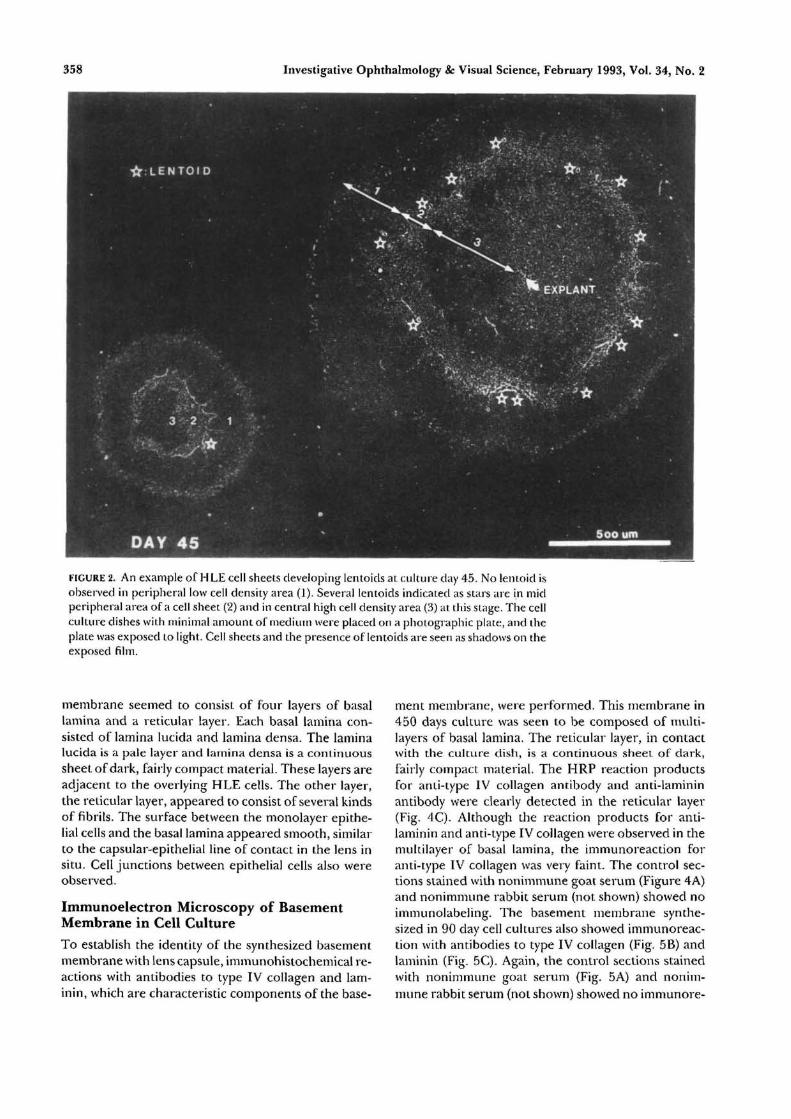
The measurement by random sampling of celldensities surrounding the dividing cells suggested thetendency toward density-dependent growth of HLEcells, similar to the corneal endothelial cells culturedin vitro.17 Cultures became confluent or cell sheetsalmost ceased to grow after 20 to 30 days of culture. Atthis stage, cells gradually overlapped or became irregu-larly aggregated, and extracellular fibrous strands be-gan to appear. In cultures of 20 to 90 days, the cellsheet could be divided into three different zones (Fig.1): (1) a central zone where compact monolayer cellsarranged regularly; (2) a mid peripheral zone wherecells aggregated irregularly with the appearance of fi-brous strands; and (3) a peripheral zone where cellselongated with a polymorphous shape and wide inter-cellular space. Spontaneous lentoids usually began toappear in the mid peripheral irregular zone after 40 to50 days of culture. Over the following 450 days thenumber of lentoids still increased. Occasionally, len-toid formation also was observed in the center of thecell sheet or in the periphery. The major cell popula-tion still was in a monolayer after 1 yr. Figure 2 showsthe pattern of growth and spontaneous lentoid forma-tion in different areas in a typical culture plate. Cellsheets and the presence of lentoids are seen asshadows.
Ultrastructure of Epithelial BasementMembrane in Cell Culture
A transparent, elastic, and delicate membrane wasseen in long-term cultures (450 days) under the com-
kFIGURE 1. Schematic diagram illustrating overall patterns ofmonolayer culture of HLE cells. (1) Developing cell sheetswith active cell proliferation. Cell densities are higher atcenter (left side) and lower at periphery (right side). Mitosisis frequently seen at mid periphery. (2) Cell sheet after 20 to30 days. Cells in mid periphery area become multilayered.(3) Early lentoid formation in the multilayering region afterabout 40 days of culture. Although center of the cell sheetsconsists of uniformly compact and regularly arranged mono-layer cells, peripheral cell sheets consist of flattened and ir-regular shaped cells. (4) Lentoid formation is observed notonly in mid periphery but also in center and periphery after2 or 3 mo. A thick basement membrane (arrow head) is seenunder the compact monolayer.
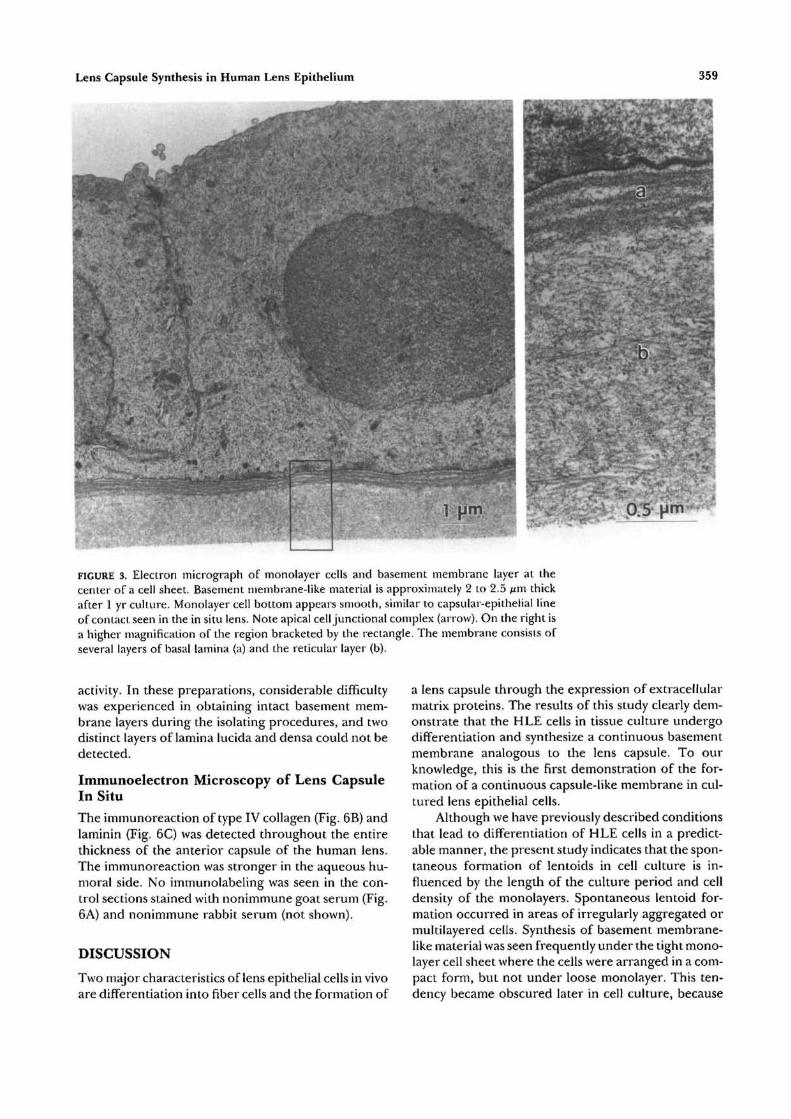
pact monolayer cells in the center encircled by an ir-regular zone. Upon careful detachment from the cul-ture dish, a compact and continuous capsule-like sheetcould be seen between the monolayer cells and theculture dish. This membrane was relatively easilypeeled off from the plastic surface with fine forcepsunder a binocular microscope. Electron micrographs(Fig. 3) revealed that the basement membrane-like ma-terial of approximately 2 to 2.5 /xm thick was formedunder the compact monolayer of HLE cells. This
Investigative Ophthalmology 8c Visual Science, February 1993, Vol. 34, No. 2
EXEUNT
3 2 1
DAY 45 5oo urn
FIGURE 2. An example of HLE cell sheets developing lentoids at culture day 45. No lentoid isobserved in peripheral low cell density area (1). Several lentoids indicated as stars are in midperipheral area of a cell sheet (2) and in central high cell density area (3) at this stage. The cellculture dishes with minimal amount of medium were placed on a photographic plate, and theplate was exposed to light. Cell sheets and the presence of lentoids are seen as shadows on theexposed film.
membrane seemed to consist of four layers of basallamina and a reticular layer. Each basal lamina con-sisted of lamina lucida and lamina densa. The laminalucida is a pale layer and lamina densa is a continuoussheet of dark, fairly compact material. These layers areadjacent to the overlying HLE cells. The other layer,the reticular layer, appeared to consist of several kindsof fibrils. The surface between the monolayer epithe-lial cells and the basal lamina appeared smooth, similarto the capsular-epithelial line of contact in the lens insitu. Cell junctions between epithelial cells also wereobserved.
Immunoelectron Microscopy of BasementMembrane in Cell Culture
To establish the identity of the synthesized basementmembrane with lens capsule, immunohistochemical re-actions with antibodies to type IV collagen and lam-inin, which are characteristic components of the base-
ment membrane, were performed. This membrane in450 days culture was seen to be composed of multi-layers of basal lamina. The reticular layer, in contactwith the culture dish, is a continuous sheet of dark,fairly compact material. The HRP reaction productsfor anti-type IV collagen antibody and anti-lamininantibody were clearly detected in the reticular layer(Fig. 4C). Although the reaction products for anti-laminin and anti-type IV collagen were observed in themultilayer of basal lamina, the immunoreaction foranti-type IV collagen was very faint. The control sec-tions stained with nonimmune goat serum (Figure 4A)and nonimmune rabbit serum (not shown) showed noimmunolabeling. The basement membrane synthe-sized in 90 day cell cultures also showed immunoreac-tion with antibodies to type IV collagen (Fig. 5B) andlaminin (Fig. 5C). Again, the control sections stainedwith nonimmune goat serum (Fig. 5A) and nonim-mune rabbit serum (not shown) showed no immunore-
Lens Capsule Synthesis in Human Lens Epithelium 359
FIGURE 3. Electron micrograph of monolayer cells and basement membrane layer at thecenter of a cell sheet. Basement membrane-like material is approximately 2 to 2.5 fim thickafter 1 yr culture. Monolayer cell bottom appears smooth, similar to capsular-epithelial lineof contact seen in the in situ lens. Note apical cell junctional complex (arrow). On the right isa higher magnification of the region bracketed by the rectangle. The membrane consists ofseveral layers of basal lamina (a) and the reticular layer (b).
activity. In these preparations, considerable difficultywas experienced in obtaining intact basement mem-brane layers during the isolating procedures, and twodistinct layers of lamina lucida and densa could not bedetected.
Immunoelectron Microscopy of Lens CapsuleIn Situ
The immunoreaction of type IV collagen (Fig. 6B) andlaminin (Fig. 6C) was detected throughout the entirethickness of the anterior capsule of the human lens.The immunoreaction was stronger in the aqueous hu-moral side. No immunolabeling was seen in the con-trol sections stained with nonimmune goat serum (Fig.6A) and nonimmune rabbit serum (not shown).
DISCUSSION
Two major characteristics of lens epithelial cells in vivoare differentiation into fiber cells and the formation of
a lens capsule through the expression of extracellularmatrix proteins. The results of this study clearly dem-onstrate that the HLE cells in tissue culture undergodifferentiation and synthesize a continuous basementmembrane analogous to the lens capsule. To ourknowledge, this is the first demonstration of the for-mation of a continuous capsule-like membrane in cul-tured lens epithelial cells.
Although we have previously described conditionsthat lead to differentiation of HLE cells in a predict-able manner, the present study indicates that the spon-taneous formation of lentoids in cell culture is in-fluenced by the length of the culture period and celldensity of the monolayers. Spontaneous lentoid for-mation occurred in areas of irregularly aggregated ormultilayered cells. Synthesis of basement membrane-like material was seen frequently under the tight mono-layer cell sheet where the cells were arranged in a com-pact form, but not under loose monolayer. This ten-dency became obscured later in cell culture, because

360 Investigative Ophthalmology & Visual Science, February 1993, Vol. 34, No. 2
a
V
cFIGURE 4. Immunoelecii on micrograph showing antibody re-action: (a) control, (b) type IV collagen, and (c) laminin inbasement membrane-like material {between two arrowheads) at the center of a cell sheet at 450th culture day.Immunoreaciion for laminin is detected in multilayers ofbasal lamina and reticnlar layer, and type IV collagen is de-tected in reticular layer. Control shows no reaction. (Bar= 1 Mm-)
an occasional lentoid also could be seen even in areasof tight monolayer after about 90 days.
During the course of cell sheet formation, activemitosis increased cell number and cell density as thecell sheet grew in size. The center of the cell sheetbecame confluent and monolayer was formed withtime. The cells at the periphery became flat and poly-morphous. The edge of the cell sheet was less compact
FIGURE 5. 1 mmunoelectron micrograph showing antibody re-action in cell sheet at 90th culture day: (a) control, (b) typeIV collagen, and (c) laminin in basement membrane-like ma-terial at the center of a cell sheet at 90th culture day. TypeIV collagen and laminin are detected in basement mem-brane-like material. Control shows no reaction. (Bar= 1 urn.)
with the loss of cell-to-cell communication. That base-ment membrane-like material was formed primarilyunder tight cell monolayer, which is homologous tothe lens epithelial cells in situ, suggests that high celldensity may be a factor in promoting the synthesis ofcapsular material.

Lens Capsule Synthesis in Human Lens Epithelium 361
a
FIGURE6. Immunoelectron micrograph showingantibody re-action in in situ anterior human lens capsule: (a) control, (b)type IV collagen, and (c) laniinin. Immunoreaction of typeIV collagen and laniinin is detected throughout the entirethickness of the lens capsule. Reaction is stronger inaqueous humoral side. Control shows no reaction. (Bar= 5 M'11-)
The identity of the basement membrane-like mate-rial synthesized in tissue culture with lens capsule insitu was examined with electron microscopy and reac-tion with antibodies to type IV collagen and laminin.Basement membranes are ubiquitous, complex multi-component structures that separate connective tissuefrom parenchymal cells. They usually are less than 0.2
^m thick and seem to consist of two layers. Laminalucida is a pale layer adjacent to the basal plasma-lemma of the associated parenchymal cells. The sec-ond layer, lamina densa, is a continuous sheet of dark,fairly compact material that makes contact with con-nective tissue. The third layer, referred to as reticularlayer, often is missing. However, when it is present, itis known to consist of scattered elements such as colla-gen fibrils, anchoring fibrils, or microfibrils in contactwith lamina densa. The lens capsule in situ, however, isa much thicker accumulation of lamina densa-like ma-terial that lacks lamina lucida.18'19
Transmission electron microscopy revealed thatthe thickness of the basement membrane-like materialsynthesized after 450 days of culture was nearly 2 to2.5 ixm. This corresponds to the thickness of lens cap-sule at posterior pole in vivo after the first decade oflife.20 The thickness of human adult anterior lens cap-sule is nearly 10 to 20 /mi.20 In contrast to newly syn-thesized basement membrane-like material in vitro,lens capsule in vivo has been reported to lack the retic-ular layers.19 The reason for these differences in theultraslructure of in situ lens capsule and the synthe-sized basement membrane-like material in our tissueculture is not known. It is not known whether laminadensa gradually gets thicker with age and lamina lucidadisappears in vivo. It must be emphasized that al-though we have referred to the two immunostainingbasement membrane layers as lamina densa and lam-ina lucida (Fig. 4), a careful examination of the ul-trathin sections (not iinmunostained; Fig. 3) show fourlayers of basal lamina (light and dark bands). The un-derlying dark layer of basement membrane sometimeshas been referred to as lamina diffusa or pars fibrore-ticularis (or reticular layer). Further studies areneeded to define these structures more precisely.
Although the molecular structure of lens capsulehas not been clearly resolved, the protein componentsof basement membrane (type IV collagen, laminin,heparan sulfate proteoglycan, entactin) were detectedin calf and bovine lens capsule by immunoblotting21
and iinmunofluorescent microscopy,18 respectively.Our studies with immunoelectron microscopy clearlyshow that type IV collagen and laminin, two majorprotein components of basement membrane, are pres-ent throughout the entire thickness of anterior humanlens capsule.
In conclusion, it is apparent that the cell culturesystem described here is a useful model for a detailedinvestigation of the expression of crystallins and extra-cellular matrix proteins of human lens during develop-ment.
Key Words
extracellular matrix, human lens epithelium, laminin, lenscapsule, tissue culture, type IV collagen.

362 Investigative Ophthalmology 8c Visual Science, February 1993, Vol. 34, No. 2
Acknowledgments
We are grateful to Dr. Michael T. Trese of the Eye ResearchInstitute of Oakland University and William Beaumont Hos-pital, Royal Oak, Michigan, for his cooperation in obtainingspecimens of human lens epithelium.
References
1. Recldy VN, Lin L-R, Arita T, Zigler JS, Huang QLCrystallins and their synthesis in human lens epithelialcells in tissue culture. Exp Eye Res. 1988;47:465-478.
2. Redcly VN, Katsura H, Arita T, et al. Study of crystal-lin expression in human lens epithelial cells duringdifferentiation in culture and in non-lenticular tissue.Exp Eye Res. 1991;53:367-374.
3. Hamada Y, Okada TS. In vitro differentiation of cellsof the lens epithelium of human fetus. Exp Eye Res.1978; 26:91-97.
4. Eguchi G, Kodama R. A study of human senile cata-ract: Growth and differentiation of lens epithelial cellsin vitro cell culture. Ophthalmic Res. 1979; J 1:308-3J5.
5. Okada TS, Eguchi G, Takeichi M. The expression ofdifferentiation of chicken lens epithelium in vitro cellculture. Development, Growth and Differentiation. 1971;13:323-335.
6. Arita T, Lin L-R, Reddy VN. Differentiation of hu-man lens epithelial cells in tissue culture. Exp Eye Res.1988;47:905-910.
7. Arita T, Lin L-R, Susan SR, Reddy VN. Enhancementof differentiation of human lens epithelium in tissueculture by change in cell-substrate adhesion. InvestOphthalmol Vis Sci. 1 990;3 1:2395-2404.
8. Papaconstantinou J. Molecular aspects of lens cell dif-ferentiation. Science. 1967; 156:338-346.
9. Bloemendal H. Lens proteins as markers of terminaldifferentiation. Ophthalmic Res. 1979; 11:243-253.
1 0. Rafferty NS, Goossens W. Growth and aging of thelens capsule. Growth. 1978;42:375-389.
1 I. Young RW, Ocumpaugh DE. Autoradiographic stud-
ies on the growth and development of the lens capsulein the rat. Invest Ophthalmol. 1966;5:583-593.
12. Haddad A, Bennet G Synthesis of lens capsule andplasma membrane giycoproteins by lens epithelialcells and fibers in the rat. American Journal of Anatomy1988;183:212-225.
13. von Sallman L, Grimes PA, Albert DM. Histogenesisof the lens capsule in tissue culture. AmJ Ophthalmol.1969; 68:435-438.
14. Iwig M, Geyer G, Marquardt I, Glaesser D. Formationof capsule like material during long-term cultivationof bovine lens epithelial cells. Ada Biol Med Genii.1976;35:413-419.
15. Laurent M, Lonchampt M-O, Regnault F, Tassin J,Courtois Y. Biochemical, ultrastructural and immuno-logical study of in vitro production of collagen by bo-vine lens epithelial cells in culture. Exp Cell Res.1978; 115:127-142.
16. Rohrbach DH, Russell P, Church RL. In vitro produc-tion of basement membrane collagen by a clonal lineof mouse lens epithelial cells. Curr Eye Res.1981; 1:267-273.
17. Arita T, Okamura R, Kodama R, Takeuchi T, KodamaY, Eguchi G. Density dependent growth of cornealendothelial cells cultured in vitro. Cell Differentiation.1987;22:61-70.
18. Cammarata PR, Cantu-Crouch D, Oakford L, MorrillA. Macromolecular organization of bovine lens cap-sule. Tissue Cell. 1986; 18:83-97.
19. Inoue S, Leblond CP. Three-dimensional network ofcords: The main component of basement membranes.American Journal of Anatomy. 1988; 181:341 -358.
20. Worgul B: Lens. In: Jakobiec FA, ed. Ocular Anatomy,Embiyology and Teratology. Philadelphia: Harper &Row Publishers; 1982:355-368.
21. Mohan PS, Spiro RG. Macromolecular organizationof basement membranes: Characterization and com-parison of glomerular basement membrane and lenscapsule components by immunochemical and lectinaffinity procedures. J Biol Chem. 1986; 261:4328-4336.




















