
The 14-3-3 Gene Expression Specificity in Response to Stress is Promoter-Dependent
11
Plant Cell Physiol. 46(10): 1635–1645 (2005) doi:10.1093/pcp/pci179, available online at www.pcp.oupjournals.org JSPP © 2005 1635 The 14-3-3 Gene Expression Specificity in Response to Stress is Promoter- Dependent Anna Aksamit 1 , Alina Korobczak 2 , Jacek Skala 1 , Marcin Lukaszewicz 1 and Jan Szopa 2, * 1 Institutes of Genetics and Microbiology, Wroclaw University, Przybyszewskiego 63/77, 51-148 Wroclaw, Poland 2 Biochemistry and Molecular Biology, Wroclaw University, Przybyszewskiego 63/77, 51-148 Wroclaw, Poland ; Genomic clone coding for the 16R isoform of 14-3-3 proteins from potato plants has recently been described. This paper reports on 20R-gene isolation and analysis, and compares two isoforms. The northern blot analysis of mRNA of the 20R 14-3-3 isoform suggests its similarity to 16R. Vascular tissue-specific expression and age-dependent synthesis in potato leaves has been detected in both promot- ers. Screening of the potato genomic library using 20R cDNA isoform resulted in identification and isolation of the corresponding gene. This gene contains four exons and three introns. Inspecting the promoter sequence of the 20R isoform revealed several boxes important for the regula- tion of gene expression. The strongest GUS expression in transgenic potato plants transformed with the uidA reporter gene under the 20R promoter has been found in young leaf and stem vascular tissue, root tips, pollen and ovules. Mature fragments exhibit a significant decrease in GUS staining, which suggests age-dependent promoter activity. The analysis of transgenic plants transformed with 20R-GUS in contrast to 16R-GUS has revealed strong acti- vation of the 20R promoter by metal ions and NaCl. Instead the 16R promoter is strongly affected by virus and salicylic acid treatments. The only factor, which strongly induced both promoters, was abscisic acid. It is thus suggested that promoter domain composition is the main factor differenti- ating the appearance of 14-3-3 isoforms. Keywords: 14-3-3 gene — GUS staining — Promoter analysis — Solanum tuberosum. The nucleotide sequences reported in this paper have been sub- mitted to the EMBL/GenBank databases under the following acces- sion numbers: 14-3-3 20R gene [AY518222.]; 20R cDNA [X87370]; 16R gene [Y070220]. Introduction Recent investigations suggest the participation of 14-3-3 proteins in numerous biological processes of eukaryotic cells. Members of the 14-3-3 protein family activate neurotransmit- ter synthesis; activate ADP-ribosylation of proteins; and regulate the activity of protein kinase C, sucrose phosphate synthase (SPS), starch synthase (SS) and nitrate reductase (NR). Moreover, they display phospholipase A 2 activity and associate with the products of proto-oncogenes, oncogenes and the cdc 25 gene (Chung et al. 1999, Szopa 2002). The broad spectrum of activities that are affected by 14-3-3 proteins sug- gests their potential in modifying plant development and metabolism (Finnie et al. 1999, Roberts 2003). While in recent years there has been substantial progress in the identification of diverse partners of 14-3-3 proteins, at least two important questions remain to be answered: (i) is there any specificity in the action of a particular 14-3-3 iso- form on the biology of the plant; and (ii) why do cells contain several isoforms and how is the expression of each particular isoform regulated? To answer these questions, we investigated a number of 14-3-3 isoforms, examining the tissue specificity of their expression, and their age-related synthesis in potato plants. By screening cDNA libraries from leaves, roots and epidermal fragments, six cDNAs encoding 14-3-3 isoforms had been pre- viously found in potato plants (Wilczynski et al. 1998). To discover the significance of 14-3-3 isoforms for plant metabolism, transgenic potatoes with repression of one, two and six mRNAs were created. The selected transgenic plants were analysed for enzyme activities known to interact with 14- 3-3 proteins, and metabolites, which are the products of the interacting enzymes. A significant increase in NR, SPS and SS activities was found for several of the analyzed enzymes (Zuk et al. 2003). The changes in enzyme activity in transgenic plants are reversible and can be complemented by heter- ogonous recombinant 14-3-3 proteins either from Cucurbita pepo or potato plants. It is interesting to note that the level of all measured activity strongly depends on 14-3-3 protein con- tent in the generated transgenic plants, thus suggesting that the quantity of 14-3-3 protein rather than the particular isoform controls the interacting enzymes. The questions now arise why the genome of eukaryotes contains several 14-3-3 isoforms and how the genes’ expression is regulated. To try to answer these questions, the gene encoding the 16R isoform of 14-3-3 proteins has recently been isolated and analysed (Szopa et al. 2003). Several motifs have been found supporting the assumption that the promoter could be tissue- specific and developmentally-regulated. Western and northern analyses as well as the histochemical and fluorometric assays * Corresponding author: E-mail, [email protected]; Fax, +48-71-3252930. at Uniwersytet Wroclawski, Biblioteka Uniwersytecka on July 26, 2012 http://pcp.oxfordjournals.org/ Downloaded from
Transcript of The 14-3-3 Gene Expression Specificity in Response to Stress is Promoter-Dependent

Plant Cell Physiol. 46(10): 1635–1645 (2005)
doi:10.1093/pcp/pci179, available online at www.pcp.oupjournals.org
JSPP © 2005
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
The 14-3-3 Gene Expression Specificity in Response to Stress is Promoter-Dependent
Anna Aksamit 1, Alina Korobczak 2, Jacek Skala 1, Marcin Lukaszewicz 1 and Jan Szopa 2, *
1 Institutes of Genetics and Microbiology, Wroclaw University, Przybyszewskiego 63/77, 51-148 Wroclaw, Poland 2 Biochemistry and Molecular Biology, Wroclaw University, Przybyszewskiego 63/77, 51-148 Wroclaw, Poland
;
Genomic clone coding for the 16R isoform of 14-3-3
proteins from potato plants has recently been described.
This paper reports on 20R-gene isolation and analysis, and
compares two isoforms. The northern blot analysis of
mRNA of the 20R 14-3-3 isoform suggests its similarity to
16R. Vascular tissue-specific expression and age-dependent
synthesis in potato leaves has been detected in both promot-
ers. Screening of the potato genomic library using 20R
cDNA isoform resulted in identification and isolation of the
corresponding gene. This gene contains four exons and
three introns. Inspecting the promoter sequence of the 20R
isoform revealed several boxes important for the regula-
tion of gene expression. The strongest GUS expression in
transgenic potato plants transformed with the uidA
reporter gene under the 20R promoter has been found in
young leaf and stem vascular tissue, root tips, pollen and
ovules. Mature fragments exhibit a significant decrease in
GUS staining, which suggests age-dependent promoter
activity. The analysis of transgenic plants transformed with
20R-GUS in contrast to 16R-GUS has revealed strong acti-
vation of the 20R promoter by metal ions and NaCl. Instead
the 16R promoter is strongly affected by virus and salicylic
acid treatments. The only factor, which strongly induced
both promoters, was abscisic acid. It is thus suggested that
promoter domain composition is the main factor differenti-
ating the appearance of 14-3-3 isoforms.
Keywords: 14-3-3 gene — GUS staining — Promoter analysis
— Solanum tuberosum.
The nucleotide sequences reported in this paper have been sub-
mitted to the EMBL/GenBank databases under the following acces-
sion numbers: 14-3-3 20R gene [AY518222.]; 20R cDNA [X87370];
16R gene [Y070220].
Introduction
Recent investigations suggest the participation of 14-3-3
proteins in numerous biological processes of eukaryotic cells.
Members of the 14-3-3 protein family activate neurotransmit-
ter synthesis; activate ADP-ribosylation of proteins; and
regulate the activity of protein kinase C, sucrose phosphate
synthase (SPS), starch synthase (SS) and nitrate reductase
(NR). Moreover, they display phospholipase A2 activity and
associate with the products of proto-oncogenes, oncogenes and
the cdc 25 gene (Chung et al. 1999, Szopa 2002). The broad
spectrum of activities that are affected by 14-3-3 proteins sug-
gests their potential in modifying plant development and
metabolism (Finnie et al. 1999, Roberts 2003).
While in recent years there has been substantial progress
in the identification of diverse partners of 14-3-3 proteins, at
least two important questions remain to be answered: (i) is
there any specificity in the action of a particular 14-3-3 iso-
form on the biology of the plant; and (ii) why do cells contain
several isoforms and how is the expression of each particular
isoform regulated?
To answer these questions, we investigated a number of
14-3-3 isoforms, examining the tissue specificity of their
expression, and their age-related synthesis in potato plants. By
screening cDNA libraries from leaves, roots and epidermal
fragments, six cDNAs encoding 14-3-3 isoforms had been pre-
viously found in potato plants (Wilczynski et al. 1998).
To discover the significance of 14-3-3 isoforms for plant
metabolism, transgenic potatoes with repression of one, two
and six mRNAs were created. The selected transgenic plants
were analysed for enzyme activities known to interact with 14-
3-3 proteins, and metabolites, which are the products of the
interacting enzymes. A significant increase in NR, SPS and SS
activities was found for several of the analyzed enzymes (Zuk
et al. 2003). The changes in enzyme activity in transgenic
plants are reversible and can be complemented by heter-
ogonous recombinant 14-3-3 proteins either from Cucurbita
pepo or potato plants. It is interesting to note that the level of
all measured activity strongly depends on 14-3-3 protein con-
tent in the generated transgenic plants, thus suggesting that the
quantity of 14-3-3 protein rather than the particular isoform
controls the interacting enzymes. The questions now arise why
the genome of eukaryotes contains several 14-3-3 isoforms and
how the genes’ expression is regulated.
To try to answer these questions, the gene encoding the
16R isoform of 14-3-3 proteins has recently been isolated and
analysed (Szopa et al. 2003). Several motifs have been found
supporting the assumption that the promoter could be tissue-
specific and developmentally-regulated. Western and northern
analyses as well as the histochemical and fluorometric assays
* Corresponding author: E-mail, [email protected]; Fax, +48-71-3252930.
1635

14-3-3 gene promoter analysis1636
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
of various potato organs from 16R-uidA-transformed plants
revealed that the expression of the 16R isoform of 14-3-3
proteins is age-dependent. The highest expression was detected
in youngest organs or organ portions, leaves and stems in
particular. The analysis of GUS staining in transgenic plants
also showed the promoter tissue-specificity. Substantial GUS
expression was characteristic not only for meristems, but also
for the primary vascular tissues of minor vascular bundles. As
16R promoter activity seems to be induced by sucrose, 14-3-3
protein activity could be involved in the phloem loading. The
data obtained strongly suggests that 14-3-3 promoter investiga-
tion is the proper way to study the function of particular iso-
forms in cell biology.
In this study we first analysed by northern blot the expres-
sion pattern of mRNA which encodes the 20R isoform of 14-3-
3 protein in different plant organs and in leaves of different
ages. Then the genomic clone encoding 14-3-3 was isolated,
analysed in detail and compared to the database of accumu-
lated sequences. Finally, the promoter characteristic was stud-
ied in transgenic plants transformed with the reporter GUS
gene under the control of the 20R promoter and the data com-
pared to other 16R properties.
The promoters contain mostly the same motifs; however,
there are also several sequences that differentiate the genes’
activity. There is in 16R a unique motif activated by salicylic
acid and virus infection, which is absent in 20R. The latter
however contains unique sequences responsive to ethylene and
metal ions. This suggests that unique domains present in its
promoter distinctly regulate each isoform.
Results
14-3-3 gene organization
Screening of a genomic library with full length cDNA
encoding 20R isoform of 14-3-3 protein (EMBL/GenBank
Database Accession No. X87370), resulted in the isolation of a
genomic 3959 bp fragment [EMBL/GenBank database acc No.
AY518222] containing 1179 bp of the putative 5′ promoter
region, the complete coding sequence, and 246 bp of the 3′
noncoding element. There are four coding regions 1240–1716,
1973–2095, 2213–2329, 2423–2473 interrupted by three
introns. The transcription start site is 60 bp upstream of the
translation start codon. The amino acid sequence of 20R iso-
form is three residues shorter than that of the 16R isoform (Fig.
1), which has the highest homology (only 12 different amino
acids).
Analysis of promoter domains
The sequence analysis with the use of a Web Signal Scan
Program resulted in the identification of several core frag-
ments of great importance for promoter function. The 1239 bp
region upstream of the translation start site of 20R contains a
common CAAT box located at the –330 bp position. Inspec-
tion of downstream fragments reveals the expected TATA box
(TTATTT) at the –107 bp position.
Elements, which could potentially be recognised by tran-
scription factors, are good guidelines for design of experi-
ments aiming to reveal physiological processes in which a
given isoform could be involved. For example the AGAAA
motif (–1065, –217, –58, –49, –28) in the 20R promoter
sequence could be responsible for expression in pollen as was
shown for the late 52 gene in tomato (Bate and Twell 1998).
Analysis of putative targets for transcription factors is use-
ful for an experimental approach to test promoter activity. For
example, in both promoters (16 and 20R) eight E-box (CAN-
NTG) motifs have been found (Table 1). The E-box is recog-
nised as a Myc binding site that participates in ABA-induction
and binds a drought-inducible Myc homologue (Busk and
Pages 1998). The additional presence of Myb (–1113, –408)
and Myc (–233) binding sites in 20R promoter sequences, con-
comitant with the strong induction of these factors upon water
stress, may suggest the involvement of 14-3-3 protein synthe-
sis in the mechanism of ABA and water stress response of
potato plants.
In contrast to the protein sequence of 14-3-3 isoforms,
which is very conserved, their promoter sequences show sev-
eral important differences. For example, promoter 16R contains
two unique sequences TTGACC (–430) and TTWTWTTWTT
(–259). The first one is the core sequence of an elicitor respon-
sive element (ElRE), required for plant defence signalling
(Chen and Chen 2000). The second is a T-box found in a SAR/
MAR sequence. The presence of an E1RE sequence in a 16R
promoter might suggest the involvement of the 14-3-3 proteins
in plant defence mechanisms upon pathogen infection and sali-
cylic acid induction. The abundance of the 14-3-3 protein in
plant tissues may result from the presence of SAR/MAR
sequences in its promoter. It is suggested that this sequence is
responsible for chromatin attachment to the nuclear matrix and
genes within regions attached to this nuclear structure are
highly active.
Two very interesting promoter elements unique to 20R are
ethylene responsive at –1028 and metal responsive at –755.
The sequence AWTTCAAA was identified as an ethylene-
responsive region in the promoter of a fruit ripening gene and
as an enhancer element involved in the senescence-related
expression of the carnation and tomato glutathione-S-trans-
ferase gene (Hwang et al. 1998). Metals represent a class of
important transcription effector molecules which regulate gene
expression. The TGCRCNC sequence at –755 position in 20R
was earlier recognized as a metal-regulated element (MRE) of
the mouse metallothionein-I gene. Heavy metal ions effec-
tively induced metallothionein gene transcription and the
induction is MRE dependent. The transcription factor Sp1 and
a zinc-inducible factor MTF-1 bind to the MRE enhancer in
vitro (Thiele 1992).
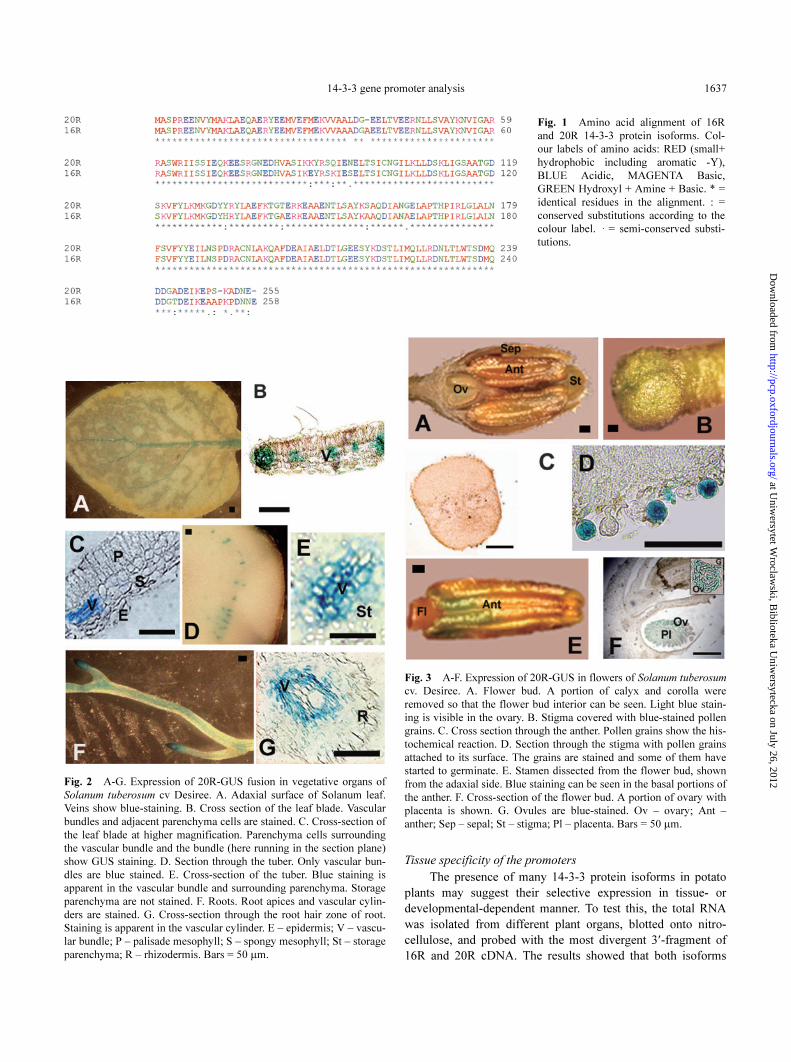
14-3-3 gene promoter analysis 1637
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
Tissue specificity of the promoters
The presence of many 14-3-3 protein isoforms in potato
plants may suggest their selective expression in tissue- or
developmental-dependent manner. To test this, the total RNA
was isolated from different plant organs, blotted onto nitro-
cellulose, and probed with the most divergent 3′-fragment of
16R and 20R cDNA. The results showed that both isoforms
Fig. 1 Amino acid alignment of 16R
and 20R 14-3-3 protein isoforms. Col-
our labels of amino acids: RED (small+
hydrophobic including aromatic -Y),
BLUE Acidic, MAGENTA Basic,
GREEN Hydroxyl + Amine + Basic. * =
identical residues in the alignment. : =
conserved substitutions according to the
colour label. · = semi-conserved substi-
tutions.
Fig. 2 A-G. Expression of 20R-GUS fusion in vegetative organs of
Solanum tuberosum cv Desiree. A. Adaxial surface of Solanum leaf.
Veins show blue-staining. B. Cross section of the leaf blade. Vascular
bundles and adjacent parenchyma cells are stained. C. Cross-section of
the leaf blade at higher magnification. Parenchyma cells surrounding
the vascular bundle and the bundle (here running in the section plane)
show GUS staining. D. Section through the tuber. Only vascular bun-
dles are blue stained. E. Cross-section of the tuber. Blue staining is
apparent in the vascular bundle and surrounding parenchyma. Storage
parenchyma are not stained. F. Roots. Root apices and vascular cylin-
ders are stained. G. Cross-section through the root hair zone of root.
Staining is apparent in the vascular cylinder. E – epidermis; V – vascu-
lar bundle; P – palisade mesophyll; S – spongy mesophyll; St – storage
parenchyma; R – rhizodermis. Bars = 50 µm.
Fig. 3 A-F. Expression of 20R-GUS in flowers of Solanum tuberosum
cv. Desiree. A. Flower bud. A portion of calyx and corolla were
removed so that the flower bud interior can be seen. Light blue stain-
ing is visible in the ovary. B. Stigma covered with blue-stained pollen
grains. C. Cross section through the anther. Pollen grains show the his-
tochemical reaction. D. Section through the stigma with pollen grains
attached to its surface. The grains are stained and some of them have
started to germinate. E. Stamen dissected from the flower bud, shown
from the adaxial side. Blue staining can be seen in the basal portions of
the anther. F. Cross-section of the flower bud. A portion of ovary with
placenta is shown. G. Ovules are blue-stained. Ov – ovary; Ant –
anther; Sep – sepal; St – stigma; Pl – placenta. Bars = 50 µm.

14-3-3 gene promoter analysis1638
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
encoded by 16R and 20R cDNAs are present in all the organs
analysed (Wilczynski et al. 1998).
Histochemical staining of transgenic potato plants trans-
formed with translational fusion of the 20R promoter with
GUS reporter protein revealed that tissue specificity is very
similar to the 16R isoform. In vegetative tissues 20R isoform
expression is localized mainly in the vascular tissue of young
organs (Fig. 2). In the leaf blade the strongest staining is lim-
ited to vascular bundles and adjacent parenchyma cells. In
tubers only vascular bundles and surrounding parenchyma are
blue, while storage parenchyma is not stained. In roots staining
is limited to vascular cylinders of root apices and the root hair
zone. In flowers (Fig. 3) GUS expression is limited to the gen-
erative organs: ovules and pollen grains which could result
from multiple AGAAA motifs.
Age-dependent analysis of 16R and 20R gene expression
Based on western analysis we have recently found that 14-
3-3 gene expression might be developmentally-regulated.
Indeed, northern blot analysis shows the highest quantity of
mRNA encoding 16R isoform in the middle stem sector. Anal-
ysis of consecutive leaves within one stem revealed the highest
mRNA level of 20R isoform in the youngest leaves. For com-
parison, the same blot when probed with 35G cDNA encoding
another 14-3-3 protein isoform showed that the expression of
this isoform was not age dependent (Wilczynski et al. 1998).
Table 1 Transcription factors binding sequences and tested factors on 14-3-3 promoters
Putative motifs recognised by transcription factors were found using Web Signal Scan Program.
+, induction of the promoter activity.
–, no effect on the promoter activity.
nd, conditions not analyzed.
TF 16R 20R
Sequence Effect Ref. Sequence Effect Ref.
ABA 1. ACGTG + 1. (Simpson et al. 2003) 1. ACGTC + 1. (Simpson et al. 2003)
2. ACGT 2. (Simpson et al. 2003) 2. ACGT 2. (Simpson et al. 2003)
3. CANNTG 3. (Stalberg et al. 1996) 3. CANNTG 3. (Stalberg et al. 1996)
4. WAACCA 4. (Abe et al. 2003) 4. WAACCA 4. (Abe et al. 2003)
5. TAACTG 5. (Urao et al. 1993) 5. YAACKG 5. (Abe et al. 2003)
6. CACATG 6. (Abe et al. 1997) 6. CTAACCA 6. (Busk and Pages 1998)
7. ACACNNG 7. (Kim et al. 1997)
Ethylene Not found – Not found 1. AWTTCAAA – 1. (Itzhaki et al. 1994)
Auxin 1. TGTCTC + 1. (Hagen and Guilfoyle 2002) 1. CATATG – 1. (Xu et al. 1997)
2. ACTTTA 2. (Baumann et al. 1999) 2. ACTTTA 2. (Baumann et al. 1999)
3. TGACG 3. (Terzaghi and Cashmore 1995)
Saliylic acid 1. TTGACC + 1. (Chen and Chen 2000) 1. TGACG – 1. (Redman et al. 2002)
Pathogen 1. GAAAAA + 1. (Park et al. 2004) 1. GAAAAA – 1. (Park et al. 2004)
2. TTGACC 2. (Chen and Chen 2000)
Metals Not found nd Not found 1. TGCRCNC + 1. (Wang et al. 2004)
NaCl 1. GAAAAA nd 1. (Park et al. 2004) 1. GAAAAA + 1. (Park et al. 2004)
Jasmonic acid 1. AACGTG – 1. (Boter et al. 2004) 1. AACGTG – 1. (Boter et al. 2004)
Light 1. GATA – 1. (Reyes et al. 2004) 1. GATA – 1. (Lam and Chua 1989)
2. GATAA 2. (Terzaghi and Cashmore 1995) 2. GATAA 2. (Terzaghi and Cashmore 1995)
3. GRWAAW 3. (Zhou 1999) 3. GRWAAW 3. (Terzaghi and Cashmore 1995)
4. GATAAG 4. (Giuliano et al. 1988) 4. TATTCT 4. (Thum et al. 2001)
5. CTCCCAC 5. (Ngai et al. 1997)
Hurt 1. AGATCCAA – 1. (Sugimoto et al. 2003) Not found – Not found
Low temp. 1. CANNTG – 1. (Chinnusamy et al. 2003) 1. CANNTG + 1. (Chinnusamy et al. 2003)
Gibberelic acid Not found – Not found 1. TAACAAR – 1. (Ogawa et al. 2003)
2. TAACAAA 2. (Gubler et al. 1995)
3. TGAC 3. (Zhang et al. 2004)
Sucrose AATAGAAAA + 1. (Grierson et al. 1994) 1. TAACARA – 1. (Huang et al. 1990)
2. TATCCAT 2. (Hwang et al. 1998)
3. TGACT 3. (Sun et al. 2003)
4. TATCCA 4. (Lu et al. 2002)
5. TATCCAY 5. (Toyofuku et al. 1998)
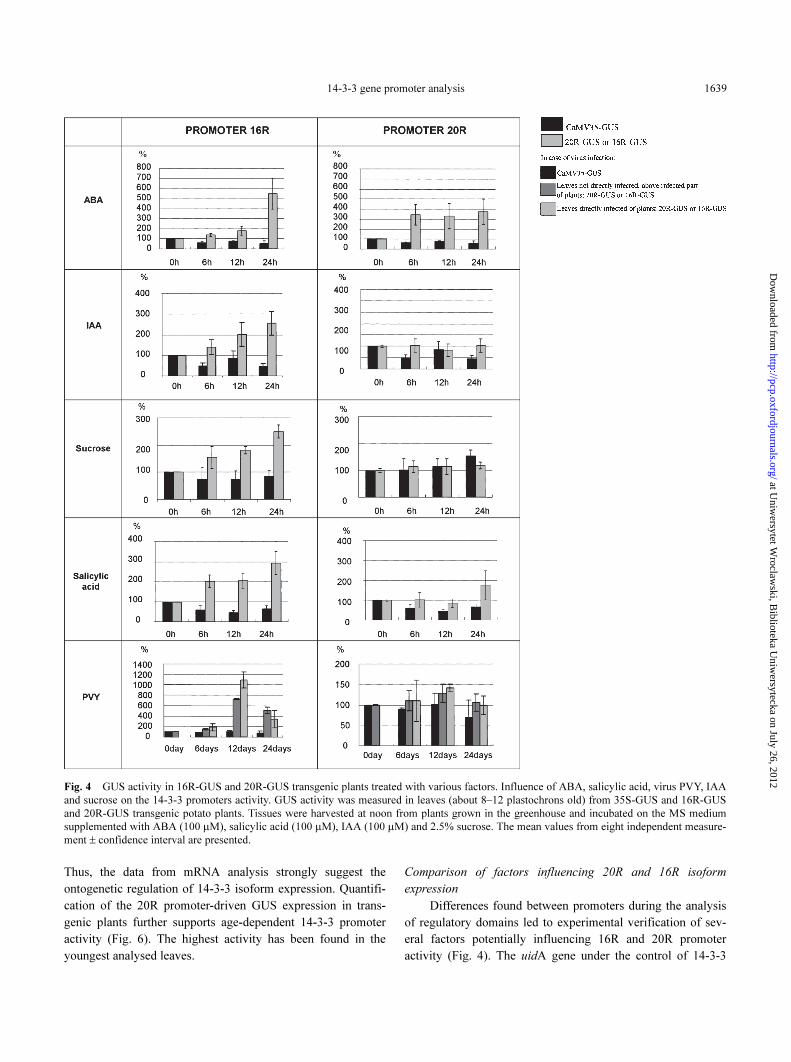
14-3-3 gene promoter analysis 1639
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
Thus, the data from mRNA analysis strongly suggest the
ontogenetic regulation of 14-3-3 isoform expression. Quantifi-
cation of the 20R promoter-driven GUS expression in trans-
genic plants further supports age-dependent 14-3-3 promoter
activity (Fig. 6). The highest activity has been found in the
youngest analysed leaves.
Comparison of factors influencing 20R and 16R isoform
expression
Differences found between promoters during the analysis
of regulatory domains led to experimental verification of sev-
eral factors potentially influencing 16R and 20R promoter
activity (Fig. 4). The uidA gene under the control of 14-3-3
Fig. 4 GUS activity in 16R-GUS and 20R-GUS transgenic plants treated with various factors. Influence of ABA, salicylic acid, virus PVY, IAA
and sucrose on the 14-3-3 promoters activity. GUS activity was measured in leaves (about 8–12 plastochrons old) from 35S-GUS and 16R-GUS
and 20R-GUS transgenic potato plants. Tissues were harvested at noon from plants grown in the greenhouse and incubated on the MS medium
supplemented with ABA (100 µM), salicylic acid (100 µM), IAA (100 µM) and 2.5% sucrose. The mean values from eight independent measure-
ment ± confidence interval are presented.
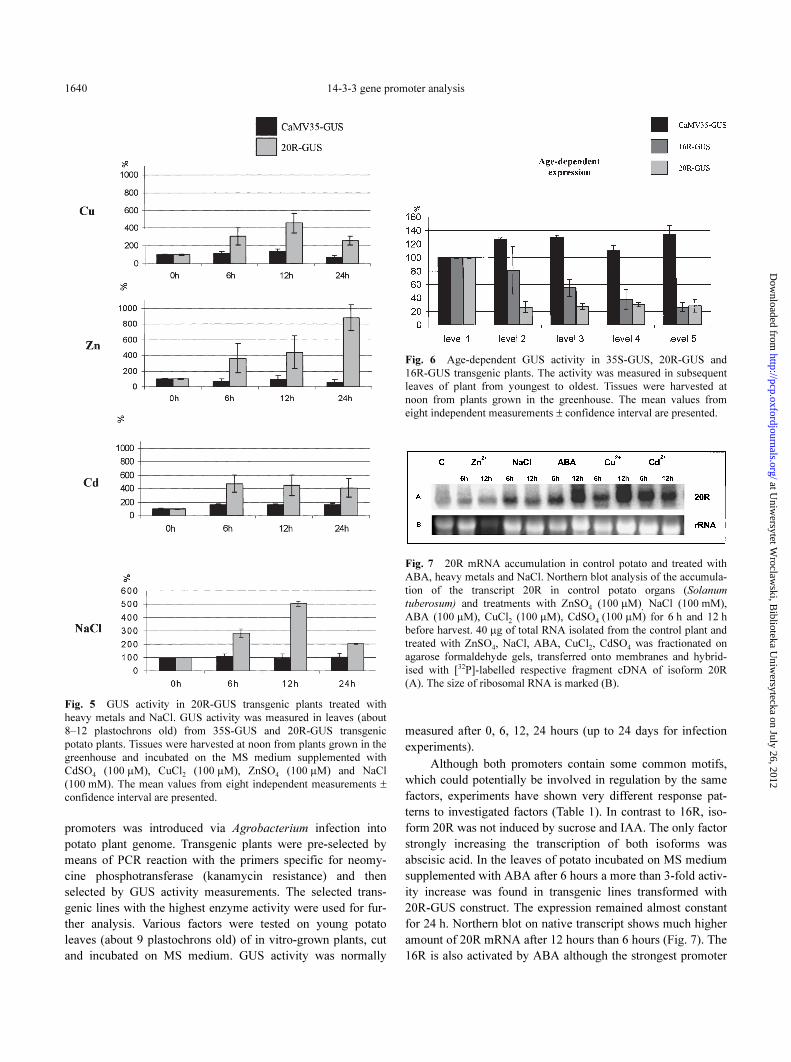
14-3-3 gene promoter analysis1640
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
promoters was introduced via Agrobacterium infection into
potato plant genome. Transgenic plants were pre-selected by
means of PCR reaction with the primers specific for neomy-
cine phosphotransferase (kanamycin resistance) and then
selected by GUS activity measurements. The selected trans-
genic lines with the highest enzyme activity were used for fur-
ther analysis. Various factors were tested on young potato
leaves (about 9 plastochrons old) of in vitro-grown plants, cut
and incubated on MS medium. GUS activity was normally
measured after 0, 6, 12, 24 hours (up to 24 days for infection
experiments).
Although both promoters contain some common motifs,
which could potentially be involved in regulation by the same
factors, experiments have shown very different response pat-
terns to investigated factors (Table 1). In contrast to 16R, iso-
form 20R was not induced by sucrose and IAA. The only factor
strongly increasing the transcription of both isoforms was
abscisic acid. In the leaves of potato incubated on MS medium
supplemented with ABA after 6 hours a more than 3-fold activ-
ity increase was found in transgenic lines transformed with
20R-GUS construct. The expression remained almost constant
for 24 h. Northern blot on native transcript shows much higher
amount of 20R mRNA after 12 hours than 6 hours (Fig. 7). The
16R is also activated by ABA although the strongest promoter
Fig. 5 GUS activity in 20R-GUS transgenic plants treated with
heavy metals and NaCl. GUS activity was measured in leaves (about
8–12 plastochrons old) from 35S-GUS and 20R-GUS transgenic
potato plants. Tissues were harvested at noon from plants grown in the
greenhouse and incubated on the MS medium supplemented with
CdSO4 (100 µM), CuCl2 (100 µM), ZnSO4 (100 µM) and NaCl
(100 mM). The mean values from eight independent measurements ±
confidence interval are presented.
Fig. 6 Age-dependent GUS activity in 35S-GUS, 20R-GUS and
16R-GUS transgenic plants. The activity was measured in subsequent
leaves of plant from youngest to oldest. Tissues were harvested at
noon from plants grown in the greenhouse. The mean values from
eight independent measurements ± confidence interval are presented.
Fig. 7 20R mRNA accumulation in control potato and treated with
ABA, heavy metals and NaCl. Northern blot analysis of the accumula-
tion of the transcript 20R in control potato organs (Solanum
tuberosum) and treatments with ZnSO4 (100 µM), NaCl (100 mM),
ABA (100 µM), CuCl2 (100 µM), CdSO4 (100 µM) for 6 h and 12 h
before harvest. 40 µg of total RNA isolated from the control plant and
treated with ZnSO4, NaCl, ABA, CuCl2, CdSO4 was fractionated on
agarose formaldehyde gels, transferred onto membranes and hybrid-
ised with [32P]-labelled respective fragment cDNA of isoform 20R
(A). The size of ribosomal RNA is marked (B).
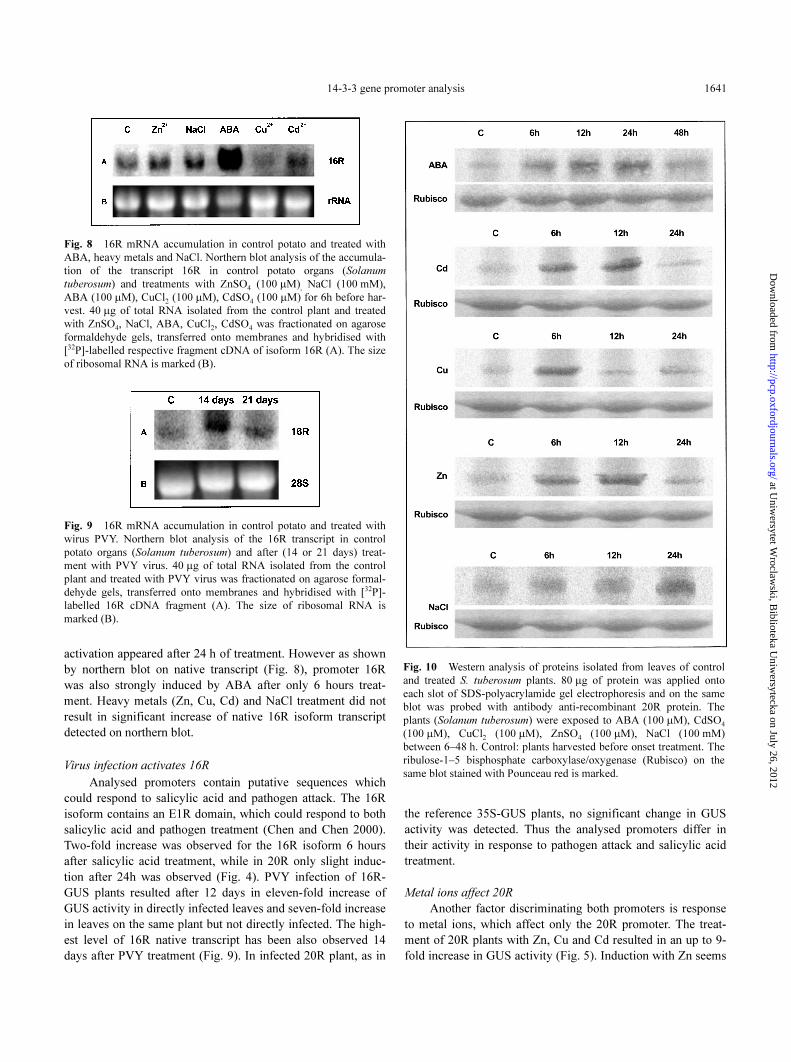
14-3-3 gene promoter analysis 1641
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
activation appeared after 24 h of treatment. However as shown
by northern blot on native transcript (Fig. 8), promoter 16R
was also strongly induced by ABA after only 6 hours treat-
ment. Heavy metals (Zn, Cu, Cd) and NaCl treatment did not
result in significant increase of native 16R isoform transcript
detected on northern blot.
Virus infection activates 16R
Analysed promoters contain putative sequences which
could respond to salicylic acid and pathogen attack. The 16R
isoform contains an E1R domain, which could respond to both
salicylic acid and pathogen treatment (Chen and Chen 2000).
Two-fold increase was observed for the 16R isoform 6 hours
after salicylic acid treatment, while in 20R only slight induc-
tion after 24h was observed (Fig. 4). PVY infection of 16R-
GUS plants resulted after 12 days in eleven-fold increase of
GUS activity in directly infected leaves and seven-fold increase
in leaves on the same plant but not directly infected. The high-
est level of 16R native transcript has been also observed 14
days after PVY treatment (Fig. 9). In infected 20R plant, as in
the reference 35S-GUS plants, no significant change in GUS
activity was detected. Thus the analysed promoters differ in
their activity in response to pathogen attack and salicylic acid
treatment.
Metal ions affect 20R
Another factor discriminating both promoters is response
to metal ions, which affect only the 20R promoter. The treat-
ment of 20R plants with Zn, Cu and Cd resulted in an up to 9-
fold increase in GUS activity (Fig. 5). Induction with Zn seems
Fig. 8 16R mRNA accumulation in control potato and treated with
ABA, heavy metals and NaCl. Northern blot analysis of the accumula-
tion of the transcript 16R in control potato organs (Solanum
tuberosum) and treatments with ZnSO4 (100 µM), NaCl (100 mM),
ABA (100 µM), CuCl2 (100 µM), CdSO4 (100 µM) for 6h before har-
vest. 40 µg of total RNA isolated from the control plant and treated
with ZnSO4, NaCl, ABA, CuCl2, CdSO4 was fractionated on agarose
formaldehyde gels, transferred onto membranes and hybridised with
[32P]-labelled respective fragment cDNA of isoform 16R (A). The size
of ribosomal RNA is marked (B).
Fig. 9 16R mRNA accumulation in control potato and treated with
wirus PVY. Northern blot analysis of the 16R transcript in control
potato organs (Solanum tuberosum) and after (14 or 21 days) treat-
ment with PVY virus. 40 µg of total RNA isolated from the control
plant and treated with PVY virus was fractionated on agarose formal-
dehyde gels, transferred onto membranes and hybridised with [32P]-
labelled 16R cDNA fragment (A). The size of ribosomal RNA is
marked (B).
Fig. 10 Western analysis of proteins isolated from leaves of control
and treated S. tuberosum plants. 80 µg of protein was applied onto
each slot of SDS-polyacrylamide gel electrophoresis and on the same
blot was probed with antibody anti-recombinant 20R protein. The
plants (Solanum tuberosum) were exposed to ABA (100 µM), CdSO4
(100 µM), CuCl2 (100 µM), ZnSO4 (100 µM), NaCl (100 mM)
between 6–48 h. Control: plants harvested before onset treatment. The
ribulose-1–5 bisphosphate carboxylase/oxygenase (Rubisco) on the
same blot stained with Pounceau red is marked.

14-3-3 gene promoter analysis1642
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
to be slower and longer lasting than other metal ions. The acti-
vation of 14-3-3 20R isoform by ABA, metal ions and NaCl,
was also confirmed by northern (Fig. 7) and western (Fig. 10)
analysis on wild-type not transformed Desiree potato plants. In
all cases a significant increase in 14-3-3 20R mRNA and pro-
tein level was detected (Fig 7, 10) after treatment with ABA,
NaCl and heavy metals. The correlation coefficient between
induction observed on northern and western blots is in most
cases very high (0.98 for ABA, 0.99 for Cd and 0.84 for Zn).
Lower correlation for Cu results from maximal induction time
(6 h on western and 12 h on northern blot). Significant activa-
tion of nitrate reductase (NR) in transgenic potato plants with
repression of six 14-3-3 protein isoformes (G3 plants) has been
previously shown (Zuk et al. 2003). This activity could be
restored at control level by addition of recombinant 14-3-3 pro-
tein. Thus, to further study in vivo 14-3-3 expression induced
by heavy metals we have analysed NR activity in control and
G3 plants treated with 100 µM metal ions solution (Fig. 11).
Treatment with metal ions resulted in decreased NR activity,
both in control and G3 plants. This further supports hypothesis
that 14-3-3 protein 20R isoform is upregulated by metal ions.
Apart ABA, all tested factors induced GUS activity in a
promoter dependent manner and the results suggest the spe-
cific appearance of particular isoforms in response to environ-
mental stimuli. While isoform 20R is specifically induced by
metal ions, isoform 16R is induced in response to pathogen
attack salicylic acid treatment, sucrose, ABA and auxin.
Discussion
14-3-3 proteins are abundant eukaryotic proteins that
interact with many other proteins, thereby modulating their
function, and thus modulating cell metabolism (Chung et al.
1999). While there has been substantial progress in the identifi-
cation of the diverse partners of 14-3-3 in recent years their
role in regulatory pathways remains to be elucidated. Promoter
studies could provide helpful guidelines how to investigate the
specificity of 14-3-3 isoforms. To answer this question, investi-
gations were made on the two 14-3-3 gene promoters. It was
previously reported that plant and human 14-3-3s could be
replaced by yeast isoforms (van Heusden et al. 1992). Such a
result was not surprising since there is a very high homology
within 14-3-3 isoforms. Our own data also suggests that at the
metabolic level each potato isoform play the same role. The
repression of any isoform or all of them in potato transgenic
plants resulted in a significant increase in NR, SPS and SS
activities and thus affected carbohydrate and amino acids
metabolism (Szopa et al. 2001, Swiedrych et al. 2002, Zuk et
al. 2003). Thus the question arises why the eukaryotic genome
accumulated several genes coding for 14-3-3 isoforms? The
data presented in this paper demonstrates clearly for the first
time that promoters specify 14-3-3 genes expression. Two iso-
lated promoters 16R and 20R differ significantly when consid-
ering their response to environmental condition. The most
important differences are 16R activation by salicylic acid and
virus infection and activation of 20R by metal ions. It was ear-
lier demonstrated that an increase in 14-3-3 protein synthesis
results from pathogen treatment of the plant (Brandt et al.
1992) and now it is clear that it is due to the presence of the
specific domain in the isoform promoter. The increase in 14-3-
3 content resulted in NR and SPS inhibition, which subse-
quently redirected metabolism to the synthesis of other com-
pounds.
Thus we conclude that for 14-3-3 genes, the main differ-
ence is the promoter region containing specific inducible
domains, and not the coding region responsible for enzymatic
specificity. Recently we have reported on phenolic compound
synthesis affected by 14-3-3 protein. Overexpression of 14-3-3
cDNA in potato resulted in an increase of phenolic acid synthe-
sis, and a reduction of 14-3-3 content showed the opposite
effect (Lukaszewicz et al. 2002). Since the 14-3-3 genes pro-
moter contains a P box (Davies and Schwinn 2003) also char-
acteristic for genes coding key enzymes (CHS, CHI, DFR) of
flavonoid biosynthesis, it might suggest that in vivo both types
of genes are regulated in parallel. In this work we show that
expression of one 20R isofom may be restricted to very spe-
cific tissue like pollen and could be specifically stimulated by
heavy metals and NaCl. It is suggested that similar genes
respond to various environmental cues and stresses (Cooper et
al. 2003). Our results of promoter studies suggest that 14-3-3
protein could be such universal stress signal regulation mole-
cule. The need for the precise regulation of the response to var-
ious environmental cues could explain existence of several
isoforms with promoters dedicated to given stimulus set. We
have shown that 16R 14-3-3protein isoform could be special-
ised in response to pathogen attack and 20R isoform in
response to metal ions. In vivo we have shown that metal ions
affected NR activity. Other putative target in response to heavy
metal ions could be plasma membrane ATPase which is sup-
posed to affect plasma membranes fluidity and active transport
through membranes. These findings could be helpful in further
Fig. 11 NR activity in leaves of plants treated with heavy metals.
Leaves of Solanum tuberosum Desiree plants-control (D) and trans-
genic plants with repression of six 14-3-3 protein isoforms (G3) were
exposed to heavy metals (100 µM) between 6–24 h. Third and fourth
leaf from the top of the plant were frozen in liquid nitrogen and
extracted as described in materials and methods, then analysed and
compared to the control (D). Results are the mean ± SE (n = 6 different
plants from G3 transgenic line).

14-3-3 gene promoter analysis 1643
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
deciphering the regulation of metabolic pathways by 14-3-3
proteins.
Materials and Methods
Plant material and bacterial strains
Potato plants (Solanum tuberosum L. cv. Desiree) were obtained
from “Saatzucht Fritz Lange KG” (Bad Schwartau, FRG). Plants in tis-
sue culture were grown under a 16 h light – 8 h dark regime on MS
medium (Murashige and Skoog 1962) containing 0.8% sucrose. Plants
in the greenhouse were cultivated in soil under 16-h light (22°)/8-h
dark (15°C) regime. Plants were grown in individual pots and were
watered daily.
Escherichia coli strain DH5a (Bethesda Research Laboratories,
Gaithersburg, USA) was cultivated using standard techniques
(Sambrook et al. 1989). Agrobacterium tumefaciens strain C58C1 con-
taining plasmid pGV2260 (Deblaere et al. 1985) was cultivated in
YEB medium (Vervliet et al. 1975).
Recombinant DNA techniques
DNA manipulations were done essentially as described by
Sambrook et al. (1989). DNA restriction and modification enzymes
were obtained from Roche (Germany) and New England Biolabs
(Beverly, MA). Escherichia coli strains DH5a and XL1-Blue were
used.
Construction of a 20R promoter GUS fusion
The 1084 bp SmaI–Asp718 fragment of the 5′ promoter region of
the 20R gene [EMBL/GenBank database acc No. AY518222.], from –
1239 to +1 (translation start site), was inserted in frame into the
BamHI/SalI site of pBI101 (Clontech), in front of the E. coli GUS
gene and the nos polyadenylation signal. The obtained vector, contain-
ing the 20R-GUS fusion, was introduced into the Agrobacterium
tumefaciens strain C58C1:pGV2260 as previously described (Rocha-
Sosa et al. 1989).
Transformation of potato plants
Young leaves of wild-type potato Solanum tuberosum L. cv.
Desiree were used for the transformation with A. tumefaciens by
immersing the leaf explants in the bacterial suspension. A.
tumefaciens-inoculated leaf explants were subsequently transferred
onto the callus induction and shoot regeneration medium (Rocha-Sosa
et al. 1989). Transgenic potato plants were screened by means of PCR
reaction with the use of the 20R promoter and kanamycin specific
primers as previously described (Wilczynski et al. 1998, Szopa et al.
2001).
Northern blot analysis
The expression of 20R and 16R 14-3-3 isoforms in potato plant
organs and developing leaves were performed by means of Northern
blot analysis. Total RNA was prepared from frozen plant material
using the guanidinium hydrochloride method. Following electro-
phoresis (1.5% [w/v] agarose, 15% [v/v] formaldehyde) RNA was
transferred to nylon membranes (Hybond N, Amersham, UK). The
membranes were hybridised overnight at 42°C in a 250 mmol sodium
phosphate buffer (pH7.2) containing 7% [w/v] SDS, 1% [w/v] bovine
serum albumin (BSA) and 1 mmol EDTA. A radioactively-labelled
hybridization probe of 16R (BamHI/PstI cDNA fragment) or 20R
(KpnI/PstI cDNA fragment) was used. The filters were washed three
times in 2× SSPE +0.1% [w/v] SDS for 10 min at 42°C; three times in
1× SSPE +0.1% [w/v] SDS for 10 min at 42°C; and three times in 0.1×
SSPE +0.1% [w/v] SDS for 10 min at 42°C.
Western blot analysis
An assessment of the expression of the 14-3-3 gene was
conducted by means of western blot analysis using the rabbit IgG
anti-recombinant 20R protein as described previously (Szopa and Rose
1986). Briefly, a solubilised protein was run on 12% SDS–poly-
acrylamide gels and blotted electrophoretically onto nitrocellulose
membranes (Schleicher and Schuell). After the transfer, the membrane
was sequentially incubated with a blocking buffer (5% dry milk)
and then with antibodies directed against the 20R recombinant protein
(1 : 2,000 dilution). The formation and detection of immune com-
plexes was performed as previously described (Szopa and Rose 1986).
Alkaline phosphatase-conjugated goat anti-rabbit IgG served as a
second antibody, and was used at a dilution of 1 : 1,500.
Histochemical analysis of the 20R promoter GUS fusion
Histochemical staining for GUS activity was performed with the
use of 5-bromo-4-chloro-3-indolyl β-D-glucuronide as a substrate
(Jefferson et al. 1987). Freshly-cut plant tissues were incubated in
GUS staining solution at 37°C, for 2–24 h depending on staining
intensity, and bleached with 70% ethanol. The material was examined
with a stereomicroscope Olympus SZX9, and anatomical sections
were prepared. The specimens were dehydrated and embedded in par-
affin (Johansen 1940). Minimal times were used for the treatment with
buthanol and buthanol–paraffin mixtures, in which the specimens were
kept in higher temperatures (60°C). Microtome paraffin sections, 25–
30 µm thick, were rinsed in xylene and ethanol for a short time.
Euparal was used as a mounting medium. The sectioned material was
examined using bright-field illumination microscopy Olympus BM50.
Fluorometric GUS assay
Potato transgenic lines with strong histochemical GUS staining
in leaves and stems were used for glucuronidase assay. The samples
were extracted with 50 mM Tris buffer, pH 8.0 containing 10 mM β-
mercaptoethanol, 10 mM EDTA, and centrifuged for 10 min at
13,000 rpm (Sigma Instrument). Aliquots of the supernatant were used
for enzyme assay with 4-methylumbelliferyl β-D-glucuronide as a sub-
strate (Jefferson 1987) and for protein determination with Bradford
reagent (Bradford 1976). The reaction product 4-methylumbelliferone
was measured fluorometrically (SFM 25 Fluorescence Spectrophotom-
eter, Kontron Instrument, Hamburg, Germany).
Treatment conditions
GUS activity was measured in leaves (about 8–12 plastochrons
old) harvested from plants grown in the greenhouse and incubated on
the MS medium supplemented with 2.5% sucrose, 100 µM ABA or
100 mM NaCl, 100 nM salicylic acid, 100 µM IAA, 100 µM CdSO4,
100 µM CuCl2, 100 µM ZnSO4. For virus infection entire greenhouse
plants were used.
Acknowledgements
The authors wish to thank Dr. G. Wilczynski for initial work on
14-3-3 gene isolation. This work is supported by grants No 2P06A 022
27 and PBZ-KBN-089/P06/2003 from National Research Committee
(KBN) and EC HPRN-CT-2002-00269.
References
Abe, H., Urao, T., Ito, T., Seki, M., Shinozaki, K. and Yamaguchi-Shinozaki,
R.A.K. (2003) Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function
as transcriptional activators in abscisic acid signalling. Plant Cell 15: 63–78.
Abe, H., Yamaguchi-Shinozaki, K., Urao, T., Iwasaki, T., Hosokawa, D.R.A.
and Shinozaki, K. (1997) Role of Arabidopsis MYC and MYB homologs in

14-3-3 gene promoter analysis1644
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
drought- and abscisic acid-regulated gene expression. Plant Cell 9: 1859–
1868.
Bate, N. and Twell, D. (1998) Functional architecture of a late pollen promoter:
pollen specific transcription is developmentally regulated by multiple stage-
specific and co-dependent activator elements. Plant Mol. Biol. 37: 859–869.
Baumann, K., De Paolis, A., Costantino, P. and Gualberti, G. (1999) The DNA
binding site of the Dof protein NtBBF1 is essential for tissue-specific and
auxin-regulated expression of the rolB oncogene in plants. Plant Cell 11:
323–333.
Boter, M., Ruiz-Rivero, O., Abdeen, A. and Prat, S. (2004) Conserved MYC
transcription factors play a key role in jasmonate signalling both in tomato
and Arabidopsis. Genes Dev. 18: 1577–1591.
Bradford, M. (1976) A rapid and sensitive method for the quantitation of micro-
gram quantities of protein utilizing the principle of protein-dye binding. Anal.
Biochem. 72: 248–254.
Brandt, J., Thordal-Christensen, H., Vad, K., Gregersen, P.L. and Collinge, D.B.
(1992) A pathogen – induced gene of barley encodes a protein showing high
similarity to a protein kinase regulator. Plant J. 2: 815–820.
Busk, P.K. and Pages, M. (1998) Regulation of abscisic-induced transcription.
Plant Mol. Biol. 37: 425–435.
Chen, C. and Chen, Z. (2000) Isolation and characterization of two pathogen-
and salicylic acid-induced genes encoding WRKY DNA-binding proteins
from tobacco. Plant Mol. Biol. 42: 387–396.
Chinnusamy, V., Ohta, M., Kanrar, S., Lee, B.H., Hong, X., Agarwal, M. and
Zhu, J.K. (2003) ICE1: a regulator of cold-induced transcriptome and freez-
ing tolerance in Arabidopsis. Genes Dev. 17: 1043–1054.
Chung, H.J., Sehnke, P.C. and Ferl, R.J. (1999) The 14-3-3 proteins: cellular
regulators of plant metabolism. Trends Plant Sci. 4: 367–371.
Cooper, B., Clarke, J.D., Budworth, P. Kreps, J., Hutchison, D., Park, S.,
Guimil, S., Dunn, M., Luginbuhl, P., Ellero, C., Goff, S.A. and Glazebrook, J.
(2003) A network of rice genes associated with stress response and seed
development. Proc. Natl Acad. Sci. USA 100: 4945–4950.
Davies, K.M. and Schwinn, K.E. (2003) Transcriptional regulation of second-
ary metabolism. Funct. Plant Biol. 30: 913–925
Deblaere, R., Bytebier, B., de Greve, H., Deboeck, F., Schell, J., Van Montagu,
M. and Leemans, J. (1985) Efficient octopine Ti plasmid-derived vectors for
Agrobacterium-mediated gene transfer to plants. Nucleic Acids Res. 13:
4777–4788.
Finnie, C., Borch, J. and Collinge, D.B. (1999) 14-3-3 proteins: eukaryotic regu-
latory proteins with many functions. Plant Mol. Biol. 40: 545–554.
Giuliano, G., Pichersky, E., Malik, V.S., Timko, M.P., Scolnik, P.A. and
Cashmore, R.A. (1988) An evolutionarily conserved protein binding
sequence upstream of a plant light-regulated gene. Proc. Natl Acad. Sci. USA
85: 7089–7093.
Grierson, C., Du, J.S., Zabala, M.T., Beggs, K., Smith, C., Holdsworth, M.R.A.
and Bevan, M. (1994) Separate cis sequences and trans factors direct meta-
bolic and developmental regulation of a potato tuber storage protein gene.
Plant J. 5: 815–826.
Gubler, F., Kalla, R., Roberts, J.K. and Jacobsen, J.V. (1995) Gibberellin-regu-
lated expression of a myb gene in barley aleurone cells: evidence for Myb
transactivation of a high-pl alpha-amylase gene promoter. Plant Cell 7: 1879–
1891.
Hagen, G. and Guilfoyle, T. (2002) Auxin-responsive gene expression: genes,
promoters and regulatory factors. Plant Mol. Biol. 49: 373–385.
Huang, N., Sutliff, T.D., Litts, J.C. and Rodriguez, R.L. (1990) Classification
and characterization of the rice alpha-amylase multigene family. Plant Mol.
Biol. 14: 655–668.
Hwang, Y.S., Karrer, E.E., Thomas, B.R., Chen, L. and Rodriguez, R.L. (1998)
Three cis-elements required for rice alpha-amylase Amy3D expression during
sugar starvation. Plant Mol. Biol. 36: 331–341.
Itzhaki, H., Maxson, J.M. and Woodson, W.R. (1994) An ethylene-responsive
enhancer element is involved in the senescence-related expression of the car-
nation glutathione-S-transferase (GSTI) gene. Proc. Natl Acad. Sci. USA 91:
8925–8929.
Jefferson, R.A. (1987) Assaying chimeric genes in plants: The GUS gene fusion
system. Plant Mol. Biol. 5: 387–405.
Jefferson, R.A., Kavanagh, T.A. and Bevan, M.W. (1987) GUS fusions: β-
glucuronidase as a sensitive and versatile gene fusion marker in higher plants.
EMBO J. 6: 3901–3907.
Johansen, D.A. (1940) Plant microtechnique. McGraw-Hill New York.
Kim, S.Y., Chung, H.J. and Thomas, T.L. (1997) Isolation of a novel class of
bZIP transcription factors that interact with ABA-responsive and embryo-
specification elements in the Dc3 promoter using a modified yeast one-hybrid
system. Plant J. 11: 1237–1251.
Lam, E. and Chua, N.H. (1989) ASF-2: A factor that binds to the cauliflower
mosaic virus 35S promoter and a conserved GATA motif in cab promoters.
Plant Cell 1: 1147–1156.
Lu, C.A., Ho, T.H.D., Ho, S.L. and Yu, S.M. (2002) Three novel MYB proteins
with one DNA binding repeat mediate sugar and hormone regulation of
alpha-amylase gene expression. Plant Cell 14: 1963–1980.
Lukaszewicz, M., Matysiak-Kata, I., Aksamit, A., Oszmianski, J. and Szopa, J.
(2002) 14-3-3 Protein regulation of the antioxidant capacity of transgenic
potato tubers. Plant Sci. 163: 125–130.
Murashige, T. and Skoog, F. (1962) A revised medium for rapid growth and bio-
assays with tobacco tissue culture. Physiol. Plant. 51: 493–497.
Ngai, N., Tsai, F.Y. and Coruzzi, G. (1997) Light-induced transcriptional repres-
sion of the pea AS1 gene: identification of cis-elements and transfactors.
Plant J. 12: 1021–1034.
Ogawa, M., Hanada, A., Yamauchi, Y., Kuwahara, A., Kamiya, Y. and
Yamaguchi, R.A.S. (2003) Gibberellin biosynthesis and response during
Arabidopsis seed germination. Plant Cell 15: 1591–1604.
Park, H.C., Kim, M.L., Kang, Y.H., Jeon, J.M., Yoo, J.H., et al. (2004) Patho-
gen- and NaCl-induced expression of the SCaM-4 promoter is mediated in
part by a GT-1 box that interacts with a GT-1-like transcription factor. Plant
Physiol. 135: 2150–2161.
Redman, J., Whiteraft, J., Jhonson, C. and Arias, J. (2002) Abiotic and biotic
stress differentially stimulate as-1 element activity in Arabidopsis. Plant Cell
Rep. 21: 180–185.
Reyes, J.C., Muro-Pastor, M.I., Florencio, F.J. (2004) The GATA family of
transcription factors in Arabidopsis and rice. Plant Physiol. 134: 1718–1732.
Roberts, M.R. (2003) 14-3-3 proteins find new partners in plant cell signalling.
Trends Plant Sci. 8: 218–223.
Rocha-Sosa, M., Sonnewald, U., Frommer, W., Stratmann, M., Schell, J. and
Willmitzer, L. (1989) Both developmental and metabolic signals activate the
promoter of a class I patatin gene. EMBO J. 8: 23–29.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) In Molecular Cloning: A
Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold
Spring Harbor, NY
Simpson, S.D., Nakashima, K., Narusaka, Y., Seki, M., Shinozaki, K. and
Yamaguchi-Shinozaki, K. (2003) Two different novel cis-acting elements of
erd1, a clpA homologous Arabidopsis gene function in induction by dehydra-
tion stress and dark-induced senescence. Plant J. 33: 259–270.
Stalberg, K., Ellerstom M., Ezcurra I., Ablov, S. and Rask, L. (1996) Disruption
of an overlapping E-box/ABRE motif abolished high transcription of the
napA storage-protein promoter in transgenic Brassica napus seeds. Planta
199: 515–519.
Sugimoto, K., Takeda, S. and Hirochika, H. (2003) Transcriptional activation
mediated by binding of a plant GATA-type zinc finger protein AGP1 to the
AG-motif (AGATCCAA) of the wound-inducible Myb gene NtMyb2. Plant
J. 36: 550–564.
Sun, C., Palmqvist, S., Olsson, H., Boren, M., Ahlandsberg, S. and Jansson, C.
(2003) A novel WRKY transcription factor, SUSIBA2, participates in sugar
signalling in barley by binding to the sugar-responsive elements of the iso1
promoter. Plant Cell 15: 2076–2092.
Szopa, J. (2002) Transgenic 14-3-3 isoforms in plants: the metabolite profiling
of repressed 14-3-3 protein synthesis in transgenic potato plants. Biochem.
Soc. Trans 30: 405–410.
Szopa, J., Lukaszewicz, M., Korobczak, A., Aksamit, A. and Kwiatkowska, D.
(2003) Structural organisation, expression, and promoter analysis of a 16R
isoform of 14-3-3 protein gene from potato. Plant Physiol. Biochem. 41: 417–
423.
Szopa, J. and Rose, K.M. (1986) Cleavage of the 190 kDa subunit of DNA-
dependent RNA polymerase I yields small polypeptides capable of degrading
DNA. J. Biol. Chem. 261: 9022–9028.
Szopa, J., Wróbel, M., Matysiak-Kata, I. and Swiedrych, A. (2001) The meta-
bolic profile of the 14-3-3 repressed transgenic potato tubers. Plant Sci. 161:
1075–1082
Swiedrych, A., Prescha. A., Matysiak-Kata, I., Biernat, J. and Szopa, J. (2002)
Repression of the 14-3-3 gene affects the amino acid and mineral composi-
tion of potato tubers. J. Agric. Food Chem. 50: 2137–2141.

14-3-3 gene promoter analysis 1645
at Uniw
ersytet Wroclaw
ski, Biblioteka U
niwersytecka on July 26, 2012
http://pcp.oxfordjournals.org/D
ownloaded from
Terzaghi, W.B. and Cashmore, A.R. (1995) Light-regulated transcription. Ann.
Rev. Plant Physiol. Plant Mol. Biol. 46: 445–474.
Thiele, D.J. (1992) Metal-regulated transcription in eukaryotes. Nucleic Acids
Res. 20: 1183–1191.
Thum, K.E., Kim, M., Morishige, D.T., Eibl, C., Koop, H.U. and Mullet, J.E.
(2001) Analysis of barley chloroplast psbD light-responsive promoter ele-
ments in transplastomic tobacco. Plant Mol. Biol. 47: 353–366.
Toyofuku, K., Umemura, T. and Yamaguchi, J. (1998) Promoter elements
required for sugar-repression of the RAmy3D gene for alpha-amylase in rice.
FEBS Lett. 428: 275–280.
Urao, T., Yamaguchi-Shinozaki, K., Urao, S. and Shinozaki, K. (1993) An
Arabidopsis myb homolog is induced by dehydration stress and its gene
product binds to the conserved MYB recognition sequence. Plant Cell 5:
1529–1539.
van Heusden, G.P.H., Wenzel, T.J., Lagendijk, E.L., de Steensma, H.Y. and van
der Berg, J.A. (1992) Characterization of the yeast BMH1 gene encoding a
putative protein homologous to mamalian protein kinase II activators and
protein kinase C inhibitors. FEBS Lett. 302: 145–150.
Vervliet, G., Holsters, M., Teuchy, H., van Montagu, M., Schell, J. (1975) Char-
acterisation of different plaque-forming and defective temperate phages in
Agrobacterium strains. J. Gen. Virol. 26: 33–48.
Wang, Y., Lorenzi, I., Georgiev, O. and Schaffner, W. (2004) Metal-responsive
transcription factor-1 (MTF-1) selects different types of metal response ele-
ments at low vs. high zinc concentration. Biol. Chem. 385: 623–632.
Wilczynski, G., Kulma, A. and Szopa, J. (1998) The expression of 14-3-3
isoforms in potato is developmentally regulated. J. Plant Physiol. 153: 118–
126.
Xu, N., Hagen, G. and Guilfoyle, T. (1997) Multiple auxin response modules in
the soybean SAUR 15A promoter. Plant Sci. 126: 193–201.
Zhang, Z.L., Xie, Z., Zou, X., Casaretto, J., Ho, T.H., Shen, Q.J. (2004) A rice
WRKY gene encodes a transcriptional repressor of the gibberellin signaling
pathway in aleurone cells. Plant Physiol. 134: 1500–1513.
Zhou, D.X. (1999) Regulatory mechanism of plant gene transcription by GT-
elements and GT-factors. Trends Plant Sci. 4: 210–214
Zuk, M., Skala, J., Biernat, J. and Szopa, J. (2003) Repression of six 14-3-3 pro-
tein isoforms resulting in the activation of nitrate and carbon fixation key
enzymes from transgenic potato plants. Plant Sci. 165: 731–741.
(Received February 11, 2005; Accepted July 24, 2005)




















