
A Qualitative Analysis of Larval Drosophila melanogaster ...

DEVELOPMENTAL BIOLOGY 89, 379-388 (1982)
The Cuticle Proteins of Drosophila melanogasfer: Stage Specificity
CAROL J. CHIHARA, DONALD J. SILVERT,**’ AND JAMES W. FRISTROM*
Department of Biology, University of San Francisco, California 94117, and *Depatiwmnt of Genetics, University of Cal~brnia, Berhzky, Califmia 91720
Received June 29, 1981; accepted in revised fm August 28, 1981
Five stage-specific cuticles are produced during the development of Drosophila Urea-soluble proteins were extracted from each developmental stage and compared by gel electrophoresis. Proteins from first and second instar cuticle are identical except for minor differences in two proteins. Each subsequent stage, third instar, pupa, and adult, has a unique set of cuticle proteins. Qualitative changes within stages are seen in proteins from third instar and adult cuticle. Third instar cuticle proteins can be divided into “early” [proteins 2a, 3, 4, 5, ‘7, and 81 and “late” [proteins 2 and l] groups. Adult cuticle proteins change in relative amounts during pharate adult development and change mobility at eclosion. The lower abdominal pupal cuticle lacks a protein found in the pupal cuticle covering the head and thorax. Cuticle proteins from each stage are immunologically related. Nonetheless, electrophoretic variants of three larval proteins do not affect any major changes in the electrophoretic mobility of proteins from other stages. We propose that each stage (except first and second instar) has proteins encoded by discrete genes.
INTRODUCTION
The study of gene regulation during development re- quires a system of readily identifiable gene products differentially expressed in time and space. Previous studies (Srivistava, 1970; Roberts and Willis, 1980; Wil- lis et aL, 1981) indicate that cuticle proteins of insects are developmentally modulated. Cuticle proteins of Drosophila melanogaster have the additional advantage of ready genetic analysis. A subset of cuticle proteins, the urea-soluble ones, are particularly advantageous. The urea-soluble proteins extracted from cuticles of late third instar larvae have been previously described (Fristrom et al., 1978). We present here a description of the urea-soluble cuticle proteins throughout devel- opment of Drosophila melanogaster and demonstrate their suitability for studies on gene regulation.
Insect cuticles have a basic structure composed of two major layers: a surface epicuticle composed of waxes and proteins and an inner procuticle composed of chitin, a nitrogenous polysaccharide, and proteins. Five sep- arate cuticles, one for each of the three larval instars, one pupal, and one adult are formed during Drosophila development. All have the basic cuticular structure. As development progresses cuticles are shed and replaced by new ones. The shedding of a cuticle is accomplished in two stages: First, apolysis occurs as the existing cu- ticle separates from an underlying epithelium. A new
‘Present address: California Institute of Technology, Division of Biology. Pasadena, California 91125.
cuticle is then deposited by the epithelium. Second, the apolysed cuticle is shed, or molted. These events are under the control of the molting hormone 20-hydrox- yecdysone (Borst et al., 1974; de Reggi et al, 1975; Hod- getts et al, 1977). The three larval cutieules are exter- nally similar in appearance. However, the third instar larval cuticle changes dramatically at the end of the instar when it becomes the puparium. After the for- mation of the puparium, metamorphosis ensues with the formation of the anterior of the adult, head and thorax, from imaginal discs. The abdominal region, with the exception of tissue contributed by the genital disc, is still primarily composed of larval epithelial cells that contain among them nests of abdominal histo- blasts that will later in development completely replace the larval cells (Roseland and Schneiderman, 1979; Madhavan and Madhavan, 1980). Beginning about 3 hr after the formation of the puparium the pupal cuticle is deposited by a combination of larval and imaginal cells (Poodry and Schneiderman. 1970; Whitten, 1968). Twenty-four hours after puparium formation apolysis of the pupal cuticle occurs. Adult cuticle deposition be- gins at about 48 hr (Mitchell et al, 1977; Reed et aZ., 1975). After completion of the adult cuticle, the pharate adult is enclosed within three cuticles: outermost, the puparium, a modified larval cuticle deposited by larval epithelial cells; next the pupal cuticle laid down by a mosaic of larval and imaginal epithelial cells; innermost the adult cuticle secreted entirely by imaginal cells.
The urea-soluble cuticle proteins of Drosophila third instar larvae have already been investigated in some
379
0012-1606/82/020379-10$02.00/O Copyright 0 1982 by Academic Press. Inc. AII rights of reproduction in any form reserved.

380 DEVELOPMENTAL BIOLOGY VOLUME 89,1982
detail. Five major and several minor components are resolved by native-gel electrophoresis. The major pro- teins range in molecular weight from about 10K to 18K. Electrophoretic variants for four of the major and two of the minor proteins have been identified. Variants for third instar larval proteins 1,2, and 3 have been mapped to a limited region on the right arm of chromosome 2 (Chihara, Kimbrell, and Fristrom, unpublished). Ge- nomic clones encoding larval proteins 1-3, and possibly 4 have been recovered (Snyder et al, 1981) and DNA and protein sequencing studies (Snyder, Hunkapiller, Yuen, Silvert, Fristrom, and Davidson, unpublished) have resulted in the identification of four coding regions located within 7.9 kb of DNA for larval proteins l-3 and probably 4. Variants for larval cuticle protein 5 have been mapped to the left arm of chromosome 3 (Chihara, Kimbrell, and Fristrom, unpublished).
Studies on urea-soluble cuticle proteins have now been extended to other developmental stages. We show here that each cuticle contains its own set of proteins with the exception of cuticles from first and second in- stars which essentially share a common set. Although many of the proteins from all stages are immunologi- cally, and thus structurally, related, we present evi- dence here indicating that the major proteins of each cuticle are in general encoded by discrete genes whose expression is stage specific.
MATERIALS AND METHODS
Stocks. An Oregon R strain of Drosophila melanogas- ter kept at the University of California, Berkeley since 1964 was obtained in 1976 and maintained at the Uni- versity of San Francisco at 25°C on standard cornmeal agar-molasses medium. This strain will hereafter be designated as “wild-type.” All third instar variants were collected from the wild (Fristrom et aZ., 1978) ex- cept for the faint-3 stock that was isolated from a b pr c px sp stock.
Culture methods. Eggs were collected from population cages over 3-5 hr intervals as described by Mitchell et al. (1964). Batch cuticles were prepared from animals collected as follows:
1. First instar: Larvae were washed from boxes 26- 36 hr after egg laying. The wash was examined to as- certain that no molted cuticles were present thereby insuring that only first instar larvae were present.
2. Second instar: Larvae were harvested 55-60 hr after egg deposition.
3. Third instar: Early third instar larvae were re- covered 72-74 hr after egg laying, late third instar lar- vae between 110 and 115 hr, or from cultures in which the larvae had migrated from the food and approxi- mately 10% had formed puparia.
4. Pupae: For the recovery of pupal cuticles, pupae were harvested between 15 and 30 hr after puparium formation. None of these pupae had visible eye color; all had everted heads.
5. Adults: Adults were recovered within 15 min of emergence and placed immediately in ice-cold Ringer’s PTU2 and then frozen.
For dissection of individual cuticles from pupae or pharate adults, individual pupae were timed from the white prepupal stage.
Cuticle recovery and extraction Cuticles were re- covered and extracted following Fristrom et al. (1978), with the following modifications: All preparations were made in the presence of 0.1 mM NaTTh and 1 mlM PMSF. First, second, pupal, and adult cuticles were col- lected on 16XX nylon screening. Pupal cuticles were separated from puparia after homogenization in 8.5% NaCl by allowing puparial fragments to settle out and collecting the floating pupal cuticles. This procedure was repeated several times until the pupal cuticles were visually free of tanned, sclerotized material. Dissected pupal cuticles were split longitudinally and scraped as clean as possible. Each dissected cuticle was then rinsed three times in 0.5 M Tris, pH 7.0, PTU, blotted dry, and groups of 10 were immediately extracted in 15 ~1 of 7 MUT. Dissected adult cuticles could not be freed ade- quately of adhering tissue and were therefore homog- enized in groups of 10 in a glass homogenizer in 0.5 ml of Ringer’s PTU to remove adhering tissue and then pelleted at high speed in a clinical, table top centrifuge. Bits of cuticle were observed to be free of contaminating tissue using a stereomicroscope at 60X.
Electrophwesis. Vertical gel electrophoresis in either nondenaturing or SDS gels as follows:
1. Nondenaturing gels: Slab acrylamide gels, (15%) 0.7 mm thick were formed using one of two formulas: 15% acrylamide, 0.4% bis-acrylamide, 0.075 M Tris- HCl, pH 8.6 or 15% acrylamide, 0.08% bis-acrylamide, 0.3 M Tris-HCl, pH 8.6. We note that although the gels contained no denaturing agents, samples were loaded in buffers containing 7 M urea.
2. Denaturing gels: SDS-gel electrophoresis was per- formed after the method of Anderson et al. (1973), in 15% acrylamide.
All gels were stained with Coomassie Brilliant Blue- G250.
’ Abbreviations: PTU, all solutions containing PTU are saturated with phenylthiourea and contain 0.7% mercaptoethanol; Na’I”Th, so- dium tetrathionate; PMSF, phenyl methyl sulfonyl fluoride; 7MUT, 7 M urea, 5 mmM Tris-HCl, pH 8.6; L&P, larval cuticle proteins from first and/or second instar; L&P, larval cuticle proteins from third instar; PCP, pupal cuticle proteins; ACP, adult cuticle proteins.
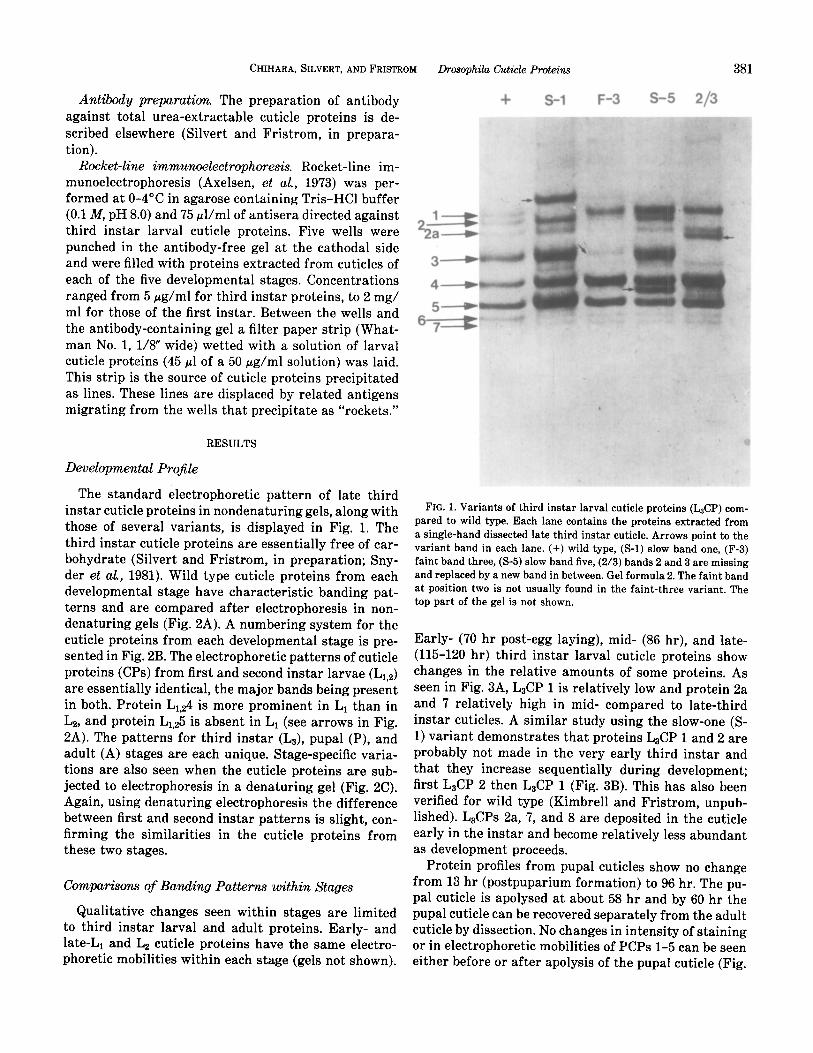
CHIHARA, SILVERT, AND FRISTROM Drosophila Cuticle Proteins 381
Antibody preparation. The preparation of antibody against total urea-extractable cuticle proteins is de- scribed elsewhere (Silvert and Fristrom, in prepara- tion).
Rocket-line immunoelectrophoresis. Rocket-line im- munoelectrophoresis (Axelsen, et al., 1973) was per- formed at 0-4°C in agarose containing Tris-HCl buffer (0.1 M, pH 8.0) and 75 J/ml of antisera directed against third instar larval cuticle proteins. Five wells were punched in the antibody-free gel at the cathodal side and were filled with proteins extracted from cuticles of each of the five developmental stages. Concentrations ranged from 5 pg/ml for third instar proteins, to 2 mg/ ml for those of the first instar. Between the wells and the antibody-containing gel a filter paper strip (What- man No. 1, l/8” wide) wetted with a solution of larval cuticle proteins (45 ~1 of a 50 pg/ml solution) was laid. This strip is the source of cuticle proteins precipitated as lines. These lines are displaced by related antigens migrating from the wells that precipitate as “rockets.”
RESULTS
Developmental Profile
The standard electrophoretic pattern of late third instar cuticle proteins in nondenaturing gels, along with those of several variants, is displayed in Fig. 1. The third instar cuticle proteins are essentially free of car- bohydrate (Silvert and Fristrom, in preparation; Sny- der et al., 1981). Wild type cuticle proteins from each developmental stage have characteristic banding pat- terns and are compared after electrophoresis in non- denaturing gels (Fig. 2A). A numbering system for the cuticle proteins from each developmental stage is pre- sented in Fig. 2B. The electrophoretic patterns of cuticle proteins (CPs) from first and second instar larvae (Lr,a) are essentially identical, the major bands being present in both. Protein Lr.24 is more prominent in L1 than in La, and protein L1,25 is absent in L1 (see arrows in Fig. 2A). The patterns for third instar (L3), pupal (P), and adult (A) stages are each unique. Stage-specific varia- tions are also seen when the cuticle proteins are sub- jected to electrophoresis in a denaturing gel (Fig. 2C). Again, using denaturing electrophoresis the difference between first and second instar patterns is slight, con- firming the similarities in the cuticle proteins from these two stages.
Comparisons of Banding Patterns within Stages
Qualitative changes seen within stages are limited to third instar larval and adult proteins. Early- and late-L1 and Lz cuticle proteins have the same electro- phoretic mobilities within each stage (gels not shown).
+ S-l F-3 S-5 2/3
FIG. 1. Variants of third instar larval cuticle proteins (L&P) com- pared to wild type. Each lane contains the proteins extracted from a single-hand dissected late third instar cuticle. Arrows point to the variant band in each lane. (+) wild type, (S-l) slow band one, (F-3) faint band three, (S-5) slow band five, (2/3) bands 2 and 3 are missing and replaced by a new band in between. Gel formula 2. The faint band at position two is not usually found in the faint-three variant. The top part of the gel is not shown.
Early- (70 hr post-egg laying), mid- (86 hr), and late- (115-120 hr) third instar larval cuticle proteins show changes in the relative amounts of some proteins. As seen in Fig. 3A, L&P 1 is relatively low and protein 2a and 7 relatively high in mid- compared to late-third instar cuticles. A similar study using the slow-one (S- 1) variant demonstrates that proteins L&P 1 and 2 are probably not made in the very early third instar and that they increase sequentially during development; first L&P 2 then L&P 1 (Fig. 3B). This has also been verified for wild type (Kimbrell and Fristrom, unpub- lished). L&Ps 2a, 7, and 8 are deposited in the cuticle early in the instar and become relatively less abundant as development proceeds.
Protein profiles from pupal cuticles show no change from 13 hr (postpuparium formation) to 96 hr. The pu- pal cuticle is apolysed at about 58 hr and by 60 hr the pupal cuticle can be recovered separately from the adult cuticle by dissection. No changes in intensity of staining or in electrophoretic mobilities of PCPs l-5 can be seen either before or after apolysis of the pupal cuticle (Fig.
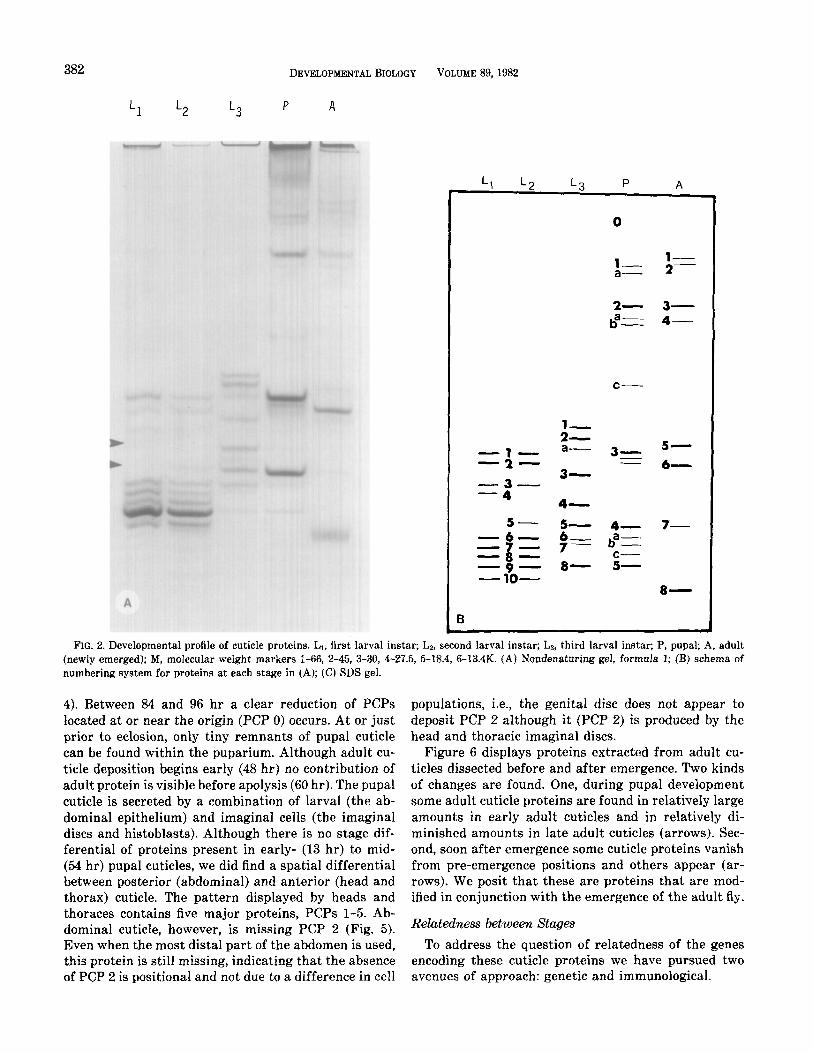
382 DEVELOPMENTAL BIOLOGY VOLUME 89, 1982
Ll L2 L3 P A
0
I--- ;= a-
c-
:= Ix:= a- 3- - 6- 5-
=i-- 3-
4- 5- 7-
-6- -- a :z 7 ;g
=g= 8- c- 5- -lO-
8-
6
FIG. 2. Developmental profile of cuticle proteins. Ll, first larval instar; Lz, second larval instar; La, third larval instar; P, pupal; A, adult (newly emerged); M, molecular weight markers l-66, 2-45, 3-30, 4-27.5, 5-18.4, 6-13.4K. (A) Nondenaturing gel, formula 1; (B) schema of numbering system for proteins at each stage in (A); (C) SDS gel.
4). Between 84 and 96 hr a clear reduction of PCPs located at or near the origin (PCP 0) occurs. At or just prior to eclosion, only tiny remnants of pupal cuticle can be found within the puparium. Although adult cu- ticle deposition begins early (48 hr) no contribution of adult protein is visible before apolysis (60 hr). The pupal cuticle is secreted by a combination of larval (the ab- dominal epithelium) and imaginal cells (the imaginal discs and histoblasts). Although there is no stage dif- ferential of proteins present in early- (13 hr) to mid- (54 hr) pupal cuticles, we did find a spatial differential between posterior (abdominal) and anterior (head and thorax) cuticle. The pattern displayed by heads and thoraces contains five major proteins, PCPs l-5. Ab- dominal cuticle, however, is missing PCP 2 (Fig. 5). Even when the most distal part of the abdomen is used, this protein is still missing, indicating that the absence of PCP 2 is positional and not due to a difference in cell
populations, i.e., the genital disc does not appear to deposit PCP 2 although it (PCP 2) is produced by the head and thoracic imaginal discs.
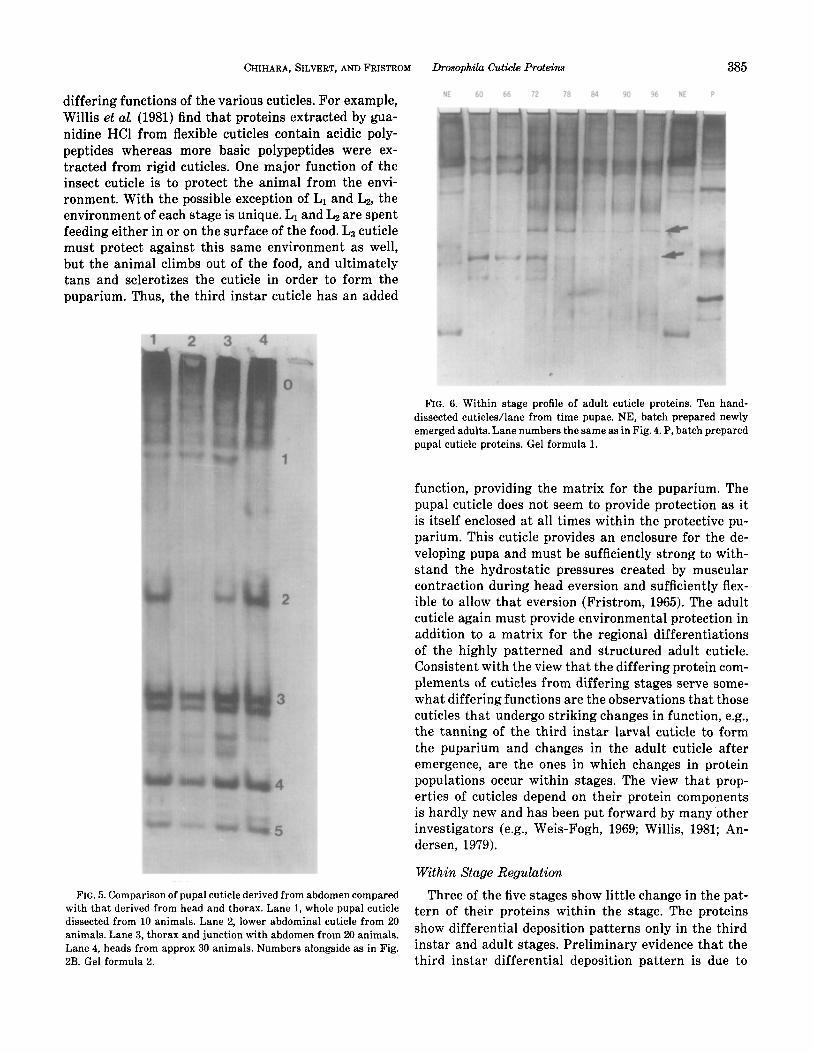
Figure 6 displays proteins extracted from adult cu- ticles dissected before and after emergence. Two kinds of changes are found. One, during pupal development some adult cuticle proteins are found in relatively large amounts in early adult cuticles and in relatively di- minished amounts in late adult cuticles (arrows). Sec- ond, soon after emergence some cuticle proteins vanish from pre-emergence positions and others appear (ar- rows). We posit that these are proteins that are mod- ified in conjunction with the emergence of the adult fly.
Relatedness between Stages
To address the question of relatedness of the genes encoding these cuticle proteins we have pursued two avenues of approach: genetic and immunological.
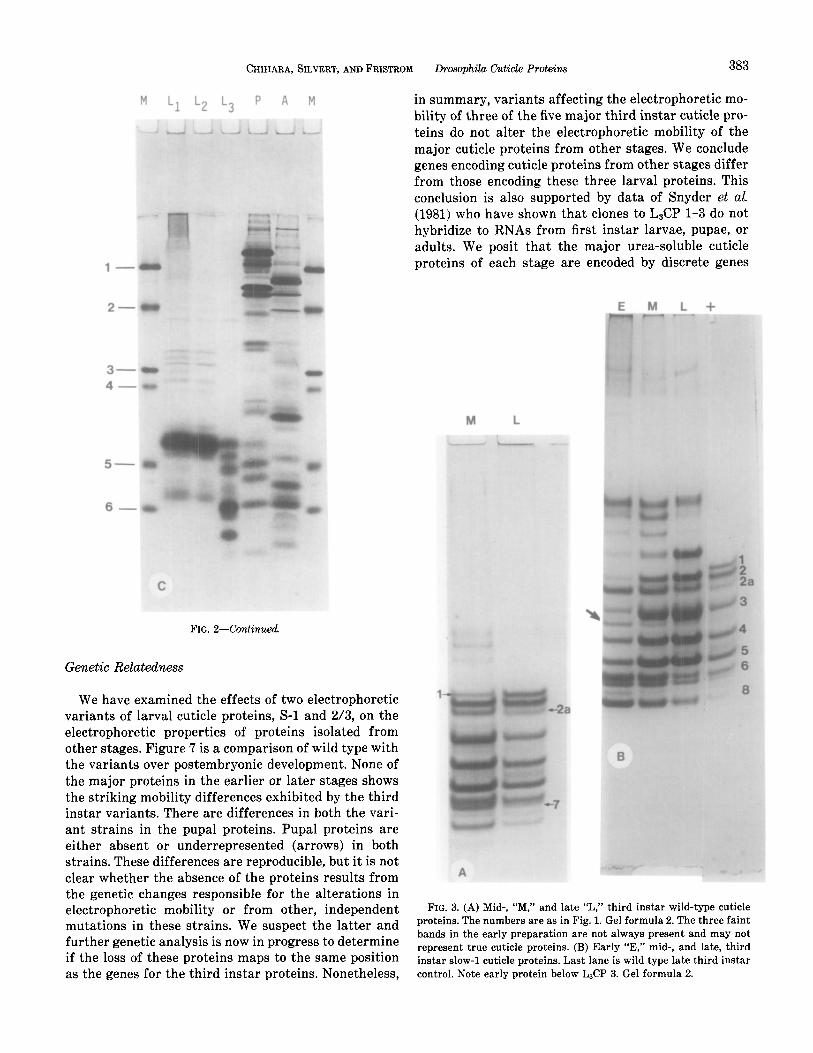
CHIHARA, SILVERT, ANDFRISTROM Drosophila Cuticle Proteins 383
M L1 L2 L3 P A M
l-
2-
3- 4-
5-
6-
FIG. 2-Continued
Genetic Relatedness
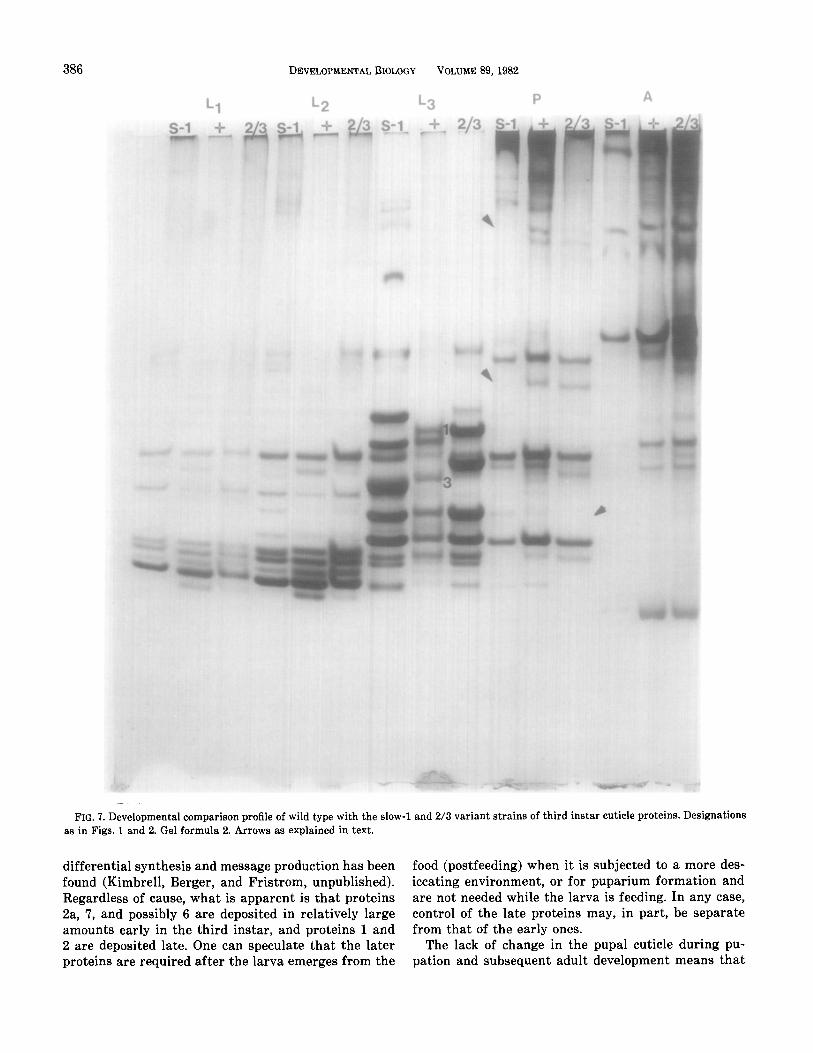
We have examined the effects of two electrophoretic variants of larval cuticle proteins, S-l and 2/3, on the electrophoretic properties of proteins isolated from other stages. Figure 7 is a comparison of wild type with the variants over postembryonic development. None of the major proteins in the earlier or later stages shows the striking mobility differences exhibited by the third instar variants. There are differences in both the vari- ant strains in the pupal proteins. Pupal proteins are either absent or underrepresented (arrows) in both strains. These differences are reproducible, but it is not clear whether the absence of the proteins results from the genetic changes responsible for the alterations in electrophoretic mobility or from other, independent mutations in these strains. We suspect the latter and further genetic analysis is now in progress to determine if the loss of these proteins maps to the same position as the genes for the third instar proteins. Nonetheless,
in summary, variants affecting the electrophoretic mo- bility of three of the five major third instar cuticle pro- teins do not alter the electrophoretic mobility of the major cuticle proteins from other stages. We conclude genes encoding cuticle proteins from other stages differ from those encoding these three larval proteins. This conclusion is also supported by data of Snyder et al. (1981) who have shown that clones to L&P 1-3 do not hybridize to RNAs from first instar larvae, pupae, or adults. We posit that the major urea-soluble cuticle proteins of each stage are encoded by discrete genes
FIG. 3. (A) Mid-, “M,” and late “L,” third instar wild-type cuticle proteins. The numbers are as in Fig. 1. Gel formula 2. The three faint bands in the early preparation are not always present and may not represent true cuticle proteins. (B) Early “E,” mid-, and late, third instar slow-l cuticle proteins. Last lane is wild type late third instar control. Note early protein below L&P 3. Gel formula 2.

384 DEVELOPMENTAL BIOLOGY VOLUME 89,1982
t 18 24 30 42 48 54 60 66 72 78 84 90 96 P
FIG. 4. Within stage profile of pupal cuticle proteins. Ten hand-dissected cuticles/lane from timed pupae. Lane numbers at top indicate hours from white puparium stage. Numbers alongside as in Fig. 2B. Pattern for 13 hr cuticle was identical to 18 hr gel not shown. Arrow indicates stage at which cuticle was first dissected separately from imaginal cuticle underneath. Gel formula 1.
(with the exception of those for the first and second instar).
Antigenic Relatedness of the Proteins between Stages
Antibody to the major third instar cuticle proteins creates three lines when reacted against L&P. This antibody when reacted against each of the develop- mental stage’s proteins in rocket-line immunoelectro- phoresis produced rockets that react with each of the three antigenic families characteristic of L&P (i.e., each stage’s proteins perturbed each of the three lines- see Fig. 8). Thus, there are antigens related to each of the major antigens of L&P present in each develop- mental stage. A more complete study of the antigenic relationships of these proteins will appear elsewhere (Silvert and Fristrom, in preparation). The complexity
of pupal and adult sets of proteins is somewhat greater than the larval proteins. If antipupal cuticle protein antiserum is added to the gel along with the anti-L&P, rockets distinct from the lines appear only in the pupal and adult cuticle protein preparations (data not shown). Thus, there are proteins in the pupal and adult stages that are not antigenically related to larval proteins.
DISCUSSION
Stage Specificity We have presented data indicating that the cuticle
of each developmental stage contains unique proteins encoded by separate and, at most, partially overlapping gene sets for the major cuticle proteins of each stage (with the exception of first and second larval instars which are essentially the same). It is likely that these differences in the cuticle proteins are involved with
CHIHARA, SILVERT, AND FRISTROM Drosophila Cuticle Proteins 385
differing functions of the various cuticles. For example, Willis et a,!+ (1981) find that proteins extracted by gua- nidine HCl from flexible cuticles contain acidic poly- peptides whereas more basic polypeptides were ex- tracted from rigid cuticles. One major function of the insect cuticle is to protect the animal from the envi- ronment. With the possible exception of L1 and Lz, the environment of each stage is unique. L1 and L2 are spent feeding either in or on the surface of the food. L3 cuticle must protect against this same environment as well, but the animal climbs out of the food, and ultimately tans and sclerotizes the cuticle in order to form the puparium. Thus, the third instar cuticle has an added
FIG. 5. Comparison of pupal cuticle derived from abdomen compared with that derived from head and thorax. Lane 1, whole pupal cuticle dissected from 10 animals. Lane 2, lower abdominal cuticle from 20 animals. Lane 3, thorax and junction with abdomen from 20 animals. Lane 4, heads from approx 30 animals. Numbers alongside as in Fig. 2B. Gel formula 2.
NE 60 66 72 78 84 90 96 NE P
FIG. 6. Within stage profile of adult cuticle proteins. Ten hand- dissected cuticles/lane from time pupae. NE, batch prepared newly emerged adults. Lane numbers the same as in Fig. 4. P, hatch prepared pupal cuticle proteins. Gel formula 1.
function, providing the matrix for the puparium. The pupal cuticle does not seem to provide protection as it is itself enclosed at all times within the protective pu- parium. This cuticle provides an enclosure for the de- veloping pupa and must be sufficiently strong to with- stand the hydrostatic pressures created by muscular contraction during head eversion and sufficiently flex- ible to allow that eversion (Fristrom, 1965). The adult cuticle again must provide environmental protection in addition to a matrix for the regional differentiations of the highly patterned and structured adult cuticle. Consistent with the view that the differing protein com- plements of cuticles from differing stages serve some- what differing functions are the observations that those cuticles that undergo striking changes in function, e.g., the tanning of the third instar larval cuticle to form the puparium and changes in the adult cuticle after emergence, are the ones in which changes in protein populations occur within stages. The view that prop- erties of cuticles depend on their protein components is hardly new and has been put forward by many other investigators (e.g., Weis-Fogh, 1969; Willis, 1981; An- dersen, 1979).
Within Stage Regulation
Three of the five stages show little change in the pat- tern of their proteins within the stage. The proteins show differential deposition patterns only in the third instar and adult stages. Preliminary evidence that the third instar differential deposition pattern is due to
386 DEVELOPMENTALBIOLOGY VOLUME 89,1982
FIG. 7. Developmental comparison profile of wild type with the slow-l and 2/3 variant strains of third instar cuticle proteins. Designations as in Figs. 1 and 2. Gel formula 2. Arrows as explained in text.
differential synthesis and message production has been food (postfeeding) when it is subjected to a more des- found (Kimbrell, Berger, and Fristrom, unpublished). iccating environment, or for puparium formation and Regardless of cause, what is apparent is that proteins are not needed while the larva is feeding. In any case, Za, 7, and possibly 6 are deposited in relatively large control of the late proteins may, in part, be separate amounts early in the third instar, and proteins 1 and from that of the early ones. 2 are deposited late. One can speculate that the later The lack of change in the pupal cuticle during pu- proteins are required after the larva emerges from the pation and subsequent adult development means that

CHIHARA, SILVERT, AND FRISTROM Drosophila Cuticle Proteins 387
A FIG. 8. Rocket-line immunoelectrophoresis of batch preparations
of cuticle proteins from each of the developmental stages against anti-L&P with lines formed by L&P. Filter paper with L&P was applied in region indicated by arrow.
the proteins of the pupal cuticle are not resorbed upon apolysis and subsequent molting events. This obser- vation is at variance with published descriptions of the molting process (Locke, 1974; Jungreis, 1979) in which enzymes in the ecdysial fluid break down components of the old cuticle. If such a breakdown occurs at or around the pupal/adult molt in Drosophila its effects were undetected. The profile of adult proteins indicates that these proteins are regulated during development and that several are modified at emergence.
conc1usicms
The cuticle proteins of Llrosophila are developmen- tally regulated and represent the products of at least one and perhaps several gene families. Snyder et al. (1981) have demonstrated that the third instar protein genes, L&P 1, 2, 3, and possibly 4, are within 7.9 kb of each other. Thus, several of the members of the gene family are clustered. Genetic mapping (Chihara et al, unpublished) and in situ hybridization of the above mentioned clone place these genes at 62 on the meiotic map or 44D in the salivaries. The L&Ps 5,6, and 8 map to the third chromosome (Chihara et a& unpublished) and experiments are underway to determine if these
form a second cluster. Such clustering of genes related in structure and/or function is exemplified by the ver- tebrate hemoglobins where several genes for /3 and p- like chains which function at different stages of devel- opment are clustered. If the genes for the proteins of stages other than third instar also cluster with their third instar counterparts they may represent a similar set of gene families. Such a system of gene families in Drosophila offers a system which could greatly increase our knowledge of gene organization and control.
This work was supported by NSF Grant PCM-7822516 to CJC, NIH Grant GM-19937 to JWF, and NIH Training Grant GM-07127 to the Department of Genetics. We thank Ms. Jennifer Hoffman for expert technical assistance.
REFERENCES
ANDERSEN, S. 0. (1979). Biochemistry of insect cuticle. Annu. Rev. Entomd 27,29-61.
ANDERSON, C. W., BAUM, P. R., and GESTELAND, R. F. (1973). Pro- cessing of adenovirus e-induced proteins. J. viral. 12,241-252.
AXELSON, N. H., KROLL, J., and WEEKE, B. (1973). “A Manual of Quan- titative Immunoelectrophoresis. Methods and Applications.” Black- well, Oxford.
BODENSTEIN, D. (1965). The postembryonic development of Drosoph- ila, In “Biology of Drosophila” (M. Demerec, ed.), pp. 275-367. Hof- ner, New York.
BORST, D. W., BOLLENBACHER, W. E., O’CONNOR, J. D., KING, D. S., and FRISTROM, J. W. (1974). Ecdysone levels during metamorphosis of Drosophila melunogaster. Develop. BioL 39,308-316.
DE REGGI, M. L., HIRN, M. H., and DELAAGE, M. A. (1975). Ecdysone release in DrosophiZa during postembryonic development. Biochem Biophhys. Res. Commun 66,1307-1315.
FRISTROM, J. W. (1965). Development of the morphological mutant cryptocephal of Drosophila melunogaster. Genetics 52,299-318.
FRISTROM, J. W., HIP, R. J., and WATT, F. (1978). The procuticle of Drosophila: Heterogeneity of urea soluble proteins. Biochemistry 17,3917-3924.
HODGE?TS, R. B., SAGE, B., and O’CONNOR, J. D. (1977). Ecdysone titers during post embryonic development of Drosophila melano- gaster. Develop. Biol 60,310-317.
JUNGREIS, A. M. (1979). Physiology of moulting in insects. Adwan Insect. Physiol. 14, 109-183.
LOCKE, M. (1974). The structure and formation of the integument in insects. In “The Physiology of Insecta” (M. Rockstein, ed.), 2nd ed., Vol. VI. Academic Press, New York.
MADHAVAN, M. M., and MADHAVAN, K. (1980). Morphogenesis of the epidermis of adult abdomen of Drosophila, J. Embryol. Exp. Mar- phol. 60,1-31.
MILNER, M. J., and SANG, J. H. (1974). Relative activities of cu-ecdysone and @-ecdysone for the differentiation in vitro of Drosophila me- Zunoguster imaginal discs. Cell 3, 141-143.
MITCHELL, H. K., and MITCHELL, A. (1967). Mass culture and age selection in Drosophila. Drosophila Iffy Serv. 39,135-137.
MITCHELL, H. K., WEBER-TRACY, U. M., and SCHAAR, G. (1971). AS- pects of cuticle formation in Drosophila melanogadw. J. Exp. Zool. 176,429-444.

388 DEVELOPMENTAL BIOLOGY VOLUME 89,1982
POODRY, C. A. (1980a). Epidermis: Morphology and development. In SILVERT, D. J., and FRISTROM, J. W. (1980). Biochemistry of imaginal “The Genetics and Biology of Drosophila” (M. Ashburner and T. discs: Retrospect and prospect. Insect &o&em 10,341-355. Wright, eds.), Vol. 2, pp. 443-497. Academic Press, New York.
POODRY, C. A. (1980b). Imaginal discs: Morphology and development. In “The Genetics and Biology of Drosophila” (M. Ashburner and T. Wright, eds.), Vol. 2, pp. 40’7-441. Academic Press, New York.
POODRY, C., and SCHNEIDERMAN, H. A. (1970). The ultrastructure of the developing leg of Drosophila melanogaster. Wilhelm Roux’ Arch. Develop Biol 166, l-44.
REED, C. T., MURPHY, C., and FRISTROM, D. (1975). The ultrastructure of the differentiating pupal leg of Drosophila melanogader. Wilhelm Roux’ Arch. Develop. Biol. 178,285-302.
ROBERTS, P. E., and WILLIS, J. H. (1980). The cuticular proteins of Tenebio molitor. I. Electrophoretic banding patterns during post- embryonic development. Develop. Biol. 75,59-69.
ROSELAND, C. R., and SCHNEIDERMAN, H. A. (1979). Regulation and metamorphosis of the abdominal histoblasts of Drosophila mela- nogastw. Wilhelm Roux’ Arch Develop. Biol. 186,235-265.
SNYDER, M., HIRSH, J., and DAVIDSON, N. (1981). The cuticle genes of Drosophila: A developmentally regulated gene cluster. Cell 25,165- 177.
SRIVASTAVA, R. P. (1970). Electrophoretic behaviour of cuticular pro- teins of different developmental stages of Galleria mellonella J. Insect Physiol. 16, 2345-2351.
WEIS-FOGH, T. (1969). Structure and formation of insect cuticle. Symp. Roy. Ent. Sot., London 5, 165-185.
WHITTEN, J. (1968). Metamorphic changes in insects. In “Metamor- phosis: A Problem in Developmental Biology” (W. Etkin and L. I. Gilbert, eds.), pp. 43-105. Appleton, Century, Crofts, New York.
WILLIS, J. H., REGIER, J. C., and DEBRUNNER, B. A. (1981). In “Current Topics in Insect Endocrinology and Nutrition” (G. Bhaskaran, S. Friedman, and J. G. Rodriguez, eds.), pp. 27-46. Plenum, New York.
Copyright © 2022 FDOKUMEN




















