
Segregation distortion in Drosophila melanogaster: genomic organization of Responder sequences
7
Copyright 0 1996 by the Genetics Society of America Segregation Distortion in Drosophila rnelanogaster: Genomic Organization of Responder Sequences Roberta Moschetti,” Ruggiero Caizzi” and Sergio Pimpinellit *Istituto di Genetica, Universita di Bari, Bari, Italy and +Dipartimento di Genetica e Biologia Molecolare, Universita di Roma “La Sapienza, ” Roma, Italy Manuscript received April 10, 1996 Accepted for publication September 14, 1996 ABSTRACT The heterochromatic Responder (Rsp) locus of Drosophila melanogaster is the target of the two distorter loci Sd and E(SD). Rsp is located in a specific heterochromatic region of the second chromosome and is made up of AT-rich satellite sequences whose abundance is related to its sensitivity to the distorter chromosomes. Here we report that a cluster of Rsp sequences is also located in the third chromosome. The third-chromosome cluster has the same flanking sequences as the clone originally used to identify the Rsp elements, and one of the flanking sequences is a rearranged 412 retrotrsansposon. The presence of a second, unlinked Rsp-sequence cluster makes re-interpretation necessary for some earlier experi- ments in whichsegregation of thethirdchromosomehadnotbeenfollowedandraisesinteresing possibilities for the origin of the Rsp locus. S EGREGATION distortion in Drosophila melunoguster is a widespread selfish genetic system that was dis- covered by HIMZUMI in 1956. Males heterozygous for a SD (Segregation Distorter) second chromosome and a SO” homologue transmitan excess of SD chromosomes due to disfunction of SD+ bearing sperm (see TEMIN et al. 1991 for review). Segregation Distortion depends on the interaction of three main elements. Two of these, Sd (Segregation distorter) and E(SD) {Enhancer of SD), are required to induce dysfunction of sperm carrying a sen- sitive allele of the third element Rsp (Responder) (GANET- ZKY 1977; SHARP et ul. 1985; TEMIN 1991). While Sd maps to a euchromatic region of the poly- tene chromosome map (3702-6) (GANETZKY 1977), both E(SD) and Rsp have heterochromatic locations (see TEMIN et al. 1991 for review). Of the three loci, the best characterized so far is the Rsp locus. Genetic analy- sis of Responderhas revealed that there is a continuum of allelic states with different sensitivities to SD (HIRAIZUMI and THOMAS 1984). Cytological analysis has shown Rsp maps to a single bright fluorescent region in 2R (h39) close to the centromere (Figure la) and that variation in sizeof the region is correlated with Rsp sensitivity (PIMPINELLI and DIMITRI 1989). Molecular analysis of Responder has revealed that the locus is composed of 120-bp AT-rich XbuI repeats organized in dimers, whose abundance also correlates with sensitivity (Wu et al. 1988). Because the Responder locus seems to be absent in sibling species of D. melanogaster (Wu et al. 1988) and Corresponding author: Sergio Pimpinelli, Dipartimento di Genetica Ado Mor0 5, 00185 Roma, Italy. e Biologia Molecolare, Universiti di Roma “La Sapienza,” Piazzale E-mail: [email protected] Genetics 144: 1665-1671 (Decernher, 1996) because survivalof the Segregation Distortion system in nature depends on the presence of sensitive chromo- somes, two main unanswered questions about the Re- sponder locus concern its origin and maintenance. WU et ul. (1989) have proposed that sensitive chromosomes are maintained in nature because Rsp has a positive impact on fitness. An effect on viability of the Rsp se- quences has been suggested by the finding that an in- sensitive chromosome, containing a deletion of h39 region, has reduced viability in population cage experi- ments (Wu et al. 1989). This chromosome, an X-ray- generated deficiency (GANETZKY 1977), however, lacks more than just Rsp. Moreover, this hypothesis is directly challenged by molecular analysis of the region (CAIZZI et al. 1993); the deficiency used for the population phage experiments is also missing an element whose striking features suggest potential function. This ele- ment, called Bari-1, is a transposon-like element with multiple euchromatic locations and a unique hetero- chromatic location in the h39 region close to the Rsp sequences. Sequence analysis has shown that Bari-1 het- erochromatic repeatsposses the same intact open read- ing frame as the euchromatic copies. Moreover, molec- ular studies on natural populations have shown that, while Rsp DNA is extremely variable, the Bari-1 hetero- chromatic array is very stable. Both the homogeneity and the potential coding capacity suggest that the het- erochromatic Ban’-1 sequences have a functional role that might better explain the results of the population cage experiments. In the present work, we report novel findings on the genomic organization of the Rsp sequences. We have found that there are clustered Rsp sequences located in the proximal 80C region of the polytene map just
Transcript of Segregation distortion in Drosophila melanogaster: genomic organization of Responder sequences

Copyright 0 1996 by the Genetics Society of America
Segregation Distortion in Drosophila rnelanogaster: Genomic Organization of Responder Sequences
Roberta Moschetti,” Ruggiero Caizzi” and Sergio Pimpinellit
*Istituto di Genetica, Universita di Bari, Bari, Italy and +Dipartimento di Genetica e Biologia Molecolare, Universita di Roma “La Sapienza, ” Roma, Italy
Manuscript received April 10, 1996 Accepted for publication September 14, 1996
ABSTRACT The heterochromatic Responder (Rsp) locus of Drosophila melanogaster is the target of the two distorter
loci Sd and E(SD). Rsp is located in a specific heterochromatic region of the second chromosome and is made up of AT-rich satellite sequences whose abundance is related to its sensitivity to the distorter chromosomes. Here we report that a cluster of Rsp sequences is also located in the third chromosome. The third-chromosome cluster has the same flanking sequences as the clone originally used to identify the Rsp elements, and one of the flanking sequences is a rearranged 412 retrotrsansposon. The presence of a second, unlinked Rsp-sequence cluster makes re-interpretation necessary for some earlier experi- ments in which segregation of the third chromosome had not been followed and raises interesing possibilities for the origin of the Rsp locus.
S EGREGATION distortion in Drosophila melunoguster is a widespread selfish genetic system that was dis-
covered by HIMZUMI in 1956. Males heterozygous for a SD (Segregation Distorter) second chromosome and a SO” homologue transmit an excess of SD chromosomes due to disfunction of SD+ bearing sperm (see TEMIN et al. 1991 for review). Segregation Distortion depends on the interaction of three main elements. Two of these, Sd (Segregation distorter) and E(SD) {Enhancer of SD), are required to induce dysfunction of sperm carrying a sen- sitive allele of the third element Rsp (Responder) (GANET- ZKY 1977; SHARP et ul. 1985; TEMIN 1991).
While Sd maps to a euchromatic region of the poly- tene chromosome map (3702-6) (GANETZKY 1977), both E(SD) and Rsp have heterochromatic locations (see TEMIN et al. 1991 for review). Of the three loci, the best characterized so far is the Rsp locus. Genetic analy- sis of Responderhas revealed that there is a continuum of allelic states with different sensitivities to SD (HIRAIZUMI and THOMAS 1984). Cytological analysis has shown Rsp maps to a single bright fluorescent region in 2R (h39) close to the centromere (Figure la ) and that variation in size of the region is correlated with Rsp sensitivity (PIMPINELLI and DIMITRI 1989). Molecular analysis of Responder has revealed that the locus is composed of 120-bp AT-rich XbuI repeats organized in dimers, whose abundance also correlates with sensitivity (Wu et al. 1988).
Because the Responder locus seems to be absent in sibling species of D. melanogaster (Wu et al. 1988) and
Corresponding author: Sergio Pimpinelli, Dipartimento di Genetica
Ado Mor0 5, 00185 Roma, Italy. e Biologia Molecolare, Universiti di Roma “La Sapienza,” Piazzale
E-mail: [email protected]
Genetics 144: 1665-1671 (Decernher, 1996)
because survival of the Segregation Distortion system in nature depends on the presence of sensitive chromo- somes, two main unanswered questions about the Re- sponder locus concern its origin and maintenance. WU et ul. (1989) have proposed that sensitive chromosomes are maintained in nature because Rsp has a positive impact on fitness. An effect on viability of the Rsp se- quences has been suggested by the finding that an in- sensitive chromosome, containing a deletion of h39 region, has reduced viability in population cage experi- ments (Wu et al. 1989). This chromosome, an X-ray- generated deficiency (GANETZKY 1977), however, lacks more than just Rsp. Moreover, this hypothesis is directly challenged by molecular analysis of the region (CAIZZI et al. 1993); the deficiency used for the population phage experiments is also missing an element whose striking features suggest potential function. This ele- ment, called Bari-1, is a transposon-like element with multiple euchromatic locations and a unique hetero- chromatic location in the h39 region close to the Rsp sequences. Sequence analysis has shown that Bari-1 het- erochromatic repeats posses the same intact open read- ing frame as the euchromatic copies. Moreover, molec- ular studies on natural populations have shown that, while Rsp DNA is extremely variable, the Bari-1 hetero- chromatic array is very stable. Both the homogeneity and the potential coding capacity suggest that the het- erochromatic Ban’-1 sequences have a functional role that might better explain the results of the population cage experiments.
In the present work, we report novel findings on the genomic organization of the Rsp sequences. We have found that there are clustered Rsp sequences located in the proximal 80C region of the polytene map just
1666 R. Moschetti, R. Caizzi and S . Pimpinelli
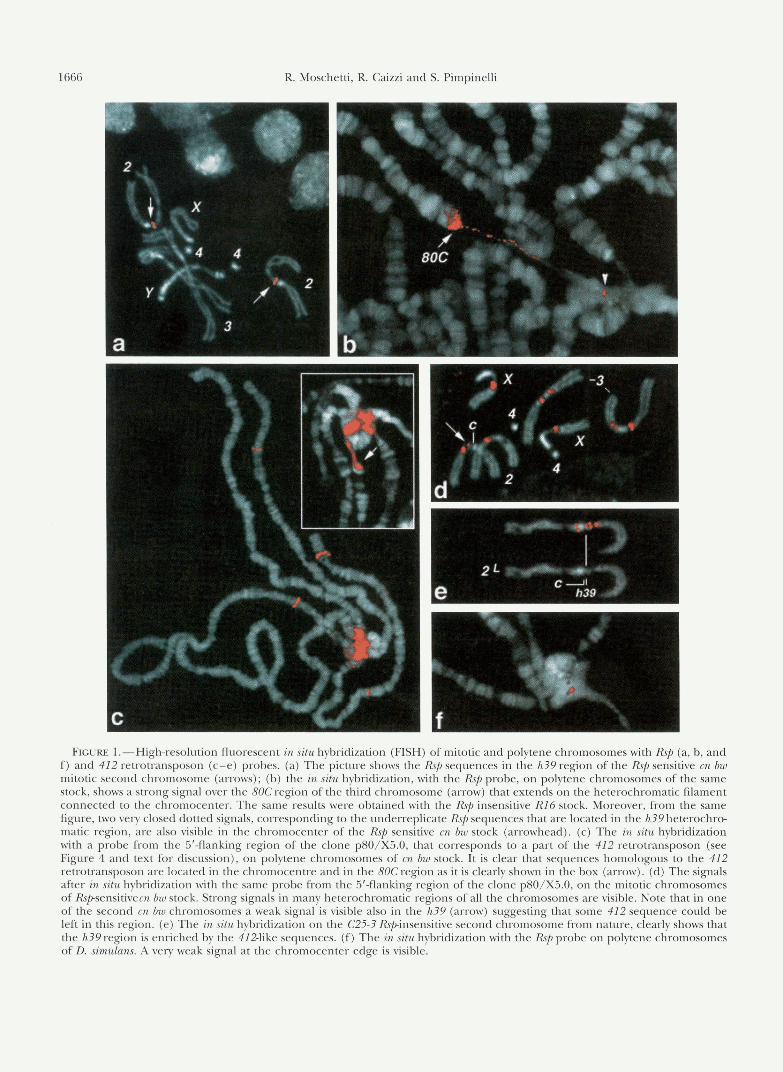
FIGURE 1.-High-resolution fluorescent in situ hybridization (FISH) of mitotic and polytene chromosomes with Rsp (a, b, and f ) and 412 retrotransposon (c-e) probes. (a) The picture shows the Rsp sequences in the h39 region of the Rsp sensitive cn bw mitotic second chromosome (arrows); (b) the in situ hybridization, with the Rsp probe, on polytene chromosomes of the same stock, shows a strong signal over the 80C region of the third chromosome (arrow) that extends on the heterochromatic filament connected to the chromocenter. The same results were obtained with the Rsp insensitive R16 stock. Moreover, from the same figure, two very closed dotted signals, corresponding to the underreplicate Rsp sequences that are located in the h39 heterochro- matic region, are also visible in the chromocenter of the Rsp sensitive cn bw stock (arrowhead). (c) The in situ hybridization with a probe from the 5"flanking region of the clone pSO/X5.0, that corresponds to a part of the 412 retrotransposon (see Figure 4 and text for discussion), on polytene chromosomes of m bw stock. It is clear that sequences homologous to the 412 retrotransposon are located in the chromocentre and in the 80C region as it is clearly shown in the box (arrow). (d) The signals after in situ hybridization with the same probe from the 5'-flanking region of the clone pSO/X5.0, on the mitotic chromosomes of Rspsensitivem bw stock. Strong signals in many heterochromatic regions of all the chromosomes are visible. Note that in one of the second cn bw chromosomes a weak signal is visible also in the h39 (arrow) suggesting that some 412 sequence could be left in this region. (e) The in situ hybridization on the C25-3 Rspinsensitive second chromosome from nature, clearly shows that the h39 region is enriched by the 412-like sequences. (f) The in situ hybridization with the Rsp probe on polytene chromosomes of D. simulans. A very weak signal at the chromocenter edge is visible.

D. mlanogaster Rsp Sequences 1667
outside the heterochromatin of the third chromosome. Intriguingly, these sequences are adjacent to a412retro- transposon which, besides its multiple euchromatic lo- cations, is also present in the h39 region. In general, experimental work on the SD system has focused on the second chromosome, and this is especially true of work on Rsp. For example, there has been little or no attempt to keep track of the origin of the third chromo- somes in stocks used to study Rsp, and evolutionary interpretations of sequence information have assumed that all of that information came from second chromo- some copies of the Rsp satellite. In the discussion, we consider the impact of identification of a second, un- linked Rsp cluster and some of the questions this raises about the origin and function of the Rsp locus.
MATERIALS AND METHODS
Drosophila strains: All genetic markers are described in LINDSLEY and ZIMM (1992).
R16, cn bw is a second chromosome carrying an X ray- induced deficiency of the Responder locus (GANETZKI 1977).
SDRoma was collected in Rome, Italy, by NICOLETTI and TRIPPA (1967).
C l l l and C25-3 are naturally occurring Rsp-insensitive chro- mosomes that were collected and genetically characterized by TEMIN and MARTHAS (1984). All of these chromosomes were cytologically characterized by PIMPINELLI and DIMITRI (1988).
To obtain strains isogenic for third chromosomes from the R16 stock the following crosses were performed: R l6 homozy- gous females were crossed en masse to males carrying the T(2;3) Ts.Tl, Tb Cy translocation. R16/Ts.T1 heterozygous males were then individually crossed to females carrying sec- ond and third chromosome balancers marked with Cy bw vi and Hu Sb. Male and female Cy Sb progeny were then crossed to generate stocks isogenic for the third chromosome in an R16 background.
Molecular techniques: Using the Promega LambdaGEM- 12 XhoI half-site arms cloning system, a genomic library from homozygous R16 flies was constructed following the protocol supplied by the manufacturer. Fifty thousand recombinant phages were screened by procedures described in MANIATIS et al. (1982) using the 240-bp XbaI fragment present in the H0 clone (WU et al. 1988) as a probe. After three rounds of plaque purification, 20 positive phages were analyzed for the presence of a 5.0- or 3.7-kb Hind111 fragment, the sizes ex- pected from genomic analysis. A 5.0-kb fragment, present in three lambda clones, was then subcloned in a pUC8 plasmid vector for further analysis. No phages with the 3.7-kb HindIII variant form were found.
Genomic DNA was prepared from adult flies by phenol/ chloroform extraction. Digestions, blotting and hybridization conditions were as in MANIATIS et al. (1982). DNA fragments used as probes for hybridization experiments were gel puri- fied and J’P-labeled by the random priming method (FEIN- BERG and VOCELSTEIN 1983).
High-resolution fluorescent in situ hybridization (FISH) and CCD camera analysis: Polytene chromosome preparations were made using standard techniques. Mitotic chromosome preparations from larval brains, fluorescent in situ hybridiza- tion and CCD camera analysis were carried out as described in GATTI et al. (1994).
Probe DNA was labeled with digoxigenin-l1dUTP (Boher- inger, Mannheim) by nick translation, and signals were de-
tected with rhodamine-conjugated sheep antidigoxigenin IgG, Fab fragments (Boheringer, Mannheim).
RESULTS
Zn situ hybridization of XbaI repeats to polytene and mitotic chromosomes: It has been reported that XbuI repeats corresponding to the original Ho clone isolated from a A phage library were always visible in Southern blots of different stocks carrying nearly complete defi- ciencies of the Responder locus (WU et al. 1988).The authors inferred that a small proximal portion of the Responder sensitive locus, complementary to part of the Hll clone, is still present in the Responder insensitive de- ficiencies. Cytological analysis of the same deficiencies however, has shown that they completely lack the h39 region as their proximal breakpoint lies in the h 3 8 re- gion. Since the Rsp is entirely within h39and h 3 8 region shows very different cytological and molecular features (PIMPINELLI and DIMITRI 1989; LOHE et al. 1993), this rules out any molecular continuity between the Re- sponder locus and the residual H1, signals and opens the possibility that Rsplike repeats may also be present in a different genomic location.
To test for the existence of XbuI repeats outside the Responder locus, we performed an in situ hybridization to polytene chromosomes of both the Rspsensitive cn bw stock and its Rspdeficient-insensitive R16 derivative. In both stocks, we found a strong hybridization signal in the 8UC region of the third chromosome (Figure 1 b). Moreover, in situ analysis of polytene chromosomes of 210 larvae from each of 10 different natural popula- tions with a global distribution invariably gave hybrid- ization signals in the same region. These results strongly suggest that Rsp sequences are stable components of the 80C region.
Genomic analysis of XbaI repeats in the Rl6 stock To study the 8UC region in more detail, we first looked at the structural organization of XbuI repeats in the Rl6 stock. Southern blot analysis, using three different enzymes and probed with XbuI sequence, in- variably shows two hybridization bands (Figure 2). The two fragments consistently show a 3:l hybridization in- tensity ratio and a difference in size of about 1.2-1.4 kb. Thus, two different clusters of the XbuI repeats are present in the R16 stock. Since the three enzymes give the same size difference between the large and the small variants, they probably differ in number of the XbuI repeats rather than in non- XbuI sequences. Thus, the larger variant contains 10 more copies of the XbuI re- peats than does the smaller. To test if these clusters are both in the 80C region of the third chromosome, we did Southern analyses of single Rl6 flies and of 13 lines homozygous for randomly selected third chromosomes using the 240 bp XbuI probe (see MATERIALS AND METH-
ODS). As shown in Figure 3a, after digestion with HindIII, we found that most randomly chosen R16 flies

1 (;(;x R. Yloschctti, R. C 1 i u . i m t l S. Pilnpinelli
FK;~.RI; . 2..-Gcnomic analysis o f t h r ,Yhd rcpc.ats i n homo- zyg011s K l h stock. Two Inicrograms o l ' DSX from homozygous / < / h stock tligcstrtl w i t h k,'rol<,iil (E), (,'///I ((:) o r l ~ i ~ ~ l l l l (1-1) ~ v c ~ c Sotlthrrn h l o t t r d together with IfintlIII digested D L \ f rom c~ h7I I (lane 4) ;\nd O w q o ~ ~ - I < (lane .5). I n c w h tligcst o f ' R l h Iancs, t h r diff+rc-ncr i n size o l ' hybridization h n d is roughly 1.2 k h and t h c upper I x w t l is thrcc times ~ n o r c intense with rcspcct t o the IOWCI- I ) m d . Prol)c is t h e S / M ~ I 23O-l)p fmg- m r n t .
were either homozygous for the :i.O-kb band o r hctcro- z\.go~rs for the two \.ariants. Of 30 single flies analyzed, 12 \vere homozygous for the large variant, 17 were het- erozygous and only one was homozygous for the small variant. Figure 31) show the results with lines isogenic for different third chromosomes. Each line is clearly homozygous for either the .5.0- or the 3.f-kb variant. The t w o variants are therefore located in the 80Cregion but do not coexist in individual chromosome. They are alleles. The same results were ;dso obtained using I.i-oRI and C ' k d (clava not sho\\~ll).
Figurc 31) also shows the patterns obtained with the 0'25-3, C I 1 I and SIl-Ro?nt/ stocks, three R~.s~)on~lrr.insensi- tivc chromosomes from natural populations (N1c:o- I.I--ITI and TRIITA 1967; Tnrls and MARTIIAS 1984). For C25-3, DNA was extracted from Rlci heterozygotes. C I 1 I and SI)-Rotnn were cxaminrd in C:y heterozygotes, but C y is also an insensitive chromosome that is com- pletely deleted for 113% the pattern o f the XhrI repeats in Rlci/Lj flies is identical t o that of homozygous Rlci flies (data not shown). Therefore, all o f the signals in lanes 15-17 of Figure 31, come from the 80C region o f the third chromosome o r from the 1139 rcgion of the second chromosomes taken from natural populations (see also M'r P/ crl. 1988). C25-3 and C I I I clearly show the two fragments coming from 80C and, as \v(. P / d .
FK;~'KI; . S.--Evitlcnccs for a tu^) allelic state o f the .Unl re- pwt i n ROC rrgion. ( a ) I<x;unplc of a Ili~ctlIIl digested DS:\ lronl I C ) single llirs I;untlolntychoosc~n i n Rlhs tock; (13) / / itall l l digests o f 13 isogrnic 3rd chron1osoInc o f X l h stock (Iancs I - 13) . R l h stock control DS..\ (I;Inc 13). ( : 2 i - i / / < l h ( I a n c t.?),
( , ' / I / / ( , : ) ( l a w 1 6 ) . S / ) / < ~ I I I I ~ / ( : ~ (lanc 17). I n I a n c IS t h r I/indIll cliq-st o l ' t h c /<r,~/):f,rn,lrlr,,.st.nsiti\c. ~ J I l m stock is shown. I n this stock t h c / I ' ~ . s / m ~ d r ~ r rcgion contains "x00 Mnl repc.ats. To ohtain a comp;lmhle signals intcnsitv, t h c quantity I ) I ' the lo;~dctl cn h7If DS,.\ NXS I O timcs Icss t h a n t h r DS;\ i n the other lanes.
(1988) previonsly showed, the)- contain X I , d copies in the 1/39 rcgion (lanes 1.5 ant l 16). In contrast, the .TI)- Ronm stock (lane Ii), shows the small 80C variant and only t w o other very weak signals. I V c do not know whether these col-respond t o a few intact I<.sj) sequences o r t o other partially homologous scqrlcnccs. Morcovcl-, by Southern analysis, we found that thc samc t w o frag- ments coming from 8OC arc also present in DNA S a m -
ples prepared from 30 flies from the same 10 n x t r w a l populations r~sctl for the iu s i / n hyl,ritlization ( d a t a not sho\vn).
Cloning and characterization of XbuI repeats from 8OC: To analyze thc organization of the X h I copies in the 8OC region, we constructed a lambda libray from the homozygous I<lh strain and screened i t with thc 240-bp X h I repeat. Several positive clones wcrc forlntl and three clones containing a . 5kh I - l i rdIII fragment identical in size t o that found in the genomic DSX o f the same strain were fr~rther charactcrizcd. Tlw .5-kb IfintlIII fragments from thcsc cloncs ~vere sltbcloncd in pLTTc8, and we obtained the clone p80/?(.5.0 whose restriction map is shown in Figure 4. From this clone, we obtained further suhcloncts (Figure 4) which were sequenced. \Ye found that one region flanking t h c ?3d- repeat cluster contains scqucnces homologous t o the 412 transposon that, besides being found in scvc~-al CII-
chromatic regions antl in the chromocenter, is also l o -

D. melanogaster Rsp Sequences 1669
1 K b
H c x I
. " " . S O E C
pao/xcl
" "
pBO/HC1 X4 X6 X0.5 p8O/XS
FIGURE 4.-Physical map of the DNA fragment p80/X5.0, harboring Responder sequences from 80C region. The map represents the 5.0-kb Hind111 fragment subcloned from lambda R2 (see MATERIALS AND METHODS). Dashed box repre- sents the region where XbaI repeats are found as determined by Southern blot hybridization and sequencing analysis. Be- low the map, the subclones used for sequencing are indicated. Symbols are: H, HindIII; C, ClaI; X, Xbd; S, Sau3A 0, XhoI; and E, EcoRI. Vertical arrowheads represent XbaI sites not finely determined. Bars below the map indicate subclones used in DNA sequencing analysis to verify the identity of XbaI repeat.
cated in 80C region (Figure IC). Intriguingly, CABOT et ul. (1993) showed that the Ho clone is also flanked by a 412 retrotransposon. Comparison of the sequences flanking both sides of the XbuI repeats of both the 8OC clusters and the Ho clone (Figure 5) reveals nearly com- plete identity. Thus, although the Ho clone was used to identify the Rsp cluster in h39, the clone itself probably came from 80C.
The non-XbuI arms of the 5.0-kb variant total -3 kb. Hence, there are about 15 copies of the 120-bp XbuI repeat in this variant and five copies in the 3.7-kb vari- ant. Since this corresponds to the 3:l intensity ratio between the two fragments, the number of copies of the entire cluster must be the same in both alleles. We didn't test directly whether each cluster is present only once, but, from the low relative intensity of the 5.0- and 3.7-kb fragments in the C25-3 and Cl l l insensitive chromosomes (lanes 15 and 16 of Figure 3) there should be, if not one, very few copies of the clusters.
In situ hybridizations to mitotic chromosomes of both the cn bwsensitive and the C25-3insensitive chromo- somes using the 412 -like flanking sequences are shown in Figure 1, d and e. In addition to several other hetero- chromatic signals in both stocks, a weak signal in the h39 region of the cn bw chromosome and a strong signal in the h39region of the C25-3chromosome are present. These results clearly show that the transposon is located in several heterochromatic regions and that, while the h39 heterochromatic block of the sensitive cn bw chro- mosome contains some 412-like sequences, the same block in the C25-3insensitive chromosome is abun- dantly enriched in such sequences.
To test for the presence of Rsplike sequences outside
pEO/XO. 7 - p a o / w x -
a 8OC
b 8OC
FIGURE 5.-Dotplot comparison of flanking sequences of the XbuI repeat clone Ho (WU et al. 1988) and the flanking sequences of the XbuI repeat clone pSO/X5.0 coming from the 80Cregion (see Figure 4). (a) Comparison between the 5'- flanking sequences. (B) Comparison between the 3'-flanking sequences. The near identity among the flanking sequences of both clones strongly suggests that the H,) clone comes from the 80C region.
D. melunoguster, we did in situ hybridization with the XbuI probe to polytene chromosomes of D. simuluns under moderately stringent conditions. As shown in Fig- ure lf , a weak hybridization signal is visible in the chro- mocentric region. This signal is not only weak in poly- tene chromosomes, but we have been unable to detect it by in situ hybridization to mitotic chromosomes. Hence, we do not know if this represent a very low number of Rsplike elements or some other sequence partially complementary to the XbuI repeat.
DISCUSSION
Clustered Rsp sequences are located outside the Re- Spender locus: The results show that XbuI sequences identical to those of the Respondm locus are a stable component of the 80C polytene region of the third chromosome and can be found in two allelic forms that

1670 R. Moschetti, R. Caizzi and S. Pimpinelli
differ in length. The analysis by both Southern blot and in situ hybridization of natural populations has shown that XbuI sequences are always present in this region. Moreover, we found that in a long-established SDRoma stock, XbaI sequences, that exclusively correspond to the cluster with the lower copy number, are also main- tained in the 80C region.
The molecular analysis of the 8UC region has also shown that one of the flanking sequences of the XbaI clusters corresponds to the 412 retrotransposon. The sequence comparison of the 80C clusters to the Ho clone previously isolated by WU et ul. (1988) shows very strong similarity, if not complete identity. All have the same genomic organization and are flanked at both sides by the same sequences. We propose, therefore, that the Ho clone was actually derived from this region. That the Ho clone could come from 80C is also sug- gested by evidence obtained in an extensive analyses of the Rsp locus in many non-SD chromosomes extracted from natural populations (Wu et ul. 1988; CAIZZI et al. 1993). These studies have shown that a very high level of polymorphism for the XbuI arrays exists except for the fragment corresponding to the H(, clone, which was always present and invariant, thus suggesting a different location. It is also possible, however, that the crosses used to isolate the second chromosomes of these popu- lations introduced and homogenized third chromo- somes derived from a common ancestor.
The relationship of the Rsp and 8UC clusters: The presence of a second XbuI cluster in the third chromo- some raises the intriguing question of whether there are evolutionary or functional relationships between the two. It also forces us to re-examine earlier infer- ences that were based on an underlying assumption that all copies of the XbuI repeat are in the second chromosome Rsp locus. With addition of the informa- tion reported here, the picture seems to be as follows:
1. We have no information at all about whether there is a functional relationship between the two clusters. The SD-Romu stock carries only the 3.7-kb allele of 8OC, but we do not know whether the third chromo- somes of that stock came from the SD-Romu fly col- lected from a natural population or from some labo- ratory stock. Knowing that the 80C cluster exists, however, certainly gives strong impetus for some ex- perimental studies.
2. From in situ hybridization studies, we know that there is widespread presence of the 80C XbaI cluster in natural populations. All larvae from the 10 popu- lations we sampled gave signals of similar intensity, suggesting a fairly stable copy number, but we don’t know if there might not be chromosomes that are missing this cluster, as is the case for the Rsp cluster in the 2.5-5% of SDchromosomes present in natural populations (see TEMIN et al. 1991 for review).
3. From the Southern analyses of the same 10 natural
populations we know that both alleles are present in all the populations, suggesting that the organization of the 8UC cluster might be constant. The data col- lected for five other stocks (cn bw, R16, C23-5, C l l l and Cy) also show that both the 5- and 3.7-kb Hind111 alleles are present. These stocks have an overlapping provenance, however, and are likely to share third chromosomes from the same laboratory strains. Wu et al. (1988) found that a 6.6-kb EcoRI fragment (cor- responding to the 5-kb Hind111 variant-see Figure 2) was present in 26 of 28 genotypes with different non-SD second chromosomes. All of these flies, how- ever, were heterozygous for Cy0 and hence all shared at least one third chromosome from the Cy0 stock.
4. Two reported observations suggest that the origin of the 80C cluster preceded speciation of D. melanogas- ter. First, the analysis of divergence of the XbuI re- peats in the Ho clone presented in WU et al. (1988) implies that this clone, whose flanking sequences are nearly identical to those of our genomic 80C clone, is quite ancient. Second, CABOT et ul. (1993), citing unpublished results of E. L. CABOT and A. T. BECK- ENBACH, report that the 412-like sequence of H(, re- sembles a D. pseudoobscuru 412 sequence more than it resembles the D. melunoguster 412 element.
5 . CABOT et ul. (1993) examined intra- and inter-array XbuI repeat sequence variation in 32 different geno- mic clones. They argue that the surprisingly low level of interchromosomal us. intrachromosomal differ- ences in Rsp sequences suggests either that the chro- mosome coalescence time is short in the region (i. e., all Rsp arrays were derived from a common ancestral array in the very recent past) or that homologous exchanges, albeit infrequent, are sufficient to oblit- erate between-chromosome differences.
Assuming that conventional crossovers fairly reflect the frequency of interchromosomal information transfer, an assumption that may not be true for heterochromatic arrays (WILLIAMS et ul. 1989), CABOT et al. favor the first of these a relatively recent origin of the Rsp array.
6. Our understanding of the mechanisms underlying the maintenance of repeated arrays is in a rather fragile state. Magnification and reduction of rDNA arrays appears to occur via several pathways (see HAWLEY and MARCUS 1989 for review). The similarity of rDNA sequences in D. melunoguster X and Y chrc- mosomes, and their marked divergence from the sequences of related species, is inconsistent with ho- mogenization by infrequent X-Y single crossovers (WILLIAMS et ul. 1989). Nearest-neighbor analysis of the D. melanoguster histone cluster has suggested that the diffusion of new variants within the cluster is consistent with unequal sister-strand exchange, but the displacements are most by only one repeat apart (COLBY and WILLIAMS 1993). The BUT--1 array in h39

D. melanogaster Rsp Sequences 1671
is nearly monomorphic ( CAIZZI et al. 1993). For the h39 Rsp cluster, there is solid evidence for unequal sister-strand exchange (CABOT et al. 1993), there is large variation in number of XbuI repeats, and there is seemingly complementary expansion and diminu- tion of other sequences in h39, including changes in number of 412-like sequences. Finally, with only 10 populations sampled, the 80C cluster of XbaI re- peats appears to be rather stable.
If there is an origin-product relationship between the 8OC and Rsp (h39) clusters at all, one can plausibly argue that the apparently ancient origin of the small 8OC array, and the short coalescence time within the large and variable Rsp array, implies evolutionary direction. If so, we might even think of the 412-like element as a vehicle for the transfer from 8OC to h39. On the other hand, one can also build plausible arguments for trans- fer by an ancient 412-like element from the variable h39 cluster to 80C where a small array became fixed. We invite our population genetics colleagues to try to decide whether extant information about the structures of these two clusters is sufficient, whether present ideas about the DNA dynamics of repeated arrays is reliable enough, and whether theories of repeated-array evolu- tion are robust enough to reach a conclusion at this point. Some data already exist that might be helpful. In particular, WU et al. (1988) collected sequence infor- mation on the several repeats of the Ho clone and CABOT et al. (1993) have extensive sequence informa- tion on genomic XbaI repeats. It may also be useful to have similar information for the repeats that are in the two 80C alleles. Nevertheless, we do not know whether comparison of intra- and inter-array variability among the Ho (80C) and genomic repeats (most of which are probably derived from h39) can actually resolve this question. It would certainly be reassuring if the type of data needed for a convincing evolutionary argument could be specified before the experiments are done rather than having evolutionary arguments constructed post hoc.
We thank CORRADO CAGCESE, MARIA BERLOCO and LAURA FANTI for help in the cytological experiments. We are particularly grateful to LXNNY ROBBINS for the invaluable help in generally improving the manuscript by thoughtful comments and suggestions on both the science and writing. We would also like to thank our two reviewers, CHUNC~I WU and an anonymous reviewer for pointed and construc- tive comments; they have forced us to not only improve our writing, but to more rigorously examine our data. This research was supported by the grants: (CR. n. 91. 00012PF99) of Progetto finalizzato “In- gegneria Genetica” of the Italian National Research Council; MURST 40%; Grandi progetti di Ateneo MURST 60% of the university of
Rome and by the “Istituto Pasteur Fondazione Cenci Bolognetti,” University of Rome “La Sapienza.”
LITERATURE CITED CABOT, E. L., P. DOSHI, M.-L. Wu and C.4. Wu, 1993 Population
genetics of tandem repeats in centromeric heterochromatin: un- equal crossing over and chromosomal divergence at the Rtsponder locus of Drosophila mrlanogaster. Genetics 135: 477-487.
CAIZLI, R., C:. CAGGESE and S . PIMPINEI.I.I, 1993 Bari-1 a new transpo- son-like family in Drosophila melanogaster with a unique hetero- chromatic organization. Genetics 133: 335-345.
COLBY, C., and S. M. WIILIAMS, 1993 The distribution and spreading of rare variants in the histone multigene family of Drorophila melanogaster. Genetics 135: 127-133.
FEINBERG, A. P., and B. VOGEI.STEIN, 1983 A technique for radiola- beling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 132: 6-13.
GANETZW, B., 1977 On the component5 of segregation distortion in Drosophila mrlanogaster. Genetics 86: 325-355.
GATTI, M., S. BONA(:CORSI and S. PIMPINEILI, 1994 Looking at Dro- sophila mitotic chromosomes. Methods Cell Biol. 44: 371-391.
HAWI.EY, R. S., and C. H. MARCUS, 1989 Recombinational controls of rDNA redundancy in Lh-osophila. Genetics 23: 87-120.
HIRAIZUMI, Y., and A. M. THOM4S, 1984 Suppressor system of Segre- gation Distorter (SD) chromosomes in natural populations of Dro- sophila mehnogaster. Genetics 106: 279-292.
LINUSIXY, D. LA., and G. ZtMh4 1992 Thr Gmomr of Drosophila melano- gaster, Academic Press, New York.
LOHE, A. R., A. J. HII.I.IKER and P. A. ROBERTS, 1993 Mapping simple repeated DNA sequences in heterochromatin of Drosophila w l a - nogaster. Genetics 134: 1149-1174.
MANIATIS, T., E .F. FRrrs(:II a n d J . SAMBROOK, I982 Molrcular Clon- ing; A Laborato?y Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
NI(:m.E:’Im, B., and G. TRIPPA, 1967 Osservazioni citologiche su un nuovo cas0 di Segregation Iktortion (SO) in una popolazione natu- rale di Drosophila mrlmogastm. Atti Assoc. Genet. Ital. 12: 361- 365.
PIMPINEI.I.I, s., and P. DIMlTRI, 1989 Cytogenetic organization of the Rsp (Responder) locus in Drosophila mrlanogaster, Genetics 121: 765-772.
SHAM’, C. B., A. J. HII.LIKER and D. G. HOLM, 1985 Further charac- terization of genetic elements associated with the segregation Distortion phenomenon in Drosophila melanogaster. Genetics 110: 671-688.
TEMIN, R. G., 1991 The indipendent distorting ability of the En- hancer of Segregation Distortion, E(<%), in Drosophila melanogaster. Genetics 128 339-356
TEMIN, R. G., and M. MARTHAS, 1984 Factors influencing the effect of Segregation Distortion in natural populations o f Drosophila melanogastrr. Genetics 187: 375-393.
TEMIN, R. G., B. GANETZKY, P. A. POWERS, T. W. LITTLE, S. PIMPINEI.I.I rt al., 1991 Segregation Distortion in Drosophila mrlanogaster: genetic and molecular analyses. Am. Nat. 137: 287-331.
WILLIAMS, S. M., J. A. KENNISON, L. G. ROBBINS and C. STROBECK, 1989 Reciprocal recombination and the evolution of the ribo- somal gene family of Drosophila melanogaster. Genetics 122: 617- 624.
WU, C.4, T. W. Lurrm, M:L. WLI and G.-F. LIN, 1988 Association between a satellite DNA sequence and the Responderof Srgrrgation distorter in Drosophila mrlanogaster. Cell 5 4 179-189.
WU, C.-I., J. R. TRUE and J. NORMAN, 1989 Fitness reduction associ- ated with the deletion of a satellite DNA array. Nature 341: 248- 251.
Communicating editor: V. G. FINNERIY




















