
Inhibiting HIV1 integrase by shifting its oligomerization equilibrium
Upload
independentCategory
view
2download
0
Molecular basis of coiled-coiloligomerization-state specificityBarbara Ciania,1,2, Saša Bjelićb,1, Srinivas Honnappab,1,3, Hatim Jawharib,1,4, Rolf Jaussib, Aishwarya Payapillya,Thomas Jowitta, Michel O. Steinmetzb,5, and Richard A. Kammerera,5,6
aWellcome Trust Centre for Cell-Matrix Research, Faculty of Life Sciences, University of Manchester, Manchester M13 9PT, United Kingdom; andbBiomolecular Research, Paul Scherrer Institut, CH-5232 Villigen PSI, Switzerland
Edited* by Thomas D. Pollard, Yale University, New Haven, CT, and approved September 14, 2010 (received for review June 23, 2010)
Coiled coils are extensively and successfully used nowadays to ra-tionally design multistranded structures for applications, includingbasic research, biotechnology, nanotechnology, materials science,and medicine. The wide range of applications as well as theimportant functions these structures play in almost all biologicalprocesses highlight the need for a detailed understanding of thefactors that control coiled-coil folding and oligomerization. Here,we address the important and unresolved question why thepresence of particular oligomerization-state determinants withina coiled coil does frequently not correlate with its topology. Wefound an unexpected, general link between coiled-coil oligomeriza-tion-state specificity and trigger sequences, elements that are indis-pensable for coiled-coil formation. By using the archetype coiled-coil domain of the yeast transcriptional activator GCN4 as a modelsystem, we show that well-established trimer-specific oligomeriza-tion-state determinants switch the peptide’s topology from a dimerto a trimer only when inserted into the trigger sequence. We suc-cessfully confirmed our results in two other, unrelated coiled-coildimers, ATF1 and cortexillin-1. We furthermore show that multipletopology determinants can coexist in the same trigger sequence,revealing a delicate balance of the resulting oligomerization stateby position-dependent forces. Our experimental results shouldsignificantly improve the prediction of the oligomerization stateof coiled coils. They therefore should have major implications forthe rational design of coiled coils and consequently many applica-tions using these popular oligomerization domains.
protein folding ∣ protein structure ∣ protein–protein interactions
Because of its simplicity, the α-helical coiled coil has been usedtraditionally in a vast number of studies aimed at understand-
ing the fundamental principles of protein stability, folding, andoligomerization (reviewed in refs. 1 and 2). As a result, coiledcoils are exploited nowadays as multipurpose tools in a steadilyincreasing number of applications ranging from basic research tomedicine (2). For example, designed two- and three-strandedcoiled coils were successfully used as lead molecules to target theadenomatous polyposis coli tumor-suppressor protein that isimplicated in colorectal cancers (3) or to inhibit HIV infection(4), respectively. For these and many other applications, knowl-edge of the factors that control the folding and oligomerizationstate appears essential to rationally engineer specific coiled-coilstructures with the desired properties.
It is well established that so-called “trigger sequences,” short,distinct amino acid sequences, play an important role in control-ling coiled-coil formation (reviewed in ref. 1). A characteristicfeature of many trigger sequences is that they fold into reasonablystable monomeric helices before the coiled-coil structure forms(5–7). We recently solved the NMR structure of a peptide span-ning the GCN4 trigger sequence and showed that its structureis stabilized by a network of hydrogen bonds and electrostaticinteractions (7). The helical conformation of the trigger sequenceprovides an effective local structural scaffold for the interactionof key residues of colliding chains. Accordingly, preformed andfolding competent helices form a nucleation site, which promotes
productive in-register chain association. Interacting helices then“zip up” along the molecule to direct formation of a stable coiled-coil structure. Trigger sequences of different proteins show con-siderable diversity, indicating that a consensus sequence is unli-kely to exist (7). As a result, the prediction of trigger sequenceson the basis of sequence information alone remains a challengeand still requires experimental verification.
With respect to coiled-coil topology, several determinantsthat control the oligomerization state of coiled coils have beenidentified and studied in detail (reviewed in refs. 1 and 2). It isgenerally acknowledged, for example, that the distribution ofhydrophobic core residues, in particular isoleucine and leucine,correlates well with the oligomerization state of coiled coils. Theoccurrence of isoleucine and leucine residues at the heptad repeata and d core positions, respectively, favors the formation of dimerswhereas the reverse arrangement results in tetrameric structures.In contrast, a more even distribution of isoleucine at both the aand d positions facilitates trimer formation. Buried polar residuesin hydrophobic interfaces also play an important role in determin-ing the number of strands in coiled coils. Many two-strandedcoiled-coil domains of transcription regulators, for example, fre-quently contain at least one conserved asparagine or lysine residueat an a position toward the center of the sequence (2).
Interhelical interactions between side chains of residues at thee and g positions as well as the packing of these amino acidsagainst the hydrophobic a and d core residues have also beenshown to contribute significantly to oligomerization-state speci-ficity of coiled coils (reviewed in refs. 1 and 2). This is exemplifiedby the trimerization motif ArgðgÞ-hðaÞ-x-x-hðdÞ-GluðeÞ (denotedR-hxxhE where hðaÞ ¼ Ile, Leu, Val, Met; hðdÞ ¼ Leu, Ile, Val;x ¼ any amino acid residue) that specifies a three-stranded, par-allel topology of coiled-coil domains (8). The trimerization driv-ing force of the motif can be explained by optimal side chain–sidechain interactions whereby the strictly conserved arginine and
Author contributions: B.C., S.B., S.H., H.J., M.O.S., and R.A.K. designed research; B.C., S.B.,S.H., H.J., R.J., A.P., and T.J. performed research; B.C., S.B., S.H., H.J., T.J., M.O.S., and R.A.K.analyzed data; and M.O.S. and R.A.K. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
Data deposition: The atomic coordinates and structure factors have been deposited in theProtein Data Bank, www.pdb.org (PDB ID codes 3GJP and 2O7H).1B.C., S.B., S.H., and H.J. contributed equally to this work.2Present address: Department of Chemistry, University of Sheffield, Sheffield S3 7HF,United Kingdom.
3Present address: Novartis Institutes for Biomedical Research, Novartis Pharma AG, 4002,Basel, Switzerland.
4Present address: Roche Applied Science Customer Support Centre, Roche DiagnosticsGmbH, D-68305 Mannheim, Germany.
5To whom correspondence should be addressed. E-mail: [email protected] [email protected].
6Present address: Biomolecular Research, Paul Scherrer Insititut, CH-5232 Villigen PSI,Switzerland.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1008502107/-/DCSupplemental.
19850–19855 ∣ PNAS ∣ November 16, 2010 ∣ vol. 107 ∣ no. 46 www.pnas.org/cgi/doi/10.1073/pnas.1008502107
glutamate residues form a distinct bifurcated interhelicalsalt-bridge network and participate in the formation of the hydro-phobic core by establishing tight packing interactions to theneighbouring residues at the a and d positions through their ali-phatic moieties. Thus, similar to contacts stabilizing thermostableproteins, the trimerization motif encompasses both networks ofsurface salt bridges and optimal internal hydrophobic packinginteractions (8).
An open issue is that the presence of a specific oligomeriza-tion-state determinant does frequently not correlate with thecorresponding coiled-coil topology. Although the trimerizationmotif R-hxxhE is predominantly found in many diverse proteinfamilies harboring parallel three-stranded coiled-coil domains,it is also present in some dimers and antiparallel trimers, suchas the transcriptional activators GCN4 (9) and ATF1 (10) orthe actin-bundling protein cortexillin-1 from Dictyostelium discoi-deum (6). To understand the molecular basis of this discrepancy,we have carried out a detailed biophysical and structural study tosystematically address how the topology of these two-strandedcoiled-coil model systems is affected when particular oligomeri-zation-state determinants are placed at different coiled-coil posi-tions. We thus address the fundamental and still unresolvedquestion how the position of specific motifs within a coiled coildefines its topology.
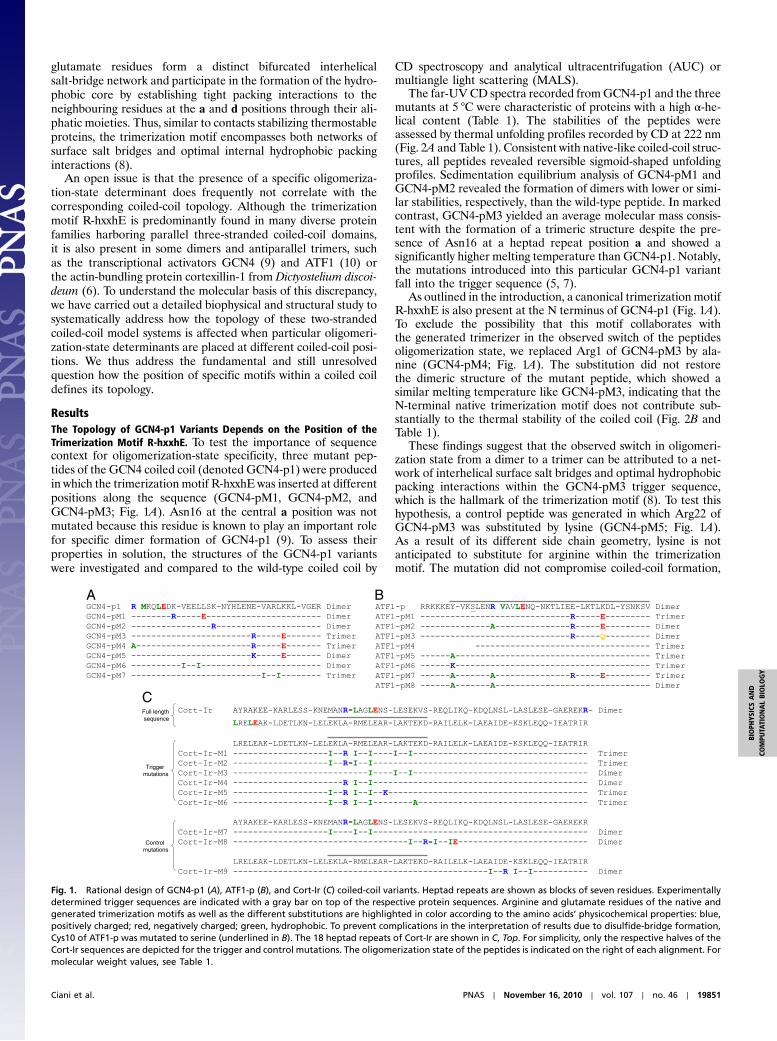
ResultsThe Topology of GCN4-p1 Variants Depends on the Position of theTrimerization Motif R-hxxhE. To test the importance of sequencecontext for oligomerization-state specificity, three mutant pep-tides of the GCN4 coiled coil (denoted GCN4-p1) were producedin which the trimerization motif R-hxxhE was inserted at differentpositions along the sequence (GCN4-pM1, GCN4-pM2, andGCN4-pM3; Fig. 1A). Asn16 at the central a position was notmutated because this residue is known to play an important rolefor specific dimer formation of GCN4-p1 (9). To assess theirproperties in solution, the structures of the GCN4-p1 variantswere investigated and compared to the wild-type coiled coil by
CD spectroscopy and analytical ultracentrifugation (AUC) ormultiangle light scattering (MALS).
The far-UV CD spectra recorded fromGCN4-p1 and the threemutants at 5 °C were characteristic of proteins with a high α-he-lical content (Table 1). The stabilities of the peptides wereassessed by thermal unfolding profiles recorded by CD at 222 nm(Fig. 2A and Table 1). Consistent with native-like coiled-coil struc-tures, all peptides revealed reversible sigmoid-shaped unfoldingprofiles. Sedimentation equilibrium analysis of GCN4-pM1 andGCN4-pM2 revealed the formation of dimers with lower or simi-lar stabilities, respectively, than the wild-type peptide. In markedcontrast, GCN4-pM3 yielded an average molecular mass consis-tent with the formation of a trimeric structure despite the pre-sence of Asn16 at a heptad repeat position a and showed asignificantly higher melting temperature than GCN4-p1. Notably,the mutations introduced into this particular GCN4-p1 variantfall into the trigger sequence (5, 7).
As outlined in the introduction, a canonical trimerization motifR-hxxhE is also present at the N terminus of GCN4-p1 (Fig. 1A).To exclude the possibility that this motif collaborates withthe generated trimerizer in the observed switch of the peptidesoligomerization state, we replaced Arg1 of GCN4-pM3 by ala-nine (GCN4-pM4; Fig. 1A). The substitution did not restorethe dimeric structure of the mutant peptide, which showed asimilar melting temperature like GCN4-pM3, indicating that theN-terminal native trimerization motif does not contribute sub-stantially to the thermal stability of the coiled coil (Fig. 2B andTable 1).
These findings suggest that the observed switch in oligomeri-zation state from a dimer to a trimer can be attributed to a net-work of interhelical surface salt bridges and optimal hydrophobicpacking interactions within the GCN4-pM3 trigger sequence,which is the hallmark of the trimerization motif (8). To test thishypothesis, a control peptide was generated in which Arg22 ofGCN4-pM3 was substituted by lysine (GCN4-pM5; Fig. 1A).As a result of its different side chain geometry, lysine is notanticipated to substitute for arginine within the trimerizationmotif. The mutation did not compromise coiled-coil formation,
A
C
B
Fig. 1. Rational design of GCN4-p1 (A), ATF1-p (B), and Cort-Ir (C) coiled-coil variants. Heptad repeats are shown as blocks of seven residues. Experimentallydetermined trigger sequences are indicated with a gray bar on top of the respective protein sequences. Arginine and glutamate residues of the native andgenerated trimerization motifs as well as the different substitutions are highlighted in color according to the amino acids’ physicochemical properties: blue,positively charged; red, negatively charged; green, hydrophobic. To prevent complications in the interpretation of results due to disulfide-bridge formation,Cys10 of ATF1-p was mutated to serine (underlined in B). The 18 heptad repeats of Cort-Ir are shown in C, Top. For simplicity, only the respective halves of theCort-Ir sequences are depicted for the trigger and control mutations. The oligomerization state of the peptides is indicated on the right of each alignment. Formolecular weight values, see Table 1.
Ciani et al. PNAS ∣ November 16, 2010 ∣ vol. 107 ∣ no. 46 ∣ 19851
BIOPH
YSICSAND
COMPU
TATIONALBIOLO
GY

with the peptide variant being more than 90% helical. Asexpected, GCN4-pM5 formed a two-stranded coiled-coil struc-ture (Fig. 2B and Table 1), demonstrating that the characteristicbifurcated ArgðgÞ-to-Gluðe0Þ (where the prime denotes a residueof a neighboring chain) salt bridge of the trimerization motif is akey element for trimer formation (8). This conclusion is furthersupported by CD thermal unfolding and sedimentation equili-brium experiments of GCN4-pM3 carried out at pH 2 where glu-tamate residues are protonated. Disruption of the network ofsurface salt bridges of the motif at low pH not only restoredthe oligomerization state of GCN4-pM3 back to a dimer but alsoresulted in a significant decrease of the Tm to 35 °C (Fig. 2B andTable 1).
Taken together, these findings demonstrate that the positionof the R-hxxhE motif within the GCN4-p1 coiled-coil domainis crucial for capitalizing on its trimerization potential. Only forthe mutant in which the trimerization motif was incorporated intothe trigger sequence a switch of the oligomerization state to thetrimer was observed, whereas the other variants were dimeric.
A General Link Between Trigger Sequence Function and Coiled-CoilOligomerization-State Specificity. The observation that incorpora-tion of the trimerization motif into the trigger sequence resultsin a switch of the oligomerization state offers the prospect thatsuch topological changes can also be generated by other inter-chain interactions. To confirm this hypothesis, we extended ourGCN4-p1 studies to the hydrophobic core. It is well establishedthat the distribution of hydrophobic core residues has an impacton the oligomerization state (see the introduction). However, in
the seminal studies by Alber, Kim, and co-workers (11, 12),all hydrophobic core positions of GCN4-p1 were simultaneouslysubstituted, and it is therefore unknown how the position ofindividual hydrophobic core residues affects the oligomerizationstate of a coiled coil.
To address this issue, we generated a GCN4-p1 variant with thepotential to fold into a trimer by substituting the central hydro-phobic core residues of the trigger sequence, Val23ðaÞ andLeu26ðdÞ, by isoleucine (GCN4-pM7; Fig. 1A). To verify that aconformational switch occurs only when mutations are intro-duced into the trigger sequence, we also generated a controlpeptide in which we made identical replacements [Val9ðaÞ andLeu12ðdÞ] in the second heptad repeat of the coiled coil (GCN4-pM6; Fig. 1A). Consistent with our previous findings (see above),introduction of isoleucine residues in the trigger sequence re-sulted in the formation of a trimer, whereas the control peptidewas dimeric (Table 1). The thermal stabilities of GCN4-pM6 andGCN4-pM7 are slightly decreased when compared with the wild-type coiled coil (Fig. 2C and Table 1), which can be explainedby an unfavorable rotamer conformation of Ile9ðaÞ in theGCN4-pM6 dimer and by the presence of Asn16ðaÞ in bothGCN4-p1 variants. In this context, it is interesting to note thatthe trigger sequence of the trimeric coiled-coil domain of thehuman macrophage scavenger receptor (173-IDEISKS) showsthe signature of a heptad repeat characteristic of trimers (173-IDEISKS) with isoleucine residues (underlined) at both the aand d positions (13), suggesting that these residues are involvedin specifying the domain’s three-stranded oligomerization state.
Our results on GCN4-p1 demonstrate that the trigger se-quence, apart from its essential role in coiled-coil folding, alsoplays an important function in controlling the protein’s oligomer-
Table 1. CD, sedimentation equilibrium AUC, and MALS data ofGCN4-p1, ATF1-p, and Cort-Ir variants
Peptide ½Θ�222103 deg cm2 dmol−1 Tm, °C * Mw , kDa†
GCN4-p1 −32.4 61 8.8GCN4-pM1 −33.4 53 9.0 ‡
GCN4-pM2 −33.3 61 8.7 ‡
GCN4-pM3 −33.1 70 (35) § 11.9 (8.5) ¶
GCN4-pM4 −33.4 69 11.3GCN4-pM5 −29.7 49 8.6GCN4-pM6 −33.4 55 7.8GCN4-pM7 −31.4 54 11.9ATF1-p −25.6 47 11.3ATF1-pM1 −25.0 57 14.0ATF1-pM2 −23.6 36 10.0ATF1-pM3 −24.5 48 10.4ATF1-pM4 −30.8 51 11.7ATF1-pM5 −21.9 47 15.0ATF1-pM6 −25.6 49 13.9ATF1-pM7 −19.6 32 13.8ATF1-pM8 −8.1 15 8.5Cort-Ir −35.0 73 28Cort-Ir-M1 −33.5 ND 43Cort-Ir-M2 −31.5 ND 44Cort-Ir-M3 −36.6 ND 28Cort-Ir-M4 −34.1 72 27Cort-Ir-M5 −38.3 ND 43Cort-Ir-M6 −35.1 ND 41Cort-Ir-M7 −34.3 71 26Cort-Ir-M8 −30.8 83 29Cort-Ir-M9 −34.1 ND 30
*Tm determined at peptide concentrations (monomer) of 35 μM.Estimated error: �1 °C. ND, not determined because protein does notunfold completely within the temperature range investigated.
†Average molecular masses determined by sedimentation equilibriumAUC. Estimated error: �10%. The calculated molecular masses of theGCN4-p1, ATF1-p, ATF1-pM4, and Cort-Ir monomers are 4.2 kDa,5.1 kDa, 3.8 kDa, and 15.0 kDa, respectively.
‡Average molecular masses determined by MALS. Error: <5%.§Tm determined at pH 2 is given in brackets.¶The average molecular mass value determined at pH 2 is given inbrackets.
-40
-35
-30
-25
-20
-15
-10
-5
0
0 20 40 60 80 100
GCN4-p1
GCN4-pM2GCN4-pM1
GCN4-pM3
-40
-35
-30
-25
-20
-15
-10
-5
0
0 20 40 60 80 100
GCN4-p1
GCN4-pM5
GCN4-pM3 at pH2GCN4-pM4
-40
-35
-30
-25
-20
-15
-10
-5
0
0 20 40 60 80 100
GCN4-pM7
GCN4-p1GCN4-pM6
-40
-35
-30
-25
-20
-15
-10
-5
0
0 20 40 60 80 100
Cort-Ir
Cort-Ir-M7Cort-Ir-M8Cort-Ir-M9
Cort-Ir-M5
-30
-25
-20
-15
-10
-5
0
0 20 40 60 80 100
ATF1-pATF1-pM1
ATF1-pM3ATF1-pM2
-35
-30
-25
-20
-15
-10
-5
0
0 20 40 60 80 100
ATF1-pM4ATF1-p
ATF1-pM5ATF1-pM6
ATF1-pM8ATF1-pM7
Temperature (°C)
[Θ] 22
2(1
03d
egcm
2d
mo
l-1)
DA
EB
FC
Fig. 2. CD analysis of GCN4-p1 (A–C), ATF1-p (D and E), and Cort-Ir (F)varaints. Thermal unfolding profiles recorded by CD at 222 nm are shown.The experiments were carried out at 35-μM peptide concentration (mono-mer) in PBS (pH 7.4) or 5-mM sodium phosphate (pH 2).
19852 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1008502107 Ciani et al.

ization state. To generalize the concept of a link between triggersequence function and oligomerization-state specificity, we inves-tigated the coiled-coil domains of the human transcriptionalactivator ATF1 and the actin-bundling protein cortexillin-1 fromD. discoideum. These two-stranded, parallel coiled coils wereselected because they also contain trimerization motifs andbecause for both domains, referred to as ATF1-p and Cort-Ir,high-resolution crystal structures are available (10, 14).
To determine the unknown ATF1-p trigger sequence, an ana-logous chimeric approach was used as previously described forGCN4-p1 (5). In these experiments, the N- and C-terminal halvesof ATF1-p were fused to the N-terminal two heptad repeats ofGCN4-p1 (Fig. S1A) and trigger sequences identified on the basisof productive coiled-coil formation. Both chimera folded intostable coiled-coil structures (Fig. S1 B and C), indicating that theentire ATF1-p coiled coil can act as a trigger sequence. Consis-tent with this hypothesis, introduction of a second trimerizationmotif into the third and fourth heptad repeat of the ATF1-psequence (ATF1-pM1; Fig. 1B) yielded a trimeric coiled coil witha significantly increased thermal stability when compared to thewild-type peptide (Fig. 2D and Table 1). However, one trimeriza-tion motif is not sufficient to switch the oligomerization state ofATF1-p from a dimer to a trimer. This is evident in the wild-typepeptide or the ATF1-pM2 and ATF1-pM3 mutants (Fig. 1B) inwhich the native or the introduced trimerization motif of ATF1-pM1, respectively, are apparently overruled (Fig. 2D and Table 1)by the presence of putatively strong dimerization determinants.
To identify determinants that could be responsible for specificdimer formation of ATF1-p apart from Asn22 at the central hep-tad repeat a position, we analyzed the protein’s crystal structure(10). The most prominent feature seen in the structure is aninterhelical hydrogen bond between Tyr7 and Glu12 at heptadrepeat positions g and e0, respectively. To elucidate whether thisdistinct structural feature is responsible for dimer formationof ATF1-p, we produced a shortened coiled-coil domain startingat Leu11 that lacks the hydrogen bond (ATF1-pM4; Fig. 1B).Molecular weight and CD measurements indicated that ATF1-pM4 forms a stable trimer (Fig. 2E and Table 1). Identical resultswere obtained after mutation of Tyr7 in the wild-type sequence(ATF1-pM5 or ATF1-pM6) or in ATF1-pM2 (ATF1-pM7)(Fig. 1B). These results indicate that the distinct hydrogen bondbetween Tyr7ðgÞ and Glu12ðe0Þ within the trigger sequencecontrols dimerization of ATF1-p (Fig. 2E and Table 1). Notably,this interaction is conserved in the ATF1 family of transcription
factors and contributes to ATF1 stability [Fig. 2E; (10)]. A cor-responding TyrðgÞ-to-Gluðe0Þ hydrogen bond is also conserved inthe coiled-coil neck of fungal kinesins where it is required forstabilization of the dimer (15). Accordingly, mutation of boththe TyrðgÞ-to-Gluðe0Þ hydrogen bond and the native trimerizationmotif of ATF1-p resulted in the formation of a significantly de-stabilized dimer (ATF1-pM8; Fig. 2E and Table 1). Dimerizationof ATF1-pM8 can be explained by the presence of Asn22 at aheptad repeat a position.
Next we performed corresponding experiments with the two-stranded Cort-Ir coiled-coil domain. This coiled coil has beenstudied extensively and its trigger sequence is well characterized(6, 14). Because of its substantial size (18 continuous heptadrepeats), we used a combination of both trimer-specific determi-nants for the design of the mutants. Accordingly, we rationallysubstituted residues to complement two ideal trimerizer motifsof the type R-IxxIE and to introduce one hydrophobic core iso-leucine residue in the trigger sequence of the Cort-Ir coiled coil(Cort-Ir-M1; Fig. 1C). Notably these substitutions were designedto span the entire trigger sequence. Consistent with our results onGCN4-p1 and ATF1-p, the trigger sequence mutant folded into astable three-stranded coiled-coil structure (Table 1 and Fig. S2).To determine the minimal number of mutations required toswitch the peptide’s oligomerization state from a dimer to a tri-mer, we produced two mutants, Cort-Ir-M2 and Cort-Ir-M3(Fig. 1C), which each contain one ideal R-IxxIE trimerizationmotif and one isoleucine substitution. Molecular weight measure-ments demonstrated that Cort-Ir-M2 formed a trimer, whereasCort-Ir-M3 was dimeric (Table 1 and Fig. S2). To exclude thatthe putative salt bridge between Arg91ðgÞ and Glu96ðe0Þ in thetrigger sequence of Cort-Ir contributes to trimer formation, itwas mutated (Cort-Ir-M5 and Cort-Ir-M6; Fig. 1C). AUC analysisdemonstrated that the introduced ideal trimerization motif issufficient to switch the dimer to a trimer (Table 1, Fig. 3, andFigs. S2 and S3). However, mutation of Glu81 to isoleucine (com-pare Cort-Ir-M5/M6 with Cort-Ir-M4; Fig. 1C) is required forthe switch of the oligomerization state (Table 1 and Fig. S2).This finding suggests that the interhelical salt bridge betweenGlu81ðdÞ and Arg85ða0Þ seen in the crystal structure of Cort-Ir(14) is a strong determinant of specific dimer formation of thecoiled coil that can overrule the presence of one ideal trimerizer.
As in the experiments described for GCN4-p1, Cort-Ir controlswere prepared in which identical replacements weremade outsideof the trigger sequence (Cort-Ir-M7, Cort-Ir-M8, and Cort-Ir-M9;
0.0
0.2
0.4
0.6
0.8
Res
idu
als
Ab
sorp
tio
n
27krpm 275nm 27krpm 235nm
Cort-Ir-M8Cort-Ir-M6 27krpm 275nm 27krpm 235nm
27krpm 275nm 27krpm 275nm
6.80 6.85 6.90 6.95 7.00 7.05 7.10
Radius (cm)
27krpm 235nm
6.80 6.85 6.90 6.95 7.00 7.05 7.10
Radius (cm)
27krpm 235nm
Fig. 3. Analysis of the oligomerization state of Cort-Ir variants by sedimentation equilibrium AUC. The sequences of Cort-Ir-M6 and Cort-Ir-M8 are shown inFig. 1C. (Upper) UV absorbance gradients as a function of the radial position (data points) and fits according to a single ideal species model (lines). (Lower)Residuals showing the difference between the experimental data and the theoretical model. The fitted values are listed in Table 1.
Ciani et al. PNAS ∣ November 16, 2010 ∣ vol. 107 ∣ no. 46 ∣ 19853
BIOPH
YSICSAND
COMPU
TATIONALBIOLO
GY

Fig. 1C). These controls were designed to cover a spectrum ofdifferent mutations and notably in Cort-Ir-M8 the same a and dresidues as in Cort-Ir-M2 were replaced by isoleucine. In agree-ment with our results on GCN4-p1, all controls were dimeric likeCort-Ir (Table 1, Figs. 2Fand 3, and Fig. S3). The observation thatC-terminal Cort-Ir mutants do not completely unfold up to 90 °Cwhereas N-terminal control peptides are less stable indicatesan important role of theC terminus in coiled-coil stability (Table 1,Fig. 2F, and Fig. S2).
Taken together, these results suggest that oligomerization-statedeterminants specify the topology of a coiled coil only when pre-sent in the trigger sequence. They therefore establish a generallink between trigger sequence function and oligomerization-statespecificity. Our data further uncovered two previously unde-scribed dimerizationmotifs, the TyrðgÞ-Gluðe0Þ interhelical hydro-gen bond and the GluðdÞ-Argða0Þ interhelical salt bridge, withinthe trigger sequences of ATF1-p and Cort-Ir, respectively. Theobservation that multiple motifs can coexist in the sametrigger sequence reveals a delicate balance of position-dependentforces that control a specific coiled-coil topology. Furthermore,we found that the trigger sequence of ATF1-p extends overfour rather than the approximately two heptad repeats character-istic of GCN4-p1 and Cort-Ir. This observation indicates thattrigger sequences can not only vary with respect to sequence (7)but also in terms of length.
Structural Basis of Trimer Formation of GCN4-p1 Variants. To under-stand the molecular basis of how the trimerization motif R-hxxhEor isoleucine substitutions at hydrophobic core positions in thetrigger sequence of the GCN4-p1 result in a switch of conforma-tion from dimer to trimer and how these mutations overrulethe dimerization potential of Asn16, we determined the crystalstructures of GCN4-pM3 and GCN4-pM7 at 1.9- and 2.0-Åresolution, respectively. The overall structures of both trimersdisplay all characteristics of left-handed, parallel, and in-registercoiled coils (Fig. S4).
The most prominent feature seen in the crystal structureof GCN4-pM3 is a network of surface salt bridges and internalhydrophobic packing interactions in the vicinity of the trimeriza-tion motif, which are similar to those described for the parallelthree-stranded coiled-coil domain of coronin-1 (Fig. 4 A–C) (8).Accordingly, the conformations of side chains and the positionof the water molecule between Arg22 and Glu27′ can be almostperfectly superimposed to the trimerization motif of the coronin-1 coiled coil, demonstrating structural conservation of the motif.In addition to the characteristic bifurcated contact betweenArg22∶Nε, Nη2 at position g of one chain and Glu270∶Oε1, Oε2at position e0 of the neighboring chain, an interaction betweenArg22∶Nη1 and Glu200∶Oε1 at position e0 extends the networkof surface salt bridges (Fig. 4A and Fig. S4).
The interior packing of GCN4-pM7 at layers a23 and d26 pro-vides a structural basis for the switch of the oligomerization statefrom a dimer to a trimer (Fig. 4D and E). In the structure the twointroduced isoleucine residues can be accommodated in the mostpreferred rotamer at the hydrophobic core positions. Accord-ingly, the acute packing geometries of Ile23 and Ile26 are verysimilar to those seen in the crystal structure of the GCN4-pIIvariant (Fig. 4 D and E) (12).
In contrast to the favorable interactions seen in the two struc-tures, the polar side chain of Asn16 at the central a positionpoints out of the hydrophobic core, generating both an energe-tically unfavorable cavity and compromised packing contactsagainst the hydrophobic moiety of Lys15 at the preceding g0 posi-tion (Fig. 4F). In contrast, in dimers, asparagine residues pointinto the hydrophobic interface and form a distinctive hydrogenbond (9). These findings therefore provide a structural explana-tion why an asparagine at core a positions favors dimer overtrimer formation.
DiscussionDetailed knowledge of the oligomerization state of coiled coils iscrucial for exploiting their potential in a wide range of applica-tions and understanding their functions in a large number ofbiological processes. Over the years, several oligomerization-state determinants have been identified and studied in detail;however, their presence does frequently not correlate with coiled-coil topology. Here, using three diverse coiled-coil model systemswe addressed this discrepancy and demonstrated that oligomer-ization-state determinants specify the topology of a coiled coilwhen present in the trigger sequence. Furthermore, we show that
Ile23
Arg22
Val23'
Ala24'
Glu27'
Leu26
Arg25
D
Arg22Arg450
Glu20'
Glu27'Glu455'
Asp452'
Ile26
Lys15
Asn16‘Tyr17'
F
C
E
BA
Fig. 4. Packing of side chains within X-ray crystal structures of GCN4-pM3and GCN4-pM7. (A) Side view of an overlay of the salt-bridge network(indicated by yellow dots) of the trimerization motifs seen in GCN4-pM3and the coiled-coil domain of coronin-1 (8) (dark gray ribbons). (B) Axial viewof the a4 layer of GCN4-pM3 showing the shielding of the Val23′ residuesfrom solvent by the aliphatic side-chain moieties of Arg22. (C) Axial viewof the d4 layer of GCN4-pM3 showing the hydrophobic packing of Ile26residues and the aliphatic side-chain moieties of the Glu27′. (D and E) Axialviews of an overlay of the acute packing of isoleucine residues at the a4 andd4 layers seen in GCN4-pM7 and GCN4-pII (dark gray) (12). (F) Axial view ofthe a3 layer showing the conformation of the Asn16 residues and the unfa-vorable packing of their polar moieties against the aliphatic portion of Lys15′as well as the formation of a large cavity in the hydrophobic core of GCN4-pM3. A similar conformation is seen in the crystal structure of GCN4-pM7.Side chains of residues are shown as stick representation (A–F) and vander Waals spheres (B, C, and F). Monomers are shown as Cα traces and theboundwater molecules as a small red (GCN4-pM3) or gray (coiled-coil domainof coronin-1) sphere (A). Oxygen and nitrogen atoms are colored in red andblue, and carbon atoms are shown in green, cyan, and yellow for GCN4-pM3and GCN4-pM7 monomers A, B, and C, respectively. The prime indicates aresidue of chain B.
19854 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1008502107 Ciani et al.

multiple motifs can coexisit in the same trigger sequence andthe resulting oligomerization state is controlled by the strengthof the determinants relative to each other. The contribution ofthe relative strength of individual motifs to final coiled-coil topol-ogy remains to be elucidated.
Our findings on a unique and general link between oligomer-ization-state specificity and trigger sequence function explainsome puzzling results reported in the literature. We previouslyshowed that a chimera in which the trigger sequence of GCN4was replaced by the one of cortexillin-1 (GCN4p-Cort T/L; notethat Thr95 of Cort-Ir has been replaced by leucine to result bychance in a canonical trimerization motif) folds into a three-stranded coiled-coil structure despite the presence of Asn16ðaÞ(5). This finding can now be rationalized by the presence of atrimerization motif (R-LAKLE) in the GCN4p-Cort T/L triggersequence. Further, the coiled-coil domain of the human dystro-phia myotonica kinase contains a motif (509-R-NRDLE), whichis very similar to the R-hxxhE consensus. Although it is currentlynot known how mutations of the hydrophobic residues of the tri-merization motif influence its function, the motif of the humandystrophia myotonica kinase is a good candidate to determine tri-merization of the coiled coil by overruling the strong dimerizationdriving force of two asparagine residues at heptad a positions inthe sequence (16). Another example is the coiled-coil domain ofthe transcription factor HY5 fromArabidopsis thaliana, which wasreported to harbor two putative and overlapping consensustrigger sequences (6-SELENR-VKDLENK and 13-KDLENK-N-SELEER) containing a trimerization motif (underlined in thefirst sequence stretch) and an asparagine residue at a heptadrepeat a position (underlined in the second sequence stretch),respectively (17). A second asparagine residue, Asn33, is presentat heptad repeat a position outside the predicted trigger se-quences. The wild-type peptide is dimeric but substitution ofone or both asparagine by leucine or valine resulted in a switchof the oligomerization state from a dimer to a trimer (17). Asimilar context-dependent effect of the trimerization motif (591-R-LNRVE) was recently also demonstrated for the K(V)7.1(KCNQ1) A-domain tail coiled coil, where C-terminal truncationresulted in a switch of the oligomerization state from a tetramerto a trimer (18).
In conclusion, our study represents an important complemen-tation and substantial extension of the sequence-to-structurerules outlined in the pioneering work of Alber, Kim, and collea-gues (11, 12). These rules together with the characteristics ofthe heptad repeat have allowed the development of a variety ofstatistics- and pattern-based methods that predict the occurrenceof coiled coils with a high degree of confidence (reviewed inref. 19). However, despite the numerous efforts during the last
three decades, a reliable prediction of the specific oligomeriza-tion state of many native coiled coils remains difficult (1). Thishighlights the need for improving existing algorithms by imple-mentation of experimentally verified sequence-to-structure ruleslike the ones based on the present study. The inclusion of theseparameters as well as the detailed dataset obtained in our studyshould therefore significantly improve and extend the kit towardpredicting and engineering the structure and function of coiled-coil proteins, which should have major implications for all appli-cations using these popular oligomerization domains.
MethodsPeptide Preparations and Crystal Structure Determination. Standard recombi-nant peptide preparation and crystal structure determination is described inSI Text.
AUC, MALS, and CD Spectroscopy. All solution studies were performed in 5mMsodium phosphate, pH 7.4, supplemented with 150 mM PBS.
AUC was carried out on an Optima XLA analytical ultracentrifuge (Beck-man) equipped with an adsorption optical system and an An-60Ti (GCN4-p1,ATF-p) or an An-50Ti (Cort-Ir) rotor. Sedimentation equilibrium data were col-lected at three different protein concentrations (0.14, 0.27, and 0.55 mg∕mLfor GCN4-p1 variants and 0.15, 0.3, and 0.6 mg∕mL for ATF1-p variants) andat rotor speeds of 28,000, 35,000, and 41,000 rpm. TheMW reported in Table 1is calculated at the highest sample concentration. Sedimentation equilibriumdata of Cort-Ir were recorded at three different protein concentrations (0.25,0.5, and 1.0 mg∕mL) and at rotor speeds of 27,000, 31,000, and 35,000 rpm.The sedimentation profiles were monitored at two wavelengths (275 nm fortyrosine and 235 nm for peptide backbone absorption). The data were fittedglobally using a single ideal species model. The partial specific volumes of thesynthetic peptides were calculated from their amino acid sequence composi-tion. Solvent density for PBS was taken as 1.004 g∕mL.
Size exclusion chromatography coupled to MALS was performed on aDAWN EOS 18-angle detector connected to an Optilab Rex refractometer(Wyatt). Peptide solutions (100 μL of 3–5 mg∕mL) were injected on a Super-dex 75 10∕30 size exclusion chromatography column equilibrated with PBS.Molecular weights were calculated by using theWyatt ASTRA version 4.90.08software package.
Far-UV CD spectroscopy was carried out on Jasco J-810 (Jasco Inc.; GCN4-p1and ATF-p variants) or Chirascan-plus (Applied Photophysics; Cort-Ir variants)spectropolarimeters equipped with temperature-controlled quartz cells of0.1-cm path length. A ramping rate of 1 °C per min was used to recordthe thermal unfolding profiles. Midpoints of the transitions, Tms, were takenas the maximum of the derivative d½θ�222∕dT .
ACKNOWLEDGMENTS. We are indebted to Ariel Lustig for performing initialAUC measurements. We thank Edward McKenzie, Sanjai Patel, KarolinaZielinska, and the Biomolecular Analysis and Crystallization Core Facilities(Faculty of Life Sciences, University of Manchester) for excellent technicalassistance. M.O.S. and R.A.K. acknowledge support by grants from the SwissNational Science Foundation and the Wellcome Trust, respectively.
1. Lupas AN, Gruber M (2005) The structure of alpha-helical coiled coils. Adv ProteinChem 70:37–38.
2. Woolfson DN (2005) The design of coiled-coil structures and assemblies. Adv ProteinChem 70:79–112.
3. Sharma VA, Logan J, King DS, White R, Alber T (1998) Sequence-based design of apeptide probe for the APC tumor suppressor protein. Curr Biol 8:823–830.
4. Bianchi E, et al. (2005) Covalent stabilization of coiled coils of the HIV gp41 N regionyields extremely potent and broad inhibitors of viral infection. Proc Natl Acad Sci USA102:12903–12908.
5. Kammerer RA, et al. (1998) An autonomous folding unit mediates the assembly oftwo-stranded coiled coils. Proc Natl Acad Sci USA 95:13419–13424.
6. Steinmetz MO, et al. (1998) A distinct 14 residue site triggers coiled-coil formation incortexillin I. EMBO J 17:1883–1891.
7. SteinmetzMO, et al. (2007) Molecular basis of coiled-coil formation. Proc Natl Acad SciUSA 104:7062–7067.
8. Kammerer RA, et al. (2005) A conserved trimerization motif controls the topology ofshort coiled coils. Proc Natl Acad Sci USA 102:13891–13896.
9. O’Shea EK, Klemm JD, Kim PS, Alber T (1991) X-ray structure of the GCN4 leucinezipper, a two-stranded, parallel coiled coil. Science 254:539–544.
10. Schumacher MA, Goodman RH, Brennan RG (2000) The structure of a CREB bZIP.so-matostatin CRE complex reveals the basis for selective dimerization and divalentcation-enhanced DNA binding. J Biol Chem 275:35242–35247.
11. Harbury PB, Zhang T, Kim PS, Alber T (1993) A switch between two-, three-, andfour-stranded coiled coils in GCN4 leucine zipper mutants. Science 262:1401–1407.
12. Harbury PB, Kim PS, Alber T (1994) Crystal structure of an isoleucine-zipper trimer.Nature 371:80–83.
13. Frank S, Lustig A, Schulthess T, Engel J, Kammerer RA (2000) A distinct seven-residue trigger sequence is indispensable for proper coiled-coil formation of thehuman macrophage scavenger receptor oligomerization domain. J Biol Chem275:11672–11677.
14. Burkhard P, Kammerer RA, SteinmetzMO, Bourenkov GP, Aebi U (2000) The coiled-coiltrigger site of the rod domain of cortexillin I unveils a distinct network of interhelicaland intrahelical salt bridges. Struct Fold Des 8:223–230.
15. Schafer F, et al. (2003) A conserved tyrosine in the neck of a fungal kinesin regulatesthe catalytic motor core. EMBO J 22:450–458.
16. Garcia P, et al. (2006) Molecular insights into the self-assembly mechanism of dystro-phia myotonica kinase. FASEB J 20:1142–1151.
17. Yoon MK, Kim HM, Choi G, Lee JO, Choi BS (2007) Structural basis for the conforma-tional integrity of the Arabidopsis thaliana HY5 leucine zipper homodimer. J BiolChem 282:12989–13002.
18. Xu Q, Minor DL, Jr (2009) Crystal structure of a trimeric form of the K(V)7.1 (KCNQ1)A-domain tail coiled-coil reveals structural plasticity and context dependent changesin a putative coiled-coil trimerization motif. Protein Sci 18:2100–2114.
19. Gruber M, Soding J, Lupas AN (2006) Comparative analysis of coiled-coil predictionmethods. J Struct Biol 155:140–145.
Ciani et al. PNAS ∣ November 16, 2010 ∣ vol. 107 ∣ no. 46 ∣ 19855
BIOPH
YSICSAND
COMPU
TATIONALBIOLO
GY
Copyright © 2022 FDOKUMEN




















