
Ein mithilfe synthetischer GCN4-Leucinzipperarrays aufgedecktes Coiled-Coil-Assoziationsnetzwerk
17
Proteinstrukturen DOI: 10.1002/ange.200603246 Ein mithilfe synthetischer GCN4-Leucinzipperarrays aufgedecktes Coiled-Coil-Assoziationsnetzwerk** Michael Portwich, Sandro Keller, HolgerM. Strauss, Carsten C. Mahrenholz, Ines Kretzschmar, Achim Kramer und Rudolf Volkmer* Das a-helicale Coiled-Coil gehɆrt zu den ersten Proteinmo- tiven, deren Strukturen im Detail aufgeklȨrt wurden. [1] Wie bereits 1953 von Crick beschrieben, [2] besteht ein Coiled-Coil aus mindestens zwei rechtsgȨngigen amphipathischen a-He- lices, die sich umeinander zu einem linksgȨngigen Supercoil winden und so ihre hydrophoben OberflȨchen miteinander in Kontakt bringen (Abbildung 1 a). Coiled-Coils kɆnnen bis zu Pentameren assoziieren und bilden homo- oder heteromere, parallele oder antiparallele Komplexe mit unterschiedlichen StɆchiometrien. [1] Ein Charakteristikum aller Coiled-Coils ist das Vorhandensein heptadischer Wiederholungssequenzen [abcdefg] i , wobei i die jeweilige Heptade bezeichnet (Abbil- dung 1a). Hydrophobe AminosȨuren finden sich normaler- weise an den Kernpositionen a und d und sind fɒr die Faltung entscheidend. [3] Die Positionen b, c, e, f und g bilden die Außenseite der Struktur und werden im Allgemeinen durch hydrophile Reste besetzt. ZusȨtzlich kɆnnen sich durch ge- ladene Reste inter- und intrahelicale Salzbrɒcken bilden, die hȨufig an den Positionen b, c, e und g gefunden werden und die Struktur stabilisieren. [4] Die „Peptid-Klettverschluss“- Hypothese [5] fasst diese Erkenntnisse zusammen und postu- liert, dass 1) a und d hydrophob, 2) e und g geladen und 3) b, c und f hydrophil sind. Obgleich Coiled-Coil-Motive mit einem hohen Maß an Sicherheit vorhergesagt werden kɆnnen, [6] ist die Vorhersage ihrer StɆchiometrie und rȨumlichen Struktur noch immer eine schwierige Aufgabe. Fɒr das Durchmustern von rekombinant hergestellten Coiled-Coil-Komplexen sind unterschiedliche Herangehens- weisen mɆglich. [7] Die Arbeitsgruppen von Plɒckthun und Michnick entwickelten einen eindrucksvollen „library-versus- library“-Ansatz zur Analyse von Polypeptiden, die ɒber ein Coiled-Coil-Motiv interagieren. [8] Kɒrzlich wurde eine Glas-Chip-Technologie angewendet, um ein Interaktionsnetzwerk aus bZIP-Transkriptionsfakto- ren auf Proteinniveau sichtbar zu machen. [9] Hier berichten wir ɒber den Einsatz optimierter synthetischer Peptidar- rays [10] mit dem Ziel, Coiled-Coil-Verbindungen auf Amino- sȨureebene zu analysieren und zu charakterisieren. Unser Ansatz beruht – im Unterschied zu rationalem Design, das meistens kurze Modellpeptide als Anschauungsobjekte nutzt – auf einem in voller LȨnge vorliegenden, homodimeren Coiled-Coil, dem GCN4-Leucinzipper (Abbildung 1 a). [11] Die Einflɒsse von AminosȨuresubstitutionen auf die Asso- ziation werden analysiert, und zusȨtzlich werden die StɆ- chiometrien mittels biophysikalischer Methoden ɒberprɒft. Es ist zu erwarten, dass die Resultate eine bessere und zu- verlȨssigere Voraussagbarkeit von Coiled-Coil-Interaktionen ermɆglichen werden. Zuerst synthetisierten wir ein Peptidarray aus 589 Ein- zelsubstitutionsvarianten des GCN4-Leucinzippers auf Zel- lulosemembranen [12] und prɒften deren FȨhigkeit, an die native GCN4-Wildtypsequenz (wt) zu binden (Tabelle 1, Abbildung 1 b). Die SignalintensitȨten der einzelnen Spots wurden gemessen und wie beschrieben ausgewertet. [13, 14] Die AustauschvariabilitȨt jeder Sequenzposition wurde berech- net [14] (Abbildung 1 c) und als niedrig (V 20 %), mittel (20 % V 50 %) oder hoch (V 50 %) eingestuft. Die im Kern liegenden Leucinreste d I -d IV zeigten erwartungsgemȨß sehr geringe VariabilitȨt hinsichtlich einer Substitution; eine Ausnahme bildete das durch Alanin ersetzbare Leu12 (d II ). Dagegen weisen die Kernpositionen a I -a V mittlere Variabili- tȨt auf; diese Klasse ist durch eine erweiterte physikochemi- sche Ɛhnlichkeit [14] der austauschbaren AminosȨuren cha- Tabelle 1: Sequenzen ausgesuchter GCN4-Varianten. g abcdefg abcdefg abcdefg abcdefg ab I II III IV wt [a] R MKQLEDK VEELLSK NYHLENE VARLKKL VG 1 R MKQLEDK VEELLSK NYHLENE KARL EKL VG 2 R MKQLEDK VEELLSK YYH TENE VARLKKL VG 3 R MKQLEDK VEELLSK IYH NENE VARLKKL VG [a] Die Sequenz entspricht den Resten 249–279 des Proteins GCN4 aus Saccharomyces cerevisiae. Informationen zur Synthese kɆnnen den Hin- tergrundinformationen, S1 und S2, entnommen werden. [*] Dr. M. Portwich, [$] C. C. Mahrenholz, I. Kretzschmar, Prof.Dr. A. Kramer, Dr. R. Volkmer Institut fɒr Medizinische Immunologie CharitȖ-UniversitȨtsmedizin Berlin Campus CharitȖ Mitte Schumannstraße 20–21, 10117 Berlin (Deutschland) Fax: (+ 49) 30-450-524942 E-Mail: [email protected] Homepage: http://www.charite.de/immunologie/research/agsm/ index.html Dr. S. Keller, [$] Dr. H. M. Strauss [+] [$] Leibniz-Institut fɒr Molekulare Pharmakologie FMP Robert-RɆssle-Straße 10, 13125 Berlin (Deutschland) [ + ] Aktuelle Adresse: Max-Planck-Institut fɒr Kolloide und GrenzflȨchen Am Mɒhlenberg 1, 14424 Golm (Deutschland) [ $ ] Diese Autoren haben gleichrangig zum Gelingen dieser Arbeit bei- getragen. [**] Wir danken der Deutschen Forschungsgemeinschaft (SFB 449, SFB 498), der Schering-Stiftung, der Jɒrgen-Manchot-Stiftung und dem UniversitȨtsklinikum CharitȖ fɒr die finanzielle Unterstɒtzung. Hintergrundinformationen zu diesem Beitrag sind im WWW unter http://www.angewandte.de zu finden oder kɆnnen beim Autor angefordert werden. Zuschriften 1682 # 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Angew. Chem. 2007, 119, 1682 –1686
-
Upload
charite-de -
Category
Documents
-
view
1 -
download
0
Transcript of Ein mithilfe synthetischer GCN4-Leucinzipperarrays aufgedecktes Coiled-Coil-Assoziationsnetzwerk

ProteinstrukturenDOI: 10.1002/ange.200603246
Ein mithilfe synthetischer GCN4-Leucinzipperarrays aufgedecktesCoiled-Coil-Assoziationsnetzwerk**Michael Portwich, Sandro Keller, Holger M. Strauss, Carsten C. Mahrenholz, Ines Kretzschmar,Achim Kramer und Rudolf Volkmer*
Das a-helicale Coiled-Coil geh�rt zu den ersten Proteinmo-tiven, deren Strukturen im Detail aufgekl�rt wurden.[1] Wiebereits 1953 von Crick beschrieben,[2] besteht ein Coiled-Coilaus mindestens zwei rechtsg�ngigen amphipathischen a-He-lices, die sich umeinander zu einem linksg�ngigen Supercoilwinden und so ihre hydrophoben Oberfl�chen miteinander inKontakt bringen (Abbildung 1a). Coiled-Coils k�nnen bis zuPentameren assoziieren und bilden homo- oder heteromere,parallele oder antiparallele Komplexe mit unterschiedlichenSt�chiometrien.[1] Ein Charakteristikum aller Coiled-Coils istdas Vorhandensein heptadischer Wiederholungssequenzen[abcdefg]i, wobei i die jeweilige Heptade bezeichnet (Abbil-dung 1a). Hydrophobe Aminos�uren finden sich normaler-weise an den Kernpositionen a und d und sind f4r die Faltungentscheidend.[3] Die Positionen b, c, e, f und g bilden dieAußenseite der Struktur und werden im Allgemeinen durchhydrophile Reste besetzt. Zus�tzlich k�nnen sich durch ge-ladene Reste inter- und intrahelicale Salzbr4cken bilden, dieh�ufig an den Positionen b, c, e und g gefunden werden unddie Struktur stabilisieren.[4] Die „Peptid-Klettverschluss“-Hypothese[5] fasst diese Erkenntnisse zusammen und postu-liert, dass 1) a und d hydrophob, 2) e und g geladen und 3) b, cund f hydrophil sind. Obgleich Coiled-Coil-Motive mit einemhohen Maß an Sicherheit vorhergesagt werden k�nnen,[6] istdie Vorhersage ihrer St�chiometrie und r�umlichen Strukturnoch immer eine schwierige Aufgabe.
F4r das Durchmustern von rekombinant hergestelltenCoiled-Coil-Komplexen sind unterschiedliche Herangehens-weisen m�glich.[7] Die Arbeitsgruppen von Pl4ckthun undMichnick entwickelten einen eindrucksvollen „library-versus-library“-Ansatz zur Analyse von Polypeptiden, die 4ber einCoiled-Coil-Motiv interagieren.[8]
K4rzlich wurde eine Glas-Chip-Technologie angewendet,um ein Interaktionsnetzwerk aus bZIP-Transkriptionsfakto-ren auf Proteinniveau sichtbar zu machen.[9] Hier berichtenwir 4ber den Einsatz optimierter synthetischer Peptidar-rays[10] mit dem Ziel, Coiled-Coil-Verbindungen auf Amino-s�ureebene zu analysieren und zu charakterisieren. UnserAnsatz beruht – im Unterschied zu rationalem Design, dasmeistens kurze Modellpeptide als Anschauungsobjekte nutzt– auf einem in voller L�nge vorliegenden, homodimerenCoiled-Coil, dem GCN4-Leucinzipper (Abbildung 1a).[11]
Die Einfl4sse von Aminos�uresubstitutionen auf die Asso-ziation werden analysiert, und zus�tzlich werden die St�-chiometrien mittels biophysikalischer Methoden 4berpr4ft.Es ist zu erwarten, dass die Resultate eine bessere und zu-verl�ssigere Voraussagbarkeit von Coiled-Coil-Interaktionenerm�glichen werden.
Zuerst synthetisierten wir ein Peptidarray aus 589 Ein-zelsubstitutionsvarianten des GCN4-Leucinzippers auf Zel-lulosemembranen[12] und pr4ften deren F�higkeit, an dienative GCN4-Wildtypsequenz (wt) zu binden (Tabelle 1,Abbildung 1b). Die Signalintensit�ten der einzelnen Spotswurden gemessen und wie beschrieben ausgewertet.[13,14] DieAustauschvariabilit�t jeder Sequenzposition wurde berech-net[14] (Abbildung 1c) und als niedrig (V� 20%), mittel(20%�V� 50%) oder hoch (V� 50%) eingestuft. Die imKern liegenden Leucinreste dI-dIV zeigten erwartungsgem�ßsehr geringe Variabilit�t hinsichtlich einer Substitution; eineAusnahme bildete das durch Alanin ersetzbare Leu12 (dII).Dagegen weisen die Kernpositionen aI-aV mittlere Variabili-t�t auf; diese Klasse ist durch eine erweiterte physikochemi-sche Jhnlichkeit[14] der austauschbaren Aminos�uren cha-
Tabelle 1: Sequenzen ausgesuchter GCN4-Varianten.
gabcdefgabcdefgabcdefgabcdefgab
I II III IV
wt[a] RMKQLEDKVEELLSKNYHLENEVARLKKLVG
1 RMKQLEDKVEELLSKNYHLENEKARLEKLVG
2 RMKQLEDKVEELLSKYYHTENEVARLKKLVG
3 RMKQLEDKVEELLSKIYHNENEVARLKKLVG
[a] Die Sequenz entspricht den Resten 249–279 des Proteins GCN4 ausSaccharomyces cerevisiae. Informationen zur Synthese k.nnen den Hin-tergrundinformationen, S1 und S2, entnommen werden.
[*] Dr. M. Portwich,[$] C. C. Mahrenholz, I. Kretzschmar,Prof. Dr. A. Kramer, Dr. R. VolkmerInstitut f8r Medizinische ImmunologieCharit9-Universit<tsmedizin BerlinCampus Charit9 MitteSchumannstraße 20–21, 10117 Berlin (Deutschland)Fax: (+49)30-450-524942E-Mail : [email protected]: http://www.charite.de/immunologie/research/agsm/
index.html
Dr. S. Keller,[$] Dr. H. M. Strauss[+] [$]
Leibniz-Institut f8r Molekulare Pharmakologie FMPRobert-R.ssle-Straße 10, 13125 Berlin (Deutschland)
[+] Aktuelle Adresse:Max-Planck-Institut f8r Kolloide und Grenzfl<chenAm M8hlenberg 1, 14424 Golm (Deutschland)
[$] Diese Autoren haben gleichrangig zum Gelingen dieser Arbeit bei-getragen.
[**] Wir danken der Deutschen Forschungsgemeinschaft (SFB 449, SFB498), der Schering-Stiftung, der J8rgen-Manchot-Stiftung und demUniversit<tsklinikum Charit9 f8r die finanzielle Unterst8tzung.
Hintergrundinformationen zu diesem Beitrag sind im WWW unterhttp://www.angewandte.de zu finden oder k.nnen beim Autorangefordert werden.
Zuschriften
1682 � 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Angew. Chem. 2007, 119, 1682 –1686
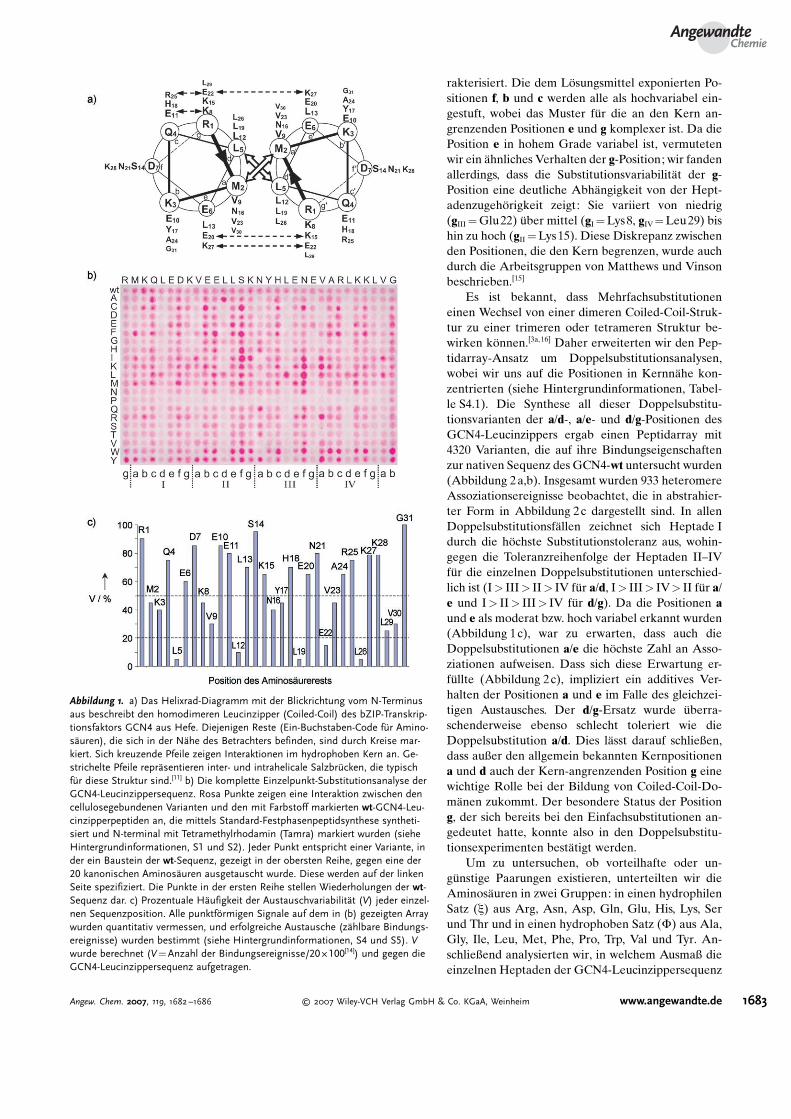
rakterisiert. Die dem L�sungsmittel exponierten Po-sitionen f, b und c werden alle als hochvariabel ein-gestuft, wobei das Muster f4r die an den Kern an-grenzenden Positionen e und g komplexer ist. Da diePosition e in hohem Grade variabel ist, vermutetenwir ein �hnliches Verhalten der g-Position; wir fandenallerdings, dass die Substitutionsvariabilit�t der g-Position eine deutliche Abh�ngigkeit von der Hept-adenzugeh�rigkeit zeigt: Sie variiert von niedrig(gIII=Glu22) 4ber mittel (gI=Lys8, gIV=Leu29) bishin zu hoch (gII=Lys15). Diese Diskrepanz zwischenden Positionen, die den Kern begrenzen, wurde auchdurch die Arbeitsgruppen von Matthews und Vinsonbeschrieben.[15]
Es ist bekannt, dass Mehrfachsubstitutioneneinen Wechsel von einer dimeren Coiled-Coil-Struk-tur zu einer trimeren oder tetrameren Struktur be-wirken k�nnen.[3a,16] Daher erweiterten wir den Pep-tidarray-Ansatz um Doppelsubstitutionsanalysen,wobei wir uns auf die Positionen in Kernn�he kon-zentrierten (siehe Hintergrundinformationen, Tabel-le S4.1). Die Synthese all dieser Doppelsubstitu-tionsvarianten der a/d-, a/e- und d/g-Positionen desGCN4-Leucinzippers ergab einen Peptidarray mit4320 Varianten, die auf ihre Bindungseigenschaftenzur nativen Sequenz des GCN4-wt untersucht wurden(Abbildung 2a,b). Insgesamt wurden 933 heteromereAssoziationsereignisse beobachtet, die in abstrahier-ter Form in Abbildung 2c dargestellt sind. In allenDoppelsubstitutionsf�llen zeichnet sich Heptade Idurch die h�chste Substitutionstoleranz aus, wohin-gegen die Toleranzreihenfolge der Heptaden II–IVf4r die einzelnen Doppelsubstitutionen unterschied-lich ist (I> III> II> IV f4r a/d, I> III> IV> II f4r a/e und I> II> III> IV f4r d/g). Da die Positionen aund e als moderat bzw. hoch variabel erkannt wurden(Abbildung 1c), war zu erwarten, dass auch dieDoppelsubstitutionen a/e die h�chste Zahl an Asso-ziationen aufweisen. Dass sich diese Erwartung er-f4llte (Abbildung 2c), impliziert ein additives Ver-halten der Positionen a und e im Falle des gleichzei-tigen Austausches. Der d/g-Ersatz wurde 4berra-schenderweise ebenso schlecht toleriert wie dieDoppelsubstitution a/d. Dies l�sst darauf schließen,dass außer den allgemein bekannten Kernpositionena und d auch der Kern-angrenzenden Position g einewichtige Rolle bei der Bildung von Coiled-Coil-Do-m�nen zukommt. Der besondere Status der Positiong, der sich bereits bei den Einfachsubstitutionen an-gedeutet hatte, konnte also in den Doppelsubstitu-tionsexperimenten best�tigt werden.
Um zu untersuchen, ob vorteilhafte oder un-g4nstige Paarungen existieren, unterteilten wir dieAminos�uren in zwei Gruppen: in einen hydrophilenSatz (x) aus Arg, Asn, Asp, Gln, Glu, His, Lys, Serund Thr und in einen hydrophoben Satz (F) aus Ala,Gly, Ile, Leu, Met, Phe, Pro, Trp, Val und Tyr. An-schließend analysierten wir, in welchem Ausmaß dieeinzelnen Heptaden der GCN4-Leucinzippersequenz
Abbildung 1. a) Das Helixrad-Diagramm mit der Blickrichtung vom N-Terminusaus beschreibt den homodimeren Leucinzipper (Coiled-Coil) des bZIP-Transkrip-tionsfaktors GCN4 aus Hefe. Diejenigen Reste (Ein-Buchstaben-Code f8r Amino-s<uren), die sich in der N<he des Betrachters befinden, sind durch Kreise mar-kiert. Sich kreuzende Pfeile zeigen Interaktionen im hydrophoben Kern an. Ge-strichelte Pfeile repr<sentieren inter- und intrahelicale Salzbr8cken, die typischf8r diese Struktur sind.[11] b) Die komplette Einzelpunkt-Substitutionsanalyse derGCN4-Leucinzippersequenz. Rosa Punkte zeigen eine Interaktion zwischen dencellulosegebundenen Varianten und den mit Farbstoff markierten wt-GCN4-Leu-cinzipperpeptiden an, die mittels Standard-Festphasenpeptidsynthese syntheti-siert und N-terminal mit Tetramethylrhodamin (Tamra) markiert wurden (sieheHintergrundinformationen, S1 und S2). Jeder Punkt entspricht einer Variante, inder ein Baustein der wt-Sequenz, gezeigt in der obersten Reihe, gegen eine der20 kanonischen Aminos<uren ausgetauscht wurde. Diese werden auf der linkenSeite spezifiziert. Die Punkte in der ersten Reihe stellen Wiederholungen der wt-Sequenz dar. c) Prozentuale H<ufigkeit der Austauschvariabilit<t (V) jeder einzel-nen Sequenzposition. Alle punktf.rmigen Signale auf dem in (b) gezeigten Arraywurden quantitativ vermessen, und erfolgreiche Austausche (z<hlbare Bindungs-ereignisse) wurden bestimmt (siehe Hintergrundinformationen, S4 und S5). Vwurde berechnet (V=Anzahl der Bindungsereignisse/20 O 100[14]) und gegen dieGCN4-Leucinzippersequenz aufgetragen.
AngewandteChemie
1683Angew. Chem. 2007, 119, 1682 –1686 � 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.angewandte.de

Kombinationen aus F/F-, x/F-, F/x- und x/x-Substi-tutionen erm�glichen. Die Ergebnisse sind als farbigeSegmente in Abbildung 2c dargestellt. Im Falle der a/e-Doppelaustausche sind alle Kombinationen zul�s-sig, mit Ausnahme der zweiten Heptade, die nur F/F-und F/x-Substitutionen erlaubt. Im Falle der a/d- undd/g-Substitutionen wurden nur sehr wenige hydro-phile Kombinationen (x/x) gefunden, die zudem aufdie erste Heptade beschr�nkt blieben. Nberraschen-derweise war zu beobachten, dass F/x-Substitutionenin den Positionen a/d erlaubt waren. Mit Ausnahmeder vierten Heptade war es 4berall m�glich, die hy-drophobe Aminos�ure an Position d durch eine hy-drophile zu ersetzen, wenn gleichzeitig in Position aeine hydrophobe Aminos�ure eingef4hrt wurde. Inden F�llen der d/g-Doppelsubstitutionen konnte keinvergleichbarer Effekt beobachtet werden: Mit Aus-nahme der ersten Heptade wurde gefunden, dassPosition d nur durch eine hydrophobe Aminos�ureersetzt werden kann, und zwar unabh�ngig von denSubstitutionsverh�ltnissen an Position g. Interessan-terweise scheint die vierte Heptade – besonders beiden a/d- und d/g-Paarungen – entscheidenden Ein-fluss auf das Assoziationsverhalten zu haben.
Insgesamt beobachteten wir drei Substitutionsef-fekte: Erstens wurden etliche Kombinationen vonvorher tolerierten Einzelmutationen (Abbildung 1b)gleichfalls in den doppelt substituierten Variantenakzeptiert (Additionseffekt). Beispielsweise enth�ltVariante 1 an Position aIV ein basisches Lys anstelledes urspr4nglichen Val und gleichzeitig an Position eIV
ein saures Glu anstelle eines Lys (Tabelle 1). Zweitenswurden einige der vorher nicht tolerierten Substitu-tionen akzeptiert, wenn sie gleichzeitig mit einerzuvor tolerierten Substitution kombiniert wurden(Nbergangseffekt). So wurde in Variante 2 ein hy-drophiles Thr an Position dIII anstatt des normaler-weise sehr empfindlichen Leu akzeptiert, wenn sichgleichzeitig ein Tyr in r�umlicher N�he an Position aIII
befand. Drittens wurde beobachtet, dass einige vorhernicht tolerierte Einzelsubstitutionen toleriert wurden,wenn sie gleichzeitig in Kombination in der GCN4-Sequenz auftraten (Umschwungeffekt). Zum Beispielenth�lt die Variante 3 Ile an Position aIII und Asn anPosition dIII, obwohl keine dieser beiden Substitutio-nen toleriert wurde, wenn sie alleine auftrat (Abbil-dung 1b).
Die beschriebenen Peptidarray-Experimenteeignen sich insbesondere zur Detektion heterospezi-fischer Assoziationen, weshalb wir f4r die Untersu-chung der homospezifischen Interaktionen derGCN4-Varianten einen anderen Ansatz w�hlten.Hierf4r wurden die synthetischen GCN4-Varianten1–3 jeweils als Triplikat auf Zellulose-Membranenimmobilisiert (siehe Hintergrundinformationen,S3).[17] Je ein Array wurde anschließend mit einer derVarianten auf Assoziation getestet, wobei sich einMuster von homo- und heteromeren Assoziationenergab (Abbildung 3). Die Variante 1 zeigte keine
Abbildung 2. a) Ein synthetischer Peptidarray, der alle GCN4-Leucinzipperse-quenzen enth<lt, die durch Doppelsubstitutionen von zwei Positionen durch allekanonischen Aminos<uren (außer Cys) in den Heptadenpositionen a/d, a/e undd/g der Heptaden I–IV erzeugt wurden. Aus praktischen Gr8nden wies jede Zu-sammenstellung 26 O 14 Syntheseorte auf, was die Einf8hrung von zus<tzlichenKontrollen erm.glichte. Jeder farbige Punkt repr<sentiert eine Variante, die hete-rospezifisch mit dem N-terminal mit Tamra markierten Wildtyp-Peptid assoziiertist (siehe Hintergrundinformationen, S2). b) Doppelsubstitutionen der Positio-nen aIV/eIV (V23/K27). Die Arrays wurden mithilfe eines Lumi-Imagers analysiert,und die Signalintensit<ten wurden in „Boehringer light units“ (BLUs) 8bersetzt,tabelliert und f8r eine bessere Auswertbarkeit in Graustufen 8berf8hrt (sieheHintergrundinformationen, S4 und S5). Einzelne Reihen spiegeln die Position aIV
wider, Spalten die Position eIV. Beide wurden mit den 19 gezeigten Aminos<urengetestet. Graue und schwarze Zellen zeigen assoziierende bzw. stark assoziie-rende Varianten. Alle Tabellen finden sich in den Hintergrundinformationen, S5.c) Kreisdiagramme der a/d-, a/e- und d/g-Substitutionen und ein Rberblick derin (a) zu beobachtenden Heteroassoziationen. Jedes Diagramm zeigt einenDoppelsubstitutionstypus f8r alle vier Heptaden, dargestellt als Viertel I–IV. DerRadius eines Viertels korreliert mit der Anzahl m.glicher Substitutionen in derentsprechenden Heptade. Die Oberfl<che jeder Scheibe ist als Funktion derAnzahl von Substitutionen skaliert, die zus<tzlich als hydrophob/hydrophob (F/F ; dunkelblau), hydrophil/hydrophob (x/F ; hellblau), hydrophob/hydrophil (F/x ; orange) oder hydrophil/hydrophil (x/x ; rot) gruppiert wurden.
Zuschriften
1684 www.angewandte.de � 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Angew. Chem. 2007, 119, 1682 –1686

Tendenz zur homospezifischen Assoziation, wohingegen 2und 3 deutlich homoassoziierten. Weiterhin wurden hetero-spezifische Assoziationen f4r die Varianten 2 und 3 beob-achtet, w�hrend 1 nur mit 3 heterospezifisch assoziierte.Durch Circulardichroismus-Spektroskopie wurde der Coiled-Coil-Charakter dieser Assoziationen best�tigt, der sich ineiner Steigerung der Helizit�t nach Mischung zweier assozi-ierender Varianten ausdr4ckte (siehe Hintergrundinforma-tionen, S6).
F4r die Charakterisierung der homospezifischen Asso-ziationsprozesse wurde analytische Ultrazentrifugation ein-gesetzt. Sowohl 2 (Abbildung 4a) als auch 3 ergaben stabilehomotrimere Strukturen (als 23 und 33 bezeichnet), w�hrend 1haupts�chlich monomer vorlag (siehe Hintergrundinforma-tionen, S7). Isotherme Titrationskalorimetrie gab einen tie-feren Einblick in die heterospezifischen Assoziationsprozes-se. Nach Mischen reorganisierten sich 23 und 33 zu einemheteromeren Coiled-Coil mit einer 1:1-St�chiometrie (Ab-bildung 4b und Hintergrundinformationen, S7). Das ur-spr4ngliche Monomer 1 bildete mit 33 ebenfalls einen 1:1-Komplex; hingegen wurde keine heterospezifische Interakti-on zwischen 1 und 23 beobachtet. Weitere Ultrazentrifugati-onsexperimente ergaben, dass alle gebildeten heteromerenKomplexe als Dimere vorlagen, und best�tigten zudem dieAbwesenheit einer Heteroassoziation zwischen den Varian-ten 1 und 2 (siehe Hintergrundinformationen, S7). Abbil-dung 4c fasst alle Interaktionen und St�chiometrien derdurch die drei Varianten gebildeten Coiled-Coil-Strukturenzusammen.
Die vorgestellten synthetischen Peptidarrays eignen sichzur Untersuchung von Coiled-Coil-Assoziationen, lassenjedoch keine Aussagen 4ber St�chiometrie und Topologie derCoiled-Coil-Strukturen zu. Unsere Experimente f4hren zueiner Neubewertung der Heptadenposition g. Das Verhaltenvon g gegen4ber einem Austausch zeigt eine deutlich h�hereEmpfindlichkeit als das der formal gleichwertigen Position e.Die simultanen Doppelsubstitutionen k�nnen einen Addi-
tions-, Nbergangs- oder Umschwungeffekt bewirken, was unserm�glichte, ein aus drei Sequenzvarianten des GCN4-Leu-cinzippers bestehendes Interaktionsnetzwerk vorzuschlagen.Dieses setzt sich aus einem helicalen Monomer sowie zweihomotrimeren und zwei heterodimeren Coiled-Coil-Struktu-ren zusammen. Die Positionen aIII und dIII fungieren alsSchalter f4r den Nbergang zwischen dem nativ homodimerenCoiled-Coil und homotrimeren Strukturen. Demnach kanndie native Funktion des GCN4-Leucinzippers, also die Bil-dung eines homodimeren Proteins, durch lediglich zwei Sub-stitutionen fundamental ver�ndert werden. Es ist denkbar,dass es diese supramolekulare Flexibilit�t ist, die das Coiled-
Abbildung 3. Arrayanalyse der homo- und heterospezifischen Assozia-tionen der GCN4-Leucinzippervarianten 1–3 (Tabelle 1). Auf einer Zel-lulosemembran wurden mittels Immobilisierung der synthetischenPeptide wt und 1–3 drei identische Arrays a)–c) hergestellt (siehe Hin-tergrundinformationen, S1 und S3). Jedes Array enthielt die Varianten1–3 in dreifacher Ausfertigung und als Kontrolle den nativen GCN4-Leucinzipper wt. Die Arrays wurden mit je einer der biotinylierten Vari-anten 1–3 auf Interaktion getestet (rechts gezeigt), wobei die Assozia-tionsereignisse 8ber Inkubation mit peroxidasemarkiertem Streptavidinvisualisiert wurden (siehe Hintergrundinformationen, S2 und S4).
Abbildung 4. a) Untersuchung zur Homoassoziation von Variante 2durch analytische Ultrazentrifugation. Einzelne Peptidproben mit Kon-zentrationen von 3 mgmL�1 (schwarze Kreise), 1.5 mgmL�1 (roteKreise) und 0.3 mgmL�1 (blaue Kreise) ergaben die lokalen Unter-schiede im Brechungsindex Dn als Funktion des quadrierten Abstandsvom Rotormittelpunkt (d 2) im chemischen und Sedimentationsgleich-gewicht bei 50 krpm. Der Rbersichtlichkeit halber ist lediglich jederzehnte Datenpunkt gezeigt. Die besten Kurvenanpassungen (durchge-zogene Linien) wurden f8r ein nichtideales Monomer-Dimer-Trimer-Modell erhalten (siehe Hintergrundinformationen, S7). b) Heteroasso-ziation der Varianten 2 und 3, untersucht durch isotherme Titrations-kalorimetrie. Proben (10 mL) einer 1.08 mm L.sung von 3 wurden bei25 8C in eine 124 mm L.sung von 2 injiziert. Die beste Kurvenanpas-sung (blaue Linie) an die normierten Reaktionsw<rmen Q (rote Kreise)ergab eine Assoziationskonstante von 1.2 O 105
m�1 bei einer St.chio-
metrie von 1:1. R ist das molare Verh<ltnis von 3 zu 2. c) Netzwerk derAssoziationen zwischen den freien Varianten 1–3, das sich aus denbiophysikalischen Experimenten a und b ergibt (siehe Hintergrundin-formationen, S7).
AngewandteChemie
1685Angew. Chem. 2007, 119, 1682 –1686 � 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.angewandte.de

Coil-Motiv zu einem so vielgestaltigen Akteur in der Naturmacht.
Eingegangen am 9. August 2006Online ver�ffentlicht am 9. Januar 2007
.Stichw�rter: Helicale Strukturen · Peptidarrays · Peptide ·Proteinstrukturen · Spot-Synthese
[1] a) J. M. Mason, K. M. Arndt, ChemBioChem 2004, 5, 170;b) A. N. Lupas, M. Gruber, Adv. Protein Chem. 2005, 70, 37.
[2] F. H. S. Crick, Acta Crystallogr. 1953, 6, 689.[3] a) P. B. Harbury, T. Zhang, P. S. Kim, T. Alber, Science 1993, 262,
1401; b) P. B. Harbury, P. S. Kim, T. Alber, Nature 1994, 371, 80;c) D. L. Akey, V. N. Malashkevich, P. S. Kim, Biochemistry 2001,40, 6352; d) B. Tripet, K. Wagschal, P. Lavigne, C. T. Mant, R. S.Hodges, J. Mol. Biol. 2000, 300, 377.
[4] a) N. E. Zhou, C. M. Kay, R. S. Hodges, J. Mol. Biol. 1994, 237,500; b) D. Krylov, I. Mikhailenko, C. Vinson, EMBO J. 1994, 13,2849; c) W. D. Kohn, C. M. Kay R. S. Hodges, J. Mol. Biol. 1998,283, 993; d) R. A. Kammerer, D. Kostrewa, P. Progias, S. Hon-nappa, D. Avila, A. Lustig, F. K. Winkler, J. Pieters, M. O.Steinmetz, Proc. Natl. Acad. Sci. USA 2005, 102, 13891.
[5] E. K. OPShea, K. J. Lumb, P. S. Kim, Curr. Biol. 1993, 3, 658.[6] a) MULTICOIL: E. Wolf, P. S. Kim, B. Berger, Protein Sci. 1997,
6, 1179; b) SOCKET: J. Walshaw, D. N. Woolfson, J. Mol. Biol.2001, 307, 1427; c) COILS: A. Lupas, M. van Dyke, J. Stock,
Science 1991, 252, 1162; d) PAIRCOIL: B. Berger, D. B. Wilson,E. Wolf, T. Tonchev, M. Milla, P. S. Kim, Proc. Natl. Acad. Sci.USA 1995, 92, 8259; e) J. M. Mason, M. A. Schmitz, K. M.M4ller, K. M. Arndt, Proc. Natl. Acad. Sci. USA 2006, 103, 8989.
[7] J. C. Hu, E. K. OPShea, P. S. Kim, R. T. Sauer, Science 1990, 250,1400.
[8] J. N. Pelletier, K. M. Arndt, A. Pl4ckthun, S. W. Michnick, Nat.Biotechnol. 1999, 17, 683.
[9] J. R. Newman, A. E. Keating, Science 2003, 300, 2097.[10] R. Frank, J. Immunol. Methods 2002, 267, 13.[11] E. K. OPShea, J. D. Klemm, P. S. Kim, T. Albert, Science 1991,
254, 539.[12] Details zur Synthese und Reinheit von Spot-synthetisierten, an
Zellulosemembranen gebundenen Peptidarrays werden in denHintergrundinformationen, S1, n�her erl�utert.
[13] A. A. Weiser, M. Or-Guil, V. Tapia, A. Leichsenring, J. Schuch-hardt, C. Fr�mmel, R. Volkmer-Engert, Anal. Biochem. 2005,342, 300.
[14] J. Przdzezdiak, S. Tremmel, I. Kretzschmar, M. Beyermann, M.Bienert, R. Volkmer-Engert, ChemBioChem 2006, 7, 780.
[15] a) B. Ibarra-Molero, J. A. Zitzewitz, C. R. Matthews, J. Mol.Biol. 2004, 336, 989; b) C. Vinson, A. Acharya, E. J. Taparowsky,Biochim. Biophys. Acta 2006, 1759, 4.
[16] X. Zeng, H. Zhu, H. A. Lashuel, J. C. Hu, Protein Sci. 1997, 6,2218.
[17] H. K. Rau, N. DeJonge, W. Haehnel, Angew. Chem. 2000, 112,256; Angew. Chem. Int. Ed. 2000, 39, 250.
Zuschriften
1686 www.angewandte.de � 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Angew. Chem. 2007, 119, 1682 –1686

1
A Network of Coiled-Coil Associations Derived from Synthetic GCN4 Leucine Zipper
Arrays
Michael Portwich, Sandro Keller, Holger M. Strauss, Carsten C. Mahrenholz, Ines
Kretzschmar, Achim Kramer, and Rudolf Volkmer
S1: Solid-phase peptide synthesis of GCN4 leucine zipper variants
Soluble GCN4 leucine zipper variants for dye labeling, biotin labeling, immobilization, and
biophysical experiments were synthesized by solid-phase peptide synthesis according to the
Fmoc strategy. Variants wt, 1, 2, and 3 were prepared with either an N-terminal (Cββ-) or a
C-terminal (-ββC) elongation (β = β-alanine). Peptides without elongation were used for
biophysical experiments. The following sequences were used for:
dye labeling: CββRMKQLEDKVEELLSKNYHLENEVARLKKLVG (Cββ-wt)
biotin labeling: CββRMKQLEDKVEELLSKNYHLENEKARLEKLVG (Cββ-1)
CββRMKQLEDKVEELLSKYYHTENEVARLKKLVG (Cββ-2)
CββRMKQLEDKVEELLSKIYHNENEVARLKKLVG (Cββ-3)
immobilization: RMKQLEDKVEELLSKNYHLENEVARLKKLVGββC (wt-ββC)
RMKQLEDKVEELLSKNYHLENEKARLEKLVGββC (1-ββC)
RMKQLEDKVEELLSKYYHTENEVARLKKLVGββC (2-ββC)
RMKQLEDKVEELLSKIYHNENEVARLKKLVGββC (3-ββC)
Automatic solid-phase peptide synthesis was performed on Tentagel-SRam resin (Rapp
Polymere, Tübingen, Germany). Reactions were performed in plastic syringes at room
temperature on a multi-peptide synthesis robot (Syro2000, MultiSynTech, Witten, Germany).
Couplings were achieved by reacting 4 equivalents Fmoc-AA-OH with 4 equivalents PyBOB
and 8 equivalents NMM in DMF. A solution of piperidine (20%) in DMF was used to remove
the Fmoc protecting group. Peptides were deprotected and cleaved from the resin using a
mixture of 10 mL TFA, 0.75 g phenol, 0.5 mL water, 0.5 mL methylphenylsulfide, and
0.25 mL 1,2-ethandithiol. After 3 h at room temperature, the cleavage solution was collected,
and the crude peptides were precipitated from the solution with dry ether at 0 °C. HPLC
purification and analysis were achieved using a linear solvent gradient (A: 0.05% TFA in
water; B: 0.05% TFA in acetonitrile; gradient: 5–60% B over 30 min; UV detector at 214 nm;

2
RP-18 column). The identities of the peptides were validated by mass spectrometry using
MALDI-TOF (VoyagerLT, Applied Biosystems, Weiterstadt, Germany) and ESI (Q-
TOFmicro, Micromass, Manchester, UK).
SPOT synthesis: Synthesis of cellulose-membrane-bound arrays was done according to: H.
Wenschuh, H. Gausepohl, L. Germeroth, M. Ulbricht, H. Matuschewski, A. Kramer, R.
Volkmer-Engert, N. Heine, T. Ast, D. Scharn, J. Schneider-Mergener in Combinatorial
Chemistry, A Practical Approach. (Ed.: H. Fenniri), Oxford University Press, Oxford, 2000,
pp. 95–116. The purity of SPOT-synthesized peptides is generally in the range of 40–85%,
which is adequate for screening assays. Even longer peptides can be SPOT-synthesized in
sufficient quality, as recently shown by Przezdziak et al.[1] The SPOT-synthesized FBP28
WW domain, containing 34 residues, was obtained in ~65% purity as determined by HPLC,
and the identity was confirmed by mass spectrometry.[1]
S2: Dye and biotin labeling
Cββ-wt was dye-labeled by Michael addition of its N-terminal Cys thiol group to
tetramethylrhodamine-6-maleimide (Molecular Probes, Eugene, USA). The peptide was
dissolved in sodium phosphate buffer (10 mM, pH 7.4, argon-sparged) at a concentration of
6.8 mM. Then, 1.5 equivalents maleimide dye compound and 1.5 equivalents tris(2-
carboxyethyl)phosphine hydrochloride were added, and the reaction mixture was stirred for
4 h at room temperature. Afterwards, 10 equivalents 2-mercaptoethanol was added to cap
unmodified maleimido groups (1 h) before adding 100 equivalents dimethylsulfoxide (1 h).
The dye-labeled peptide was purified by HPLC as described in S1; detection was carried out
at 540 and 214 nm. The identity of the peptide was confirmed by mass spectrometry (see S1).
According to the same protocol, variants Cββ-1, Cββ-2, and Cββ-3 were N-terminally
labeled with biotin using the Nα-(3-maleimidylpropionyl)biocytin reagent (Molecular Probes,
Eugene, USA), purified, and confirmed.
S3: Immobilization of wt-ββC, 1-ββC, 2-ββC, and 3-ββC on cellulose membranes
The purified and analytically characterized peptides (see S1) were immobilized via their C-
terminal Cys to a maleimide-functionalized Whatman 50 cellulose support as described by
Otte et al.[2]
S4: Binding assay on cellulose membranes

3
General: Membrane-bound GCN4 variants were washed with DMF, ethanol, and three times
with TBS buffer (13.7 mM NaCl, 0.27 mM KCl, 5 mM Tris (tris(hydroxymethyl)amino-
methane), pH 8.0) for 10 min each. Membranes were incubated for 3 h with blocking buffer
(for 50 mL blocking buffer: 5 mL blocking buffer (Sigma–Aldrich, Steinheim, Germany),
2.5 g sucrose, 5 mL 10 × TBS buffer (137 mM NaCl, 2.7 mM KCl, 50 mM Tris, pH 8.0),
filled up to 50 mL with water).
Single-substitution arrays: Arrays were incubated with a solution of N-terminally tetra-
methylrhodamine-labeled Cββ-wt at a concentration of 10 µM in blocking buffer at room
temperature overnight. Arrays were washed three times with TBS, and the resulting spot
patterns were recorded with a flatbed scanner (Snapscan e40, Agfa, Mortsel, Belgium).
Subsequently, arrays were analyzed densitometrically to yield spot signal intensities in terms
of Boehringer light units using the Lumi-Analyst software (Roche, Mannheim, Germany; see
also S5). Arrays were regenerated by washing with DMF (4 times, 20 min each) followed by
washing with ethanol (3 times, 3 min each) and drying. As a negative control, arrays were
probed with 2-mercaptoethanol coupled to tetramethylrhodamine-6-maleimide (prepared
according to S2).
Double-substitution arrays: Cellulose membranes were incubated with a solution of N-
terminally tetramethylrhodamine-labeled Cββ-wt at a concentration of 50 µM in blocking
buffer at room temperature overnight before proceeding as described above. Results are
shown in tabular form in S5.
Table S4.1: GCN4 heptad positions and residues selected for double substitutions.
Position Substituted residues Location a/dI Met2/Leu5 core/core aII/dII Val9/Leu12 core/core aIII/dIII Asn16/Leu19 core/core aIV/dIV Val23/Leu26 core/core aI/eI Met2/Glu5 core/flank aII/eII Val9/Leu13 core/flank aIII/eIII Asn16/Glu20 core/flank aIV/eIV Val23/Lys27 core/flank dI/gI Leu5/Lys8 core/flank dII/gII Leu12/Lys15 core/flank dIII/gIII Leu19/Glu22 core/flank dIV/gIV Leu26/Leu29 core/flank
Mutual associations between variants wt, 1, 2, and 3: Three identical arrays were incubated
simultaneously, each with one of the biotin-labeled variants (Cββ-1, Cββ-2, and Cββ-3) at
10 µM in blocking buffer at room temperature overnight. After washing 3 times with TBS,
4
arrays were incubated for 30 min with horse-radish-peroxidase-labeled streptavidin (Sigma,
St. Louis, USA) according to the supplier’s instructions and subsequently washed five times
with TBS. Binding was visualized using a chemoluminescent substrate (Uptilight HRP,
Uptima, Interchim, Montluçon, France) and a Lumi-Imager (Roche, Mannheim, Germany).
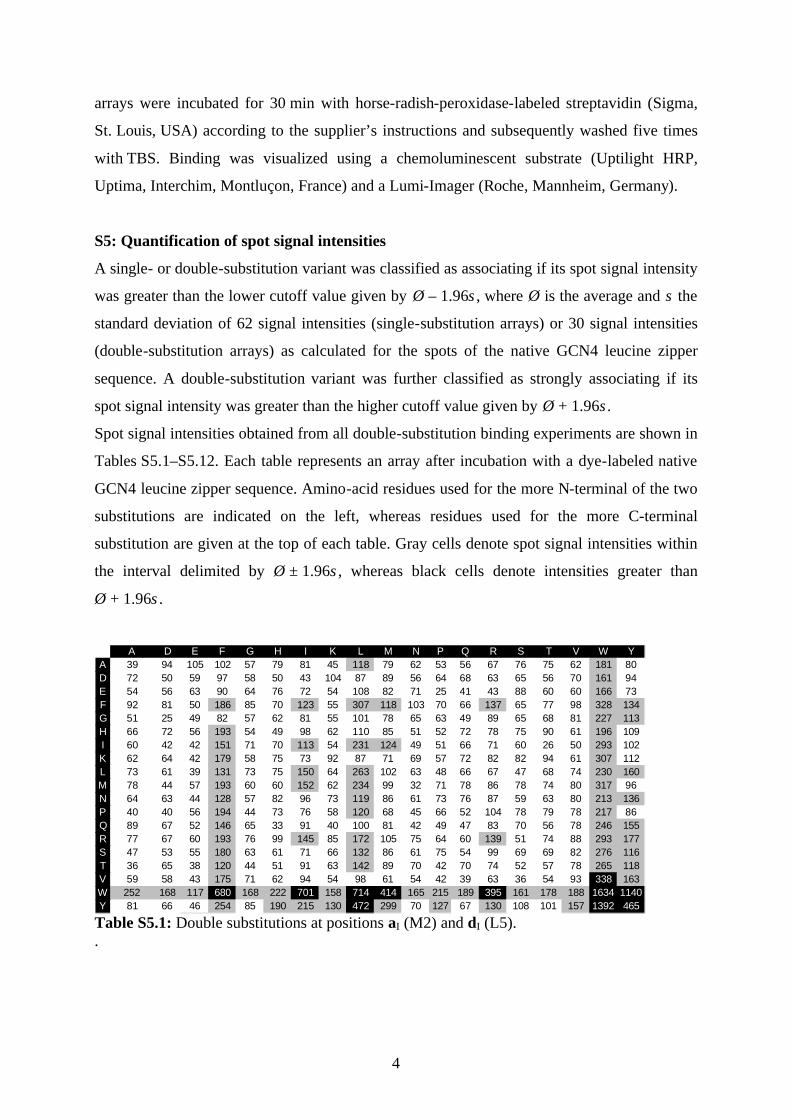
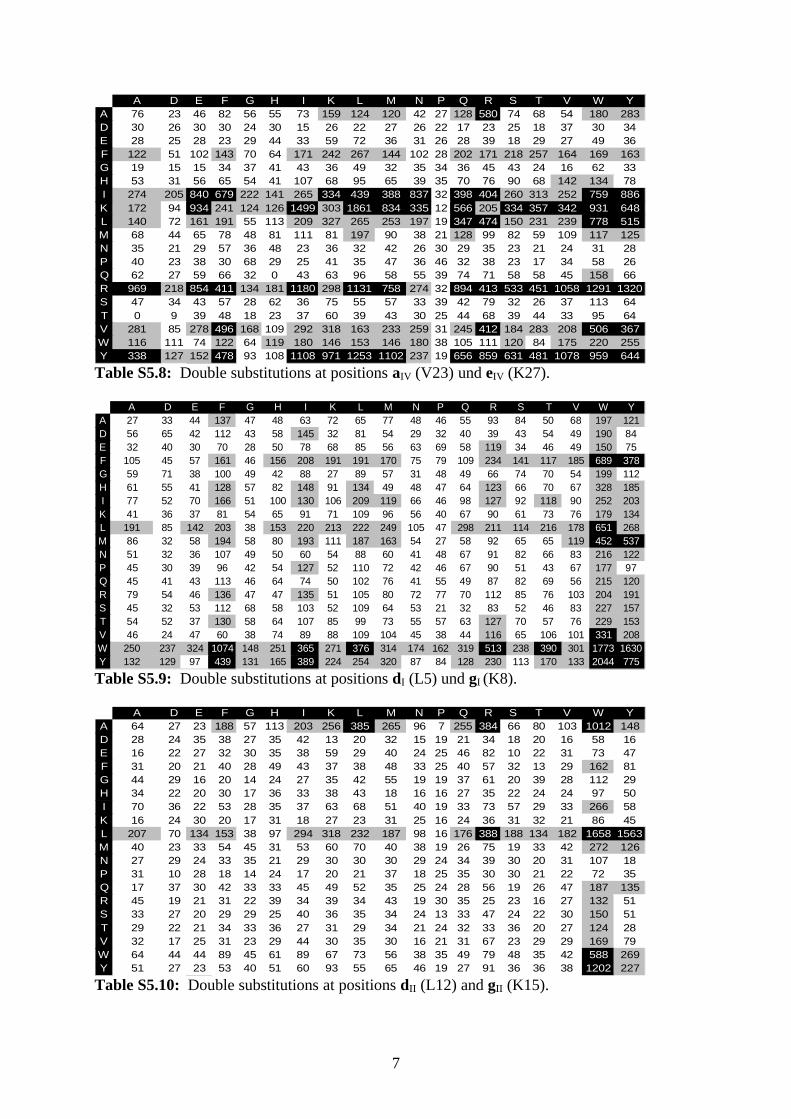
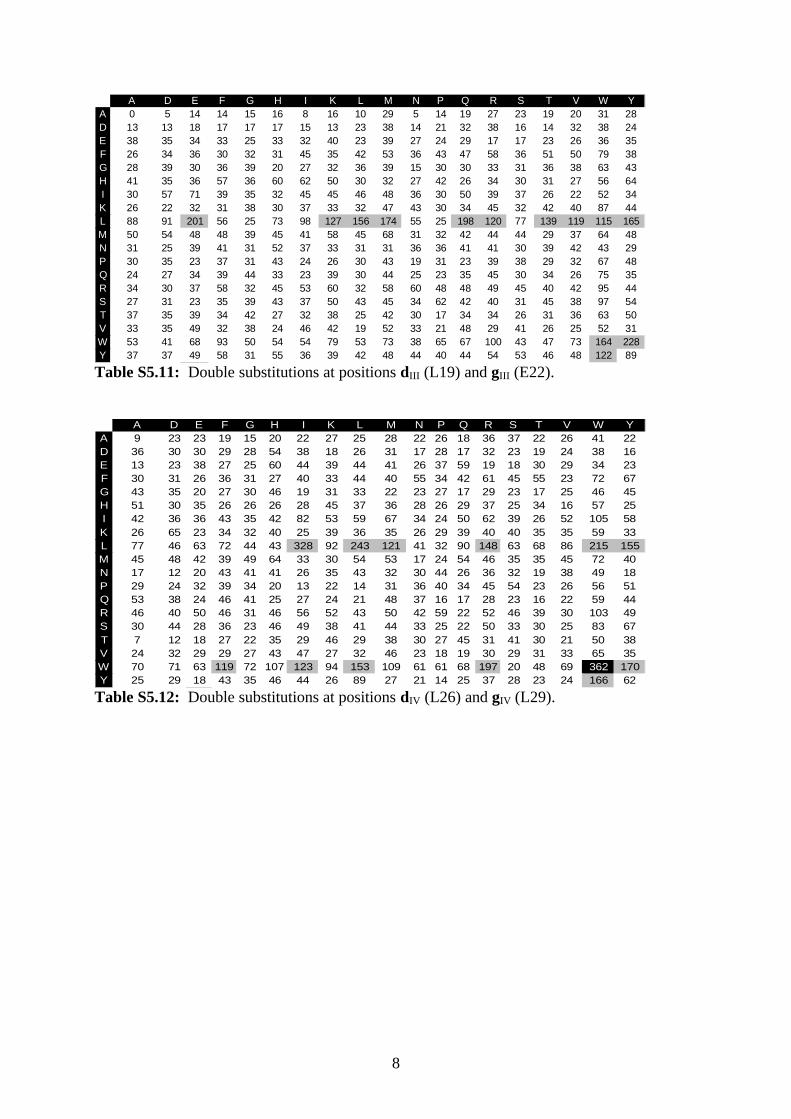
S5: Quantification of spot signal intensities
A single- or double-substitution variant was classified as associating if its spot signal intensity
was greater than the lower cutoff value given by Ø – 1.96s , where Ø is the average and s the
standard deviation of 62 signal intensities (single-substitution arrays) or 30 signal intensities
(double-substitution arrays) as calculated for the spots of the native GCN4 leucine zipper
sequence. A double-substitution variant was further classified as strongly associating if its
spot signal intensity was greater than the higher cutoff value given by Ø + 1.96s .
Spot signal intensities obtained from all double-substitution binding experiments are shown in
Tables S5.1–S5.12. Each table represents an array after incubation with a dye-labeled native
GCN4 leucine zipper sequence. Amino-acid residues used for the more N-terminal of the two
substitutions are indicated on the left, whereas residues used for the more C-terminal
substitution are given at the top of each table. Gray cells denote spot signal intensities within
the interval delimited by Ø ± 1.96s , whereas black cells denote intensities greater than
Ø + 1.96s .
A D E F G H I K L M N P Q R S T V W Y
A 39 94 105 102 57 79 81 45 118 79 62 53 56 67 76 75 62 181 80D 72 50 59 97 58 50 43 104 87 89 56 64 68 63 65 56 70 161 94E 54 56 63 90 64 76 72 54 108 82 71 25 41 43 88 60 60 166 73F 92 81 50 186 85 70 123 55 307 118 103 70 66 137 65 77 98 328 134G 51 25 49 82 57 62 81 55 101 78 65 63 49 89 65 68 81 227 113H 66 72 56 193 54 49 98 62 110 85 51 52 72 78 75 90 61 196 109I 60 42 42 151 71 70 113 54 231 124 49 51 66 71 60 26 50 293 102K 62 64 42 179 58 75 73 92 87 71 69 57 72 82 82 94 61 307 112L 73 61 39 131 73 75 150 64 263 102 63 48 66 67 47 68 74 230 160M 78 44 57 193 60 60 152 62 234 99 32 71 78 86 78 74 80 317 96N 64 63 44 128 57 82 96 73 119 86 61 73 76 87 59 63 80 213 136P 40 40 56 194 44 73 76 58 120 68 45 66 52 104 78 79 78 217 86Q 89 67 52 146 65 33 91 40 100 81 42 49 47 83 70 56 78 246 155R 77 67 60 193 76 99 145 85 172 105 75 64 60 139 51 74 88 293 177S 47 53 55 180 63 61 71 66 132 86 61 75 54 99 69 69 82 276 116T 36 65 38 120 44 51 91 63 142 89 70 42 70 74 52 57 78 265 118V 59 58 43 175 71 62 94 54 98 61 54 42 39 63 36 54 93 338 163W 252 168 117 680 168 222 701 158 714 414 165 215 189 395 161 178 188 1634 1140Y 81 66 46 254 85 190 215 130 472 299 70 127 67 130 108 101 157 1392 465
Table S5.1: Double substitutions at positions aI (M2) and dI (L5). .

5
A D E F G H I K L M N P Q R S T V W YA 21 30 26 35 19 18 35 25 103 48 31 37 34 42 43 41 40 73 27D 26 25 18 34 23 29 21 48 28 36 38 28 16 26 32 35 30 49 51E 30 28 40 43 45 29 26 40 53 30 24 36 29 13 29 24 38 28 33F 103 22 322 40 46 36 74 39 237 59 46 44 99 36 40 31 63 60 93G 36 3 23 29 22 38 37 37 26 40 22 42 42 40 23 15 40 59 35H 41 34 40 50 21 48 36 36 35 40 50 42 43 43 27 47 41 51 42I 288 25 74 53 54 42 80 31 403 55 43 24 69 55 48 8 58 56 71K 30 31 35 27 21 27 42 31 76 36 19 43 43 53 29 49 20 66 46L 86 48 76 29 61 24 56 24 177 41 26 22 44 30 47 31 61 60 74M 68 67 50 45 41 47 51 47 107 29 29 58 101 44 29 23 28 70 41N 31 38 36 38 23 24 35 39 29 44 23 30 37 50 47 41 32 31 43P 30 27 22 31 29 35 19 29 26 51 33 27 27 37 42 36 36 54 48Q 26 48 32 52 26 11 59 31 39 30 31 27 38 36 44 23 35 62 42R 44 38 45 47 43 45 35 37 81 29 47 35 0 61 53 32 40 142 39S 19 36 24 23 35 38 33 36 39 36 49 41 30 30 28 43 31 75 38T 5 52 42 21 38 37 39 34 37 42 18 35 38 43 33 19 37 63 41V 198 24 43 33 56 38 55 16 158 67 35 21 33 22 33 28 29 45 64W 121 28 262 44 54 70 80 66 123 94 61 41 119 84 29 38 71 155 111Y 122 19 507 53 22 36 43 29 475 43 23 14 137 43 49 43 89 114 417
Table S5.2: Double substitutions at positions aII (V9) and dII (L12).
A D E F G H I K L M N P Q R S T V W YA 33 32 38 61 23 26 19 6 76 36 29 10 29 67 26 31 17 33 48D 19 18 19 16 17 21 1 32 30 31 35 30 21 26 21 18 28 37 25E 30 35 25 23 27 28 28 39 49 34 31 29 39 14 69 47 34 39 29F 35 30 32 40 34 46 49 39 133 56 40 37 49 43 42 141 55 72 49G 25 4 27 21 41 23 39 24 73 41 32 32 31 35 30 30 29 41 38H 37 34 35 45 36 30 120 19 857 69 26 46 27 47 41 50 41 49 41I 52 28 22 60 45 236 40 65 41 65 186 31 72 37 38 37 57 62 60K 32 26 18 51 26 39 31 27 173 43 30 36 21 49 38 27 31 62 38L 20 26 30 23 25 34 38 33 48 29 32 24 44 32 22 40 32 43 36M 45 35 22 41 31 57 58 34 395 73 42 34 51 52 43 78 47 68 44N 27 44 15 48 26 38 59 24 223 48 43 31 18 30 29 21 36 65 37P 41 37 20 32 20 34 20 25 35 24 30 34 23 31 27 19 22 54 27Q 35 23 23 43 29 23 97 44 342 93 29 18 42 52 27 41 109 52 46R 27 28 26 72 38 66 49 22 405 50 41 22 26 37 48 20 43 59 48S 20 17 18 52 35 39 126 28 328 69 36 26 25 57 23 41 74 62 51T 30 30 42 69 41 44 275 28 736 215 73 29 29 46 50 95 156 68 28V 40 12 23 23 36 47 28 11 34 55 52 44 42 42 40 49 15 42 35W 34 44 47 57 79 59 83 46 180 83 47 60 69 49 26 106 76 166 107Y 44 40 30 90 21 36 251 45 1363 316 68 25 45 47 39 237 124 100 101
Table S5.3: Double substitutions at positions aIII (N16) and dIII (L19).
A D E F G H I K L M N P Q R S T V W YA 54 36 24 33 26 33 39 18 100 27 23 38 20 34 35 14 17 35 43D 24 23 23 33 24 33 6 40 56 36 19 24 18 29 26 17 20 61 31E 33 20 41 27 23 26 27 20 36 35 25 22 26 24 24 45 31 36 30F 32 35 33 32 38 23 27 38 123 32 27 19 23 19 22 13 26 73 33G 31 2 39 31 23 38 30 29 30 28 14 28 22 27 36 17 17 39 34H 34 12 22 24 37 36 25 24 39 37 43 29 23 35 31 23 34 73 37I 32 23 31 41 29 27 110 28 227 40 27 18 21 25 18 4 30 62 63K 29 16 23 28 30 35 50 39 173 47 38 38 19 33 42 30 41 84 60L 29 27 30 0 45 62 54 27 157 42 35 43 40 45 44 37 45 110 65M 37 32 41 45 39 41 27 36 91 34 0 91 62 45 44 23 25 54 63N 28 42 39 28 21 42 36 20 25 39 22 21 32 44 22 43 26 21 47P 58 31 16 30 27 32 27 30 32 44 36 34 36 39 26 24 26 72 35Q 39 28 22 50 33 0 41 58 60 31 33 22 39 49 44 28 42 76 48R 47 32 23 45 28 41 52 41 178 44 25 39 0 61 69 33 38 92 49S 36 29 29 47 42 56 44 33 45 45 32 31 30 37 28 37 29 80 39T 0 35 66 48 50 32 40 25 39 39 39 38 34 41 34 23 32 79 36V 24 24 32 63 38 47 69 1 155 87 60 43 32 49 26 36 47 93 85W 33 37 37 64 43 46 37 66 138 61 49 61 60 95 18 54 76 123 90Y 18 22 20 41 35 46 42 34 829 33 30 32 18 28 16 21 32 243 252
Table S5.4: Double substitutions at positions aIV (V23) and dIV (L26).

6
A D E F G H I K L M N P Q R S T V W YA 186 56 112 214 118 134 204 136 241 204 103 115 142 193 186 110 164 1546 358D 107 53 120 211 108 103 117 90 120 100 101 87 66 168 124 143 123 485 189E 93 56 98 277 102 122 150 195 129 157 97 88 94 112 127 82 131 503 223F 329 161 251 498 303 307 441 368 374 1063 303 206 322 477 861 487 698 1034 1168G 191 63 87 260 152 160 216 145 253 224 94 131 146 154 144 182 265 679 426H 224 120 202 807 198 220 440 167 238 218 181 125 240 174 222 200 227 1019 516I 216 69 270 400 288 305 363 398 347 375 213 172 205 398 428 193 286 876 841K 158 97 119 508 142 166 236 145 213 204 158 157 161 321 226 194 308 874 573L 283 107 278 378 246 331 430 286 294 342 188 159 298 320 388 358 320 1632 1211M 296 97 270 530 289 307 443 293 530 412 169 99 247 377 177 213 290 934 1180N 173 105 123 328 142 166 238 212 229 187 186 146 134 230 240 132 170 501 220P 134 65 140 233 176 140 182 197 171 212 98 154 108 238 213 197 200 1214 353Q 136 105 140 344 162 127 264 119 197 174 110 167 97 205 239 166 212 888 316R 303 160 222 594 260 265 461 142 434 350 153 237 161 222 221 216 317 908 494S 167 80 174 411 194 160 272 168 312 280 135 150 210 230 220 214 167 788 366T 170 60 138 304 123 161 237 139 304 284 140 172 149 201 169 164 261 827 419V 324 96 179 457 162 203 209 206 281 251 112 134 132 267 230 220 202 1059 522W 920 1384 1350 1688 1554 1670 1497 1123 1296 1713 1501 546 1791 667 1515 1351 1269 1846 2092Y 371 190 842 850 455 735 1048 769 502 1358 475 313 924 1194 1232 1242 1229 2121 1800
Table S5.5: Double substitutions at positions aI (M2) und eI (E5).
A D E F G H I K L M N P Q R S T V W YA 50 22 50 41 31 33 60 43 65 86 27 15 41 96 47 44 60 227 146D 34 21 29 26 22 40 5 27 30 21 20 28 17 19 13 15 8 36 23E 19 30 21 32 29 22 37 30 31 47 14 24 38 37 14 34 29 64 22F 79 38 57 179 27 51 195 118 258 204 58 33 121 120 217 69 134 485 295G 26 23 18 30 33 28 19 17 21 17 25 22 18 32 14 19 14 86 39H 27 16 28 56 19 33 63 48 69 34 21 22 30 61 21 33 46 120 35I 252 255 378 394 93 263 389 279 263 484 283 17 237 359 243 149 218 566 682K 37 26 55 57 35 40 61 37 70 42 37 29 34 72 31 28 26 112 67L 195 70 109 255 38 59 229 143 231 239 75 32 106 213 83 86 195 525 630M 85 65 92 133 48 58 155 84 105 82 32 20 59 82 54 38 70 234 188N 41 27 23 37 29 20 25 47 31 41 23 22 42 32 27 35 47 49 25P 28 33 22 28 28 36 28 42 27 51 30 33 28 39 28 26 26 66 23Q 32 40 17 34 22 19 23 31 34 44 23 25 26 37 34 40 33 121 49R 39 32 45 62 26 45 59 61 68 74 34 32 51 85 35 38 67 176 98S 40 31 30 44 34 43 36 41 52 35 44 25 36 42 68 38 30 102 57T 28 15 19 32 30 39 52 34 48 46 17 32 30 36 35 36 42 170 42V 334 144 123 312 94 71 273 164 188 155 70 12 155 230 111 79 184 1334 606W 134 53 68 164 72 132 258 298 243 140 105 37 109 246 136 88 112 1635 454Y 124 9 43 439 36 86 288 290 755 349 74 22 204 378 215 82 222 1372 629
Table S5.6: Double substitutions at positions aII (V9) und eII (L13).
A D E F G H I K L M N P Q R S T V W YA 16 40 44 26 45 29 20 35 28 47 29 27 25 27 30 24 19 78 32D 24 14 32 14 26 18 8 34 14 36 31 39 52 42 56 30 18 67 35E 36 23 42 44 27 46 45 60 45 82 47 12 42 62 38 35 40 108 64F 61 128 171 56 93 57 59 125 54 116 90 31 131 129 199 227 64 95 74G 61 19 171 56 56 65 52 73 49 120 46 34 73 78 65 57 33 104 57H 445 164 459 207 85 264 340 1256 466 719 377 23 478 1398 774 1065 944 509 315I 36 41 31 32 25 32 30 37 51 29 33 29 26 41 42 19 25 64 54K 62 47 280 71 37 60 102 97 219 199 33 37 166 75 96 152 79 122 72L 30 46 35 24 35 46 40 40 43 46 33 23 42 42 43 30 23 68 44M 177 120 600 93 116 109 160 303 307 230 102 30 341 396 173 218 192 293 149N 254 82 301 170 142 331 183 443 222 497 473 25 405 1034 293 340 271 409 312P 19 17 28 32 23 23 25 28 36 41 39 22 31 41 30 28 18 67 45Q 314 311 1210 413 480 146 434 516 264 887 191 14 740 926 774 448 257 1068 762R 168 62 419 128 106 107 275 193 466 424 94 34 305 176 134 233 216 230 175S 130 176 674 83 110 68 61 93 68 155 86 54 109 112 135 109 76 135 112T 101 254 971 84 135 70 122 98 102 306 87 39 204 126 207 152 77 297 118V 39 41 46 42 54 56 33 20 15 31 25 41 32 43 38 38 48 97 57W 170 230 230 136 184 228 139 358 150 471 383 71 271 253 326 202 169 187 222Y 1028 1185 1756 546 1563 631 191 1232 530 1372 986 47 1339 1313 1247 1545 502 1092 1180
Table S5.7: Double substitutions at positions aIII (N16) und eIII (E20).
7
A D E F G H I K L M N P Q R S T V W YA 76 23 46 82 56 55 73 159 124 120 42 27 128 580 74 68 54 180 283D 30 26 30 30 24 30 15 26 22 27 26 22 17 23 25 18 37 30 34E 28 25 28 23 29 44 33 59 72 36 31 26 28 39 18 29 27 49 36F 122 51 102 143 70 64 171 242 267 144 102 28 202 171 218 257 164 169 163G 19 15 15 34 37 41 43 36 49 32 35 34 36 45 43 24 16 62 33H 53 31 56 65 54 41 107 68 95 65 39 35 70 76 90 68 142 134 78I 274 205 840 679 222 141 265 334 439 388 837 32 398 404 260 313 252 759 886K 172 94 934 241 124 126 1499 303 1861 834 335 12 566 205 334 357 342 931 648L 140 72 161 191 55 113 209 327 265 253 197 19 347 474 150 231 239 778 515M 68 44 65 78 48 81 111 81 197 90 38 21 128 99 82 59 109 117 125N 35 21 29 57 36 48 23 36 32 42 26 30 29 35 23 21 24 31 28P 40 23 38 30 68 29 25 41 35 47 36 46 32 38 23 17 34 58 26Q 62 27 59 66 32 0 43 63 96 58 55 39 74 71 58 58 45 158 66R 969 218 854 411 134 181 1180 298 1131 758 274 32 894 413 533 451 1058 1291 1320S 47 34 43 57 28 62 36 75 55 57 33 39 42 79 32 26 37 113 64T 0 9 39 48 18 23 37 60 39 43 30 25 44 68 39 44 33 95 64V 281 85 278 496 168 109 292 318 163 233 259 31 245 412 184 283 208 506 367W 116 111 74 122 64 119 180 146 153 146 180 38 105 111 120 84 175 220 255Y 338 127 152 478 93 108 1108 971 1253 1102 237 19 656 859 631 481 1078 959 644
Table S5.8: Double substitutions at positions aIV (V23) und eIV (K27).
A D E F G H I K L M N P Q R S T V W YA 27 33 44 137 47 48 63 72 65 77 48 46 55 93 84 50 68 197 121D 56 65 42 112 43 58 145 32 81 54 29 32 40 39 43 54 49 190 84E 32 40 30 70 28 50 78 68 85 56 63 69 58 119 34 46 49 150 75F 105 45 57 161 46 156 208 191 191 170 75 79 109 234 141 117 185 689 378G 59 71 38 100 49 42 88 27 89 57 31 48 49 66 74 70 54 199 112H 61 55 41 128 57 82 148 91 134 49 48 47 64 123 66 70 67 328 185I 77 52 70 166 51 100 130 106 209 119 66 46 98 127 92 118 90 252 203K 41 36 37 81 54 65 91 71 109 96 56 40 67 90 61 73 76 179 134L 191 85 142 203 38 153 220 213 222 249 105 47 298 211 114 216 178 651 268M 86 32 58 194 58 80 193 111 187 163 54 27 58 92 65 65 119 452 537N 51 32 36 107 49 50 60 54 88 60 41 48 67 91 82 66 83 216 122P 45 30 39 96 42 54 127 52 110 72 42 46 67 90 51 43 67 177 97Q 45 41 43 113 46 64 74 50 102 76 41 55 49 87 82 69 56 215 120R 79 54 46 136 47 47 135 51 105 80 72 77 70 112 85 76 103 204 191S 45 32 53 112 68 58 103 52 109 64 53 21 32 83 52 46 83 227 157T 54 52 37 130 58 64 107 85 99 73 55 57 63 127 70 57 76 229 153V 46 24 47 60 38 74 89 88 109 104 45 38 44 116 65 106 101 331 208W 250 237 324 1074 148 251 365 271 376 314 174 162 319 513 238 390 301 1773 1630Y 132 129 97 439 131 165 389 224 254 320 87 84 128 230 113 170 133 2044 775
Table S5.9: Double substitutions at positions dI (L5) und gI (K8).
A D E F G H I K L M N P Q R S T V W YA 64 27 23 188 57 113 203 256 385 265 96 7 255 384 66 80 103 1012 148D 28 24 35 38 27 35 42 13 20 32 15 19 21 34 18 20 16 58 16E 16 22 27 32 30 35 38 59 29 40 24 25 46 82 10 22 31 73 47F 31 20 21 40 28 49 43 37 38 48 33 25 40 57 32 13 29 162 81G 44 29 16 20 14 24 27 35 42 55 19 19 37 61 20 39 28 112 29H 34 22 20 30 17 36 33 38 43 18 16 16 27 35 22 24 24 97 50I 70 36 22 53 28 35 37 63 68 51 40 19 33 73 57 29 33 266 58K 16 24 30 20 17 31 18 27 23 31 25 16 24 36 31 32 21 86 45L 207 70 134 153 38 97 294 318 232 187 98 16 176 388 188 134 182 1658 1563M 40 23 33 54 45 31 53 60 70 40 38 19 26 75 19 33 42 272 126N 27 29 24 33 35 21 29 30 30 30 29 24 34 39 30 20 31 107 18P 31 10 28 18 14 24 17 20 21 37 18 25 35 30 30 21 22 72 35Q 17 37 30 42 33 33 45 49 52 35 25 24 28 56 19 26 47 187 135R 45 19 21 31 22 39 34 39 34 43 19 30 35 25 23 16 27 132 51S 33 27 20 29 29 25 40 36 35 34 24 13 33 47 24 22 30 150 51T 29 22 21 34 33 36 27 31 29 34 21 24 32 33 36 20 27 124 28V 32 17 25 31 23 29 44 30 35 30 16 21 31 67 23 29 29 169 79W 64 44 44 89 45 61 89 67 73 56 38 35 49 79 48 35 42 588 269Y 51 27 23 53 40 51 60 93 55 65 46 19 27 91 36 36 38 1202 227
Table S5.10: Double substitutions at positions dII (L12) and gII (K15).
8
A D E F G H I K L M N P Q R S T V W YA 0 5 14 14 15 16 8 16 10 29 5 14 19 27 23 19 20 31 28D 13 13 18 17 17 17 15 13 23 38 14 21 32 38 16 14 32 38 24E 38 35 34 33 25 33 32 40 23 39 27 24 29 17 17 23 26 36 35F 26 34 36 30 32 31 45 35 42 53 36 43 47 58 36 51 50 79 38G 28 39 30 36 39 20 27 32 36 39 15 30 30 33 31 36 38 63 43H 41 35 36 57 36 60 62 50 30 32 27 42 26 34 30 31 27 56 64I 30 57 71 39 35 32 45 45 46 48 36 30 50 39 37 26 22 52 34K 26 22 32 31 38 30 37 33 32 47 43 30 34 45 32 42 40 87 44L 88 91 201 56 25 73 98 127 156 174 55 25 198 120 77 139 119 115 165M 50 54 48 48 39 45 41 58 45 68 31 32 42 44 44 29 37 64 48N 31 25 39 41 31 52 37 33 31 31 36 36 41 41 30 39 42 43 29P 30 35 23 37 31 43 24 26 30 43 19 31 23 39 38 29 32 67 48Q 24 27 34 39 44 33 23 39 30 44 25 23 35 45 30 34 26 75 35R 34 30 37 58 32 45 53 60 32 58 60 48 48 49 45 40 42 95 44S 27 31 23 35 39 43 37 50 43 45 34 62 42 40 31 45 38 97 54T 37 35 39 34 42 27 32 38 25 42 30 17 34 34 26 31 36 63 50V 33 35 49 32 38 24 46 42 19 52 33 21 48 29 41 26 25 52 31W 53 41 68 93 50 54 54 79 53 73 38 65 67 100 43 47 73 164 228Y 37 37 49 58 31 55 36 39 42 48 44 40 44 54 53 46 48 122 89
Table S5.11: Double substitutions at positions dIII (L19) and gIII (E22).
A D E F G H I K L M N P Q R S T V W YA 9 23 23 19 15 20 22 27 25 28 22 26 18 36 37 22 26 41 22D 36 30 30 29 28 54 38 18 26 31 17 28 17 32 23 19 24 38 16E 13 23 38 27 25 60 44 39 44 41 26 37 59 19 18 30 29 34 23F 30 31 26 36 31 27 40 33 44 40 55 34 42 61 45 55 23 72 67G 43 35 20 27 30 46 19 31 33 22 23 27 17 29 23 17 25 46 45H 51 30 35 26 26 26 28 45 37 36 28 26 29 37 25 34 16 57 25I 42 36 36 43 35 42 82 53 59 67 34 24 50 62 39 26 52 105 58K 26 65 23 34 32 40 25 39 36 35 26 29 39 40 40 35 35 59 33L 77 46 63 72 44 43 328 92 243 121 41 32 90 148 63 68 86 215 155M 45 48 42 39 49 64 33 30 54 53 17 24 54 46 35 35 45 72 40N 17 12 20 43 41 41 26 35 43 32 30 44 26 36 32 19 38 49 18P 29 24 32 39 34 20 13 22 14 31 36 40 34 45 54 23 26 56 51Q 53 38 24 46 41 25 27 24 21 48 37 16 17 28 23 16 22 59 44R 46 40 50 46 31 46 56 52 43 50 42 59 22 52 46 39 30 103 49S 30 44 28 36 23 46 49 38 41 44 33 25 22 50 33 30 25 83 67T 7 12 18 27 22 35 29 46 29 38 30 27 45 31 41 30 21 50 38V 24 32 29 29 27 43 47 27 32 46 23 18 19 30 29 31 33 65 35W 70 71 63 119 72 107 123 94 153 109 61 61 68 197 20 48 69 362 170Y 25 29 18 43 35 46 44 26 89 27 21 14 25 37 28 23 24 166 62
Table S5.12: Double substitutions at positions dIV (L26) and gIV (L29).
9
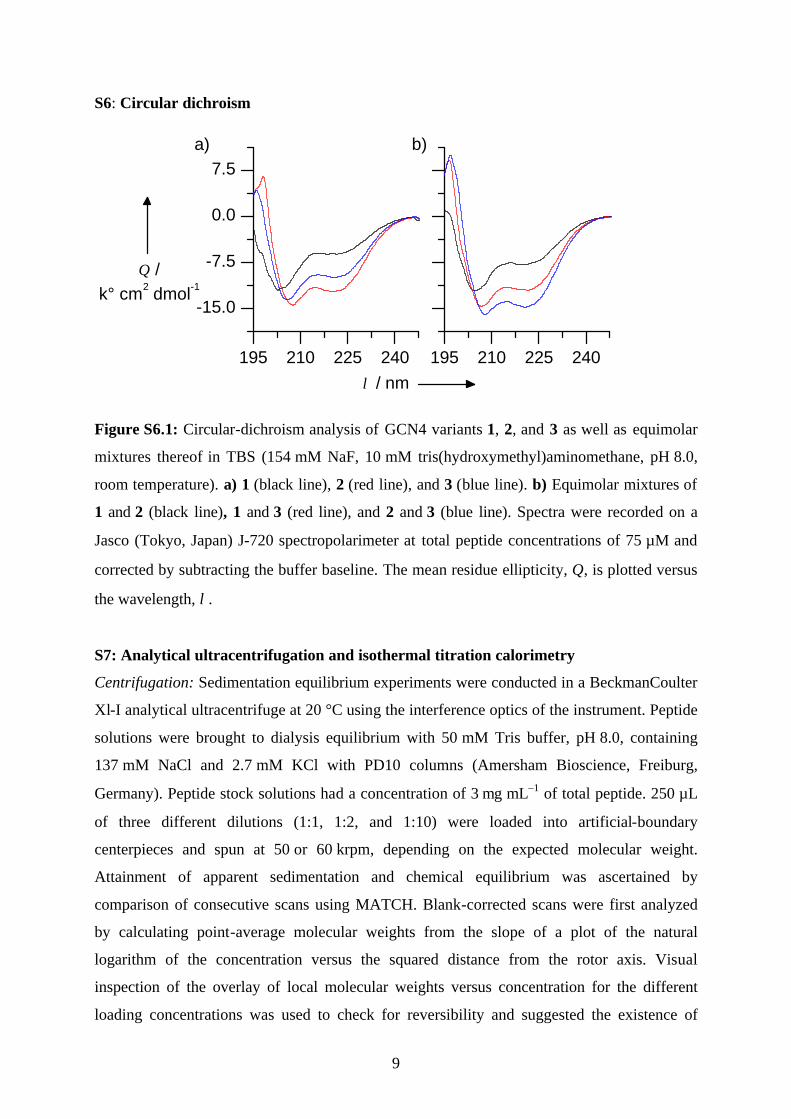
S6: Circular dichroism
195 210 225 240
-15.0
-7.5
0.0
7.5
195 210 225 240
a) b)
Θ /k° cm2 dmol-1
λ / nm
Figure S6.1: Circular-dichroism analysis of GCN4 variants 1, 2, and 3 as well as equimolar
mixtures thereof in TBS (154 mM NaF, 10 mM tris(hydroxymethyl)aminomethane, pH 8.0,
room temperature). a) 1 (black line), 2 (red line), and 3 (blue line). b) Equimolar mixtures of
1 and 2 (black line), 1 and 3 (red line), and 2 and 3 (blue line). Spectra were recorded on a
Jasco (Tokyo, Japan) J-720 spectropolarimeter at total peptide concentrations of 75 µM and
corrected by subtracting the buffer baseline. The mean residue ellipticity, Θ, is plotted versus
the wavelength, λ.
S7: Analytical ultracentrifugation and isothermal titration calorimetry
Centrifugation: Sedimentation equilibrium experiments were conducted in a BeckmanCoulter
Xl-I analytical ultracentrifuge at 20 °C using the interference optics of the instrument. Peptide
solutions were brought to dialysis equilibrium with 50 mM Tris buffer, pH 8.0, containing
137 mM NaCl and 2.7 mM KCl with PD10 columns (Amersham Bioscience, Freiburg,
Germany). Peptide stock solutions had a concentration of 3 mg mL–1 of total peptide. 250 µL
of three different dilutions (1:1, 1:2, and 1:10) were loaded into artificial-boundary
centerpieces and spun at 50 or 60 krpm, depending on the expected molecular weight.
Attainment of apparent sedimentation and chemical equilibrium was ascertained by
comparison of consecutive scans using MATCH. Blank-corrected scans were first analyzed
by calculating point-average molecular weights from the slope of a plot of the natural
logarithm of the concentration versus the squared distance from the rotor axis. Visual
inspection of the overlay of local molecular weights versus concentration for the different
loading concentrations was used to check for reversibility and suggested the existence of

10
limiting species at the lower and upper ends of the concentration scale. Where appropriate,
this information was used to select distinct models to describe the data. These models were
then directly fitted to the data using NONLIN. A model was judged to be an adequate
description of the data if the residuals were random by visual inspection and if no other model
explained the data significantly better as judged by the variances of the fits.[3]
The density of the buffer used and the partial specific volume (PSV) of the different peptides
were measured using a DMA 5000 densitometer (Anton Paar, Graz, Austria) as described
elsewhere.[4] For mixtures of two peptides, the weighted average of the individual values of
the PSV was used to calculate point-average molecular weights. A conversion factor of
3.29 fringes mg–1 mL–1 was used to convert values from fringe units to molar quantities.
Tables S7.1–S7.3 display the fitting results returned by different homoassociation models for
variants 1, 2, and 3 as reported by NONLIN. Values are given in fringe units rather than
molar quantities. Where appropriate, 95%-confidence limits are given below the best-fit
parameters. Parameters for which no confidence limits are given were held fixed. Best-fit
values are shown in bold (d.o.f.: degrees of freedom, σ: reduced buoyant molecular weight as
defined in Reference 4, B: colligative 2nd virial coefficient as reported by NONLIN, n.a.: not
applicable).
variance d.o.f. σ ln(K2 × M) ln(K3 × M2) B
3.9156e–3 2536 1.666 n.a. n.a. –6.22e–3 –6.48e–3/–6.0e–3
7.5418e–4 2536 1.666 –3.602 –3.633/–3.571 n.a. 0
7.2672e–4 2535 1.666 –3.519 –3.617/–3.421
n.a. 2.71e–4 –1.59e–5/5.57e–4
Table S7.1: Homoassociation of variant 1. PSV: 0.720 mL g–1, ρ: 1.007 g mL–1, Mw: 3741 Da, σ60krpm, theoretical: 1.666.
variance d.o.f. σ ln(K2 × M) ln(K3 × M2) B
1.0915e–2 2645 1.232 n.a. –2.518 –2.67/–2.362 0
9.1454e–4 2644 1.232 n.a. –1.644 –1.719/–1.556
1.66e–3 1.58e–3/1.75e–3
1.7804e–3 2644 1.232 –0.244 –0.451/–0.029
–1.204 –1.367/–1.035 0
7.1164e–4 2643 1.232 –1.885 –2.087/–1.692
–1.443 –1.485/–1.401
1.28e–3 1.18e–3/1.38e–3
Table S7.2: Homoassociation of variant 2. PSV: 0.703 mL g–1, ρ: 1.007 g mL–1, Mw: 3748 Da, σ50krpm, theoretical: 1.232.
11
variance d.o.f. σ ln(K2 × M) ln(K3 × M2) B
1.0481e–2 2684 1.216 n.a. –3.474 –3.587/–3.358 0
7.3351e–4 2683 1.216 n.a. –2.823 –2.879/–2.766
1.55e–3 1.47e–3/1.62e–3
2.8006e–3 2683 1.216 –1.506 –1.693/–1.320
–2.9 –2.985/–2.806 0
7.7608e–4 2682 1.216 –17.28 –1.3e4/–9.26e3
–2.836 –2.841/–2.834
1.55e–3 1.56e–3/1.58e–3
Table S7.3: Homoassociation of variant 3. PSV: 0.704 mL g–1, ρ: 1.007 g mL–1, Mw: 3711 Da, σ50krpm, theoretical: 1.216.
Table S7.4 summarizes the homoassociation constants determined for variants 1, 2, and 3.
Again, 95%-confidence limits are given below the best-fit parameters.
variant K2 (M–1) K3 (M–2)
1 182 165/201 n.a.
2 936 765/1136
1.2 × 107 1.15/1.25 × 107
3 n.a. 0.3 × 107 0.28/0.31 × 107
Table S7.4: Homoassociation constants of variants 1, 2, and 3.
Calorimetry: High-sensitivity isothermal titration calorimetry (ITC) was performed at 25 °C
on a VP-ITC (MicroCal, Northampton, USA) by injecting 10-µL aliquots of a concentrated
solution (~1 mM) of variant 1, 2, or 3 into a dilute solution (~100 µM) of another variant.
Both samples were gently degassed under stirring before the experiment to avoid air bubbles.
Baseline subtraction and peak integration were accomplished using Origin 5.0 as described by
the manufacturer (MicroCal Software, Northampton, USA). All reaction heats were
normalized with respect to the molar amount of peptide injected and corrected by subtracting
the heat of dilution (taken as the constant heat measured toward the end of the titration).
Binding constants, K, and binding stoichiometries, N, were calculated with Origin 5.0 using
the “One Set of Sites” model. Table S7.5 lists the K and N values determined by ITC for all
possible combinations of variants 1, 2, and 3.
variants K (M–1) N 1/2 n.a. n.a. 1/3 5.0 × 104 1.0 2/3 1.2 × 105 1.0
Table S7.5: Heteroassociation constants of variants 1, 2, and 3.
12
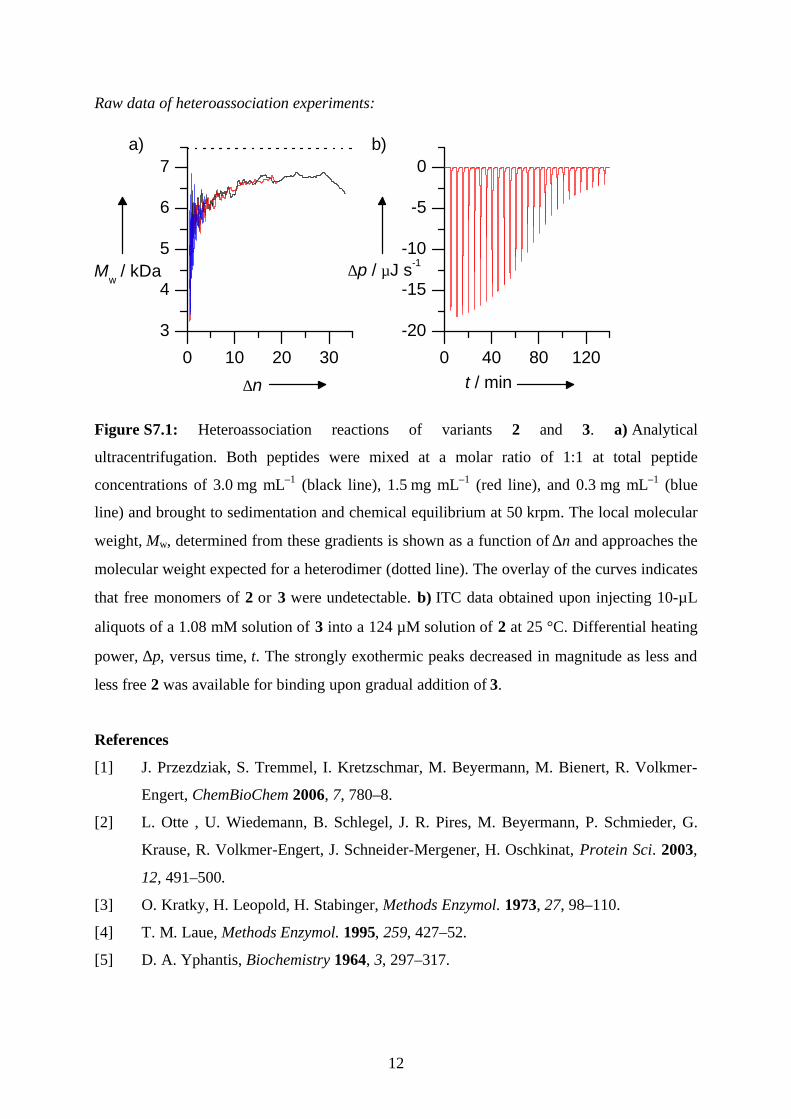
Raw data of heteroassociation experiments:
0 10 20 30
3
4
5
6
7
0 40 80 120
-20
-15
-10
-5
0
∆p / µJ s-1
a) b)
Mw / kDa
∆n t / min
Figure S7.1: Heteroassociation reactions of variants 2 and 3. a) Analytical
ultracentrifugation. Both peptides were mixed at a molar ratio of 1:1 at total peptide
concentrations of 3.0 mg mL–1 (black line), 1.5 mg mL–1 (red line), and 0.3 mg mL–1 (blue
line) and brought to sedimentation and chemical equilibrium at 50 krpm. The local molecular
weight, Mw, determined from these gradients is shown as a function of ∆n and approaches the
molecular weight expected for a heterodimer (dotted line). The overlay of the curves indicates
that free monomers of 2 or 3 were undetectable. b) ITC data obtained upon injecting 10-µL
aliquots of a 1.08 mM solution of 3 into a 124 µM solution of 2 at 25 °C. Differential heating
power, ∆p, versus time, t. The strongly exothermic peaks decreased in magnitude as less and
less free 2 was available for binding upon gradual addition of 3.
References
[1] J. Przezdziak, S. Tremmel, I. Kretzschmar, M. Beyermann, M. Bienert, R. Volkmer-
Engert, ChemBioChem 2006, 7, 780–8.
[2] L. Otte , U. Wiedemann, B. Schlegel, J. R. Pires, M. Beyermann, P. Schmieder, G.
Krause, R. Volkmer-Engert, J. Schneider-Mergener, H. Oschkinat, Protein Sci. 2003,
12, 491–500.
[3] O. Kratky, H. Leopold, H. Stabinger, Methods Enzymol. 1973, 27, 98–110.
[4] T. M. Laue, Methods Enzymol. 1995, 259, 427–52.
[5] D. A. Yphantis, Biochemistry 1964, 3, 297–317.




















