
Gankyrin, a biomarker for epithelial carcinogenesis, is overexpressed in human oral cancer

Research ArticlePromoter Hypermethylation and Suppression ofGlutathione Peroxidase 3 Are Associated with InflammatoryBreast Carcinogenesis
Mona M Mohamed1 Salwa Sabet1 Dun-Fa Peng2 M Akram Nouh3
Mohamed El-Shinawi4 and Wael El-Rifai2
1 Department of Zoology Faculty of Science Cairo University Giza 12613 Egypt2 Department of Surgery Vanderbilt University Medical Center Nashville TN 37232 USA3Department of Pathology National Cancer Institute Cairo University Giza 12613 Egypt4Department of General Surgery Faculty of Medicine Ain Shams University Cairo 11566 Egypt
Correspondence should be addressed to Mona M Mohamed mmostafascicuedueg
Received 25 November 2013 Revised 22 January 2014 Accepted 30 January 2014 Published 20 March 2014
Academic Editor Jeannette Vasquez-Vivar
Copyright copy 2014 Mona M Mohamed et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Reactive oxygen species (ROS) play a crucial role in breast cancer initiation promotion and progression Inhibition of antioxidantenzymes that remove ROS was found to accelerate cancer growth Studies showed that inhibition of glutathione peroxidase-3(GPX3) was associated with cancer progression Although the role of GPX3 has been studied in different cancer types its rolein breast cancer and its epigenetic regulation have not yet been investigated The aim of the present study was to investigateGPX3 expression and epigenetic regulation in carcinoma tissues of breast cancer patientsrsquo in comparison to normal breasttissues Furthermore we compared GPX3 level of expression and methylation status in aggressive phenotype inflammatory breastcancer (IBC) versus non-IBC invasive ductal carcinoma (IDC) We found that GPX3 mRNA and protein expression levels weredownregulated in the carcinoma tissues of IBC compared to non-IBC However we did not detect significant correlation betweenGPX3 and patientsrsquo clinical-pathological prosperities Promoter hypermethylation of GPX3 gene was detected in carcinoma tissuesnot normal breast tissues In addition IBC carcinoma tissues showed a significant increase in the promoter hypermethylationof GPX3 gene compared to non-IBC Our results propose that downregulation of GPX3 in IBC may play a role in the diseaseprogression
1 Introduction
The breast tumor microenvironment is characterized bythe release of endogenous reactive oxygen species (ROS)that resulted from accumulation of different metabolic andpathological changes such as glucose deprivation steroid hor-monesmetabolism by lactoperoxidase [1] mitochondrial dis-order [2] infiltration of macrophages [3] and angiogenesisand reperfusion of blood vessels [4 5] Removal of ROS fromtissues is achieved by the family of glutathione peroxidases(GPXs) (Enzyme Commission number 11119) expressedcytoplasmically and their expression is tissue specific [6]GPXs are known to protect cells against oxidative stress by
catalyzing the reduction of H2O2 organic hydroperoxide and
lipid peroxides by reduced glutathione [6]Among the family of GPXs the isoenzymeGPX3 is found
to play a crucial role in the removal of ROS and healthy tissuedetoxification [6] In addition studies showed that expressionand activity of GPX3 contribute to prevention of cancerinitiation [7 8] Paradoxically GPX3 inhibition is suggestedto be associated with different stages of cancer progressionincluding initiation promotion and metastasis [6] GPX3was found to be downregulated in the plasma of breastgastric and colorectal cancer patients [9] Moreover GPX3downregulation was reported in prostate cancer tissues [10]thyroid cancer [11] and esophageal cancer [12 13] Promoter
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2014 Article ID 787195 9 pageshttpdxdoiorg1011552014787195
2 Oxidative Medicine and Cellular Longevity
hypermethylation mechanism which is a ldquofrequent event inhuman cancersrdquo may result in GPX3 gene silencing and inhi-bition of GPX3 expression [14] GPX3 promoter hypermethy-lation and downregulation were detected in prostate cancer[15] endometrial adenocarcinoma [16] cervical thyroid andlung cancer [17] head and neck carcinoma [14] gastric cancer[18] and multiple myeloma [19] GPX3 hypermethylationcorrelates with disease poor prognosis and resistance tochemotherapy in head and neck cancer patients [14] andmul-tiplemyeloma [19] Recently we found that inactivation of theGPX3 gene by promoter hypermethylation in gastric cancer isassociatedwith high incidence of lymphnodemetastasis [20]
Inflammatory breast cancer (IBC) is an aggressive andhighly metastatic form of breast cancer most prominentamong premenopausal women [21] IBC is characterized byrapid onset over a period of only weeks to a few months andpatients presented with erythema edema of the breast anda ldquopeau drsquoorangerdquo appearance of the skin [22 23] Almost allIBC patients had lymph node metastasis at time of diagnosisDespite the distinct clinical features associated with IBC thegenetic and epigenetic signature underlining the aggressivemetastatic behavior of IBC remains poorly understood [24]Studies showed that in carcinoma tissues ROS induce cellproliferation motility invasion angiogenesis and the escapefrom apoptotic mechanism [25] ROS was found to stimulatecancer cell motility and invasion by activating protein kinase-C (PKC) and the mitogen-activated protein kinase (MAPK)and extracellular signal-regulated kinases (ERKs) signalingpathways thus increasing the risk of metastasis [26 27]Furthermore we found that promoter methylation and lossin copy number of GPX3 gene are associatedwith the numberof lymph nodemetastases in gastric cancer [20] Since almostall IBC patients presented with positive axillary lymph nodemetastasis [21] herein we investigated whether expressionand epigenetic regulation of GPX3 may contribute to theaggressive phenotype IBC versus non-IBC
2 Materials and Methods
21 Reagents DNeasy tissue kit was purchased from Qiagen(Valencia CA USA) EZ DNA Methylation-Gold Kit waspurchased from ZYMO Research (Orange CA USA) DakoAntibody Diluent-Reduce Background and Dako Cytoma-tion EnVision+ Dual Link System-HRP (DAB+) kit (catalognumber K4065) were purchased from Dako (CarpinteriaCA USA) Mouse anti-GPX3 monoclonal antibody Clone23B1 was purchased from Abcam Cambridge MA USAPermount was purchased from Fisher Scientific (Fair LawnNJ USA) Unless otherwise stated all other reagents werefrom Sigma (St Louis MO USA)
22 Patients and Tissues Patients were enrolled from BreastClinics of Ain Shams University Hospitals Ain ShamsUniversity Cairo Egypt after obtaining the InstitutionalReview Board (IRB) approval from the ethics committee ofAin Shams University All patients signed informed consentbefore participating in the study Breast cancer diagnosis was
achieved by clinical examination ultrasound mammogra-phy and biopsy In the present study we enrolled 40 womenclinically diagnosed with breast cancer Patients were dividedinto non-IBC (119899 = 20) and IBC (119899 = 20) subgroups Patientswere considered to harbor IBC when they presented with aswollen breast skin inflammation and edema as previouslydescribed [21] In addition 6 normal breast tissue samplesdonated by healthy volunteers undergoing mammoplastywere used as control
Tissue samples were collected either from preadjuvantchemotherapy tissue biopsies or during modified radicalmastectomy Tissue samples were divided into two piecesone snap-frozen at minus80∘C in RNAlater (Qiagen) and onefixed in 10 neutral buffered formalin and processed forsectioning for routine pathological examination as well as forimmunohistochemistry Pathological data regarding tumorsize tumor grade [28] presence and absence of lymphovascu-lar invasion and dermal and tumor stromal lymphatic emboliwere assessed for routine diagnosis as we described before[21]
23 Assessment of mRNA Expression of GPX3 Using Quan-titative Real-Time Polymerase Chain Reaction (qRT-PCR)We isolated total RNA from frozen breast tissue samplesusing an RNeasy Mini kit (Qiagen) following the manufac-toryrsquos instructions Single-stranded cDNA was synthesizedusing the iScript cDNA Synthesis Kit (Bio-Rad HerculesCA USA) We assessed transcription levels of GPX3 inbreast cancer samples versus normal tissues (control) frombreast of healthy volunteers using qRT-PCR GPX3 primerswere used as we described before (forward 51015840-GCCGGG-GACAAGAGAAGT-31015840 and reverse 51015840-GAGGACGTATTT-GCCAGCAT-31015840) [20] Quantitative real-time PCR (qRT-PCR) reactions were carried out using Bio-Rad IQ SYBRGREEN Supermix (Bio-Rad CA USA) on iCycler (Bio-Rad CA USA) with the threshold cycle number determinedby use of iCycler software version 30 All samples wererun in triplicate and the threshold cycle (Ct) was deter-mined using the iCycler software and then was averagedResults were normalized to internal control Hypoxanthinephosphoribosyltransferase-1 (HPRT1) and GPX3 expressionfold was calculated according to the formula 2(119877
119905minus119864119905)2(119877119899minus
119864119899) as previously described [29] where 119877
119905is the threshold
cycle number for the reference gene observed in the tumor119864119905is the threshold cycle number for the experimental gene
observed in the tumor 119877119899is the threshold cycle number for
the reference gene observed in the normal samples and 119864119899is
the threshold cycle number for the reference gene observedin the tumor 119877
119899and 119864
119899values were calculated as an average
of the 6 normal samples For all primary BACs the gene wasconsidered to be downregulated if the mRNA expression foldwas le05 in comparison with the normal samples [20]
24 Immunohistochemistry Tissue sections of 5 120583m thick-ness were prepared from paraffin blocks of breast carcinomaand normal breast tissues and stained with hematoxylinand eosin to select sections suitable for immunostainingand scoring Tissue sections were first deparaffinized and
Oxidative Medicine and Cellular Longevity 3
rehydrated through graded concentrations of ethanol Forantigen retrieval slides were incubated in citrate buffer (pH60) in a water bath for 1 h at 99∘C Slides were cooledby incubation in Tris-buffered saline (TBS 005molL Tris-HCl pH 76 015molL NaCl and 005 Tween 20) for20min Endogenous peroxidase activity was blocked byusing Dako Dual Endogenous Enzyme Block for 10min Forimmunohistochemical stain of GPX3 tissue sections wereincubated for 1 h at room temperature with the primaryantibody mouse anti-GPX3 (mouse anti-GPX3 monoclonalantibody Clone 23B1 Abcam)Themonoclonal antibodywasdiluted 1 100 inDakoAntibodyDiluent-Reduce Backgroundproduct (DAKO) Detection was carried out by incubatingsections with 100 120583L of Horseradish Peroxidase (HRP) Rab-bitMouse (EnVision+ Dual Link System-HRP diaminoben-zidine (DAB+)) for 45min Staining was achieved by adding100 120583L of DAB+ diluted 1 50 in substrate buffer [EnVision+Dual Link System-HRP (DAB+)] Staining was progressedfor 15min until the development of brown color Nucleiwere counterstained with hematoxylin rinsed in PBS andmounted using Permount formicroscopic examination Neg-ative control slides were run in parallel with each markerwhere the primary antibody is replaced by PBS The level ofexpression of GPX3 protein was scored according to both theintensity and the positivity of the stain of the cells within theentire slide negative no immunostaining is observed score(+) less than 10 of cells showed no or weak staining score(++) 10ndash50 of cells showed moderate to strong stainingand score (+++) more than 50 of cells showed strongstaining [21]
25 DNA Extraction from Fresh Tissue and Identificationof CpG Islands in the Promoter Region of GPX3 GeneGenomic DNA was extracted from 25mg of fresh tissueof biopsy or modified radical mastectomy using DNeasytissue kit Qiagen In the last step of the protocol DNAwas eluted in 200 120583L elution buffer We used Universityof Californiarsquos (UCSC) Genome Browser website (httpgenomeucscedu) to obtain DNA sequences around thepromoter region Identified sequences were confirmed to beidentical to that from the DBTSS (database of transcriptionalstart sites httpdbtsshgcjp) as described [29] The CpGisland in the promoter region of GPX3 was defined usingCpG island searcher online tool (httpwwwuscnorriscomcpgislands2cpgaspx)
26 DNA Bisulfite Treatment and Methylation-Specific PCR(MSP) Bisulfite modification of the purified DNA wasachieved by using an EZ DNAmethylation-Gold kit (ZYMOResearch Orange CA USA) following the manufactoryrsquosinstructionsWe used 1120583g of extracted genomic DNA of eachsample from normal breast tissue samples IBC and non-IBC breast carcinoma tissue samples [12 29] The bisulfitetreatedDNAwas subjected toMSP in a final reaction of 50120583LWe designed primers for MSP targeting CpG-rich promoterregion using online software ldquoMethPrimerrdquo (httpwwwurogeneorgmethprimer)The forward and reverse primers
used for methylated GPX3 were 51015840-GTTGAGGGTAAG-TCGCGTTC-31015840 and 51015840-GTCCGTCTAAAATATCCGACG-31015840 and those for unmethylated GPX3 were 51015840-GAGTTG-AGGGTAAGTTGTGTTTGT-31015840 and 51015840-CCATCCATC-TAAAATATCCAACACT-31015840 PCR mixture included thePlatinumPCR SuperMixHigh Fidelity (Invitrogen CarlsbadCA USA) and the program was adjusted as follows initialdenaturation at 94∘C for 5min then 35 cycles consisting eachof 94∘C for 1min annealing at 54∘C for 1min and finallyextension at 72∘C for 1min PCR products were subjectedto electrophoresis on 15 agarose gel using GelRed nucleicacid stain (Biotium Hayward CA USA) and amplified DNAwas visualized by 300 nm transillumination For quantitativeanalysis visualized bands of the agarose gel were analyzedby ImageJ (National Institutes of Health Bethesda MAUSA) software For each carcinoma tissue specimen the bandintensity of the methylated and unmethylated MSP productswas quantified and normalized against gel background asdescribed elsewhere [30]
27 Statistical Analysis The data was analyzed using SPSSsoftware version 160 Differences were evaluated by Studentrsquos119905-test and Fisherrsquos exact test 119875 lt 005 was considered asstatistically significant
3 Results
31 Clinical and Pathological Features of IBC and Non-IBCPatients The clinical and pathological features of non-IBCand IBC patients are presented in Table 1 All of IBC patientswere premenopausal one decade younger than non-IBCpatients (119875 lt 0001) Statistical analysis revealed that IBCpatients showed a significant higher incidence (119875 lt 001)of positive metastatic lymph nodes compared to non-IBCpatients In addition a significant difference (119875 lt 0001)in lymphovascular invasion and dermal lymphatic emboliwas highly detected in IBC versus non-IBC patients tissuesections
32 Downregulation of the GPX3 Protein Expression in BreastCarcinoma Tissue Level of expression of GPX3 proteinwas assessed in normal and carcinoma breast tissues usingIHC GPX3 expression staining results were scored for thepositivity and intensity (Table 2) Statistical analysis using theChi-square test revealed that normal breast tissues expresssignificantly high level of GPX3 protein (Figure 1(a)) com-pared to breast carcinoma tissues of non-IBC (Figure 1(b))and IBC (Figure 1(c)) patients (119875 = 0001 and 119875 lt 0001resp) In addition non-IBC tissues showed a significantincrease (119875 = 0043) in the level of expression of GPX3 ascompared to those of IBC patients
33 Suppression of GPX3mRNAExpression in IBC CarcinomaTissue Samples Inhibition of GPX3 protein expression in thecarcinoma tissues of breast cancer patients was confirmedat mRNA level by using qRT-PCR We found that the levelof GPX3 mRNA expression in normal breast tissues wassignificantly higher than that in breast carcinoma tissue
4 Oxidative Medicine and Cellular Longevity
Table 1 Patientsrsquo clinical and pathological data
Characteristics Non-IBC119899 = 20 ()
IBC119899 = 20 () 119875 value
AgeMean plusmn SE 5435 plusmn 21 4055 plusmn 17
le50 7 (35) 18 (90)lt0001lowast
gt50 13 (65) 2 (10)Tumor sizelowastlowast
Mean plusmn SE 512 plusmn 05 708 plusmn 06
le2 cm 1 (5) 0gt005
gt2 cm 19 (95) 18 (90)Tumor grade
G2 15 (75) 14 (70)gt005
G3 5 (25) 6 (30)Lymph node statuslowastlowast
Positive 17 (85) 18 (90)gt005
Negative 3 (15) 0Number of lymph nodeslowastlowast
le6 16 (80) 6 (333)lt001lowast
gt6 4 (20) 12 (667)Lymphovascular invasion
Positive 2 (10) 14 (70)lt0001lowast
Negative 18 (90) 6 (30)Dermal lymphatic emboli
Positive 2 (10) 18 (90)lt0001lowast
Negative 18 (90) 2 (10)ER status
Positive 7 (35) 6 (30)gt005
Negative 13 (65) 14 (70)PR status
Positive 9 (45) 7 (35)gt005
Negative 11 (55) 13 (65)HER-2 status
Positive 2 (10) 4 (20)gt005
Negative 18 (90) 16 (80)lowastSignificant 119875 value calculated by Studentrsquos 119905-testlowastlowast119899 = 18 in IBC patients
Table 2 Scoring of GPX3 expression in normal non-IBC and IBC breast tissues
Score Normal (119899 = 6) Non-IBC (119899 = 16)a IBC (119899 = 16)bc
119899 () 119899 (asymp) 119899 (asymp)Negative 0 (0) 5 (312) 11 (687)+ 0 (0) 7 (438) 3 (187)++ 3 (50) 4 (25) 2 (125)+++ 3 (50) 0 0Significant 119875 value calculated by the Chi-square testaSignificant 119875 value (119875 = 0001) when normal tissues were compared to non-IBC tissuesbSignificant 119875 value (119875 lt 0001) when normal tissues were compared to IBC tissuescSignificant 119875 value (119875 = 0043) when non-IBC tissues were compared to IBC tissues119899 = number of patients
Oxidative Medicine and Cellular Longevity 5
(a) (b) (c)
Figure 1 GPX3 protein is downregulated in IBC Microscopic images representative of IHC stain of GPX3 (brown color) in (a) normalbreast tissues showing moderate to marked intensity of GPX3 (b) non-IBC tissue sections showing mild intensity of GPX3 and (c) IBCtissue sections with no immunostaining of GPX3 by carcinoma cells within tumor emboli (magnification upper panel 10x lower panel40x)
samples of non-IBC and IBC (119875 = 0001 and 119875 lt0001 resp) We compared the level of mRNA expressionof GPX3 in non-IBC and IBC and our results revealedthat mRNA expression level of GPX3 expression in non-IBC carcinoma tissues was statistically significant (119875 =0036) higher than that in IBC (Figure 2) Using the Pearsoncorrelation statistical analysis we did not detect significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties (tumor size tumorgrade and number of axillary metastatic lymph nodes) innon-IBC and IBC patients However downregulation ofGPX3 in IBC versus non-IBC carcinoma tissues suggests thatGPX3 may play a role in IBC disease progression
34 Methylation Profile of GPX3 Promoter Region in BreastCancer Tissue Samples Analysis of the promoter region ofGPX3 indicated that CpG islands were present within minus1000to +300 bp of the gene We assessed the GPX3 promoterregion methylation in 40 breast cancer tissue samples and 6normal breast tissue samples Agarose gel electrophoresis ofall normal breast tissues showed only one band correspond-ing to unmethylated GPX3-MSP products (Figure 3(a)) Onthe contrary agarose gel electrophoresis of all GPX3-MSPproducts breast carcinoma tissue samples showed bands of200 bp corresponding to both unmethylated (Figure 3(a))and methylated GPX3-MSP products (Figure 3(b)) Thepresent results showed that GPX3 methylation was detectedin breast carcinoma tissues and not in normal breast tissuesThus GPX3 promoter methylation is responsible for thedownregulation of GPX3 mRNA in breast carcinoma tissues
since it was not detected in normal breast tissues It shouldbe noted that detection of methylated and unmethylatedproducts in some breast carcinoma tissue homogenates maybe due to heterogeneous population of breast carcinoma cellsandor the presence of other normal stromal cells from breasttumor microenvironment
For quantitative analysis visualized bands of agarosegel were analyzed by ImageJ (National Institutes of HealthBethesda MA USA) software For each carcinoma tissuespecimen the band intensity of themethylated and unmethy-lated GPX3-MSP products was quantified and normalizedagainst gel background as described elsewhere [30] Intensityvalues of methylatedunmethylated (MU) ratio for GPX3-MSP products for non-IBC (119899 = 20) and IBC (119899 = 20)patients were analyzed by Studentrsquos 119905-test Normal breasttissues (N) with anMU ratio = 0 (no methylated bands weredetected from MSP) were assessed as unmethylated Resultsrevealed a significant increase (119875 = 004) in the MU ratiosof GPX3-MSP products in IBC versus non-IBC carcinomatissues (Figure 3(c)) Significant increase inMU ratio in IBCversus non-IBC suggests a potential role for GPX3 in IBCdisease progression
4 Discussion
Elevated levels of ROS detected in breast cancer were foundto play a crucial role in the disease progression [4] Forinstance carcinoma cells utilize ROS to stimulate cancercell proliferation motility invasion angiogenesis and escapeof apoptotic mechanism [25] In addition ROS augment
6 Oxidative Medicine and Cellular Longevity
ControlNon-IBCIBC
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100Re
lativ
e mRN
A ex
pres
sion
Control Non-IBC IBC
P = 0036
Figure 2 Expression of GPX3mRNA is downregulated in IBC Blotrepresents the mean fold change of GPX3 mRNA measured by RT-PCR in normal breast tissues IBC and non-IBC carcinoma tissuesStatistical analysis revealed a significant increase in expression ofGPX3 mRNA in normal breast tissues compared to non-IBC andIBC carcinoma tissues (119875 = 0001 and 119875 lt 0001 resp) Incarcinoma tissues of non-IBC patients the level of expression ofGPX3 mRNA is significantly higher (119875 = 0036) than that of IBCResults are representative of at least three independent experimentsData are expressed as mean plusmn SD and 119875 value was determined byStudentrsquos 119905-test
carcinoma cell motility and invasion by activating proteinkinase-C (PKC) and the ERKMAPK signaling pathwaysthus increasing the risk of metastasis [26 27] Failure toremove exogenous and endogenous ROS may occur due todefect in the cellular antioxidant system of carcinoma cellsrepresented by the inhibition of the activity of antioxidantenzymes as GPXs familymembersThus drugs which induceexpression of antioxidant enzymes were suggested for cancertreatment [29] GPXs are categorized into two types onetype is selenium-dependent catalytic activity (GPX-1 -2 -3-4 and 6) and the second type is non-selenium-dependent(GPX-5 and -7) [6] Downregulation and genetic imbalanceamong GPXs were found to play a key role in breast cancerFor example GPX1 gene allelic variants and loss of heterozy-gosity (LOH) at 3p213p region contribute to breast cancerdevelopment [31] Low expression of GPX4 in breast-invasiveductal carcinoma correlated with high tumor grade and poorprognosis of breast cancer patients [32] Low expression ofGPX3 significantly correlates with high risk of breast cancerlocal recurrence among early-stage invasive breast cancerpatients regardless of patientsrsquo clinic-pathological criteria[33]
In fact GPX3 is an essential enzyme responsible for theremoval of ROS in healthy tissues On the contrary GPX3was found to be downregulated in carcinoma tissues of breastgastric and colorectal cancer patients [9] prostate cancer[10] thyroid cancer [11] and esophageal cancer [12] Theimpaired function of GPX3 would result in the accumulationof an increased amount of hydrogen peroxide and other ROSwhich may induce breast carcinogenesis via induction ofoxidative DNA damage genetic instability neoplastic trans-formation [34] and mutation of the p53 tumor suppressorgene [35] Epigenetic mechanisms such as DNA hypermethy-lation and histonemodificationmay repress the expression ofGPX3 For instance treatment of SKGT4 esophageal cancercell lines [29] and endometrial tumor cell lines (NUT12 andNUT81) [16] that show GPX3 promoter hypermethylationwith the demethylating agent 5-aza-21015840-deoxycytidine (5Aza-dC) and the histone deacetylace inhibitor trichostatin A(TSA) results in re-expression of GPX3 mRNAThese resultssuggest that DNA hypermethylation and histone deacetyla-tion may act together to regulate the expression of GPX3mRNA Paradoxically downregulation of GPX3 in absenceof promoter hypermethylation is associated with GPX3 genedeletion in endometrial tumor cell line (NUT84) [16] In factGPX3 promoter hypermethylation is linked to downregula-tion of GPX3 expression in different types of cancer cells andtreatment with 5-Aza to human esophageal adenocarcinomacancer cells SKGT4 [29] and human myeloma cells KMS11[19] restores GPX3 gene expression
Our previous studies demonstrated that GPX3was down-regulated in Barrettrsquos carcinoma due to hypermethylation ofthe promoter region [12] In addition we found that lossin DNA copy number hypermethylation of the promoterregion and downregulation of mRNA expression of GPX3are associated with lymph node metastasis in gastric car-cinomas Reactivation of GPX3 in gastric adenocarcinomacell line AGS inhibits cell motility as assessed by woundhealing assay [20] Herein we studied the role of GPX3 inbreast carcinogenesis We analyzed the level of expressionof GPX3 in normal breast tissues obtained from healthyvolunteers during mammoplasty and non-IBC and IBCbreast carcinoma tissues Our results revealed that GPX3protein level and mRNA were significantly expressed innormal breast tissues and downregulated in breast carcinomatissues The present results are consistent with other studieswhich proved that GPX3 is downregulated in carcinomatissues such as prostate [10] thyroid [11] and esophageal [12]When we compared carcinoma tissues of IBC with non-IBCsamples we detected a significant decrease in mRNA andprotein expression of GPX3 in IBC tissue samples versusnon-IBC tissue samples We did not detect any significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties The present resultsagree with other studies that showed downregulation ofGPX3 in endometrial adenocarcinoma [16] and early invasivebreast carcinoma [33] regardless of patientsrsquo clinical andpathological criteria
IBC is an aggressive phenotype characterized by highmetastatic potential disease recurrence and resistance tochemotherapy [36] Furthermore recently we found that IBC
Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

2 Oxidative Medicine and Cellular Longevity
hypermethylation mechanism which is a ldquofrequent event inhuman cancersrdquo may result in GPX3 gene silencing and inhi-bition of GPX3 expression [14] GPX3 promoter hypermethy-lation and downregulation were detected in prostate cancer[15] endometrial adenocarcinoma [16] cervical thyroid andlung cancer [17] head and neck carcinoma [14] gastric cancer[18] and multiple myeloma [19] GPX3 hypermethylationcorrelates with disease poor prognosis and resistance tochemotherapy in head and neck cancer patients [14] andmul-tiplemyeloma [19] Recently we found that inactivation of theGPX3 gene by promoter hypermethylation in gastric cancer isassociatedwith high incidence of lymphnodemetastasis [20]
Inflammatory breast cancer (IBC) is an aggressive andhighly metastatic form of breast cancer most prominentamong premenopausal women [21] IBC is characterized byrapid onset over a period of only weeks to a few months andpatients presented with erythema edema of the breast anda ldquopeau drsquoorangerdquo appearance of the skin [22 23] Almost allIBC patients had lymph node metastasis at time of diagnosisDespite the distinct clinical features associated with IBC thegenetic and epigenetic signature underlining the aggressivemetastatic behavior of IBC remains poorly understood [24]Studies showed that in carcinoma tissues ROS induce cellproliferation motility invasion angiogenesis and the escapefrom apoptotic mechanism [25] ROS was found to stimulatecancer cell motility and invasion by activating protein kinase-C (PKC) and the mitogen-activated protein kinase (MAPK)and extracellular signal-regulated kinases (ERKs) signalingpathways thus increasing the risk of metastasis [26 27]Furthermore we found that promoter methylation and lossin copy number of GPX3 gene are associatedwith the numberof lymph nodemetastases in gastric cancer [20] Since almostall IBC patients presented with positive axillary lymph nodemetastasis [21] herein we investigated whether expressionand epigenetic regulation of GPX3 may contribute to theaggressive phenotype IBC versus non-IBC
2 Materials and Methods
21 Reagents DNeasy tissue kit was purchased from Qiagen(Valencia CA USA) EZ DNA Methylation-Gold Kit waspurchased from ZYMO Research (Orange CA USA) DakoAntibody Diluent-Reduce Background and Dako Cytoma-tion EnVision+ Dual Link System-HRP (DAB+) kit (catalognumber K4065) were purchased from Dako (CarpinteriaCA USA) Mouse anti-GPX3 monoclonal antibody Clone23B1 was purchased from Abcam Cambridge MA USAPermount was purchased from Fisher Scientific (Fair LawnNJ USA) Unless otherwise stated all other reagents werefrom Sigma (St Louis MO USA)
22 Patients and Tissues Patients were enrolled from BreastClinics of Ain Shams University Hospitals Ain ShamsUniversity Cairo Egypt after obtaining the InstitutionalReview Board (IRB) approval from the ethics committee ofAin Shams University All patients signed informed consentbefore participating in the study Breast cancer diagnosis was
achieved by clinical examination ultrasound mammogra-phy and biopsy In the present study we enrolled 40 womenclinically diagnosed with breast cancer Patients were dividedinto non-IBC (119899 = 20) and IBC (119899 = 20) subgroups Patientswere considered to harbor IBC when they presented with aswollen breast skin inflammation and edema as previouslydescribed [21] In addition 6 normal breast tissue samplesdonated by healthy volunteers undergoing mammoplastywere used as control
Tissue samples were collected either from preadjuvantchemotherapy tissue biopsies or during modified radicalmastectomy Tissue samples were divided into two piecesone snap-frozen at minus80∘C in RNAlater (Qiagen) and onefixed in 10 neutral buffered formalin and processed forsectioning for routine pathological examination as well as forimmunohistochemistry Pathological data regarding tumorsize tumor grade [28] presence and absence of lymphovascu-lar invasion and dermal and tumor stromal lymphatic emboliwere assessed for routine diagnosis as we described before[21]
23 Assessment of mRNA Expression of GPX3 Using Quan-titative Real-Time Polymerase Chain Reaction (qRT-PCR)We isolated total RNA from frozen breast tissue samplesusing an RNeasy Mini kit (Qiagen) following the manufac-toryrsquos instructions Single-stranded cDNA was synthesizedusing the iScript cDNA Synthesis Kit (Bio-Rad HerculesCA USA) We assessed transcription levels of GPX3 inbreast cancer samples versus normal tissues (control) frombreast of healthy volunteers using qRT-PCR GPX3 primerswere used as we described before (forward 51015840-GCCGGG-GACAAGAGAAGT-31015840 and reverse 51015840-GAGGACGTATTT-GCCAGCAT-31015840) [20] Quantitative real-time PCR (qRT-PCR) reactions were carried out using Bio-Rad IQ SYBRGREEN Supermix (Bio-Rad CA USA) on iCycler (Bio-Rad CA USA) with the threshold cycle number determinedby use of iCycler software version 30 All samples wererun in triplicate and the threshold cycle (Ct) was deter-mined using the iCycler software and then was averagedResults were normalized to internal control Hypoxanthinephosphoribosyltransferase-1 (HPRT1) and GPX3 expressionfold was calculated according to the formula 2(119877
119905minus119864119905)2(119877119899minus
119864119899) as previously described [29] where 119877
119905is the threshold
cycle number for the reference gene observed in the tumor119864119905is the threshold cycle number for the experimental gene
observed in the tumor 119877119899is the threshold cycle number for
the reference gene observed in the normal samples and 119864119899is
the threshold cycle number for the reference gene observedin the tumor 119877
119899and 119864
119899values were calculated as an average
of the 6 normal samples For all primary BACs the gene wasconsidered to be downregulated if the mRNA expression foldwas le05 in comparison with the normal samples [20]
24 Immunohistochemistry Tissue sections of 5 120583m thick-ness were prepared from paraffin blocks of breast carcinomaand normal breast tissues and stained with hematoxylinand eosin to select sections suitable for immunostainingand scoring Tissue sections were first deparaffinized and
Oxidative Medicine and Cellular Longevity 3
rehydrated through graded concentrations of ethanol Forantigen retrieval slides were incubated in citrate buffer (pH60) in a water bath for 1 h at 99∘C Slides were cooledby incubation in Tris-buffered saline (TBS 005molL Tris-HCl pH 76 015molL NaCl and 005 Tween 20) for20min Endogenous peroxidase activity was blocked byusing Dako Dual Endogenous Enzyme Block for 10min Forimmunohistochemical stain of GPX3 tissue sections wereincubated for 1 h at room temperature with the primaryantibody mouse anti-GPX3 (mouse anti-GPX3 monoclonalantibody Clone 23B1 Abcam)Themonoclonal antibodywasdiluted 1 100 inDakoAntibodyDiluent-Reduce Backgroundproduct (DAKO) Detection was carried out by incubatingsections with 100 120583L of Horseradish Peroxidase (HRP) Rab-bitMouse (EnVision+ Dual Link System-HRP diaminoben-zidine (DAB+)) for 45min Staining was achieved by adding100 120583L of DAB+ diluted 1 50 in substrate buffer [EnVision+Dual Link System-HRP (DAB+)] Staining was progressedfor 15min until the development of brown color Nucleiwere counterstained with hematoxylin rinsed in PBS andmounted using Permount formicroscopic examination Neg-ative control slides were run in parallel with each markerwhere the primary antibody is replaced by PBS The level ofexpression of GPX3 protein was scored according to both theintensity and the positivity of the stain of the cells within theentire slide negative no immunostaining is observed score(+) less than 10 of cells showed no or weak staining score(++) 10ndash50 of cells showed moderate to strong stainingand score (+++) more than 50 of cells showed strongstaining [21]
25 DNA Extraction from Fresh Tissue and Identificationof CpG Islands in the Promoter Region of GPX3 GeneGenomic DNA was extracted from 25mg of fresh tissueof biopsy or modified radical mastectomy using DNeasytissue kit Qiagen In the last step of the protocol DNAwas eluted in 200 120583L elution buffer We used Universityof Californiarsquos (UCSC) Genome Browser website (httpgenomeucscedu) to obtain DNA sequences around thepromoter region Identified sequences were confirmed to beidentical to that from the DBTSS (database of transcriptionalstart sites httpdbtsshgcjp) as described [29] The CpGisland in the promoter region of GPX3 was defined usingCpG island searcher online tool (httpwwwuscnorriscomcpgislands2cpgaspx)
26 DNA Bisulfite Treatment and Methylation-Specific PCR(MSP) Bisulfite modification of the purified DNA wasachieved by using an EZ DNAmethylation-Gold kit (ZYMOResearch Orange CA USA) following the manufactoryrsquosinstructionsWe used 1120583g of extracted genomic DNA of eachsample from normal breast tissue samples IBC and non-IBC breast carcinoma tissue samples [12 29] The bisulfitetreatedDNAwas subjected toMSP in a final reaction of 50120583LWe designed primers for MSP targeting CpG-rich promoterregion using online software ldquoMethPrimerrdquo (httpwwwurogeneorgmethprimer)The forward and reverse primers
used for methylated GPX3 were 51015840-GTTGAGGGTAAG-TCGCGTTC-31015840 and 51015840-GTCCGTCTAAAATATCCGACG-31015840 and those for unmethylated GPX3 were 51015840-GAGTTG-AGGGTAAGTTGTGTTTGT-31015840 and 51015840-CCATCCATC-TAAAATATCCAACACT-31015840 PCR mixture included thePlatinumPCR SuperMixHigh Fidelity (Invitrogen CarlsbadCA USA) and the program was adjusted as follows initialdenaturation at 94∘C for 5min then 35 cycles consisting eachof 94∘C for 1min annealing at 54∘C for 1min and finallyextension at 72∘C for 1min PCR products were subjectedto electrophoresis on 15 agarose gel using GelRed nucleicacid stain (Biotium Hayward CA USA) and amplified DNAwas visualized by 300 nm transillumination For quantitativeanalysis visualized bands of the agarose gel were analyzedby ImageJ (National Institutes of Health Bethesda MAUSA) software For each carcinoma tissue specimen the bandintensity of the methylated and unmethylated MSP productswas quantified and normalized against gel background asdescribed elsewhere [30]
27 Statistical Analysis The data was analyzed using SPSSsoftware version 160 Differences were evaluated by Studentrsquos119905-test and Fisherrsquos exact test 119875 lt 005 was considered asstatistically significant
3 Results
31 Clinical and Pathological Features of IBC and Non-IBCPatients The clinical and pathological features of non-IBCand IBC patients are presented in Table 1 All of IBC patientswere premenopausal one decade younger than non-IBCpatients (119875 lt 0001) Statistical analysis revealed that IBCpatients showed a significant higher incidence (119875 lt 001)of positive metastatic lymph nodes compared to non-IBCpatients In addition a significant difference (119875 lt 0001)in lymphovascular invasion and dermal lymphatic emboliwas highly detected in IBC versus non-IBC patients tissuesections
32 Downregulation of the GPX3 Protein Expression in BreastCarcinoma Tissue Level of expression of GPX3 proteinwas assessed in normal and carcinoma breast tissues usingIHC GPX3 expression staining results were scored for thepositivity and intensity (Table 2) Statistical analysis using theChi-square test revealed that normal breast tissues expresssignificantly high level of GPX3 protein (Figure 1(a)) com-pared to breast carcinoma tissues of non-IBC (Figure 1(b))and IBC (Figure 1(c)) patients (119875 = 0001 and 119875 lt 0001resp) In addition non-IBC tissues showed a significantincrease (119875 = 0043) in the level of expression of GPX3 ascompared to those of IBC patients
33 Suppression of GPX3mRNAExpression in IBC CarcinomaTissue Samples Inhibition of GPX3 protein expression in thecarcinoma tissues of breast cancer patients was confirmedat mRNA level by using qRT-PCR We found that the levelof GPX3 mRNA expression in normal breast tissues wassignificantly higher than that in breast carcinoma tissue
4 Oxidative Medicine and Cellular Longevity
Table 1 Patientsrsquo clinical and pathological data
Characteristics Non-IBC119899 = 20 ()
IBC119899 = 20 () 119875 value
AgeMean plusmn SE 5435 plusmn 21 4055 plusmn 17
le50 7 (35) 18 (90)lt0001lowast
gt50 13 (65) 2 (10)Tumor sizelowastlowast
Mean plusmn SE 512 plusmn 05 708 plusmn 06
le2 cm 1 (5) 0gt005
gt2 cm 19 (95) 18 (90)Tumor grade
G2 15 (75) 14 (70)gt005
G3 5 (25) 6 (30)Lymph node statuslowastlowast
Positive 17 (85) 18 (90)gt005
Negative 3 (15) 0Number of lymph nodeslowastlowast
le6 16 (80) 6 (333)lt001lowast
gt6 4 (20) 12 (667)Lymphovascular invasion
Positive 2 (10) 14 (70)lt0001lowast
Negative 18 (90) 6 (30)Dermal lymphatic emboli
Positive 2 (10) 18 (90)lt0001lowast
Negative 18 (90) 2 (10)ER status
Positive 7 (35) 6 (30)gt005
Negative 13 (65) 14 (70)PR status
Positive 9 (45) 7 (35)gt005
Negative 11 (55) 13 (65)HER-2 status
Positive 2 (10) 4 (20)gt005
Negative 18 (90) 16 (80)lowastSignificant 119875 value calculated by Studentrsquos 119905-testlowastlowast119899 = 18 in IBC patients
Table 2 Scoring of GPX3 expression in normal non-IBC and IBC breast tissues
Score Normal (119899 = 6) Non-IBC (119899 = 16)a IBC (119899 = 16)bc
119899 () 119899 (asymp) 119899 (asymp)Negative 0 (0) 5 (312) 11 (687)+ 0 (0) 7 (438) 3 (187)++ 3 (50) 4 (25) 2 (125)+++ 3 (50) 0 0Significant 119875 value calculated by the Chi-square testaSignificant 119875 value (119875 = 0001) when normal tissues were compared to non-IBC tissuesbSignificant 119875 value (119875 lt 0001) when normal tissues were compared to IBC tissuescSignificant 119875 value (119875 = 0043) when non-IBC tissues were compared to IBC tissues119899 = number of patients
Oxidative Medicine and Cellular Longevity 5
(a) (b) (c)
Figure 1 GPX3 protein is downregulated in IBC Microscopic images representative of IHC stain of GPX3 (brown color) in (a) normalbreast tissues showing moderate to marked intensity of GPX3 (b) non-IBC tissue sections showing mild intensity of GPX3 and (c) IBCtissue sections with no immunostaining of GPX3 by carcinoma cells within tumor emboli (magnification upper panel 10x lower panel40x)
samples of non-IBC and IBC (119875 = 0001 and 119875 lt0001 resp) We compared the level of mRNA expressionof GPX3 in non-IBC and IBC and our results revealedthat mRNA expression level of GPX3 expression in non-IBC carcinoma tissues was statistically significant (119875 =0036) higher than that in IBC (Figure 2) Using the Pearsoncorrelation statistical analysis we did not detect significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties (tumor size tumorgrade and number of axillary metastatic lymph nodes) innon-IBC and IBC patients However downregulation ofGPX3 in IBC versus non-IBC carcinoma tissues suggests thatGPX3 may play a role in IBC disease progression
34 Methylation Profile of GPX3 Promoter Region in BreastCancer Tissue Samples Analysis of the promoter region ofGPX3 indicated that CpG islands were present within minus1000to +300 bp of the gene We assessed the GPX3 promoterregion methylation in 40 breast cancer tissue samples and 6normal breast tissue samples Agarose gel electrophoresis ofall normal breast tissues showed only one band correspond-ing to unmethylated GPX3-MSP products (Figure 3(a)) Onthe contrary agarose gel electrophoresis of all GPX3-MSPproducts breast carcinoma tissue samples showed bands of200 bp corresponding to both unmethylated (Figure 3(a))and methylated GPX3-MSP products (Figure 3(b)) Thepresent results showed that GPX3 methylation was detectedin breast carcinoma tissues and not in normal breast tissuesThus GPX3 promoter methylation is responsible for thedownregulation of GPX3 mRNA in breast carcinoma tissues
since it was not detected in normal breast tissues It shouldbe noted that detection of methylated and unmethylatedproducts in some breast carcinoma tissue homogenates maybe due to heterogeneous population of breast carcinoma cellsandor the presence of other normal stromal cells from breasttumor microenvironment
For quantitative analysis visualized bands of agarosegel were analyzed by ImageJ (National Institutes of HealthBethesda MA USA) software For each carcinoma tissuespecimen the band intensity of themethylated and unmethy-lated GPX3-MSP products was quantified and normalizedagainst gel background as described elsewhere [30] Intensityvalues of methylatedunmethylated (MU) ratio for GPX3-MSP products for non-IBC (119899 = 20) and IBC (119899 = 20)patients were analyzed by Studentrsquos 119905-test Normal breasttissues (N) with anMU ratio = 0 (no methylated bands weredetected from MSP) were assessed as unmethylated Resultsrevealed a significant increase (119875 = 004) in the MU ratiosof GPX3-MSP products in IBC versus non-IBC carcinomatissues (Figure 3(c)) Significant increase inMU ratio in IBCversus non-IBC suggests a potential role for GPX3 in IBCdisease progression
4 Discussion
Elevated levels of ROS detected in breast cancer were foundto play a crucial role in the disease progression [4] Forinstance carcinoma cells utilize ROS to stimulate cancercell proliferation motility invasion angiogenesis and escapeof apoptotic mechanism [25] In addition ROS augment
6 Oxidative Medicine and Cellular Longevity
ControlNon-IBCIBC
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100Re
lativ
e mRN
A ex
pres
sion
Control Non-IBC IBC
P = 0036
Figure 2 Expression of GPX3mRNA is downregulated in IBC Blotrepresents the mean fold change of GPX3 mRNA measured by RT-PCR in normal breast tissues IBC and non-IBC carcinoma tissuesStatistical analysis revealed a significant increase in expression ofGPX3 mRNA in normal breast tissues compared to non-IBC andIBC carcinoma tissues (119875 = 0001 and 119875 lt 0001 resp) Incarcinoma tissues of non-IBC patients the level of expression ofGPX3 mRNA is significantly higher (119875 = 0036) than that of IBCResults are representative of at least three independent experimentsData are expressed as mean plusmn SD and 119875 value was determined byStudentrsquos 119905-test
carcinoma cell motility and invasion by activating proteinkinase-C (PKC) and the ERKMAPK signaling pathwaysthus increasing the risk of metastasis [26 27] Failure toremove exogenous and endogenous ROS may occur due todefect in the cellular antioxidant system of carcinoma cellsrepresented by the inhibition of the activity of antioxidantenzymes as GPXs familymembersThus drugs which induceexpression of antioxidant enzymes were suggested for cancertreatment [29] GPXs are categorized into two types onetype is selenium-dependent catalytic activity (GPX-1 -2 -3-4 and 6) and the second type is non-selenium-dependent(GPX-5 and -7) [6] Downregulation and genetic imbalanceamong GPXs were found to play a key role in breast cancerFor example GPX1 gene allelic variants and loss of heterozy-gosity (LOH) at 3p213p region contribute to breast cancerdevelopment [31] Low expression of GPX4 in breast-invasiveductal carcinoma correlated with high tumor grade and poorprognosis of breast cancer patients [32] Low expression ofGPX3 significantly correlates with high risk of breast cancerlocal recurrence among early-stage invasive breast cancerpatients regardless of patientsrsquo clinic-pathological criteria[33]
In fact GPX3 is an essential enzyme responsible for theremoval of ROS in healthy tissues On the contrary GPX3was found to be downregulated in carcinoma tissues of breastgastric and colorectal cancer patients [9] prostate cancer[10] thyroid cancer [11] and esophageal cancer [12] Theimpaired function of GPX3 would result in the accumulationof an increased amount of hydrogen peroxide and other ROSwhich may induce breast carcinogenesis via induction ofoxidative DNA damage genetic instability neoplastic trans-formation [34] and mutation of the p53 tumor suppressorgene [35] Epigenetic mechanisms such as DNA hypermethy-lation and histonemodificationmay repress the expression ofGPX3 For instance treatment of SKGT4 esophageal cancercell lines [29] and endometrial tumor cell lines (NUT12 andNUT81) [16] that show GPX3 promoter hypermethylationwith the demethylating agent 5-aza-21015840-deoxycytidine (5Aza-dC) and the histone deacetylace inhibitor trichostatin A(TSA) results in re-expression of GPX3 mRNAThese resultssuggest that DNA hypermethylation and histone deacetyla-tion may act together to regulate the expression of GPX3mRNA Paradoxically downregulation of GPX3 in absenceof promoter hypermethylation is associated with GPX3 genedeletion in endometrial tumor cell line (NUT84) [16] In factGPX3 promoter hypermethylation is linked to downregula-tion of GPX3 expression in different types of cancer cells andtreatment with 5-Aza to human esophageal adenocarcinomacancer cells SKGT4 [29] and human myeloma cells KMS11[19] restores GPX3 gene expression
Our previous studies demonstrated that GPX3was down-regulated in Barrettrsquos carcinoma due to hypermethylation ofthe promoter region [12] In addition we found that lossin DNA copy number hypermethylation of the promoterregion and downregulation of mRNA expression of GPX3are associated with lymph node metastasis in gastric car-cinomas Reactivation of GPX3 in gastric adenocarcinomacell line AGS inhibits cell motility as assessed by woundhealing assay [20] Herein we studied the role of GPX3 inbreast carcinogenesis We analyzed the level of expressionof GPX3 in normal breast tissues obtained from healthyvolunteers during mammoplasty and non-IBC and IBCbreast carcinoma tissues Our results revealed that GPX3protein level and mRNA were significantly expressed innormal breast tissues and downregulated in breast carcinomatissues The present results are consistent with other studieswhich proved that GPX3 is downregulated in carcinomatissues such as prostate [10] thyroid [11] and esophageal [12]When we compared carcinoma tissues of IBC with non-IBCsamples we detected a significant decrease in mRNA andprotein expression of GPX3 in IBC tissue samples versusnon-IBC tissue samples We did not detect any significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties The present resultsagree with other studies that showed downregulation ofGPX3 in endometrial adenocarcinoma [16] and early invasivebreast carcinoma [33] regardless of patientsrsquo clinical andpathological criteria
IBC is an aggressive phenotype characterized by highmetastatic potential disease recurrence and resistance tochemotherapy [36] Furthermore recently we found that IBC
Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

Oxidative Medicine and Cellular Longevity 3
rehydrated through graded concentrations of ethanol Forantigen retrieval slides were incubated in citrate buffer (pH60) in a water bath for 1 h at 99∘C Slides were cooledby incubation in Tris-buffered saline (TBS 005molL Tris-HCl pH 76 015molL NaCl and 005 Tween 20) for20min Endogenous peroxidase activity was blocked byusing Dako Dual Endogenous Enzyme Block for 10min Forimmunohistochemical stain of GPX3 tissue sections wereincubated for 1 h at room temperature with the primaryantibody mouse anti-GPX3 (mouse anti-GPX3 monoclonalantibody Clone 23B1 Abcam)Themonoclonal antibodywasdiluted 1 100 inDakoAntibodyDiluent-Reduce Backgroundproduct (DAKO) Detection was carried out by incubatingsections with 100 120583L of Horseradish Peroxidase (HRP) Rab-bitMouse (EnVision+ Dual Link System-HRP diaminoben-zidine (DAB+)) for 45min Staining was achieved by adding100 120583L of DAB+ diluted 1 50 in substrate buffer [EnVision+Dual Link System-HRP (DAB+)] Staining was progressedfor 15min until the development of brown color Nucleiwere counterstained with hematoxylin rinsed in PBS andmounted using Permount formicroscopic examination Neg-ative control slides were run in parallel with each markerwhere the primary antibody is replaced by PBS The level ofexpression of GPX3 protein was scored according to both theintensity and the positivity of the stain of the cells within theentire slide negative no immunostaining is observed score(+) less than 10 of cells showed no or weak staining score(++) 10ndash50 of cells showed moderate to strong stainingand score (+++) more than 50 of cells showed strongstaining [21]
25 DNA Extraction from Fresh Tissue and Identificationof CpG Islands in the Promoter Region of GPX3 GeneGenomic DNA was extracted from 25mg of fresh tissueof biopsy or modified radical mastectomy using DNeasytissue kit Qiagen In the last step of the protocol DNAwas eluted in 200 120583L elution buffer We used Universityof Californiarsquos (UCSC) Genome Browser website (httpgenomeucscedu) to obtain DNA sequences around thepromoter region Identified sequences were confirmed to beidentical to that from the DBTSS (database of transcriptionalstart sites httpdbtsshgcjp) as described [29] The CpGisland in the promoter region of GPX3 was defined usingCpG island searcher online tool (httpwwwuscnorriscomcpgislands2cpgaspx)
26 DNA Bisulfite Treatment and Methylation-Specific PCR(MSP) Bisulfite modification of the purified DNA wasachieved by using an EZ DNAmethylation-Gold kit (ZYMOResearch Orange CA USA) following the manufactoryrsquosinstructionsWe used 1120583g of extracted genomic DNA of eachsample from normal breast tissue samples IBC and non-IBC breast carcinoma tissue samples [12 29] The bisulfitetreatedDNAwas subjected toMSP in a final reaction of 50120583LWe designed primers for MSP targeting CpG-rich promoterregion using online software ldquoMethPrimerrdquo (httpwwwurogeneorgmethprimer)The forward and reverse primers
used for methylated GPX3 were 51015840-GTTGAGGGTAAG-TCGCGTTC-31015840 and 51015840-GTCCGTCTAAAATATCCGACG-31015840 and those for unmethylated GPX3 were 51015840-GAGTTG-AGGGTAAGTTGTGTTTGT-31015840 and 51015840-CCATCCATC-TAAAATATCCAACACT-31015840 PCR mixture included thePlatinumPCR SuperMixHigh Fidelity (Invitrogen CarlsbadCA USA) and the program was adjusted as follows initialdenaturation at 94∘C for 5min then 35 cycles consisting eachof 94∘C for 1min annealing at 54∘C for 1min and finallyextension at 72∘C for 1min PCR products were subjectedto electrophoresis on 15 agarose gel using GelRed nucleicacid stain (Biotium Hayward CA USA) and amplified DNAwas visualized by 300 nm transillumination For quantitativeanalysis visualized bands of the agarose gel were analyzedby ImageJ (National Institutes of Health Bethesda MAUSA) software For each carcinoma tissue specimen the bandintensity of the methylated and unmethylated MSP productswas quantified and normalized against gel background asdescribed elsewhere [30]
27 Statistical Analysis The data was analyzed using SPSSsoftware version 160 Differences were evaluated by Studentrsquos119905-test and Fisherrsquos exact test 119875 lt 005 was considered asstatistically significant
3 Results
31 Clinical and Pathological Features of IBC and Non-IBCPatients The clinical and pathological features of non-IBCand IBC patients are presented in Table 1 All of IBC patientswere premenopausal one decade younger than non-IBCpatients (119875 lt 0001) Statistical analysis revealed that IBCpatients showed a significant higher incidence (119875 lt 001)of positive metastatic lymph nodes compared to non-IBCpatients In addition a significant difference (119875 lt 0001)in lymphovascular invasion and dermal lymphatic emboliwas highly detected in IBC versus non-IBC patients tissuesections
32 Downregulation of the GPX3 Protein Expression in BreastCarcinoma Tissue Level of expression of GPX3 proteinwas assessed in normal and carcinoma breast tissues usingIHC GPX3 expression staining results were scored for thepositivity and intensity (Table 2) Statistical analysis using theChi-square test revealed that normal breast tissues expresssignificantly high level of GPX3 protein (Figure 1(a)) com-pared to breast carcinoma tissues of non-IBC (Figure 1(b))and IBC (Figure 1(c)) patients (119875 = 0001 and 119875 lt 0001resp) In addition non-IBC tissues showed a significantincrease (119875 = 0043) in the level of expression of GPX3 ascompared to those of IBC patients
33 Suppression of GPX3mRNAExpression in IBC CarcinomaTissue Samples Inhibition of GPX3 protein expression in thecarcinoma tissues of breast cancer patients was confirmedat mRNA level by using qRT-PCR We found that the levelof GPX3 mRNA expression in normal breast tissues wassignificantly higher than that in breast carcinoma tissue
4 Oxidative Medicine and Cellular Longevity
Table 1 Patientsrsquo clinical and pathological data
Characteristics Non-IBC119899 = 20 ()
IBC119899 = 20 () 119875 value
AgeMean plusmn SE 5435 plusmn 21 4055 plusmn 17
le50 7 (35) 18 (90)lt0001lowast
gt50 13 (65) 2 (10)Tumor sizelowastlowast
Mean plusmn SE 512 plusmn 05 708 plusmn 06
le2 cm 1 (5) 0gt005
gt2 cm 19 (95) 18 (90)Tumor grade
G2 15 (75) 14 (70)gt005
G3 5 (25) 6 (30)Lymph node statuslowastlowast
Positive 17 (85) 18 (90)gt005
Negative 3 (15) 0Number of lymph nodeslowastlowast
le6 16 (80) 6 (333)lt001lowast
gt6 4 (20) 12 (667)Lymphovascular invasion
Positive 2 (10) 14 (70)lt0001lowast
Negative 18 (90) 6 (30)Dermal lymphatic emboli
Positive 2 (10) 18 (90)lt0001lowast
Negative 18 (90) 2 (10)ER status
Positive 7 (35) 6 (30)gt005
Negative 13 (65) 14 (70)PR status
Positive 9 (45) 7 (35)gt005
Negative 11 (55) 13 (65)HER-2 status
Positive 2 (10) 4 (20)gt005
Negative 18 (90) 16 (80)lowastSignificant 119875 value calculated by Studentrsquos 119905-testlowastlowast119899 = 18 in IBC patients
Table 2 Scoring of GPX3 expression in normal non-IBC and IBC breast tissues
Score Normal (119899 = 6) Non-IBC (119899 = 16)a IBC (119899 = 16)bc
119899 () 119899 (asymp) 119899 (asymp)Negative 0 (0) 5 (312) 11 (687)+ 0 (0) 7 (438) 3 (187)++ 3 (50) 4 (25) 2 (125)+++ 3 (50) 0 0Significant 119875 value calculated by the Chi-square testaSignificant 119875 value (119875 = 0001) when normal tissues were compared to non-IBC tissuesbSignificant 119875 value (119875 lt 0001) when normal tissues were compared to IBC tissuescSignificant 119875 value (119875 = 0043) when non-IBC tissues were compared to IBC tissues119899 = number of patients
Oxidative Medicine and Cellular Longevity 5
(a) (b) (c)
Figure 1 GPX3 protein is downregulated in IBC Microscopic images representative of IHC stain of GPX3 (brown color) in (a) normalbreast tissues showing moderate to marked intensity of GPX3 (b) non-IBC tissue sections showing mild intensity of GPX3 and (c) IBCtissue sections with no immunostaining of GPX3 by carcinoma cells within tumor emboli (magnification upper panel 10x lower panel40x)
samples of non-IBC and IBC (119875 = 0001 and 119875 lt0001 resp) We compared the level of mRNA expressionof GPX3 in non-IBC and IBC and our results revealedthat mRNA expression level of GPX3 expression in non-IBC carcinoma tissues was statistically significant (119875 =0036) higher than that in IBC (Figure 2) Using the Pearsoncorrelation statistical analysis we did not detect significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties (tumor size tumorgrade and number of axillary metastatic lymph nodes) innon-IBC and IBC patients However downregulation ofGPX3 in IBC versus non-IBC carcinoma tissues suggests thatGPX3 may play a role in IBC disease progression
34 Methylation Profile of GPX3 Promoter Region in BreastCancer Tissue Samples Analysis of the promoter region ofGPX3 indicated that CpG islands were present within minus1000to +300 bp of the gene We assessed the GPX3 promoterregion methylation in 40 breast cancer tissue samples and 6normal breast tissue samples Agarose gel electrophoresis ofall normal breast tissues showed only one band correspond-ing to unmethylated GPX3-MSP products (Figure 3(a)) Onthe contrary agarose gel electrophoresis of all GPX3-MSPproducts breast carcinoma tissue samples showed bands of200 bp corresponding to both unmethylated (Figure 3(a))and methylated GPX3-MSP products (Figure 3(b)) Thepresent results showed that GPX3 methylation was detectedin breast carcinoma tissues and not in normal breast tissuesThus GPX3 promoter methylation is responsible for thedownregulation of GPX3 mRNA in breast carcinoma tissues
since it was not detected in normal breast tissues It shouldbe noted that detection of methylated and unmethylatedproducts in some breast carcinoma tissue homogenates maybe due to heterogeneous population of breast carcinoma cellsandor the presence of other normal stromal cells from breasttumor microenvironment
For quantitative analysis visualized bands of agarosegel were analyzed by ImageJ (National Institutes of HealthBethesda MA USA) software For each carcinoma tissuespecimen the band intensity of themethylated and unmethy-lated GPX3-MSP products was quantified and normalizedagainst gel background as described elsewhere [30] Intensityvalues of methylatedunmethylated (MU) ratio for GPX3-MSP products for non-IBC (119899 = 20) and IBC (119899 = 20)patients were analyzed by Studentrsquos 119905-test Normal breasttissues (N) with anMU ratio = 0 (no methylated bands weredetected from MSP) were assessed as unmethylated Resultsrevealed a significant increase (119875 = 004) in the MU ratiosof GPX3-MSP products in IBC versus non-IBC carcinomatissues (Figure 3(c)) Significant increase inMU ratio in IBCversus non-IBC suggests a potential role for GPX3 in IBCdisease progression
4 Discussion
Elevated levels of ROS detected in breast cancer were foundto play a crucial role in the disease progression [4] Forinstance carcinoma cells utilize ROS to stimulate cancercell proliferation motility invasion angiogenesis and escapeof apoptotic mechanism [25] In addition ROS augment
6 Oxidative Medicine and Cellular Longevity
ControlNon-IBCIBC
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100Re
lativ
e mRN
A ex
pres
sion
Control Non-IBC IBC
P = 0036
Figure 2 Expression of GPX3mRNA is downregulated in IBC Blotrepresents the mean fold change of GPX3 mRNA measured by RT-PCR in normal breast tissues IBC and non-IBC carcinoma tissuesStatistical analysis revealed a significant increase in expression ofGPX3 mRNA in normal breast tissues compared to non-IBC andIBC carcinoma tissues (119875 = 0001 and 119875 lt 0001 resp) Incarcinoma tissues of non-IBC patients the level of expression ofGPX3 mRNA is significantly higher (119875 = 0036) than that of IBCResults are representative of at least three independent experimentsData are expressed as mean plusmn SD and 119875 value was determined byStudentrsquos 119905-test
carcinoma cell motility and invasion by activating proteinkinase-C (PKC) and the ERKMAPK signaling pathwaysthus increasing the risk of metastasis [26 27] Failure toremove exogenous and endogenous ROS may occur due todefect in the cellular antioxidant system of carcinoma cellsrepresented by the inhibition of the activity of antioxidantenzymes as GPXs familymembersThus drugs which induceexpression of antioxidant enzymes were suggested for cancertreatment [29] GPXs are categorized into two types onetype is selenium-dependent catalytic activity (GPX-1 -2 -3-4 and 6) and the second type is non-selenium-dependent(GPX-5 and -7) [6] Downregulation and genetic imbalanceamong GPXs were found to play a key role in breast cancerFor example GPX1 gene allelic variants and loss of heterozy-gosity (LOH) at 3p213p region contribute to breast cancerdevelopment [31] Low expression of GPX4 in breast-invasiveductal carcinoma correlated with high tumor grade and poorprognosis of breast cancer patients [32] Low expression ofGPX3 significantly correlates with high risk of breast cancerlocal recurrence among early-stage invasive breast cancerpatients regardless of patientsrsquo clinic-pathological criteria[33]
In fact GPX3 is an essential enzyme responsible for theremoval of ROS in healthy tissues On the contrary GPX3was found to be downregulated in carcinoma tissues of breastgastric and colorectal cancer patients [9] prostate cancer[10] thyroid cancer [11] and esophageal cancer [12] Theimpaired function of GPX3 would result in the accumulationof an increased amount of hydrogen peroxide and other ROSwhich may induce breast carcinogenesis via induction ofoxidative DNA damage genetic instability neoplastic trans-formation [34] and mutation of the p53 tumor suppressorgene [35] Epigenetic mechanisms such as DNA hypermethy-lation and histonemodificationmay repress the expression ofGPX3 For instance treatment of SKGT4 esophageal cancercell lines [29] and endometrial tumor cell lines (NUT12 andNUT81) [16] that show GPX3 promoter hypermethylationwith the demethylating agent 5-aza-21015840-deoxycytidine (5Aza-dC) and the histone deacetylace inhibitor trichostatin A(TSA) results in re-expression of GPX3 mRNAThese resultssuggest that DNA hypermethylation and histone deacetyla-tion may act together to regulate the expression of GPX3mRNA Paradoxically downregulation of GPX3 in absenceof promoter hypermethylation is associated with GPX3 genedeletion in endometrial tumor cell line (NUT84) [16] In factGPX3 promoter hypermethylation is linked to downregula-tion of GPX3 expression in different types of cancer cells andtreatment with 5-Aza to human esophageal adenocarcinomacancer cells SKGT4 [29] and human myeloma cells KMS11[19] restores GPX3 gene expression
Our previous studies demonstrated that GPX3was down-regulated in Barrettrsquos carcinoma due to hypermethylation ofthe promoter region [12] In addition we found that lossin DNA copy number hypermethylation of the promoterregion and downregulation of mRNA expression of GPX3are associated with lymph node metastasis in gastric car-cinomas Reactivation of GPX3 in gastric adenocarcinomacell line AGS inhibits cell motility as assessed by woundhealing assay [20] Herein we studied the role of GPX3 inbreast carcinogenesis We analyzed the level of expressionof GPX3 in normal breast tissues obtained from healthyvolunteers during mammoplasty and non-IBC and IBCbreast carcinoma tissues Our results revealed that GPX3protein level and mRNA were significantly expressed innormal breast tissues and downregulated in breast carcinomatissues The present results are consistent with other studieswhich proved that GPX3 is downregulated in carcinomatissues such as prostate [10] thyroid [11] and esophageal [12]When we compared carcinoma tissues of IBC with non-IBCsamples we detected a significant decrease in mRNA andprotein expression of GPX3 in IBC tissue samples versusnon-IBC tissue samples We did not detect any significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties The present resultsagree with other studies that showed downregulation ofGPX3 in endometrial adenocarcinoma [16] and early invasivebreast carcinoma [33] regardless of patientsrsquo clinical andpathological criteria
IBC is an aggressive phenotype characterized by highmetastatic potential disease recurrence and resistance tochemotherapy [36] Furthermore recently we found that IBC
Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013
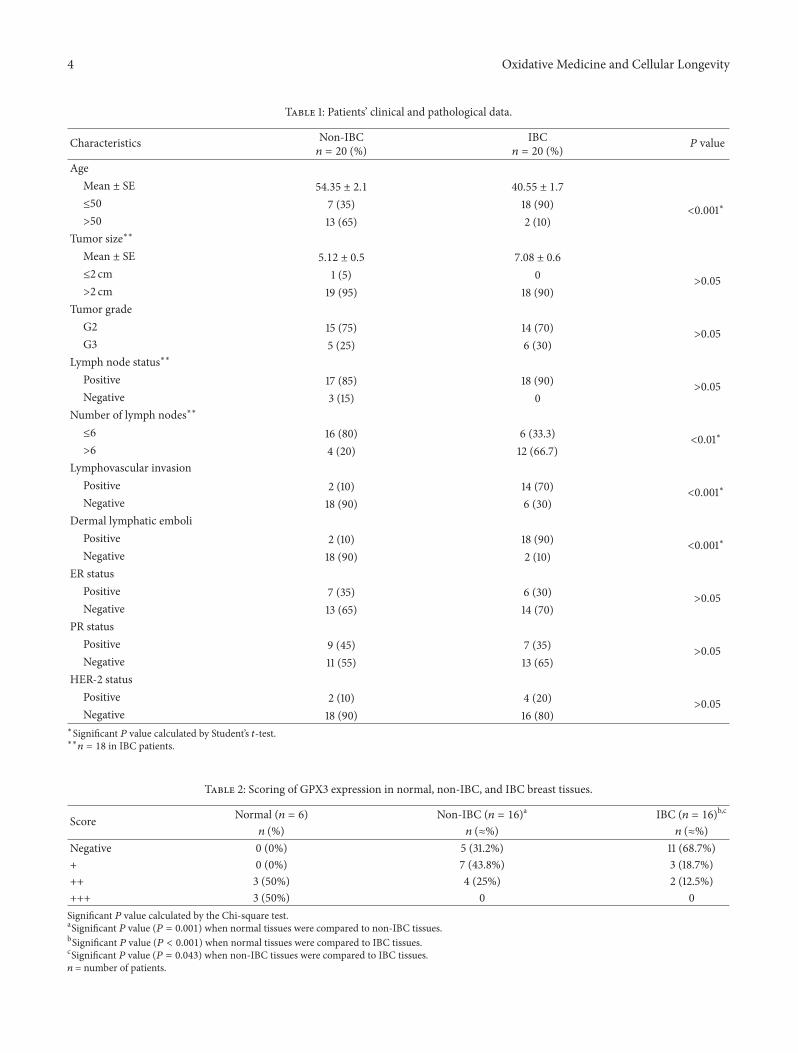
4 Oxidative Medicine and Cellular Longevity
Table 1 Patientsrsquo clinical and pathological data
Characteristics Non-IBC119899 = 20 ()
IBC119899 = 20 () 119875 value
AgeMean plusmn SE 5435 plusmn 21 4055 plusmn 17
le50 7 (35) 18 (90)lt0001lowast
gt50 13 (65) 2 (10)Tumor sizelowastlowast
Mean plusmn SE 512 plusmn 05 708 plusmn 06
le2 cm 1 (5) 0gt005
gt2 cm 19 (95) 18 (90)Tumor grade
G2 15 (75) 14 (70)gt005
G3 5 (25) 6 (30)Lymph node statuslowastlowast
Positive 17 (85) 18 (90)gt005
Negative 3 (15) 0Number of lymph nodeslowastlowast
le6 16 (80) 6 (333)lt001lowast
gt6 4 (20) 12 (667)Lymphovascular invasion
Positive 2 (10) 14 (70)lt0001lowast
Negative 18 (90) 6 (30)Dermal lymphatic emboli
Positive 2 (10) 18 (90)lt0001lowast
Negative 18 (90) 2 (10)ER status
Positive 7 (35) 6 (30)gt005
Negative 13 (65) 14 (70)PR status
Positive 9 (45) 7 (35)gt005
Negative 11 (55) 13 (65)HER-2 status
Positive 2 (10) 4 (20)gt005
Negative 18 (90) 16 (80)lowastSignificant 119875 value calculated by Studentrsquos 119905-testlowastlowast119899 = 18 in IBC patients
Table 2 Scoring of GPX3 expression in normal non-IBC and IBC breast tissues
Score Normal (119899 = 6) Non-IBC (119899 = 16)a IBC (119899 = 16)bc
119899 () 119899 (asymp) 119899 (asymp)Negative 0 (0) 5 (312) 11 (687)+ 0 (0) 7 (438) 3 (187)++ 3 (50) 4 (25) 2 (125)+++ 3 (50) 0 0Significant 119875 value calculated by the Chi-square testaSignificant 119875 value (119875 = 0001) when normal tissues were compared to non-IBC tissuesbSignificant 119875 value (119875 lt 0001) when normal tissues were compared to IBC tissuescSignificant 119875 value (119875 = 0043) when non-IBC tissues were compared to IBC tissues119899 = number of patients
Oxidative Medicine and Cellular Longevity 5
(a) (b) (c)
Figure 1 GPX3 protein is downregulated in IBC Microscopic images representative of IHC stain of GPX3 (brown color) in (a) normalbreast tissues showing moderate to marked intensity of GPX3 (b) non-IBC tissue sections showing mild intensity of GPX3 and (c) IBCtissue sections with no immunostaining of GPX3 by carcinoma cells within tumor emboli (magnification upper panel 10x lower panel40x)
samples of non-IBC and IBC (119875 = 0001 and 119875 lt0001 resp) We compared the level of mRNA expressionof GPX3 in non-IBC and IBC and our results revealedthat mRNA expression level of GPX3 expression in non-IBC carcinoma tissues was statistically significant (119875 =0036) higher than that in IBC (Figure 2) Using the Pearsoncorrelation statistical analysis we did not detect significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties (tumor size tumorgrade and number of axillary metastatic lymph nodes) innon-IBC and IBC patients However downregulation ofGPX3 in IBC versus non-IBC carcinoma tissues suggests thatGPX3 may play a role in IBC disease progression
34 Methylation Profile of GPX3 Promoter Region in BreastCancer Tissue Samples Analysis of the promoter region ofGPX3 indicated that CpG islands were present within minus1000to +300 bp of the gene We assessed the GPX3 promoterregion methylation in 40 breast cancer tissue samples and 6normal breast tissue samples Agarose gel electrophoresis ofall normal breast tissues showed only one band correspond-ing to unmethylated GPX3-MSP products (Figure 3(a)) Onthe contrary agarose gel electrophoresis of all GPX3-MSPproducts breast carcinoma tissue samples showed bands of200 bp corresponding to both unmethylated (Figure 3(a))and methylated GPX3-MSP products (Figure 3(b)) Thepresent results showed that GPX3 methylation was detectedin breast carcinoma tissues and not in normal breast tissuesThus GPX3 promoter methylation is responsible for thedownregulation of GPX3 mRNA in breast carcinoma tissues
since it was not detected in normal breast tissues It shouldbe noted that detection of methylated and unmethylatedproducts in some breast carcinoma tissue homogenates maybe due to heterogeneous population of breast carcinoma cellsandor the presence of other normal stromal cells from breasttumor microenvironment
For quantitative analysis visualized bands of agarosegel were analyzed by ImageJ (National Institutes of HealthBethesda MA USA) software For each carcinoma tissuespecimen the band intensity of themethylated and unmethy-lated GPX3-MSP products was quantified and normalizedagainst gel background as described elsewhere [30] Intensityvalues of methylatedunmethylated (MU) ratio for GPX3-MSP products for non-IBC (119899 = 20) and IBC (119899 = 20)patients were analyzed by Studentrsquos 119905-test Normal breasttissues (N) with anMU ratio = 0 (no methylated bands weredetected from MSP) were assessed as unmethylated Resultsrevealed a significant increase (119875 = 004) in the MU ratiosof GPX3-MSP products in IBC versus non-IBC carcinomatissues (Figure 3(c)) Significant increase inMU ratio in IBCversus non-IBC suggests a potential role for GPX3 in IBCdisease progression
4 Discussion
Elevated levels of ROS detected in breast cancer were foundto play a crucial role in the disease progression [4] Forinstance carcinoma cells utilize ROS to stimulate cancercell proliferation motility invasion angiogenesis and escapeof apoptotic mechanism [25] In addition ROS augment
6 Oxidative Medicine and Cellular Longevity
ControlNon-IBCIBC
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100Re
lativ
e mRN
A ex
pres
sion
Control Non-IBC IBC
P = 0036
Figure 2 Expression of GPX3mRNA is downregulated in IBC Blotrepresents the mean fold change of GPX3 mRNA measured by RT-PCR in normal breast tissues IBC and non-IBC carcinoma tissuesStatistical analysis revealed a significant increase in expression ofGPX3 mRNA in normal breast tissues compared to non-IBC andIBC carcinoma tissues (119875 = 0001 and 119875 lt 0001 resp) Incarcinoma tissues of non-IBC patients the level of expression ofGPX3 mRNA is significantly higher (119875 = 0036) than that of IBCResults are representative of at least three independent experimentsData are expressed as mean plusmn SD and 119875 value was determined byStudentrsquos 119905-test
carcinoma cell motility and invasion by activating proteinkinase-C (PKC) and the ERKMAPK signaling pathwaysthus increasing the risk of metastasis [26 27] Failure toremove exogenous and endogenous ROS may occur due todefect in the cellular antioxidant system of carcinoma cellsrepresented by the inhibition of the activity of antioxidantenzymes as GPXs familymembersThus drugs which induceexpression of antioxidant enzymes were suggested for cancertreatment [29] GPXs are categorized into two types onetype is selenium-dependent catalytic activity (GPX-1 -2 -3-4 and 6) and the second type is non-selenium-dependent(GPX-5 and -7) [6] Downregulation and genetic imbalanceamong GPXs were found to play a key role in breast cancerFor example GPX1 gene allelic variants and loss of heterozy-gosity (LOH) at 3p213p region contribute to breast cancerdevelopment [31] Low expression of GPX4 in breast-invasiveductal carcinoma correlated with high tumor grade and poorprognosis of breast cancer patients [32] Low expression ofGPX3 significantly correlates with high risk of breast cancerlocal recurrence among early-stage invasive breast cancerpatients regardless of patientsrsquo clinic-pathological criteria[33]
In fact GPX3 is an essential enzyme responsible for theremoval of ROS in healthy tissues On the contrary GPX3was found to be downregulated in carcinoma tissues of breastgastric and colorectal cancer patients [9] prostate cancer[10] thyroid cancer [11] and esophageal cancer [12] Theimpaired function of GPX3 would result in the accumulationof an increased amount of hydrogen peroxide and other ROSwhich may induce breast carcinogenesis via induction ofoxidative DNA damage genetic instability neoplastic trans-formation [34] and mutation of the p53 tumor suppressorgene [35] Epigenetic mechanisms such as DNA hypermethy-lation and histonemodificationmay repress the expression ofGPX3 For instance treatment of SKGT4 esophageal cancercell lines [29] and endometrial tumor cell lines (NUT12 andNUT81) [16] that show GPX3 promoter hypermethylationwith the demethylating agent 5-aza-21015840-deoxycytidine (5Aza-dC) and the histone deacetylace inhibitor trichostatin A(TSA) results in re-expression of GPX3 mRNAThese resultssuggest that DNA hypermethylation and histone deacetyla-tion may act together to regulate the expression of GPX3mRNA Paradoxically downregulation of GPX3 in absenceof promoter hypermethylation is associated with GPX3 genedeletion in endometrial tumor cell line (NUT84) [16] In factGPX3 promoter hypermethylation is linked to downregula-tion of GPX3 expression in different types of cancer cells andtreatment with 5-Aza to human esophageal adenocarcinomacancer cells SKGT4 [29] and human myeloma cells KMS11[19] restores GPX3 gene expression
Our previous studies demonstrated that GPX3was down-regulated in Barrettrsquos carcinoma due to hypermethylation ofthe promoter region [12] In addition we found that lossin DNA copy number hypermethylation of the promoterregion and downregulation of mRNA expression of GPX3are associated with lymph node metastasis in gastric car-cinomas Reactivation of GPX3 in gastric adenocarcinomacell line AGS inhibits cell motility as assessed by woundhealing assay [20] Herein we studied the role of GPX3 inbreast carcinogenesis We analyzed the level of expressionof GPX3 in normal breast tissues obtained from healthyvolunteers during mammoplasty and non-IBC and IBCbreast carcinoma tissues Our results revealed that GPX3protein level and mRNA were significantly expressed innormal breast tissues and downregulated in breast carcinomatissues The present results are consistent with other studieswhich proved that GPX3 is downregulated in carcinomatissues such as prostate [10] thyroid [11] and esophageal [12]When we compared carcinoma tissues of IBC with non-IBCsamples we detected a significant decrease in mRNA andprotein expression of GPX3 in IBC tissue samples versusnon-IBC tissue samples We did not detect any significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties The present resultsagree with other studies that showed downregulation ofGPX3 in endometrial adenocarcinoma [16] and early invasivebreast carcinoma [33] regardless of patientsrsquo clinical andpathological criteria
IBC is an aggressive phenotype characterized by highmetastatic potential disease recurrence and resistance tochemotherapy [36] Furthermore recently we found that IBC
Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

Oxidative Medicine and Cellular Longevity 5
(a) (b) (c)
Figure 1 GPX3 protein is downregulated in IBC Microscopic images representative of IHC stain of GPX3 (brown color) in (a) normalbreast tissues showing moderate to marked intensity of GPX3 (b) non-IBC tissue sections showing mild intensity of GPX3 and (c) IBCtissue sections with no immunostaining of GPX3 by carcinoma cells within tumor emboli (magnification upper panel 10x lower panel40x)
samples of non-IBC and IBC (119875 = 0001 and 119875 lt0001 resp) We compared the level of mRNA expressionof GPX3 in non-IBC and IBC and our results revealedthat mRNA expression level of GPX3 expression in non-IBC carcinoma tissues was statistically significant (119875 =0036) higher than that in IBC (Figure 2) Using the Pearsoncorrelation statistical analysis we did not detect significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties (tumor size tumorgrade and number of axillary metastatic lymph nodes) innon-IBC and IBC patients However downregulation ofGPX3 in IBC versus non-IBC carcinoma tissues suggests thatGPX3 may play a role in IBC disease progression
34 Methylation Profile of GPX3 Promoter Region in BreastCancer Tissue Samples Analysis of the promoter region ofGPX3 indicated that CpG islands were present within minus1000to +300 bp of the gene We assessed the GPX3 promoterregion methylation in 40 breast cancer tissue samples and 6normal breast tissue samples Agarose gel electrophoresis ofall normal breast tissues showed only one band correspond-ing to unmethylated GPX3-MSP products (Figure 3(a)) Onthe contrary agarose gel electrophoresis of all GPX3-MSPproducts breast carcinoma tissue samples showed bands of200 bp corresponding to both unmethylated (Figure 3(a))and methylated GPX3-MSP products (Figure 3(b)) Thepresent results showed that GPX3 methylation was detectedin breast carcinoma tissues and not in normal breast tissuesThus GPX3 promoter methylation is responsible for thedownregulation of GPX3 mRNA in breast carcinoma tissues
since it was not detected in normal breast tissues It shouldbe noted that detection of methylated and unmethylatedproducts in some breast carcinoma tissue homogenates maybe due to heterogeneous population of breast carcinoma cellsandor the presence of other normal stromal cells from breasttumor microenvironment
For quantitative analysis visualized bands of agarosegel were analyzed by ImageJ (National Institutes of HealthBethesda MA USA) software For each carcinoma tissuespecimen the band intensity of themethylated and unmethy-lated GPX3-MSP products was quantified and normalizedagainst gel background as described elsewhere [30] Intensityvalues of methylatedunmethylated (MU) ratio for GPX3-MSP products for non-IBC (119899 = 20) and IBC (119899 = 20)patients were analyzed by Studentrsquos 119905-test Normal breasttissues (N) with anMU ratio = 0 (no methylated bands weredetected from MSP) were assessed as unmethylated Resultsrevealed a significant increase (119875 = 004) in the MU ratiosof GPX3-MSP products in IBC versus non-IBC carcinomatissues (Figure 3(c)) Significant increase inMU ratio in IBCversus non-IBC suggests a potential role for GPX3 in IBCdisease progression
4 Discussion
Elevated levels of ROS detected in breast cancer were foundto play a crucial role in the disease progression [4] Forinstance carcinoma cells utilize ROS to stimulate cancercell proliferation motility invasion angiogenesis and escapeof apoptotic mechanism [25] In addition ROS augment
6 Oxidative Medicine and Cellular Longevity
ControlNon-IBCIBC
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100Re
lativ
e mRN
A ex
pres
sion
Control Non-IBC IBC
P = 0036
Figure 2 Expression of GPX3mRNA is downregulated in IBC Blotrepresents the mean fold change of GPX3 mRNA measured by RT-PCR in normal breast tissues IBC and non-IBC carcinoma tissuesStatistical analysis revealed a significant increase in expression ofGPX3 mRNA in normal breast tissues compared to non-IBC andIBC carcinoma tissues (119875 = 0001 and 119875 lt 0001 resp) Incarcinoma tissues of non-IBC patients the level of expression ofGPX3 mRNA is significantly higher (119875 = 0036) than that of IBCResults are representative of at least three independent experimentsData are expressed as mean plusmn SD and 119875 value was determined byStudentrsquos 119905-test
carcinoma cell motility and invasion by activating proteinkinase-C (PKC) and the ERKMAPK signaling pathwaysthus increasing the risk of metastasis [26 27] Failure toremove exogenous and endogenous ROS may occur due todefect in the cellular antioxidant system of carcinoma cellsrepresented by the inhibition of the activity of antioxidantenzymes as GPXs familymembersThus drugs which induceexpression of antioxidant enzymes were suggested for cancertreatment [29] GPXs are categorized into two types onetype is selenium-dependent catalytic activity (GPX-1 -2 -3-4 and 6) and the second type is non-selenium-dependent(GPX-5 and -7) [6] Downregulation and genetic imbalanceamong GPXs were found to play a key role in breast cancerFor example GPX1 gene allelic variants and loss of heterozy-gosity (LOH) at 3p213p region contribute to breast cancerdevelopment [31] Low expression of GPX4 in breast-invasiveductal carcinoma correlated with high tumor grade and poorprognosis of breast cancer patients [32] Low expression ofGPX3 significantly correlates with high risk of breast cancerlocal recurrence among early-stage invasive breast cancerpatients regardless of patientsrsquo clinic-pathological criteria[33]
In fact GPX3 is an essential enzyme responsible for theremoval of ROS in healthy tissues On the contrary GPX3was found to be downregulated in carcinoma tissues of breastgastric and colorectal cancer patients [9] prostate cancer[10] thyroid cancer [11] and esophageal cancer [12] Theimpaired function of GPX3 would result in the accumulationof an increased amount of hydrogen peroxide and other ROSwhich may induce breast carcinogenesis via induction ofoxidative DNA damage genetic instability neoplastic trans-formation [34] and mutation of the p53 tumor suppressorgene [35] Epigenetic mechanisms such as DNA hypermethy-lation and histonemodificationmay repress the expression ofGPX3 For instance treatment of SKGT4 esophageal cancercell lines [29] and endometrial tumor cell lines (NUT12 andNUT81) [16] that show GPX3 promoter hypermethylationwith the demethylating agent 5-aza-21015840-deoxycytidine (5Aza-dC) and the histone deacetylace inhibitor trichostatin A(TSA) results in re-expression of GPX3 mRNAThese resultssuggest that DNA hypermethylation and histone deacetyla-tion may act together to regulate the expression of GPX3mRNA Paradoxically downregulation of GPX3 in absenceof promoter hypermethylation is associated with GPX3 genedeletion in endometrial tumor cell line (NUT84) [16] In factGPX3 promoter hypermethylation is linked to downregula-tion of GPX3 expression in different types of cancer cells andtreatment with 5-Aza to human esophageal adenocarcinomacancer cells SKGT4 [29] and human myeloma cells KMS11[19] restores GPX3 gene expression
Our previous studies demonstrated that GPX3was down-regulated in Barrettrsquos carcinoma due to hypermethylation ofthe promoter region [12] In addition we found that lossin DNA copy number hypermethylation of the promoterregion and downregulation of mRNA expression of GPX3are associated with lymph node metastasis in gastric car-cinomas Reactivation of GPX3 in gastric adenocarcinomacell line AGS inhibits cell motility as assessed by woundhealing assay [20] Herein we studied the role of GPX3 inbreast carcinogenesis We analyzed the level of expressionof GPX3 in normal breast tissues obtained from healthyvolunteers during mammoplasty and non-IBC and IBCbreast carcinoma tissues Our results revealed that GPX3protein level and mRNA were significantly expressed innormal breast tissues and downregulated in breast carcinomatissues The present results are consistent with other studieswhich proved that GPX3 is downregulated in carcinomatissues such as prostate [10] thyroid [11] and esophageal [12]When we compared carcinoma tissues of IBC with non-IBCsamples we detected a significant decrease in mRNA andprotein expression of GPX3 in IBC tissue samples versusnon-IBC tissue samples We did not detect any significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties The present resultsagree with other studies that showed downregulation ofGPX3 in endometrial adenocarcinoma [16] and early invasivebreast carcinoma [33] regardless of patientsrsquo clinical andpathological criteria
IBC is an aggressive phenotype characterized by highmetastatic potential disease recurrence and resistance tochemotherapy [36] Furthermore recently we found that IBC
Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

6 Oxidative Medicine and Cellular Longevity
ControlNon-IBCIBC
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100Re
lativ
e mRN
A ex
pres
sion
Control Non-IBC IBC
P = 0036
Figure 2 Expression of GPX3mRNA is downregulated in IBC Blotrepresents the mean fold change of GPX3 mRNA measured by RT-PCR in normal breast tissues IBC and non-IBC carcinoma tissuesStatistical analysis revealed a significant increase in expression ofGPX3 mRNA in normal breast tissues compared to non-IBC andIBC carcinoma tissues (119875 = 0001 and 119875 lt 0001 resp) Incarcinoma tissues of non-IBC patients the level of expression ofGPX3 mRNA is significantly higher (119875 = 0036) than that of IBCResults are representative of at least three independent experimentsData are expressed as mean plusmn SD and 119875 value was determined byStudentrsquos 119905-test
carcinoma cell motility and invasion by activating proteinkinase-C (PKC) and the ERKMAPK signaling pathwaysthus increasing the risk of metastasis [26 27] Failure toremove exogenous and endogenous ROS may occur due todefect in the cellular antioxidant system of carcinoma cellsrepresented by the inhibition of the activity of antioxidantenzymes as GPXs familymembersThus drugs which induceexpression of antioxidant enzymes were suggested for cancertreatment [29] GPXs are categorized into two types onetype is selenium-dependent catalytic activity (GPX-1 -2 -3-4 and 6) and the second type is non-selenium-dependent(GPX-5 and -7) [6] Downregulation and genetic imbalanceamong GPXs were found to play a key role in breast cancerFor example GPX1 gene allelic variants and loss of heterozy-gosity (LOH) at 3p213p region contribute to breast cancerdevelopment [31] Low expression of GPX4 in breast-invasiveductal carcinoma correlated with high tumor grade and poorprognosis of breast cancer patients [32] Low expression ofGPX3 significantly correlates with high risk of breast cancerlocal recurrence among early-stage invasive breast cancerpatients regardless of patientsrsquo clinic-pathological criteria[33]
In fact GPX3 is an essential enzyme responsible for theremoval of ROS in healthy tissues On the contrary GPX3was found to be downregulated in carcinoma tissues of breastgastric and colorectal cancer patients [9] prostate cancer[10] thyroid cancer [11] and esophageal cancer [12] Theimpaired function of GPX3 would result in the accumulationof an increased amount of hydrogen peroxide and other ROSwhich may induce breast carcinogenesis via induction ofoxidative DNA damage genetic instability neoplastic trans-formation [34] and mutation of the p53 tumor suppressorgene [35] Epigenetic mechanisms such as DNA hypermethy-lation and histonemodificationmay repress the expression ofGPX3 For instance treatment of SKGT4 esophageal cancercell lines [29] and endometrial tumor cell lines (NUT12 andNUT81) [16] that show GPX3 promoter hypermethylationwith the demethylating agent 5-aza-21015840-deoxycytidine (5Aza-dC) and the histone deacetylace inhibitor trichostatin A(TSA) results in re-expression of GPX3 mRNAThese resultssuggest that DNA hypermethylation and histone deacetyla-tion may act together to regulate the expression of GPX3mRNA Paradoxically downregulation of GPX3 in absenceof promoter hypermethylation is associated with GPX3 genedeletion in endometrial tumor cell line (NUT84) [16] In factGPX3 promoter hypermethylation is linked to downregula-tion of GPX3 expression in different types of cancer cells andtreatment with 5-Aza to human esophageal adenocarcinomacancer cells SKGT4 [29] and human myeloma cells KMS11[19] restores GPX3 gene expression
Our previous studies demonstrated that GPX3was down-regulated in Barrettrsquos carcinoma due to hypermethylation ofthe promoter region [12] In addition we found that lossin DNA copy number hypermethylation of the promoterregion and downregulation of mRNA expression of GPX3are associated with lymph node metastasis in gastric car-cinomas Reactivation of GPX3 in gastric adenocarcinomacell line AGS inhibits cell motility as assessed by woundhealing assay [20] Herein we studied the role of GPX3 inbreast carcinogenesis We analyzed the level of expressionof GPX3 in normal breast tissues obtained from healthyvolunteers during mammoplasty and non-IBC and IBCbreast carcinoma tissues Our results revealed that GPX3protein level and mRNA were significantly expressed innormal breast tissues and downregulated in breast carcinomatissues The present results are consistent with other studieswhich proved that GPX3 is downregulated in carcinomatissues such as prostate [10] thyroid [11] and esophageal [12]When we compared carcinoma tissues of IBC with non-IBCsamples we detected a significant decrease in mRNA andprotein expression of GPX3 in IBC tissue samples versusnon-IBC tissue samples We did not detect any significantcorrelation between downregulation of GPX3 mRNA andpatients clinical-pathological properties The present resultsagree with other studies that showed downregulation ofGPX3 in endometrial adenocarcinoma [16] and early invasivebreast carcinoma [33] regardless of patientsrsquo clinical andpathological criteria
IBC is an aggressive phenotype characterized by highmetastatic potential disease recurrence and resistance tochemotherapy [36] Furthermore recently we found that IBC
Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

Oxidative Medicine and Cellular Longevity 7
Non-IBC IBC
400bp
300bp
200 bp
100 bp
M N1 N2 1 2 3 4 5 6 7 8
Non-IBC IBC
M
(a)
(b)
(c)
N1 N2 1 2 3 4 5 6 7 8
0
02
04
06
08
1
12
14
Normal Non-IBC IBC
NormalNon-IBCIBC
lowast
Ratio
of b
and
inte
nsiti
es
Figure 3 Gel electrophoresis of GPX3 methylation-specific PCR (GPX3-MSP) products Representative results of MSP using (a)unmethylated primers and (b) methylated primers M is the DNA marker lanes N1 and N2 represent normal breast tissues lanes 1ndash4 represent non-IBC carcinoma tissues and lanes 4ndash8 represent IBC breast carcinoma tissues (c) Bars represent intensity values ofmethylatedunmethylated (MU) ratios as quantified by ImageJ software Normal breast tissues (119899 = 6) with anMU ratio = 0 (nomethylatedbands were detected from MSP) were recognized as unmethylated We detected a significant increase (119875 = 004) in the MU ratio in IBC(119899 = 20) carcinoma tissues compared to non-IBC (119899 = 20) carcinoma tissues
8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

8 Oxidative Medicine and Cellular Longevity
carcinoma tissues are characterized by high infiltration oftumor associated macrophages that enhance carcinoma cellsinvasion and motility [37] Interestingly loss of GPX3 con-tributes to high infiltration of tumor associated macrophagesthat support tumor survival in GPX3 knockout mice model[38] Thus inhibition of GPX3 in IBC carcinoma tissuesmay be associated with the high infiltration of macrophagesIn addition quantitative analysis of the band intensities ofthe corresponding methylatedunmethylated MSP productsrevealed a significant increase in GPX3 promoter hyperme-thylation in IBC carcinoma tissues versus non-IBC carci-noma tissues We assumed that GPX3 may contribute to IBCmolecular phenotype
5 Conclusion
Our results suggest that epigenetic regulation of GPX3occurred widely in breast cancer tissues compared to normalbreast tissues and this may be due to GPX3 promoterhypermethylation in breast cancer cells and not in normalbreast tissues In addition methylation silencing of GPX3in IBC may contribute to invasion of IBC carcinoma cellsinto lymphatic vessels formation of tumor emboli andIBC chemoresistance as suggested in other cancers [14 19]Further studies to validate the role of GPX3 as a prognosticmaker in IBC and identify the mechanisms by which GPX3is involved in IBC carcinogenesis are essential
Conflict of Interests
The authors declare that they have no competing interests
Authorsrsquo Contribution
Mona M Mohamed and Salwa Sabet contributed equally tothis work
Acknowledgments
This work was supported by Science and Technology Devel-opment Funds (STDF) Egypt Grant no 408 (Wael El-RifaiandMonaMMohamed) and Avon Foundation USA (MonaMMohamed) Awards nos 02-2007-049 and 02-2009-085b
References
[1] H J Sipe Jr S J Jordan P M Hanna and R P Mason ldquoThemetabolism of 17120573-estradiol lactoperoxidase a possible sourceof oxidative stress in breast cancerrdquo Carcinogenesis vol 15 no11 pp 2637ndash2643 1994
[2] K Plak AM Czarnecka T Krawczyk P Golik and E BartnikldquoBreast cancer as a mitochondrial disorder (review)rdquo OncologyReports vol 21 no 4 pp 845ndash851 2009
[3] AM Fulton andY C Chong ldquoThe role ofmacrophage-derivedTNFa in the induction of sublethal tumor cell DNA damagerdquoCarcinogenesis vol 13 no 1 pp 77ndash81 1992
[4] N S Brown and R Bicknell ldquoOxidative stress its effects on thegrowth metastatic potential and response to therapy of breastcancer Oxidative stress its effects on the growth metastatic
potential and response to therapy of breast cancerrdquo BreastCancer Research vol 3 no 5 pp 323ndash327 2001
[5] D-H Kang ldquoOxidative stress DNAdamage and breast cancerrdquoAACN clinical issues vol 13 no 4 pp 540ndash549 2002
[6] R Brigelius-Flohe and A Kipp ldquoGlutathione peroxidases indifferent stages of carcinogenesisrdquo Biochimica et BiophysicaActa vol 1790 no 11 pp 1555ndash1568 2009
[7] M P Rayman ldquoSelenium in cancer prevention a review of theevidence andmechanism of actionrdquo Proceedings of the NutritionSociety vol 64 no 4 pp 527ndash542 2005
[8] J-Y Wang I-P Yang D-C Wu S-W Huang J-Y Wu and S-HH Juo ldquoFunctional glutathione peroxidase 3 polymorphismsassociated with increased risk of Taiwanese patients with gastriccancerrdquo Clinica Chimica Acta vol 411 no 19-20 pp 1432ndash14362010
[9] Z Pawlowicz B A Zachara U Trafikowska A Maciag EMarchaluk and A Nowicki ldquoBlood selenium concentrationsand glutathione peroxidase activities in patients with breastcancer and with advanced gastrointestinal cancerrdquo Journal ofTrace Elements and Electrolytes in Health and Disease vol 5 no4 pp 275ndash277 1991
[10] Y P YuG YuG Tseng et al ldquoGlutathione peroxidase 3 deletedor methylated in prostate cancer suppresses prostate cancergrowth and metastasisrdquo Cancer Research vol 67 no 17 pp8043ndash8050 2007
[11] Y Hasegawa T Takano A Miyauchi et al ldquoDecreased expres-sion of glutathione peroxidase mRNA in thyroid anaplasticcarcinomardquo Cancer Letters vol 182 no 1 pp 69ndash74 2002
[12] O-J Lee R Schneider-Stock P A McChesney et al ldquoHyper-methylation and loss of expression of glutathione peroxidase-3in Barrettrsquos tumorigenesisrdquo Neoplasia vol 7 no 9 pp 854ndash8612005
[13] Y He Y Wang P Li S Zhu J Wang and S Zhang ldquoIdenti-fication of GPX3 epigenetically silenced by CpG methylationin human esophageal squamous cell carcinomardquo DigestiveDiseases and Sciences vol 56 no 3 pp 681ndash688 2011
[14] B Chen X Rao M G House K P Nephew K J Cullenand Z Guo ldquoGPx3 promoter hypermethylation is a frequentevent in human cancer and is associatedwith tumorigenesis andchemotherapy responserdquo Cancer Letters vol 309 no 1 pp 37ndash45 2011
[15] D Lodygin A Epanchintsev A Menssen J Diebold andH Hermeking ldquoFunctional epigenomics identifies genes fre-quently silenced in prostate cancerrdquo Cancer Research vol 65no 10 pp 4218ndash4227 2005
[16] E Falck S Karlsson J Carlsson G Helenius M Karlsson andK Klinga-Levan ldquoLoss of glutathione peroxidase 3 expression iscorrelated with epigenetic mechanisms in endometrial adeno-carcinomardquo Cancer Cell International vol 10 article 46 2010
[17] X Zhang J-J Yang Y S Kim K-Y Kim W S Ahn and SYang ldquoAn 8-gene signature including methylated and down-regulated glutathione peroxidase 3 of gastric cancerrdquo Interna-tional Journal of Oncology vol 36 no 2 pp 405ndash414 2010
[18] Y Qu S Dang and P Hou ldquoGene methylation in gastriccancerrdquo Clinica Chimica Acta vol 424 pp 53ndash65 2013
[19] M F Kaiser D C Johnson P Wu et al ldquoGlobal methylationanalysis identifies prognostically important epigenetically inac-tivated tumor suppressor genes in multiple myelomardquo Bloodvol 122 no 2 pp 219ndash226 2013
[20] D F Peng T L Hu B G Schneider Z Chen Z K Xu and WEl-Rifai ldquoSilencing of glutathione peroxidase 3 through DNA
Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013

Oxidative Medicine and Cellular Longevity 9
hypermethylation is associated with lymph node metastasis ingastric carcinomasrdquo PLoS ONE vol 7 no 10 Article ID e462142012
[21] M A Nouh M M Mohamed M El-Shinawi et al ldquoCathepsinb a potential prognostic marker for inflammatory breast can-cerrdquo Journal of Translational Medicine vol 9 article 1 2011
[22] P Bonnier C Charpin C Lejeune et al ldquoInflammatorycarcinomas of the breast a clinical pathological or a clinicaland pathological definitionrdquo International Journal of Cancervol 62 no 4 pp 382ndash385 1995
[23] Y Gong ldquoPathologic aspects of inflammatory breast cancerpart 2 Biologic insights into its aggressive phenotyperdquo Seminarsin Oncology vol 35 no 1 pp 33ndash40 2008
[24] M M Mohamed D Al-Raawi S F Sabet and M El-ShinawildquoInflammatory breast cancer new factors contribute to diseaseetiology (Review)rdquo Journal of Advanced Research 2013
[25] P Storz ldquoReactive oxygen species in tumor progressionrdquo Fron-tiers in Bioscience vol 10 no 2 pp 1881ndash1896 2005
[26] I-C Lo J-M Shih and M J Jiang ldquoReactive oxygenspecies and ERK 12 mediate monocyte chemotactic protein-1-stimulated smoothmuscle cellmigrationrdquo Journal of BiomedicalScience vol 12 no 2 pp 377ndash388 2005
[27] W-S Wu ldquoThe signaling mechanism of ROS in tumor progres-sionrdquoCancer andMetastasis Reviews vol 25 no 4 pp 695ndash7052006
[28] C Genestie B Zafrani B Asselain et al ldquoComparison of theprognostic value of Scarff-Bloom-Richardson and Nottinghamhistological grades in a series of 825 cases of breast cancermajorimportance of themitotic count as a component of both gradingsystemsrdquo Anticancer Research vol 18 no 1 pp 571ndash576 1998
[29] D F Peng M Razvi H Chen et al ldquoDNA hypermethylationregulates the expression ofmembers of theMu-class glutathioneS-transferases and glutathione peroxidases in Barrettrsquos adeno-carcinomardquo Gut vol 58 no 1 pp 5ndash15 2009
[30] A Christians C Hartmann A Benner et al ldquoPrognostic valueof three different methods of MGMT promoter methylationanalysis in a prospective trial on newly diagnosed glioblastomardquoPLoS ONE vol 7 no 3 Article ID e33449 2012
[31] Y J Hu and A M Diamond ldquoRole of glutathione peroxidase 1in breast cancer loss of heterozygosity and allelic differences inthe response to seleniumrdquo Cancer Research vol 63 no 12 pp3347ndash3351 2003
[32] P Cejas M A Garcıa-Cabezas E Casado et al ldquoPhospholipidHydroperoxide Glutathione Peroxidase (PHGPx) expression isdownregulated in poorly differentiated breast invasive ductalcarcinomardquo Free Radical Research vol 41 no 6 pp 681ndash6872007
[33] C M Woolston A Al-Attar S J Storr I O Ellis D A LMorgan and S G Martin ldquoRedox protein expression predictsradiotherapeutic response in early-stage invasive breast cancerpatientsrdquo International Journal of Radiation Oncology BiologyPhysics vol 79 no 5 pp 1532ndash1540 2011
[34] C B Ambrosone ldquoOxidants and antioxidants in breast cancerrdquoAntioxidants and Redox Signaling vol 2 no 4 pp 903ndash9172000
[35] S Perwez Hussain M H Hollstein and C C Harris ldquop53tumor suppressor gene at the crossroads of molecular carcino-genesis molecular epidemiology and human risk assessmentrdquoAnnals of the New York Academy of Sciences vol 919 pp 79ndash852000
[36] F M Robertson M Bondy W Yang et al ldquoInflammatorybreast cancer the disease the biology the treatmentrdquoCACancerJournal for Clinicians vol 60 no 6 pp 351ndash375 2010
[37] M M Mohamed E A El-Ghonaimy M A Nouh R JSchneider B F Sloane and M El-Shinawi ldquoCytokines secretedby macrophages isolated from tumor microenvironment ofinflammatory breast cancer patients possess chemotactic prop-ertiesrdquoThe International Journal of Biochemistry amp Cell Biologyvol 46 pp 138ndash147 2014
[38] C W Barrett W Ning X Chen et al ldquoTumor suppressorfunction of the plasma glutathione peroxidase gpx3 in colitis-associated carcinomardquo Cancer Research vol 73 no 3 pp 1245ndash1255 2013
Copyright © 2022 FDOKUMEN




















